![]()
[Abridged English version]
[I - Introduction]
[II La carrière de Larcan] [III L'hadrosaure de Larcan]
[IV Le niveau à iridium : argument de corrélation ou argument de datation ?]
[Conclusion] et ...
[Références bibliographiques]
Laboratoire Mécanismes et Transfert en Géologie (LMTG), UMR 5563 CNRS, Université Paul Sabatier, 39 allées Jules-Guesde, 31400 Toulouse (France)
Association Paléontologique du Sud-Ouest (APSO), 2 boulevard d'Arcole, 31000 Toulouse (France)
1 chemin de Castillon, 31140 Pechbonnieu (France)
Manuscrit en ligne depuis le 31 Mai 2010
![]()
La découverte de deux fragments d'hadrosaure (maxillaire et carré) dans des sédiments marins de la fin du Crétacé (Maastrichtien terminal) est relatée. Si ce type de taphocoenose n'est pas exceptionnel pour des organismes continentaux, sa situation à environ 1 m sous la zone à iridium de la limite K/T méritait d'être soulignée. Cette présentation permet en outre de faire le point sur des données bio et chronostratigraphiques récemment formulées dans cette région.
Dinosaure ; hadrosaure ; Maastrichtien ; iridium ; Pyrénées ; France.
M., Y. & D. (2010).- Restes d'hadrosaure dans le Crétacé terminal marin de Larcan (Petites Pyrénées, Haute-Garonne, France).- Carnets de Géologie / Notebooks on Geology, Brest, Note brève / Letter 2010/02 (CG2010_L02)
Hadrosaur remains in the Late Cretaceous marine sediments of Larcan (Petites Pyrenees, Haute-Garonne, France).- The discovery of two hadrosaur fragments (maxilla and quadrate) in marine sediments of the end of the Cretaceous (uppermost Maastrichtian) is reported. Although this type of taphocoenosis is not exceptional for continental organisms, its situation at approximately 1 m under the iridium zone of the K/T boundary deserves to be emphasized.
Dinosaur; hadrosaur; Maastrichtian; iridium; Pyrenees; France.
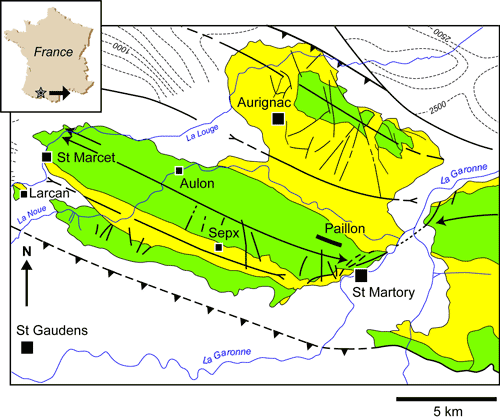
Located south of Saint-Marcet, on the right bank of the Save River, the Larcan outcrop is in the westernmost portion of the Latoue-Sepx subpyrenean syncline (Fig. 1
![]() ). Beneath a Miocene unconformity, the strata are tilted up to the vertical. The stratigraphic succession, largely exposed in a quarry, is composed of the following units
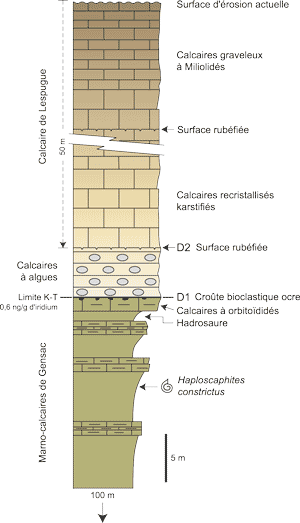
(Fig. 2
). Beneath a Miocene unconformity, the strata are tilted up to the vertical. The stratigraphic succession, largely exposed in a quarry, is composed of the following units
(Fig. 2 ![]() ). The Upper Cretaceous (approximately 160 m) is represented by two formations referred to as Maastrichtian. They are the blue marls of Saint-Loup that are underlain by the yellow marly limestones of Gensac. A very sharply defined discontinuity (D1), marks the upper limit of these deposits and is accompanied by a small concentration of iridium (0.6 ng/g).
). The Upper Cretaceous (approximately 160 m) is represented by two formations referred to as Maastrichtian. They are the blue marls of Saint-Loup that are underlain by the yellow marly limestones of Gensac. A very sharply defined discontinuity (D1), marks the upper limit of these deposits and is accompanied by a small concentration of iridium (0.6 ng/g).
Above the discontinuity that marks the Cretaceous-Tertiary boundary, the Paleocene (50 m visible) is represented by two formations: the lower one, the algal limestones of Larcan (6 m), is terminated by another discontinuity (D2) which is overlain by the Miliolid limestones of Lespugue. The Larcan quarry exploits only the algal limestones. To get to them exploitation has exposed locally the upper part of the yellow marly limestones of Gensac thus providing conditions favourable to paleontologic investigations. A marine fauna and microfauna (sea-urchins, bivalves, gastropods, cephalopods, bryozoans, foraminifers,
) have been collected from the higher levels of the Maastrichtian, and recently two hadrosaurid dinosaur remains were discovered only 1 m below the discontinuity marking the K-T boundary in this area
(Pl. 1
![]() ).
).
The two cranial elements of a hadrosaur discovered at Larcan are from the area of the quarry currently exploited. The bones were discovered in the uppermost level of the Gensac marly limestones superposed one upon the other
(Fig. 3 ![]() ). The matrix
there is a dark calcareous siltstone in which many benthic and planktonic micro-organisms are present, among them Hellenocyclina beotica, Praestorsella roestae, Siderolites calcitrapoides, calcispheres,
The D1 discontinuity with the iridium concentration of 0.6 ng/g is at most 1 m above the hadrosaur specimens. The material is now deposited in the private collection of Dominique
.
). The matrix
there is a dark calcareous siltstone in which many benthic and planktonic micro-organisms are present, among them Hellenocyclina beotica, Praestorsella roestae, Siderolites calcitrapoides, calcispheres,
The D1 discontinuity with the iridium concentration of 0.6 ng/g is at most 1 m above the hadrosaur specimens. The material is now deposited in the private collection of Dominique
.
The maxilla measuring 115 mm in length and 42 mm in height; it is a right maxilla probably that of a juvenile hadrosaur. Because of the matrix its description is possible only from the lateral aspect. The teeth are preserved except at the anterior extremity. The presence of the quadrate (probably of the same individual), sited in the central portion of the maxilla, makes it impossible to determine the exact number of tooth alveoli. The teeth have a prominent true median ridge developed on the enamelled labial surface. It is not possible to determine whether or not the crown has marginal denticles and whether or not secondary ridges are present on both sides of the median ridge.
The ectopterygoid shelf is wide and extends to about half the length of the maxilla; it is bordered by a strong ectopterygoid ridge which ends in a prominent bulge overhung by the jugal articulation. At its base, the bulge shows two large aligned neurovascular foramina.
The dorsal process of the maxilla, slightly truncated in our specimen, is broad and its maximum height appears to be located at the mid-length of the bone (the general shape of the maxilla is triangular and symmetrical).
The anterior part of the dorsal process is developed into a very elevated rounded lateral crest.
Dorsally, the anterior part of the maxilla shows a broad plate which tapers gradually and forms the antero-ventral process.
The maxilla resembles the specimens MDE-Ma3-15 and MDE-Ma3-16 from the Tricouté 3 site, located southwest of the town of Marignac-Laspeyres (Aurignac anticline) where the Auzas Marls Formation has yielded many hadrosaurid remains (, 2003). The specimens of Tricouté 3 were provisionally referred to the genus Pararhabdodon from the Upper Cretaceous of Spain ( et alii, 1993, 1999; & , 2009).
In the current state of our knowledge the specimens of Larcan and Tricouté 3 are the remains of a new hadrosaurid dinosaur different from Pararhabdodon (pers. comm. P. ). Indeed, the great development of the lateral crest in front of the dorsal process has no equivalent in the Hadrosauridae described to date, with the exception of Aralosaurus tuberiferus ("curved lateral crest" of et alii, 2004, Fig. 4).
Another lambeosaurine hadrosaurid, Arenysaurus ardevoli, from the upper Maastrichtian of Arén (Huesca, Spain) has also been described recently ( et alii, 2009). However its maxilla is not known, which makes comparison with our specimen impossible.
The quadrate is probably strongly eroded. It is 80 mm long and its general shape suggests that of a hadrosaurid. The preservation of the specimen does not allow a detailed description. In any case, its size and its association with the maxilla just described makes the attribution of these remains to the same juvenile hadrosaurid dinosaur almost unquestionable.
The discovery of cranial hadrosaurid remains in the upper Maastrichtian marine levels just below the iridium level found at the K/T boundary is significant; among other things the discovery made it possible to support more rigorously the idea that the laguno-continental Auzas marls and the marine marly limestones of
Gensac are, at least in part, equivalent in time
(Fig. 4 ![]() ). It also provides evidence of the existence of dinosaurs at the highest levels of the Upper Maastrichtian on the northern flank of the Pyrenees.
). It also provides evidence of the existence of dinosaurs at the highest levels of the Upper Maastrichtian on the northern flank of the Pyrenees.
Au sud de Saint-Marcet, sur la rive droite de La Save, l'affleurement de Larcan s'intègre au flanc sud du synclinal sous-pyrénéen de Latoue-Sepx, dont il constitue le pointement le plus occidental (Fig. 1
![]() ). Sous une couverture miocène discordante, les couches sont relevées à la verticale ; le Crétacé supérieur (160 m environ) y est représenté par deux formations connues régionalement pour représenter les dépôts du Maastrichtien ; ce sont les Marnes bleues de Saint-Loup et les Marno-calcaires jaunes de Gensac. Une très nette discontinuité, présentant une faible concentration en iridium, couronne ces dépôts ; au-dessus le Paléocène (50 m visible) est représenté par deux autres
formations : d'abord, les Calcaires à algues de Larcan
(6 m), puis les Calcaires de Lespugue à miliolidés. La carrière de Larcan recherche exclusivement le faciès des Calcaires à algues. Pour les atteindre, l'exploitation a dégagé par place la partie supérieure des Marno-calcaires jaunes de Gensac et crée ainsi des surfaces d'affleurements favorables à la recherche d'indices paléontologiques. Alors que généralement ce sont des faunes et microfaunes marines (oursins, bivalves, gastéropodes, céphalopodes, bryozoaires, foraminifères, ...) qui sont extraites de ces niveaux supérieurs du Maastrichtien, deux fragments d'un dinosaure
hadrosaure viennent d'y être découverts à moins de
1 m sous la discontinuité de la limite K/T. La présentation de cette trouvaille est mise à profit pour dénoncer quelques incohérences qui accompagnent des travaux stratigraphiques régionaux récents.
). Sous une couverture miocène discordante, les couches sont relevées à la verticale ; le Crétacé supérieur (160 m environ) y est représenté par deux formations connues régionalement pour représenter les dépôts du Maastrichtien ; ce sont les Marnes bleues de Saint-Loup et les Marno-calcaires jaunes de Gensac. Une très nette discontinuité, présentant une faible concentration en iridium, couronne ces dépôts ; au-dessus le Paléocène (50 m visible) est représenté par deux autres
formations : d'abord, les Calcaires à algues de Larcan
(6 m), puis les Calcaires de Lespugue à miliolidés. La carrière de Larcan recherche exclusivement le faciès des Calcaires à algues. Pour les atteindre, l'exploitation a dégagé par place la partie supérieure des Marno-calcaires jaunes de Gensac et crée ainsi des surfaces d'affleurements favorables à la recherche d'indices paléontologiques. Alors que généralement ce sont des faunes et microfaunes marines (oursins, bivalves, gastéropodes, céphalopodes, bryozoaires, foraminifères, ...) qui sont extraites de ces niveaux supérieurs du Maastrichtien, deux fragments d'un dinosaure
hadrosaure viennent d'y être découverts à moins de
1 m sous la discontinuité de la limite K/T. La présentation de cette trouvaille est mise à profit pour dénoncer quelques incohérences qui accompagnent des travaux stratigraphiques régionaux récents.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Situation géographique et géologique de la localité de Larcan au SW des Petites Pyrénées de la Haute-Garonne.
Figure 1: Geographical and geological location of the Larcan locality to the SW of the Petite Pyrenees (Haute-Garonne area).
Dans la carrière de Larcan
(Fig. 2 ![]() ) se succèdent, du sud vers le nord, le sommet des Marno-calcaires de Gensac (15 m environ) puis les Calcaires à algues de Larcan
(6 m). L'exploitation en tranchée
(Pl. 1
) se succèdent, du sud vers le nord, le sommet des Marno-calcaires de Gensac (15 m environ) puis les Calcaires à algues de Larcan
(6 m). L'exploitation en tranchée
(Pl. 1
![]() , fig. 3) de cette dernière unité lithologique est facilitée par la présence de deux importantes discontinuités sédimentaires situées, respectivement, au toit des Marno-calcaires de Gensac et au toit des Calcaires à algues de Larcan ; les Calcaires de Lespugue qui font suite ne sont de ce fait pas (ou peu) affectés par le front de taille. Par contre, la partie supérieure des Marno-calcaires de Gensac est fréquemment dégagée pour des raisons d'accessibilité.
, fig. 3) de cette dernière unité lithologique est facilitée par la présence de deux importantes discontinuités sédimentaires situées, respectivement, au toit des Marno-calcaires de Gensac et au toit des Calcaires à algues de Larcan ; les Calcaires de Lespugue qui font suite ne sont de ce fait pas (ou peu) affectés par le front de taille. Par contre, la partie supérieure des Marno-calcaires de Gensac est fréquemment dégagée pour des raisons d'accessibilité.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 :
La série stratigraphique de la carrière de Larcan au passage Crétacé Tertiaire.
Figure 2: The stratigraphic sequence of the Larcan quarry at the Cretaceous - Tertiary boundary.
Les recherches effectuées antérieurement sur ce site (, 1985), dans la partie ancienne de l'exploitation, toujours visible dans un niveau inférieur de la colline de Lapeyre, ont précisé l'organisation des unités lithologiques en présence. Par rapport à cette étude, la position du passage Crétacé-Tertiaire a été modifiée. Dans l'état actuel de nos connaissances, la succession, du sud vers le nord, est chronologiquement la suivante :
Le sommet des Marno-calcaires de Gensac s'achève par trois séquences métriques grano et strato-croissantes (marnes argileuses grises silts calcareux bioturbés calcaires bioclastiques) qui rendent compte de l'évolution régressive des dépôts. Parmi les marqueurs indiscutables du Maastrichtien supérieur nous citerons (,
1985), dans l'antépénultième séquence, Hoploscaphites constrictus (détermination W.J. ) ; dans les calcaires beiges sommitaux, l'association : Lepidorbitoides socialis, Orbitoides apiculata, Clypeorbis mamillata, Hellenocyclina beotica, Praestorssella roestae, Siderolites calcitrapoides, S. denticulatus, ... Le toit de ce dernier banc est couronné par une surface d'arrêt de sédimentation (D1,
Fig. 2 ![]() ) particulièrement bien marquée ; cette surface est affectée par une intense corrosion qui pénètre souvent sur plusieurs centimètres les calcaires qui la supportent.
) particulièrement bien marquée ; cette surface est affectée par une intense corrosion qui pénètre souvent sur plusieurs centimètres les calcaires qui la supportent.
Une croûte ocre bioclastique qui remplit les poches de corrosions (Pl. 1
![]() , fig. 4) ; il est possible d'y observer des débris de foraminifères du Maastrichtien tels, Hellenocyclina beotica, Praestorssella roestae, Orbitoides sp., ... associés à d'abondants débris d'échinodermes. C'est dans cette croûte ocre qu'a été mesurée une anomalie en iridium de 0,6 ng/g ( et alii,
1998).
, fig. 4) ; il est possible d'y observer des débris de foraminifères du Maastrichtien tels, Hellenocyclina beotica, Praestorssella roestae, Orbitoides sp., ... associés à d'abondants débris d'échinodermes. C'est dans cette croûte ocre qu'a été mesurée une anomalie en iridium de 0,6 ng/g ( et alii,
1998).
Les Calcaires à algues de Larcan : aucune organisation sédimentaire nette ne se dégage de cette formation qui apparaît comme constituée d'un empilement de bancs décimétrique à métriques de calcaires de teinte beige clair. Une attribution au Maastrichtien terminal (, 1985 ; , 1991) a été remise en cause par et alii (1997) suite à la reconnaissance d'une microfaune planctonique composée de "Globanomalina aff. compressa et Subbotina triloculinoides". Aucune illustration n'accompagne ces déterminations effectuées sur des sections en lames minces, et aucune microfaune benthique n'est citée. Cette attribution au Paléocène se fonde donc sur une aff. et sur un taxon connu pour franchir la limite K/T !
Outre les microfaunes benthiques apparemment remaniées de niveaux inférieurs (Hellenocyclina beotica, Planorbulina cretae), le contenu le plus visible de ces calcaires, qui leur confère un intérêt ornemental et justifie leur exploitation, est l'abondance des thalles de Lithothamniées associés à Gypsina cf. o'gormani (Solenomeris auct.). Une nouvelle étude micropaléontologique (L. et Y. , inédit) y fait état seulement d'une microfaune benthique constituée de "Planorbulina" cretae, "Mississipina cf. binkhorsti", Gyroidina sp., Anomalina sp., Thalmannita sp. et Coscinophragma cribosum. Pour compléter l'inventaire paléontologique de cette formation, citons la présence dans son 1/3 inférieur d'une faunule de brachiopodes attribués à Erymnaria rauschi (détermination D. ) et de bryozoaires cyclostomes. En définitive, si le cachet de cette faune plaide en faveur d'un âge Paléocène des Calcaires de Larcan, aucun des éléments cités ne semble déterminant ; seule leur situation, au-dessus du niveau à iridium conforte cette attribution.
La discontinuité sommitale D2 : elle correspond à une surface très nette et parallèle à la discontinuité D1 ; elle se marque par place par la présence d'un "horizon brèchique à éléments anguleux de calcaires silteux laminés, de calcaires finement cristallisés et de microconglomérats à éléments quartzeux" (, 1985). Il semble en outre qu'elle ait servi de drain aux fluides responsables de la recristallisation des calcaires sus-jacents. Curieusement, les Calcaires de Larcan ne sont pas affectés par cette recristallisation.
Les Calcaires de Lespugue : ils débutent par une masse fortement cristallisée
(30 m) au sein de laquelle il n'a été possible de reconnaître, dans des microsparites et des dolosparites, que des fantômes de mollusques, de polypiers et d'algues dasycladales. Au-dessus et sur
20 m environ, les calcaires deviennent graveleux et renferment une riche microfaune benthique composée surtout de Miliolidés, d'Ataxophragmiidés et de Rotalidés. Une surface rubéfiée s'observe dans ce dernier ensemble
(Fig. 2 ![]() ). Mentionnons enfin qu'un important réseau karstique, développé au Pléistocène, affecte les calcaires cristallisés de la base de la formation.
). Mentionnons enfin qu'un important réseau karstique, développé au Pléistocène, affecte les calcaires cristallisés de la base de la formation.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 1 : La carrière de
Larcan.
Fig. 1. La discontinuité D1 (limite K/T) au toit des Marno-calcaires de Gensac, dans la partie ancienne de l'exploitation.
Fig. 2. Situation de l'hadrosaure de Larcan (F) par rapport à la discontinuité D1.
Fig. 3. L'excavation entre les discontinuités D1 (limite K/T) et D2 (intra-paléogène) matérialise la position des Calcaire à algues de Larcan.
Fig. 4. Détail de la discontinuité D1 ; 1 : calcaire à orbitoididés ; 2 : remplissages bioclastiques des poches de corrosions, dans lesquels a été mesurée l'anomalie en iridium.
Plate 1: The Larcan quarry.
Fig. 1. D1 discontinuity (K/T boundary) at the top of Gensac marly-limestones, in the old part of the exploitation.
Fig. 2. Situation of the Larcan hadrosaur (F) compared to the D1 discontinuity.
Fig. 3. The excavation between discontinuities D1 (K/T boundary) and D2 (intra-Paleogene) materializes the position of the Larcan Calcaire à algues.
Fig. 4. Detail of D1 discontinuity; 1: limestone with orbitoids; 2: bioclastic fillings of the corrosion pockets in which the iridium anomaly was measured.
Les deux éléments crâniens d'hadrosaure découverts à Larcan proviennent de la partie récente de l'exploitation. C'est dans la dernière séquence des Marno-calcaires de Gensac que les ossements ont été découverts, superposés l'un à l'autre
(Fig. 3 ![]() ). La matrice qui les liait correspond à des silts calcareux sombres dans lesquels sont présents de nombreux micro-organismes benthiques et planctoniques : Hellenocyclina beotica, Praestorssella roestae, Siderolites calcitrapoides,
calcisphères, ... La discontinuité D1 qui a livré une concentration en iridium de
0,6 ng/g couronne cette dernière séquence ; elle se situe au plus à
1 m au-dessus de la découverte.
). La matrice qui les liait correspond à des silts calcareux sombres dans lesquels sont présents de nombreux micro-organismes benthiques et planctoniques : Hellenocyclina beotica, Praestorssella roestae, Siderolites calcitrapoides,
calcisphères, ... La discontinuité D1 qui a livré une concentration en iridium de
0,6 ng/g couronne cette dernière séquence ; elle se situe au plus à
1 m au-dessus de la découverte.
Le matériel figuré est déposé dans la collection particulière de Dominique sous le n° REP-LCR-k6-001.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 3 :
Maxillaire et carré de l'hadrosaure de Larcan (n° REP-LCR-k6-001).
Figure 3: Maxilla and quadrate of the Larcan hadrosaur.
Il s'agit d'un maxillaire droit ; il mesure 115 mm de longueur et 42 mm de hauteur et appartient certainement à un individu juvénile. Étant encore dans sa gangue de roche seule sa face latérale peut être observée.
Les dents sont préservées, sauf à l'extrémité antérieure, et la présence du carré (appartenant vraisemblablement au même individu) positionné au beau milieu de l'échantillon ne nous permet pas de compter le nombre exact d'alvéoles dentaires. Les dents sont munies d'une véritable crête médiane proéminente qui se développe sur la face labiale émaillée. Il n'est pas possible de voir si la couronne possède des denticules marginaux et si des rides secondaires sont présentes de part et d'autre de la crête médiane.
Le plateau ectoptérygoïde est large et s'étend sur la moitié postérieure de la longueur du maxillaire ; latéralement le processus ectoptérygoïde forme un fort bourrelet et se termine en un renflement proéminent latéral au-dessus duquel s'élève la surface articulaire du jugal. Ce renflement présente à sa base deux gros foramens neurovasculaires alignés.
Le processus dorsal du maxillaire, légèrement tronqué sur notre spécimen, est large et son maximum de hauteur semble situé au milieu de la longueur de l'os (forme générale du maxillaire triangulaire et symétrique). Le processus dorsal se prolonge antérieurement par une véritable crête latérale arrondie fortement développée.
La partie antérieure du maxillaire montre dorsalement un large plateau qui s'affine progressivement pour se terminer en pointe et forme le processus antéro-ventral qui seul subsiste.
Ce maxillaire ressemble aux spécimens MDE-Ma3-15 et MDE-Ma3-16 provenant du site de Tricouté 3, situé au Sud-Ouest du Village de Marignac-Laspeyres (Anticlinal d'Aurignac) où la Formation des Marnes d'Auzas a livré de nombreux restes d'hadrosaures (, 2003). Les spécimens de Tricouté 3 avaient été provisoirement attribués au genre Pararhabdodon du Crétacé supérieur espagnol ( et alii, 1993, 1999 ; & , 2009). Il semblerait, toutefois, que notre spécimen, ainsi que ceux de Tricouté 3 appartiennent à un nouvel hadrosauridé différent de Pararhabdodon (comm. pers. P. ). En effet, le fort développement de la crête latérale qui prolonge antérieurement le processus dorsal n'a pas d'équivalent chez les hadrosauridés décrits à ce jour, à l'exception d'Aralosaurus tuberiferus ("curved lateral crest" de et alii, 2004, Fig. 4).
Un autre hadrosauridé, Arenysaurus ardevoli, du Maastrichtien supérieur d'Arén (Huesca, Espagne) a récemment été décrit ( et alii, 2009). Toutefois son maxillaire n'étant pas connu, toute comparaison avec notre spécimen est impossible.
Il s'agit vraisemblablement d'un carré fortement érodé, mesurant 80 mm de longueur, dont la forme générale rappelle celui des hadrosaures. L'état de conservation du spécimen ne nous permet pas une description rigoureuse.
Quoiqu'il en soit, sa taille et son association au maxillaire décrit ci-dessus, permettent l'attribution quasi certaine de ces deux restes d'hadrosaure au même individu juvénile.
La découverte de restes d'organismes continentaux, autres que végétaux, dans le milieu marin, sans être très fréquent, n'est pas non plus un fait exceptionnel (,
1994). Il a été mentionné ces dernières années des découvertes comparables : restes de dinosauriens dans le Campanien de Tercis ( et alii,
2001), reste du plus ancien proboscidien (Phosphatherium escuilliei) dans le Thanétien marin du Maroc ( et alii,
1998) ; coquille de Lychnus elongatus, gastéropode terrestre, dans un niveau à ammonites hétéromorphes du Campanien de l'Ariège ( et alii,
1998). Dans la plupart des cas il s'agit de corps flottés post-mortem ou dans le cas de Lychnus elongatus d'un organisme qui a dérivé sur un "radeau" végétal. L'intérêt de ces découvertes est divers, souvent anecdotique mais parfois de premier plan comme pour Phosphatherium par exemple. Dans le cas de l'hadrosaure de Larcan l'intérêt est double. En effet, les restes d'hadrosaures sont fréquents dans les Petites Pyrénées de la Haute-Garonne : Jadet ( et ,
1973) ; Lestaillats ( et alii,
1999) ; Marnes d'Auzas ( et alii,
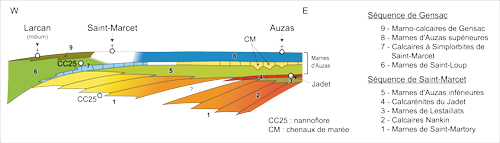
2002 ; , 2003). Les Marnes d'Auzas sont considérées comme l'équivalent laguno-continental des Marno-calcaires de Gensac, dans une séquence de dépôt désignée Séquence de Gensac
: Fig. 4 ![]() (, 1991 ; et ,
2006). Il paraît donc logique d'admettre que c'est de ce milieu laguno-continental et donc des Marnes d'Auzas que provient l'hadrosaure de Larcan. Les nombreux chenaux fluviaux qui parcouraient cette formation ont permis le transport jusqu'à la mer de la dépouille de ce jeune spécimen dont on ignore tout des causes de la mort (noyade, maladie ou prédation ?). L'autre intérêt réside dans l'âge Maastrichtien terminal de la taphocoenose. Jusqu'à présent, seuls des arguments essentiellement indirects permettaient de soutenir l'âge Maastrichtien supérieur des faunes des Marnes d'Auzas ( et alii,
2002). Cette découverte clôt le débat et permet d'affirmer, contrairement à une opinion récemment exprimée ( et ,
2003), que les dinosaures sont présents dans le Maastrichtien supérieur du versant nord des Pyrénées, au moins jusqu'à
1 m sous la limite K/T.
(, 1991 ; et ,
2006). Il paraît donc logique d'admettre que c'est de ce milieu laguno-continental et donc des Marnes d'Auzas que provient l'hadrosaure de Larcan. Les nombreux chenaux fluviaux qui parcouraient cette formation ont permis le transport jusqu'à la mer de la dépouille de ce jeune spécimen dont on ignore tout des causes de la mort (noyade, maladie ou prédation ?). L'autre intérêt réside dans l'âge Maastrichtien terminal de la taphocoenose. Jusqu'à présent, seuls des arguments essentiellement indirects permettaient de soutenir l'âge Maastrichtien supérieur des faunes des Marnes d'Auzas ( et alii,
2002). Cette découverte clôt le débat et permet d'affirmer, contrairement à une opinion récemment exprimée ( et ,
2003), que les dinosaures sont présents dans le Maastrichtien supérieur du versant nord des Pyrénées, au moins jusqu'à
1 m sous la limite K/T.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 :
Organisation séquentielle des dépôts du Maastrichtien dans la partie occidentale des Petites Pyrénées de la Haute-Garonne.
Figure 4: Sequential organization of the Maastrichtian deposits in the western part of the Petite Pyrenees (Haute-Garonne area).
Pour (1969), auteur d'une première étude de ce site, deux formations s'observaient à Larcan : les Marnes d'Auzas du Maastrichtien et le Calcaire à algues attribuée au Danien ; une discontinuité séparait les deux et constituait donc la limite entre le Crétacé et le Tertiaire. C'est cette conception qui a été adoptée sur les documents cartographiques régionaux, feuilles de Saint-Gaudens (, 1970) et de Boulogne ( et , 1977) et par et (1981). Ultérieurement, les travaux biostratigraphiques et sédimentologiques de (1985), ont démontré l'existence d'une seconde discontinuité au toit des Calcaires à algues et proposé pour toutes ces formations un cadre lithologique et stratigraphique formel. En particulier, la reconnaissance dans le 1/3 inférieur des Calcaires à algues, d'une rare microfaune crétacée à "Hellenocyclina beotica, H. gr. visserae-charentensis, Planorbulina cretae, associée à Solenomeris o'gormani et à de nombreuses Mélobésiées telles Archaeolithothamnium sp., Lithothamnium aff. villaegranis", a entrainé l'attribution au Maastrichtien terminal des Calcaires à algues et positionné la limite Crétacé-Tertiaire au niveau de la seconde discontinuité. Les travaux de ont aussi démontré les relations existant entre ces calcaires à algues et les calcaires à bryozoaires des Dômes annexes des Petites Pyrénées (vallée de la Gesse, sources de Lerd et Bois de la Taillade), les réunissant sous l'appellation de Calcaires à algues et bryozoaires de Larcan-Nizan et leur attribuant par voie de conséquence le même âge Maastrichtien terminal. Cependant, (1985) constate que, dans les Dômes annexes, la base des Calcaires à bryozoaires, séparée comme à Larcan des Marno-calcaires de Gensac par une nette discontinuité, ne renferme que des microfaunes (Orbitoididés) remaniées. C'est sur ces bases stratigraphiques que (1991), et (1991) ont par la suite proposé une reconstitution en termes de stratigraphie séquentielle du Crétacé terminal sous-pyrénéen.
La découverte dans la vallée de la Gesse ("Source de Pichore" in et alii, 1997, 1998) d'un niveau enrichi en iridium à la limite entre les Marno-calcaires de Gensac et les Calcaires à bryozoaires de Nizan et, dans ces mêmes calcaires comme dans ceux de Larcan, de microfaunes planctoniques "dano-sélandiennes" déterminées spécifiquement sur lames minces, a réhabilité la position de la limite K/T, telle que classiquement admise. et alii (1998) ont par la suite rapporté la présence de l'iridium à Larcan, dans une position similaire (cf. supra).
Il est incontestable que cette découverte de l'iridium, dans deux sites distants l'un de l'autre de 25 km environ, constitue un élément important de corrélation et que dans l'état actuel des connaissances, établies essentiellement en domaine pélagique, il paraît raisonnable de placer à ce niveau la célèbre limite K/T. Malgré tout, les travaux de et alii (1997, 1998) ont introduit d'importantes confusions. Dans les Dômes annexes, les microfaunes maastrichtiennes considérées comme remaniées à la base des Calcaires à bryozoaires (, 1985, cf. ante), sont considérées comme autochtones par ces auteurs ; mieux, ils ont proposé pour l'"Association Orbitoididés-Sidérolites" le concept de "Taxons franchissant la limite K/T". Une autre ambiguïté de ce travail est la citation, à Taillade (marnière de Saint-Loup), de taxons du Paléocène, Eoglobigerina eobulloides, Globanomalina planocompressa, associés à Pseudoguembelina hariaensis à la partie sommitale des Marno-calcaires de Gensac ; ils n'étaient pas mentionnés dans l'étude de 1997 qui fait état jusqu'au sommet des "Marno-calcaires jaunes de Gensac" (Marnes bleues de Saint-Loup en réalité) uniquement de taxons du Maastrichtien supérieur ("Zone à P. hariaensis"). Toutes ces contradictions laissent perplexe, d'autant que les auteurs considèrent comme "très faible (ou nulle)" l'érosion des Marnes bleues de Saint-Loup par le Calcaire à bryozoaires sus-jacents, ce que dément le remaniement de microfaunes crétacées dans ces même Calcaires à bryozoaires. Nulle part, à notre connaissance, il n'a été mentionné la coexistence de microfaunes benthiques du Crétacé terminal et planctoniques du Paléocène, à moins que les premières ne soient remaniées dans des niveaux plus récents ; si tel n'était pas le cas, quels arguments micropaléontologiques prévaudraient dans la datation de l'horizon à iridium : la disparition des microfaunes du Crétacé ou l'apparition des microfaunes du Paléocène ? Dans l'un et l'autre cas, le niveau à iridium perdrait sa valeur universelle de limite K/T. Il apparaît donc que hormis la mention de l'horizon à iridium dans la vallée de la Gesse (Pichore), les propositions biostratigraphiques de et alii (1997, 1998) relatives à la limite K/T, sont en désaccord avec les observations établies dans les nombreux sites où cette limite a été étudiée.
La découverte de fragments crâniens d'hadrosaure dans les derniers niveaux marins du Maastrichtien qui précédent immédiatement le niveau à iridium de la limite K/T n'est pas qu'un simple fait
anecdotique. Cette découverte a permis :
(1) de conforter l'idée que les Marnes d'Auzas laguno-continentales et les Marno-calcaires de
Gensac, marins, sont, en partie au moins, des équivalents latéraux ;
(2) d'apporter la preuve de la présence des dinosaures jusque dans les niveaux les plus élevés du Maastrichtien du versant nord des Pyrénées ;
(3) de relever les incompatibilités relatives à la dernière conception biostratigraphique de la limite K/T dans le
"Comminges occidental".
Les auteurs remercient : Mme L. et M. N. pour leur aide dans la mise en forme de la version anglaise, Mme C. pour son implication dans la réalisation des illustrations, M. Marc , découvreur du spécimen fossile, et les rapporteurs E. et P. dont les remarques autorisées ont grandement permis d'améliorer cette contribution.
R. & X. (2003).- Dinosaurs of France.- Comptes Rendus Palevol, Paris, vol. 2, p. 27-44.
M. (1991).- Séquences de dépôt et limite de blocs dans le Crétacé terminal et Paléocène basal du bassin d'avant-pays sous-pyrénéen (Petites Pyrénées Dômes annexes, France).- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, vol. 15, n° 2, p. 411-437.
M. & B. (2006).- Les Marnes d'Auzas (Maastrichtien supérieur sous-pyrénéen). Stratigraphie et paléoenvironnements, associations d'ostracodes.- Revista Española de Micropaleontologia, Madrid, vol. 38, n° 2-3, p. 309-320.
M. & F. (1991).- Stratigraphie séquentielle des séries du Sénonien supérieur dans la zone sous-pyrénéenne (Petites Pyrénées et Dômes annexes, SW France).- Comptes Rendus de l'Académie des Sciences, Paris, vol. 312, p. 393-398.
M., M. & J. (1998).- Découverte de Lychnus elongatus 1884, Gastéropode continental, dans le Campanien marin nord-pyrénéen.- Revue de Paléobiologie, Genève, vol. 17, n° 2, p. 541-547.
E. (1994).- The significance of dinosaur remains in marine sediments : an investigation based on the French record.- Berliner geowissenschaftliche Abhandlungen, vol. 13, p. 125-133.
A. & Y. (1977).- Boulogne-sur-Gesse.- Carte géologique au 1/50 000, Notice, BRGM, Orléans, feuille XVIII-45, 25 p.
M.L., J.V. & A. (1993). Pararhabdodon isonense n.gen. n.sp. (Dinosauria). Morphology, radio-tomographic study and biomechanic considerations.- Paleontologia i Evolucio, Barcelona, vol. 26-27, p. 121-131.
M.L., X., J.V. & D.B. (1999). First lambeosaurine hadrosaurid from Europe : paleobiogegraphical implications.- Geological Magazine, Cambridge, vol. 136, n° 2, p. 205-211.
E., J., H. & G. (1998).- Phosphatherium escuilliei du Thanétien du bassin des Ouled Abdoun (Maroc), plus ancien proboscidien (Mammalia) d'Afrique.- Geobios, Villeurbanne, vol. 30, p. 247-269.
P., V. & Y. (2004).- A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, (Sciences de la Terre), vol. 74 (supplément), p. 139-154.
Y. (2003) Les faunes de vertébrés continentaux du Maastrichtien supérieur d'Europe. Systématique et biodiversité.- Strata, Toulouse, série 2, vol. 41, 81 p.
Y., L. & M. (1999).- Découverte d'un gisement à vertébrés dans le Maastrichtien supérieur des Petites Pyrénées.- Comptes Rendus de l'Académie des Sciences, Paris, vol. 328, p. 781-787.
Y., M. & J. (2002).- Late Maastrichtian continental vertebrates from southwestern France: correlation with marine fauna.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 187, p. 121-135.
B. (1985).- Le Crétacé terminal et le Paléocène basal dans les Petites Pyrénées et les Dômes annexes. Biostratigraphie Sédimentologie.- Strata, Toulouse, série 2, vol. 4, 276 p.
G.S., E., J., C. & J.-P. (2001).- Reptilian remains from the late Campanian at Tercis les Bains (Landes) and Vanxains (Dordogne). In : G.S. (Éd.), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, p. 665-674.
J.P. (1970).- Saint-Gaudens.- Carte géologique au 1/50 000, Notice, BRGM, Orléans, feuille XIX-46, 23 p.
J.P. & P. (1973).- Découverte d'un fragment de dentaire d'Hadrosaurien (Reptile Dinosaurien) dans le Crétacé supérieur des Petites Pyrénées (Haute-Garonne).- Bulletin du Muséum National d'Histoire Naturelle de Paris, série 3, n° 130, Sciences de la Terre 22, p. 17-27.
X., J.I., P., J.L., N., O. & J.I. (2009).- The last hadrosaurid dinosaurs of Europe: a new lambeosaurine from the uppermost Cretaceous of Aren (Huesca, Spain).- Comptes Rendus Palevol, Paris, vol. 8, p. 559-572.
B., M.J. & P. (1997).- Identification de microfaunes dano-sélandiennes des sous-zones P1c à P3b dans les "Calcaires à Algues / Calcaires à Bryozaires de Larcan-Nizan" ex-crétacés, du Comminges occidental (Zone sous-pyrénéenne, Haute-Garonne).- Bulletin de la Société d'Histoire Naturelle de Toulouse, tome 133, p. 83-90.
B., M.J., P., E. & R. (1998).- La limite Crétacé-Tertiaire en domaine marin dans les Pyrénées centrales (zone sous-pyrénéenne, France).- Comptes Rendus de l'Académie des Sciences, Paris, tome 326, p. 647-653.
A. & J.R. (2009).- Parahabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia.- Cretaceous Research, London, vol. 30, p. 1238-1246.
R., E., Y., J. & M. (1998).- Iridium at the K/T boundary in the Larcan quarry (Sub-pyrenean Zone, Southern France).- 2ème Congrès Français de Stratigraphie, Paris, p. 143.
E. (1969).- Un contact intéressant entre le Maastrichtien et le Danien dans les Petites Pyrénées. In : P. & H.H. (Éds.), Proceedings of the 1st International Conference on Planktonic Microfossils, Geneva, 1967.- E.J. Brill, Leiden, II, p. 672-674.
Y. & J. (1981).- Itinéraire géologique pédestre Nord-Sud dans les Petites Pyrénées de la Haute-Garonne.- Revue des Comminges et des Pyrénées centrales, Saint-Gaudens, vol. 94, p. 351-364.