![]()
Sommaire
[Introduction] [Objectifs]
[Matériels et méthodes]
[Cadre géomorphologique et géologique]
[Résultats] [Discussion]
[Conclusions] [ Références bibliographiques] et ... [Figures]
* Auteure correspondante - Corresponding author
Unité GEOGLOB AD10-02, Département des Sciences de la Terre, Faculté des Sciences, Université de Sfax, 3038 Sfax (Tunisie)
Unité GEOGLOB AD10-02, Département des Sciences de la Terre, Faculté des Sciences, Université de Sfax, 3038 Sfax (Tunisie)
16, rue de Mégret, F-33400 Talence (France)
Unité GEOGLOB AD10-02, Département des Sciences de la Terre, Faculté des Sciences, Université de Sfax, 3038 Sfax (Tunisie)
Manuscrit en ligne depuis le 05 Juillet 2011
![]()
L'étude quantitative et qualitative des ostracodes des carottes SG1 et BSC2, prélevées au niveau de la Sebkha El-Guettiate de Skhira (Sud - Est Tunisien), nous a permis de reconnaître les trois associations suivantes : (1) association de milieu marin s.l. composée de Aurila prasina , 1971, Cushmanidea elongata (, 1868), Urocythereis oblonga (, 1866), U. favosa (, 1838) (marin côtier), Basselerites berchoni (, 1869), Semicytherura incongruens (G.W. , 1894), S. paradoxa (G.W. , 1894), S. sella (, 1866), S. ruggierii (, 1955), Carinocythereis carinata (, 1838) et Paracytheridea depressa (G.W. , 1894) (marin ouvert) ; (2) association de milieu lagunaire, à salinité variable, constituée, de Xestoleberis aurantia (, 1838), Leptocythere fabaeformis (G.W. , 1894) et Cytherois fischeri (, 1866), (3) association de milieu saumâtre estuarien où se développent Cyprideis torosa (, 1850) et Loxoconcha elliptica , 1868. Ces associations révèlent les caractéristiques des paléo-environnements qui se sont succédés, au cours de l'Holocène supérieur, à l'emplacement de la Sebkha El-Guettiate. Trois phases peuvent être reconnues : i) la première phase à caractère lagunaire estuarien dominant (6595 ± 120 à 6055 ± 30 ans B.P.) comprend un environnement lagunaire ouvert riche en ostracodes marins dominants évoluant brutalement vers un environnement lagunaire estuarien. Ce dernier est caractérisé par un évènement de haute énergie, vers 6595 ± 120 ans B.P., révélé par des apports détritiques riches en galets, synchrones à l'enrichissement des ostracodes saumâtres. Ensuite, une lagune s'installe vers 6055 ± 30 ans B.P. permettant le développement des ostracodes principalement saumâtres dominants mais aussi lagunaires. ii) la phase suivante (6055 ± 120 à 5150 ± 50 ans B.P.) se distingue par la réapparition des taxons marins associés aux espèces lagunaires et saumâtres traduisant une lagune estuarienne largement ouverte soumise à l'influence des courants de dérive littorale. Ces courants, en édifiant des cordons littoraux, ont entraîné la fermeture du milieu et la fin de la dominance des ostracodes saumâtres. iii) la dernière phase (5150 ± 50 ans B.P. à l'actuel) comprend un milieu caractérisé par la richesse des taxons lagunaires évoluant vers un milieu laguno-saumâtre recevant le dépôt de washover suite à un événement climatique extrême. La lagune, de plus en plus fermée, évoluera progressivement vers la sebkha actuelle.
Sebkha El-Guettiate ; Sud-Est tunisien ; Holocène ; variations du niveau marin ; ostracodes.
C., F., P. & M. (2011).- Distribution des ostracodes dans les sédiments de subsurface de la Sebkha El-Guettiate (Skhira, golfe de Gabès). Intérêt pour la reconstitution des paléo-environnements de l'Holocène.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2011/03 (CG2011_A03), p. 63-81.
Ostracods distribution in the subsurface sediments of El-Guettiate Sebkha (Skhira, gulf of Gabes). Interest for Holocene palaeoenvironmental reconstruction.- A quantitative and qualitative study of the ostracods from the SG1 and BSC2 cores of El-Guettiate Sebkha of Skhira (southeastern Tunisia) has allowed the identification of different associations and the reconstruction of the Holocene palaeoenvironments. Ostracods, rich in species, provide three associations: (i) a marine association containing Aurila prasina , 1971, Cushmanidea elongata (, 1868), Urocythereis oblonga (, 1866), U. favosa (, 1838) (coastal marine), Basselerites berchoni (, 1869), Semicytherura incongruens (G.W. , 1894), S. paradoxa (G.W. , 1894), S. sella (, 1866), S. ruggierii (, 1955), Carinocythereis carinata (, 1838) and Paracytheridea depressa (G.W. , 1894) (open marine); (ii) lagoonal association, of variable salinity, comprising Xestoleberis aurantia (, 1838), Leptocythere fabaeformis (G.W. , 1894) and Cytherois fischeri (, 1866); (iii) brackish estuarine association subject to estuarine influence as indicated by Cyprideis torosa (, 1850) and Loxoconcha elliptica , 1868. The ostracods from El-Guettiate Sebkha allow us to recognize discrete palaeoenvironments that can be grouped in three phases: i) the first (6595 ± 120 - 6055 ± 30 years BP) begins with an open lagoon rich in marine ostracods followed by an estuarine lagoon (towards 6595 years BP) characterized by the richness of brackish ostracods and high energy. In the last lagoonal environment (installed towards 6055 years BP), brackish species are dominant and associated with lagoonal taxa; ii) the second phase (6055 ± 30 years BP - 5150 ± 30 years BP) is an estuarine lagoon, subject to a large marine influence in which brackish marine and lagoonal species are present in the same percentages. In this environment, littoral drift currents were responsible for the genesis of sand spits, the termination of environments and the dominance of brackish-water ostracods; iii) The last phase (5150 ± 50 years BP - recent) represents a restricted lagoon (about 5150 years BP) and finally a brackish lagoon evolved towards the sebkha environment under investigation here. This phase includes an unstable period with washover deposits resulting from an extreme climatic event.
El-Guettiate Sebkha; South - Eastern Tunisia; Holocene; sea level change; ostracods.
Plusieurs auteurs se sont intéressés à l'évolution des systèmes côtiers tunisiens au cours de l'Holocène. Ils ont conclus que la frange littorale du Sud Est tunisien, relativement stable et sans activité tectonique significative depuis les derniers 130 000 ans, a été contrôlée essentiellement par les variations eustatiques ( & , 1977 ; et alii, 1982 ; , 1986 ; et alii, 2003). Concernant les études micropaléontologiques, elles ont porté aussi bien sur les séries holocènes du littoral du golfe de Gabès que sur celui du nord du pays. Pour le littoral nord, plusieurs travaux ont mis en évidence l'impact de la communication des eaux des lagunes avec la Méditerranée sur la composition des associations des ostracodes et des foraminifères (, 1979 ; , 1980 ; & , 1981 ; et alii, 2006 ; et alii, 2006).
Pour les sédiments de surface et de subsurface du golfe de Gabès, les études paléontologiques et micropaléontologiques réalisées sont relativement anciennes et correspondent à l'analyse des mollusques (, 1979), des phanérogames (, 1979) et des bryozoaires ( & , 1979). La microflore a intéressé (1979) pour les dinoflagellés et (1979) pour les pollens. Les foraminifères ont été étudiés par et alii (1979), les nannofossiles calcaires par (1979) et les ostracodes par et alii (1979). Plus tard, plusieurs synthèses ont été proposées pour les forages du Golfe. (1989) a fait l'inventaire des ostracodes holocènes des carottes offshore du golfe de Gabès. Il a décrit différentes associations d'ostracodes, essentiellement pléistocènes, contenant des phytaux, des opportunistes ubiquistes, des opportunistes phytaux et des nageurs. Les associations holocènes lui ont permit de caractériser un environnement de basse énergie aux alentours de 5480 ans B.P. (1992) grâce à l'analyse palynologique des carottes marines dans le golfe de Gabès et sur le plateau des Kerkennah, a pu caractériser les modifications du paysage végétal en Tunisie depuis 20 000 ans. Il met en évidence que dans le Tardiglaciaire et la première moitié de l'Holocène, trois événements botaniques majeurs pouvant être considérés comme des signaux climatiques :
i) l'extension d'Artemisia corrélativement avec une régression de Quercus à feuillage caduc et la prééminence d'Olea sur Pistacia entre la fin de l'époque romaine et le Xème siècle ;
ii) la suprématie d'Olea au cours des derniers siècles de notre ère. (2002) en se basant sur les dinokystes a reconstitué la variation relative du niveau marin au niveau du Golfe de Gabès. Elle indique, qu'après 13 000 ans B.P., la transgression postglaciaire progresse rapidement avec une vitesse "conforme aux résultats présentés par (1989) pour la Barbade". Les datations radiocarbone montrent que, dans le golfe de Gabès, le niveau de la mer est à -75 m vers 12 580 ans B.P. La montée des eaux s'accentue à partir de 9930 ans B.P. Ce niveau est à -26 m vers 5480 ans B.P. et atteint un maximum vers 4630 ans B.P. avant de se stabiliser.
Plusieurs études sédimentologiques ont porté sur les séries connues à l'affleurement et celles recoupées par les forages des sebkhas paraliques et endoréiques du golfe de Gabès. La variation relative du niveau marin ( et alii, 2006), la dynamique littorale et le réajustement hydro-isostatique ( et alii, 2000 ; & , 2005), les évènements extrêmes ( et alii, 2010) et les changements climatiques ( et alii, 2008) seraient les facteurs responsables de l'évolution des environnements holocènes.
La frange côtière ici étudiée n'a fait l'objet d'aucune étude micropaléontologique et sédimentologique aussi bien pour les sédiments de surface que de subsurface. En effet, le peu de travaux micropaléontologiques réalisés ont porté exclusivement sur les forages du golfe de Gabès, d'ailleurs, implantés à plusieurs km de la ligne de côte. De même, si le littoral sud du golfe de Gabès (littoral de l'île de Jerba, des sebkhas El Melah, Boujmal et Mhabeul, et de la lagune El Biban), pour lequel nombre d'études sédimentologiques et géomorphologiques ont été menées, la frange littorale à l'emplacement de la Sebkha El-Guettiate reste très peu documentée.
L'objectif de ce travail est : (1) de caractériser les paléo-environnements qui se sont succédés à l'emplacement de la Sebkha El-Guettiate et (2) de distinguer l'impact des facteurs se rapportant à la dynamique littorale, aux variations climatiques, au réajustement hydro-isostatique, ŕ l'eustatisme et aux événements climatiques extrêmes, sur l'évolution de la ligne de rivage durant les derniers 6500 ans B.P. Dans ce travail, qui intervient dans un contexte de réchauffement climatique global accompagné d'un soulèvement général du niveau marin, nous nous baserons principalement sur l'analyse des faunes d'ostracodes présentes dans les sédiments de subsurface pour caractériser les paléo-environnements et éclairer les mécanismes naturels de forçage responsables des changements environnementaux.
La Sebkha El-Guettiate, de type paralique et caractérisée par des flux détritiques généralement réduits, est soumise à une alimentation alternée en eaux marines et continentales, ce qui a pour effet d'entraîner des variations du spectre faunistique et sédimentaire dans le temps et dans l'espace. Elle offre, ainsi, un terrain de choix pour la caractérisation des environnements passés à travers la microfaune de ses sédiments de subsurface.
Les travaux sur les ostracodes récents et fossiles ont démontré l'importance de ce peuplement pour la caractérisation des changements environnementaux holocènes. En effet, vivant à différentes profondeurs, sur tous types de substratums et dans les eaux à salinités variables, les ostracodes constituent d'excellents proxies pour la reconnaissance des paramètres physiques et chimiques des paléo-environnements. Ainsi l'analyse des biocénoses permet de caractériser les environnements où ils habitent et l'étude des thanatocœnoses autorise de retracer les schémas de circulation littorale (, 1980, 1982 ; et alii, 1981 ; et alii, 1988, 1990 ; , 1989 ; et alii, 2001 ; et alii, 2003, 2006 ; et alii, 2005 ; et alii, 2005).
Le matériel étudié correspond aux ostracodes présents dans les sédiments traversés par les deux carottes SG1 et BSC2 de longueurs respectives 122 et 150 cm. Les deux carottes ont été prélevées à proximité du chenal de marée principal de la sebkha étudiée. Au laboratoire, après la photographie des sédiments, les différentes unités lithologiques identifiées dans les carottes sont décrites et comparées à la série sédimentaire affleurant sur les rives du chenal de marée de la sebkha. L'échantillonnage a été effectué tout les centimètres. Les échantillons, préalablement séchés et pesés, sont lavés en utilisant deux tamis à mailles respectives 150 µm et 63 µm. Après séchage, les refus des tamis ont été observés à la loupe binoculaire pour la détermination des espèces. Le comptage du nombre d'espèces et de la densité a été réalisé sur le résidu total. Les valeurs obtenues sont rapportées à 10 grammes de sédiment sec. Pour chaque niveau les paramètres biocénotiques suivants sont calculés : 1) densité faunique ou abondance absolue (nombre d'individus) (∑) ; 2) richesse spécifique (NS). Ces paramètres expriment la stabilité du milieu et sa richesse en éléments nutritifs. Les associations microfaunistiques inventoriées nous ont permis de caractériser le type d'environnement et ce, par comparaison aux associations citées au niveau du littoral tunisien et méditerranéen. L'étude sédimentologique a concerné essentiellement le comptage et l'étude morphoscopique des grains de quartz présents dans la fraction supérieure à 150 µm. La variation de la richesse des différents niveaux de la carotte en grains quartz traduit l'exposition du milieu aux apports continentaux et/ou marins. La morphoscopie des grains nous informerait sur la proximité de leur source et leur mode de transport. La datation des deux carottes a été réalisée, au laboratoire des Sciences du Climat et de l'Environnement du CNRS de Gif-sur-Yvette (France), grâce à la malacofaune et la microfaune d'ostracodes et des foraminifères.
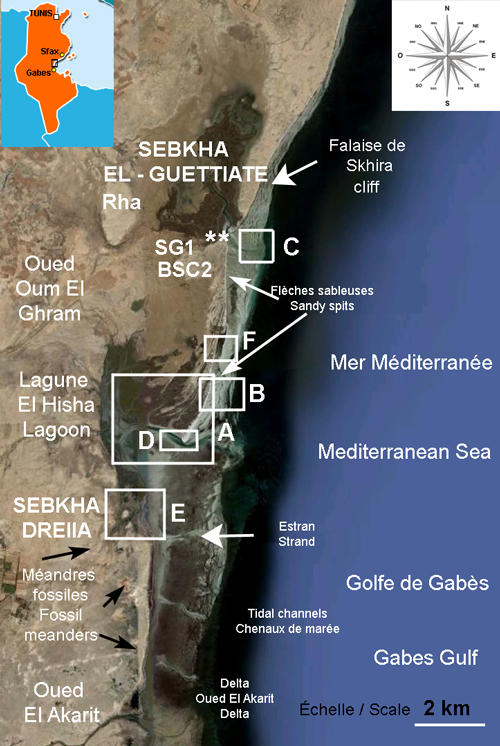
Le littoral étudié, délimité au Nord par la falaise de Skhira et vers le Sud par oued El Akarit, est une unité déployant environ 15 Km de linéaire côtier. Il constitue la partie la plus concave du golfe de Gabès. Ce linéaire côtier correspond dans sa partie nord à la falaise de Skhira, en cours d'écroulement, et dans sa partie Sud à une succession de cordons littoraux sableux de direction N-S ancrés sur la limite Sud de la falaise (Fig. 1 ![]() ). Ces cordons littoraux, sous l'effet de la dérive littorale, isolent la Sebkha El-Guettiate, la partie septentrionale de la Sebkha Dreîaa et la lagune El Hisha. La partie sud de la
Sebkha Dreîaa ouverte sur la mer est envahie au cours des marées exceptionnelles de
vives eaux. Le littoral de Skhira est soumis à un fort marnage pouvant atteindre 1,3 m. Ceci est mis en évidence par la présence d'accumulations de matériels détritiques, sableux et graveleux, exposées aux dynamiques côtières agressives. Ainsi du nord vers le sud les traits morphologiques suivants peuvent être reconnus :
). Ces cordons littoraux, sous l'effet de la dérive littorale, isolent la Sebkha El-Guettiate, la partie septentrionale de la Sebkha Dreîaa et la lagune El Hisha. La partie sud de la
Sebkha Dreîaa ouverte sur la mer est envahie au cours des marées exceptionnelles de
vives eaux. Le littoral de Skhira est soumis à un fort marnage pouvant atteindre 1,3 m. Ceci est mis en évidence par la présence d'accumulations de matériels détritiques, sableux et graveleux, exposées aux dynamiques côtières agressives. Ainsi du nord vers le sud les traits morphologiques suivants peuvent être reconnus :
1- La Sebkha El-Guettiate d'une superficie d'environ 15 km2 communique avec la Méditerranée grâce à un étroit chenal de marée, de 10 m de largeur et de 7 m de profondeur. Il constitue l'exutoire des deux oueds Oum El Ghram et Bou Saïd dont l'écoule se limite aux périodes de fortes crues. Les sédiments superficiels de la sebkha sont constitués par des limons rouges, de quelques dizaine de centimètres d'épaisseur, montrant parfois de grands cristaux de gypse, surmontant des argiles vertes plastiques. De nombreux chenaux, creusés sous l'influence de la marée et rarement des apports fluviatiles, entaillent les sédiments superficiels de la sebkha faisant apparaître les sédiments plus profonds. En effet, le chenal de marée principal montre plusieurs bifurcations pouvant se poursuivre très loin à l'intérieur de la sebkha. Un système de schorre et slikke caractérise la partie centrale de la sebkha.
2- La Sebkha El-Guettiate est relayée, sans limites claires, vers le Sud par la Sebkha Dreîaa. Cette dernière d'une superficie plus importante est séparée de la mer ouverte dans sa partie nord par
un système de flèches sableuses littorales dénotant une dérive
Sud des sables littoraux. Dans sa partie sud, cette sebkha se trouve largement ouverte à la mer.
Il s'individualise une petite lagune (lagune d'El Hisha) soumise aux influences de la marée surtout lors des vives eaux. Cette lagune est occupée dans sa partie nord par un système
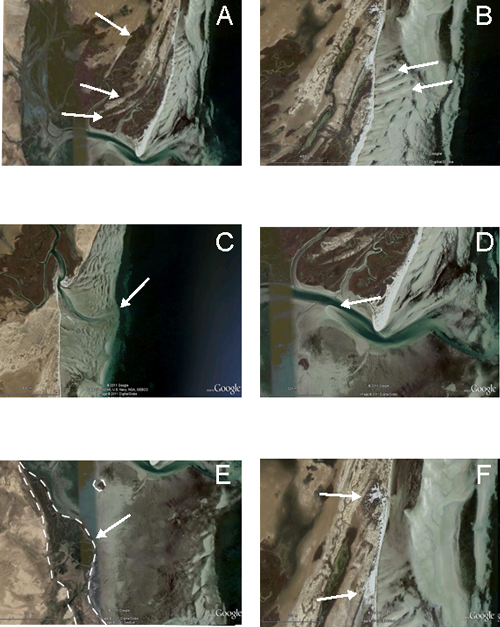
de schorres et de slikkes laissant apparaître en photographies aériennes 5 flèches littorales fossiles orientées NE-SO (Fig. 2A ![]() ). Ces flèches littorales dénotent d'une part la fermeture progressive de la lagune et
de la sebkha par rapport à la Méditerranée et l'importance de la dérive littorale dans cette partie du golfe de Gabès d'autre part. Les schorres et les slikkes sont parcourus par un système de chenaux sous l'influence de la marée et/ou des apports fluviatiles. Les photographies satellites montrent
que deux bras de l'oued El Ghram se déversent dans la lagune oů se
dessinent plusieurs méandres abandonnés.
). Ces flèches littorales dénotent d'une part la fermeture progressive de la lagune et
de la sebkha par rapport à la Méditerranée et l'importance de la dérive littorale dans cette partie du golfe de Gabès d'autre part. Les schorres et les slikkes sont parcourus par un système de chenaux sous l'influence de la marée et/ou des apports fluviatiles. Les photographies satellites montrent
que deux bras de l'oued El Ghram se déversent dans la lagune oů se
dessinent plusieurs méandres abandonnés.
Le littoral adjacent aux deux sebkhas est caractérisé par un estran très développé d'une largeur moyenne qui varie de 400 m au Nord à environ 1,4 km vers le Sud. L'estran montre des mégarides (Fig. 2B ![]() ) dont certaines sont orientées parallèlement à ligne côte et d'autres y sont obliques avec un angle de l'ordre de 15°,
environ. Associés aux accumulations des galets décimétriques, visibles sur la ligne de côte en face du chenal de marée de la
Sebkha El-Guettiate, ces figures sédimentaires témoignent de l'importance des courants marins. Dans sa partie
externe, l'estran montre un delta de jusant (Fig. 2C
) dont certaines sont orientées parallèlement à ligne côte et d'autres y sont obliques avec un angle de l'ordre de 15°,
environ. Associés aux accumulations des galets décimétriques, visibles sur la ligne de côte en face du chenal de marée de la
Sebkha El-Guettiate, ces figures sédimentaires témoignent de l'importance des courants marins. Dans sa partie
externe, l'estran montre un delta de jusant (Fig. 2C ![]() ), se prolongeant dans le subtidal, à l'embouchure des chenaux de marée principaux. Des accumulations de sables, pouvant correspondre à des deltas de flot, s'observent également à l'embouchure du chenal de marée de la
Sebkha Dreîaa du côté de la lagune (Fig. 2D
), se prolongeant dans le subtidal, à l'embouchure des chenaux de marée principaux. Des accumulations de sables, pouvant correspondre à des deltas de flot, s'observent également à l'embouchure du chenal de marée de la
Sebkha Dreîaa du côté de la lagune (Fig. 2D ![]() ). Le littoral de la
Sebkha Dreîaa montre la présence d'un haut fond, complètement émergé en basses marées
de vives eaux, colonisé par les végétaux Ammophilia arenaria. Au niveau de la ligne de côte et en face de l'un des méandres abandonné, ces mêmes végétaux occupent une grande partie du littoral (Fig. 2E
). Le littoral de la
Sebkha Dreîaa montre la présence d'un haut fond, complètement émergé en basses marées
de vives eaux, colonisé par les végétaux Ammophilia arenaria. Au niveau de la ligne de côte et en face de l'un des méandres abandonné, ces mêmes végétaux occupent une grande partie du littoral (Fig. 2E ![]() ).
).
L'impact des tempêtes sur la morphologie du littoral est visible sur les photos aériennes. En effet plusieurs dépôts de washover matérialisés par des corps sableux dont l'épaisseur diminue en s'éloignant du cordon littoral sont observés dont certains sont situés au niveau des embouchures tidales (Fig. 2F ![]() ). Les deux sebkhas sont donc soumises aussi bien aux influences marines, suite aux
tempętes induisant une surélévation du plan d'eau et un
renforcement de l'érosion, qu'aux influences continentales lors des crues exceptionnelles.
C'est ainsi que les sebkhas sont des lieux d'accumulation de matériel terrigène provenant des bassins versants mais aussi de sables venus de la mer en période de tempêtes.
). Les deux sebkhas sont donc soumises aussi bien aux influences marines, suite aux
tempętes induisant une surélévation du plan d'eau et un
renforcement de l'érosion, qu'aux influences continentales lors des crues exceptionnelles.
C'est ainsi que les sebkhas sont des lieux d'accumulation de matériel terrigène provenant des bassins versants mais aussi de sables venus de la mer en période de tempêtes.
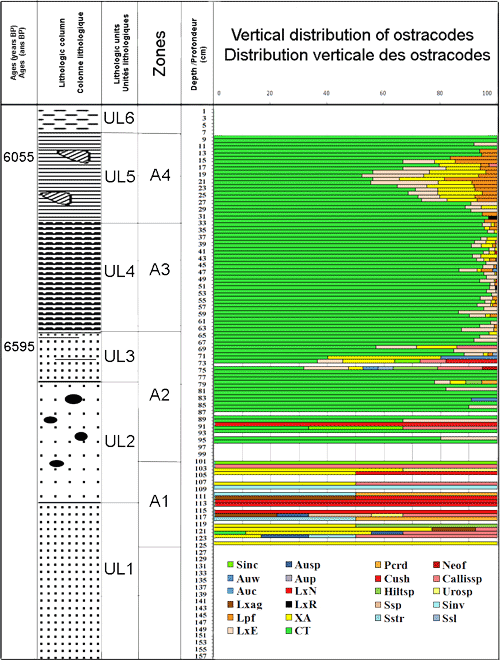
La carotte BSC2 d'une épaisseur totale de 157 cm a recoupé, de la base vers le sommet, les six unités lithologiques suivantes (Fig. 3 ![]() ) : (UL1) des sables fins entre 157 et 113 cm ; (UL2) des sables grossiers de plus en plus riches à galets entre 113 et 78 cm ; (UL3) des sables silteux gris riches en niveaux à végétaux et en mollusques entre 78 et 64cm ; (UL4) des sables silteux gris entre 64 et 33 cm ; (UL5) des argiles jaunes verdâtres renfermant des mollusques lagunaires entre 33 et 7 cm et enfin (UL6) des argiles vertes azoïques riches en cristaux de gypse.
) : (UL1) des sables fins entre 157 et 113 cm ; (UL2) des sables grossiers de plus en plus riches à galets entre 113 et 78 cm ; (UL3) des sables silteux gris riches en niveaux à végétaux et en mollusques entre 78 et 64cm ; (UL4) des sables silteux gris entre 64 et 33 cm ; (UL5) des argiles jaunes verdâtres renfermant des mollusques lagunaires entre 33 et 7 cm et enfin (UL6) des argiles vertes azoïques riches en cristaux de gypse.
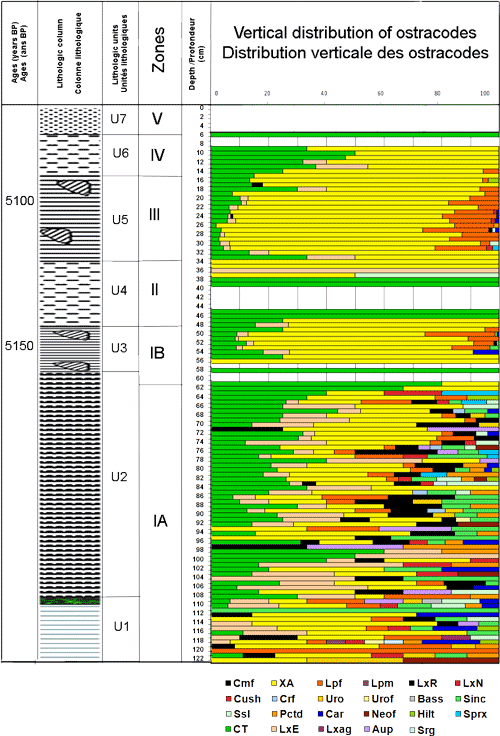
La carotte SG1, plus riche en macrofossiles et en meiofaune, a révélé, de la base vers le sommet, les unités lithologiques suivantes (Fig. 4 ![]() ) : (U1) des sables fins argileux surmontés par un niveau centimétrique de végétaux, entre 122 et 107 cm ; (U2) des sables silteux beiges, entre 107 cm et 58 cm ; (U3) des argiles jaunes verdâtres riches en mollusques, entre 58 cm et 48 cm ; (U4) des argiles vertes, entre 48 cm et 34 cm ; (U5) des argiles jaunes verdâtres riches en débris de mollusques entre 34 cm et 15 cm ; (U6) des argiles vertes plastiques, entre 15 et 6 cm et enfin (U7) des limons rouges à cristaux de gypses de 6 cm à la surface.
) : (U1) des sables fins argileux surmontés par un niveau centimétrique de végétaux, entre 122 et 107 cm ; (U2) des sables silteux beiges, entre 107 cm et 58 cm ; (U3) des argiles jaunes verdâtres riches en mollusques, entre 58 cm et 48 cm ; (U4) des argiles vertes, entre 48 cm et 34 cm ; (U5) des argiles jaunes verdâtres riches en débris de mollusques entre 34 cm et 15 cm ; (U6) des argiles vertes plastiques, entre 15 et 6 cm et enfin (U7) des limons rouges à cristaux de gypses de 6 cm à la surface.
Du point de vue chronologique, des datations absolues par 14C réalisées au centre de Gif / Yvette ont fournit les âges conventionnels suivants :
Les carottes SG1 et BSC2 révèlent la présence de 23 espèces d'ostracodes représentées par un nombre d'individus atteignant 300 individus par espèce par 10g de sédiment sec. Toutes ces espèces prolifèrent aujourd'hui à la fois au niveau des côtes Nord et Sud méditerranéennes et des côtes Est atlantiques. Les espèces reconnues caractérisent les trois associations :
Les espèces d'ostracodes qui dominent le long de SG1 sont les lagunaires (Xestoleberis aurantia (17%) ; Leptocythere fabaeformis (9 %) suivies par les saumâtres Cyprideis torosa (16%) et Loxoconcha elliptica (11%) et enfin les marines (6%). Le nombre d'individus maximal inventorié est de 329 indiv. / 10g de sédiment sec observé dans l'unité U5.
Les sédiments recoupés par la carotte BSC2 se distinguent par la dominance de C. torosa (49%) suivie par la forme lagunaire X. aurantia (5%) et enfin l'espèce marine Cushmanidea elongata (3%). Les autres formes ne dépassant pas le 1%. Le nombre d'individus maximal est analogue à celui de la carotte SG1 atteignant 328 indiv. / 10g de sédiment sec.
En se basant sur les variations de la distribution relative des taxons dominants, de la richesse spécifique, des abondances absolues, et des abondances relatives de chaque association des ostracodes, les zones suivantes peuvent être distinguées au niveau des différentes unités lithologiques des carottes BSC2 (Fig. 5 ![]() ) et SG1 (Fig. 6
) et SG1 (Fig. 6 ![]() ) :
) :
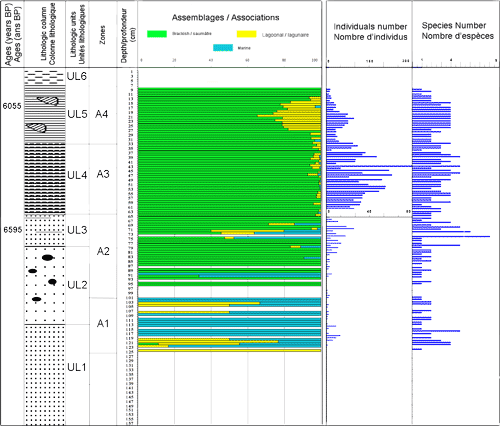
Zone A1, de 125 à 100 cm, correspond au sommet de l'unité UL1 et la partie inférieure de UL2. Elle se caractérise par les faibles valeurs de la densité (ne dépassant pas 20 indiv. / 10g), par la richesse spécifique des ostracodes (1 et 5 espèces) et par la dominance des ostracodes marins, par rapport aux taxons lagunaires. Les espèces marines phytales, dénotant l'ouverture du milieu, sont : Aurila woodwardii (, 1868), Aurila sp., Loxoconcha agillis , 1967, L. napoliana , 1963, Urocythereis sp., Semicytherura incongruens, S. inversa (, 1880), S. striata (, 1866), Paracytheridea depressa, Hiltermannicythere sp. et Callistocythere sp. Les formes lagunaires se limitent à l'espèce Xestoleberis aurantia. Tous ces paramètres traduisent un milieu lagunaire de forte énergie et soumis à d'importantes influences marines.
Zone A2, de 100 à 63cm, couvre la partie supérieure des sables moyens à galets (UL2) et la totalité des sables silteux gris riches en niveaux à végétaux et en mollusques (UL3). Elle se distingue par l'apparition brutale des deux espèces saumâtres Loxoconcha elliptica et C. torosa, largement dominantes, et par l'appauvrissement des taxons marins. L'enrichissement des taxons euryhalins est associé à la richesse des sédiments en galets. Le sommet de cette zone, constitué par les silts à végétaux et mollusques (UL3) d'âge 6595 ans B.P., se caractérise par une augmentation de la densité des ostracodes atteignant 30 individus et la richesse spécifique (8 espèces). Il montre également la présence des ostracodes marins dénotant une influence marine. La dominance d'ostracodes saumâtres, le long de la zone A2, indique des eaux euryhalines et un environnement lagunaire à influence estuarienne.
Zone A3, de 63 à 33 cm, se développant dans les sables silteux gris riches en mollusques (UL4), elle est marquée par une importante augmentation de la densité des ostracodes (326 individus / 10g de sédiment sec) témoignant la présence d'éléments nutritifs (, 1980). Les valeurs de la richesse spécifique demeurent comparables à celles de la zone sous-jacente. Cyprideis torosa domine le peuplement (90%) suivie de L. elliptica, Xestoleberis aurantia et Leptocythere fabaeformis. Le long de la zone A3, la rareté des ostracodes marins et la dominance des espèces saumâtres dénotent la pérennité du caractère euryhalin des eaux d'une lagune estuarienne.
Zone A4, de 33 cm à 7 cm, correspond aux argiles à mollusques lagunaires. On y trouve Cerastoderma glaucum et Cerithium vulgatum d'âge 6055 ans B.P. (UL5). Les ostracodes montrent une réduction de leur densité, et leur richesse spécifique reste comparable à celle de la zone sous - jacente. L'enrichissement de L. elliptica, X. aurantia et L. fabaeformis associé à l'appauvrissement relative de C. torosa traduit l'installation d'un environnement lagunaire.
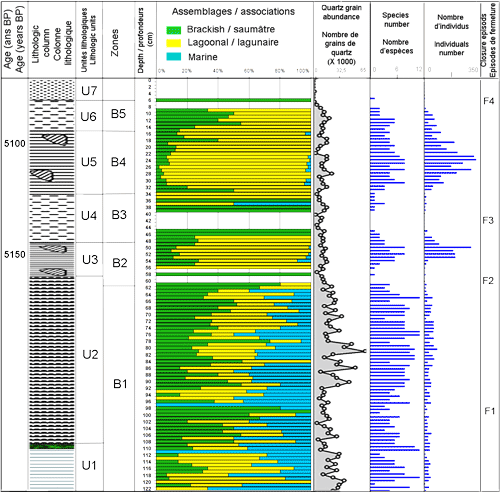
Zone B1, de 122 à 61 cm : très riche en débris d'algues vertes Halimeda, se caractérise par une importante richesse spécifique (10 espèces) et une faible densité des ostracodes (entre 4 et 44 indiv. / 10g de sédiment sec). Les espèces marines, marquant l'ouverture du milieu (, 1980) : Aurila prasina, Cushmanidea elongata, Urocythereis oblonga, U. favosa, Semicytherura incongruens, S. sella, Carinocythereis carinata, Paracytheridea depressa, Loxoconcha rhomboîdea (, 1855), Hiltermannicythere emaciata (, 1867) et Callistocythere sp., sont les plus abondantes dans les parties inférieure et moyenne de la zone. Leur richesse est corrélée positivement avec les fortes valeurs de l'abondance des grains de quartz. Les espèces lagunaires Xestoleberis aurantia et Leptocythere fabaeformis dominantes à la base de la zone, gardent le long de la zone la même richesse avant de s'appauvrir, en même temps que les espèces marines, jusqu'à disparaître au sommet. La coexistence le long de la zone B1 des trois associations d'ostracodes, avec des abondances relatives variables, traduit généralement un milieu lagunaire estuarien ouvert. À la base de la zone, de 111 à 98 cm, le détritisme décroît et les taxons marins sont progressivement remplacés par les espèces euryhalines, caractéristiques des environnements paraliques, de plus en plus dominantes. L'enrichissement des espèces saumâtres et la baisse de la richesse spécifique marqueraient un épisode de fermeture du milieu (F1). À partir de 75 cm de profondeur, C. torosa s'enrichit de nouveau et s'associe à l'appauvrissement des taxons marins et lagunaires. À la fin de la zone, C. torosa domine le peuplement sans ostracodes marins et lagunaires. Cette évolution signerait un deuxième épisode de fermeture (F2).
Zone B2, de 61 cm à 48 cm : se distingue par pauvreté en grains de quartz et par sa richesse en lamellibranches et gastéropodes, en position de vie. Ces derniers, représentés respectivement par les espèces lagunaires Cerastoderma glaucum et Cerithium vulgatum, ont fourni un âge 5150 ans B.P. L'intervalle 61 - 57 cm, se caractérise, pour la plupart de ses niveaux, par l'absence des ostracodes.
À partir de 56 cm, les ostracodes réapparaissent de nouveau et montrent une densité beaucoup plus importante que dans la zone A (atteignant 297 indiv. / 10g). Dans cet intervalle X. aurantia domine (75%), viennent ensuite C. torosa (14%), L. fabaeformis (7%), L. elliptica (3%) et les ostracodes marins (1%). Xestoleberis aurantia dominera le peuplement des ostracodes jusqu'au sommet de la carotte.
La structure des populations des ostracodes révèle que les espèces lagunaires et saumâtres sont représentées par aussi bien les individus adultes que juvéniles. Par contre, les formes marines ne fournissent que des individus juvéniles pour certaines espèces et que des individus adultes pour d'autres témoignant de leur caractère allochtone. En effet (2001) remarque que les associations fossiles dites thanatocœnoses s.l. peuvent comprendre autant d'individus autochtones (paléobiocénose) que d'individus amenés par des courants et qui ne vivaient pas dans le milieu (thanatocœnose de transport ou allothanatocœnose). Selon ce même auteur, la structure populationnelle des espèces et le pourcentage des valves juvéniles par rapport aux valves adultes (rapport J/A) sont des critères pour la reconnaissance de faune fossilisée sur place et de faune déplacée. En effet, les espèces in situ doivent être représentées à la fois par des valves juvéniles et des valves adultes. Selon (1988), les populations ayant des rapports J/A compris entre 3/1 et 5/1 ont une grande probabilité d'être in situ. Ainsi, pour les ostracodes de la zone B2, les espèces lagunaires et saumâtres sont ici considérées comme en place (paléobiocénose) et les espèces marines seraient déplacées (allothanatocénose) suite à l'hydrodynamisme du milieu. Tous ces éléments plaident en faveur d'un environnement lagunaire pour la zone B2. La présence des taxons marins, rares et synchrones à l'enrichissement du milieu en grains de quartz, indique des communications occasionnelles avec le milieu marin par l'intermédiaire d'un chenal de marée ou suite à des marées exceptionnelles.
Zone B3, de 48 cm à 34 cm : est marquée par l'absence des ostracodes dans la plupart de ses niveaux. Dans les premiers niveaux de la zone sont présentes les espèces lagunaire Xestoleberis aurantia (75%) et Loxoconcha elliptica associées l'espèce saumâtre Cyprideis torosa, vers le haut il ne subsiste que C. torosa. Après les niveaux exempts d'ostracodes (44 à 40 cm), réapparaissent successivement C. torosa puis X. aurantia et enfin L. elliptica. La fin de la zone se caractérise par la présence exclusive de l'espèce lagunaire X. aurantia. L'absence des ostracodes dans l'intervalle 44 - 40 cm peut être expliquée par la dissolution complète de leurs valves à l'interface eau - sédiment où aux conditions du milieu qui ne permettaient pas la survie de l'ostracofaune. Si la première hypothèse est la bonne, la dissolution aurait affectée aussi bien les valves des ostracodes que les tests des foraminifères. La préservation totale de ces derniers et l'absence de traces de dissolution sur leurs tests témoignent que l'absence des ostracodes serait due à la fermeture du milieu lagunaire (épisode F3) induisant son émersion. Au cours de cette phase de fermeture, caractérisée par l'absence des ostracodes et par la réduction du nombre des grains de quartz, les foraminifères ont cependant pu survivre, tout en enregistrant une réduction de leur richesse spécifiques et de leur densité. Les foraminifères sont en effet capables de résister pendant les périodes d'assèchement ( & , 2005). La fermeture complète de la lagune peut être rapportée à l'ancrage d'un cordon dunaire sur le littoral coupant les connexions avec le domaine marin. Par la suite, le milieu devient moins agité et s'y développent C. torosa et X. aurantia. La réapparition de X. aurantia (atteignant 100%) indiquerait l'ouverture de la lagune, probablement via un chenal de marée alimentant la lagune derrière un cordon littoral.
Zone B4, de 34 cm à 16 cm, correspond à l'unité unités U5. Elle renferme les mêmes espèces de lamellibranches et de gastéropodes inventoriées dans la zone B2. Ces mollusques sont d'âge 5100 ans B.P. comparable à celui de l'unité U3. Les valves des lamellibranches et les coquilles des gastéropodes sont brisées et organisées en amas signant une sédimentation de forte énergie. La densité et la richesse spécifique des ostracodes sont plus élevées par rapport à la zone B. À ces différents paramètres s'ajoutent la présence des foraminifères planctoniques indice de pélagisme. Tous ces éléments plaident en faveur d'un dépôt de haute énergie de l'unité U5 probablement suite à une période de tempête ou de tsunami. L'ostracofaune de l'unité U5 serait donc remaniée. Il s'agit probablement d'une thanatocœnose de transport sous haute énergie à partir de séries plus anciennes, pouvant ętre l'unité U3. En effet, les âges des mollusques des unités U3 et U5 étant comparables.
Zone B5, de 16 cm à la surface, se caractérise par l'enrichissement progressif de C. torosa au détriment de X. aurantia et L. fabaeformis. La réduction de la densité et de la richesse spécifique et l'enrichissement de C. torosa marquent un nouvel épisode de fermeture de la lagune.
Les données micropaléontologiques associées à l'analyse de la structure et de la diversité des populations des ostracodes permettent de reconstituer l'évolution des paléo-environnements à l'emplacement de la Sebkha El-Guettiate. La frange littorale s'est de plus en plus isolée par rapport à la mer ouverte. La progression de la proportion des ostracodes saumâtres, lagunaires et marins associée à la variation de leur densité et richesse spécifique peut avoir plusieurs explications. Il pourrait s'agir : (1) de causes sédimentologiques, liées à une dynamique littorale, telle que la fermeture des lagunes ouvertes suite à la formation d'un cordon littoral par le biais des courants de dérive littorale ; (2) de causes climatiques telle que l'aridification provoquant la sursalure du milieu où au contraire un apport d'eau continentale diluant les eaux de la lagune favorisant le développement des ostracodes saumâtres ; (3) de causes eustatiques ayant engendré l'isolement de la lagune ; (4) de causes hydro-isostatiques provoquant l'émersion de la frange côtière.
L'analyse morphoscopique des grains de quartz présents dans les sédiments des carottes SG1 et BSC2, révèle qu'ils sont en majorité luisants indiquant un apport aquatique. L'aspect anguleux à sub-anguleux de ces grains traduit la proximité de leur source. Les grains peuvent avoir une origine continentale (érosion des terrains mio-pliocènes constituant la dune de Rha) et transportés par les chenaux fluviatiles. L'origine côtière des apports détritiques est aussi possible suite à la destruction de la falaise de Skhira et/ou des anciennes dunes littorales, et les courants de marée et/ou de dérive littorale auraient pu également assurer le transport. De la base vers le sommet de la carotte SG1, la corrélation positive entre la richesse des ostracodes marins et l'abondance des grains de quartz suggère que les apports en grains de quartz diminuent avec la réduction des échanges avec le milieu marin. De même l'analyse granulométrique des sables des unités U1 et U2, recoupés par la carotte SG1, révèle qu'il s'agit de sables fins moyennement classés à courbe de fréquence bimodale témoignant d'un mélange d'éléments ayant deux origines différentes.
Pour la carotte BSC2, la dominance des ostracodes estuariens dans les sables à galets de UL2 traduit leur origine fluviatile probable, conséquente à des périodes de forte pluviométrie, responsables de l'euryhalinité du milieu, favorisant le développement de la faune saumâtre.
Concernant la morphologie du littoral, le développement pendant l'Holocène supérieur d'îles-barrières, de flèches et de bancs sableux responsables de l'isolement des environnements lagunaires du littoral tunisien a été mentionné par de nombreux auteurs. Citons (1979) pour la genèse du lac Ghar El Melah et de la Sebkha Ariana à partir de 6000 ans et et alii (2006) pour la genèse des faciès bioclastiques et des sédiments riches en matière organique de la Sebkha Boujmel de 6800 à 4000 ans B.P. D'ailleurs la présence de plusieurs flèches littorales fossiles, dont la plus récente est d'âge ca 877 ± 30 ans B.P., au niveau de la Sebkha Dreîaa, adjacente à Sebkha El-Guettiate, témoigne de l'existence des courants de dérive littorale à l'Holocène supérieur. Toutes ces données permettent d'envisager la présence, au cours de l'Holocène supérieur, d'un milieu marin riche en sables quartzeux. Ces grains seraient remaniés par le biais des courants de dérive littorale permettant la genèse des flèches littorales aujourd'hui visibles à l'intérieur de la Sebkha Dreîaa. Les différentes phases de fermeture de la lagune seraient en liaison avec l'action des courants de dérive littorale responsables de la genèse de flèches sableuses et l'isolement du milieu par rapport au milieu marin.
(2002) signale une augmentation du volume des océans due à la fonte de la glace antarctique aboutissant à une élévation du niveau des mers d'une moyenne de 3 mètres entre 6000 et 3000 ans. Les études réalisées au niveau de la frange côtière du SE tunisien témoignent paradoxalement d'un abaissement relatif du niveau marin et l'évolution de la frange littorale tunisienne vers la fermeture au cours de la période de temps ici considérée ( et alii, 2006 ; & , 1977). Au niveau de la Sebkha El-Guettiate, l'évolution des environnements d'un milieu lagunaire ouvert (Unité UL3 d'âge 6595 ± 120 ans B.P.) vers un milieu lagunaire fermé (unité U3 d'âge 5150 ± 50 ans B.P.) traduirait, comme c'est le cas pour les sebkhas Boujmel et El Melah de Zarzis, une fermeture du milieu.
Selon plusieurs auteurs, la frange côtière du golfe de Gabès était soumise au cours de l'Holocène supérieur à un soulèvement hydro-isostatique. En effet et alii (1998) citent des dépôts bioclastiques de plage, sur le littoral de Djerba, couvrant l'intervalle 5900 - 4000 ans B.P., situés à des altitudes variables entre + 40 à +100 cm ainsi que des dépôts d'origine marine plus récents, d'âge 2100 ans B.P affleurant au même niveau que la mer. La régularité des niveaux bioclastiques, de même âge, par rapport au niveau marin confirme pour ces auteurs l'absence de la tectonique. L'élévation, réalisée entre 5900 ans B.P. et 2000 ans B.P., serait donc d'origine hydro-isostatique. Pour & (2005) l'émersion de la frange côtière aurait atteint une altitude plus importante soit de 186 ± 11 cm et serait produite entre 6000 ans et 5000 ans B.P. Pour la Sebkha El-Guettiate, l'unité d'origine lagunaire (U3) est située au même niveau que le niveau marin. Si le réajustement hydro-isostatique avait touché la frange côtière à l'emplacement de la Sebkha El-Guettiate, l'altitude de (U3) ne pourrait être expliquée que par une subsidence ultérieure de la sebkha.
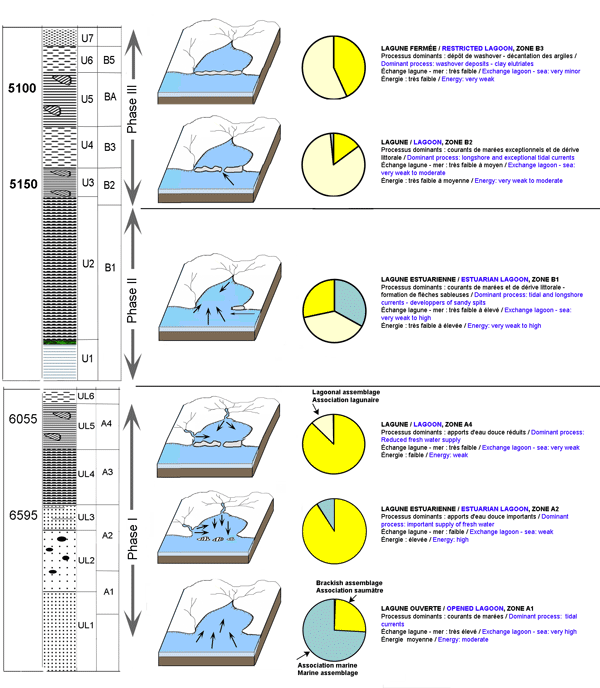
L'évolution dans le temps des abondances relatives des associations des ostracodes nous permet d'esquisser l'évolution de la frange côtière de Skhira à l'emplacement des carottes BSC2 et SG1 au cours de l'Holocène supérieur (Fig. 7 ![]() ). Trois phases peuvent être reconnues dans la séquence environnementale des deux carottes prélevées.
). Trois phases peuvent être reconnues dans la séquence environnementale des deux carottes prélevées.
Elle débute par l'installation d'une lagune ouverte à dynamique marine où se déposent, en premier lieu, les derniers sables fins d'UL1, riches en mollusques lagunaires, puis la partie inférieure des sables moyens de UL2. Ils s'y développent des ostracodes marins phytaux de la zone A1 traduisant l'ouverture de la lagune vers le milieu marin franc avec essentiellement : Aurila woodwardi, Aurila sp., Loxoconcha agillis, L. napoliana, Urocythereis sp. et Semicytherura incongruens. Les formes lagunaires se limitent à l'espèce Xestoleberis aurantia. Après cet épisode de lagune ouverte, les indicateurs environnementaux et sédimentaires changent brusquement et les influences du domaine marin deviennent plus rares. Le milieu connaît, au début, un épisode de haute énergie révélé par le dépôt des sables moyens à galets de plusieurs centimètres de diamètre (sommet de UL2). L'ostracofaune (zone A2) voit le développement de Cyprideis torosa, espèce vivant dans des eaux à caractéristiques chimiques variables (fortes variations de salinité avec phases de sursalure). Cette espèce dénote donc le caractère perturbé du milieu. En effet, l'apparition brutale des ostracodes saumâtres, dominants et à forte densité, est synchrone à l'enrichissement des sables de l'unité UL2 en galets. Cela montre que la lagune est soumise à une influence fluviatile. Les apports d'eau douce seraient responsables de l'euryhalinité des eaux de la lagune.
Ensuite la lagune devient moins agitée, et s'y déposent les silts à végétaux riches en mollusques lagunaires fournissant un âge 6595 ans B.P. Les galets disparaissent et les sables sont de plus en plus fins et s'enrichissent en végétaux. Les ostracodes marins réapparaissent. Toutefois, la pérennité et la dominance des ostracodes saumâtres (sommet de la zone A2) témoignent de la persistance des influences fluviatiles synchrones à une période humide. En effet pour la période ici considérée, plusieurs auteurs ont fait état d'un climat relativement humide au niveau du golfe de Gabès et en Tunisie Centrale. Une phase d'augmentation de l'humidité et une amélioration des conditions environnementales s'est développée entre 7,5 et 7,2 ka B.P. en Tunisie Centrale dans la région de Kasserine (, 2006), aux piémonts du chaînon Goubrar/Khechem entre 8 ka et 7,5 ka (, 2010) ainsi que dans la région de Maknassy entre 7 et 10 ka B.P. ( et alii, 1998). À une échelle plus grande, les travaux réalisés sur les environnements lacustres et les terrasses fluviatiles africains, ont révélé que les conditions étaient plus humides que l'actuel entre 10000 - 9500 et 7000 - 6000 ans B.P pour la partie Est du Sahara ( & , 1979 ; & , 1980 ; et alii, 1985 ; & , 1987).
Le dépôt des vases sableuses riches en coquilles lagunaires (UL4) traduit un milieu relativement calme. Les valeurs de la densité et de la richesse spécifiques des ostracodes (zone A3) sont les plus importantes de toute la carotte signant la disponibilité des éléments nutritifs dans la lagune (, 1980) et la stabilité relative des conditions du milieu. La rareté des ostracodes marins et lagunaires associée à la dominance des espèces saumâtres dénotent la persistance du caractère euryhalin des eaux d'une lagune estuarienne.
La phase I s'achève par l'installation d'un environnement lagunaire, vers 6055 ans B.P., permettant le dépôt des argiles de l'unité UL5 traduisant un milieu encore plus calme. La présence des mollusques lagunaires Cerastoderma glaucum et Cerithium vulgatum associée à l'enrichissement de L. elliptica X. aurantia et L. fabaeformis au dépend de C. torosa corroborent l'installation d'un environnement lagunaire.
Elle se distingue par l'influence marine dont témoigne la présence, au niveau des unités U1 et U2, des trois associations saumâtre, lagunaire et marine à parts égales (zone B1). L'ouverture de l'environnement est aussi argumentée par l'importante richesse spécifique et la faible densité des ostracodes. La richesse des espèces marines est associée à l'abondance des grains de quartz. La phase II peut être subdivisée en deux épisodes :
i) le premier permet le dépôt, en premier lieu, des sables fins argileux gris (U1) riches en Cerithium et en débris de lamellibranches puis des premiers sables silteux beiges de l'unité (U2). Ces sables contiennent des taxons marins (30%), lagunaires (60%) et saumâtres (10%). Ces derniers s'enrichissent progressivement, jusqu'à dominer le peuplement, et les taxons lagunaires et marins s'appauvrissent signant la fermeture de la lagune et son isolement par rapport au milieu marin ouvert. La réduction du nombre des grains de quartz, de l'abondance absolue et de la richesse spécifique des ostracodes corrobore la fermeture du milieu (F1). Cette fermeture est probablement conséquente à la formation d'un cordon littoral ;
ii) au cours du deuxième épisode, permettant le dépôt des derniers sables d'U2, la lagune s'ouvre brutalement, suite à la rupture du cordon littoral lors de périodes de tempêtes. L'ouverture est argumentée par l'augmentation brutale de la richesse des ostracodes marins (partie supérieure de la zone B1) et par l'arrivée des foraminifères planctoniques. L'augmentation du nombre de grains de quartz signe la présence de courants de dérive littorale enrichissant le milieu microfaune marine. Tous ces paramètres indiquent un milieu lagunaire à salinité variable, probablement estuarien, largement ouvert.
Cette phase débuterait par l'appauvrissement du milieu en grains de quartz et la raréfaction des ostracodes traduisant une nouvelle période de fermeture de la lagune (F2) suite à la formation d'un nouveau cordon littoral. Elle permet la décantation des argiles jaunes verdâtres à mollusques (U3) caractérisées par la dominance de l'espèce lagunaire X. aurantia. En effet, après la période de fermeture précédente (F2) le milieu s'ouvre et permet le développement des taxons lagunaires, dominants, marins et saumâtres (zone B2). Les espèces Cerastoderma glaucum et Cerithium vulgatum, présents dans U3, ont fournit un âge 5150 ans B.P. Ces mollusques lagunaires sont caractéristiques des environnements paraliques abrités aussi bien actuels que pléistocènes du Golfe de Gabès (, 1974 ; , 1998 ; et alii, 2004 ; , travaux en cours). La persistance des ostracodes marins témoigne de la communication temporaire de la lagune avec le milieu marin par l'intermédiaire d'un chenal de marée ou suite à des marées exceptionnelles. Les courants de marée auraient transportés et dispersés les espèces d'ostracodes marins entrées accidentellement dans la lagune.
Le maximum de fermeture de la lagune et son comblement, par les argiles vertes de l'unité (U4), entraînera son émersion et la disparition totale des ostracodes. L'absence des ostracodes peut être rapportée à leur sensibilité aux périodes d'émersion. En effet & (2005) indiquent que les périodes d'émersion et d'assèchement constituent un danger pour les ostracodes vivant en zone intertidale au dessous du niveau moyen de la marée. Ces petits crustacés ne peuvent survivre pendant les phases émersions que durant 1 à 2 cycles de marées en fermant leurs valves et en diminuant leur respiration. Cette phase d'émersion de la lagune (F3) peut correspondre à la période d'aridité identifiée dans golfe de Gabès par (1992) sur la base de la régression des hygrophytes marquant le début d'une aridification vers 5000 B.P. Récemment, & (2008) en étudiant la dynamique fluviale en Tunisie ont caractérisé une période généralement aride entrecoupée par des périodes d'inondation à partir de 4700 ans B.P. Cette phase d'aridité serait aussi synchrone à la précipitation des saumures, à partir de 5000 ans B.P., de la Sebkha El Melah de Zarzis ( & , 1977).
Durant la période de fermeture (F3) la lagune permet le dépôt de l'unité (U5). Cette unité, riche en accumulations de débris de mollusques, d'âge 5100 ans B.P., serait déposée suite probablement à une période de haute énergie. En effet les sédiments et les taxons d'ostracodes présents dans U5 sont similaires à ceux de U3 et le pourcentage des bioclastes s'avère légèrement plus élevé. L'ostracofaune de U5 correspondrait à une thanatocœnose de transport (zone B4), avec la conservation de la structure de population originale, à partir probablement de l'unité U3. L'unité U5 caractérisée par sa base érosive, visible au niveau du chenal de marée, et dont l'épaisseur diminue vers le centre de la sebkha, montre des stratifications en mamelons témoin d'une sédimentation de forte énergie. Toutes ces données dénotent d'un dépôt probablement de type washover ( et alii, 2003 ; et alii, 2008) pour l'unité (U5). Ce dépôt serait formé suite à une tempęte en arrière du cordon littoral sableux précédemment formé (période F3). Il est donc formé par l'activité de la houle et du vent qui érodent le cordon dunaire et déposent l'unité U5 dans une lagune. L'épaisseur de l'unité (U5) diminue en s'éloignant du cordon littoral vers le centre de la sebkha. Il s'agit de la première caractérisation de dépôt de type washover dans la série holocène du golfe de Gabès. En effet les faciès de haute énergie, cités par et alii (2010) et observés au sommet de l'éolianite de Sidi Salem (d'âge controversé 6530, 6980, 7750 ou 8045 ans B.P.), sont rapportés à une tempête ou un tsunami d'âge 6500 ou 8000 ans B.P. ayant affecté le littoral de Jerba.
Après cet épisode de haute énergie, le milieu devient moins agité et s'y décantent les argiles de l'unité U6. Ils s'y développent essentiellement C. torosa et X. aurantia (zone B5). L'enrichissement de C. torosa et la réduction de X. aurantia témoigne de la fermeture du milieu (F4). Au passage U6/U7 l'ostracofaune disparaît confirmant le passage vers le milieu actuel de type sebkha.
L'ostracofaune présente dans les sédiments des carottes SG1 et BSC2 nous a permis de caractériser trois phases d'évolution des paléo-environnements à l'emplacement de la Sebkha El-Guettiate de Skhira et de préciser et les forçages naturels responsables de cette évolution. Ces phases sont mises en évidence par les variations de l'abondance relative des différentes associations, de la densité, de la richesse spécifique et des indices de diversité. Une évolution en trois phases peut y ętre reconnue :
i) la première phase, d'âge > 6595 ans B.P. et 6055 ans, présente ŕ la base un milieu lagunaire largement ouvert à ostracodes marins dominants, et caractérisé par une influence marine. Le développement et la dominance de Cyprideis torosa, espèce euryhaline et eurytherme, signe le caractère perturbé du milieu. Puis, l'apparition brutale des ostracodes saumâtres montre que la lagune estuarienne est soumise ŕ des influences fluviatiles responsables de l'euryhalinité des eaux de la lagune. Cet environnement serait synchrone à une période de forte pluviométrie induisant l'euryhalinité des eaux de la lagune et sa richesse en grains de quartz et en espèces saumâtres. Il évoluera par la suite vers un milieu lagunaire moins agité (aux environs de 6055 ans B.P.) où les ostracodes saumâtres, toujours dominants, s'associent aux taxons lagunaires ;
ii) la deuxième phase, non datée directement, serait située entre environs 6055 et 5150 ans B.P.). Elle marque l'installation d'une lagune estuarienne soumise aux influences marines et aux courants de dérive littorale responsables de la présence des ostracodes marins. Ces courants amčnent la fermeture de la lagune et la dominance des espèces saumâtres ;
iii) la troisième phase, de 5150 ans B.P. à l'actuel, débute avec l'installation d'un environnement lagunaire, où les ostracodes lagunaires sont dominants. Au cours de son évolution, cette lagune, passant par des périodes d'émersion, subira des dépôts de type washover représentés par des faciès bioclastiques de haute énergie, traduction probable d'un évènement climatique extrême. Enfin le développement des ostracodes saumâtres signe la fermeture du milieu et son passage vers l'état actuel de sebkha.
L. (2010).- Morphogenèse et paléoenvironnements depuis le Pléistocène supérieur en Tunisie centrale : l'exemple des piémonts du chaînon Goubrar/Khechem.- Zeitschrift für Geomorphologie, Berlin, vol. 54, n° 1, p. 65-92.
J. (1979).- The ecology and distribution of the littoral ostracods of Cyprus.- Journal of Natural History, London,vol. 13, n° 2, p. 135–160.
L., P. & P. (1979).- Les foraminifčres.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 171–209.
G., M., G. & N. (1979).- Les ostracodes benthiques.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 280-284.
S., Y., E. & J. (2003).- Néotectonique affectant les dépôts marins tyrrhéniens du littoral sud-est tunisien : implication pour les variations du niveau marin.- Comptes Rendus Géoscience, Paris, vol. 335, p. 247-254.
E.M. (1988).- Palaeobathymetry on the continental shelf based on examples using ostracods from the Gulf of Alaska. In : P., J.P. & J.P. (eds.), Ostracoda in the Earth Sciences.- Elsevier, Amsterdam, p. 55–76.
A. (1979).- Recherches palynologiques sur les sédiments du Golfe de Gabčs : résultats préliminaires.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 247–264.
A. (1992).- Pollens dans les séries marines du Golfe de Gabčs et du plateau des Kerkennah (Tunisie): signaux climatiques et anthropiques.– Quaternaire, Paris, vol. 3, n° 1, p. 31–39.
G. & J.P. (1977).- Intérêt de la Sebkha El Melah (Sud tunisien) pour l'interprétation des séries évaporitiques anciennes.- Sedimentary Geology, Amsterdam, vol. 19, p. 139-164.
P. (1980).- Les ostracodes et leur intérêt dans la définition des écosystèmes estuariens et de plateforme continentale. Essais d'application à des domaines anciens.- Thèse Doctorat čs Sciences naturelles, Université de Bordeaux I ; Mémoires de l'Institut de Géologie du Bassin d'Aquitaine, Talence, n° 11, 350 p.
P. (1982).- Les Ostracodes, traceurs des variations hydrologiques dans les systèmes de transition eau douce-eau salée.- Mémoires de la Société Géologique de France, Paris, vol. 144, p. 117-128.
P., P.H., M., A.M., C. & M. (1981).- Évolution du Lac du Tunis. Un modèle de passage du milieu littoral à un milieu lagunaire.- Actes du Premier Congrès National des Sciences de la Terre, Tunis, p. 91-100.
P. & M. (1981).- Comportement des microfaunes benthiques en milieu lagunaire.- Actes du Premier Congrès National des Sciences de la Terre, Tunis, p. 127-139.
L. (1979).- Les peuplements de phanérogames de la zone euphotique.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 271–274.
L. & S. (1979).- Bryozoaires.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 265–270.
R.G. (1989).- A 17000-year glacio-eustatic sea level record: influence of glacial melting rates on the Younger Dryas event and deep-ocean circulation.- Nature, London, vol. 342, p. 637-642.
G., C.A. & E. (2010).- Catastrophic event recorded among Holocene eolianites (Sidi Salem Formation, SE Tunisia).- Sedimentary Geology, Amsterdam, vol. 224, p. 38–48.
P. & I. (2005).- The use of ostracods from marginal marine, brackish waters as bioindicators of modern and Quaternary environmental change.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 225, n° 1-4, p. 68-92.
A.Z., R., M., S. & K. (2007).- Sedimentary dynamics and ecological state of Nakta tidal flat (littoral), South of Sfax, Gulf of Gabès, Tunisia).- Chinese Journal of Geochemistry, Guizhon, vol. 26, n° 3, p. 244-251.
H.I. (2001).- Ostracod evolution and extinction fits biostratigraphic value in the European Quaternary.- Quaternary Science Reviews, Amsterdam, vol. 20, p. 1743-1751.
Y. (2000, inédit).- Sédimentologie et géochronologie des dépôts littoraux quaternaires : reconstitution des variations des paléoclimats et du niveau marin dans le Sud-est tunisien.- Thèse de Doctorat d'État čs Sciences géologiques, Université de Tunis II, 338 p.
Y., N., M., M.H., A. & M. (1998).- A high relative sea level stand in the middle Holocene of southeastern Tunisia.- Marine Geology, Amsterdam, vol. 147, p. 123-130.
M. (1982, inédit).- Faciès sédimentaires et processus dynamique dans la formation d'un milieu lagunaire : évolution holocène et actuel du lac de Tunis et de ses abords.- Thèse de Doctorat de 3ème cycle, Université de Bordeaux I, 218 p.
A.M. (1989).- Écologie des Ostracodes du domaine méditerranéen : application au Golfe de Gabes (Tunisie Orientale). Les variations du niveau marin depuis 30000 ans.- Documents des Laboratoires de Géologie, Lyon, vol. 108, 239 p.
A.M. & A.M. (1988).- Les ostracodes et l'évolution paléogéographique au Quaternaire récent du site d'Ashtart (Golfe de Gabès, Tunisie Orientale).- Geobios, Villeurbanne, vol. 21, p. 73-80.
A.M. & A.M. (1990).- Les ostracodes et les variations paléoeustatiques du golfe de Gabès (Méditerranée) depuis 30000 ans.- Bulletin de la Société géologique de France, Paris, vol. 8, p. 113-122.
R., M., M.H. & A. (2006).- A Mediterranean Holocene restricted coastal lagoon under arid climate: Case of the sedimentary record of Sabkha Boujmel (SE Tunisia).- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 241, p. 177-191.
K. (2002).- Sea level change from mid Holocene to Recent time: an Australian example with global implications. In : J.X. & B.L.A. (eds.), Ice Sheets, Sea Level and the Dynamic Earth.- American Geophysical Union, Geodynamics Series, Washington, vol. 29, p. 33-50.
C., N.G., R.A. & R.H. (2007).- Mid-Holocene evolution and paleoenvironments of the shoreface-offshore transition, north-eastern Argentina: New evidence based on benthic microfauna.- Marine Geology, Amsterdam, vol. 240, p. 43-56.
R. (1979, inédit).- Contribution à l'étude de la sédimentation littorale historique et actuelle au voisinage du delta de la Mejerda : étude du lac Rhar el Melah et de la Sebkha de l'Ariana.- Thèse de 3ème cycle, Université de Bordeaux, 248 p.
L., S., A., E., D. & E. (2008).- Late Holocene high resolution palaeoclimatic reconstruction inferred from Sebkha Mhabeul, southeast Tunisia.- Quaternary Research, Washington, vol. 70, n° 2, p. 240–250.
V., J., J.M., O., E., C., J.M. & F. (2007).- A multiproxy approach of the Holocene evolution of shelf-slope circulation on the NW Iberian continental shelf.- Marine Geology, Amsterdam, vol. 239, p. 1-18.
I., P., M., F., L., E., M. & E. (1999).- Late Quaternary sea-level changes along the Tyrrhenian coast near Orbetello (Tuscany, central Italy).- Marine Micropaleontology, Amsterdam, n° 37, p. 289–311.
S., C., F. & M. (2007).- Impact sur les biophases marines des rejets anthropiques notamment des métaux lourds rejetés sur le littoral nord de la ville de Sfax (Tunisie).- Techniques Sciences Méthodes, Paris, n° 10, p. 71-85.
C. & P.A. (2005).- Mid-Holocene emergence of southern Tunisian coasts.- Marine Geology, Amsterdam, vol. 220, p. 205-213.
M.T. (1979).- Les kystes de Dinoflagellés.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 221–246.
M.T. (2002).- Evolution of the sebkhas in the gulf of Gabes (Tunisia) at the upper Pleistocene-Holocene transition.- Quaternaire, Paris, vol. 13, n° 2, p. 111-123.
C. (1979).- Les nannofossiles calcaires.- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 210–220.
S. & H. (1980).- African environmental and climatic changes and the general atmospheric circulation in late Pleistocene and Holocene.- Climatic Change, Boston, vol. 2, n° 4, p. 313-348.
B., K., H., N. & C. (1998).- Nouvelles données paléoenvironnementales pour le Quaternaire récent en Tunisie centrale (bassin de Maknassy).- Comptes Rendus de l'Académie des Sciences, Paris, (IIA), vol. 326, n° 12, p. 855–861.
A. (1986).- Jerba et Kerkena (îles de la côté orientale de la Tunisie) : leur évolution géomorphologique au cours du Quaternaire.- Publication de la Faculté de Lettres de Tunis, vol. XXI, 210 p.
A., R. & P. (1982).- Le Tyrrhénien de la Tunisie : essai de synthèse.- Bulletin de la Société géologique de France, Paris, (7čme série), t. XXIV, n° 2, p. 173-178.
T. & T. (2005).- Ecological analysis of benthic ostracods in the northern Japan Sea, based on water properties of modern habitats and late Cenozoic fossil records.- Marine Micropaleontology, Amsterdam, vol. 55, p. 255-276.
A. & P. (1992).- Distribution and annual variations of Loxoconcha elliptica in the Gernika estuary (Bay of Biscay).- Geobios, Villeurbanne, vol. 25, n° 4, p. 495-503.
R. & P. (1977).- Les formations quaternaires de l'île de Jerba (Tunisie) : essai sur les lignes de rivage.- Compte Rendu sommaire des Séances de la Société géologique de France, année 1977, fasc. 4, p. 217-219.
E., A. & J. (2003).- Stratigraphy of washover deposits in Florida: implications for recognition in the stratigraphic record.- Marine Geology, Amsterdam, vol. 200, p. 31-48.
N., P.F., J.L., M., J.C. & A. (1991).- Paléoclimats holocènes du Sahara septentrional. Dépôts lacustres et terrasses alluviales en bordure du Grand Erg Oriental à l'extrême-Sud de la Tunisie.- Comptes Rendus de l'Académie des Sciences, Paris, vol. 312, n° 13, p. 1661-1666.
E.C. (1966).- The measurement of diversity in different types of biological collections.- Journal of Theoretical Biology, Amsterdam, vol. 13, p. 131-144.
J.C., C.H. & C.V. (1985).- Sediment and pollen evidence for an early to mid-Holocene humid period in the eastern Sahara.- Nature, London, vol. 314, p. 352-355.
J.C. & C.V. (1987).- Holocene vegetation zonation in the eastern Sahara.- Nature, London, vol. 330, p. 645-647.
J.C. (1979).- Mollusques testacés (Macrofaune).- Géologie Méditerranéenne, Marseille, t. VI, n° 1, p. 143–170.
F., M.L., J.M., J.G., A., L. & J. (2003).- Population age structure techniques and ostracods: Applications in coastal hydrodynamics and paleoenvironmental analysis.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 199, p. 51-69.
F., M., E., I., I., M., J.L. & M. (2006).- The present environmental scenario of El Melah Lagoon (NE Tunisia) and its evolution to a future sebkha.- African Earth Sciences Review, Oxford, vol. 44, p. 289-302.
P., L., M., L., C., F., M. & P. (2008).- Reconstruction of paleostorm events in a coastal lagoon (Herault, South of France).- Marine Geology, Amsterdam, vol. 251, p. 224–232.
M.A., H.F., S.M. & W.H. (2003).- Analyse d'ensemble de Foraminifères benthiques actuels de la côte nord-ouest méditerranéenne de l'Égypte.- Revue de Micropaléontologie, Paris, vol. 46, p. 111-130.
F.A. & A.T. (1979).- Global maps of lake-level fluctuations since 30000 yr. B.P.- Quaternary Research, Washington, vol. 12, p. 83-118.
I.P., B S.A. & S.Z. (2005).- The impact of late Holocene environmental change on lacustrine ostracoda in Armenia.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 225, p. 187-202.
J.E., A. & W. (2007).- Benthic foraminiferal biodiversity response to a changing Arctic palaeoclimate in the last 24.000 years.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 255, n° 3, p. 195-222.
C., F., D., Y. & M. (2006).- Impact de la genèse et de l'évolution de la flèche sableuse de Rass Kaboudia sur les environnements littoraux de la région de Chebba (Tunisie). Apport des foraminifères benthiques et des mollusques.- Notes du Service Géologique de Tunisie, Tunis, vol. 74, p. 93-106.
J. (1974, inédit).- Les peuplements malacologiques dans les biocénoses lagunaires tunisiennes. Étude de la biologie de l'espèce pionnière Cerastoderma glaucum .- Thèse de Doctorat d'État es Sciences naturelles, Université de Caen, 345 p.
K. (2006).- Les environnements holocčnes et actuels dans le bassin versant de l'Oued Sbeďtla (Tunisie centrale).- Thčse de doctorat, Université Denis-Diderot (Paris 7), 333 p.
C. & D. (2008).- Mid and Late Holocene fluvial chronology of Tunisia.- Quaternary Science Reviews, New York, vol. 27, p. 580-588.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Carte de localisation de la Sebkha El–Guettiate et emplacement des carotte SG1 et BSC2. A à F : positions des photos détaillées de la
Fig. 2 ![]() [Droits réservés : Image © 2011 DigitalGlobe - Data SIO NOAA, U.S. Navy, NGA, GEBCO
- © 2011 Google - Image © 2011 GeoEye].
[Droits réservés : Image © 2011 DigitalGlobe - Data SIO NOAA, U.S. Navy, NGA, GEBCO
- © 2011 Google - Image © 2011 GeoEye].
Note éditoriale : "Les auteurs seront tenus pour 'seul responsable' de
l'utilisation faite des textes, illustrations (tableaux, dessins), photographies
et vidéos qu'ils auront fournis et utilisés dans leurs publications
respectives."
Figure 1: Location map of El–Guettiate Sebkha and location of SG1 and BSC2
cores. A to F: Positions of the detailed photos of the Fig. 2 ![]() [Some rights reserved: Image © 2011 DigitalGlobe - Data SIO NOAA, U.S. Navy,
NGA, GEBCO - © 2011 Google - Image © 2011 GeoEye].
[Some rights reserved: Image © 2011 DigitalGlobe - Data SIO NOAA, U.S. Navy,
NGA, GEBCO - © 2011 Google - Image © 2011 GeoEye].
Editorial note: "The authors are the 'sole responsible' for the usage made
of texts, illustrations (tables and drawings), photos and videos provided and
used in their respective publications."
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : A : cordons littoraux fossiles orientées NE-SO au niveau de la Sebkha Dreîaa ; B : mégarides obliques par rapport à ligne côte ; C : delta de flot au niveau du chenal de marée de la
Sebkha Dreîaa ; D : delta de jusant au niveau du chenal de marée de la
Sebkha Dreîaa ; E : Hauts-fonds colonisés par les végétaux Ammophilia arenaria; F : dépôt de washover entaillant le cordon littoral
actuel
[Droits réservés : © 2011 Google - Imagerie © DigitalGlobe, Cnes/spot Image,
GeoEye].
Note éditoriale : "Les auteurs seront tenus pour 'seul responsable' de
l'utilisation faite des textes, illustrations (tableaux, dessins), photographies
et vidéos qu'ils auront fournis et utilisés dans leurs publications
respectives."
Figure 2: A: fossilized dune ridge oriented NE-SO (Dreîaa Sebkha);
B: Wrinkles oblique to coastline; C: flood tidal delta observed at outlet of tidal channel (Dreîaa Sebkha);
D: Washover fan and tidal channel ; E: strand and ebb tidal delta at the inlet of the tidal channel (El-Guettiate Sebkha) ;
F: washover deposits cutting the offshore bar
[Some rights reserved: © 2011 Google - Imagerie © DigitalGlobe, Cnes/spot Image,
GeoEye].
Editorial note: "The authors are the 'sole responsible' for the usage made
of texts, illustrations (tables and drawings), photos and videos provided and
used in their respective publications."
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 3 : Répartition verticale des ostracodes de la carotte BSC2, âge, colonnes et unités lithologiques, répartition verticales des zones.
Figure 3: Ostracods vertical repartition along BSC2 core, lithologic units and columns, vertical distribution of zones.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 : Données sédimentologiques et microfaunistiques (ostracodes) de la carotte SG1 : Colonne lithologique, abondance des grains de quartz, répartition des taxons et des zones/sous-zones ainsi que du nombre des espèces et de l'abondance absolue.
Figure 4: Sedimentologic and microfaunistic (ostracods) data of SG1 core: Lithologic column, quartz grain abundance, vertical distribution of taxa, zones/sub-zones, species and individual number.
CT: Cyprideis torosa ; LxE: Loxoconcha elliptica ; Cmf: Cytheromorpha fuscata ; XA: Xestoleberis aurantia ; Lpf: Leptocythere fabaeformis ; Lpm: Leptocythere macella ; LxR: Loxoconcha rhomboîdea ; LxN: Loxoconcha napoliana ; Lxag: Loxoconcha agillis ; Aup: Aurila prasina ; Cush: Cushamanidea ; Crf: Cytherois fischeri ; Uro: Urocythereis oblonga ; Urof: Urocythereis favosa ; Bass: Basselerites berchoni ; Sinc: Semicytherura incongruens ; Srg: Semicytherura ruggierii ; Sprx: Semicytherura paradoxa ; Ssl: Semicytherura sella ; Pctd: Paracytheridea depressa ; Car: Carinocythereis carinata ; Neof: Neocytherideis fasciata ; Hilt: Hiltermannicythere emaciata
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 5 : Évolution des associations, du nombre d'individus et du nombre d'espèces des ostracodes le long de la carotte BSC2.
Figure 5: Evolution of ostracod assemblages, individuals and species number along SBSC2 core.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 6 : Évolution des associations, du nombre d'individus et du nombre d'espèces des ostracodes le long de la carotte SG1.
Figure 6: Evolution of ostracods assemblages, individuals and species number along SG1 core.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 7 : Représentation schématique des étapes de la formation de la Sebkha El-Guettiate à partir de l'étude des assemblages des ostracodes.
Figure 7: Main stages of formation of the El-Guettiate Sebkha deduced from ostracods assemblages.