![]()
Contents
[Introduction] [Methodology of integrating biostratigraphic data]
[Integrated Upper Albian chronostratigraphy]
[Chronostratigraphic correlation of key sections]
[Re-examination of the rationale for a 'Vraconnian Stage']
[Conclusions] and ... [Bibliographic references]
Precision Stratigraphy Associates & University of Tulsa, RR3 Box 103-3, Cleveland, Oklahoma 74020 (U.S.A.)
Manuscript online since April 22, 2009
![]()
The Albian Stage is the highest chronostratigraphic unit of the Lower Cretaceous Series and underlies the Cenomanian Stage of the Upper Cretaceous Series. The Albian is divided into three substages, each of which is composed of two or three zones based on distinctive and phylogenetically related ammonite assemblages. The uppermost zone of the Upper Albian Substage, the Stoliczkaia dispar Zone, is found in many Western European condensed sections. The ammonite assemblage in the thin glauconitic sandstone near La Vraconne, Switzerland, was defined as the 'Vraconnian Stage' in 1868. However this concept has been little used and was abandoned in 1963 as part of the Cretaceous chronostratigraphic scale. A recent proposal to resurrect and redefine this stage is based on a number of criteria and very detailed and reliable stratigraphic data. A quantitative biostratigraphic analysis of the ammonite ranges in the key sections shows that the proposed subzones of the S. dispar Zone have discordant ranges. Furthermore, the utility of a 'Vraconnian Stage' between the Albian and Cenomanian stages is geographically limited and the concept embraces one of many depositional sequence cycles of the Albian. The reinstatement of a 'Vraconnian Stage' is not recommended.
Albian; Vraconnian; ammonites; planktic foraminifers; graphic correlation; age calibration.
R.W. (2009).- Uppermost Albian biostratigraphy and chronostratigraphy.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2009/03 (CG2009_A03)
Biostratigraphie et chronostratigraphie de l'Albien sommital/terminal.- L'étage Albien constitue l'unité chronostratigraphique la plus élevée du Crétacé inférieur et repose sous l'étage Cénomanien du Crétacé supérieur. L'Albien est divisé en trois sous-étages, chacun comprenant deux ou trois zones établies sur des associations d'ammonites distinctes mais phylogénétiquement reliées. La zone sommitale du sous-étage Albien supérieur, la Zone à Stoliczkaia dispar, a été identifiée au sein de nombreuses séries condensées en Europe occidentale. L'association d'ammonite reconnue dans le mince niveau de grès glauconieux des environs de La Vraconne (Suisse) a été définie comme 'étage Vraconnien' en 1868. Toutefois ce concept a été peu usité et fut abandonné en 1963 en tant qu'unité de l'échelle chronostratigraphique du Crétacé. Il a été récemment proposé de réhabiliter et de redéfinir cet étage sur la base d'un certain nombre de critères et de données stratigraphiques très détaillées et fiables. Or une analyse biostratigraphique quantitative des répartitions des ammonites dans les coupes clefs révèle que les sous-zones proposées pour la subdivision de la Zone à S. dispar correspondent à des intervalles non concordants. En outre l'intérêt de placer un 'étage Vraconnien' entre les étages Albien et Cénomanien apparaît géographiquement limité, et le concept correspond à une seule des nombreuses séquences de dépôt de l'Albien. Il n'est donc pas recommandé de restaurer un 'étage Vraconnien'.
Albien ; Vraconnien ; ammonites ; foraminifères planctoniques ; correlation graphique ; calibration des âges.
Recently (2002, 2008), & (2008) and et alii (2007) proposed to reinstate the 'Vraconnian Stage' between the Albian and Cenomanian stages. This proposition, which would significantly modify the Cretaceous geologic column, merits careful and thorough analysis. Our objectives are to provide a testable global biostratigraphic database with which to evaluate the ranges of key uppermost Albian species and to calibrate their ranges to a numerical time scale.
The Cretaceous System is composed of twelve stages that can be correlated world-wide and are of different durations (,
2003). The Albian Stage is the youngest chronostratigraphic unit of the Lower Cretaceous Series and one of the longest Cretaceous stages, about 13 to 15 myr. The Albian Stage was defined by d'
(1840-1842) to include the fossil assemblages in shale and glauconite sands cropping out along the Aube River in the Department of Aube on the eastern margin of the Paris Basin ( & ,
1980). A stratotype section along the Aube River was composited from outcrop sections and nearby boreholes ( et alii,
1965; , 1992). The Albian biota in the Paris Basin is transitional between Boreal and Tethyan realms. The concepts of Albian ammonite zones have evolved over a period of time beginning in
1868 when the
"Ammonites mammillaris" zone (de ,
1868) (now the
Douvilleiceras mammillatum Zone) and the "zone of Ammonites inflatus" (de , 1868) (now considered equivalent to the Stoliczkaia dispar Zone) were proposed ( et alii,
1978). The current zonal scheme
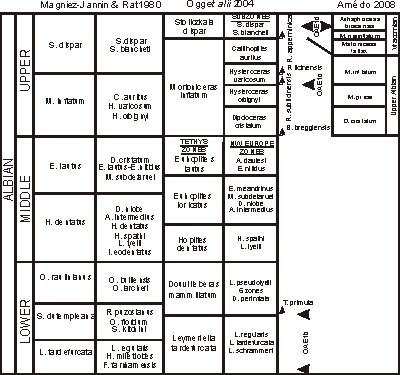
(Fig. 1 ![]() ) was composed by
(1923) and (1947) and modified by
(1971) and (1992). The zonal succession has been relatively stable since
1947 and most are interval zones defined by
the first appearance of an ammonite species (FO; ,
1992).
) was composed by
(1923) and (1947) and modified by
(1971) and (1992). The zonal succession has been relatively stable since
1947 and most are interval zones defined by
the first appearance of an ammonite species (FO; ,
1992).
The Albian Stage is divided into three substages, Lower, Middle and Upper. Although stratotype sections for these substages are yet to be agreed upon, here they are used formally following et alii
(1996) and are capitalized. The zonal boundaries of the substages have been used consistently since
1947 (Fig. 1 ![]() ). Prior to
1947, however, two different criteria were used to define the base of the Upper Albian Substage.
(1923, 1941) placed the boundary at the top of the
Dipoloceras cristatum Subzone, which directly overlies the Euhoplites lautus Zone (,
1971). Alternatively
(1947) placed the Cristatum Subzone in the basal Upper Albian Mortoniceras inflatum Zone. This later opinion has been followed since ( et alii,
1984; , 1984a,
1984b; , 1991;
et alii, 1996; & ,
1999; & ,
2000; et alii,
2003). The rationale for this change is reviewed by
(1971) and et alii
(1996). In North America a regional transgressive unconformity coincides with the top boundary of the Cristatum Subzone, which is a widespread mappable contact ( et alii,
2003).
). Prior to
1947, however, two different criteria were used to define the base of the Upper Albian Substage.
(1923, 1941) placed the boundary at the top of the
Dipoloceras cristatum Subzone, which directly overlies the Euhoplites lautus Zone (,
1971). Alternatively
(1947) placed the Cristatum Subzone in the basal Upper Albian Mortoniceras inflatum Zone. This later opinion has been followed since ( et alii,
1984; , 1984a,
1984b; , 1991;
et alii, 1996; & ,
1999; & ,
2000; et alii,
2003). The rationale for this change is reviewed by
(1971) and et alii
(1996). In North America a regional transgressive unconformity coincides with the top boundary of the Cristatum Subzone, which is a widespread mappable contact ( et alii,
2003).
The Albian Stage comprises the evolutionary origins and/or diversification of seven families of ammonites. Zones of the Lower, Middle and Upper Albian substages are based mainly on species of the families of Lyelliceratidae, Hoplitidae, and Brancoceratidae. Four heteromorph families first appear in the Albian, Anisoceratidae, Baculitidae, Hamitidae, and Turrilitidae, but are not used to define zones until their appearance in the Upper Cretaceous.
Click on thumbnail to enlarge the image.
Figure 1: Chart comparing three stage and substage concepts of the Albian (P. , 1979 in & , 1980; et alii, 2004, and , 2008). The positions of Albian anoxic events and key microfossils were interpolated by et alii (2004).
The 'Vraconnian Stage' was proposed by
(1868) for a 2 m-thick condensed interval of green glauconitic sand with a distinctive ammonite fauna between the Upper Albian and Lower Cenomanian substages near La Vraconne in western Switzerland. The ammonite assemblage is part of the Stoliczkaia dispar Zone and the lithostratigraphic unit correlates with the Upper Gault and Upper Greensand in England
(Fig. 1 ![]() ). This stage was discarded in 1963 (,
1965; et alii,
1978; , 1991,
2003, among many others). However the 'Vraconnian Stage' has been resurrected and redefined by
(2002,
2008), &
(2008) and by et alii
(2007). cites five reasons for recognizing 'Vraconnian' sedimentary strata as a stage between the Albian and Cenomanian stages: (1) the interval is mappable in Western Europe, (2) the interval is a third-order depositional cycle that records an important eustatic event, (3) the interval has a distinctive and diverse fossil assemblage that can be recognized outside Europe, (4) its 2 to 3 myr duration is equivalent to that of the Santonian Stage, and (5) in the Vocontian Basin the interval is more than 100 m thick, which is thicker than the underlying part of the Albian (,
2002,
2008). has presented detailed lithostratigraphic and biostratigraphic data of twelve sections in Europe, Tunisia, Madagascar, and California to support his proposal. For the first time he presents a regional lithostratigraphic correlation of many of these sections.
(2003) reviewed the history of the rejection of the 'Vraconnian' concept by the community of Lower Cretaceous stratigraphers beginning in 1963 and believed that reasons to revive 'Vraconnian' were "trivial". He noted that most 'Vraconnian' sections are condensed intervals.
). This stage was discarded in 1963 (,
1965; et alii,
1978; , 1991,
2003, among many others). However the 'Vraconnian Stage' has been resurrected and redefined by
(2002,
2008), &
(2008) and by et alii
(2007). cites five reasons for recognizing 'Vraconnian' sedimentary strata as a stage between the Albian and Cenomanian stages: (1) the interval is mappable in Western Europe, (2) the interval is a third-order depositional cycle that records an important eustatic event, (3) the interval has a distinctive and diverse fossil assemblage that can be recognized outside Europe, (4) its 2 to 3 myr duration is equivalent to that of the Santonian Stage, and (5) in the Vocontian Basin the interval is more than 100 m thick, which is thicker than the underlying part of the Albian (,
2002,
2008). has presented detailed lithostratigraphic and biostratigraphic data of twelve sections in Europe, Tunisia, Madagascar, and California to support his proposal. For the first time he presents a regional lithostratigraphic correlation of many of these sections.
(2003) reviewed the history of the rejection of the 'Vraconnian' concept by the community of Lower Cretaceous stratigraphers beginning in 1963 and believed that reasons to revive 'Vraconnian' were "trivial". He noted that most 'Vraconnian' sections are condensed intervals.
As a framework for discussing the wisdom of revising the Albian Stage by
reinstating the 'Vraconnian Stage', a review of the stage concept is relevant. "A stage is a chronostratigraphic unit of smaller scope and rank than a series. It is most commonly of greatest use in intra-continental classification and correlation, although it has the potential for worldwide recognition" (NACSN,
2005, p. 1582). As a chronostratigraphic unit the Albian Stage has synchronous boundaries and is the physical evidence or 'material referent' of a time interval, the Albian Age. The Albian Stage has traditionally been defined by a set of ammonite biozones
(Fig. 1 ![]() ):
d', 1840-1842; ,
1923;
P. & J.-P. ,
1965; , 1984a,
1984b; , 1991; et alii,
1996, among
others. The Global Stratotype Section and Point (GSSP) have yet to be selected to define the basal boundary although an excellent section has been proposed ( et alii,
2002). The upper boundary of the Albian Stage is defined by the base Cenomanian Stage GSSP at Mont Risou, France ( et alii,
1996; et alii,
2004).
):
d', 1840-1842; ,
1923;
P. & J.-P. ,
1965; , 1984a,
1984b; , 1991; et alii,
1996, among
others. The Global Stratotype Section and Point (GSSP) have yet to be selected to define the basal boundary although an excellent section has been proposed ( et alii,
2002). The upper boundary of the Albian Stage is defined by the base Cenomanian Stage GSSP at Mont Risou, France ( et alii,
1996; et alii,
2004).
The integrated ranges of select ammonites in seven key sections defined three zones
of the Vraconnian (
2008,
Fig. 4  ;
Fig. 1
;
Fig. 1 ![]() ,
Table 1). The ranges of key planktic foraminifera in the Tunisian sections and the French Mont Risou section were added to that set of species by et alii
(2007). However most of the ammonite species are found in only one or two sections (,
2008). To test the accuracy of the integration of species ranges the quantitative technique of graphic correlation is used in this report. Graphic correlation (GC) provides an objective method to compare the ranges of species in multiple sections and the outcome can be tested independently.
,
Table 1). The ranges of key planktic foraminifera in the Tunisian sections and the French Mont Risou section were added to that set of species by et alii
(2007). However most of the ammonite species are found in only one or two sections (,
2008). To test the accuracy of the integration of species ranges the quantitative technique of graphic correlation is used in this report. Graphic correlation (GC) provides an objective method to compare the ranges of species in multiple sections and the outcome can be tested independently.
Graphic correlation (GC) is a quantitative, non-statistical, technique that determines the coeval relationships between two sections by comparing the ranges of event records in both sections ( & , 1995). A graph of any pair of sections is an X/Y plot of the FOs (first appearances) and LOs (last appearances) of taxa found in both sections. The interpreter places a line of correlation (LOC) through the tops and bases that are at their maximum range in both sections. This LOC is the most constrained hypothesis of synchroneity between the two sections and extends the fewest bioevents. The LOC also accounts for hiatuses or faults at stratal discontinuities indicated by the lithostratigraphic record. The position of the LOC is defined by the equation for a regression line. Explanation and examples of the graphic technique are illustrated by (1977) and & (1995). By iteratively graphing successive sections a database of species ranges is compiled. The accuracy of these ranges depends on the number of sections, preservation and correct identification of the species. Such a database is testable and the process is transparent so that the fossil occurrence in each section can be evaluated to determine its accuracy. This process compiles data of many specialists who have studied numerous global sections.
| Mont Risou, France, Section | FO meters | LO meters |
| Base limestone interval | 330 m | |
| Actinoceramus sulcatus | 50 | 78 |
| Actinoceramus subsulcatus | 25 | |
| Anisoceras salei | 78 | |
| Anisoceras perarmatum | 78 | |
| Arrhaphoceras briacensis | 305 | |
| Dipoloceras cristatum | 18 | 25 |
| Hysteroceras orbignyi (as sp.) | 78 | |
| Lechites moreti | 155 | |
| Mantelliceras mantelli | 205 | |
| Mariella gresslyi | 155 | |
| Mortoniceras perinflatum | 205 | |
| Rota globotruncanoides | 295 | |
| Turrilitoides hugardianus | 155 | |
| Folkestone, UK, Section | ||
| Base cenomanian ammonites | 38 | |
| Inoc concentricus | 0 | 10.5 |
| Actinoceramus sulcatus | 9.7 | 13 |
| Anisoceras perarmatum | 26 | 38 |
| Arrhaphoceras substuderi | 26 | 38 |
| Callihoplites auritus | 14.5 | 24.5 |
| Callihoplites cantabrigense | 25 | |
| Callihoplites leptus | 25 | |
| Callihoplites tetragonus | 26 | 38 |
| Callihoplites vraconensis | 26 | 38 |
| Hyphoplites coelonotus | 25 | |
| Lepthoplites falcoides | 26 | 38 |
| Mortoniceras inflatum | 14.5 | 25 |
| Pleurohoplites renauxianus | 26 | 38 |
| Merstham, UK | ||
| Anisoceras picteti | 4.7 | 5 |
| Callihoplites seeleyi | 7 | 7.2 |
| Callihoplites vraconensis | 4.7 | 7.2 |
| Idohamites elegantulus | 4.7 | 5 |
| Lechites gaudini | 7 | |
| Lepthoplites falcoides | 4.7 | 5 |
| Lepthoplites pseudoplanus | 4.7 | 7.2 |
| Mortoniceras alstonensis | 4.7 | 5 |
| Mortoniceras fallax | 4.7 | 5 |
| Neophlycticeras blancheti | 4.7 | 6.6 |
| Ostlingoceras puzosianum | 7 | 7.2 |
| Pleurohoplites renauxianus | 7 | 7.2 |
| Harchies, Belgium | ||
| Anisoceras perarmatum | -87.1 | |
| Anisoceras pseudoelegans | -95.5 | |
| Callihoplites vraconensis | -87.1 | |
| Hamites virgulatus | -98.4 | |
| Hyphoplites subfalcatus (ID cf.) | -86.1 | |
| Lepthoplites cantabrigiensis | -102.1 | |
| Pleurohoplites subvarians | -87.1 | |
| Strépy, Belgium, Outcrop | ||
| Callihoplites seeleyi | 30 | |
| Callihoplites pulcher | 30 | |
| Callihoplites tetragonus | 20 | |
| Cantabrigites subsimplex | 30 | |
| Hyphoplites valbonnensis | 20 | |
| Lechites gaudini | 20 | |
| Mortoniceras fallax | 20 | |
| Neophlycticeras blancheti | 20 | |
| ANDRA MAR 203 Core, France | ||
| Hamites virgulatus (ID cf.) | -523.23 | |
| Hyphoplites coelonotus | -776.75 | -523.38 |
| Hyphoplites falcatus | -455.85 | |
| Hyphoplites valbonnensis | -487.17 | |
| Lechites gaudini | -487.17 | |
| Mariella bergeri (ID cf.) | -467.25 | |
| Ostlingoceras puzosianum | -561.54 | |
| Pleurohoplites renauxianus | -581.8 | |
| Schloenbachia varians | -430.8 | |
| Diégo core, Madagascar | ||
| Lechites gaudini | -50 | |
| Mantelliceras mantelli | -23 | -5 |
| Mariella bergeri | -20 | |
| Neostlingoceras carcitanense | -23 | -5 |
| Scaphites simplex | -54 | |
| Sciponoceras roto | -23 | -5 |
| Stoliczkaia dispar | -50 | |
| Biti breggiensis | -215 | -118 |
| Planomalina buxtorfi | -106 | -10 |
| Planomalina praebuxtorfi | -118 | -90 |
| Rota appenninica | -131 | -10 |
| Rota brotzeni | -23 | -10 |
| Rota globotruncanoides | -23 | -10 |
| Tici praeticinensis | -215 | -180 |
| Tici primula | -236 | -135 |
| Tici subticinensis | -215 | -155 |
| Tici ticinensis | -180 | -106 |
Table 1: Biostratigraphic species and ranges in meters of each section from (2008).
To begin the GC experiment the Mont Risou section, southeastern France ( et alii,
1996;
Fig. 2 ![]() ) was selected as the standard reference section because it records continuous basin deposition at a uniform rate of accumulation and it yields diverse ammonites, inoceramids, planktic foraminfera, and nannofossils. This section was cross plotted to itself setting its thickness in meters as the relative time scale. Subsequently seven other sections, which yielded precise and abundant biostratigraphic data, were plotted to it through multiple rounds and the ranges of the fossils extended to the thickness scale at Mont Risou. The first section plotted was the Kalaat Senan, Tunisia (,
2008,
Fig. 20 ), which spans from uppermost Albian to Cenomanian; this section also represents uniform deposition. The result is that the scale was extended from the base of the Upper Albian to the Cenomanian/Turonian contact.
) was selected as the standard reference section because it records continuous basin deposition at a uniform rate of accumulation and it yields diverse ammonites, inoceramids, planktic foraminfera, and nannofossils. This section was cross plotted to itself setting its thickness in meters as the relative time scale. Subsequently seven other sections, which yielded precise and abundant biostratigraphic data, were plotted to it through multiple rounds and the ranges of the fossils extended to the thickness scale at Mont Risou. The first section plotted was the Kalaat Senan, Tunisia (,
2008,
Fig. 20 ), which spans from uppermost Albian to Cenomanian; this section also represents uniform deposition. The result is that the scale was extended from the base of the Upper Albian to the Cenomanian/Turonian contact.
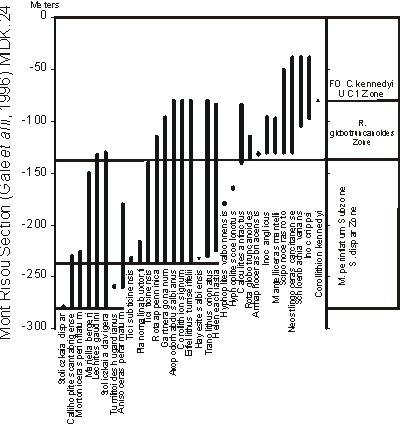
Click on thumbnail to enlarge the image.
Figure 2: Biostratigraphic range chart of Mont Risou Albian-Cenomanian reference section showing key ammonites, planktic foraminifera and nannofossils (from et alii, 1996).
The second graphed section was the Diégo well, N Madagascar (,
2008,
Fig. 22 ). This 300 m cored interval is mainly marl that spans an interval from lowermost Cenomanian to middle Albian
(Fig. 3D ![]() ). The section yields several key planktic foraminifera and in three
samples a few ammonites. The LOC of the graph is constrained by a small number of first occurrences and it extends the bases of many foraminifera. The section appears to record continuous Late Albian deposition.
). The section yields several key planktic foraminifera and in three
samples a few ammonites. The LOC of the graph is constrained by a small number of first occurrences and it extends the bases of many foraminifera. The section appears to record continuous Late Albian deposition.
The third section graphed was the cored interval in the ANDRA MAR 203 well (,
2008,
Fig.
8 ). This interval is about 560 m thick and spans from the Cenomanian to the Aptian stages
(Fig. 3C ![]() ). A significant unconformity, probably a sequence boundary, is at the base of the Albian Valbonne Formation (-827 m), which is a
glauconitic sandstone that overlies Aptian marls. A second unconformity, a transgressive contact, is at the base of a phosphatic interval in the upper part of the Valbonne at -797.9
m. The Valbonne grades up into the Marcoule Formation. Biostratigraphic data appear 18 m above this upper break and
span an interval about 350 m thick. In this section placed the base of the 'Vraconnian' at the unconformity in the upper part of the Valbonne so that the 'Vraconnian' is 334 m thick. The LOC of the section is constrained by the FO and LO of
Hyphoplites coelonotus and the LOs of three other ammonites (Fig. 3C
). A significant unconformity, probably a sequence boundary, is at the base of the Albian Valbonne Formation (-827 m), which is a
glauconitic sandstone that overlies Aptian marls. A second unconformity, a transgressive contact, is at the base of a phosphatic interval in the upper part of the Valbonne at -797.9
m. The Valbonne grades up into the Marcoule Formation. Biostratigraphic data appear 18 m above this upper break and
span an interval about 350 m thick. In this section placed the base of the 'Vraconnian' at the unconformity in the upper part of the Valbonne so that the 'Vraconnian' is 334 m thick. The LOC of the section is constrained by the FO and LO of
Hyphoplites coelonotus and the LOs of three other ammonites (Fig. 3C ![]() ). A short hiatus is suggested by the offset of the LOC in the basal Cenomanian. An alternative interpretation is that the interval recorded a reduced rate of sediment accumulation beginning in latest
Albian times. At the Marcoule locality the complete section, from its base at -781 m to the base of the Cenomanian at about -464 m, correlates with zones XII-XIII in the near shore Folkestone section, where these
same zones are only 13.8 m thick.
). A short hiatus is suggested by the offset of the LOC in the basal Cenomanian. An alternative interpretation is that the interval recorded a reduced rate of sediment accumulation beginning in latest
Albian times. At the Marcoule locality the complete section, from its base at -781 m to the base of the Cenomanian at about -464 m, correlates with zones XII-XIII in the near shore Folkestone section, where these
same zones are only 13.8 m thick.
The fourth section to be added was the Mont Risou section in
(2008,
Fig. 11 ), which was based on data from multiple sources including et alii
(1996). Thus the slope of the LOC is the same as that of the standard reference section. This plot is well constrained by several first and last occurrences; it extends some ranges and it adds three new species not
found in other sections (Fig. 3A ![]() ).
).
The next section plotted was the Gault Clay in the Folkestone section in
(2008,
Fig. 14 ). This section spans several significant unconformities
(Fig. 3B ![]() ), one at the base of the Gault Clay overlying the Lower Greensand, a second at the condensed interval VIII zone, a third at the XII zone, and the highest at the Albian/Cenomanian contact between the Gault and the glauconitic marl (,
2000). The LOC of the lower interval of zones I-VII is poorly constrained at the top by the FO of Actinoceramus sulcatus and its slope is the same as that of the higher LOCs. The unconformity break is placed at the top of zone VIII. The LOC in the next higher interval of zones IX-XI is constrained by the range of
Callihoplites auritus. In the next higher section of zones XII-XIII the LOC is constrained by several FO and LO bioevents
(Fig. 3B
), one at the base of the Gault Clay overlying the Lower Greensand, a second at the condensed interval VIII zone, a third at the XII zone, and the highest at the Albian/Cenomanian contact between the Gault and the glauconitic marl (,
2000). The LOC of the lower interval of zones I-VII is poorly constrained at the top by the FO of Actinoceramus sulcatus and its slope is the same as that of the higher LOCs. The unconformity break is placed at the top of zone VIII. The LOC in the next higher interval of zones IX-XI is constrained by the range of
Callihoplites auritus. In the next higher section of zones XII-XIII the LOC is constrained by several FO and LO bioevents
(Fig. 3B ![]() ). This section plot defines the relative duration of the hiatus of two unconformities as meter thicknesses in the Mont Risou section; these intervals can now be projected into other graphed sections.
). This section plot defines the relative duration of the hiatus of two unconformities as meter thicknesses in the Mont Risou section; these intervals can now be projected into other graphed sections.
An older ammonite data set from Folkestone by
(1992) is graphed by several FO and LO's. The unconformities are well constrained as in the newer section in
(2008). The Merstham, England (,
2008,
Fig. 15 ) repeats the uppermost part of the Upper Greensand Formation that is present at Folkestone. The data are confined to two horizons so the LOC is not well constrained. However
these data adjust nine FO's and nine LO's. The Strépy and Harches sections were graphed next (,
2008,
Fig. 17 ) and have limited data so that the LOC of each is not tightly constrained.
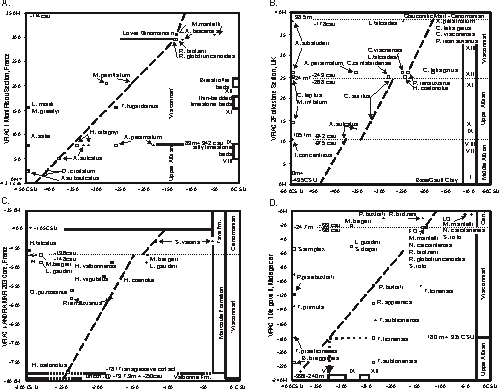
Click on thumbnail to enlarge the image.
Figure 3: Graphic correlation interpretations of four key 'Vraconnian' sections. First occurrences (FO) of species are indicated by a square symbol (□) and the last occurrence (LO) by a plus sign (+). Superposed symbols of a species indicates a single occurrence. Symbols on the left of the Y axis indicate that the occurrence is only in the section they accompany. CSU is a composite standard unit calibrated in meters to the thickness of the Mont Risou section (file MIDK24). Horizontal solid and dotted lines are stratigraphic contacts; inclined dashed lines project a position in the section into the position of a species in the set of sections in the database of "Vraconnian" sections; these lines are 'lines of correlation' (LOC). Roman numerals indicate ammonite zones in the English sections.
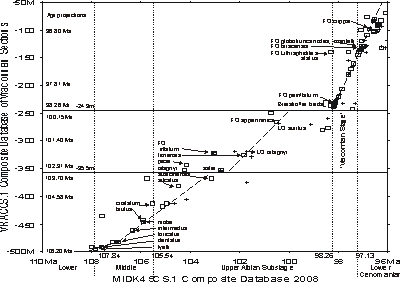
The graphic correlation experiment resulted in a list of species and their ranges relative to each other in the metric scale of the Mont Risou section. The next step in the experiment was to convert the metric scale to a numerical age scale in mega-annum units. This was accomplished by plotting the composited range data set of 's 'Vraconnian' data set, VRACCS.1, to the MIDK45CS.1 composited range data set
(Fig. 4 ![]() ). The MIDK45CS.1 data set is the next development stage beyond MIDK3 ( et alii,
2000) and MIDK42CS.1 (,
in press; see also data of MIDK42 on website precisionstratigraphy.com). It is composed of more than one hundred sections and nearly 3000 bioevents, geochemical events, magnetochrons, and sequence stratigraphic contacts. The scale of this range data set is the numerical time scale of et alii
(1990),
as revised by et alii (2004), with the exception
of the age ascribed the base Cenomanian, 97.13 Ma, because correlations of radiometrically dated bentonites and bioevents
have been re-evaluated ( et alii,
2007).
). The MIDK45CS.1 data set is the next development stage beyond MIDK3 ( et alii,
2000) and MIDK42CS.1 (,
in press; see also data of MIDK42 on website precisionstratigraphy.com). It is composed of more than one hundred sections and nearly 3000 bioevents, geochemical events, magnetochrons, and sequence stratigraphic contacts. The scale of this range data set is the numerical time scale of et alii
(1990),
as revised by et alii (2004), with the exception
of the age ascribed the base Cenomanian, 97.13 Ma, because correlations of radiometrically dated bentonites and bioevents
have been re-evaluated ( et alii,
2007).
The two unconformities in the uppermost Albian Upper Greensand Formation are well defined by the data plot (Fig. 4 ![]() ). The two hiatuses separate the VRACCS.1 section into three intervals; the lower break is at -355 meters and the upper break is at -249 meters, which is the base of the 'Vraconnian Stage'. The duration of the lower hiatus is 103.70 to 102.91 Ma, and
that of the upper hiatus 100.15 to 98.26 Ma. The LOC of the lower interval is well constrained by numerous bioevents. This lower interval represents two rates of sediment accumulation, a slower basal interval and a faster upper interval. This rate change is indicated by the change in the LOC slope. The LOC of the middle interval is tightly constrained by ammonites and planktic foraminifera. The upper interval includes numerous bioevents and the LOC is very tightly constrained. The age of the 'Vraconnian Stage' spans from 98.26 to 97.13 Ma. The black shale, organic-rich marker beds were deposited at the beginning of the 'Vraconnian'.
). The two hiatuses separate the VRACCS.1 section into three intervals; the lower break is at -355 meters and the upper break is at -249 meters, which is the base of the 'Vraconnian Stage'. The duration of the lower hiatus is 103.70 to 102.91 Ma, and
that of the upper hiatus 100.15 to 98.26 Ma. The LOC of the lower interval is well constrained by numerous bioevents. This lower interval represents two rates of sediment accumulation, a slower basal interval and a faster upper interval. This rate change is indicated by the change in the LOC slope. The LOC of the middle interval is tightly constrained by ammonites and planktic foraminifera. The upper interval includes numerous bioevents and the LOC is very tightly constrained. The age of the 'Vraconnian Stage' spans from 98.26 to 97.13 Ma. The black shale, organic-rich marker beds were deposited at the beginning of the 'Vraconnian'.
Click on thumbnail to enlarge the image.
Figure 4: Graphic correlation plot of VRACCS.1 database to the global integrated MIDK45CS.1 database.
The stratigraphic ranges of 46 ammonites that were recorded in the seven key sections of
(2008) were integrated into a single range chart at the scale of the thickness of the Mont Risou section
(Fig. 5 ![]() ). This scale was converted to a numerical age scale by graphing the composited VRACCS.1 data set to the MIDK45CS.1 data set as explained above. The three Middle Albian Substage zones are defined by the FO of the nominate species
(Fig. 1
). This scale was converted to a numerical age scale by graphing the composited VRACCS.1 data set to the MIDK45CS.1 data set as explained above. The three Middle Albian Substage zones are defined by the FO of the nominate species
(Fig. 1 ![]() ;
Table 2). The base of the Upper Albian Substage is defined by the FO of
Dipoloceras cristatum, which is the basal subzone of the Inflatum Zone. Two of the next overlying subzones are defined in this dataset by the FOs of
Hysteroceras orbignyi and Callihoplites auritus; Hysteroceras varicosum is not included in this dataset. The base of Stoliczkaia dispar defines the base of the uppermost Upper Albian Dispar Zone.
;
Table 2). The base of the Upper Albian Substage is defined by the FO of
Dipoloceras cristatum, which is the basal subzone of the Inflatum Zone. Two of the next overlying subzones are defined in this dataset by the FOs of
Hysteroceras orbignyi and Callihoplites auritus; Hysteroceras varicosum is not included in this dataset. The base of Stoliczkaia dispar defines the base of the uppermost Upper Albian Dispar Zone.
(2008) proposed that the base of the 'Vraconnian Stage' be defined by the FOs of Mortoniceras fallax and Neophlycticeras blancheti. He included three ammonite zones in this stage in ascending order: Mortoniceras (Mortoniceras) fallax Interval Zone, Mortoniceras (Subschloenbachia) perinflatum Total Range Zone, and the Arrhaphoceras (Praeschloenbachia) briacensis Interval Zone
(Fig. 1 ![]() ). The composited dataset of his sections shows that M. inflatum first appears lower in the section than M. fallax
(Fig. 5
). The composited dataset of his sections shows that M. inflatum first appears lower in the section than M. fallax
(Fig. 5 ![]() ) because et alii
(1996) reported it lower in their Mont Risou section than did
(2008,
Fig. 11 ). The FO of A. briacensis is at the base of the Cenomanian Stage based on this set of sections. The ranges of 's key zonal species (,
2008,
Fig. 4 ) are dashed lines on
Fig. 5
) because et alii
(1996) reported it lower in their Mont Risou section than did
(2008,
Fig. 11 ). The FO of A. briacensis is at the base of the Cenomanian Stage based on this set of sections. The ranges of 's key zonal species (,
2008,
Fig. 4 ) are dashed lines on
Fig. 5 ![]() ; some ranges are similar and others are quite different from the ranges derived by graphic correlation of his data. S. dispar has a short range because it occurs in two sections, the
Diégo well and 's et alii Mont Risou section. Both Mariella bergeri and
Stoliczkaia clavigera are low in the Mont Risou section ( et alii,
1996). Some species have longer ranges in 's dataset because they occur with species of other zones in condensed intervals in other sections not graphed, such as at the 'Vraconnian' stratotype ( & ,
1978) and near Drap in the Alpes-Maritimes, France ( & ,
1988).
; some ranges are similar and others are quite different from the ranges derived by graphic correlation of his data. S. dispar has a short range because it occurs in two sections, the
Diégo well and 's et alii Mont Risou section. Both Mariella bergeri and
Stoliczkaia clavigera are low in the Mont Risou section ( et alii,
1996). Some species have longer ranges in 's dataset because they occur with species of other zones in condensed intervals in other sections not graphed, such as at the 'Vraconnian' stratotype ( & ,
1978) and near Drap in the Alpes-Maritimes, France ( & ,
1988).
This graphic correlation experiment shows that the base of the S. dispar Zone and the FO of M. perinflatum are significantly lower (31 m) and older (380 kyr) than the FO of either M. fallax or N. blancheti. Thus the concept of a 'Vraconnian Stage' is not equal to the Dispar Zone. However the FOs of M. perinflatum, M. fallax/N. blancheti, and A. briacensis may be used to divide the Dispar Zone into three subzones.
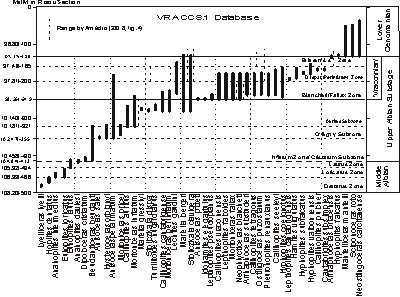
Click on thumbnail to enlarge the image.
Figure 5:
Chronostratigraphic range chart of ammonites in 'Vraconnian' composited database of key 'Vraconnian' sections. Ages interpolated by graphic correlation of the VRACCS.1 database with the MIDK45CS.1 database. The ages of some ranges are younger in the subset of 'Vraconnian' sections than in the complete set of sections in the MIDK45CS database used in
Figure 7 ![]() .
.
| TAXA | VRACCS: FO - LO | MIDK45CS: FO - LO | ||
| Acanthoceras amphibolum | 65.0472 | 65.0472 | 94.3454 | 92.6212 |
| Acanthoceras rhotomagense | 5.9825 | 43.151 | 94.9483 | 92.5959 |
| Actinoceramus subsulcatus | -406.00 | -406.00 | 104.79* | 104.79* |
| Actinoceramus sulcatus | -381.00 | -329.0148 | 104.4776 | 101.5134 |
| Algericeras boghariense | -103.4988 | -93.7467 | 96.5874 | 96.2374 |
| Algericeras proratum | -92.6427 | -64.6744 | 96.4245 | 95.578 |
| Amphizygus brooksii | -236.00 | -80.00 | 124.717 | 65.505 |
| Anahoplites daviesi | -414.8517 | -410.6029 | 105.1156 | 104.8296 |
| Anahoplites intermedius | -465.8373 | -450.2584 | 107.1613 | 106.32 |
| Anisoceras perarmatum | -353.00 | -180.0123 | 98.426 | 97.5888 |
| Anisoceras picteti | -248.88 | -247.2 | 94.8091 | 94.3577 |
| Anisoceras pseudoelegans | -188.2678 | -188.2678 | 97.68* | 97.68* |
| Anisoceras salei | -353.00 | -353.00 | 102.7656 | 102.7656 |
| Arrhaphoceras briacensis | -136.00 | -132.00 | 97.0865 | 97.0865 |
| Arrhaphoceras substuderi | -236.8768 | -180.3551 | 98.18* | 97.6* |
| Axopodorhabdus albianus | -236.00 | 102.7677 | 109.77 | 90.6992 |
| Axopodorhabdus dietzmannii | -139.916 | 35.7909 | 128.6386 | 82.56 |
| Beudanticeras beudanti | -410.6029 | -321.1158 | 104.9638 | 103.0425 |
| Biticinella breggiensis | -367.6759 | -250.0185 | 105.757 | 97.5059 |
| Calculites anfractus | -140.00 | -84.00 | 97.1702 | 96.5842 |
| Callihoplites auritus | -321.1924 | -269.043 | 103.0146 | 100.4255 |
| Callihoplites cantabrigense | -280.00 | -230.00 | 98.6353 | 98.1121 |
| Callihoplites leptus | -241.587 | *** | 98.23* | *** |
| Callihoplites pulcher | -170.00 | -170.00 | 97.49* | 97.49* |
| Callihoplites seeleyi | -236.00 | -170.00 | 98.17* | 97.49* |
| Callihoplites tetragonus | -236.8768 | -180.00 | 98.18* | 97.60* |
| Callihoplites vraconensis | -248.88 | -180.0123 | 97.49* | 97.60* |
| Cantabrigites subsimplex | -170.00 | -170.00 | 97.49* | 97.49* |
| Carbon peak OAE 2 | 100.9276 | 104.9757 | 93.52 | 93.45 |
| Chiastozygus bifarius | -236.00 | -84.00 | 98.1749 | 65.505 |
| Chiastozygus litterarius | -236.00 | -80.00 | 128.2573 | 65.5535 |
| Chiastozygus platyrhethus | -236.00 | -80.00 | 125.5054 | 88.3575 |
| Corollithion kennedyi | -80.00 | 101.6637 | 96.55 | 92.00 |
| Corollithion madagaskarensis | -236.00 | -80.00 | 122.358 | 65.3262 |
| Corollithion signum | -236.00 | -80.00 | 114.0455 | 70.4437 |
| Cretarhabdus conicus | -236.00 | -80.00 | 133.1464 | 64.5992 |
| Cretarhabdus striatus | -236.00 | -80.00 | 117.1136 | 65.505 |
| Cribrosphaerella ehrenbergii | -236.00 | -80.00 | 116.0163 | 94.3685 |
| Dicarinella algeriana | 38.73 | 118.2239 | 95.105 | 89.7362 |
| Dicarinella hagni | 80.3194 | 118.2239 | 93.8363 | 89.1675 |
| Dimorphoplites niobe | -449.5502 | -436.8038 | 106.3502 | 105.8965 |
| Dipoloceras cristatum | -413.00 | -406.00 | 105.5386 | 104.1574 |
| Discorhabdus ignotus | -236.00 | -80.00 | 128.6386 | 65.505 |
| Eiffellithus turriseiffelii | -236.00 | -80.00 | 101.856 | 64.4399 |
| Ellipsagelosphaera ovata | -216.00 | -112.00 | 121.959 | 88.1021 |
| Eprolithus apertior | -236.00 | -80.00 | 121.1235 | 96.5423 |
| Eprolithus floralis | -236.00 | -84.00 | 122.8468 | 85.5995 |
| Euhoplites lautus | -433.9713 | -410.6 | 105.9188 | 104.6453 |
| Euhoplites loricatus | -465.8373 | -436.8038 | 106.9309 | 105.8965 |
| Euhystrichoceras nicaisei | -103.4988 | -19.04 | 96.5874 | 94.5395 |
| Euomphaloceras septemseriatum | 108.6557 | 108.6557 | 93.88 | 93.15 |
| Favusella washitensis | -139.916 | 11.8706 | 114.1975 | 94.84 |
| Forbesiceras beaumontianum | -127.8239 | -128.8911 | 96.9524 | 96.2621 |
| Gartnerago nanum | -236.00 | -96.00 | 98.296 | 94.23 |
| Gartnerago obliquum | 107.1837 | 128.16 | 99.2536 | 64.743 |
| Gartnerago praeobliquum | -84.00 | *** | 96.5842 | *** |
| Gartnerago stenostauron | -139.916 | -128.3391 | 97.588 | 96.864 |
| Gartnerago theta | -108.00 | -80.00 | 96.8353 | 89.0722 |
| Graysonites azregensis | -127.82 | -127.2379 | 96.95* | 96.94* |
| Graysonites cobbani | -126.0738 | -122.2903 | 96.92* | 96.87* |
| Hamites virgulatus | -191.1179 | -165.9805 | 97.71* | 97.45* |
| Hayesites albiensis | -232.00 | *** | 118.6477 | 97.3296 |
| Hedbergella libyca | -236.00 | -212.00 | 100.6217 | 94.66 |
| Helenea chiastia | -224.00 | 102.7677 | 133.9695 | 92.7 |
| Helicolithus trabeculatus | -236.00 | -80.00 | 122.8533 | 65.505 |
| Hoplites dentatus | -472.9187 | -465.8373 | 107.5987 | 106.93 |
| Hyphoplites coelonotus | -242.942 | -165.00 | 97.4319 | 97.4319 |
| Hyphoplites falcatus | -132.0523 | -132.0523 | 96.2621 | 96.08 |
| Hyphoplites subfalcatus | -179.0295 | -179.0295 | 97.51* | 97.51* |
| Hyphoplites valbonnensis | -180.00 | -155.0337 | 97.5888 | 97.5888 |
| Hypoturrilites gravesianus | -128.8911 | -112.3309 | 96.9684 | 94.84 |
| Hypoturrilites schneegansi | -127.8239 | -98.9 | 96.9524 | 96.3536 |
| Hysteroceras orbignyi | -353.00 | -318.414 | 104.226 | 101.71 |
| Idohamites elegantulus | -248.88 | -247.2 | 98.31* | 98.29* |
| Inoceramus anglicus | -130.00 | -96.00 | 104.1574 | 96.7098 |
| Inoceramus concentricus | -435.00 | -375.0571 | 107.5807 | 101.71 |
| Inoceramus crippsi | -97.00 | -38.00 | 96.7471 | 94.2 |
| Lechites gaudini | -236.00 | -151.2174 | 98.6353 | 96.9133 |
| Lechites moreti | -276.00 | -276.00 | 98.59* | 98.59* |
| Lepthoplites cantabrigiensis | -194.7543 | -194.7543 | 97.75* | 97.75* |
| Lepthoplites falcoides | -248.88 | -180.3551 | 98.58* | 98.31* |
| Lepthoplites pseudoplanus | -248.88 | -234.88 | 98.31* | 98.16* |
| Lithraphidites acutus | -90.8027 | 101.6637 | 96.3969 | 92.7 |
| Lithraphidites alatus | -139.916 | -105.5228 | 98.296 | 93.2 |
| Lithraphidites carniolensis | -236.00 | -80.00 | 134.1294 | 65.4901 |
| Lithraphidites pseudoquadratus | -236.00 | 21.0707 | 98.1749 | 93.87 |
| Lyelliceras lyelli | -475.7512 | -475.7512 | 107.84 | 107.5987 |
| Manivitella pemmatoidea | -236.00 | -80.00 | 133.9695 | 65.2514 |
| Mantelliceras dixoni | -41.4901 | -41.49 | 95.6585 | 95.4 |
| Mantelliceras mantelli | -132.00 | -38.00 | 97.0656 | 95.65 |
| Mantelliceras saxbii | -127.8239 | -54.0022 | 96.9643 | 95.5 |
| Mariella bergeri | -280.00 | -129.6378 | 98.6353 | 96.8887 |
| Mariella cenomanensis | -127.8239 | -112.3952 | 96.9524 | 95.7203 |
| Mariella gresslyi | -276.00 | -276.00 | 98.59* | 98.59* |
| Markalius circumradiatus | -228.00 | -80.00 | 134.0985 | 82.2933 |
| Marker bed Breistroffer | -235.00 | -224.00 | 98.1644 | 98.0493 |
| Mojsisovicsia subdelaruei | -436.8038 | -436.8038 | 105.8965 | 105.8965 |
| Mortoniceras alstonensis | -248.88 | -247.2 | 98.31* | 98.29* |
| Mortoniceras fallax | -248.88 | -180.00 | 98.31* | 97.58* |
| Mortoniceras inflatum | -321.1158 | -241.59 | 103.0425 | 99.1149 |
| Mortoniceras perinflatum | -280.00 | -226.00 | 98.6353 | 98.0702 |
| Mortoniceras pricei | -342.7298 | -318.414 | 104.1574 | 102.1977 |
| Neolobites vibrayeanus | 101.6637 | 102.7677 | 93.7514 | 93.12 |
| Neophlycticeras blancheti | -248.88 | -180.00 | 98.31* | 97.58* |
| Neostlingoceras carcitanense | -132.00 | -38.00 | 97.0656 | 96.1028 |
| Ostlingoceras puzosianum | -236.00 | -129.6378 | 98.17* | 96.97* |
| Oxytropidoceras roissyanum | -472.9187 | -472.9187 | 108.0147 | 107.58 |
| Percivalia hauxtonensis | -144.00 | -144.00 | 97.2121 | 93.47 |
| Placozygus fibuliformis | -230.00 | -80.00 | 98.1121 | 65.505 |
| Planomalina buxtorfi | -236.00 | -119.00 | 100.606 | 96.0093 |
| Planomalina praebuxtorfi | -250.0185 | -216.0556 | 100.7341 | 97.9467 |
| Pleurohoplites renauxianus | -236.8768 | -180.3551 | 98.18* | 97.60* |
| Pleurohoplites subvarians | -180.0123 | -180.0123 | 97.59* | 97.59* |
| Praeglobotruncana delrioensis | -222.9565 | 101.6637 | 108.0343 | 92.00 |
| Praeglobotruncana stephani | -235.7826 | 118.2239 | 100.509 | 90.41 |
| Prediscosphaera columnata | -236.00 | -80.00 | 122.8533 | 90.7 |
| Prediscosphaera cretacea | -84.00 | -80.00 | 119.3508 | 64.523 |
| Prediscosphaera spinosa | -220.00 | -80.00 | 121.9656 | 64.4953 |
| Pseudaspidoceras flexuosum | 109.7598 | 109.7598 | 93.044 | 93.00 |
| Quadrum gartneri | 107.1837 | 128.16 | 93.5451 | 68.3833 |
| Rhagodiscus achlyostaurion | -236.00 | -80.00 | 124.6741 | 92.00 |
| Rhagodiscus angustus | -236.00 | -80.00 | 123.278 | 65.505 |
| Rhagodiscus asper | -236.00 | -80.00 | 128.6386 | 92.95 |
| Rhagodiscus splendens | -228.00 | -80.00 | 123.278 | 65.551 |
| Rotalipora appenninica | -265.787 | 23.6467 | 100.5159 | 94.48 |
| Rotalipora brotzeni | -136.00 | -119.00 | 97.0842 | 94.62 |
| Rotalipora cushmani | 33.9508 | 101.6637 | 96.17 | 93.07 |
| Rotalipora deeckei | 23.6467 | 37.9989 | 95.3597 | 93.2 |
| Rotalipora evoluta | -139.916 | -52.1622 | 97.588 | 95.05 |
| Rotalipora gandolfi | -222.9565 | 88.7835 | 98.9078 | 93.52 |
| Rotalipora globotruncanoides | -136.00 | 101.6637 | 97.13 | 93.15 |
| Rotalipora greenhornensis | -100.0028 | 99.8236 | 97.0842 | 93.07 |
| Rotalipora montsalvensis | -59.5223 | 101.6637 | 97.384 | 93.15 |
| Rotalipora reicheli | -1.0096 | 32.1108 | 96.706 | 94.42 |
| Rotelapillus crenulatus | -236.00 | -80.00 | 124.612 | 65.597 |
| Rotelapillus laffittei | -224.00 | -80.00 | 134.0402 | 66.407 |
| Scaphites simplex | -172.3889 | *** | 97.51* | *** |
| Schloenbachia varians | -119.9188 | -38.00 | 97.0648 | 95.12 |
| Sciponoceras roto | -132.00 | -50.00 | 97.0656 | 96.2284 |
| Sharpeiceras laticlavicum | -76.0825 | -42.2261 | 96.8389 | 95.45 |
| Sharpeiceras schlueteri | -127.8239 | -103.4988 | 96.9524 | 96.4478 |
| Staurolithites glaber | -232.00 | -112.00 | 98.133 | 96.8772 |
| Stoliczkaia africana | -222.9565 | -131.2738 | 98.04* | 97.00* |
| Stoliczkaia clavigera | -280.00 | -132.00 | 98.6353 | 97.0656 |
| Stoliczkaia dispar | -280.00 | -280.00 | 98.6353 | 97.116 |
| Stoverius achylosus | -172.00 | -80.00 | 123.3079 | 97.116 |
| Tegumentum stradneri | -230.00 | -80.00 | 129.3543 | 83.3875 |
| Tetrapodorhabdus coptensis | -230.00 | -80.00 | 128.4545 | 93.05 |
| Ticinella praeticinensis | -367.6759 | -325.2222 | 110.9318 | 97.4606 |
| Ticinella primula | -393.1481 | -270.6389 | 111.614 | 97.3667 |
| Ticinella subticinensis | -367.6759 | -232.00 | 103.1399 | 97.82 |
| Ticinella ticinensis | -325.2222 | -140.00 | 101.8279 | 96.8779 |
| Tranolithus gabalus | -230.00 | -80.00 | 123.9877 | 83.5584 |
| Tranolithus minimus | -220.00 | -132.00 | 98.0074 | 65.505 |
| Tranolithus orionatus | -230.00 | -80.00 | 110.9137 | 69.1743 |
| Turrilites acutus | 26.5908 | 71.6713 | 94.64 | 93.98 |
| Turrilites costatus | -23.8259 | 26.5908 | 95.3943 | 94.14 |
| Turrilites scheuchzerianus | -14.8097 | -0.6416 | 96.1343 | 94.07 |
| Turrilitoides hugardianus | -276.00 | -260.00 | 98.426 | 98.426 |
Table 2: Biostratigraphic ranges of Albian Stage ammonites, planktic foraminifera and nannofossils zones in 'Vraconnian' sections in metric units in the Mont Risou section (VRACCS.1) and numerical ages in the global MIDK45CS.1 database; ages with asterisk are interpolated by plotting the 'Vraconnian' database, VRACCS.1 to the MIDK45CS.1 database.
Integration of key planktic foraminifera and calcareous nannofossils (Fig. 6 ![]() ) based on the limited dataset of
(2008) shows that R. appenninica is below the base of Mortoniceras fallax, and the first occurrence of E. turriseiffelii is slightly
above it. In the larger MIDK45 dataset the FO of E. turriseiffelii is projected at 101.86 Ma and R. appenninica at 100.37 Ma, both of which are significantly older than the FO of M. fallax at 98.26 Ma. So neither pelagic species is useful as a proxy guide to the base of a 'Vraconnian
Stage'.
) based on the limited dataset of
(2008) shows that R. appenninica is below the base of Mortoniceras fallax, and the first occurrence of E. turriseiffelii is slightly
above it. In the larger MIDK45 dataset the FO of E. turriseiffelii is projected at 101.86 Ma and R. appenninica at 100.37 Ma, both of which are significantly older than the FO of M. fallax at 98.26 Ma. So neither pelagic species is useful as a proxy guide to the base of a 'Vraconnian
Stage'.
Five inoceramid species are recorded in the sections at Mont Risou and at Folkestone
(Fig. 6 ![]() ). Actinoceramus concentricus, Actinoceramus sulcatus, and Actinoceramus subsulcatus characterize the lower part of the Upper Albian and
Inoceramus anglicus and Inoceramus crippsi characterize the Lower Cenomanian.
). Actinoceramus concentricus, Actinoceramus sulcatus, and Actinoceramus subsulcatus characterize the lower part of the Upper Albian and
Inoceramus anglicus and Inoceramus crippsi characterize the Lower Cenomanian.
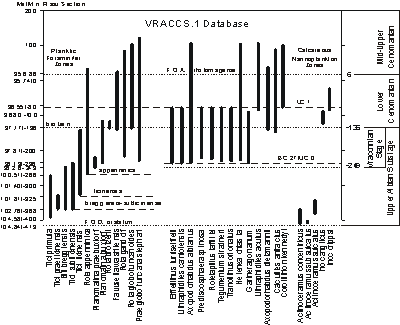
Click on thumbnail to enlarge the image.
Figure 6: Chronostratigraphic range chart of planktic foraminifera, calcareous nannofossils, and inoceramids in 'Vraconnian' composited database of key 'Vraconnian' sections. Vertical axis is meters in the Mont Risou section. Ages in Ma are interpolated by graphic correlation of the VRACCS.1 database with the MIDK45CS.1 database. Highest part of composite section not converted to ages. Foraminifer zones are defined by FO of named taxa. Nannofossil zones are defined by FO of E. turriseiffelii and C. kennedyi respectively. The FO of Lithraphidites acutus is in the middle part of the range of the Lower Cenomanian ammonite Mantelliceras mantelli in the Kalaat Senan section, Tunisia and 103.5 m above the base of Rotalipora globotruncanoides (MIDK.10 section; et alii, 1994).
The Albian stage is divided into seven zones and twenty-five subzones. The ranges of key fossils that define these zones can be calibrated to numerical Ma ages by graphic plots to sections bearing dated bentonites and geochemical events. This process measures the durations of the zones as proposed by and (2008). This database is anchored to bentonites in the U.S. Western Interior dated by (1993) and projected to the age of Magnetochron M0 at the base of the Aptian. As new radiometric ages are accrued this database can be tested and adjusted to accommodate new data.
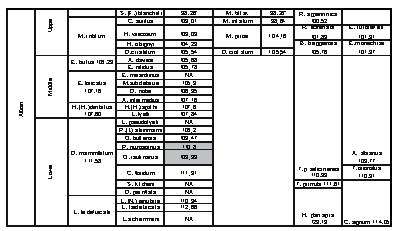
Click on thumbnail to enlarge the image.
Figure 7: Chronostratigraphic chart of the original Albian Stage. Ammonite zones from et alii (2004), et alii (2003), & (2000), et alii (1993), (1991), and (1984a, 1984b). Planktic foraminifera zones based on FOs from & (2002). Nannofossil zones based on FOs from et alii (1998) and et alii (1995). Numerical ages interpolated by graphic correlation of MIDK45 database ( et alii, 2000). NA indicates species not in the MIDK45 database.
The graphic correlation method provides data for chronostratigraphic correlation of the key 'Vraconnian' sections
(Fig. 8 ![]() ). The proposed 'Vraconnian Stage' correlates with the upper part of the Upper Albian Substage
(Fig. 4
). The proposed 'Vraconnian Stage' correlates with the upper part of the Upper Albian Substage
(Fig. 4 ![]() ). The cross section datum is the Albian/Cenomanian stage boundary as defined by the FO of R. globotruncanoides. used the FOs of Mortoniceras fallax and Neophlycticeras blancheti to define the base of the 'Vraconnian', which is approximated by the -250 m position in the VRACCS.1 database. This position correlates with the unconformity at the base of zones XII-XIII in the Folkestone section and with the transgressive facies between -800 and -781 m in the MAR 203 core at
Marcoule. The same time line also correlates within the lowstand limestone bundles in the Mont Risou and
Diégo core. The entire interval of the Bracquegnies Formation in the Strépy boatlift section and in the Harchies
N° 1 well (,
2008,
Fig. 17 ) correlates with Folkestone zones XII-XIII. The organic-rich interval in the Mont Risou section lies within the lower part of the transgressive interval equivalent to zone XII; it may represent maximum flooding. In the Mont Risou section a lower bundle of bioclastic-glauconitic limestone from about 50 to 80 m (,
2008,
Fig. 11 ) correlates with the lower condensed section between zones
VIII and IX at
Folkestone.
). The cross section datum is the Albian/Cenomanian stage boundary as defined by the FO of R. globotruncanoides. used the FOs of Mortoniceras fallax and Neophlycticeras blancheti to define the base of the 'Vraconnian', which is approximated by the -250 m position in the VRACCS.1 database. This position correlates with the unconformity at the base of zones XII-XIII in the Folkestone section and with the transgressive facies between -800 and -781 m in the MAR 203 core at
Marcoule. The same time line also correlates within the lowstand limestone bundles in the Mont Risou and
Diégo core. The entire interval of the Bracquegnies Formation in the Strépy boatlift section and in the Harchies
N° 1 well (,
2008,
Fig. 17 ) correlates with Folkestone zones XII-XIII. The organic-rich interval in the Mont Risou section lies within the lower part of the transgressive interval equivalent to zone XII; it may represent maximum flooding. In the Mont Risou section a lower bundle of bioclastic-glauconitic limestone from about 50 to 80 m (,
2008,
Fig. 11 ) correlates with the lower condensed section between zones
VIII and IX at
Folkestone.
The thickness of the 'Vraconnian Stage' varies greatly among the sections as noted by (, 2008). In the MAR 203 core at Marcoule the equivalent 'Vraconnian' interval is 317 m thick, eight times than thicker than at the Strépy section, more than twenty-three times thicker than at Folkestone, and nearly 160 times thicker than in the 'Vraconnian' reference section in Switzerland. The rates of sediment accumulation varied from 4.5 m/myr at Folkestone to 37.6 m/myr at Mont Risou, and 111.6 m/myr at Marcoule. The rate of accumulation is based on the compacted section and is not a sedimentation rate. These rates are based on the duration of about 3 myr for the 'Vraconnian' interval. This great range in rates is based on very different basin subsidence histories and tectonic conditions of each section.
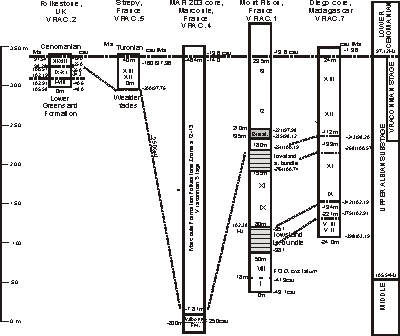
Click on thumbnail to enlarge the image.
Figure 8: Stratigraphic correlation of key 'Vraconnian' sections using base of Cenomanian Stage as datum. CSU values are meters in the Mont Risou section of et alii (1996, MIDK.24). Ages interpolated by graphic correlation of the VRACCS.1 database with the MIDK45CS.1 database. Age of lower part of Valbonne Formation below intra-formational unconformity at -800 m not projected because of the absence of fossils.
This criterion is essential to the definition of lithostratigraphic units such as formations (NACSN, 2005). However, it is not part of the definition of a stage, in which lithofacies change from basin to basin. The lithologies that comprise the 'Vraconnian' interval are quite different from section to section (, 2008), and thus do not make up a mappable lithostratigraphic unit.
Indeed the 'Vraconnian' interval records a third-order three myr depositional cycle of transgression and regression on a regional even global scale. This feature, however, defines sequence stratigraphic units, not stages (NASCN, 2005). The Upper Albian Stage records five such sequences of this scale ( et alii, 2003), but it would be impractical to divide the Albian into five stages.
This property is an essential feature of a stage concept. Three species of Mortoniceras and one species of Stoliczkaia comprise the 'Vraconnian Stage' according to
(2008) and &
(2008).
(2008) characterizes the 'Vraconnian' by the abrupt diversification of heteromorph ammonites of the
Turrilitidae, Hamitidae, Anisoceratidae, and Baculitidae in the condensed La Vraconne section, where they comprise 60% of the specimens. However, species of three of these families appear earlier in the Late Albian between 102.47 and 98.64 Ma
(Fig. 5 ![]() ) earlier than the FO of Mortoniceras fallax and Neophlycticeras blancheti.
) earlier than the FO of Mortoniceras fallax and Neophlycticeras blancheti.
The three heteromorph species restricted to the 'Vraconnian' in 's database
(2008) are primarily found in Western Europe. In the
Carpatho-Balkan region of Eastern Europe the 'Vraconnian' is represented by a condensed interval of glauconitic and phosphatic sandy limestone less than one meter thick; the only ammonite species of the zone is Stoliczkaia notha ( & ,
1996). One North American section, Dry Creek in northern California (,
2008,
Fig. 24 ), yields three ammonite species of the Dispar Zone, only one of which, Lechites gaudini, is found at the type section of the 'Vraconnian' in Switzerland ( & ,
1978). Few species characteristic of the 'Vraconnian' are found outside of Western Europe. The ammonite assemblage is not widespread in the Tethyan or Boreal Realms.
The proposed duration of the 'Vraconnian Stage' is 2-3 myr, which is equivalent to that of the Santonian Stage. However the durations of Cretaceous stages vary from 2.3 to 13 myr ( et alii, 2004) and duration is not a criterion for defining a stage.
Sections bearing the 'Vraconnian' fauna are locally thicker than the underlying part of the Albian. The thickness difference is highly variable from basin to basin and within basins. Such thickness differences are found between condensed sections and coeval basin margin and basin center sections of many zones.
The diverse ammonite assemblage in the uppermost part of the Upper Albian Substage comprises a distinctive zone in Western Europe that can be subdivided into three subzones. These subzones are recognized in many Tethyan and transitional Boreal sections in Europe. However, at the 'Vraconnian' type section in Switzerland Stoliczkaia dispar and Mortoniceras perinflatum are the only zonal named taxa present. The strata bearing the S. dispar Zone are bounded by unconformities in many sections and they are but one of five Upper Albian depositional sequences. The thickness of this sequence varies from condensed sections of two meters to expanded basinal sections more than 300 meters thick. The S. dispar Zone represents a time interval of about three myr, which is about the same duration as the briefest Cretaceous ages. This interval is very useful as a biostratigraphic unit and a third-order sequence stratigraphic unit. However this interval is not a practical chronostratigraphic unit such as a stage because its boundaries cannot be demonstrated to be synchronous and the interval is not isochronous, nor is it globally recognizable. Defining this interval as a stage equivalent to the Albian and Cenomanian stages would materially alter the concept of the Albian Stage by deleting its uppermost zone. The concept of a 'Vraconnian Stage' is not a practical subdivision of the Cretaceous System.
Michael and anonymous referees offered very useful and constructive suggestions that clarified this contribution.
F. (1992).- L'Albien du bassin anglo-parisien : ammonites, zonation phylétique, séquences.- Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Pau, vol. 16, n° 1, p. 187-233.
F. (2002).- Plaidoyer pour un étage Vraconnien entre l'Albien sensu stricto et le Cénomanien (système Crétacé).- Académie Royale de Belgique, Mémoire, Bruxelles, (Classe des Sciences), t. IV, 128 p.
F. (2008).- Support for a Vraconnian Stage between the Albian sensu stricto and the Cenomanian (Cretaceous System).- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2008/02 (CG2008_M02), 83 p.
F. & F. (2008).- Zonation by ammonites and foraminifers of the Vraconnian-Turonian interval: A comparison of the Boreal and Tethyan domains (NW Europe / Central Tunisia).- Carnets de Géologie / Notebooks on Geology, Memoir 2008/02 (CG2008_L02), 5 p.
T., J.M., M.B., P.F., J., F., F. & F. (1984).- Cretaceous stage boundaries. Proposals.- Bulletin of the Geological Society of Denmark, Copenhagen, vol. 33, p. 3-20.
P.R., D.C., J.A. & L.T. (1998).- Lower Cretaceous. In: P.R. (ed.), Calcareous nannofossil biostratigraphy.- Kluwer Academic Publishers, London, p. 86-130.
T.J. & R.M. (1995).- An integrated Cretaceous microfossil biostratigraphy. In: W.W., D.V., M.-P. & J. (eds.), Geochronology, time scales and global stratigraphic correlation.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication n° 54, p. 65-79.
M. (1947).- Sur les zones d'ammonites de l'Albien de France et d'Angleterre.- Travaux du Laboratoire de Géologie de l'Université de Grenoble, t. 26, p. 17-104.
J.L. & R.W. (1995).- Graphic correlation and composite standard databases as tools for the exploration biostratigrapher. In: K.O. & H.R. (eds.), Graphic correlation.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication n° 53, p. 23-43.
M. (1965).- Rapport sur l'étage Albien. In: Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoire du Bureau de Recherches Géologiques et Minières, Orléans, n° 34, p. 313-318.
G. & J.-L. (1988).- Découverte d'un nouveau gisement albien dans les environs de Drap (Alpes-Maritimes, France) et description d'une riche ammonitofaune d'âge Albien terminal.- Geobios, Villeurbanne, n° 21, fasc. 6, p. 749-771.
P. & J.-P. (1965).- Distribution zonale des ammonites dans l'Albien du bassin de Paris. In: Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoire du Bureau de Recherches Géologiques et Minières, Orléans, n° 34, p. 255-270.
A.S., W.J., J.A., M. & B.E. (1996).- The Late Albian to Early Cenomanian succession at Mont Risou near Rosans (Drôme, SE France): an integrated study (ammonites, inoceramids, planktonic foraminifera, nannofossils, oxygen and carbon isotopes).- Cretaceous Research, London, vol. 17, n° 5, p. 515-606.
J.M. (1991).- Ammonite scales for the Cretaceous system.- Cretaceous Research, London, vol. 12, n° 3, p. 259-291.
J.M. (2003).- Book review: "Plaidoyer pour un étage Vraconnien entre l'Albien sensu stricto et le Cénomanien (système Crétacé)" Francis 2002, Académie Royale de Belgique, Publication de la Classe des Sciences, 3rd series, 4, 128 pp., 9 pls; ISBN 2-8031-0186-6, Euro 21.- Cretaceous Research, London, vol. 24, n° 1, p. 95-96.
W.B., R.L., A.V., L.E., A.G. & D.G. (1990).- A geological time scale 1989.- Cambridge University Press, Cambridge, 263 p.
M.B. (2000).- Foraminifera, sequence stratigraphy and regional correlation; an example from the Uppermost Albian of Southern England.- Revue de Micropaléontologie, Paris, vol. 43, n° 1, p. 27-45.
M.B., F. & H. (1996).- The Albian stage and substage boundaries.- Institut royal des Sciences naturelles de Belgique, Bulletin Sciences de la Terre, Bruxelles, vol. 66, suppl., p. 45-56.
P.J. & M. (reporters), M.B., E., T.N., L., L., F., G., M., L., H., P.F., J., J.M., J.-P., S.Z. & Z. (1993).- Ammonites zonation for the Lower Cretaceous of the Mediterranean region; basis for the stratigraphic correlation within IGCP-Project 262.- Revista Española de Paleontologia, Madrid, vol. 8, n° 1, p. 117-120.
P.J. & P.F. (reporters), M.B., T.N., I., P., C., M., J., A., N., O. & Z. (2000).- Report on the 5th International Workshop of the Lower Cretaceous Cephalopod team (Vienna, 5 September 2000).- Cretaceous Research, London, vol. 21, n° 6, p. 857-860.
P.J. & S. (reporters), M.B., P., M., F., R., M., C., J., A., I., N., P.F., P., Z., J. & M.G.E. (2003).- Report on the 1st International Workshop of the IUGS Lower Cretaceous Ammonite Working Group, the ' Group' (Lyon, 11 July 2002).- Cretaceous Research, London, vol. 24, n° 1, p. 89-94 ; erratum, vol. 24, n° 6, p. 805.
W.J., A.S., P.R., M., R.J., D. & D.S. (2002).- Integrated stratigraphy across the Aptian-Albian boundary in the Marne Bleus, at Col de Pré-Guittard, Arnayon (Drôme), and at Tartonne (Alpes-de-Haute-Provence), France: a candidate Global Boundary Stratotype Section and Boundary Point for the base of the Albian Stage.- Cretaceous Research, London, vol. 21, n° 5, p. 591-720.
W.J., A.S., J.A. & M. (2004).- Definition of a Global Boundary Stratotype Section and Point (GSSP) for the base of the Cenomanian Stage, Mont Risou, Hautes-Alpes, France.- Episodes, Beijing, vol. 27, n° 1, p. 21-32.
, J. & R. (1996).- The Mid-Cretaceous Boreal/Tethyan biogeographical boundary in the Carpatho-Balkan area.- Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, Heft 77, p. 141-147.
A. de (1868).- Note sur l'étage de la gaize.- Bulletin de la Société géologique de France, Paris, (2ème série), t. XXV, p. 868-871.
C., P. & M. (1965).- Documents paléontologiques et stratigraphiques sur l'Albien de l'Aube. In: Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoire du Bureau de Recherches Géologiques et Minières, Orléans, n° 34, p. 237-253.
F. & P. (1980).- Albien. In: C. & J. (eds.), Les étages français et leurs stratotypes.- Mémoire du Bureau de Recherches Géologiques et Minières, Orléans, n° 109, p. 126-129.
F.X. (1977).- The graphic correlation method in biostratigraphy. In: E.G. & J.E. (eds.), Concepts and methods of biostratigraphy.- Dowden, Hutchinson & Ross, Inc., Stroudsburg, Pennsylvania, p. 165-186.
NACSN - North American Commission on Stratigraphic Nomenclature (2005).- North American stratigraphic code.- American Association of Petroleum Geologists Bulletin, Tulsa, vol. 89, n° 11, p. 1547-1591.
F.E., D.G. Jr, R.W., J.M., M.J. & J. (2007).- Re-evaluation of the Albian-Cenomanian boundary in the U.S. Western Interior based on dinoflagellate cysts.- Review of Palaeobotany and Palynology, Amsterdam, vol. 144, n° 1-2, p. 77-97.
J.D. (1993).- A Cretaceous time scale.- Geological Association of Canada, Special Paper, Toronto, n° 39, p. 379-396.
J.G., F.P. & F.M. (2004).- The Cretaceous Period. In: F., J.G. & A. (eds.), A geologic time scale.- Cambridge University Press, Cambridge, p. 344-383.
A. d' (1840-42).- Paléontologie française. Terrains crétacés. t. I, Céphalopodes.- Masson, Paris, 662 p.
H.G. (1971).- Middle Albian stratigraphy in the Anglo-Paris basin.- Bulletin of the British Museum (Natural History), London, Supplement 8, 164 p.
H.G. (1984a).- The Albian stage: European province chronology and ammonite zonation.- Cretaceous Research, London, vol. 5, n° 4, p. 329-344.
H.G. (1984b).- Albian stage and substage boundaries.- Bulletin of the Geological Society of Denmark, Copenhagen, vol. 33, p. 183-189.
I. & W.V. (2002).- Practical manual of Cretaceous planktonic foraminifera.- Dipartmento di Scienze della Terra, University of Perugia (Italy), International School on Planktonic Foraminifera, I° course: Cretaceous, 462 p.
C.E. de (1868).- On the Albian or Gault of Folkestone.- Geological Magazine, London, vol. 5, p. 163-171.
P.F., D., F.C., J.M., W.J., J.W., C.J. & B.C. (1978). A correlation of Cretaceous rocks in the British Isles.- Geological Society London, Special Report, n° 9, 70 p.
P.F & P.J. (reporters), M.B, E., M., S.R.A., J., E., H.G., P., M.R.A., M. & Z. (1999).- Report on the 4th International Workshop of the Lower Cretaceous Cephalopod Team (IGPC-Project 362).- Scripta Geologica, Leiden, Special Issue 3, p. 313.
E. (1868).- Notices géologiques et paléontologiques sur les Alpes Vaudoises et les régions environnantes. V. Complément de la faune de Cheville.- Bulletin de la Société Vaudoise des Sciences Naturelles, Lausanne, t. 9, n° 58, p. 389-482.
O. & P. (1978).- Aptian to Maastrichtian in the Swiss Jura Mountains.- Eclogae geologicae Helvetiae, Basel, vol. 71, p. 1-18.
F., M., F., C., J., J.M., D. & S. (1994).- Le Cénomanien de la région de Kalaat Senan (Tunisie centrale) : Litho-biostratigraphie et interprétation séquentielle.- Revue de Paléobiologie, Genève, vol. 12, n° 2, p. 351-505.
F., F., J.M. & D. (2007).- Les bioévénements de la limite Albien (Vraconnien)-Cénomanien aux marges nord et sud de la Téthys (S.E. de la France et Tunisie centrale). In: L.G., S. & D. (eds.), Relations entre les marges septentrionale et méridionale de la Téthys au Crétacé [Relations between the northern and southern margins of the Tethys ocean during the Cretaceous period].- Carnets de Géologie / Notebooks on Geology, Mémoire 2007/02, Résumé 01 (CG2007_M02/01), p. 3-15.
R.W. (2009, in press).- Chronostratigraphic database for Upper Cretaceous oceanic Red Beds (CORBs). In: X., C., L., R. & M. (eds), Cretaceous oceanic Red Beds: A clue to ocean/climate change.- SEPM (Society for Sedimentary Geology), Special Publication, n° 91.
R.W., W., B. & S.A. (2000).- Are Mid-Cretaceous eustatic events recorded in Middle East carbonate platforms?. In: A.S. & R.W. (eds.), Middle East Models of Jurassic/Cretaceous Carbonate Systems.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication n° 69, p. 77-88.
R.W., D.G., R.W., B.L. & F.E. (2003).- Integrated Albian-Lower Cenomanian chronostratigraphy standard, Trinity River section, Texas. In: R.W. (ed.), Cretaceous stratigraphy and paleoecology, Texas and Mexico: Perkins Memorial Volume.- GCSSEPM Foundation, Houston, Special Publications in Geology, n° 1, CD book, p. 277-334.
L.F. (1923).- On the ammonite horizons of the Gault and contiguous deposits.- Geological Survey U.K., London, Summary Progress for 1922, p. 139-149.
L.F. (1941).- A monograph of the Ammonoidea of the Gault. Part XIV.- Palaeontological Society, London, Monograph, p. 609-668.