![]()
Contents - Sommaire
English version ![]()
[1. Introduction] [2. Lithostratigraphy
of ...]
[3. The planktonic foraminifera] [4. The location of the ... boundary]
[5. Conclusions] [Bibliographic references]
and ... [Plates]
Version française ![]()
[1. Introduction] [2. lithostratigraphie
de ...]
[3. Les foraminifčres planctoniques] [4. La position de la limite ...]
[5. Conclusions] [Références bibliographiques] et ... [Planches]
[The Abiod Formation at Ellčs (Tunisia):
tripartite lithology,
biohorizons based on globotruncanids and ammonites,
duration, location of Campanian-Maastrichtian boundary,
correlation with Kalaat Senan and the Tercis (France) stratotype]
Université de Mons, Faculté Polytechnique, 9 rue de Houdain, 7000 Mons
(Belgium)
ETAP, Entreprise Tunisienne d'Activités Pétroličres, Laboratoire de
Biostratigraphie, Centre de Recherche et de Développement Pétroliers La Charguia II, Tunis
(Tunisia)
Manuscript online since August 27, 2010
![]()
In central Tunisia near the village of Ellčs is Wadi Ed Dam. There the Abiod Formation, underlain by the Kef marls and capped by the El Haria marls, crops out in an almost uninterrupted exposure about 286 m thick. In view of its possible use as a regional litho- and bio- stratigraphic reference section for the southern border of the Tunisian trough it is examined in detail here. Fossils studied include foraminifera (115 samples representing 6 micropaleontologic zones, see chart) and ammonites. The biohorizons established are based on the first (FO) or last occurrence (LO) of certain species of planktonic foraminifers in addition to the FO of already established foraminiferal zonal markers. Ammonite markers found include both the FO and LO of some forms, or just one or the other.
The location of the Campanian-Maastrichtian (Cp-Ma) boundary time line at Ellčs is established by comparison with that determined at Kalaat-Senan and with that of the international stratotype of Tercis (France), ratified by the International Union of Geological Sciences.
At Kalaat-Senan the Cp-Ma boundary is set at the lower third of the upper indurated limestone bar of the Abiod Formation - the Ncham Member. Its location at Kalaat-Senan is based on the upper limit of occurrence of Nostoceras (Nostoceras) hyatti and Pseudokossmaticeras brandti together with the FO of Nostoceras magdadiae, a Maastrichtian ammonite. At Ellčs several of these same markers are present, along with the respective planktonic foraminiferal zones.
The length in years of some lithologic units and paleontologic zones in the Upper Cretaceous succession of wadi Ed Dam was calculated using cyclostratigraphy. The Abiod Formation represents 13.2 Ma, the Radotruncana calcarata Zone 0.79 Ma, and the Nostoceras (Bostrychoceras) polyplocum Zone 0.97 Ma. A proposed subdivision of the Upper Campanian is recommended at the base of the Bostrychoceras Zone.
et alii's 1999 proposal for fixing the location of the Cp-Ma boundary at the top of a limestone bed in the El Haria marls is reviewed.
Campanian-Maastrichtian; Abiod Formation; Central Tunisia; Ellčs; Tercis; Kalaat Senan; globotruncanids; ammonites.
F. & M. (2010).- The Abiod at Ellčs (Tunisia): stratigraphies, Campanian-Maastrichtian boundary, correlation [L'Abiod d'Ellčs (Tunisie) : stratigraphies, limite Campanien-Maastrichtien et corrélation].- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/04 (CG2010_A04)
L'Abiod d'Ellčs (Tunisie) : stratigraphies, limite Campanien-Maastrichtien et corrélation.- En Tunisie centre-nord, prčs d'Ellčs, l'oued Ed Dam expose un enregistrement sédimentaire quasi continu de la Formation Abiod encadrée par la Formation Kef au-dessous et la Formation El Haria au-dessus. Cette Formation Abiod, tripartite, a été étudiée pour servir de référence litho-biostratigraphique régionale sur la bordure sud-est du Sillon tunisien oů elle atteint prčs de 286 m. Le contenu en foraminifčres planctoniques d'une série d'échantillons est reporté sur un tableau de distribution et soutient une division en 6 biozones micropaléontologiques. Les positions des ammonites récoltées sont repérées par rapport ŕ la succession lithologique. Plusieurs biohorizons sont décrits qui correspondent aux premičres et derničres présences de foraminifčres, par exemple de Radotruncana calcarata, et aux émergences des marqueurs de zones comme Globotruncana ventricosa, Gansserina gansseri et Contusotruncana contusa ainsi que les premičres et/ou les derničres présences de plusieurs ammonites marqueurs comme Nostoceras (Bostrychoceras) polyplocum, Pseudokossmaticeras brandti et Nostoceras (Nostoceras) hyatti.
L'un des points d'ancrage de la succession stratigraphique étant la limite Campanien-Maastrichtien (Cp-Ma), on définira la position de cette ligne-temps ŕ Ellčs par comparaison avec la coupe de Kalaat Senan et celle du stratotype international de Tercis (France) ratifiée par l'Union internationale des sciences géologiques.
À Kalaat Senan, la limite Cp-Ma est placée au tiers inférieur de la barre supérieure de l'Abiod ou Membre Ncham, au sommet de la Zone ŕ N. (N.) hyatti, contenant P. brandti, ammonites de la partie supérieure du Campanien, et ŕ l'apparition de Nostoceras magdadiae, ammonite de la partie inférieure du Maastrichtien. À Ellčs, plusieurs de ces marqueurs ont été retrouvés comme aussi les zones de foraminifčres planctoniques ŕ Radotruncana calcarata, ŕ Globotruncana falsostuarti, ŕ Gansserina gansseri et ŕ Contusotruncana contusa.
En utilisant l'outil cyclostratigraphique, la durée des diverses unités lithologiques et celle de quelques zones paléontologiques sont précisées : la durée totale de dépôt de la Formation Abiod ŕ Ellčs est mesurée ŕ 13,2 Ma, celle de la Zone ŕ R. calcarata ŕ 0,79 Ma et celle de la Zone ŕ N. (B.) polyplocum ŕ 0,97 Ma. Une limitation du Campanien supérieur ŕ la base de la Zone d'ammonite ŕ N. (Bostrychoceras) polyplocum est proposée.
La position de la limite Cp-Ma proposée par et alii en 1999 au sommet d'une série de bancs calcaires dans la Formation El Haria est discutée.
Campanien-Maastrichtien ; Formation Abiod ; Tunisie centrale ; Ellčs ; Tercis ; Kalaat Senan ; globotruncanidae ; ammonites.
In north central Tunisia the Abiod Formation marks its location in the upper portion of the Late Cretaceous very clearly by its effect on the topography: a double line of low hills separated by a slight median depression. This morphology reflects the general tripartite composition of the Formation: a lower bar of indurated white limestone, intermediate turquoise marls and an upper bar again of white limestone. The assemblage is of Campanian-Maastrichtian age and therefore includes the boundary between the two geologic stages. In accordance with changes in criteria during the last century this limit was at first placed near the top of the lower limestone, then at about the middle of the intermediate marls or at the base or at the top of the upper limestone. These fluctuations in position were due to the absence of an international agreement regarding the criteria for the location of the boundary between the stages.
Today, the section at Tercis (Landes, France) contains the international stratotype of the Campanian-Maastrichtian boundary. A number of discrete but confirmatory criteria were involved in the selection of this site and approval of the exact location of the boundary. In September, 1999, they were approved by the Working Group on the Maastrichtian; in June, 2000, they were recommended by the International Subcommission on Cretaceous Stratigraphy; in December, 2000, they were adopted by the International Commission on Stratigraphy and ratified by the International Union of Geological Sciences in February 2001 (, 2002).
Using this section for comparison, and employing several biostratigraphic indices, it is now possible to locate more precisely the Campanian-Maastrichtian boundary in north central Tunisia. It is in the lower third of the upper limestone bar of the Abiod Formation.
This location is well defined in the areas where the Abiod Formation approaches an almost unbroken exposure which is the case for several sections in north central Tunisia (the "Sillon tunisien" "Tunisian trough" of et alii, 1952). Good examples occur at El Kef (, 1956) where, as macrofossils are relatively rare, the distribution of microfossils, in particular that of the planktonic foraminifers was emphasized (, 1955, 1956). They served also at Kalaat Senan where the ammonites seem to be more numerous and planktonic foraminifers too are abundant ( et alii, 2000).
In localities other than those of north central Tunisia the Abiod Formation presents itself less commonly in its "classic" tripartite aspect: sometimes the lower bar is less well developed, sometimes the upper bar is less prominent, and in other localities the formation is an almost monolithic entity, as is the case in certain sections north of Kairouan and in wells in the plain southeast of Kairouan. In order to date more precisely these different lithologies of the Abiod Formation, a section at the southeast edge of the "Tunisian trough" in which sedimentation appears to have been continuous was studied first. It is the Ellčs section, about 45 km southeast of El Kef. There, as at Kalaat Senan the beds of limestone and marly limestone contain macrofossils, among them ammonites, along with planktonic foraminifers visible in thin sections, while the marls yield profuse numbers of isolated planktonic foraminifers. The vertical distribution of the planktonic species and the zonation derived therefrom constitutes the basis for stratigraphic attributions in successions where the thickness of the Abiod Formation is greatly reduced, either by lacunae or by the development of a uniformity in lithology caused by an increase in the content of limestones. Later, more precise attributions will make possible isochronic paleogeographic maps that may serve to find lithologic or structural events favorable to the entrapment of fluids such as hydrocarbons.
In what follows, these points are taken up successively:
In a statement of conclusions, the value of several macro- and micropaleontologic horizons will be emphasized for they constitute time lines usable for improving the correspondence of outcrop sections at least in central Tunisia as well as their help in the correlation of petroleum research wells.
Rakia (now Mme ) studied the Cretaceous-Paleogene boundary of the Ellčs section and presented the findings in her thesis of 1978. She analyzed the limestone-marl alternances at the top of the upper bar of the Abiod Formation along with the marls at the base of the El Haria Formation. She noted the presence of planktonic foraminifera near the top of the upper bar (the upper limit of our Ncham Member) and in the alternances above (our Gouss Member) among which we retain the species "Globotruncana stuarti, falsostuarti, gagnebini (= aegyptiaca), arca, contusa-patelliformis, fornicata, linneiana, ventricosa and Rugoglobigerina hexacamerata" the association characterizing her "Lower Maastrichtian" (an interpretation still valid today, at least in part) and, higher, in her "Upper Maastrichtian" in addition to these species, the presence of "Globotruncana gansseri, G. contusa-galeoidis, Globotruncanella havanensis and Trinitella scotti and then Abathomphalus mayaroensis". Note that no "Globotruncana calcarata" is reported in the El Haria Formation marls, a point that will be useful later in our § 4.6.
In the explanation accompanying the Ebba Ksour 1: 50,000 geologic sheet, (1979) gives a succinct description of a good outcrop of the Abiod Formation in the sector called El Magoula (or El Mazoula, his page 11). It is undoubtedly the succession that we studied in detail. considers it to be representative, and capable of "serving as an example" of the Abiod Formation in the northeastern portion of the sheet. For him, the thickness of the entire Formation is 235 m, while our measurement is in the neighborhood of 286 m. The author had clearly recognized its tripartite aspect and noted the presence of ammonites of the genus Bostrychoceras near the top of the lower bar and as well the presence of "Globotruncana calcarata" in the "membre marneux moyen" (middle marly member). The chronostratigraphic interpretation is based on the micropaleontologic interpretations of A.-L. and P. of the Tunisian Geologic Survey. They called the lower bar "Upper Campanian", the "membre marneux moyen" "Upper Campanian (upper part)" and the upper bar "Lower Maastrichtian". This evaluation, as will be seen further on, agrees in part with our interpretation, the difference being that we put the Campanian-Maastrichtian boundary well within the upper bar, where it is located on the basis of ammonite collections.
The more recent publications of & (1998) and of et alii (1999) must also be mentioned. The authors state that in the Ellčs section -exactly the same one as that discussed below- the upper limit of the "Globotruncanita calcarata" Zone (for them the Campanian-Maastrichtian boundary) is near the top of the "limestones" that must be the upper bar of the Abiod. Later they put the boundary in "grey marly shales" with "limestone layers", that is to say probably in the El Haria marls. Both are contestable as we shall discuss farther on.
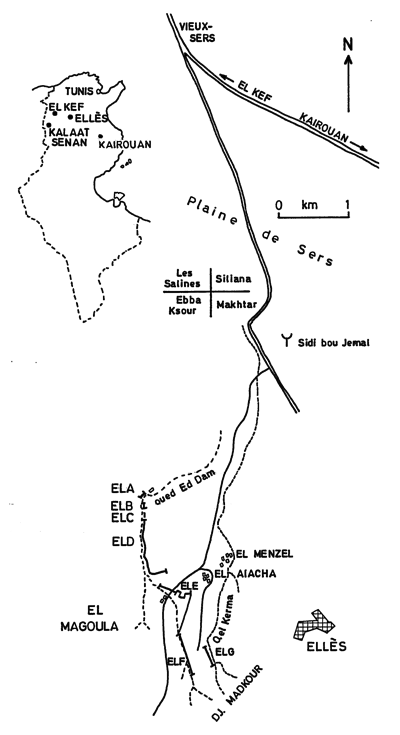
Click on thumbnail to enlarge the image.
Figure 1: Geographic location of the Ellčs section of the Abiod Formation (F.R.).
About 2 km west of the village of Ellčs, wadi Ed Dam offers an almost continuous section of the Abiod Formation. It rests on the Kef marls Formation and is capped by the El Haria marls, that are also completely exposed (Fig. 1 ![]() ). The section was measured using a Jacob's staff 1.50 m long with a sighting device capable of measuring thicknesses to within few per cent. Marls and limestones were sampled regularly for the collection of isolated foraminifera or the preparation of thin sections. In all 115 samples were collected from the 359 m of partial measured sections of which 286 m represent the Abiod Formation. During and after the lithologic examination of the succession there was an active search for macrofossils and several of them that are particularly significative will be reported on.
). The section was measured using a Jacob's staff 1.50 m long with a sighting device capable of measuring thicknesses to within few per cent. Marls and limestones were sampled regularly for the collection of isolated foraminifera or the preparation of thin sections. In all 115 samples were collected from the 359 m of partial measured sections of which 286 m represent the Abiod Formation. During and after the lithologic examination of the succession there was an active search for macrofossils and several of them that are particularly significative will be reported on.
Remarks. Among the macrofossils we shall emphasize particularly the ammonites we collected during lithologic studies. At certain levels they are not rare at Ellčs. They will provide, in part, support for the establishment of correlations with the Kalaat Senan section which is some 60 km southwest of Ellčs. However, the Ellčs section has been the source of collections of ammonites much more important and more systematic than ours. They were made by Stijn (KLU, Leuven, Belgium) for his thesis on the Campanian-Maastrichtian ammonites of Tunisia and will have numerous plates illustrating the species found (unpublished, 2010).
In addition, some inoceramids, also collected during field work, were submitted to the late Annie (Royal Institute of Natural Sciences of Brussels). The several species cited here have determinations that she told us about before her disappearance on 1 September 2006.
Other macrofossils were collected during field work (some echinoids, bivalve and brachiopod fragments) but they are sparse in number and their poor preservation has resulted in their not having been submitted to specialists for examination.
The lithologic description has benefited from previous work by et alii (2000) in the Kalaat Senan region where the Abiod Formation was subdivided into seven members, from top to bottom:
| Formations | Members | Previous names |
| El Haria Fm.: marls | ||
| Abiod Formation | Gouss Mb.: upper alternances | upper bar |
| Ncham Mb.: limestone bar | ||
| Gourbeuj Mb.: alternances | ||
| Akhdar Mb.: intermediate marls | middle marls | |
| Mahdi Mb.: alternances | lower bar | |
| Haraoua Mb.: limestone bar | ||
| Assila Mb.: basal alternances | ||
| Kef Fm.: marls | ||
In the literature the "lower bar" corresponds to the Assila and Haraoua
members to which the Mahdi Member is often attached, while the "upper bar" corresponds to the Ncham and Gouss
members (often with the Gourbeuj Member). According to the authors the "middle marls" refers to the Akhdar Member (and sometimes, but rarely or in part to the alternances of the Mahdi and Gourbeuj
members). Seven partial sections have been described and designated ELA to
ELG. The first three do not have any beds in common whereas the four others do, so on the ground they permit easy matching, so establishing continuity in the lithologic succession is not difficult (Fig. 2 ![]() ).
).
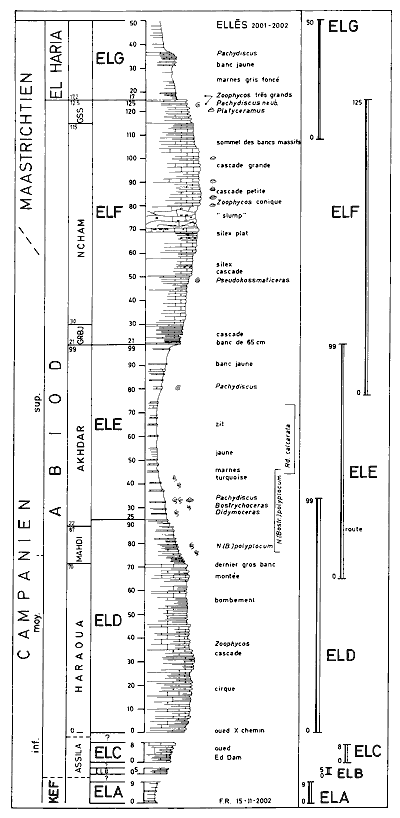
Click on thumbnail to enlarge the image.
Figure 2: Synthetic lithologic succession derived from the partial sections of the Abiod Formation at Ellčs (F.R.).
Kef Formation (named by ,
1978). Section ELA (Fig. 2 ![]() ): marls with limestone beds. Seen for 9 m on the left bank of wadi Ed Dam. They are greenish-gray marls, almost flat-lying, with a 60 cm limestone at the base and farther up several beds of marly limestone ranging from 10 to 30 cm in thickness.
): marls with limestone beds. Seen for 9 m on the left bank of wadi Ed Dam. They are greenish-gray marls, almost flat-lying, with a 60 cm limestone at the base and farther up several beds of marly limestone ranging from 10 to 30 cm in thickness.
Abiod Formation (named by et alii,
1954). Partial sections ELB, ELC, ELD, ELE, ELF and a part of ELG (Fig. 2 ![]() ): two bars of limestone bounded at the base and top by alternances framing intermediate marls. et alii
(2000) distinguish several members.
): two bars of limestone bounded at the base and top by alternances framing intermediate marls. et alii
(2000) distinguish several members.
Assila Member (sections ELB and ELC): basal alternances, about 13 m accessible, probably a total of 17 to 20 m.
These are the first limestone beds visible at the downstream end of wadi Ed Dam, above the Kef marls. Stratigraphic continuity is not assured visually between the three partial sections ELA, ELB and ELC. However, as the dip is very low, it may be assumed (if the existence of important faulting is excluded because the limestones are not fractured more than is normal and the succession is not incoherent) that the interruptions in visibility do not represent more than a few meters. The limestones grayish-white, well-indurated with a conchoidal fracture are interbedded with marly limestones, their thicknesses measured in decimeters.
Haraoua Member (section ELD, from 0 to 70 m): the lower limestone bar, 70 m.
Section ELD in wadi Ed Dam starts at a cross-road. From there the section is almost unbroken until its contact with the El Haria Formation. Southeast dip increases regularly to reach about 10°, and locally a little more up to 12°.
In its topographic expression the Haraoua Member is a belt of hills made up of several tens of meters of white limestones in beds 20 to 90 cm thick, more or less amalgamated and with stratification commonly slightly undulant. These limestones are separated by interbeds of marly limestone 10 to 20-30 cm thick. The limestones, grayish white to white, are compact with a conchoidal fracture. In thin section foraminifers are numerous in a wackestone matrix. Impressions of inoceramids from 10 to 20 cm in diameter and trace fossils of the Zoophycos group are not rare.
Mahdi Member (section ELD, from 70 to 87 m): 17 meters of alternances
The thick limestones of the Haraoua Member grade progressively down to 20 and 30 cm, already a little more marly and grayer, alternating with beds of gray marly limestone 0 to 30 cm thick, generally altered and leached. The first occcurence of the ammonite Nostoceras (Bostrychoceras) polyplocum was found in ELD 76 and two levels were recognized and followed laterally, in particular the one at 79.4 m.
Akhdar Member (section ELE from 22 to 99 m): turquoise greenish blue marls, 77 m.
The argillaceous content of the marl interbeds increases progressively, along with their thickness which attains 50 to 80 cm between the limestones that become more marly too. Several levels yield more ammonite fragments N. (B.) polyplocum, Nostoceras (Didymoceras) sp. and Pachydiscus (P.) sp. found between ELE 26.80 and ELE 45.00 with very numerous examples at level ELE 32.70, which is near a paved road. The last occurrence of N. (B.) polyplocum is at about ELE 45.00.
From 40 to 80 m the marl beds increase in thickness, up to several meters. Above 80 m, limestone beds are more numerous. Several ammonites, among them Pachydiscus sp., were still present at ELE 80.50 and 81.00. Note that the planktonic foraminifer Radotruncana calcarata is present at least between ELE 43.50 and 73.50, that is more than 30 m at least.
Gourbeuj Member (section ELF from 21 to 30m): 9 m of alternances.
The succession begins with a bed of white compact limestone 65 cm thick, succeeded by 18 or 19 beds of limestone ranging in thickness from 20 to 50 cm separated by interbeds of about 30 cm that decrease in thickness upward to 10 cm and eventually to 5 cm. The wadi Ed Dam waterfall exposes all of it very clearly. An ammonite fragment of the genus Pseudokossmaticeras was collected at the base of the member, another specimen, out of place, was found between ELF 13 and 25. With reservations, another fragment at the base of the Gourbeuj Member, its identification requiring confirmation, appears to be Nostoceras (Nostoceras) cf. hyatti (personal communication of Stijn , unpublished, 2010).
Ncham Member (ELF section, 30 to 115 m): upper limestone bar.
The limestones are of four different types, from bottom to top:
In thin sections all of these limestones have a micritic matrix in which numerous planktonic foraminifera and some benthonic forms are preserved in a wackestone type texture along with various other bioclasts.
Gouss Member (ELF section from 115 to 125 m and the ELG section from 6 to 17.20 m): upper alternances.
The interbeds thicken and become more marly, thus characterizing the Gouss Member. At the base another specimen of P. neubergicus is noted while near the top of the ELF section a rather worn impression of an ammonite 20 cm in diameter is found at ELF 122.40 in limestone eroded by the wadi. It is a Pachydiscus sp.
At about 300 m east of the ELF section, wadi El Kerma offers a well exposed section of the two upper facies of the Ncham bar (that is around 10 m of limestones with slumping containing flint nodules and about 35 m of meter-sized limestones without flint) above which are Gouss alternances which grade upward to the El Haria Formation. The upper part of the last limestone of the Gouss Member is well exposed because the overlying marls of the El Haria Formation have been removed by erosion. So this last bed provides a fine structural surface exposing impressions of inoceramids and Zoophycos as well as several types of bioturbation. In summary, the upper limit of this bed is not recrystallized, has no perforations and is not glauconitized or phosphatized, phenomena which would have allowed its surface to be interpreted as a true "hardground" (as et alii, 1998, mistakenly called it).
El Haria Formation (named by ,
1956). Section ELG from 17.20 to 50 m and higher (cf.
Fig. 2 ![]() ): gray marls with some limestone beds.
): gray marls with some limestone beds.
To summarize, several biohorizons with a time-line value are of great importance in making regional or larger scale correlations:
A recent publication has shown that the cyclostratigraphic method is a tool worthy of confidence as a measure of the time required for the deposition of a litho- or biostratigraphic unit when the sedimentation was rhythmic, as it is in the Abiod Formation ( et alii, 2009). We shall review successively the three entities: upper bar, intermediate marls and lower bar. The time values given are based on fig. 8 in et alii (2009). They are summarized in Table 1 where the T100 column represents the time based on the ± 100,000 years eccentricity cycles and the T20 column the time deduced from precession cycles of ± 20,000 years. Line "66" indicates the most probable position of the Campanian-Maastrichtian boundary and line "76" shows the level at which N. (Bostrychoceras) polyplocum appears.
| Lithological units | T100 | T20 | (T100+T20)/2 | length | length | |||
| inferred | inferred | calculated | calculated | accepted | ||||
| thickness | (Ma) | (Ma) | (Ma) | (Ma) | (Ma) | |||
| index | ELHARIA | |||||||
| 125 | 2.65 | 2.55 | 2.60±0.05 | |||||
| 10m | GOUSS | 0.325±0.025 | 0.30 to 0.35 | |||||
| 115 | 2.35 | 2.20 | 2.275±0.075 | |||||
| NCHAM | 2.275±0.075 | 2.20 to 2.35 | ||||||
| 85m | 66 | 0 | 0 | 0 | ||||
| 1.70±0.10 | 1.60 to 1.80 | |||||||
| 30 | -1.80 | -1.60 | -1.70±0.10 | |||||
| 9m | GOURBEUJ | 0.575±0.012 | 0.56 to 0.58 | |||||
| 21/99 | -2.30 | -2.05 | -2.175±0.125 | |||||
| 77m | AKHDAR | 2.40±0.05 | 2.35 to 2.45 | |||||
| 87/22 | -4.65 | -4.50 | -4.475±0.075 | |||||
| MAHDI | 0.275 | |||||||
| 17m | 76 | 0.425±0.175 | 0.25 to 0.60 | |||||
| 0.15 | ||||||||
| 70 | -5.25 | -4.75 | -5.00±0.25 | |||||
| 70m | HARAOUA | 5.175±0.50 | 4.67 to 5.67 | |||||
| 0 | -10.90 | -9.45 | -10.175±0.75 | |||||
| (17m) | ASSILA | (0.425±0.15) | (0.27 to 0.57) | |||||
| (-11.50) | (-9.70) | (-10.60±0.90) | ||||||
| KEF | ||||||||
Table 1: Duration of the members of the Abiod Formation based on a cyclostratigraphic analysis of the alternances that compose them (according to et alii, 2009). T100 and T20 columns show the location of the boundaries of the members as they relate to geologic time. 0 (zero) is set at the Campanian-Maastrichtian boundary. Time is considered positive above that level and negative below it (F.R.).
Remark: In Table 1 the values of T100 and T20 restated in their respective columns are taken from a reading of the graphic of Fig. 8 of et alii (2009) and appear with two significant figures after the period. In the 2 columns that follow the mean values (T100+T20)/2 and the durations of deposition of the members derived therefrom are produced by a simple calculation which sometimes produces 3 figures after the comma, the last obviously without significance. The last column posts a probable duration of deposition for each member of the Abiod Formation while taking into account the uncertainty between the durations derived from T100 and those from T20.
Upper bar, mean duration in Ma (= million years) (T100+T20)/2
Gouss Member (10 m): 0.325 Ma
Ncham Member (85 m): 3.975 Ma
Gourbeuj Member (9 m): 0.475 Ma
That is a total mean duration of 4.775 Ma for the upper limestone bar, which represents a sedimentation rate of uncompacted strata of 10400 cm ÷ 4775 ky = 2.17 cm/ky
Intermediate marls
Akhdar Member (77 m): 2.40 Ma
This represents rate of sedimentation for uncompacted strata of 7700 cm ÷ 2400 ky = 3.20 cm/ky
Lower bar
Mahdi Member (17 m): 0.425 Ma
Haraoua Member (70 m): 5.175 Ma
Assila Member ("18" m): 0.425 Ma
(the Assila Member was not taken into account in the measurement of alternances because it is poorly exposed at the bottom of wadi Ed Dam; here we assume that its aspect and its thickness, apparently approximating those of the Mahdi Member, make it likely that its duration too may have been similar). In this way a total mean duration for the deposition of the lower limestone bar of 6.025 Ma may be accepted (whether 5.20 to 6.85 Ma when taking uncertainties into account), so the mean rate of deposition of uncompacted sediments of 10500 cm ÷ 6025 ky = 1.74 cm/ky
In total, the time required for the deposition of the Abiod Formation, consisting of the lower bar, the intermediate marls and the upper bar is in the neighborhood of 13.2 Ma for 286 m of strata.
Radotruncana calcarata Zone
Again using fig. 8 in et alii (2009) for calculations concerning calcarata:
T100: 3.90 – 3. 30 = 0.60 Ma
T20: 3. 60 –2.65 = 0.95 Ma
That gives a mean of 0.77 Ma for the minimum duration of the zone.
However, going back to the table of basic calculations (unpublished) that served to construct Fig. 8, T100 is 765 ky long and T20 is 830 ky long, which is in accordance with the measurements based on Fig. 8 as closely as they can be read and give a mean value of 0.79 Ma which will be used here.
Nostoceras (Bostrychoceras) polyplocum Zone
Many specimens of the polyplocum group were collected in the partial sections ELD and ELE with a range from ELD 76 to ELE 45. Using fig. 8 or better the basic calculations table, T100 is 988 ky and T20 is 964 ky, for a mean of 976 ky or 0.97 Ma.
In sections ELA to ELG 115 samples were taken in 395 m of partial sections, that when synthesized totaled 323 m of which 281 m of the Abiod Formation were sampled. Knowing that there are some gaps in visibility at the base, the total thickness of the Abiod Formation can be estimated to be about 286 m. 82 of the samples were sufficiently marly to be washed, preceded by wetting in hydrogen peroxide to insure that the microfossils were freed. The other samples of limestone or marly limestones were thin-sectioned or treated with acetic acid. If all the information obtained from the overlapping partial sections is grouped, the isolated specimens and those in thin section are distributed over 86 sampling levels all indexed on
Fig. 3 ![]() .
.
The determination of the whole specimens were made by M.M. and F.R. using the figures and the criteria in the Atlas of Upper Cretaceous Planktonic Foraminifera ( et alii, 1984). The preparation of specimens for scanning by the ETAP electronic microscope in Tunis was done by F.R.
In the entire section 28 main taxa were indexed of which 6 were very rare or of uncertain determination. Certain species are distributed throughout the whole of the succession like Globotruncanita stuartiformis (), Globotruncana orientalis , Globotruncana arca (), Globotruncana linneiana (d'), Globotruncana bulloides , Contusotruncana fornicata (), while others have a sharply defined range and so constitute important stratigraphic points of reference (Fig. 3 ![]() ). Among these marker species may be cited:
). Among these marker species may be cited:
Plates 1 to 5 (Pl. 1 ![]() - Pl. 2
- Pl. 2 ![]() - Pl. 3
- Pl. 3 ![]() - Pl. 4
- Pl. 4 ![]() - Pl. 5
- Pl. 5 ![]() ) figure the species encountered in the whole of the Ellčs section.
) figure the species encountered in the whole of the Ellčs section.
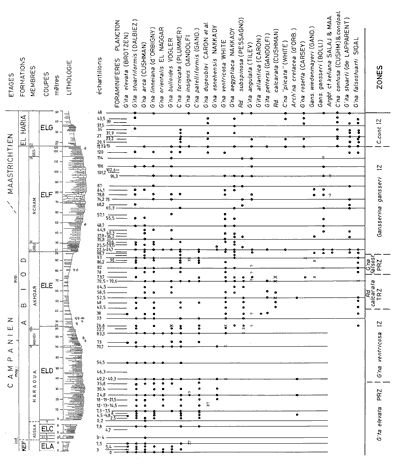
Click on thumbnail to enlarge the image.
Figure 3: Vertical ranges of the principal planktonic foraminifera of the Ellčs section (F.R. & M.M.). Arch'na: Archaeoglobigerina; Argbt: Archaeoglobitruncana; C'na: Contusotruncana; Gans.: Gansserina; G'ita: Globotruncanita; G'na: Globotruncana; Rd.: Radotruncana.
Using the first and last occurrences of marker species, a zonation by planktonic foraminifera can be made that is very close to the one proposed by et alii (2000) for the Kalaat Senan region.
The abbreviations for zones as defined in the et alii, Atlas, 1984.
From bottom to top there are thus:
Remark: it must be made clear that in the Kalaat Senan section ( et alii, 2000), the first Globotruncana ventricosa were already present at the top of the Kef marls, which thus involves a Globotruncana ventricosa Zone that extends farther downward.
To summarize, among the biostratigraphic events described above, if the biohorizons with an international value are to be pointed out, three time-lines with identical positions in the Ellčs and Kalaat Senan sections may be selected:
The location of the Campanian-Maastrichtian boundary (Cp-Ma) in the Abiod Formation has fluctuated considerably in accordance with the views of the successive authors.
Originally for de (1901) working outside of Tunisia, "Turrilites polyplocus" is found in the Campanian, but toward the base of his "upper Campanian" because this author did not recognize the existence of the Maastrichtian.
(1907) considers that the zone with Bostrychoceras polyplocum is in limestones that "correspond to the Upper Campanian beds" but that "several authors place it even in the Maastrichtian".
A little later (1911) restores the B. polyplocum Zone but places it at the base of his "Maestrichtien" (without explaining exactly what criterion he had chosen to support his choice).
In 1941 (p. 270) is the first to make popular the use of "Globotruncana calcarata" as a marker of the "Lower Campanian". Later the taxon would move up to the terminal portion of the Upper Campanian (, 1946; , 1945, 1947; & , 1947; 1952; cf. & de , 1954).
In his thesis (1956) took up again the problem of the location of the Cp-Ma boundary, bringing up 's concept (1911: a Zone of B. polyplocum at the base of the "Maëstrichtien") and that of the micropaleontologists (the Campanian ends with the extinction of "Globotruncana calcarata", that is a little above abundant Bostrychoceras polyplocum).
The concept of the micropaleontologists ("the last Globotruncana calcarata = end of the Campanian") will be adopted by most of the geologists working on North African sections among them (1955), & (1956), (1967, 1977, 1987), (1969), (1979), et alii (1983), et alii (1984), et alii (1990), et alii (1995).
However, starting in 1980 (p. 101) , reviewing the Kef section, places the base of the Maastrichtian not at the extinction of "Globotruncana calcarata" but just above the level that yielded the ammonite Pseudokossmaticeras brandti found at the bottom of the upper bar and at that time considered by as characteristic of a "high Campanian" (, 1974, in litteris cited by , p. 101).
Continuing in this sense, & (1982) bring up the small planktonic foraminifer Rugotruncana kefiana n. sp. (attributed later to to the genus Archaeoglobitruncana by & (1984) which cohabits with Pseudokossmaticeras brandti, the two taxa thus indicative of the "Campanian". At that time they put the Cp-Ma boundary near the top of the upper bar of the Abiod, there where, according to them, the first Globotruncana falsostuarti appear. (1952) cited this species. in his "Maestrichtien" (but according to us is already present lower down in the Akhdar marls).
This positioning of the Cp-Ma boundary at the top or around the middle of the upper bar was restated by (1983) and by & (1984).
Returning to the presence of Pseudokossmaticeras brandti at the bottom of the upper bar, this time considers it a marker of the basal Maastrichtian, referring to & (1976) in & (1989, p. 303). This age determination affects that of Archaeoglobitruncana kefiana -it cohabits with P. brandti- so that it too must indicate a basal Maastrichtian age.
This determination was retained by & (1992) for the Kalaat Senan section where the base of the A. kefiana Zone designates the base of the Maastrichtian and corresponds fairly closely to the base of the upper bar.
Nevertheless, three years later, et alii (1995) support the opinion that the Campanian ends with the upper limit of the "Globotruncana calcarata" Zone, although (1995) in the explanation of the Gafour sheet, puts the Cp-Ma boundary near the top of the lower bar and so attributes to the Maastrichtian the intermediate alternances in which he does not report the presence of G. calcarata.
In the Kalaat Senan area, the Upper Cretaceous was revised by et alii (2000) who place the Cp-Ma boundary in the lower third of the Ncham upper bar, at the articulation between the extinction of the ammonite Nostoceras (Nostoceras) cf. hyatti (uppermost Campanian) and the appearance of the ammonite Nostoceras (Nostoceras) aff. magdadiae & (basal Maastrichtian). In so doing the criteria recommended by the International Subcommission on Cretaceous Stratigraphy ( et alii, 1996) were strictly observed. Remember that Nostoceras magdadiae is a species defined from the base of the Maastrichtian in the Prairie Bluff Chalk of Alabama and Mississippi ( & , 1995). For these two authors, in Europe the base of the Maastrichtian is at the first occurrence of Belemnella lanceolata where this belemnite immediately overlies an assemblage of ammonites with N. hyatti. This indicates that it is the extinction of N. hyatti that constitutes a time line representing the base of the Maastrichtian in the Prairie Bluff Chalk and it is followed immediately by the entry of N. magdadiae.
This limit for the Cp-Ma boundary which corresponds to the top of the Nostoceras hyatti Zone is a little higher than the last Pseudokossmaticeras brandti (this species is manifestly Late Campanian according to the study of & , 1993 and et alii, 2001).
In the Ncham wadi section, P. brandti has been collect along with N. hyatti at the base of the upper bar, in the alternances of the Gourbeuj Member. In addition, at Kalaat Senan N. hyatti has been found higher up - in the Ncham limestones- in the lower third of the upper bar ( et alii, 2000).
This conception of the location of the Cp-Ma boundary was accepted and used by et alii (2002) in their study of the Abiod sequence from El Djebil at El Kef.
In 1995, at the Brussels Symposium dedicated to the limits of the stages of the Cretaceous, the International Subcommission on Cretaceous stratigraphy recommended the choice of the former quarry at Tercis as the location of a GSSP (Global Standard Stratotype, Section and Point) –named PSM (Point Stratigraphique Mondial) by et alii, 2005- for the Campanian-Maastrichtian boundary ( et alii, 1996), adapting a proposal by the Working Group on the Maastrichtian. In February 2001 the decision was ratified by the International Union of Geological Sciences (in & , 2001). At the same time an 881 pages volume was published presenting a multi-disciplined characterization of the Cp-Ma transition at Tercis (edited by with 70 authors, 2001) which was succeeded by several notes summarizing the principal results ( & , 2001; , 2002; cf. , 2002).
The succession at Tercis is interesting for it houses coexistent groups of paleontologic key species: ammonites, planktonic foraminifers, nannofossils, dinoflagellates, etc. in a continuous limestone facies.
Among the biostratigraphic criteria made evident at Tercis those involving ammonites and planktonic foraminifers are referred to here because some of them are found in central Tunisia, cf.
Fig. 4 ![]() , a fact that renders it feasible to make comparisons and correlations between Tercis and the two Tunisian sections at Kalaat Senan and Ellčs:
, a fact that renders it feasible to make comparisons and correlations between Tercis and the two Tunisian sections at Kalaat Senan and Ellčs:
On the other hand, several biohorizons of planktonic foraminifers in the Ellčs section are not proposed as criteria for long distance correlation, as follows.
The biohorizon based on the appearance of Rugoglobigerina scotti at Tercis
& (2001) cite the species and consider it "an excellent index fossil" of proximity to the Campanian-Maastrichtian boundary and propose it as a zonal index. However, several factors militate against the use of this species as an international zonal index although it is usable at a regional level as for example Tercis and the nearby areas of Spain:
The biohorizon based on the appearance of Contusotruncana contusa at Tercis
Here the problem of specific determination lies essentially in the concept of species. Indeed, C. contusa is the final term in an anagenetic evolutionary series in which three morphologic "species" were erected to demarcate this continuum. C. fornicata—>C. patelliformis—>C. contusa (cf. text-fig. 5 in et alii, 1984; , 1988a). In this series the convexity of the spiral side increases gradually from the Santonian to the end of the Maastrichtian, that is for almost 20 Ma. In the species C. fornicata the spire is low to moderately high and the diameter is generally between 0.3 and 0.5 mm; in C. patelliformis the spire becomes moderately high to high and the diameter ranges from 0.5 to 1 mm; and finally in C. contusa the spire is very high with a large number of whorls and an undulated surface, with a very large diameter, commonly between 0.7 and 1.5 mm. As in all anagenetic lineages there are many intermediaries between these "type species", that are particularly numerous between C. patelliformis and C. contusa. Furthermore, the morphotypes evolved from fornicata coexist after the first patelliformis are differentiated and the two forms continue to coexist after the first differentiation of contusa and of other closely related species like C. walfischensis and C. plicata (see phylogenetic table of text-fig. 8 in et alii, 1984). A "profile index" could be established to categorize the morphologic limits of the forms discussed using the Height (H) and the Diameter (D) relationships of the spire, but this would not take into account possible variations in morphology caused by paleobathymetric, paleoecologic and paleo- geographic conditions. Even a method of "grade dating" has been tried in order to establish a "numerical age" using the measurements of such indices (, 1988a, 1988b), a method that has been contested several times because it is "irreconcilable with the paleontologic data and regional observations" and is not capable of giving ages with a precision of around 0.1 Ma, for it did not detect a tectonic redoubling of several Ma in Pyrenean sections (, 1994; et alii, 1999).
Thus, any paleontologist can determine the typical morphotypes of fornicata, patelliformis and contusa ... but he will always have difficulties in naming the intermediate forms associated with the species types: such is the case when a form is to be named that is described qualitatively as an "evolved patelliformis" or a "first contusa".
As an example we have calculated the H/D index of a certain number of morphotypes considered by several authors as representatives of "type species" thus constituting reference points in the anagenetic series fornicata—>patelliformis—>contusa and chosen to be photographed and published. The index is calculated below using measurements taken on specimens figured in the Atlas of planktonic foraminifers of the Upper Cretaceous ( et alii, 1984) and in the plates of & (2001) listed as I. & O.
C. fornicata
| Atlas: | pl. 38, | fig. 1b | H/D = 0.34 |
| fig. 2b | H/D = 0.40 | ||
| fig. 3b | H/D = 0.33 | ||
| fig. 5b | H/D = 0.34 | ||
| mean in the Atlas: 0.35 | |||
| I. & O.: | pl. VI, | fig. 129 | H/D = 0.37 ("fornicata A") |
| fig. 136 | H/D = 0.51 ("fornicata B") |
C. patelliformis
| Atlas: | pl. 39, | fig. 2b | H/D = 0.51 |
| fig. 1b | H/D = 0.55 | ||
| fig. 3b | H/D = 0.58 | ||
| mean in the Atlas: 0.54 | |||
| I. & O.: | pl. VII, | fig. 148 | H/D = 0.57 |
| pl. VI, | fig. 140 | H/D = 0.55 ("fornicata B / contusa") | |
| fig. 137 | H/D = 0.62 ("fornicata B / contusa") |
C. contusa
| Atlas: | pl. 36, | fig. 2b | H/D = 0.55 |
| fig. 1b | H/D = 0.61 | ||
| pl. 37, | fig. 1b | H/D = 0.71 | |
| fig. 2b | H/D = 0.76 | ||
| fig. 3b | H/D = 0.86 | ||
| mean in the Atlas: 0.70 | |||
| I. & O.: | pl. VI, | fig. 144 | H/D = 0.58 |
| pl. VII, | fig. 146 | H/D = 0.59 |
Although the results have no statistical value because the number of specimens is too small, it can be seen, nevertheless, that the authors chose morphotypes that qualitatively are quite typical.
The figures posted above are worthy of commentary:
Consequently, it is understandable that the profile index may give an immediate indication of the convexity of a form but the final determination cannot be made solely on the basis of this index, particularly for C. contusa: other specific criteria must also be taken into account such as size, the number of whorls, the aspect of the chamber surfaces, the elongation of chambers, the presence or absence of sutures in relief between the chambers, etc.
Moreover, to make possible a more complete comparison with the other planktonic elements in the Tercis section, it would have been interesting for the "intermediaries" to have been figured, for example the "patelliformis/contusa" reported by & (2001, Fig. 1, p. 336), or the "contusa" immediately below and above the Campanian-Maastrichtian boundary of & (2001, Fig. 1, p. 342), or again the "fornicata B" and the "patelliformis" near the Campanian-Maastrichtian boundary of & (2001, Fig. 2, p. 353). It would then have been feasible to ascertain that the different authors all had the same concept of contusa.
In no way do these remarks denigrate the value of the Campanian-Maastrichtian boundary as it was conceived: a limit set by many criteria, defined by the arithmetic mean of the position of a dozen biohorizons from six important paleontologic groups. Even if the concepts of species has been somewhat modified by later paleontologic studies, the position of the "Golden Spike" must always set the base of the Maastrichtian at Tercis at level 115.2. It is surely there where the interest of this concept of limits based on many criteria proposed by (2002) - that is, not to rely on the appearance of a "magic" taxon, in this case the occurrence of the ammonite Pachydiscus neubergicus- is most appropriate. The earliest occurrence of this species was noted at level 116.1 by & (2001), but a morphologic continuum exists between P. neubergicus and its ancestor P. perfidus ( & , 1986; in et alii, § 9, 2001), in the same fashion as that linking C. patelliformis and C. contusa, and everyone knows how difficult it is to set a limit in a continuum. It is in this way that after the first comparisons of the Tercis sections with those of the Vistula (, 2004), (2005), repeated in et alii (2007), using his personal criteria was led to consider that "P. neubergicus" might be present at Tercis at level 98.2 (he finds the form figured on Pl. VI, 45 that & in 2001 called P. perfidus/neubergicus to be a "P. neubergicus"). If the limit remains at 115.2 that determination would have P. neubergicus appear in the terminal beds of the Campanian and that N. (N.) hyatti would encroach the basal Maastrichtian. Thus for him, as for all of those favorable to the use of the appearance of one taxon to define the base of a stage, the fact of making the appearance of P. neubergicus older would make obligatory a change in the position of the "Golden Spike", that is to go against the decision of the IUGS to stabilize the limits of the stage. Furthermore, each new collection or each modification in the concept of species would bring changes that would cause a perpetual yoyo. But in any case, before questioning a decision of the IUGS that would have heavy consequences involving the stability of many limits and in particular that of the Campanian-Maastrichtian, would it not be better to review what is essentially the criteria that define a species, particularly where anagenetic evolution is involved, as is the case for perfidus and neubergicus ?. For that review a large team of specialists on this group of fossils gather using for discussion a collection of the species concerned.
The Gansserina gansseri Zone at Tercis
In the standard zonation of planktonic foraminifers ( coord. in et alii, 1998), the first occurrence of the species G. gansseri is reported at the base of the N. hyatti Zone. The G. gansseri Zone extends throughout the upper part of the Campanian and the lower part of the Maastrichtian; the species continues upward almost to the top of the Maastrichtian. This distribution of the species in the upper part of the Campanian is seen in Tunisia where it is controlled by its relationships to the ammonites at both Kalaat Senan and Ellčs.
At Tercis, the first occurrence of the species G. gansseri seems to be younger when the contributions of the several authors who worked on the section are examined.
& (2001) do not report the species to be present (but the sampling is a preliminary study and the few washed samples available indicate that "index species" were rare).
& (2001) do not report the species either.
& (2001) report the species present but 50 meters above the Campanian-Maastrichtian boundary.
Remember that previously et alii (1996) did not note G. gansseri in the Tercis section and that & (1997) cite it only very high in the section, well above the Campanian-Maastrichtian limit as it was later defined in ed. (2001).
The only record of G. gansseri in the vicinity of the Campanian-Maastrichtian boundary in the Tercis quarry appears in the distribution table of planktonic foraminifers in fig. 3 of the note by et alii (1997). The species is listed there at level 112, that is a little below that of the GSSP -or the PSM of et alii, 2005- of the Campanian-Maastrichtian limit which is at level 115.2, then much higher in the vicinity of levels 132 and 158. In any event, in the absence of illustrations or of specimens of the reported presence at level 112 (the number of specimens is not given) and in hopes that this or these figures will be published, it is not possible to consider this first citation of G. gansseri as confirmed.
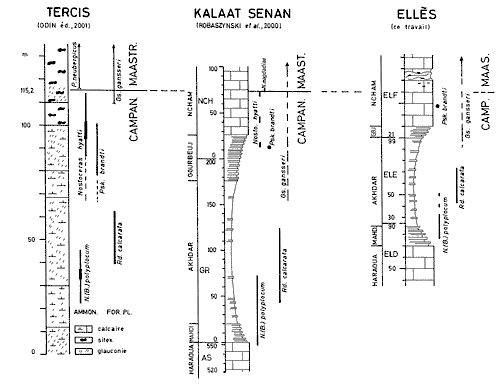
Click on thumbnail to enlarge the image.
Figure 4: Location of the Campanian-Maastrichtian boundary at Ellčs as related to that of the International stratotype of Tercis (Landes, France) and to that of the Kalaat Senan section in Tunisia (F.R.).
Near the village of Kalaat Senan in the sections of wadies Assila, Gourbeuj and Ncham the Abiod Formation is strongly developed with a thickness of nearly 500 m ( et alii, 2000). From the biostratigraphic findings the vertical ranges of the markers found also at the Tercis stratotype can be extracted for comparison. It is this list that appears in Figure 4:
So at Kalaat Senan the Cp-Ma limit is placed where the Campanian ammonite Nostoceras cf. hyatti, is replaced by the Maastrichtian ammonite Nostoceras aff. magdadiae, that is appreciably in the lower third of the Ncham upper bar.
Several biostratigraphic markers of short vertical range were located in the section
(Fig. 4 ![]() ):
):
According to the concepts developed at the Brussels Symposium of 1995 ( & , 1996), the appearance of the ammonite Placenticeras bidorsatum was proposed as a criterion for the base of the Campanian, near the extinction of the crinoid Marsupites testudinarius which in turn is slightly before the appearance of Broinsonia parca and after the appearance of Globotruncanita elevata. As for the top of the Campanian it is coincident with the base of the Maastrichtian, a limit based on multicriteria where the appearance of Pachydiscus neubergicus - near the first occurrence of Belemnella lanceolata takes over as Nostoceras hyatti disappears at the top of the Campanian. A Campanian bounded in this way would have a duration of about 11 Ma ( & , 1990; , 1994; in et alii, 2000; & , 2003), for 12.2 Ma ± 1 (in et alii, 1998) and for ± 12.9 Ma ± 1.3 (stratigraphy.org, chart 2009, where it is indicated as lying between 70.6 Ma ± 0.6 and 83.5 Ma ± 0.7), which averages out as about 12 Ma. In addition, if the division of the stage into three substages (Lower, Middle and Upper) of approximately equal length is accepted ( & , 1996), the length of a substage should be in the neighborhood of 4 Ma.
If the Nostoceras (Bostrychoceras) polyplocum ammonite Zone is assigned the upper part of the Campanian (cf. for example "Upper" with Nostoceras (Bostrychoceras) polyplocum in & , 2001; or in et alii, 2001, fig. 8), it is possible at Ellčs to measure the time elapsed between the biohorizon based on the first occurrence of N. (B.) polyplocum at about ELD 76 (or ELE 12) and the Campanian-Maastrichtian boundary at about ELF 66 (cf. § 4.4). On Table 1 this length of time is the sum of that part of the Mahdi Member above level 76 (11/17 x 0.425 = 0.275 Ma) + the Akhdar Member (2.40 Ma) + the Gourbeuj Member (0.475 Ma) + that part of the Ncham Member below level 66 (1.70 Ma), a total of 4.85 Ma, a value that is more than 4 Ma but is not incompatible with what was anticipated.
Such a proposal adds to a suggestion expressed and supported by (2004) who writes: "The cosmopolitan N. (B.) polyplocum seems to be the ideal index for the definition of the base of the Upper Campanian Substage in Europe and the Russian Platform."
If a closer approach to the 4 Ma value is wanted, instead of selecting the first occurrence of N. (B.) polyplocum as the base of the Campanian, its level of disappearance could be used, that is at about ELD 45 about 0.9 to 1 Ma higher. But that case abandons the conception of an Upper Campanian as generally understood in a large part of the literature concerning Boreal and Tethyan domains. In general these works include the N. (B.) polyplocum Zone in the upper part of the Campanian.
In summary, if recognition of the base of the Upper Campanian is restricted to the first occurrence of Nostoceras (Bostrychoceras) polyplocum such an "Upper Campanian" would have a length of about 4.8 Ma which to begin with would satisfy the proposal for a tripartite division of the stage into substages of approximately equal length and secondly agree with current usage of this biohorizon in literature.
Placement of the boundary between Lower and Middle Campanian remains to be accomplished. If the Middle Campanian is to have a duration of about 4 Ma, at Ellčs that would be represented by 60 to 70 m of Haraoua limestones and the Lower/ Middle contact would be between ELD 10 and 15. It is unfortunate that at Ellčs there is neither an ammonite nor a planktonic foraminifer biohozizon at these levels: the biohorizon of the first occurrence of Globotruncana ventricosa is 30 m higher, at ELD 42, and at Kalaat Senan, the entry of the nannofossil Ceratolithoides aculeus in AS 474 is lower, at the bottom of the upper bar. (AS 474 base of the coccolith Zone CC20 according to in et alii, 2000, p. 431).
Two publications concerning the Cretaceous of Ellčs have been published quite recently, one in 1998 by & , the other in 1999 by et alii. In these two communications based on planktonic foraminifers the Campanian-Maastrichtian boundary is placed at two discrete levels neither one corresponding to that proposed here. The indigence of lithologic columns and the fact that facies are not related to formations makes it difficult to understand clearly some of the biozonal and chronostratigraphic interpretations proposed by the authors. Let us see about these successively in the two publications.
In & (1998), the Campanian-Maastrichtian boundary is set at the top of the "Globotruncanita calcarata" Zone. However, at the very least several elements seem to be expediencies.
In et alii (1999, p. 239) the Campanian-Maastrichtian boundary is placed this time at the appearance of the foraminifer Rugoglobigerina hexacamerata (while referring to the magnetic reversal C32N at the DSPD 525A site, which is several thousands of kilometers distant in the Atlantic of the southern hemisphere) that is well above the base of the marls (El Haria) at the last of "5 beds" as depicted on their fig. 4b. This choice would indicate that all of the upper bar of the Abiod as well as 15 to 20 m of the basal El Haria marls are Campanian (while macropaleontologic data from ammonites clearly support a much lower Cp-Ma boundary).
As for the appearance of Rugoglobigerina hexacamerata, this species was reported from Ellčs as early as 1978 near the base of the El Haria marls (, sample 33B) as "lower Maastrichtian". It was also found at Kalaat Senan at the base of the El Haria marls, and even lower still at the top of the Akhdar marls and in the Gourbeuj alternances ( & in et alii, 2000). Unless intended for use as an acme zone, it does not appear judicious to choose this appearance as a proxy for a stage limit, a limit that in any event is manifestly lower than that proposed by the authors.
In lithologic terms, the Ellčs section reveals that the Abiod Formation is tripartite, bounded below by the Kef marls and above by the El Haria marls. In the countryside it is expressed as two resistant limestone bars separated by a marl-filled depression. The lower bar, about 105 m thick, is made up of the basal Assila alternances, followed by the Haraoua limestones (70 m) and ends with Mahdi alternances. The intermediate unit, the Akhdar Member, is 77 m of characteristically tinted turquoise marls. The upper bar, 104 m thick, begins with the Gourbeuj alternances, followed by the thick-bedded Ncham limestones (85 m) and ends with Gouss alternances.
Several biohorizons, defined by micro- and macrofossils constituting time lines, are excellent tools for correlation.
Among the microfossils, the first occurrences of marker planktonic foraminifers establish a zonation of international scope with the succession from oldest to youngest as follows: IZ of Globotruncanita elevata, IZ of Globotruncana ventricosa, TRZ of Radotruncana calcarata, PRZ of Globotruncana falsostuarti, IZ of Gansserina gansseri, IZ of Contusotruncana contusa. To these must be added the first occurrences of Radotruncana subspinosa (a little before that of R. calcarata), of Globotruncanita angulata (a little before that of G. gansseri) and of Archaeoglobitruncana cf. kefiana (near the base of the upper bar).
Among the macrofossils the ammonites were the prime target for collection and several biohorizons are well defined: the short stratigraphic range of Nostoceras (Bostrychoceras) polyplocum from the middle of the Mahdi alternances to the end of the first quarter of the Akhdar marls; the Nostoceras (N.) cf. hyatti level at the bottom of the upper bar, the beds with Pseudokossmaticeras brandti in the lower quarter of the Ncham limestones; the beds with Pachydiscus (P.) neubergicus in the upper third of the Ncham limestones.
The conjunction in the occurrence of several of these time lines permits a relatively more precise dating of the several lithologic units and particularly a correlation of series in far-distant basins.
Thus, when paleogeographic maps are constructed -for example those that might be established in Tunisia for each of the principal members of the Abiod- the use of well-controlled time lines synchronous with and usable on an international scale would permit well constrained time-times to be drawn with results capable of locating surfaces favorable to exploration for petroleum.
Taking into consideration the several marker biohorizons, the Campanian-Maastrichtian boundary is about 50 m above the first occurrences of G. gansseri and of N. hyatti and 15-20 m above the beds with P. brandti, that is at the top of the first third of the Ncham bar. The position of this limit is compared to and correlated with that established in the Kalaat Senan section (the same markers and appreciably the same vertical ranges of macro- and microfossils) and with that of the Tercis stratotype in France (thanks to the biohorizon markers N. (B.) polyplocum, N. hyatti, P. brandti and R. calcarata).
In Tunisia, combining information derived from lithologic units and biohorizons, the tripartite Abiod at Ellčs constitutes an ensemble nearly 286 thick which is very clearly expressed in the morphology of the terrain. It represents pelagic marine sedimentation that lasted for about 13.2 million years, according to cyclostratigraphic calculations.
The lower bar (Assila + Haraoua + Mahdi), 105 m thick, represents deposition during about 6 Ma with a mean rate of non-compacted sedimentation of 1.7 cm/ky. The Akhdar intermediate marls, 77 m thick, represent a deposition time of about 2.4 Ma, with the deposition of non-compacted sediments at the rate of 3.2 cm/ky. For the upper bar (Gourbeuj + Ncham + Gouss) its 104 m thickness represents deposition for about 4.7 Ma with a mean rate of accumulation of non-compacted sediments of 2.1 cm/ky. The Gourbeuj alternances and the lower third of the Ncham limestones are Upper Campanian while the upper two-thirds of the Ncham limestones are already Lower Maastrichtian.
If a ternary division of the Campanian stage with subdivisions of approximately equal length is accepted, it is proposed that an Upper Campanian be defined of which the base would be taken at the appearance of the ammonite N. (B.) polyplocum and the top, that is the lower limit of the Maastrichtian, would be coincident with the extinction of Nostoceras hyatti. Measured by cyclostratigraphy such an Upper Campanian would have a length of about 4.8 Ma of a total length for the Campanian of about 12 Ma.
Several paleontologic zones indicate the total vertical distribution of several markers and have a global value. These are the TRZ with N. (B.) polyplocum which includes the top of the lower bar and the lower third of the Akhdar marls, about 34 to 36 m and 0.9 to 1 Ma; the TRZ with P. brandti from the Gourbeuj alternances to the lower quarter of the Ncham; the TRZ with N. hyatti which also starts in the Gourbeuj alternances, and continues into the lower third of the Ncham where its extinction marks the Campanian-Maastrichtian boundary; the TRZ with R. calcarata, from the middle of the Akhdar marls where it is known for at least 30 m with a minimum deposition time of about 0.8 Ma.
Taking these contributions into account, a critical reading of two notes published a decade ago was carried out regarding a part of the Ellčs section and in particular the controversial placement of the Campanian-Maastrichtian boundary.
The determination of planktonic foraminifera in thin sections benefited from the vast experience of Madam Michčle who received one of us (M.M.) at the University of Friburg (Switzerland). The generic relationships of ammonites collected from the partial sections were confirmed or defined in the course of discussions with the doctoral candidate Stijn (University of Leuven, Belgium). In the field, geological surveys, sampling and the collecting of macrofauna was completed by the two authors assisted by Mourad and Taoufikh (ETAP) as well as by Moheddine (Kalaat Senan). Thanks to all of them for their assistance. The scanning electron microscope photographs were made by one of us (F.R.) using the apparatus of the ETAP Research Center in Tunis and we thank that company for authorizing its use. The draft benefited from constructive remarks and suggestions by Jochen , by an anonymous reader and by Dr Gilles Serge who participated in clarifying discussions at several stages in the revision of the final text. We thank all three and Nestor for the translation in English.
[La Formation Abiod ŕ Ellčs (Tunisie) :
composition lithologique tripartite,
biohorizons de globotruncanidés et d'ammonites,
durée, position de la limite Campanien-Maastrichtien,
corrélation avec Kalaat Senan et le stratotype de Tercis (France)]
Université de Mons, Faculté Polytechnique, 9 rue de Houdain, 7000 Mons
(Belgique)
ETAP, Entreprise Tunisienne d'Activités Pétroličres, Laboratoire de
Biostratigraphie, Centre de Recherche et de Développement Pétroliers La Charguia II, Tunis
(Tunisie)
Manuscrit en ligne depuis le 27 Août 2010
![]()
En Tunisie centre-nord, prčs d'Ellčs, l'oued Ed Dam expose un enregistrement sédimentaire quasi continu de la Formation Abiod encadrée par la Formation Kef au-dessous et la Formation El Haria au-dessus. Cette Formation Abiod, tripartite, a été étudiée pour servir de référence litho-biostratigraphique régionale sur la bordure sud-est du Sillon tunisien oů elle atteint prčs de 286 m. Le contenu en foraminifčres planctoniques d'une série d'échantillons est reporté sur un tableau de distribution et soutient une division en 6 biozones micropaléontologiques. Les positions des ammonites récoltées sont repérées par rapport ŕ la succession lithologique. Plusieurs biohorizons sont décrits qui correspondent aux premičres et derničres présences de foraminifčres, par exemple de Radotruncana calcarata, et aux émergences des marqueurs de zones comme Globotruncana ventricosa, Gansserina gansseri et Contusotruncana contusa ainsi que les premičres et/ou les derničres présences de plusieurs ammonites marqueurs comme Nostoceras (Bostrychoceras) polyplocum, Pseudokossmaticeras brandti et Nostoceras (Nostoceras) hyatti.
L'un des points d'ancrage de la succession stratigraphique étant la limite Campanien-Maastrichtien (Cp-Ma), on définira la position de cette ligne-temps ŕ Ellčs par comparaison avec la coupe de Kalaat Senan et celle du stratotype international de Tercis (France) ratifiée par l'Union internationale des sciences géologiques.
À Kalaat Senan, la limite Cp-Ma est placée au tiers inférieur de la barre supérieure de l'Abiod ou Membre Ncham, au sommet de la Zone ŕ N. (N.) hyatti, contenant P. brandti, ammonites de la partie supérieure du Campanien, et ŕ l'apparition de Nostoceras magdadiae, ammonite de la partie inférieure du Maastrichtien. À Ellčs, plusieurs de ces marqueurs ont été retrouvés comme aussi les zones de foraminifčres planctoniques ŕ Radotruncana calcarata, ŕ Globotruncana falsostuarti, ŕ Gansserina gansseri et ŕ Contusotruncana contusa.
En utilisant l'outil cyclostratigraphique, la durée des diverses unités lithologiques et celle de quelques zones paléontologiques sont précisées : la durée totale de dépôt de la Formation Abiod ŕ Ellčs est mesurée ŕ 13,2 Ma, celle de la Zone ŕ R. calcarata ŕ 0,79 Ma et celle de la Zone ŕ N. (B.) polyplocum ŕ 0,97 Ma. Une limitation du Campanien supérieur ŕ la base de la Zone d'ammonite ŕ N. (Bostrychoceras) polyplocum est proposée.
La position de la limite Cp-Ma proposée par et alii en 1999 au sommet d'une série de bancs calcaires dans la Formation El Haria est discutée.
F. & M. (2010).- L'Abiod d'Ellčs (Tunisie) : stratigraphies, limite Campanien-Maastrichtien et corrélation [The Abiod at Ellčs (Tunisia): stratigraphies, Campanian-Maastrichtian boundary, correlation].- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/04 (CG2010_A04)
Campanien-Maastrichtien ; Formation Abiod ; Tunisie centrale ; Ellčs ; Tercis ; Kalaat Senan ; globotruncanidae ; ammonites.
The Abiod at Ellčs (Tunisia): stratigraphies, Campanian-Maastrichtian boundary, correlation.- In central Tunisia near the village of Ellčs is Wadi Ed Dam. There the Abiod Formation, underlain by the Kef marls and capped by the El Haria marls, crops out in an almost uninterrupted exposure about 286 m thick. In view of its possible use as a regional litho- and bio- stratigraphic reference section for the southern border of the Tunisian trough it is examined in detail here. Fossils studied include foraminifera (115 samples representing 6 micropaleontologic zones, see chart) and ammonites. The biohorizons established are based on the first (FO) or last occurrence (LO) of certain species of planktonic foraminifers in addition to the FO of already established foraminiferal zonal markers. Ammonite markers found include both the FO and LO of some forms, or just one or the other.
The location of the Campanian-Maastrichtian (Cp-Ma) boundary time line at Ellčs is established by comparison with that determined at Kalaat-Senan and with that of the international stratotype of Tercis (France), ratified by the International Union of Geological Sciences.
At Kalaat-Senan the Cp-Ma boundary is set at the lower third of the upper indurated limestone bar of the Abiod Formation - the Ncham Member. Its location at Kalaat-Senan is based on the upper limit of occurrence of Nostoceras (Nostoceras) hyatti and Pseudokossmaticeras brandti together with the FO of Nostoceras magdadiae, a Maastrichtian ammonite. At Ellčs several of these same markers are present, along with the respective planktonic foraminiferal zones.
The length in years of some lithologic units and paleontologic zones in the Upper Cretaceous succession of wadi Ed Dam was calculated using cyclostratigraphy. The Abiod Formation represents 13.2 Ma, the Radotruncana calcarata Zone 0.79 Ma, and the Nostoceras (Bostrychoceras) polyplocum Zone 0.97 Ma. A proposed subdivision of the Upper Campanian is recommended at the base of the Bostrychoceras Zone.
et alii's 1999 proposal for fixing the location of the Cp-Ma boundary at the top of a limestone bed in the El Haria marls is reviewed.
Campanian-Maastrichtian; Abiod Formation; Central Tunisia; Ellčs; Tercis; Kalaat Senan; globotruncanids; ammonites.
En Tunisie centre-nord, la Formation Abiod marque trčs bien la partie élevée du Crétacé supérieur en organisant les paysages sous forme de doubles collines séparées par une partie médiane en légčre dépression. Cette morphologie correspond ŕ la composition généralement tripartite de la Formation avec une barre inférieure de calcaires blancs, des marnes intermédiaires turquoise et une barre supérieure de calcaires blancs. L'ensemble est d'âge Campanien-Maastrichtien et recčle donc la limite entre ces deux étages géologiques. Suivant des critčres qui ont varié au cours du dernier sičcle, cette limite a d'abord été située vers le sommet de la barre inférieure, puis vers le milieu des marnes intermédiaires, ou encore ŕ la base de la barre supérieure puis ŕ son sommet. Ces fluctuations étaient dues ŕ ce qu'un accord international n'existait pas sur la position de la frontičre entre les deux étages.
Aujourd'hui, aprčs avoir été approuvée par le Groupe de travail sur le Maastrichtien en septembre 1999, recommandée par la Sous-Commission internationale de stratigraphie du Crétacé en juin 2000, adoptée par la Commission internationale de stratigraphie en décembre 2000 et ratifiée par l'Union internationale des Sciences géologiques en février 2001 (in & , 2001), la coupe de Tercis (Landes, France) contient le stratotype international de la limite multicritčre entre le Campanien et le Maastrichtien (, 2002).
Par comparaison avec cette coupe, en utilisant plusieurs index biostratigraphiques on peut maintenant repérer plus précisément la limite Campanien-Maastrichtien en Tunisie centre-nord et la placer au tiers inférieur de la barre calcaire supérieure de la Formation Abiod.
Cette position est bien définie dans les régions oů l'enregistrement sédimentaire de la Formation Abiod est le plus continu possible, ce qui est le cas de plusieurs coupes situées en Tunisie centre-nord ("Sillon tunisien" de et alii, 1952). De bons exemples se trouvent localisés ŕ El Kef (, 1956) oů, les macrofossiles étant relativement rares, l'accent avait été mis sur la répartition des microfossiles, en particulier les foraminifčres planctoniques (, 1955, 1956), et aussi ŕ Kalaat Senan oů les ammonites semblent plus nombreuses en plus des foraminifčres planctoniques qui sont également abondants ( et alii, 2000).
En dehors de la Tunisie centre-nord, la Formation Abiod ne se présente plus forcément sous l'aspect tripartite classique : parfois c'est la barre inférieure qui est réduite, parfois la barre supérieure, ou, dans d'autres cas, la Formation présente une succession quasi monolithique comme dans certaines coupes au nord de Kairouan ou dans des sondages de la plaine au sud-est de Kairouan. Afin de dater plus précisément ces différents types d'Abiod, on a d'abord étudié une coupe ŕ sédimentation apparemment continue située sur la bordure sud-est du "Sillon tunisien" : c'est la coupe d'Ellčs, ŕ 45 km environ au sud-est d'El Kef. Lŕ, comme ŕ Kalaat Senan, les bancs de calcaires et de calcaires marneux contiennent des macrofossiles, dont des ammonites, ainsi que des foraminifčres planctoniques visibles en plaques minces, tandis que les marnes livrent en dégagé des foraminifčres planctoniques ŕ profusion. La distribution verticale des espčces planctoniques et la zonation qui en découle constitueront la base pour les attributions stratigraphiques dans des successions oů la Formation Abiod est marquée par de fortes réductions d'épaisseur, par des lacunes ou par une uniformisation de la lithologie vers un facičs majoritairement carbonaté. Par la suite, des attributions plus précises permettront de réaliser des cartes paléogéographiques isochrones pouvant servir ŕ localiser des événements lithologiques ou structuraux, favorables ŕ un piégeage de fluides tels les hydrocarbures.
Dans ce qui suit, on abordera successivement :
Dans une conclusion, on fera ressortir la valeur de plusieurs biohorizons macro- et micropaléontologiques qui constituent des lignes-temps utilisables pour relier des coupes ŕ l'affleurement au moins pour la Tunisie centrale ainsi que leur intéręt pétrolier dans la corrélation des sondages de recherche.
Dans la coupe d'Ellčs, la limite Crétacé-Paléogčne a été étudiée par Rakia (devenue Mme ) et présentée dans sa thčse en 1978. À cette occasion, elle avait analysé les alternances calcaires-marnes du sommet de la barre supérieure de la Formation Abiod ainsi que les marnes de la base de la Formation El Haria. Elle avait noté la présence de foraminifčres planctoniques dans le haut de la barre supérieure (sommet de notre Membre Ncham) et dans les alternances supérieures (notre Membre Gouss) parmi lesquels on retient les espčces : "Globotruncana stuarti, falsostuarti, gagnebini (= aegyptiaca), arca, contusa-patelliformis, fornicata, linneiana, ventricosa et Rugoglobigerina hexacamerata", association de son "Maastrichtien inférieur" (interprétation toujours valable aujourd'hui, au moins en partie) et, plus haut, dans son "Maastrichtien supérieur", en plus de ces espčces, la présence de "Globotruncana gansseri, G. contusa-galeoidis, Globotruncanella havanensis et Trinitella scotti puis d'Abathomphalus mayaroensis". À noter un élément qui nous servira par la suite au § 4.6 : aucune "Globotruncana calcarata" n'est signalée dans les marnes de la Formation El Haria.
Dans la notice de la feuille géologique au 1 : 50 000 d'Ebba Ksour, (1979) décrit succinctement un bon affleurement de la Formation Abiod dans le secteur dénommé El Magoula (ou El Mazoula, dans sa p. 11). Il s'agit certainement de la succession que nous avons levée de façon détaillée. Il la considčre comme représentative et pouvant "servir d'exemple" de la Formation Abiod pour le nord-est de la feuille. Pour lui, la puissance de l'ensemble de la Formation est évaluée ŕ 235 m tandis que nos mesures avoisinent 286 m. L'auteur avait parfaitement observé sa composition tripartite et avait bien noté la présence d'ammonites du genre Bostrychoceras vers le sommet de la barre inférieure ainsi que la présence de "Globotruncana calcarata" dans le "membre marneux moyen". Quant ŕ l'interprétation chronostratigraphique, celle-ci était fondée sur les déterminations micropaléontologiques de A.-L. et de P. du Service géologique de Tunis, qui attribuent au "Campanien supérieur" la barre inférieure, au "Campanien supérieur (partie supérieure)" le "membre marneux moyen" et au "Maastrichtien inférieur" la barre supérieure. Ce qui, comme on le verra plus loin, correspond en partie ŕ notre interprétation, ŕ la différence prčs que nous plaçons la limite Campanien-Maastrichtien au sein męme de la barre supérieure sur base de récoltes d'ammonites.
Il faut également signaler les publications plus récentes de & (1998) et de et alii (1999) oů les auteurs affirment que, dans la coupe d'Ellčs -correspondant exactement ŕ celle qui est présentée ci-aprčs- le sommet de la Zone ŕ "Globotruncanita calcarata" (représentant pour eux la limite Campanien-Maastrichtien) se trouve vers le sommet des "limestones" devant représenter la barre supérieure de l'Abiod, et placent, plus tard, cette limite dans les "grey marly shales" ŕ "limestones layers" c'est ŕ dire probablement au sein des marnes El Haria, interprétations contestables que nous discuterons plus loin au paragraphe 4.6.
Cliquer sur la vignette pour agrandir l'image.
Figure 1: Situation géographique de la coupe de la Formation Abiod d'Ellčs (F.R.).
À 2 km environ ŕ l'ouest du village d'Ellčs, l'oued Ed Dam offre une coupe quasi continue de la Formation
Abiod, reposant sur la Formation des marnes Kef et supportant la Formation des marnes El Haria complčtement exposée elle aussi (Fig. 1 ![]() ). Le métrage de la section a été effectué au moyen d'un bâton de Jacob de 1,50 m muni d'une équerre de visée permettant des mesures d'épaisseurs ŕ quelques pour-cent
prčs. Des marnes et des calcaires ont été réguličrement prélevés pour l'observation de foraminifčres dégagés et en plaques minces. Au total, 115 échantillons ont été collectés sur les 395 m des coupes partielles levées dont environ 286 m pour la Formation
Abiod. Pendant et aprčs l'examen lithologique de la succession, des macrofossiles ont été recherchés et plusieurs d'entre
eux, particuličrement significatifs seront signalés.
). Le métrage de la section a été effectué au moyen d'un bâton de Jacob de 1,50 m muni d'une équerre de visée permettant des mesures d'épaisseurs ŕ quelques pour-cent
prčs. Des marnes et des calcaires ont été réguličrement prélevés pour l'observation de foraminifčres dégagés et en plaques minces. Au total, 115 échantillons ont été collectés sur les 395 m des coupes partielles levées dont environ 286 m pour la Formation
Abiod. Pendant et aprčs l'examen lithologique de la succession, des macrofossiles ont été recherchés et plusieurs d'entre
eux, particuličrement significatifs seront signalés.
Remarques : parmi les macrofossiles, on insistera plus spécialement sur les ammonites que nous avons récoltées au cours des levés lithologiques et qui, dans certains niveaux, ne sont pas rares ŕ Ellčs. Elles fourniront, en partie, des arguments positifs dans l'établissement de corrélations avec la coupe de Kalaat Senan située ŕ une soixantaine de km au sud-ouest d'Ellčs. Toutefois, il faut signaler que la coupe d'Ellčs a fait l'objet de récoltes ammonitologiques beaucoup plus importantes et plus systématiques de la part du doctorant Stijn (KUL, Leuven, Belgique) dont les résultats feront partie intégrante de sa thčse en cours sur les ammonites du Campanien-Maastrichtien de Tunisie qui comportera de nombreuses planches illustrant les espčces rencontrées.
En outre, quelques inocérames, également récoltés au cours des levés, ont été soumis ŕ feue Annie (Institut royal des Sciences naturelles de Bruxelles) et les quelques espčces citées ici rapportent des déterminations dont elle nous a fait part oralement avant sa disparition le 1er septembre 2006.
D'autres macrofaunes ont été repérées lors des levés (quelques échinides, des fragments de lamellibranches et de brachiopodes) mais leur faible nombre et leur mauvais état de préservation font qu'elles n'ont pas encore été proposées ŕ l'examen des spécialistes.
La description lithologique a bénéficié des travaux menés antérieurement par et alii (2000) dans la région de Kalaat Senan oů la Formation Abiod a été subdivisée en sept membres soit, du haut vers le bas :
| Formations | Membres | Dénominations antérieures |
| Fm. El Haria : marnes | ||
| Formation Abiod | Mb. Gouss : alternances supérieures | barre supérieure |
| Mb. Ncham : barre calcaire | ||
| Mb. Gourbeuj : alternances | ||
| Mb. Akhdar : marnes intermédiaires | marnes moyennes | |
| Mb. Mahdi : alternances | barre inférieure | |
| Mb. Haraoua : barre calcaire | ||
| Mb. Assila : alternances de base | ||
| Fm. Kef : marnes | ||
Dans la littérature, la "barre inférieure" correspond aux
membres Assila et Haraoua auxquels est souvent joint le Membre Mahdi, tandis que la "barre supérieure" correspond aux membres Ncham et Gouss (avec souvent le Membre
Gourbeuj). Suivant les auteurs, les "marnes moyennes" réfčrent au Membre Akhdar (et comprennent
parfois, mais rarement - ou pro parte -, les alternances des membres Mahdi et
Gourbeuj). Sept coupes partielles ont été décrites et nommées ELA ŕ ELG. Les trois premičres n'ont pas de parties communes tandis que les quatre suivantes ont toutes des parties chevauchantes qui, de visu sur le terrain, permettent des raccords faciles pour établir une succession lithologique continue (Fig. 2 ![]() ).
).
Cliquer sur la vignette pour agrandir l'image.
Figure 2: Succession lithologique synthétique ŕ partir de coupes partielles de la Formation Abiod ŕ Ellčs (F.R.).
Formation Kef (nommée par ,
1978). Coupe ELA (Fig. 2 ![]() )
: marnes ŕ bancs calcaires.
)
: marnes ŕ bancs calcaires.
Vues sur 9 m en rive gauche de la partie basse de l'oued Ed Dam, ce sont des marnes gris verdâtre, subhorizontales, admettant un banc calcaire de 60 cm ŕ la base et, plus haut, plusieurs bancs de calcaire marneux de 10 ŕ 30 cm.
Formation Abiod (nommée par et alii,
1954). Coupes partielles ELB, ELC, ELD, ELE, ELF et ELG pro parte (Fig. 2 ![]() )
: deux barres calcaires limitées ŕ leur base et ŕ leur sommet par des alternances encadrent des marnes
intermédiaires. On y distingue plusieurs membres selon et alii
(2000).
)
: deux barres calcaires limitées ŕ leur base et ŕ leur sommet par des alternances encadrent des marnes
intermédiaires. On y distingue plusieurs membres selon et alii
(2000).
Membre Assila (coupes ELB et ELC) : alternances de base, environ 13 m accessibles, 17 ŕ 20 m probables.
Ce sont les premiers bancs calcaires visibles ŕ l'aval de l'oued Ed Dam, au-dessus des marnes Kef. La continuité stratigraphique visuelle n'est pas assurée entre les trois coupes partielles ELA, ELB et ELC. Toutefois, les pendages étant trčs faibles, on peut admettre (si l'on exclut l'existence de failles importantes par le fait que les calcaires ne sont pas anormalement fracturés et que la succession n'est pas incohérente) que les lacunes de visibilité ne dépassent pas quelques mčtres. Il s'agit de calcaires blanc-gris, tenaces, ŕ cassure conchoďdale, avec des interbancs de calcaire marneux décimétriques.
Membre Haraoua (coupe ELD, de 0 ŕ 70 m) : barre calcaire inférieure de 70 m.
La coupe ELD commence dans l'oued Ed Dam au croisement avec un chemin qui le traverse. À partir de lŕ, la succession est quasiment ininterrompue jusqu'ŕ la Formation El Haria. Le pendage vers le sud-est augmente réguličrement pour atteindre 10° environ et localement parfois un peu plus, jusqu'ŕ 12°.
Le Membre Haraoua apparaît dans le paysage comme une bande de collines formée de plusieurs dizaines de mčtres de calcaires en bancs de 20 ŕ 90 cm d'épaisseur, plus ou moins amalgamés et ŕ stratification souvent un peu onduleuse, séparés par des interbancs de calcaire marneux de 10 cm ŕ 20-30 cm. Les calcaires, blanc-gris ŕ blanc, compacts, ŕ cassure conchoďdale, montrent en plaque mince de nombreux foraminifčres en texture wackestone. Des empreintes d'inocérames de 10 ŕ 20 cm de diamčtre ainsi que des trace-fossiles du groupe des Zoophycos n'y sont pas rares.
Membre Mahdi (coupe ELD, de 70 ŕ 87 m) : alternances sur 17 m.
Les gros bancs calcaires du Membre Haraoua passent progressivement ŕ des bancs calcaires de 20 ŕ 30 cm, déjŕ un peu plus marneux et plus gris, alternant avec des bancs de calcaire marneux gris de 20 ŕ 50 cm généralement altérés et en creux. La premičre présence de l'ammonite Nostoceras (Bostrychoceras) polyplocum est repérée ŕ ELD 76 et deux niveaux sont notés puis suivis latéralement, en particulier celui ŕ 79,40 m.
Membre Akhdar (coupe ELE, de 22 ŕ 99 m) : marnes vert-bleu turquoise sur 77 m.
Progressivement, le caractčre argileux des interbancs marneux augmente comme aussi l'épaisseur des interbancs qui atteint 50 ŕ 80 cm et sépare les bancs carbonatés devenant eux-męmes plus marneux. Plusieurs niveaux livrent encore des fragments d'ammonites N. (B.) polyplocum, Nostoceras (Didymoceras) sp. et Pachydiscus (P.) sp. entre ELE 26,80 et ELE 45,00 avec de trčs nombreux exemplaires ŕ la cote ELE 32,70 m située prčs de la route goudronnée. Le niveau de la derničre présence de N. (B.) polyplocum se trouve vers ELE 45,00.
À partir du mčtre 40 jusqu'au mčtre 80, les passées marneuses s'épaississent pour atteindre plusieurs mčtres. Au-dessus de ELE 80, les bancs calcaires redeviennent plus fréquents. Plusieurs ammonites, dont Pachydiscus sp., sont encore récoltées ŕ ELE 80,50 et 81,00. À noter que le foraminifčre planctonique Radotruncana calcarata est présent au moins entre ELE 43,50 et 73,70 m soit sur 30 mčtres d'épaisseur au minimum.
Membre Gourbeuj (coupe ELF, de 21 ŕ 30 m) : alternances sur 9 m.
La succession commence par un gros banc de calcaire blanc et compact, de 65 cm d'épaisseur, suivi de 18 ŕ 19 bancs de calcaires de 20 ŕ 50 cm séparés par des interbancs d'abord de 30 cm puis de 10 cm et jusqu'ŕ 5 cm d'épaisseur. L'ensemble est traversé en cascade par l'oued Ed Dam. Un fragment d'ammonite appartenant au genre Pseudokossmaticeras a été récolté ŕ la base du Membre et un exemplaire, non en place, entre ELF 13 et 25. Sous réserve, un autre fragment situé ŕ la base du Membre Gourbeuj et dont la détermination demande confirmation, correspondrait ŕ Nostoceras (Nostoceras) cf. hyatti (comm. pers., Stijn , inédit, 2010).
Membre Ncham (coupe ELF, de 30 ŕ 115 m) : barre calcaire supérieure.
Les calcaires présentent quatre aspects, successivement du bas vers le haut:
En plaques minces, tous ces calcaires montrent un fond micritique oů sont conservés en texture wackestone de nombreux foraminifčres planctoniques et quelques formes benthiques ainsi que des bioclastes divers.
Membre Gouss (coupes ELF de 115 ŕ 125 m et ELG de 6 ŕ 17,20 m) : alternances supérieures.
Les interbancs deviennent plus épais et plus marneux, ce qui caractérise le Membre Gouss. À la base, on note encore un exemplaire de P. neubergicus tandis que, vers le sommet de ELF, une empreinte d'ammonite de 20 cm de diamčtre, assez usée, est trouvée dans les bancs érodés par l'oued ŕ ELF 122,40 m et correspond ŕ un Pachydiscus sp.
À 300 m environ ŕ l'est de la coupe ELF, l'oued El Kerma offre une section trčs bien exposée des deux aspects supérieurs de la barre Ncham (c'est-ŕ-dire environ 10 m de calcaires montrant des glissements synsédimentaires avec des silex et environ 35 m de gros bancs métriques de calcaires sans silex) surmontés des alternances Gouss qui, elles, passent ŕ la Formation El Haria. La partie supérieure du dernier banc calcaire au sommet du Membre Gouss est bien dégagée grâce au déblaiement par l'érosion des premičres marnes El Haria sus-jacentes. Ce dernier banc constitue une belle surface structurale qui expose des empreintes d'inocérames et de Zoophycos ainsi que diverses bioturbations. En revanche, le sommet de ce banc ne présente aucune recristallisation, ni perforation, ni glauconitisation ou phosphatisation qui pourraient le faire interpréter comme un véritable hardground (comme l'ont malencontreusement qualifié et alii, 1998).
Formation El Haria (nommée par ,
1956). Coupe ELG, de 17,20 m ŕ 50,00 m et
au-delŕ (cf.
Fig. 2 ![]() ) : marnes grises avec quelques bancs
carbonatés.
) : marnes grises avec quelques bancs
carbonatés.
Pour résumer, plusieurs biohorizons ŕ valeur de lignes-temps auront une grande importance pour effectuer des corrélations régionales ou ŕ plus grande échelle avec les ammonites :
Il a été montré dans une publication récente que la méthode cyclostratigraphique constituait un outil digne de confiance de la mesure du temps de dépôt d'une unité litho- ou biostratigraphique dans le cas d'une sédimentation rythmique comme l'est celle de l'Abiod ( et alii, 2009). Nous verrons successivement les trois ensembles : barre supérieure, marnes intermédiaires et barre inférieure. Les valeurs présentées sont issues de la fig. 8 in et alii (2009). Elles sont résumées dans le tableau 1 oů T100 représente le temps déduit des cycles d'excentricité de ± 100 000 ans et T20 le temps déduit des cycles de précession de ± 20 000 ans. La ligne "66" indique la position la plus probable de la limite Campanien-Maastrichtien et la ligne "76" indique le niveau d'émergence de Nostoceras (Bostrychoceras) polyplocum.
| unités lithologiques | T100 | T20 | (T100+T20)/2 | durée | durée | |||
| lu | lu | calculé | calculée | admise | ||||
| épaisseur | (Ma) | (Ma) | (Ma) | (Ma) | (Ma) | |||
| index | ELHARIA | |||||||
| 125 | 2,65 | 2,55 | 2,60±0,05 | |||||
| 10m | GOUSS | 0,325±0,025 | 0,30 à 0,35 | |||||
| 115 | 2,35 | 2,20 | 2,275±0,075 | |||||
| NCHAM | 2,275±0,075 | 2,20 à 2,35 | ||||||
| 85m | 66 | 0 | 0 | 0 | ||||
| 1,70±0,10 | 1,60 à 1,80 | |||||||
| 30 | -1,80 | -1,60 | -1,70±0,10 | |||||
| 9m | GOURBEUJ | 0,575±0,012 | 0,56 à 0,58 | |||||
| 21/99 | -2,30 | -2,05 | -2,175±0,125 | |||||
| 77m | AKHDAR | 2,40±0,05 | 2,35 à 2,45 | |||||
| 87/22 | -4,65 | -4,50 | -4,475±0,075 | |||||
| MAHDI | 0,275 | |||||||
| 17m | 76 | 0,425±0,175 | 0,25 à 0,60 | |||||
| 0,15 | ||||||||
| 70 | -5,25 | -4,75 | -5,00±0,25 | |||||
| 70m | HARAOUA | 5,175±0,50 | 4,67 à 5,67 | |||||
| 0 | -10,90 | -9,45 | -10,175±0,75 | |||||
| (17m) | ASSILA | (0,425±0,15) | (0,27 à 0,57) | |||||
| (-11,50) | (-9,70) | (-10,60±0,90) | ||||||
| KEF | ||||||||
Tableau 1 : Durées des membres de la Formation Abiod ŕ partir de l'analyse cyclostratigraphique des alternances qui les composent (selon et alii, 2009). Colonnes T100 et T20 : position des limites des membres dans l'échelle du temps, le 0 étant posé ŕ la limite Campanien-Maastrichtien. Le temps est compté en positif vers le haut et en négatif vers le bas (F.R.)
Remarque : dans le tableau 1, les valeurs de T100 et de T20 reprises dans leurs colonnes respectives proviennent de la lecture du graphique de la fig. 8 de et alii (2009) et apparaissent avec 2 chiffres significatifs aprčs la virgule. Dans les 2 colonnes suivantes les valeurs moyennes (T100 + T20)/2 et les durées de dépôt des membres qui en sont déduites sont issues d'un simple calcul qui amčne ŕ avoir parfois 3 chiffres aprčs la virgule, le dernier n'étant évidemment pas significatif. La derničre colonne affiche une durée de dépôt vraisemblable qui peut ętre admise pour chacun des membres de la Formation Abiod en tenant compte de l'incertitude entre les durées issues de T100 et de T20.
Barre supérieure, durées moyennées (T100+T20)/2
Membre Gouss (10 m) : 0,325 Ma
Membre Ncham (85 m) : 3,975 Ma
Membre Gourbeuj (9 m) : 0,475 Ma
soit une durée moyenne totale de 4,775 Ma pour les 104 m de la barre calcaire supérieure, ce qui représente un taux moyen de sédimentation non décompactée de 10400 cm : 4775 ka = 2,17 cm/ka (ka = kilo-années)
Marnes intermédiaires
Membre Akhdar (77 m) : 2,40 Ma
ce qui représente un taux moyen de sédimentation non décompactée de 7700 cm : 2400 ka = 3,20 cm/ka
Barre inférieure
Membre Mahdi (17 m) : 0,425 Ma
Membre Haraoua (70 m) : 5,175 Ma
Membre Assila ("18 m") : 0,425 Ma
(en réalité le Mb. Assila n'a pas été pris en compte dans les relevés des alternances par le fait d'une mauvaise exposition au fond de l'oued Ed Dam ; on a admis ici que son allure et son épaisseur étant apparemment voisines de celles du Mb. Mahdi, sa durée pouvait également ętre voisine de celle de ce dernier). De cette façon, une durée moyenne totale de 6,025 Ma (soit 5,20 ŕ 6,85 Ma en tenant compte des incertitudes) pourrait ętre acceptée pour le dépôt des calcaires de la barre inférieure, soit un taux moyen de sédimentation non décompactée de 10500 cm : 6025 ka = 1,74 cm/ka.
Au total, le temps de dépôt de la Formation Abiod comprenant barre inférieure, marnes intermédiaires et barre supérieure avoisine les 13,2 Ma pour 286 mčtres de couches.
Zone ŕ Radotruncana calcarata
En retournant ŕ la fig. 8 in et alii (2009), on trouve pour calcarata :
T100 : 3,90 – 3,30 = 0,60 Ma
T20 : 3,60 – 2,65 = 0,95 Ma
soit une moyenne de 0,77 Ma pour la durée minimale de la zone.
Toutefois, en reprenant le tableau des calculs de base (non publié) ayant servi ŕ élaborer cette fig.8, on relčve pour T100 une durée de 765 ka et pour T20 de 830 ka, ce qui comprend la valeur obtenue ŕ partir de la fig. 8, aux approximations de lecture prčs et donne cette fois une valeur moyenne de 0,79 Ma. C'est celle que nous utiliserons.
Zone ŕ Nostoceras (Bostrychoceras) polyplocum
De nombreux spécimens du groupe polyplocum ont été repérés dans les coupes partielles ELD et ELE avec une distribution allant dans nos levés de ELD 76 ŕ ELE 45. En partant de la fig.8, ou mieux, ŕ partir du tableau de calcul, on note pour T100 une durée de 988 ka et pour T20 de 964 ka, soit une moyenne de 976 ka ou 0,97 Ma.
Dans les sections ELA ŕ ELG, 115 échantillons ont été prélevés sur 395 m de coupes partielles levées représentant une suite synthétique de 323 m dont 281 m d'Abiod échantillonnés. Sachant qu'il existe quelques lacunes de visibilité ŕ la base, on peut estimer l'épaisseur totale de la Formation Abiod ŕ environ 286 m. Sur l'ensemble des échantillons, 82 étaient suffisamment marneux pour subir un traitement par lavage précédé d'un trempage ŕ l'eau oxygénée afin d'assurer le dégagement des microfossiles. Les autres prélčvements de calcaires ou de calcaires marneux ont suivi la filičre de préparation en plaques minces ou de traitement ŕ l'acide acétique. Si l'on regroupe les informations fournies par les échantillons des parties chevauchantes des coupes partielles, les spécimens dégagés et en plaques minces se répartissent sur 86 niveaux d'échantillonnage, tous indexés dans la
Fig. 3 ![]() .
.
Les déterminations du matériel dégagé, réalisées par M.M. et F.R., suivent les figurations et les critčres exposés dans l'Atlas des foraminifčres planctoniques du Crétacé supérieur ( et alii, 1984). La préparation des spécimens et leur photographie au Microscope électronique ŕ balayage de l'ETAP ŕ Tunis ont été effectuées par F.R.
Pour toute la section, 28 taxons principaux ont été répertoriés dont 6 trčs rares ou ŕ détermination incertaine. Certaines espčces sont réparties sur toute la succession comme Globotruncanita stuartiformis (), Globotruncana orientalis , Globotruncana arca (), Globotruncana linneiana (d'), Globotruncana bulloides , Contusotruncana fornicata (), tandis que d'autres ont leur premičre et/ou leur derničre présence bien localisées et constituent des repčres biostratigraphiques importants (Fig. 3 ![]() ). Parmi ces espčces marqueurs on peut citer :
). Parmi ces espčces marqueurs on peut citer :
Les planches 1 ŕ 5 (Pl. 1 ![]() - Pl. 2
- Pl. 2 ![]() - Pl. 3
- Pl. 3 ![]() - Pl. 4
- Pl. 4 ![]() - Pl. 5
- Pl. 5 ![]() ) figurent des espčces rencontrées dans l'ensemble de la coupe d'Ellčs.
) figurent des espčces rencontrées dans l'ensemble de la coupe d'Ellčs.
Cliquer sur la vignette pour agrandir l'image.
Figure 3: Distribution verticale des principaux foraminifčres planctoniques de la coupe d'Ellčs (F.R. & M.M.). Arch'na : Archaeoglobigerina ; Argbt : Archaeoglobitruncana ; C'na : Contusotruncana ; Gans. : Gansserina ; G'ita : Globotruncanita ; G'na : Globotruncana ; Rd. : Radotruncana.
En utilisant les premičres et derničres présences des espčces marqueurs, on est amené ŕ élaborer une zonation avec les foraminifčres planctoniques qui est trčs proche de celle proposée par et alii (2000) pour la région de Kalaat Senan.
Zones telles que définies dans l'Atlas de et alii, 1984 :
Du bas vers le haut on a ainsi :
Remarque : il faut relever que dans la coupe de Kalaat Senan ( et alii, 2000), les premičres Globotruncana ventricosa étaient déjŕ présentes au sommet des marnes Kef, ce qui entraînait une Zone ŕ Globotruncana ventricosa plus développée vers le bas.
Pour résumer, parmi les événements biostratigraphiques décrits ci-dessus, si l'on veut pointer les biohorizons ŕ valeur internationale, on peut retenir trois lignes-temps ayant une position identique dans les coupes d'Ellčs et de Kalaat Senan :
La position de la limite Campanien-Maastrichtien (Cp-Ma) a beaucoup fluctué dans la Formation Abiod selon les auteurs successifs.
À l'origine, en dehors de la Tunisie, pour de (1901), "Turrilites polyplocus" se trouve dans le Campanien, mais vers le bas de son "Campanien supérieur", puisque l'auteur ne reconnaît pas l'existence du Maastrichtien.
(1907) considčre que la Zone ŕ Bostrychoceras polyplocum se trouve dans des calcaires qui "correspondent aux couches supérieures du Campanien" mais que "divers auteurs placent męme dans le Maëstrichtien".
Un peu plus tard (1911) reprend la Zone ŕ B. polyplocum… mais la place ŕ la base de son "Maëstrichtien" (sans que l'on sache exactement quel est le critčre qu'il a retenu pour décider de ce choix).
En 1941 (p. 270), est le premier auteur ŕ populariser l'espčce de foraminifčre "Globotruncana calcarata" pour marquer un "Campanien inférieur". Le taxon remontera ensuite dans le Campanien supérieur terminal (, 1946 ; , 1945, 1947 ; & , 1947; , 1952 ; cf. & de , 1954).
Dans sa thčse, (1956) reprend le problčme de la position de la limite Cp-Ma suivant que soit utilisé le concept de (1911 : Zone ŕ B. polyplocum ŕ la base du "Maëstrichtien") ou celui des micropaléontologistes (le Campanien se termine avec l'extinction de "Globotruncana calcarata", c'est ŕ dire un peu au-dessus de Bostrychoceras polyplocum abondant).
Le concept des micropaléontologistes (derničre présence de "Globotruncana calcarata = fin du Campanien") sera adopté par la plupart des géologues ayant travaillé sur des coupes d'Afrique du Nord parmi lesquels (1955), & (1956), (1967, 1977, 1987), (1969), (1979), et alii (1983), et alii (1984), et alii (1990), et alii (1995).
Toutefois, dčs 1980 (p. 101), , reprenant la coupe du Kef, place la base du Maastrichtien non plus ŕ l'extinction de "Globotruncana calcarata" mais juste au-dessus du niveau ayant livré l'ammonite Pseudokossmaticeras brandti trouvée ŕ la base de la barre supérieure et considérée alors par comme caractéristique du "Campanien élevé" (, 1974, in litteris cité par , p. 101).
Poursuivant dans ce sens, & (1982) repčrent le petit foraminifčre planctonique Rugotruncana kefiana n. sp. (attribué plus tard au genre Archaeoglobitruncana par & , 1984), qui cohabite avec Pseudokossmaticeras brandti, les deux taxons indiquant ainsi le "Campanien". À ce moment, ils placent la limite Cp-Ma vers le haut de la barre supérieure de l'Abiod, lŕ oů apparaîtraient selon eux les premičres Globotruncana falsostuarti que (1952) citait dans son "Maëstrichtien" (espčce qui, selon nous, est déjŕ présente plus bas, dans les marnes Akhdar).
Cette position de la limite Cp-Ma au sommet de la barre supérieure ou vers son milieu est reprise par (1983) et & (1984).
Revenant sur la présence de Pseudokossmaticeras brandti ŕ la base de la barre supérieure, l'ammonite est cette fois interprétée comme marquant le Maastrichtien basal par se référant ŕ & (1976) in & (1989, p.303), âge qui rejaillit sur Archaeoglobitruncana kefiana -qui cohabite avec P. brandti- et qui signerait alors, elle aussi, le Maastrichtien basal.
C'est ce qui a été retenu par & (1992) pour la coupe de Kalaat Senan oů la base de la Zone ŕ A. kefiana désigne la base du Maastrichtien et correspond ŕ peu prčs ŕ la base de la barre supérieure.
Trois années plus tard, et alii (1995) se rallient pourtant ŕ l'opinion que le Campanien se termine avec la Zone ŕ "Globotruncana calcarata", tandis que (1995), dans la notice de la feuille de Gafour, place la limite Cp-Ma vers le sommet de la barre inférieure et attribue au Maastrichtien les alternances intermédiaires dans lesquelles il ne signale pas la présence de G. calcarata, ...
Dans la région de Kalaat Senan, le Crétacé supérieur a été révisé par et alii (2000) qui placent la limite Cp-Ma au tiers inférieur de la barre supérieure Ncham, ŕ l'articulation entre l'extinction de l'ammonite Nostoceras (Nostoceras) cf. hyatti (Campanien terminal) et l'apparition de l'ammonite Nostoceras (Nostoceras) aff. magdadiae & (Maastrichtien basal), suivant en cela les critčres recommandés par la Sous-commission internationale de stratigraphie du Crétacé ( et alii, 1996). Rappelons que Nostoceras magdadiae est une espčce définie dčs la base du Maastrichtien dans la Prairie Bluff Chalk d'Alabama et du Mississipi ( & , 1995).
Pour ces deux auteurs, la base du Maastrichtien est prise en Europe ŕ l'émergence de Belemnella lanceolata oů cette bélemnite surmonte immédiatement l'assemblage d'ammonites ŕ N. hyatti. Cela veut dire que c'est l'extinction de N. hyatti qui constitue la ligne-temps représentant la base du Maastrichtien dans la Prairie Bluff Chalk et elle est suivie par l'entrée de N. magdadiae.
Cette limite Cp-Ma qui correspond au sommet de la Zone ŕ Nostoceras hyatti se place un peu plus haut que les derniers Pseudokossmaticeras brandti (cette derničre espčce étant manifestement dans un Campanien élevé selon l'étude de & , 1993 et et alii, 2001).
Dans la coupe de l'oued Ncham, P. brandti a été récolté avec N. cf. hyatti ŕ la base de la barre supérieure, dans les alternances du Membre Gourbeuj. En outre, N. cf. hyatti a été trouvé plus haut dans les calcaires Ncham, au tiers inférieur de la barre supérieure ŕ Kalaat Senan ( et alii, 2000).
Cette conception de la limite Cp-Ma a été retenue et suivie par et alii (2002) dans leur étude de la coupe de l'Abiod d'El Djebil au Kef.
En 1995, au Symposium de Bruxelles dédié aux limites des étages du systčme Crétacé, sur proposition du Groupe de travail sur le Maastrichtien, la Sous-commission internationale de stratigraphie du Crétacé a recommandé de choisir l'ancienne carričre de Tercis pour placer le GSSP (Global standard boundary Stratotype, Section and Point) de la limite Campanien-Maastrichtien ( et alii, 1996), encore dénommé PSM (Point Stratotypique Mondial) par et alii (2005). La décision a été ratifiée par l'International Union of Geological Sciences en février 2001 (in & , 2001). En męme temps, un volume fort de 881 pages a été publié pour caractériser ŕ Tercis une limite Cp-Ma multicritčre ( rédacteur et 70 auteurs, 2001), suivi de plusieurs notes synthétisant les principaux résultats ( & , 2001 ; , 2002 ; cf. , 2002).
La coupe de Tercis est intéressante par la coexistence de groupes et d'espčces paléontologiques-clés comme les ammonites, les foraminifčres planctoniques, les nannofossiles, dinoflagellés, etc. dans une succession ŕ facičs calcaire continu.
Parmi les critčres biostratigraphiques mis en évidence ŕ Tercis, on retiendra ceux correspondant aux ammonites et aux foraminifčres planctoniques par le fait qu'on retrouve certains d'entre eux en Tunisie centrale, cf.
Fig. 4 ![]() , ce qui permettra d'effectuer des comparaisons et des corrélations entre Tercis et les deux coupes tunisiennes de Kalaat Senan et d'Ellčs :
, ce qui permettra d'effectuer des comparaisons et des corrélations entre Tercis et les deux coupes tunisiennes de Kalaat Senan et d'Ellčs :
En revanche, on ne retiendra pas ici comme critčres de corrélation ŕ grande distance de la coupe d'Ellčs plusieurs biohorizons relatifs aux foraminifčres planctoniques suivants.
Le biohorizon de l'apparition de Rugoglobigerina scotti ŕ Tercis
La citation de l'espčce est donnée par & (2001) qui la considčrent comme un "excellent index fossil" de la proximité de la limite Campanien-Maastrichtien et la proposent comme index de zone. Toutefois, plusieurs éléments modčrent l'utilisation de cette espčce en tant qu'index de zone international, quoique valable au niveau régional comme ŕ Tercis et dans l'Espagne proche :
Le biohorizon de l'apparition de Contusotruncana contusa ŕ Tercis
Ici, le problčme de détermination spécifique réside essentiellement dans le concept de l'espčce. En effet, C. contusa constitue le dernier terme d'une série évolutive anagénétique oů trois "espčces" morphologiques ont été retenues pour jalonner ce continuum : C. fornicata—>C. patelliformis—>C. contusa (cf. text-fig. 5 in et alii, 1984 ; , 1988a). Dans cette série la convexité de la face spirale croît graduellement depuis le Santonien jusqu'ŕ la fin du Maastrichtien, soit pendant prčs de 20 Ma. Dans l'espčce C. fornicata la spire est basse ŕ moyennement haute et le diamčtre est généralement compris entre 0,3 et 0,5 mm ; chez C. patelliformis la spire devient modérément haute ŕ haute et le diamčtre est compris entre 0,5 et 1 mm ; enfin, C. contusa se caractérise par une spire trčs haute, ŕ grand nombre de tours et ŕ surface ondulée, avec un diamčtre ŕ valeur élevée, souvent compris entre 0,7 et 1,5 mm. Comme dans toute lignée anagénétique, il existe de nombreux intermédiaires entre ces "espčces-types" et, en particulier entre C. patelliformis et C. contusa. Par ailleurs, les morphotypes évolués de fornicata coexistent aprčs la différenciation des premičres patelliformis et les deux formes coexistent encore aprčs la différenciation des premičres contusa et d'autres espčces proches comme C. walfischensis et C. plicata (cf. tableau phylogénétique de la text-fig. 8 in et alii, 1984). Pour fixer les limites morphologiques des espčces en cause, idéalement, on pourrait envisager de déterminer un "indice de profil" représenté par le rapport hauteur (H) / diamčtre (D) de la spire, mais ce serait sans tenir compte des variations possibles des morphologies qui dépendent des conditions paléobathymétriques, paléoécologiques, paléogéographiques etc. Une méthode de "grade datation" a męme été tentée pour déduire un "âge numérique" ŕ partir de la mesure d'un tel indice (, 1988a, 1988b), méthode qui a été contestée plusieurs fois car "inconciliable avec les données paléontologiques et les observations régionales" et ne pouvant donner des âges ŕ 0,1 Ma prčs alors qu'un redoublement tectonique de plusieurs Ma n'avait pas été décelé dans des coupes pyrénéennes (, 1994 ; et alii, 1999).
Ainsi, tout micropaléontologiste peut déterminer des morphotypes typiques de fornicata, de patelliformis et de contusa, ... mais il aura toujours des difficultés ŕ nommer les intermédiaires situés au voisinage des espčces-types : c'est le cas quand il s'agit de nommer une forme que l'on décrit qualitativement comme une "patelliformis évoluée" ou "premičre contusa".
À titre d'exemple, nous avons calculé l'indice H/D d'un certain nombre de morphotypes considérés par différents auteurs comme représentatifs des "espčces-types" constituant les jalons de la série anagénétique fornicata—>patelliformis—>contusa et choisis pour ętre figurés et publiés. L'indice est calculé ci-dessous ŕ partir de mesures effectuées sur des spécimens figurés dans l'Atlas de foraminifčres planctoniques du Crétacé supérieur ( et alii, 1984) et dans les planches de & (2001) : I. & O.
C. fornicata
| Atlas : | pl. 38, | fig. 1b | H/D = 0,34 |
| fig. 2b | H/D = 0,40 | ||
| fig. 3b | H/D = 0,33 | ||
| fig. 5b | H/D = 0,34 | ||
| moyenne pour l'Atlas : 0,35 | |||
| I. & O. : | pl. VI, | fig. 129 | H/D = 0,37 ("fornicata A") |
| fig. 136 | H/D = 0,51 ("fornicata B") |
C. patelliformis
| Atlas : | pl. 39, | fig. 2b | H/D = 0,51 |
| fig. 1b | H/D = 0,55 | ||
| fig. 3b | H/D = 0,58 | ||
| moyenne pour l'Atlas : 0,54 | |||
| I. & O. : | pl. VII, | fig. 148 | H/D = 0,57 |
| pl. VI, | fig. 140 | H/D = 0,55 ("fornicata B / contusa") | |
| fig. 137 | H/D = 0,62 ("fornicata B / contusa") |
C. contusa
| Atlas : | pl. 36, | fig. 2b | H/D = 0,55 |
| fig. 1b | H/D = 0,61 | ||
| pl. 37, | fig. 1b | H/D = 0,71 | |
| fig. 2b | H/D = 0,76 | ||
| fig. 3b | H/D = 0,86 | ||
| moyenne pour l'Atlas : 0,70 | |||
| I. & O. : | pl. VI, | fig. 144 | H/D = 0,58 |
| pl. VII, | fig. 146 | H/D = 0,59 |
Bien que les résultats n'aient pas de valeur statistique, le nombre de spécimens mesurés étant trop faible, on voit néanmoins que les auteurs ont sélectionné des morphotypes qualitativement assez typiques.
Les relevés ci-dessus appellent quelques commentaires :
De la sorte, on peut comprendre que l'indice de profil peut donner une indication immédiate sur la convexité d'une forme mais la détermination finale ne peut pas dépendre que de cet indice, notamment pour C. contusa : elle doit tenir compte également des autres critčres spécifiques comme la taille, le nombre de tours, l'aspect de la surface des loges, l'allongement des loges, l'existence ou non de sutures en relief entre les loges, etc.
En outre et pour réaliser des comparaisons plus complčtes avec les autres intervenants planctoniciens sur la coupe de Tercis, il aurait été intéressant que des "intermédiaires" aient été figurés, par exemple les "patelliformis/contusa" signalés par & (2001, Fig. 1, p. 336), ou bien les "contusa" juste au-dessous et au-dessus de la limite Campanien/Maastrichtien de & (2001, Fig. 1, p. 342), ou encore les "fornicata B" et les "patelliformis" proches de la limite Campanien/Maastrichtien de & (2001, Fig. 2, p. 353). On aurait alors pu s'assurer que c'est bien le męme concept de "contusa" qui était pratiqué par les différents auteurs.
Ces remarques n'entachent d'ailleurs en aucune façon la valeur de la limite Campanien-Maastrichtien telle qu'elle a été conçue en tant que limite multicritčre, définie par la moyenne arithmétique de la position d'une douzaine de biohorizons issus de six groupes paléontologiques importants. Męme si les concepts d'espčce venaient ŕ ętre quelque peu modifiés par des études paléontologiques ultérieures, la position du "Clou d'Or" (le "Golden Spike" des auteurs anglo-saxons) fixée ŕ Tercis ŕ la cote 115,2 devrait toujours marquer la base du Maastrichtien. C'est bien lŕ tout l'intéręt de ce concept de limite multicritčre imaginé par (2002) de ne pas se focaliser sur "l'apparition" d'un "taxon magique", en l'occurrence l'ammonite Pachydiscus neubergicus. En effet, la premičre présence de cette espčce a été notée ŕ 116,1 par & (2001) mais il existe un continuum morphologique entre P. neubergicus et son ancętre P. perfidus ( & , 1986 ; in et alii, § 9, 2001), de la męme façon qu'entre C. patelliformis et C. contusa, et l'on sait combien il est difficile de fixer une limite dans un continuum ! C'est ainsi qu'aprčs les premičres comparaisons entre les coupes de Tercis et de la Vistule (, 2004), (2005), repris in et alii (2007) en arrive, avec ses critčres personnels, ŕ considérer que P. neubergicus serait déjŕ présent ŕ Tercis ŕ 98,2 (pour lui, la forme figurée Pl. VI, 45 comme P. perfidus/neubergicus par & , 2001, serait un P. neubergicus). En conservant la limite ŕ 115,2 cela impliquerait que P. neubergicus apparaîtrait dans le Campanien terminal tandis que N. (N.) hyatti empičterait fortement dans le Maastrichtien si la base du Maastrichtien était descendue ŕ 98,2. Aussi, pour lui comme pour tous ceux favorables ŕ l'utilisation de l'apparition d'un seul taxon pour définir la base d'un étage, le fait de vieillir l'émergence de P. neubergicus impliquerait obligatoirement de modifier la position du "Clou d'Or", c'est ŕ dire d'aller ŕ l'encontre de la volonté de l'IUGS de stabiliser les limites d'étage. En outre, chaque nouvelle collecte ou chaque changement de concept d'espčce amčnerait ŕ modifier la position des limites qui seraient entraînées alors dans un yoyo perpétuel. Mais, de toute façon, avant de plonger dans une remise en question d'une décision de l'IUGS qui serait lourde de conséquences quant ŕ la stabilité de nombreuses limites et en particulier de celle du Campanien-Maastrichtien, ne faudrait-il pas revoir ce qu'il en est réellement des critčres de définition de l'espčce, surtout dans le cas d'une évolution anagénétique comme c'est le cas entre perfidus et neubergicus et, pour cela qu'un large éventail de spécialistes de ce groupe fossile se réunisse pour une discussion en face des collections de spécimens.
La Zone ŕ Gansserina gansseri ŕ Tercis
Dans la zonation standard des foraminifčres planctoniques ( coord. in et alii, 1998), la premičre présence de l'espčce gansseri étant signalée ŕ la base de la Zone ŕ N. hyatti, la Zone ŕ G. gansseri recouvre la partie supérieure élevée du Campanien et la partie inférieure du Maastrichtien, l'espčce se poursuivant ensuite presque jusqu'au sommet du Maastrichtien. Cette distribution de l'espčce dčs la partie supérieure du Campanien est reconnue en Tunisie oů elle est contrôlée par rapport aux ammonites tant dans la coupe de Kalaat Senan que dans celle d'Ellčs.
À Tercis, la premičre présence de l'espčce G. gansseri semble plus jeune quand on examine les contributions des divers auteurs ayant travaillé sur la coupe.
& (2001) ne signalent pas la présence de l'espčce (mais l'échantillonnage correspondait ŕ une étude préliminaire avec peu de matériel lavé disponible qui a montré que les "espčces-indice" étaient rares).
& (2001) ne signalent pas non plus l'espčce.
& (2001) remarquent la présence de l'espčce mais prčs de 50 m au-dessus de la limite Campanien-Maastrichtien.
Rappelons qu'antérieurement et alii (1996) ne notent pas de G. gansseri dans la section de Tercis et que & (1997) ne la citent que trčs haut dans la coupe, largement au-dessus de la limite Campanien-Maastrichtien telle que celle-ci fut définie ultérieurement dans réd. (2001).
La seule évocation de "G. gansseri" aux alentours de la limite Campanien-Maastrichtien dans la carričre de Tercis n'apparaît que dans le tableau de distribution des foraminifčres planctoniques de la fig.3 de la note de et alii (1997). L'espčce y est notée vers le niveau 112, soit un peu en dessous du GSSP -ou PSM selon et alii, 2005- de la limite Campanien-Maastrichtien (qui se trouve ŕ 115,2), puis nettement plus haut, vers 132 et 158. Toutefois, en l'absence de figuration du ou des spécimens de la premičre présence vers le niveau 112 (le nombre de spécimens n'est pas indiqué) et en souhaitant la publication de cette (ces) figuration(s), il n'est pas possible de considérer cette premičre citation comme avérée.
Cliquer sur la vignette pour agrandir l'image.
Figure 4: Position de la limite Campanien-Maastrichtien ŕ Ellčs par comparaison avec le stratotype international de Tercis, Landes, France et avec la coupe de Kalaat Senan en Tunisie (F.R.).
Prčs du village de Kalaat Senan, dans les coupes des oueds Assila, Gourbeuj et Ncham, la Formation Abiod montre un fort développement avec une puissance de prčs de 500 m ( et alii, 2000). Parmi les résultats biostratigraphiques on peut extraire pour comparaison la distribution verticale des marqueurs identifiés dans le stratotype de Tercis. C'est ce qui apparaît dans la figure 4 oů l'on voit :
Ainsi, ŕ Kalaat Senan, la limite Cp-Ma est placée lŕ oů s'arręte l'ammonite campanienne Nostoceras cf. hyatti, prise en relais par l'ammonite maastrichtienne Nostoceras aff. magdadiae, c'est-ŕ-dire sensiblement au tiers inférieur de la barre calcaire supérieure Ncham.
Plusieurs marqueurs biostratigraphiques ŕ distribution verticale courte ont été repérés dans la coupe
(Fig. 4 ![]() )
:
)
:
Ainsi, par comparaison avec la coupe trčs équivalente de Kalaat Senan, on peut envisager une limite Campanien-Maastrichtien quelque peu au-dessus de P. brandti, aux environs du niveau ELF 66 (comme dans et alii, 2009, fig. 3), soit environ au tiers inférieur de la barre calcaire supérieure Ncham.
Selon les concepts exposés au Symposium de Bruxelles en 1995 ( & , 1996), la base du Campanien a été proposée ŕ l'apparition de l'ammonite Placenticeras bidorsatum, proche de l'extinction du crinoďde Marsupites testudinarius qui, elle, est légčrement antérieure ŕ l'apparition de Broinsonia parca et postérieure ŕ l'apparition de Globotruncanita elevata. Quant au sommet du Campanien, il correspond ŕ la base du Maastrichtien qui est une limite multicritčre oů l'émergence de Pachydiscus neubergicus -proche de l'apparition de Belemnella lanceolata- prend en relais la disparition de Nostoceras hyatti au sommet du Campanien. Un Campanien limité de cette façon aurait une durée d'environ 11 Ma ( & , 1990 ; , 1994 ; in et alii, 2000 ; & , 2003), de 12,2 Ma ± 1 (in et alii, 1998) et de 12,9 Ma ± 1,3 (stratigraphy.org, chart 2009, oů elle est comprise entre 70,6 Ma ± 0,6 et 83,5 Ma ± 0,7), ce qui représente une moyenne d'environ 12 Ma. Par ailleurs, si l'on accepte la proposition d'une subdivision en trois sous-étages (inférieur, moyen et supérieur), de durées sensiblement équivalentes ( & , 1996), la durée d'un sous-étage devrait avoisiner les 4 Ma.
En admettant que la Zone d'ammonite ŕ Nostoceras (Bostrychoceras) polyplocum soit partie prenante de la partie supérieure du Campanien (cf. par exemple "Upper" in & , 2001 ; ou in et alii, 2001, fig. 8), on peut mesurer ŕ Ellčs le temps compris entre le biohorizon de l'émergence de N. (B.) polyplocum aux environs de ELD 76 (ou ELE 12) et la limite Campanien-Maastrichtien placée aux environs de ELF 66 (cf. § 4.4). Sur le tableau 1, ce temps est la somme de : la part du Mb. Mahdi au-dessus de la cote 76 (11/17 x 0,425 = 0,275 Ma) + Mb. Akhdar (2,40 Ma) + Mb. Gourbeuj (0,475 Ma) + la part du Mb. Ncham en dessous de la cote 66 (1,70 Ma), ce qui représente un total de 4,85 Ma, valeur qui dépasse 4 Ma mais n'est pas encore incompatible avec celle attendue.
Une telle proposition rejoint une suggestion exprimée et argumentée par (2004) qui écrit "The cosmopolitan N. (B.) polyplocum seems to be the ideal index for the definition of the base of the Upper Campanian Substage in Europe and the Russian Platform."
Si l'on voulait se rapprocher plus de la valeur 4 Ma, au lieu de fixer le niveau d'émergence de N. (B.) polyplocum comme base du Campanien supérieur, on pourrait prendre son niveau de disparition c'est ŕ dire aux environs de ELD 45, soit ŕ peu prčs 0,9 ŕ 1 Ma au-dessus. Mais, dans ce cas, on s'éloignerait résolument de la conception d'une partie supérieure du Campanien telle qu'elle a été comprise dans une grande partie des littératures traitant des domaines boréal ou téthysien qui, elles, incluent généralement la Zone ŕ N. (B.) polyplocum dans la partie supérieure du Campanien.
En résumé, en limitant la base d'un Campanien supérieur au biohorizon de l'émergence de Nostoceras (Bostrychoceras) polyplocum, un tel "Campanien supérieur" aurait une durée mesurée d'environ 4,8 Ma, ce qui satisferait assez bien d'abord ŕ la proposition d'une division tripartite de l'étage en sous-étages de durées ŕ peu prčs équivalentes et ensuite ŕ l'usage de ce biohorizon dans la littérature.
Il resterait maintenant ŕ placer une coupure entre un Campanien inférieur et moyen. Si l'on voulait un Campanien moyen d'une durée d'environ 4 Ma, ŕ Ellčs cela correspondrait ŕ 65 ŕ 70 m de calcaires Haraoua et une limite Campanien inférieur/moyen se situant entre ELD 10 et 15. Malheureusement, ŕ Ellčs, il n'y a pas de biohorizon d'ammonite ou de foraminifčre planctonique remarquable ŕ ces niveaux (le biohorizon d'apparition de Globotruncana ventricosa est environ 30 m plus haut, ŕ ELD 40,2 et, ŕ Kalaat Senan l'entrée de Ceratolithoides aculeus en AS 474, base de la Zone ŕ coccolithes CC 20 selon in et alii, 2000, p. 431, se produit plus bas, ŕ la base de la barre supérieure).
Deux études ŕ propos du Crétacé d'Ellčs ont été publiées assez récemment, l'une en 1998 par & , l'autre en 1999 par et alii Dans ces deux communications, sur base de foraminifčres planctoniques, la limite Campanien-Maastrichtien est placée ŕ deux niveaux différents ne correspondant ni l'un ni l'autre ŕ celui que nous proposons ici. L'indigence des colonnes lithologiques et la non définition des facičs en termes de formations ne permettent pas de comprendre toujours trčs clairement les interprétations biozonales et chronostratigraphiques proposées par les auteurs. Voyons ce qu'il en est successivement dans les deux publications.
Dans & (1998), la limite Campanien-Maastrichtien est placée au sommet de la Zone ŕ "Globotruncanita calcarata". Pourtant, plusieurs éléments semblent pour le moins intrigants.
Dans et alii (1999, p. 239), la limite Campanien-Maastrichtien est placée cette fois ŕ l'apparition du foraminifčre Rugoglobigerina hexacamerata (en faisant référence ŕ l'inversion magnétique C32N du site DSDP 525A … qui se trouve ŕ plusieurs milliers de km dans l'Atlantique de l'hémisphčre sud), c'est-ŕ-dire largement au-dessus de la base des marnes (El Haria), au dernier des "5 bancs" comme l'indique leur fig. 4B. Ce qui laisserait entendre que la totalité de la barre supérieure de l'Abiod ainsi que les 15 ŕ 20 m de base des marnes El Haria appartiendraient au Campanien (alors que les données macropaléontologiques relatives aux ammonites argumentent clairement une limite Cp-Ma beaucoup plus bas).
Quant ŕ l'apparition de Rugoglobigerina hexacamerata, cette espčce a été signalée ŕ Ellčs dčs 1978 vers la base des marnes El Haria (, éch. 33B) dans un "Maastrichtien inférieur". Elle a été trouvée aussi ŕ Kalaat Senan dans la base de la Formation des marnes El Haria et męme plus bas encore, au sommet des marnes Akhdar et dans les alternances Gourbeuj ( & in et alii, 2000). À moins d'utiliser le taxon dans une zone d'acmé, il ne paraît pas judicieux de choisir cette apparition en tant que proxy de limite d'étage, limite qui, de toute façon, se trouve manifestement plus bas que proposé par les auteurs.
En termes lithologiques, la coupe d'Ellčs a révélé une Formation Abiod tripartite, limitée vers le bas par les marnes Kef et, vers le haut, par les marnes El Haria. La Formation se marque dans le paysage par deux barres calcaires séparées par une dépression marneuse. La barre inférieure, épaisse d'environ 105 m est constituée des alternances de base Assila, suivies des bancs calcaires Haraoua (70 m) et se termine par les alternances Mahdi. Les marnes intermédiaires, ou Membre Akhdar, se développent sur 77 m avec leur teinte turquoise caractéristique. La barre supérieure, épaisse de 104 m, commence par les alternances Gourbeuj, suivies des gros bancs calcaires Ncham (85 m) et se termine par les alternances Gouss.
Plusieurs biohorizons définis ŕ partir de micro- et de macrofossiles constituent des lignes-temps qui sont d'excellents outils de corrélation.
Parmi les microfossiles, les premičres présences de foraminifčres planctoniques marqueurs fondent une zonation ŕ valeur internationale avec la succession suivante, du plus ancien au plus récent : IZ ŕ Globotruncanita elevata, IZ ŕ Globotruncana ventricosa, TRZ ŕ Radotruncana calcarata, PRZ ŕ Globotruncana falsostuarti, IZ ŕ Gansserina gansseri, IZ ŕ Contusotruncana contusa. Il faut y ajouter les niveaux d'émergence de Radotruncana subspinosa (un peu avant celui de R. calcarata), de Globotruncanita angulata (un peu avant celui de G. gansseri) et de Archaeoglobitruncana cf. kefiana (vers la base de la barre supérieure).
Parmi les macrofossiles, des ammonites ont été surtout récoltées et plusieurs biohorizons sont bien délimités : la courte distribution verticale de Nostoceras (Bostrychoceras) polyplocum ŕ partir du milieu des alternances Mahdi jusqu'ŕ la fin du premier quart des marnes Akhdar ; le niveau ŕ Nostoceras (N.) cf. hyatti ŕ la base de la barre supérieure, les bancs ŕ Pseudokossmaticeras brandti dans le quart inférieur des calcaires Ncham ; les bancs ŕ Pachydiscus (P.) neubergicus dans le tiers supérieur des calcaires Ncham.
La conjonction de plusieurs de ces lignes-temps permet une datation relative plus précise des diverses unités lithologiques et surtout une corrélation des séries appartenant ŕ des bassins éloignés.
Ainsi, lors de la construction de cartes paléogéographiques – par exemple celles que l'on pourrait établir en Tunisie pour chacun des membres principaux de l'Abiod – l'utilisation de lignes-temps bien contrôlées, synchrones et valables au plan international permettrait des tracés bien contraints temporellement avec des résultats susceptibles de localiser des surfaces favorables ŕ des recherches pétroličres.
En tenant compte des divers biohorizons marqueurs, la limite Campanien-Maastrichtien, se situe ŕ environ 50 m au-dessus des apparitions de G. gansseri et de N. hyatti et ŕ 15-20 m au-dessus des bancs ŕ P. brandti, c'est-ŕ-dire au sommet du premier tiers de la barre Ncham. La position de cette limite est comparée et corrélée avec celle établie dans la coupe de Kalaat Senan (męmes marqueurs et sensiblement męmes distributions verticales des macro- et des microfossiles) et avec celle du stratotype de Tercis en France (grâce aux biohorizons marqueurs de N. (B.) polyplocum, N. hyatti, P. brandti et R. calcarata).
En Tunisie, en combinant les informations issues des unités lithologiques et des biohorizons, l'Abiod tripartite d'Ellčs constitue un ensemble puissant de prčs de 286 m se marquant trčs clairement dans la morphologie et représentant une sédimentation marine pélagique durant environ 13,2 millions d'années selon les mesures réalisées avec l'outil cyclostratigraphique.
La barre inférieure (Assila + Haraoua + Mahdi), épaisse de 105 m, représente une durée de dépôt d'environ 6 Ma avec un taux moyen de sédimentation non décompactée de 1,7 cm/ky. Les marnes intermédiaires Akhdar, épaisses de 77 m représentent une durée de dépôt d'environ 2,4 Ma avec un taux moyen de sédimentation non décompactée de 3,2 cm/ka. Pour la barre supérieure (Gourbeuj + Ncham + Gouss), ses 104 m représentent une durée de dépôt d'environ 4,7 Ma avec un taux moyen de sédimentation non décompactée de 2,1 cm/ka. Les alternances Gourbeuj et le tiers inférieur des calcaires Ncham correspondent ŕ la partie supérieure du Campanien supérieur tandis que les deux tiers supérieurs des calcaires Ncham appartiennent déjŕ ŕ la partie inférieure du Maastrichtien.
Si l'on accepte une division ternaire de l'étage Campanien avec des sous-étages de durées ŕ peu prčs équivalentes, il est proposé de définir un Campanien supérieur dont la base serait prise ŕ l'apparition de l'ammonite N. (Bostrycoceras) polyplocum et le sommet, c'est ŕ dire la base du Maastrichtien, correspondrait ŕ l'extinction de Nostoceras hyatti. Un tel Campanien supérieur aurait une durée mesurée par cyclostratigraphie d'environ 4,8 Ma pour un étage Campanien d'un peu plus de 12 Ma.
Plusieurs zones paléontologiques désignent la distribution verticale totale de plusieurs marqueurs et ont une valeur globale. Ce sont la TRZ ŕ N. (B.) polyplocum qui comprend le sommet de la barre inférieure et le tiers inférieur des marnes Akhdar : environ 34 ŕ 36 m et 0,9 ŕ 1 Ma ; la TRZ ŕ P. brandti, ŕ partir des alternances Gourbeuj au quart inférieur de Ncham ; la TRZ ŕ N. hyatti qui commence également dans les alternances Gourbeuj, se poursuit jusqu'au tiers inférieur de Ncham oů sa derničre présence marque la limite Campanien-Maastrichtien, la TRZ ŕ R. calcarata, de la partie moyenne des marnes Akhdar, connue sur au moins 30 m avec un temps de dépôt minimum d'environ 0,79 Ma.
En tenant compte de ces apports, une lecture critique a été effectuée de deux notes publiées il y a une décennie ŕ propos d'une partie de la coupe d'Ellčs et en particulier sur la position controuvée de la limite Campanien-Maastrichtien.
La détermination des foraminifčres planctoniques en plaques minces a bénéficié de la grande expérience de Madame Michčle qui a reçu l'un d'entre nous (M.M.) ŕ l'Université de Fribourg (Suisse). L'appartenance générique des ammonites récoltées dans les coupes partielles a été confirmée ou définie lors de discussions avec le doctorant Stijn (Université de Leuven, Belgique). Sur le terrain, les travaux de levés géologiques, d'échantillonnage et de collecte de macrofaunes ont été réalisés par les deux auteurs avec l'aide de Mourad et de Taoufikh (ETAP) ainsi que de Moheddine (Kalaat Senan). Qu'ils soient tous remerciés de leur intervention. Les photographies au Microscope électronique ŕ balayage ont été effectuées par l'un d'entre nous (F.R.) sur l'appareil du Centre de Recherche de l'ETAP ŕ Tunis et nous remercions cet organisme de nous avoir autorisé ŕ l'utiliser. Le manuscrit a bénéficié des remarques constructives et des suggestions de Jochen , d'un lecteur anonyme et du Dr Gilles-Serge qui est intervenu dans des discussions éclairantes ŕ divers stades de la révision du texte final, nous les en remercions tous trois ainsi que Nestor qui a bien voulu accepter la tâche ingrate de la traduction en anglais.
J.A. & E. (2001).- Planktic foraminiferal quantitative analysis across the Campanian-Maastrichtian boundary at Tercis. In: G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. C5b, p. 338-348.
G. & I. de (1954).- Contribution ŕ la connaissance du niveau stratigraphique de "Globotruncana calcarata". In : Association française pour l'Avancement des Sciences, Actes du 73e Congrčs.- Annales de l'Université de Poitiers, 2, n° 5, 4 p.
J.-P., M., P., D., A.L. & J. (1983).- Le Campanien sommital et le Maastrichtien de la coupe du Kef (Tunisie septentrionale) : zonation sur base des foraminifčres planctoniques.- Zitteliana, München, vol. 10, p. 609-611.
A., P.F. & F. (1990).- Petroleum geology in Tunisia.- ETAP (Entreprise Tunisienne d'Activités Pétroličres), Mémoire, Tunis, n° 1, 194 p.
M. (1994).- Faunes d'ammonites et interprétation des successions sédimentaires du Campanien-Maastrichtien sous-pyrénéen (Aričge, Haute-Garonne). Réfutation d'un modčle fondé sur des âges numériques hypothétiques.- Géologie de la France, Orléans, 1994, n° 3, p. 71-80.
M., G.S. & P. (1999).- Arguments ŕ l'encontre des estimations d'âge biograduelles (et de leur application en chronologie et stratigraphie séquentielle) dans le Sud-Ouest de la France.- Bulletin de la Société géologique de France, Paris, vol. 170, n° 4, p. 465-473.
J., P.F. & G. (1952).- Le Sillon tunisien. In : XIXe Congrčs Géologique International, Monographies régionales, (2e Série : Tunisie), n° 5, 112 p.
P.F. (1956).- Contribution ŕ l'étude stratigraphique de la Tunisie centrale.- Annales des Mines et de la Géologie, Tunis, vol. 18, 350 p.
P.F., A., D. & A. (1954).- Unités stratigraphiques en Tunisie centrale.- Compte rendu du 19čme Congrčs géologique international d'Alger, XXI, p. 243-254.
P.F. & P. (1956).- Notice explicative de la carte géologique de Tunisie au 50 000e, feuille n° 51 : Tajerouine.- Service de la Carte géologique de la Tunisie, Tunis.
M. & G.S. (2001).- Report on a preliminary study of the planktonic foraminifera of the Campanian-Maastrichtian succession at Tercis (Landes, France). In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. C5a, p. 334-337.
W.A. & W.J. (1995).- Maastrichtian ammonites chiefly from the Prairie Bluff Chalk in Alabama and Mississippi.- The Paleontological Society, Memoir, London, 44, 40 p.
P. & G.S. (2001).- Les ammonites spiralées du Campanien et du Maastrichtien de Tercis les Bains (Landes, France). In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, p. 529-549.
J.A. (1946).- Upper Cretaceous Foraminifera of the Gulf Coastal Plain of the United States and adjacent areas.- U.S. Geological Survey, Professional Papers, Washington, 206, 241 p.
F. (1955).- The genus Globotruncana in Tunisia.- Micropaleontology, New York, vol. 1, p. 161-171.
F. (1956).- Étude sommaire des microfaunes de la région du Kef. Notice explicative de la Carte géologique de la Tunisie au 50 000e, feuille n° 44 : Le Kef, 33-38.- Service de la Carte géologique de la Tunisie, Tunis.
T. (1995).- Notice explicative de la Carte géologique de la Tunisie au 50 000e, feuille n° 40 : Gafour.- Service de la Carte géologique de la Tunisie, Tunis, 16 p.
J.-P. & P. (1947).- Résultats stratigraphiques d'un sondage ŕ Peyrehorade (Landes).- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, 1947, p. 330-333.
M.-J. (1988a).- Grade-datations sénoniennes : application aux séquences de dépôt du versant sud des Pyrénées centrales.- Thčse de Doctorat es Sciences, Université de Toulouse, 835 p.
M.-J. (1988b).- L'évolution graduelle du profil du test dans la lignée de Rosita fornicata. Une nouvelle possibilité de grade-datation au Crétacé supérieur.- Comptes Rendus des Séances de l'Académie des Sciences, Paris, (II), t. 306, p. 1379-1384.
D. (1978).- Nomenclature stratigraphique des séries du Crétacé supérieur au Tertiaire de Tunisie.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, vol. 2, n° 1, p. 97-148.
S. (2010, inédit/unpublished).- Late Cretaceous ammonites from Tunisia: chronology and causes of their extinction and extrapolation to other areas.- PhD, K.U. Leuven, 388 p., atlas: 84 pls.
A. de (1901).- Recherches sur la craie supérieure.- Mémoires explicatifs de la Carte géologique détaillée de la France, Ministčre des Travaux Publics, Paris, fasc. 1 : 559 p. ; fasc. 2 : p. 560-1013.
J.M. & A.S. (1996).- The Campanian stage.- Institut royal des Sciences naturelles de Belgique, Bulletin, Bruxelles, Sciences de la Terre, vol. 66, Supplement, p. 103-109.
J.M. & W.J. (1993).- The high Cretaceous ammonite fauna from Tercis, Landes, France.- Institut royal des Sciences naturelles de Belgique, Bulletin, Bruxelles, Sciences de la Terre, vol. 63, p. 149-209.
J., J., M.B., T., P.-C. de & P.R. (eds., 1998).- Cretaceous biochronohorizons and zones. In : Cretaceous biochronostratigraphy, Chart 5, Mesozoic and Cenozoic Sequence Stratigraphy of European Basins.- SEPM (Society for Sedimentary Geology), Special Publication, Tulsa, n° 60, 786 p.
E. (1911).- Traité de géologie. II : les périodes géologiques.- A. Colin, Paris, 2021 p.
M., F. & S. (2009).- Cyclostratigraphy and chronometric scale in the Campanian – Lower Maastrichtian: the Abiod Formation at Ellčs, central Tunisia.- Cretaceous Research, London, vol. 30, n° 2, p. 325-338.
J. & G.S. (2001).- Planktonic Foraminifera from the Campanian-Maastrichtian at Tercis les Bains (Landes, France). In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. C5c, p. 349-370.
I., A., R.T.J. & S. de (2002).- Late Cretaceous (Campanian) carbon isotope events, sea-level change and correlation of the Tethyan and Boreal realms.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 188, p. 215-248.
W.J., M. & A. (2001).- Pseudokossmaticeras brandti , 1873, an Upper Campanian marker fossil in northern Aquitaine, France.- Cretaceous Research, London, vol. 22, n° 2, p. 259-262.
W.J. & H. (1986).- Lower Maastrichtian ammonites from Neuberg, Steiermark, Austria.- Beiträge zür Paläontologie von Österreich, Wien, Band 12, p. 181-242.
J. (1945).- Étude sur les Globotruncana du Bassin d'Aquitaine.- Diplôme d'Études Supérieures, Université de Toulouse, 95 + 4 p.
J. (1947).- Les Globotruncana du Crétacé supérieur Nord-Pyrénéen.- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, 1947, n° 1, p. 19-22.
T., A. & M. (2001).- The Campanian-Maastrichtian boundary in northern Spain (Navarra Province): the Imiscoz and Erro sections. In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. E3, p. 723-744.
T. & G.S. (2001).- Upper Campanian-Maastrichtian ammonites (Nostoceratidae, Diplomoceratidae) from Tercis les Bains (Landes, France). In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. D4e, p. 500-528.
I. (1979).- Carte géologique au 50 000e et notice explicative de la Carte géologique de la Tunisie au 50 000e, feuille n° 52 : Ebba Ksour.- Service de la Carte géologique de la Tunisie, Tunis, 36 p.
L. & G. (1998).- Diversification and extinction in Campanian-Maastrichtian planktic foraminifera of Northwestern Tunisia.- Eclogae geologicae Helvetiae, Basel, vol. 91, p. 75-102.
L., G. & W. (1999).- The Late Campanian and Maastrichtian in northwestern Tunisia : palaeoenvironmental inferences from lithology, macrofauna and benthic foraminifera.- Cretaceous Research, London, vol. 20, n° 2, p. 231-252.
M. (2005).- Late Maastrichtian and earliest Danian scaphitid ammonites from central Europe: taxonomy, evolution and extinction.- Acta Palæontologica Polonica, Warszawa, vol. 50, p. 653-696.
M., J.W.M., N.H. & N. (2007).- The highest records of North American scaphitid ammonites in the European Maastrichtian (Upper Cretaceous) and their stratigraphic implications.- Acta Geologica Polonica, Warszawa, vol. 57, p. 169-185.
P. (1941).- Les foraminifčres de la craie ŕ Belemnitella mucronata du Bassin de Paris.- Mémoires du Muséum, Muséum national d'Histoire naturelle de Paris, (Nouv. Série), t. XII, fasc. 1, 296 p., 37 pls.
A., F., P.F., L. & H. (1995).- Recueil des coupes-types de Tunisie.- ETAP, Mémoire, Tunis, n° 8B, 123 p.
M.H. & A. (1992).- Les facičs riches en matičre organique des formations Aleg et Abiod dans le Centre-Nord de la Tunisie. In : Actes des 3e Journées de Géologie tunisienne appliquée ŕ la recherche des hydrocarbures.- ETAP, Mémoire, Tunis, n° 5, p. 277-294.
B. (2004).- Late Campanian nostoceratid ammonites from Lehrte West Syncline near Hannover, northern Germany.- Acta Geologica Polonica, Warszawa, vol. 54, p. 473-487.
G.S. (1994).- Geological time scale, 1994.- Comptes Rendus des Séances de l'Académie des Sciences, Paris, (II), t. 318, p. 59-71
G.S. (Éditeur, 2001).- The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, xxviii + 881 p.
G.S. (2002).- Définition d'une limite multicritčre ; stratigraphie du passage Campanien-Maastrichtien du site géologique de Tercis (Landes, SW France).- Comptes rendus Géosciences, Paris, vol. 334, p. 409-414.
G.S., P., W.A. & M. (2001).- The Campanian-Maastrichtian ammonite fauna from Tercis (Landes, France); a synthetic view. In : G.S. (ed., 2001), The Campanian - Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents.- IUGS Special Publication (monograph) Series, 36 ; Developments in Palaeontology and Stratigraphy Series, Amsterdam, 19, chap. D4g, p. 550-561.
G.S., S., F. & J. (2005).- Les unités de l'histoire géologique ; discussion sur les conventions et l'échelle des temps.- Annales de la Société géologique du Nord, Lille, (2čme série), t. 12, p. 63-68.
G.S. & M.A. (2001).- The global Campanian-Maastrichtian stage boundary at Tercis les Bains, Landes, SW France.- Episodes, Beijing, vol. 24, n° 4, p. 229-238.
G.S. & Ch. (1990).- Échelle numérique des temps géologiques.- Géochronique, Paris, n° 35, p. 12-21.
G.S. & I. (2003).- Sur les inocérames de Tercis (Landes, France) : le meilleur outil corrélatif entre Europe et Amérique du Nord autour de la limite Campanien-Maastrichtien / Inoceramids of the site of Tercis (Landes, France): the best correlative tool between Europe and North America across the Campanian-Maastrichtian boundary.- Comptes rendus Géosciences, Paris, vol. 335, n° 2, p. 239-246.
L. (1907).- Études de paléontologie tunisienne. I. Céphalopodes des terrains secondaires, systčme crétacique.- Carte géologique de Tunisie, de Rudeval, Paris, v + 438 p.
B., M.-J., Y. & P. (1997).- Stratigraphie séquentielle, biozonation par les foraminifčres planctoniques, grade-datation et évaluation des taux de sédimentation dans les calcaires crayeux campano-maastrichtiens de Tercis (SW de la France).- Bulletin de la Société géologique de France, Paris, vol. 168, p. 143-153.
P.F., A.V., J.M. & W.J. (eds., 1996).- Proceedings of the 2nd International Symposium on Cretaceous Stage Boundaries, Brussels 8-16 sept. 1995.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, 66 (Supplement), 177 p.
J., ICS Subcommissions, A. & G.S. (2000).- International Stratigraphic Chart and Explanatory note.- UNESCO-IUGS Special edition, 31st International Geological Congress, Rio.
F. (1998).- Cretaceous biochronohorizons and zones. Planktonic foraminifera. In : J., J., M.B., T., P.-C. de & P.R. (eds.), Cretaceous biochronostratigraphy, Chart 5, Mesozoic and Cenozoic Sequence Stratigraphy of European Basins.- SEPM (Society for Sedimentary Geology), Special Publication, Tulsa, n° 60.
F. (2002).- The Campanian-Maastrichtian Stage Boundary. Edited by G.S. . Book review.- Episodes, Beijing, vol. 25, p. 215-216.
F. & M. (1995).- Foraminifčres planctoniques du Crétacé : commentaires de la zonation Europe-Méditerranée.- Bulletin de la Société géologique de France, Paris, vol. 166, p. 681-692.
F., M., J.M., A.H. (eds.) & European Working Group on Planktonic Foraminifera (1984).- Atlas of Late Cretaceous globotruncanids.- Revue de Micropaléontologie, Paris, vol. 26, p. 145-305.
F., J.M., D., F., M., C., A. & S. (2000).- Le Crétacé Supérieur de la Région de Kalaat Senan, Tunisie Centrale. Litho-biostratigraphie intégrée : zones d'ammonites, de foraminifčres planctoniques et de nannofossiles du Turonien supérieur au Maastrichtien.- Bulletin des Centres de Recherche Exploration Production Elf-Aquitaine, Pau, vol. 22, n° 2, p. 359-490.
R. (1978).- Étude stratigraphique et micropaléontologique du passage Crétacé-Tertiaire du synclinal d'Ellčs (région Siliana-Sers), Tunisie centrale.- Thčse 3čme cycle Université Pierre et Marie Curie, Paris VI, 275 + I-XV p.
J. (1969).- Zones planctoniques du Crétacé et du Paléogčne de Tunisie. In : Proceedings of the First International Conference on Planktonic Microfossils, Geneva (1967).- E.J. Brill, Leiden, vol. II, p. 588-593.
J. (1980).- Microbiostratigraphie du Crétacé et du Paléogčne de la Tunisie septentrionale et orientale (hypostratotypes tunisiens).- Geologicky Ustav Dionyza Stura, Bratislava, 238 p.
J. (1983).- Quelques problčmes taxinomiques concernant les foraminifčres planctiques et la zonation du Crétacé supérieur d'El Kef.- Geologicky Zbornik, Geologica Carpathica, Bratislava, vol. 34, p. 187-211.
J. & A.L. (1982).- Rugotruncana kefiana n.sp. (Foraminifera) du Sénonien supérieur d'El Kef.- Geologicky Zbornik, Geologica Carpathica, Bratislava, vol. 33, p. 463-479.
J. & A.L. (1984).- Campanian-Maastrichtian boundary in Tunisia.- Geologicky Zbornik, Geologica Carpathica, Bratislava, vol. 35, p. 551-558.
J. & A.L. (1987).- Archaeoglobitruncana kefiana foraminiferal zone of the basal Tethyan Maastrichtian.- Geologicky Zbornik, Geologica Carpathica, Bratislava, vol. 38, p. 677-689.
J. & J. (1989).- The Campanian-Maastrichtian boundary in the El Kef Section, Tunisia. In : J. (ed.), Cretaceous of the Western Tethys.- Proceedings of the 3nd International Cretaceous Symposium, Tübingen (1987).- E. Schweizerbart'sche Verlagbuch, Stuttgart, p. 299-315.
J. (1952).- Aperçu stratigraphique sur la micropaléontologie du Crétacé.- XIXe Congrčs Géologique International, Monographies régionales, (1e Série : Algérie), Alger, n° 26, 47 p.
J. (1967).- Essai sur l'état actuel d'une zonation stratigraphique ŕ l'aide des principales espčces de Rosalines (Foraminifčres).- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, 1967, n° 2, p. 48-50.
J. (1977).- Essai de zonation du Crétacé méditerranéen ŕ l'aide des foraminifčres planctoniques.- Géologie Méditerranéenne, Marseille, t. 4, n° 2, p. 99-108.
J. (1987).- Une échelle zonale du Crétacé méditerranéen et quelques réflexions suscitées par son établissement particuličrement ŕ propos du Danien.- Revue de Micropaléontologie, Paris, vol. 30, n° 1, p. 32-51.
M.D., C.L. & J.M. (1996).- Planktonic foraminifera across the Campanian-Maastrichtian boundary at Tercis, SW France.- Newsletters on Stratigraphy, Hannover, vol. 34, p. 65-80.
F. & J. (1976).- Ammoniten und Alter der höheren Kreide (Gosau) des Krappfeldes in Kärnten (Österreich).- Mitteilungen aus dem Geologisch-Paläontologischen Instituts der Universität Hamburg, Heft 45, p. 9-27.
I. (2004).- Inoceramids and inoceramid biostratigraphy of the Upper Campanian to basal Maastrichtian of the Middle Vistula Valley, central Poland.- Acta Geologica Polonica, Warszawa, vol. 54, p. 95-168.
P.D. & W. (1997).- Campanian-Maastrichtian ammonite and planktonic foraminiferal biostratigraphy from Tercis, France.- Journal of Paleontology, Tulsa, vol. 71, p. 407-418.
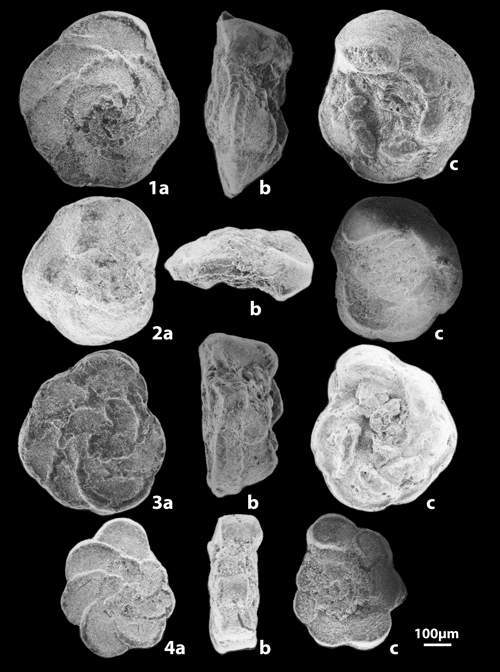
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 1: Planktonic foraminifera of the Abiod Formation (a: spiral side, b: lateral view, c: umbilical side).
figure 1: Globotruncanita elevata (): section ELA 5.4 m; top of the marls of the Kef Formation.
figure 2: Contusotruncana fornicata (): section ELD 83.3 m; top of the lower limestone bar, alternances of the Mahdi Member.
figure 3: Globotruncana ventricosa (): section ELE 86.2 m; upper part of the Akhdar intermediate marls.
figure 4: Globotruncana linneiana (d'): section ELE 43.50 m; lower part of Akhdar Member.
Planche 1 : Foraminifčres planctoniques de la Formation Abiod ŕ Ellčs (a : face spirale ; b : profil ; c : face ombilicale).
figure 1 : Globotruncanita elevata () : coupe ELA 5,4 m ; sommet des marnes de la Formation Kef.
figure 2 : Contusotruncana fornicata () : coupe ELD 83,3 m ; sommet de la barre calcaire inférieure, alternances du Membre Mahdi.
figure 3 : Globotruncana ventricosa () : coupe ELE 86,2 m ; partie élevée des marnes intermédiaires Akhdar.
figure 4 : Globotruncana linneiana (d') : coupe ELE 43,50 m ; partie inférieure du Membre Akhdar.
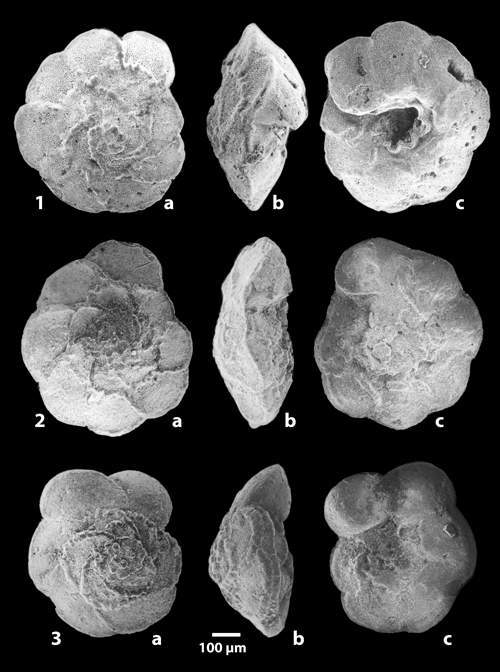
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 2: Planktonic foraminifers of the Abiod Formation at Ellčs. (a: spiral side, b: lateral view, c: umbilical side).
figure 1: Globotruncana dupeublei , , & : section ELG 27 m; lower part of the El Haria Formation.
figure 2: Globotruncana falsostuarti : section ELE 26.6 m; base of the intermediate Akhdar Member.
figure 3: A form transitional between Globotruncana arca and Globotruncana orientalis = Globotruncana stephensoni (1 to 2 final chambers with one keel): section ELE 82 m; upper part of the Akhdar marls.
Planche 2 : Foraminifčres planctoniques de la Formation Abiod ŕ Ellčs (a : face spirale ; b : profil ; c : face ombilicale).
figure 1 : Globotruncana dupeublei , , & : coupe ELG 27 m ; partie basse de la Formation El Haria.
figure 2 : Globotruncana falsostuarti : coupe ELE 26,6 m ; base des marnes intermédiaires Akhdar.
figure 3 : Intermédiaire entre Globotruncana arca et Globotruncana orientalis = Globotruncana stephensoni (1 ŕ 2 derničres loges unicarénées) : coupe ELE 82 m ; partie élevée des marnes intermédiaires Akhdar.

Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 3: Planktonic foraminifers of the Abiod Formation at Ellčs. (a: spiral side, b: lateral view, c: umbilical side).
figure 1: Radotruncana calcarata (): section ELE 43.5 m; lower part of the Akhdar intermediate marls.
figure 2: Radotruncana subspinosa (): section ELE 70.5 m; upper part of the Akhdar marls.
figure 3: Globotruncanita stuartiformis (): section ELE 90 m; higher part of the Akhdar marls.
Planche 3 : Foraminifčres planctoniques de la Formation Abiod ŕ Ellčs (a : face spirale ; b : profil ; c : face ombilicale).
figure 1 : Radotruncana calcarata () : coupe ELE 43,5 m ; partie inférieure des marnes intermédiaires Akhdar.
figure 2 : Radotruncana subspinosa () : coupe ELE 70,5 m ; partie supérieure des marnes intermédiaires Akhdar.
figure 3 : Globotruncanita stuartiformis () : coupe ELE 90 m ; partie élevée des marnes intermédiaires Akhdar.

Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 4: Planktonic foraminifers of the Abiod Formation at Ellčs (a: spiral side, b: lateral view, c: umbilical side).
figure 1: Globotruncana insignis : section ELE 22.7 m; basal part of the Akhdar marls; note that its first occurrence is lower than that in the et alii (2000) Kalaat Senan section.
figure 2: Contusotruncana contusa () or a transition between C. contusa and C. patelliformis () but closer to C. contusa: section ELG 27 m; lower part of the El Haria marls; profile index 0.54; a morphotype very close to that of C. contusa in the Atlas of Upper Cretaceous Planktonic Foraminifers, pl. 36, figs. 2 abc, index of this profile: 0.55 ( et alii, 1984).
figure 3: Globotruncana stuarti (de ): section ELG 31.1 m, lower part of the El Haria marls.
figure 4: Globotruncana aegyptiaca : section ELG 31.1 m, lower part of the El Haria marls.
Planche 4 : Foraminifčres planctoniques de la Formation Abiod ŕ Ellčs (a : face spirale ; b : profil ; c : face ombilicale).
figure 1 : Globotruncana insignis : coupe ELE 22,7 m ; partie basale des marnes intermédiaires Akhdar. À noter que cette premičre présence se manifeste plus bas que dans la coupe de Kalaat Senan de et alii (2000).
figure 2 : Contusotruncana contusa () ou intermédiaire entre C. contusa et C. patelliformis () mais plus proche de C. contusa : coupe ELG 27 m ; partie inférieure des marnes El Haria, indice de profil : 0,54. Morphotype trčs proche de la représentation de C. contusa de l'Atlas des foraminifčres planctoniques du Crétacé supérieur, pl. 36, figs. 2abc, indice de profil : 0,55 ( et alii, 1984).
figure 3 : Globotruncana stuarti (de ) : coupe ELG 31,1 m ; partie inférieure des marnes El Haria.
figure 4 : Globotruncana aegyptiaca : coupe ELG 31,1 m ; partie inférieure des marnes El Haria.
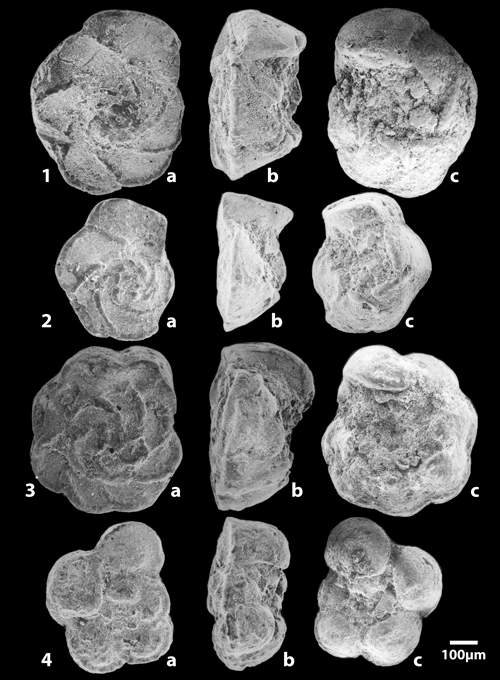
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 5: Planktonic foraminifers of the Abiod Formation at Ellčs. (a: spiral side, b: lateral view, c: umbilical side).
figure 1: Globotruncanita angulata (): section ELE 26.6 m, base of the Akhdar intermediate marls. A single-keeled form, a homeomorph of Gansserina gansseri, found very low in the Akhdar marls (lower than these morphs in the Kalaat Senan section).
figure 2: Globotruncanita pettersi (): section ELE 33 m; base of the Akhdar intermediate marls.
figure 3: Globotruncana ventricosa : section ELD 83.3 m; alternances of the Mahdi Member below the Haraoua bar.
figure 4: Gansserina wiedenmayeri (): section ELE 80 m; form with two closely-spaced keels then on the last chamber only one, thus tending toward Gansserina gansseri.
Planche 5 : Foraminifčres planctoniques de la Formation Abiod ŕ Ellčs (a : face spirale ; b : profil ; c : face ombilicale).
figure 1 : Globotruncanita angulata () : coupe ELE 26,6 m, base des marnes intermédiaires Akhdar. Forme unicarénée, homéomorphe de Gansserina gansseri, se trouvant trčs bas dans les marnes Akhdar (plus bas que les morphes de la coupe de Kalaat Senan).
figure 2 : Globotruncanita pettersi () : coupe ELE 33 m ; base des marnes intermédiaires Akhdar.
figure 3 : Globotruncana ventricosa : coupe ELD 83,3 m ; alternances Mahdi au-dessus de la barre Haraoua.
figure 4 : Gansserina wiedenmayeri () : coupe ELE 80 m ; forme montrant deux carčnes proches puis une carčne sur la derničre loge et conduisant ŕ Gansserina gansseri.

Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 6: The lower part of the Abiod Formation at Ellčs (photos F. ).
figure 1 (negative 265-26). The lower bar of the Abiod in partial section ELD (in the eastern thalweg of wadi Ed Dam; upper portion of the lower part of the Campanian.
In the foreground crop out the last beds of the massive limestones of the Haraoua Member (H) of the lower bar of the Abiod Formation. The passage from the limestones of the Haraoua Member to the alternances of limestones-marly limestones of the Mahdi Member (M) takes place around ELD 70 where the first impressions of ammonites of the Nostoceras (Bostrychoceras) group occur.
To the right of the pylon, the three small white arrows indicate the location of the paved road where there is a vehicle.
figure 2 (negative 267-16). Intermediate marls of the Akhdar Member in partial section ELE (in the northern thalweg of wadi Ed Dam, southeast of the paved road) upper part of the Campanian.
The green turquoise marls are dominant in the Akhdar Member (A) with respect to the occurrence of marly limestones and limestones. Note that the planktonic foraminifer Radotruncana calcarata is present at least from ELE 43.50 m and continues up to 73.70 m. The lower limit quoted is about 2 m below the limestone bed at 45.50 m in the foreground. In the background at the top of the Akhdar marls are the first limestone banks of the Gourbeuj Member (G).
figure 3 (negative 267-37). The transition between the Akhdar marls (A) and the alternances of the Gourbeuj Member (G) in the partial section ELE; Upper Campanian.
The upper limit of the Akhdar marls is well-exposed in this outcrop which shows the transition to the alternances of the Gourbeuj Member, at the base of the upper bar of the Abiod Formation. It is in the final beds of these Akhdar marls that the first examples of the planktonic foraminifer Gansserina gansseri appear (formerly reputed to mark the "middle" Maastrichtian).
Planche 6 : La partie inférieure de la Formation Abiod ŕ Ellčs (photos F. ).
figure 1 (cliché 265-26). La barre inférieure de l'Abiod dans la coupe partielle ELD (sur le thalweg Est de l'oued Ed Dam) ; partie inférieure élevée du Campanien.
Au premier plan affleurent les derniers bancs de calcaires massifs du Membre Haraoua (H) de la barre inférieure de la Formation Abiod. Aux environs de ELD 70 se situe le passage entre les calcaires du Membre Haraoua et les alternances calcaires-calcaires marneux du Membre Mahdi (M) oů se signalent les premičres empreintes d'ammonites du groupe Nostoceras (Bostrychoceras).
À droite du pylône électrique, les trois petites flčches blanches indiquent la position de la route goudronnée oů se trouve un véhicule.
figure 2 (cliché 267-16). Les marnes intermédiaires du Membre Akhdar dans la coupe partielle ELE (sur le thalweg Nord de l'oued Ed Dam, au sud-est de la route goudronnée) ; partie supérieure du Campanien.
Les marnes vert turquoise sont dominantes dans le Membre Akhdar (A) par rapport aux bancs de calcaire marneux et de calcaire. À noter que le foraminifčre planctonique Radotruncana calcarata est présent au moins dčs ELE 43,50 m - soit 2m environ sous le banc ŕ 45,50 m au premier plan - et se poursuit au moins jusqu'ŕ 73,70 m. À l'arričre plan, au sommet des marnes Akhdar viennent les premiers bancs calcaires du Membre Gourbeuj (G).
figure 3 (cliché 267-37). La transition entre les marnes Akhdar (A) et le Membre des alternances Gourbeuj (G) dans la coupe partielle ELE ; partie supérieure du Campanien.
Le sommet des marnes Akhdar est bien exposé dans cet affleurement qui montre le passage aux alternances du Membre Gourbeuj, ŕ la base de la barre supérieure de la Formation Abiod. C'est dans les derniers bancs de ces marnes Akhdar qu'apparaissent les premiers exemplaires de l'espčce de foraminifčre planctonique Gansserina gansseri (autrefois réputée pour marquer le Maastrichtien "moyen").

Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Plate 7: The upper part of the Abiod Formation and the transition to the marls of the El Haria Formation (photos F. ).
figure 1 (negative 268-15). The upper bar of the Abiod Formation in partial section ELF (upstream end of wadi Ed Dam), upper part of the Campanian and basal Maastrichtian.
The waterfall shows a progressive change at around the ELF 30 level from the alternances of the Gourbeuj Member (G) to the limestone beds with very thin intercalations of marly limestone of the Ncham Member (N); it is on this G/N line that Moheddine stands to give the scale. Going up from that line in the bed of wadi Ed Dam (past the photo) level ELE 48 yielded the ammonite Pseudokossmaticeras brandti characteristic of the upper part of the Campanian. By comparison with the section at Kalaat Senan. ( et alii, 2000), the Campanian (Cp)- Maastrichtian (Ma) boundary is above this level, between ELF 60 and 70, probably around ELF 66.
figure 2 (negative 275-18). The boundary between the Abiod Formation (AB) and the El Haria Formation (EH) in partial section ELG (upper part of wadi El Kerma); middle portion of the Maastrichtian.
The upstream portion of wadi El Kerma is in the immediate foreground, running obliquely toward the lower right corner of the photo, following a narrow band of fairly continuous vegetation.
On the left bank of wadi El Kerma in the foreground on the right are the limestone-marl alternances of the Gouss Member (G), the highest of which yielded ammonites, among them a Pachydiscus neubergicus (von ). At level ELG 17.20 m, the blue-black marls of the El Haria Formation rest on the last limestone bed of the Gouss Member. The very weak resistance to erosion of the El Haria marls caused the last bed of limestone to be well exposed and to crop out over a large surface, however without any of the characteristics of a hardground (it is the upper surface of this bed that et alii, 1999, fig. 4a, p. 240 erroneously interpreted as a hardground. The lowermost levels of the El Haria marls too contain no glauconite, phosphate or quartz grains but have millimeter-sized bioclasts of bivalves and echinoderms which makes the marls feel grainy.
Higher up, five thin beds of limestone resting on two small beds of marly limestone form a ridge in the topography (the last of these beds is at level 36) that et alii, 1999, fig. 4B, p. 240 place erroneously their "Campanian-Maastrichtian boundary". That is, above the upper bar of the Abiod, and about 150 m above the levels with Radotruncana calcarata in the lower and middle portions of the Akhdar marls.
Above level 36 the El Haria marls continue, their upper portion of Paleocene age.
Planche 7 : La partie supérieure de la Formation. Abiod et la transition aux marnes de la Formation El Haria (photos F. ).
figure 1 (cliché 268-15). La barre supérieure de la Formation Abiod dans la coupe partielle ELF (amont de l'oued Ed Dam) ; partie supérieure du Campanien et base du Maastrichtien.
La cascade de base montre un passage progressif - aux environs du niveau ELF 30 - entre les alternances du Membre Gourbeuj (G) et les bancs calcaires ŕ trčs minces interbancs de calcaire marneux du Membre Ncham (N) ; c'est sur cette ligne G/N que se trouve Moheddine qui donne l'échelle. En remontant sur la droite le lit de l'oued Ed Dam (hors photo), le niveau ELE 48 a livré l'ammonite Pseudokossmaticaras brandti caractéristique de la partie supérieure du Campanien. Par comparaison avec la coupe de Kalaat Senan ( et alii, 2000), la limite Campanien (Cp)- Maastrichtien (Ma) se trouve au-dessus de ce niveau, entre ELF 60 et 70, soit vers ELF 66.
figure 2 (cliché 275-18). La limite entre les Formations Abiod (AB) et El Haria (EH) dans la coupe partielle ELG (partie amont de l'oued El Kerma) ; partie moyenne du Maastrichtien.
L'amont de l'oued El Kerma se trouve au tout premier plan, allant obliquement de la gauche vers le coin inférieur droit de la photo, en suivant une petite bande de végétation assez continue.
En rive gauche de l'oued El Kerma, affleurent au premier plan ŕ droite les alternances calcaires-marnes du Membre Gouss (G) dont les derničres ont livré des ammonites dont un Pachydiscus neubergicus (von ). Au niveau ELG 17,20m, les marnes bleu-noir de la Formation El Haria reposent sur le dernier banc calcaire du Membre Gouss. La trčs faible résistance ŕ l'érosion des marnes El Haria fait que le sommet du dernier banc calcaire est bien dégagé et affleure sur une grande surface, sans présenter toutefois les caractčres d'un hardground : pas de recristallisation, pas de glauconie ni de grains phosphatés (c'est le sommet de ce banc qui est interprété erronément comme un hardground par et alii, 1999, fig. 4a, p. 240). La base des marnes El Haria ne montre pas non plus de glauconie ni de phosphate ou de grains de quartz mais contient des bioclastes millimétriques de lamellibranches et d'échinodermes, ce qui lui confčre un toucher graineux.
Plus haut, cinq petits bancs de calcaires superposés ŕ deux petits bancs de calcaires marneux forment un ressaut dans la topographie (le dernier de ces bancs se trouve au niveau 36) et ont livré plusieurs empreintes d'ammonites appartenant au genre Pachydiscus (c'est au sommet de ce banc 36 que et alii, 1999, fig. 4b, p. 240, placent erronément leur "Campanian-Maastrichtian boundary" c'est-ŕ-dire au-dessus de la barre supérieure de l'Abiod, ŕ environ 150 m au-dessus des niveaux ŕ Radotruncana calcarata de la partie inférieure ŕ moyenne des marnes Akhdar).
Au-dessus du niveau 36 on voit encore la suite des marnes El Haria dont la partie supérieure appartient au Paléocčne.