![]()
Contents
[Introduction and geological setting] [Phycological
content of the ...]
[Systematic paleontology] [Conclusions] and ...
[Bibliographic references]
* Corresponding author
Department of Geology, Faculty of Sciences, Islamic Azad University-Mashhad branch, Mashhad (I.R. Iran);
Young Researchers Club
71 chemin de Planta, 1223 Cologny (Switzerland)
Department of Geology, Faculty of Sciences, Islamic Azad University-Mashhad branch, Mashhad (I.R. Iran)
Center of Excellence for Paleontology, Ferdowsi University of Mashhad, Mashhad (I.R. Iran)
Manuscript online since August 27, 2010
![]()
Abundant, diversified algal assemblages comprising more than 25 species of Dasycladales are described from Barremian-Aptian limestone deposits of the Tirgan Formation in NE Iran. New species are described: one of them possibly belongs to a new endosporate genus, tentatively assigned the Triploporellaceae; two others, Clypeina ? sp. 1 and Rajkaella ? sp. 1, are left in open nomenclature. The widely distributed, locally abundant Montiella ? elitzae is revisited, because of the presence of a complete, exceptionally well preserved specimen. Yet to be demonstrated, the species is a junior synonym of Turkmenaria adducta , also originally described from the Kopet Dagh. Other species found in the Arkan section have already been reported from remote locations, primarily in Europe, in areas corresponding to the Northern and/or Southern Tethyan domains. Some of them are known only from the Hauterivian and/or the Barremian, excluding the Aptian, thus dating the lower part of the Arkan section as Barremian, but not excluding the Late Hauterivian.
Calcareous algae; Dasycladales; new taxon; Lower Cretaceous; Tirgan Formation; Iran.
M., M.A., A.A. & A.R. (2010).- Barremian-Aptian Dasycladalean algae, new and revisited from the Tirgan Formation in the Kopet Dagh, NE Iran.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/05 (CG2010_A05)
Algues dasycladales du Barrémien-Aptien, nouvelles ou révisées, de la Formation Tirgan dans le Kopet Dagh, NE Iran.- Des relevés détaillés entrepris près d'Arkan, dans la region de Bojnurd, au Kopet Dagh, montrent la présence de plus de 25 espèces de Dasycladales dans les calcaires de la Formation de Tirgan. En grande partie, ces algues sont déjà connues en Europe, dans l'Hauterivien supérieur, le Barrémien et l'Aptien des domaines Nord- et Sud-Téthysiens. Grâce à elles, la partie inférieure de la coupe d'Arkan peut être datée du Barrémien, sans pour autant exclure l'Hauterivien supérieur. Un nouveau taxon, sans doute endospore, est décrit sur la base d'une seule section. Deux espèces, Clypeina ? sp. 1 et Rajkaella ? sp. 1, sont décrites de manière informelle. Enfin, une espèce bien connue, Montiella ? elitzae, est particulièrement abondante. Peut-être s'agit-il d'un synonyme récent de Turkmenaria adducta , provenant également du Barrémien de la même région. Ici, l'espèce fait l'objet d'une description plus détaillée, fondée en particulier sur un spécimen complet permettant de mettre en question l'interprétation taxonomique proposée jusqu'ici.
Algues calcaires ; Dasycladales ; nouveau taxon ; Crétacé inférieur ; Formation de Tirgan ; Iran.
Abundant, diversified algal assemblages are described from Barremian-Aptian limestone deposits of the Tirgan Formation in NE Iran. The site is at Arkan, near Bojnourd, in the Kopet Dagh fold belt, at the SW margin of the Turan Platform.
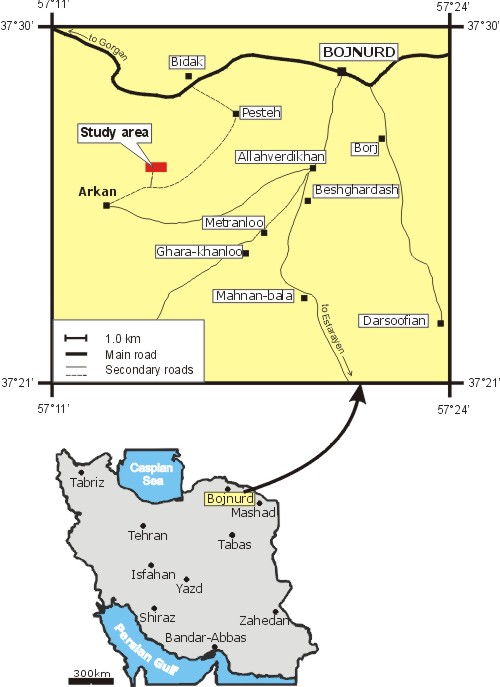
Click on thumbnail to enlarge the image.
Figure 1: Location of the studied section.
Fundamental differences in the crustal character and age of basement consolidation allow three major structural units to be distinguished, separated by ophiolitebearing sutures. Other criteria such as structural style, the age and intensity of deformation, the age and nature of magmatism are used to subdivide these major zones into smaller elements. The three major units and their main constituents are as follows:
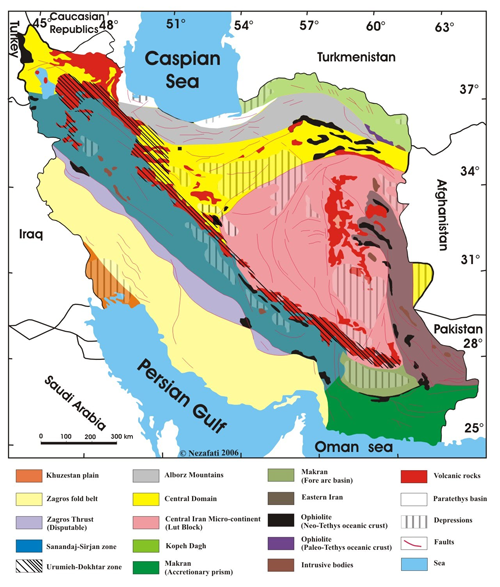
Click on thumbnail to enlarge the image.
Figure 2: Major geological subdivisions of Iran after (1968) and (1976), modified [copyright N. (2006), duplicated with permission].
The active fold belt of NE Iran, Kopet Dagh, was formed on a Hercynian metamorphosed basement, at the SW margin of the Turan Platform. The belt consists of about 10 km of Mesozoic and Tertiary sediments, mostly carbonates. Like the Zagros, it was rucked into long, linear NW-SE trending folds during the last Plio-Pleistocene phase of Alpine orogenesis. No magmatic rocks crop out in the Kopet Dagh except those in the basement of the Aghdarband Window and some Triassic basic dikes.
Barremian deposits are up to 300 m thick. They consist of alternating beds of calcareous sand and sandy oolitic limestone. Microfossils include bryozoa, algae and foraminifers with Gavelinella barremiana, Praehedbergella infracretacea, Globorotalites intercedens, Lenticulina ouachensis, Marssonella oxycona, Tritaxia tricarinata and T. pyramidata.
Aptian deposits consist of up to 1100 m of mainly massive limestones. Quoting and (1991), foraminifera include Ammobaculites reophacoides, Astacolus cf. schloenbachi, Brotzenia cretosa, B. spinulifera polypioides, Citharina aptiensis, Dentalina distincta, Gaudryina dividens, Globorodrospira djaffaensis, Textularia foedu, Verneuilinoides subfiliformis, Orbitolina kurdica, O. discoidea, and O. conica.
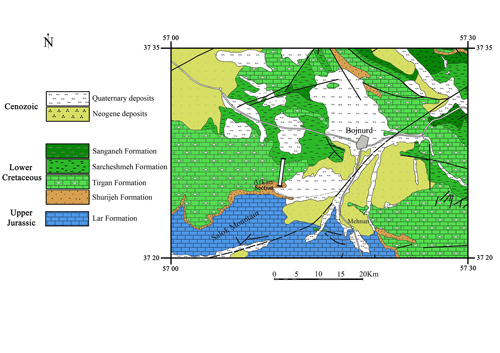
Click on thumbnail to enlarge the image.
Figure 3: Geological map of the Bojnurd area, with the location of the Arkan section.
This formation is named from its exposure in the Tirgan Valley of Kopet Dagh. It was introduced by geologists of the National Iranian Oil Company for a feature-forming unit of massively bedded, oolitic and organodetrital limestone occurring throughout the Kopet Dagh ranges. The 700 m Tirgan type-section is 39 km southeast of Dargaz and consists mainly of mid-sized to thick beds of grey fossiliferous limestone (, 1994). For the eastern part of Kopet Dagh, (1969) indicates the thickness of Tirgan Formation to be only 50 m. It overlies the Shurijeh Formation and underlies the Sarcheshmeh Formation; both contacts are conformable but a transitional interfingering between the Tirgan and the Shurijeh exists in southeastern Kopet Dagh. The Tirgan Formation is essentially Neocomian, locally extending as high as the Aptian.
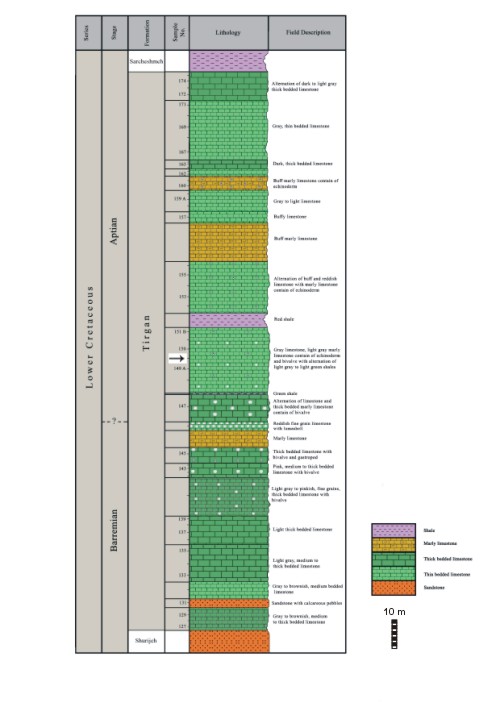
Click on thumbnail to enlarge the image.
Figure 4: Geological field section at Arkan, Tirgan Formation. Arrow: stratum typicum of Steinmanniporella ? parsica, n.sp.: sample 149b.
The Bojnourd study area (Fig. 3 ![]() )
of the Kopet Dagh is located in the Northern Khorasan province. In this area, several outcrops of the Cretaceous Shurijeh, Tirgan, Sarcheshmeh, Sanganeh and Abderaz formations are
present. The section (Fig. 4
)
of the Kopet Dagh is located in the Northern Khorasan province. In this area, several outcrops of the Cretaceous Shurijeh, Tirgan, Sarcheshmeh, Sanganeh and Abderaz formations are
present. The section (Fig. 4 ![]() ) is near Arkan, 10 km southwest of
Bojnourd, Khorasan-e-shomali province (Figs. 1
) is near Arkan, 10 km southwest of
Bojnourd, Khorasan-e-shomali province (Figs. 1 ![]() - 2
- 2 ![]() ). In this area, the strike is E-W with a
dip of 15-35 degrees. The 195 m Tirgan Formation rests on sandstones of the Shurijeh Formation.
The contact is concordant and transitional. The Tirgan Formation, characterized
mainly by the presence of orbitolinids, consists of discrete levels of oolitic limestone, fossiliferous limestone,
and some marly limestones, marls and argillaceous limestones. It is overlain by
the shales and limestones of the Sarcheshmeh Formation. Lithologies found in the Tirgan type-section are also found in the Arkan section (Fig. 3
). In this area, the strike is E-W with a
dip of 15-35 degrees. The 195 m Tirgan Formation rests on sandstones of the Shurijeh Formation.
The contact is concordant and transitional. The Tirgan Formation, characterized
mainly by the presence of orbitolinids, consists of discrete levels of oolitic limestone, fossiliferous limestone,
and some marly limestones, marls and argillaceous limestones. It is overlain by
the shales and limestones of the Sarcheshmeh Formation. Lithologies found in the Tirgan type-section are also found in the Arkan section (Fig. 3 ![]() ). In
the lower half of this section up to 50% of the biota consists of calcareous algae (Fig. 5
). In
the lower half of this section up to 50% of the biota consists of calcareous algae (Fig. 5 ![]() ), essentially Dasycladales, Halimedaceae and Bryopsidales. The
alphabetically-listed Dasycladales that follow were identified under the microscope, mainly in the thick limestone
beds of the Tirgan Formation, but not all of them are illustrated. The generalized range
of each is also given.
), essentially Dasycladales, Halimedaceae and Bryopsidales. The
alphabetically-listed Dasycladales that follow were identified under the microscope, mainly in the thick limestone
beds of the Tirgan Formation, but not all of them are illustrated. The generalized range
of each is also given.
Other algae and incertae sedis include: abundant Halimedaceae, Arabicodium sp., Boueina sp., Cayeuxia
sp., Coptocampylodon lineolatus (Fig. 6 (d) ![]() ), Ethelia cf. alba, Palaeosiphonium sp., Permocalculus sp., Vermiporella ? sp.
(Fig. 6 (d)
), Ethelia cf. alba, Palaeosiphonium sp., Permocalculus sp., Vermiporella ? sp.
(Fig. 6 (d) ![]() ) and Juraella bifurcata.
This last species belongs to the Bryopsidales. It is dealt with separately, in et alii
(2009).
) and Juraella bifurcata.
This last species belongs to the Bryopsidales. It is dealt with separately, in et alii
(2009).
Based solely on the known distribution of these Dasycladales at Arkan, the lower part of the Tirgan Formation is Barremian but not excluding the Late Hauterivian. Higher, its age is Barremian-Aptian, as confirmed by the echinoids including Heteraster spp. and Toxaster spp., in marls and marly limestones. The depositional environment is open marine shallow water throughout.
Click on thumbnail to enlarge the image.
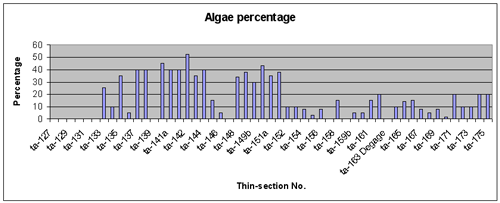
Figure 5:
Arkan section (Kopet Dagh), Tirgan Formation, Barremian / Aptian: quantitative distribution of the dasycladalean and other calcareous green algae. Values are expressed in relative
percentages with respect to the total biotic content. Counts were made using the
microscope. Sample distribution is shown in Fig. 4 ![]() .
.
Click on thumbnail to enlarge the image.
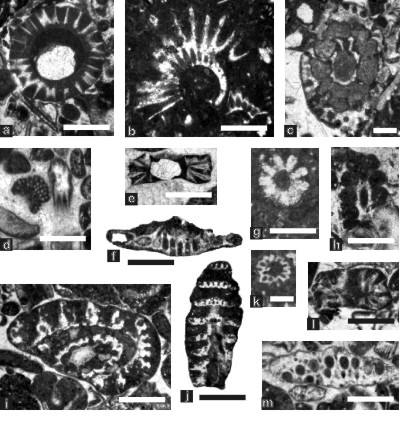
Figure 6: Dasycladalean algae from the Tirgan Formation, Kopet Dagh; fig. a: transversal section of Triploporella ? sp. (secondary laterals uncalcified or missing), sample 174, scale bar 0.7 mm; fig. b: incomplete, almost transversal section of Clypeina cf. gigantea (another, smaller specimen is visible above the scale bar), sample 140, scale bar 0.7 mm; fig. c: oblique-transversal section of Conradella bakalovae, sample 135, scale bar 0.4 mm; fig. d: two incertae sedis, with Vermiporella ? sp. (left) and a tangential section of Coptocampylodon cf. lineolatus (right), sample 152, scale bar 0.7 mm; fig. e: axial section of Praturlonella cf. dalmatica, sample 133, scale bar 0.6 mm; fig. f: tangential section of Actinoporella podolica, sample 135, scale bar 0.75 mm; fig. g: almost transversal section of Clypeina ? cf. C. solkani, sample 150, scale bar 0.7 mm; fig. h: oblique section of Holosporella sp., sample 135, scale bar 0.75 mm; fig. i: broken oblique section of Clypeina ? n.sp. 1 enclosing a smaller specimen of Otternstella ? sp., sample 142, scale bar 0.4 mm; fig. j: oblique-tangential section of Clypeina ? n.sp. 1, sample 142, scale bar 0.74 mm; fig. k: transversal section of Praturlonella nerae, sample 150, scale bar 0.6 mm; fig. l: axial section of Praturlonella aff. P. nerae, sample 135, scale bar 0.6 mm; fig. m: tangential section of Neomeris ? cf. N. cretacea; sample 138, scale bar 0.6 mm.
Click on thumbnail to enlarge the image.
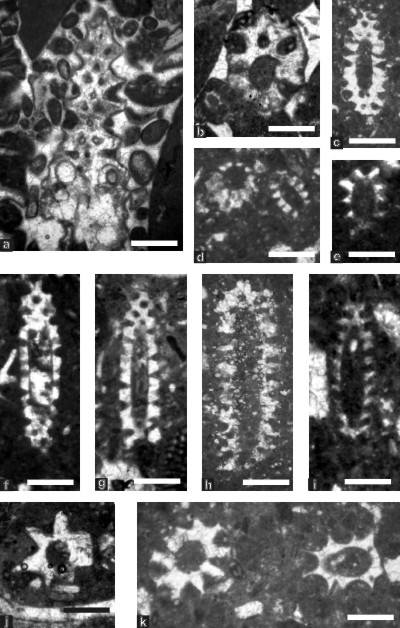
Figure 7: Species of genus Salpingoporella (Dasycladales) from the Tirgan Formation, Kopet Dagh; fig. a: oblique-tangential section of Salpingoporella cemi, sample 152, scale bar 0.7 mm; fig. b: oblique section of S. cemi, sample 166, scale bar 0.7 mm; fig. c: oblique section of Salpingoporella cf. S. piriniae, sample 151a, scale bar 0.55 mm; fig. d: two sections of Salpingoporella cf. S. piriniae, sample 150, scale bar 0.7 mm; fig. e: oblique section of Salpingoporella cf. S. parapiriniae sample 140, scale bar 0.6 mm; fig. f: oblique section of Salpingoporella muehlbergii, sample 140, scale bar 0.6 mm; fig. g: oblique section of S. muehlbergii, sample 148, scale bar 0.7 mm; fig. h: oblique section of Salpingoporella sp., sample 151b, scale bar 0.7 mm; fig. i: oblique section of Salpingoporella sp., sample 145, scale bar 0.75 mm; fig. j: almost transversal section of Salpingoporella aff. S. hasi; sample 145, scale bar 0.8 mm; fig. k: to the left, oblique section of Salpingoporella aff. S. hasi; to the right, oblique section of Holosporella ?, sample 150, scale bar 0.7 mm.
Altogether 85 samples were collected and 108 thin-sections were prepared. Materials (rock samples and thin-sections) are housed in the department of geology, Islamic Azad University, Mashhad branch.
Ordo Dasycladales , 1931
Currently, the nomenclature and assignments of fossil Dasycladales with a rank higher than genus necessitates an ad hoc revision. The following tentative hierarchy is provisional.
Familia Dasycladaceae (, 1843) & , 1992
Tribus Dasycladeae , 1920
Genus Montiella (L. & J. , 1922) , 1987
The genus Montiella was assigned by et alii (1979) to the Neomereae (). Here, the genus is placed in the Dasycladeae, in compliance with and (1992, p. 50). Currently, Montiella includes six species, ranging in age from Hauterivian to Paleocene ( & , 1993; & , 1994). In the Paleocene type-species M. munieri, the presence of two types of secondary laterals, respectively fertile and sterile, terminal to first order laterals is inferred, yet not physically demonstrated. In view of this equivocal situation, the genus Bakalovaella , 1993 (type-species, B. elitzae; see synonymy below) will possibly prove to be a well-founded taxon rather than a junior synonym of Montiella.
Montiella ? elitzae (, 1971) , 1980
Selected citations:
? 1960 Turkmenaria adducta gen. et sp. n. V.P., 1960, p. 1036, fig. 1b, fig. 2.
1971 Cylindroporella elitzae sp. n. D., p. 126, Pl. 3.
1978 Cylindroporella barbui n.sp. O., p. 126, text-fig. 2 (a-e).
1980 Montiella? elitzae () nov. komb. R., p. 114, pls. I-IV; figs. 1, 2 [reconstruction].
1993 Bakalovaella elitzae () nov. comb. I.I., p. 100, pl. 7. [Inclusive of a comprehensive synonymy].
1995 Montiella elitzae. J.-P. & A., pl. 1, figs. 4-5.
2007 Montiella elitzae. I.I., p. 11.
Turkmenaria adducta comes from Barremian deposits in the Sviryuzinskoye Gorge, Turkmen Central Kopet Dagh. Although formally valid, it unfortunately remains ill-described, so Montiella? elitzae might be a junior synonym. The species ranges from the Late Hauterivian to the Albian, usually in open marine deposits. Taxonomically, it is closely related to the Barremian-Aptian Montiella benizarensis (), and the Cenomanian-Turonian Montiella filipovici .
The following, simplified description is based on (1993, p. 100-102, under Bakalovaella elitzae): Thallus cylindrical, non-constricted, bearing numerous, contiguous whorls of short tubular primary laterals, disposed horizontally and form quincunxes. Main axis cylindrical or intusannulated, somewhat larger between the whorls. Two types of secondary laterals are clustered at the tip of the primaries: (1) one (single) large sub-spherical or egg-like fertile appendage (gametophore), oblique to the axis, free or occasionally slightly deformed, on top of a short and narrow proximal attachment; (2) below the gametophore, one (single) secondary lateral, horizontal, commonly slightly twisted, first slender and tubular, then progressively expanding, occasionally compressed by the adjacent gametophores, triangular in section. Gametangia uncalcified. Dimensions, partly modified from (1993): L = up to 7 mm, D = 0.51-1.24 mm, d = 0.12- 0.34 mm, d/D = 13-37%, h = 0.17-0.23 with up to 42 whorls, l = 0.09-0.15 mm, p = 0.09-0.11 mm, l' (sterile) = 0.23-0.25 mm, p' (sterile) = 0.09 mm, l' (fertile) = 0.23-0.25 mm, p' (fertile) = 0.19-0.26 mm, w = 6-9.
At Arkan, numerous sections of Montiella? elitzae are found in
several thick-bedded intervals throughout the Tirgan Formation. Of special interest is the
fully 7 mm-long, flexuous specimen depicted in Fig. 8 (a) ![]() . In the lower, right hand side of the picture, gametophores are missing in the narrow,
basal part of the thallus presumably close to the rhizoid. A similar section is depicted in
Fig. 8 (d)
. In the lower, right hand side of the picture, gametophores are missing in the narrow,
basal part of the thallus presumably close to the rhizoid. A similar section is depicted in
Fig. 8 (d) ![]() . At top of the Fig. 8
(a)
. At top of the Fig. 8
(a) ![]() , the longitudinal section cuts the acuminated, fertile tip of the lime skeleton. The sterile secondary laterals
appear to be missing, causing a striking resemblance with certain sections of Holosporella spp. Apparently, the uncalcified, sterile vertex of the alga decayed before
the vegetative phase ended, prior to the formation of gametophores.
, the longitudinal section cuts the acuminated, fertile tip of the lime skeleton. The sterile secondary laterals
appear to be missing, causing a striking resemblance with certain sections of Holosporella spp. Apparently, the uncalcified, sterile vertex of the alga decayed before
the vegetative phase ended, prior to the formation of gametophores.
Here, the traditional interpretation of the taxonomy of Montiella? elitzae is retained, although elaborated. However, many of the sections of this alga also suggest the following alternative interpretation: the fertile appendages (gametophores) are lateral in position to the proximal part of the primaries. In other terms, no sterile secondary laterals would exist, a situation found, for example, in the Recent Bornetella nitida.
Click on thumbnail to enlarge the image.
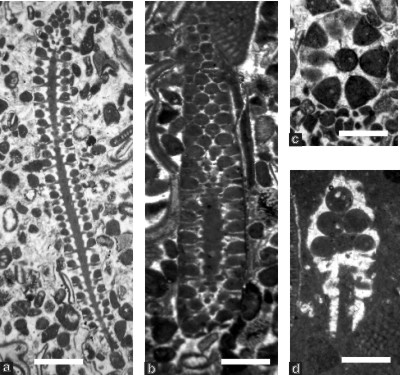
Figure 8: Montiella ? elitzae (Dasycladales), from the Tirgan Formation, Kopet Dagh; fig. a: full longitudinal section, sample 149b, scale bar 2.3 mm; fig. b: oblique-tangential section, sample 174, scale bar 0.7 mm; fig. c: almost transversal section, sample 140, scale bar 0.6 mm; fig. d: oblique-tangential section cutting the basal part of the alga, sample 165, scale bar 0.8 mm.
Familia Triploporellaceae (, 1920) & , 1992, sensu et alii, 2008
Quoting and (1992, table 2.4) "This family is characterized by euspondyl arrangement of laterals and cladospore formation of gametangia". According to et alii (2008) "Although the kind of reproduction (endosporous or cladosporous) is a high taxonomic character, their evidence is lacking in most of fossil taxa (...) the occurrence of thin and delicate laterals, irrespectively of whether trichophorous or phloiophorous, often associated with a bulky central stem, is considered a proof of an endosporous reproduction (for a discussion see , 1997). (...) Consequently, the diagnosis of Triploporellaceae ought to include both endosporous and cladosporous reproduction."
Tribus Salpingoporelleae et alii, 1979
Genus Salpingoporella ( in , 1918) et alii, 2006
At least eight species of Salpingoporella (Fig. 7
![]() ) are present in the Tirgan Formation.
Of them only two, S. cemi and S. muehlbergii, are identified with
certainty.
) are present in the Tirgan Formation.
Of them only two, S. cemi and S. muehlbergii, are identified with
certainty.
Genus Clypeina , 1845
Clypeina ? sp. 1
The two illustrated specimens, the only ones available, apparently are the same taxon. Provisional description: wide axial main axis, cylindrical and/or intusannulated, weakly calcified, presumably fertile (endosporate), bearing spaced out whorls of numerous, strongly calcified primary laterals, flexuous, almost horizontal, progressively widening out (phloiophorous), open (uncalcifed) at tip; irregular presence of slender filaments, coating the axis and/or the laterals. Dimensions: D = ab. 0.9 mm; d = ab. 0.47 mm; d/D = ab. 50%; h = ab. 0.3 mm; w = 28-32; p max. = 0.07 mm.
Tribus Triploporelleae (, 1920) et alii, 1979
Genus Rajkaella & , 1988
The lastest, emended diagnosis was introduced by and (1993), under the name Radoiciciella. Excerpts of the description are as follows: "(...) slightly calcified whorls spaced out along an uncalcified (or extremely poorly calcified) axial cavity. Whorls made up of primary and secondary branches. Primaries, long and cylindrical (tubular), sometimes slightly dilated distally; they are perpendicular or more or less inclined to the axial cavity. Secondaries, shorter and widening distally; they occur as a bush around the distal end of primaries. Sometimes the branches may be coalescent either proximally or on a longer part of their tract (...). Due to the poor calcification of the axial cavity, the alga is fragile and may be found only as fragments representing branches or part of them." So far, there are six valid species of Rajkaella, ranging in age from Tithonian to Aptian.
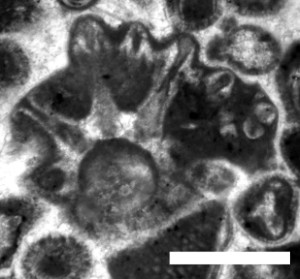
Rajkaella ? sp. 1
A provisional description is based on the one specimen (illustrated) available: narrow main axis bearing whorls of weakly calcified primary laterals, dumb-bell shaped, presumably fertile (cladosporate), bent at tip, forming imbricate (overlapping) calcified quincunxes. Short, phloiophorous secondary laterals are clustered at tip of the primaries. Dimensions: D = 1.7 mm; d = 0.42 mm; d/D = 25%; w = 9; w' = approx. 9; p = 0.15 mm; p' = 0.06 mm; l = 0.65 mm; l' = 0.06 mm. These values do not widely differ from the known species of Rajkaella. However, in Rajkaella? sp. 1 the secondary laterals are fewer and shorter.
Click on thumbnail to enlarge the image.
Figure 9: Nearly transversal section of Rajkaella ? sp. 1 (Dasycladales), Tirgan Formation, Kopet Dagh, sample 153, scale bar 0.7 mm.
Genus Steinmanniporella et alii, 2009
The genus Steinmanniporella with its type-species the Late Jurassic S. kapelensis ( & ), was recently created to include four species ranging in age from Late Jurassic to Paleocene originally assigned the genus Linoporella . Only two orders of laterals are found in these species, instead of the three in Linoporella. The diagnosis in et alii (2009) is as follows: "Cylindrical thallus with euspondyl tubular primary laterals giving rise to a tuft of secondaries. The tubular secondary laterals enlarge towards the distal end (phloiophorous type), that most probably, formed a cortex at the outer end of the thallus. Reproductive structures were situated very probably inside the central stem (endosporate)."
In terms of formal description, these general characters fit well with Steinmanniporella? parsica, n.sp., but the secondary laterals are horn-shaped, that is phloiophorous throughout their length, instead of being tubular first and phloiophorous at the tip. Such a minor difference is considered significant only at the species level. Conversely, when at the species recently assigned to Steinmanniporella, e.g. S. kapelensis illustrated in et alii (2009, fig. 1a), are examined, differences from Steinmanniporella? parsica, n.sp., are striking: in all of these previously described species the primary laterals are arranged in rather close whorls, are rather short and stout, with quite long, thin secondary laterals that are phloiophorous at tip. In other terms, our new species may also be a new genus. Here, for convenience, it is provisionally assigned Steinmanniporella and thus avoids the creation of a genus based on a single specimen.
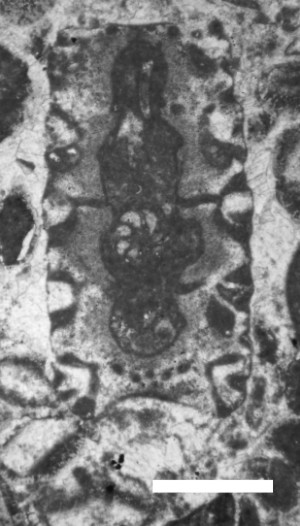
Steinmanniporella ? parsica, n.sp.
Holotype: the sole specimen is a slightly oblique, sub-axial section depicted in
Fig. 10 ![]() . Sample 149b, Arkan section,
in the middle of the Tirgan Formation assigned the Aptian.
. Sample 149b, Arkan section,
in the middle of the Tirgan Formation assigned the Aptian.
Derivatio nominis: after Persia (= Pars).
Diagnosis: thallus cylindrical, euspondyl. Axial cavity intusannulated. Spaced out whorls of numerous, slender, acrophorous primary laterals, perpendicular to or slightly tilted upward from the main axis. Secondary laterals horn-shaped (phloiophorous), distally uncalcified, approximately eight in number, clustered at the tip of the primaries, forming a rosette. Calcareous skeleton conspicuous, forming a sleeve, originally aragonitic. Gametangia or gametophores uncalcified, assumed to have been located in the main axis (endosporate-type).
Description: The specimen is about 1 mm wide (D value), with five whorls of about twenty primary laterals. The wide axial cavity is clearly intusannulated, although the width (d value) between the whorls is increased by some process of microbioerosion that possibly destroyed an original proximal attachment of the primary laterals. The approximately 0.17 mm-long primary laterals are only 0.02 to 0.03 mm-wide. They are flexuous, increase very slightly in diameter, or maintain their width throughout their length. Approximately eight horn-shaped (phloiophorous) secondary laterals are clustered at the tip of the primaries, forming a conspicuous, bowl-shaped and gently-tilted downward rosette. Except at the periphery of the rosettes, the mucilage coat of the alga is entirely calcified, and even more heavily around the laterals. The specimen is irregularly worn at its periphery. In the better preserved lower part of the section (the lower part of Fig. 10), the rosettes touch suggestive of a distal, polygonal cortex like that of the Recent genus Bornetella. However, this is unlikely, because in the living alga the rosettes are irregularly bent downward by their weight so are probably not connected to each other.
Comparisons: Steinmanniporella ? parsica, n.sp., is difficult to compare with any other species, except perhaps Balkhanella hurkai , 1993. The so far mono-specific genus Balkhanella is possibly also endosporate. It was established on the basis of a single nice specimen from Hauterivian bioclastic deposits in the Bolshoi Balkhan, Turkmenia. In Balkhanella hurkai, the thallus is clearly articulated, as shown by the orientation of the laterals that form clusters. The wide main axis is slightly intusannulated, heavily calcified, somewhat narrower between the whorls. Possibly it has bulging proximal features, perhaps more like vestibules than slender primary laterals.
Click on thumbnail to enlarge the image.
Figure 10: Sub-axial section of Steinmanniporella ? parsica, n.sp. (Dasycladales), Tirgan Formation, Kopet Dagh, sample 149b, scale bar 0.7 mm.
Barremian - Aptian limestones of the Tirgan Formation yield rich assemblages of calcareous algae, including over 25 species of Dasycladales. In the lower part of the Arkan section, three species, namely Actinoporella podolica, Praturlonella dalmatica and Clypeina gigantea, suggest a Barremian age, but do not exclude the Late Hauterivian. In the upper part of the section the presence of Praturlonella nerae is indicative of the Aptian. Taxonomy of the particularly abundant Montiella ? elitzae, is reviewed based on the finding of a complete specimen. Two species, Clypeina ? sp. 1 and Rajkaella ? sp. 1 are informally described and a new, presumably endosporate taxon, Steinmanniporella ? parsica, n.sp., is introduced. The one specimen available has a unique combination of characters, possibly requiring the erection of a new genus of Triploporellaceae. As mentioned by other authors, the family should be emended to include endosporate taxa.
The authors wish to thank Prof. Ioan (University of Cluj-Napoca) for his careful review of the manuscript and helpful suggestions. We thank M.Sc Abbas from the Center of Excellence for Paleontology, Ferdowsi University of Mashhad for his contributions in the field. Also, we thank the Department of geology, Islamic Azad University, Mashhad branch for supplying field equipment. Dr. Nicolaos (IGME, Athens) advised on the taxonomy of certain species. Reviews of the English by Mr. (language school head of studies) and Hasmik (Ecocenter NAS RA) are gratefully acknowledged.
A. (1969).- History of oil exploration and brief description of the geology of the Sarakhs area and the anticline of Khangiran.- Bulletin of the Iranian Petroleum Institute, Tehran, vol. 37, p. 86-94 (in Persian).
A. (1994).- Geology of Kopet-Dagh.- Geological Survey of Iran publication, Tehran, 275 p. (in Persian).
D. (1971).- Nouvelles espèces de Dasycladaceae (Algae) dans les sédiments urgoniens du Prébalkan central.- Bulgarian Academy of Sciences - Committee of Geology, Bulletin of the Geological Institute, Sofia, (Series Paleontology), vol. XX, p. 123-127.
F., A. & R. (2008).- New dasycladalean algae from the Middle Norian (Upper Triassic) of northern Italy (Mt. Pramaggiore, Carnic Prealps).- Facies, Erlangen, vol. 54, p. 549-580.
J.-P., P., R., P., M. & D. (1979).- Essai de classification des Dasycladales en tribus. In : 2ème Symposium international sur les Algues fossiles (Paris, 23-26 avril 1979).- Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Pau, vol. 3, n° 2, p. 429-442.
S. & M.J. (1992).- Dasycladales - An illustrated monograph of a fascinating algal order.- Georg Thieme Verlag, Stuttgart - New York, 247 p.
I.I. (1993).- Some new or poorly known calcareous algae (Dasycladales, Gymnocodiaceae) in the Lower Cretaceous deposits from the Resita-Moldava Noua Zone (Southern Carpathians, Romania).- Revista Española de Micropaleontologia, Madrid, vol. 25, p. 93-126.
I.I. (2007).- Salpingoporella ? popgrigorei, a new species of Dasycladales (calcareous algae) from the Lower Aptian deposits of Padurea Craiului (Northern Apuseni Mountains, Romania).- Facies, Erlangen, vol. 53, p. 377-388.
I.I., B. & F. (2009).- Steinmanniporella, a new dasycladale genus name for "Linoporella" with two orders of laterals.- Facies, Erlangen, vol. 56, n° 2, p. 317-321.
N., M.A. & R. (2006).- Salpingoporella, a common genus of Mesozoic Dasycladales (calcareous green algae).- Revue de Paléobiologie, Genève, vol. 25, n° 2, p. 457-517.
P. (1997).- Introduzione allo studio in sezione sottile delle Dasicladali fossili.- Quaderni dell'Accademia Pontaniana, Napoli, n° 22, 262 p.
R. (1988).- Nouvelle taxonomie des Algues Dasycladales - New taxonomy of Dasycladale Algae.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, vol. 12, p. 165-217.
R. & B. (1993).- Inventaire des Algues Dasycladales fossiles. I° partie- Les Algues Dasycladales du Tertiaire.- Revue de Paléobiologie, Genève, vol. 11, n° 2 (1992), p. 331-356.
O. (1978).- Microfacies de la serie calcaires, cretacée inférieure d'Aliman (Dobrogea de sud).- Dari de Seama ale Sedintelor, Bucuresti, t. LXIV (1976-1977), 4. Stratigrafie, p. 107-136, 7 pls.
O. & I.I. (1988).- Comments on the genus Radoiciciella.- Analele Universitatii Bucuresti Geologie, vol. 37, p. 97-99.
O. & I.I. (1993).- The dasyclad genus Radoiciciella and its representatives from Romania.- Revista Española de Micropaleontologia, Madrid, vol. 25, p. 5-23.
P. (1987).- Les Chlorophycées calcaires du Paléogène d'Europe nord-occidentale (Bassin de Paris, Bretagne, Cotentin, Bassin de Mons).- Thèse de Doctorat d'État, Université de Nantes, 500 p.
B. & R. (1994).- Inventaire des Algues Dasycladales fossiles. II° partie- Les Algues Dasycladales du Jurassique et du Crétacé.- Revue de Paléobiologie, Genève, vol. 12, n° 1 (1993), p. 19-65.
J. (1994).- Cretaceous (Albian-Turonian) calcareous algae from Egypt and Jordan - Systematics, stratigraphy and paleogeography.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 50, p. 295-317.
F.T. (1843).- Phycologia generalis oder Anatomie, Physiologie und Systemkunde der Tange.- F.A. Brockhaus, Leipzig, p. 311-313.
V.P. (1960).- Novoye Vodorosli Mela Kopet-Daga (Turkmeniya) [New cretaceous algae of Kopet Dag (Turkmenia)].- Doklady Akademii Nauk SSSR, Moscow, Tom 134, n° 4, p. 939-941.
J.-P. & A. (1995).- Early Cretaceous calcareous algae of the Northwest Pacific guyots.- Proceedings of the Ocean Drilling Program, Scientific Results, College Station, vol. 144, p. 221-230.
H. (1840-1847).- Iconographie zoophytologique. Description par localités et terrains des Polypiers fossiles de France et pays environnants.- P. Bertrand, Paris, 348 p., 79 pls.
L. & J. (1922).- Nouvelle contribution à l'étude des Dasycladacées tertiaires.- Mémoires de la Société géologique de France, Paris, (Paléontologie), t. XXV, fasc. 2, N° 58, 33 p.
M.H. (1976).- An introduction to geology of Iran.- Geological Survey of Iran publication, Tehran, Iran, 110 p. (in Persian).
N. (2006).- Au-Sn-W-Cu-Mineralization in the Astaneh-Sarband Area, West Central Iran including a comparison of the ores with ancient bronze artifacts from Western Asia.- PhD, Eberhard-Karls-Universität Tübingen, 116 p.
A. (1931).- Systematische Übersicht über die mit Flagellaten in Zusammenhang stehenden Algenreihen und Versuch einer Einreihung dieser Algenstämme in die Stämme des Pflanzenreiches.- Beihefte zum Botanischen Centralblatt, Jena, Band XLVIII, Heft 3, p. 17-332.
J. von (1918).- Dasycladaceae. In: F. (ed.), Das Eozänvorkommen bei Radstat im Pongau und seine Beziehungen zu den Gleichalterigen Ablagerungen bei Kirchberg am Wechsel und Wimpassing am Leithagebirge.- Denkschriften der mathem.-naturw. Klasse, Wien, vol. 95, p. 171-278.
J. von (1920).- Die Siphoneae verticillatae vom Karbon bis zur Kreide.- Abhandlungen der zoologisch-botanischen Gesellschaft in Wien, Band XI, Heft 2, 259 p., 8 pls. [Les Siphonées verticillées du Carbonifère au Crétacé (1961). Editions Technip, Rueil-Malmaison, 258 p., 8 pls.].
R. (1980).- Contribution to the reconsideration of a group of cylindroporelliform dasyclads.- Bulletin de l'Académie Serbe des Sciences et des Arts, Beograd, (Classe des Sciences mathématiques et naturelles, Sciences naturelles), vol. 20, p. 109-115.
N.K. (1973).- Neocomian calcareous algae from Bolshoi Balkhan, USSR.- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, 1973, n° 11, p. 690-708.
J. (1968).- Structural history and tectonics of Iran; a review.- The American Association of Petroleum Geologists, Bulletin, Tulsa, vol. 52, n° 7, p. 1229-1258.
J. & A. (1991).- Stratigraphic Lexicon of Iran, Report No. 18.- Geological Survey of Iran publication, Tehran, Iran, 376 p.
M., F., A.R. & A.A. (2009).- Juraella bifurcata , 1984 (Calcareous alga, Gymnocodiaceae?) from the Lower Cretaceous (Barremian) Tirgan Formation of the Kopet Dagh basin, north-east Iran.- Journal of Alpine Geology, Leoben, vol. 51, p. 79-86.