![]()
Sommaire
[1 - Cadre géographique] [2 - Cadre géologique et structural]
[3 - Présentation du gisement]
[4 - Biostratigraphie]
[5 - Paléoécologie des ostracodes]
[6 - Paléobiogéographie des ostracodes]
[7 - Taxonomie des ostracodes]
[Références bibliographiques]
[Appendice] et ...
[Planches]
Laboratoire de Paléontologie Stratigraphique et Paléoenvironnement, Université d'Oran, BP 1524,
El M'naouer, Oran
3100 (Algérie) ;
Département des Sciences de la Terre, Faculté des Sciences Exactes des Sciences de la Nature et de la Vie,
Université de Jijel, BP 98 Ouled Aissa, 18000 Jijel (Algérie)
* Auteur correspondant - Corresponding author
Impasse des Biroulayres, 33610 Cestas (France) ;
Universidade de Lisboa, Faculdade de Ciências, Centro de Geologia, Campo Grande, C-6, 3° 1749-016
Lisboa (Portugal)
Laboratoire de Paléontologie Stratigraphique et Paléoenvironnement, Université d'Oran, BP 1524, El M'naouer, Oran 3100 (Algérie)
Manuscrit en ligne depuis le 5 Juillet 2011
![]()
Un gisement bien daté par une riche microflore de charophytes caractéristique de la zone à Peckichara disermas, de l'Éocène inférieur (Yprésien inférieur = Sparnacien = Ilerdien) a livré une intéressante association d'ostracodes non-marins dans le Djebel Amour, Atlas Saharien occidental, en Algérie. Cette faune est caractérisée par la dominance du genre Neocyprideis avec l'espèce nouvelle Neocyprideis meguerchiensis nov. sp. et la présence occasionnelle des genres Hemicyprideis, Perissocytheridea (P. algeriensis nov. sp.) et Limnocythere. Il s'agit de la seconde découverte du genre Neocyprideis dans le Paléogène du continent africain, la première étant dans l'Éocène inférieur du Sénégal. Les Neocyprideis sont représentés à 75 % par des formes lisses, normalement calcifiées, à 20 % par des individus réticulés et tuberculés et à 5 % par des spécimens uniquement réticulés. Ce polymorphisme suggère une saisonnalité marquée avec une salinité variable. La dominance des morphes lisses bien calcifiés indique un milieu avec un équilibre entre le Ca++ et le Mg++, les morphes réticulés caractérisant un milieu plus riche en Mg++. La présence de forme tuberculées-réticulées suggère des apports saisonniers organo-siliceux continentaux et une salinité pouvant temporairement atteindre des valeurs inférieures ou égales à 5 psu.
Ostracodes ; milieu lagunaire ; Éocène inférieur ; Algérie ; Atlas saharien ; taxonomie ; paléoécologie ; paléobiogéographie.
F., J.-P. & F. (2011).- Un gisement d'ostracodes non-marins dans l'Éocène inférieur du Djebel Amour, Atlas saharien central, Algérie : taxonomie, paléoécologie et paléobiogéographie.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2011/04 (CG2011_A04), p. 83-97.
An outcrop with non-marine ostracodes in the Early Eocene of Djebel Amour, central Saharian Atlas, Algeria: taxonomy, palaeoecology and palaeobiogeography.- An outcrop with non-marine ostracods in the Early Eocene of Djebel Amour, an outcrop well dated by a rich charophyta microflora characterizing the Peckichara disermas Zone of the Early Eocene (Lower Ypresian = Ilerdian), has yielded an interesting non-marine ostracode fauna in the Djebel Amour, Western Saharian Atlas, in Algeria. This fauna is characterized by the dominance of the genus Neocyprideis with the new species Neocyprideis meguerchiensis nov. sp., and the occasional occurrence of the genera Hemicyprideis, Perissocytheridea (P. algeriensis nov. sp.) and Limnocythere. It is the second discovery of the genus Neocyprideis in the Palaeogene of the African continent, the first one being in the Early Eocene of Senegal. The Neocyprideis are represented by 75 % of smooth and normally calcified carapaces, 20 % by reticulate-noded specimens and 5 % of only reticulate forms. This polymorphism suggests a marked seasonality with fluctuating salinity. The dominance of smooth and well calcified morphs indicates an environment with an equilibrium Ca++ / Mg++, the reticulate morphs characterizing waters richer in Mg++. The presence of noded-reticulate forms suggests seasonal influxes of organo-siliceous material from the continent and a salinity which could temporarily reach values lower or equal to 5 psu.
Ostracodes; brackish environment; Early Eocene; Algeria; Saharan Atlas; taxonomy; palaeoecology; palaeobiogeography.
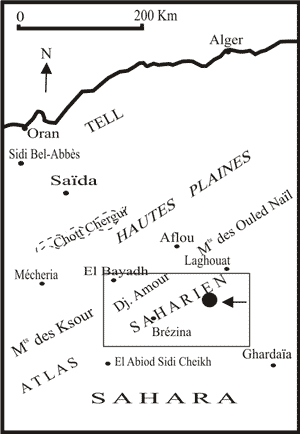
Le Djebel Amour (Atlas saharien central) constitue la partie centrale de l'Atlas saharien. Il est situé à 200 km au SE de la ville d'Oran et est limité, à l'W par les Monts des Ksour, à l'E par les Monts de Ouled Naïl, au S par le Sahara, et au N par les Hautes plaines et les Hauts Plateaux
(Fig. 1 ![]() ).
).
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 1 : Carte de localisation de la région étudiée.
Figure 1: Location map of the studied region.
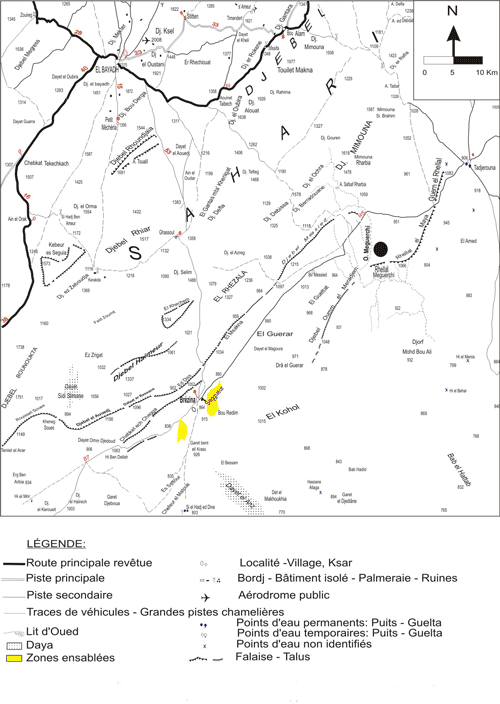
Notre secteur d'étude intéresse le gisement de l'Oued
Meguerchi ; celui-ci est situé à la bordure méridionale de l'Atlas saharien central, à 80 km environ à l'E de Brézina. L'Oued Meguerchi constitue un des principaux cours d'eau qui traversent le Djebel Amour et se perdent
vers le Sud aux abords du Grand Erg occidental
(Fig. 2 ![]() ).
).
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 : Carte de localisation du gisement de l'Oued Meguerchi (Atlas saharien central). Extrait de la carte topographique de Laghouat au
1/50 000.
Figure 2: Location map of the Oued Meguerchi outcrop (Central Saharan Atlas). Extracted from the
1/50,000 topographic map of Laghouat.
L'Atlas saharien correspond à une chaîne intracratonique plissée, orientée du SW au NE, qui s'interpose entre deux domaines
stables : le domaine des Hautes Plaines Oranaises au N, la plate-forme saharienne au S.
Cette chaîne fait 2550 km de long et est formée d'W vers l'E des Monts des Ksour,
du Djebel Amour et des Monts des Ouled Naïl
(Fig. 1 ![]() ).
Elle est limitée latéralement par deux grands accidents :
).
Elle est limitée latéralement par deux grands accidents :
Sous l'effet de la subsidence s'y sont accumulés durant le Mésozoïque et une partie du Cénozoïque de puissantes séries néritiques et continentales (, 1952). Les sédiments sont de plus en plus jeunes quand on se dirige vers l'E. Les faciès continentaux dominent dans les régions occidentales de la chaîne. Pendant le Cénozoïque, lors de la phase orogénique pyrénéenne (ou atlasique), les dépôts de ce sillon furent énergiquement plissés et donnèrent naissance à des plis simples mais de forte amplitude, qui constituent l'un des traits caractéristiques de l'Atlas saharien. D'une façon générale, les anticlinaux présentent le type coffré, à flanc fortement redressé et à charnière plate, les synclinaux à fond plat et large sont souvent perchés.
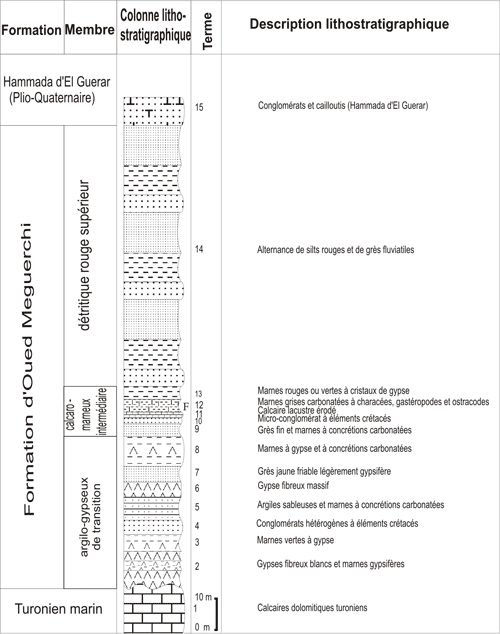
L'Oued Meguerchi oů se situe le gisement étudié recoupe les terrains continentaux néogènes de la Hammada d'El Guerar et les terrains mésozoïques et cénozoïques du Djebel Messied.
Le gisement est situé sur la rive droite de l'Oued
Meguerchi a été découvert par M.
(Fig. 3 ![]() ). La partie inférieure de
la formation étudiée repose en concordance sur des calcaires dolomitiques turoniens
marins dont elle est séparée par une surface de ravinement. La partie supérieure
est coiffée par les dépôts gréso-carbonatés continentaux
discordants d'âge plio-quaternaire (, 1952) qui appartiennent à la Formation de la Hammada d'El Guerar.
). La partie inférieure de
la formation étudiée repose en concordance sur des calcaires dolomitiques turoniens
marins dont elle est séparée par une surface de ravinement. La partie supérieure
est coiffée par les dépôts gréso-carbonatés continentaux
discordants d'âge plio-quaternaire (, 1952) qui appartiennent à la Formation de la Hammada d'El Guerar.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 3 : Coupe lithostratigraphique du gisement de l'Oued
Meguerchi.
Figure 3: Lithostratigraphic log of the Oued Meguerchi outcrop.
La Formation de l'Oued Meguerchi est subdivisée, de bas en haut, en trois membres : un membre argilo-gypseux inférieur (dit "de transition"), un membre calcaréo-marneux intermédiaire et un membre détritique rouge supérieur.
a. Le membre argilo-gypseux de transition (40-50 m)
Neocyprideis sp. 1, sp. 2, sp. 3 , 1999 (très probablement morphotypes d'une seule espèce) : Éocène moyen.
Ce membre est constitué par des argiles roses à la base, dont les fentes de dessiccation sont remplies de gypse diagénétiques blanc, passant vers le haut à un ensemble de marnes verdâtres à gypses de quelques dizaines de mètres dans lequel s'intercale un banc de conglomérat. La partie supérieure de ce membre comporte un banc de gypse fibreux surmonté par des grès jaunes et des marnes gypsifères. Notons qu'aucun fossile n'a été trouvé dans ce membre.
b. Le membre calcaro-marneux intermédiaire (15 m)
Il affleure sur une partie de la rive droite de l'Oued. Il débute par des bancs de grès fins séparés par des marnes verdâtres ou grisâtres légèrement gypsifères, rarement carbonatées. Dans ces niveaux marneux, s'intercalent un mince passage conglomératique et un banc de calcaire lacustre. Ce banc de calcaire est surmonté par une
série de marnes carbonatées de couleur grisâtre. Ces niveaux marneux sont très riches en gastéropodes, ostracodes et charophytes
(Fig. 3 ![]() , terme 12).
, terme 12).
c. Le membre détritique rouge supérieur (100 m)
C'est une puissante série, constituée par une alternance métrique de silts rouges gréseux et des grès fluviatiles de couleur rougeâtre. Cet ensemble devient de plus en plus détritique vers le sommet. Notons également que ces bancs gréseux sont souvent mal cimentés et contiennent des dragées de quartz; ils montrent par endroit quelques fragments de bois fossilisés très érodés, sans structure apparente.
La flore de charophytes se compose de huit espèces (, 1993; et alii, 1997) dont cinq décrites dans ce gisement ( & , 1999), Peckisphaera bessediki & , 1999, Harrisichara meguerchiensis & , 1999, Peckichara atlasensis & , 1999, Neochara ameuriorium & , 1999, Gyrogona caudata & , 1999, Nitellopsis (T.) thaleri ( & , 1969) & , 1972, Maedleriella pachycera & , 1969, Maedleriella cristellata , 1977.
Grâce à la présence des espèce Maedleriella pachycera et Nitellopsis (T.) thaleri, la flore de Meguerchi peut être corrélée avec la zone de Monze, elle-même rattachée à la zone cuisienne à Nitellopsis (T.) thaleri de (1986). Cependant, la nouvelle datation de cette flore de Monze, fondée sur les foraminifères ( et alii, 1989), conduit à la considérer comme représentant plutôt la zone à Peckichara disermas du Sparnacien/Ilerdien.
L'association de l'ensemble de ces espèces de charophytes nous a permis de proposer un âge Sparnacien-Ilerdien (Éocène inférieur) à la formation de l'Oued Meguerchi ( et alii, 1997).
Les ostracodes sont très abondants et bien conservés dans le niveau étudié (plus de 200 spécimens). L'espèce dominante (environ 200 valves et carapaces) est une espèce nouvelle, Neocyprideis meguerchiensis nov. sp. Les autres espèces sont accessoires : Perissocytheridea algeriensis nov. sp. (15 valves), Limnocythere sp. (une seule valve mal préservée) et Hemicyprideis sp. (2 valves). Les espèces étant nouvelles, elles ne permettent pas d'apporter des éléments de datation.
Pratiquement tous les genres représentés dans ce niveau sont eury- et mesohalins : Neocyprideis, Perisssocytheridea, Hemicyprideis. Un seul genre, plus strictement dulçaquicole ou oligohalin, a été trouvé. Il s'agit d'un Limnocythere, représenté par un seul spécimen. Ceci est en accord avec (1972), qui avance que Neocyprideis n'a jamais été trouvé avec des ostracodes d'eau douce.
Le genre dominant est clairement Neocyprideis, avec Neocyprideis meguerchiensis nov. sp. (95 % de la population). Ceci, par contre, contredit les données de (1972, 1977, 1990) qui avance que Neocyprideis forme exceptionnellement des assemblages monospécifique contrairement à Hemicyprideis, et qu'il est plus courant comme un constituant mineur (5-10 %) d'associations d'ostracodes méso- ou polyhalins. Pour cet auteur, Neocyprideis est un genre euryhalin moins adaptable qu'Hemicyprideis.
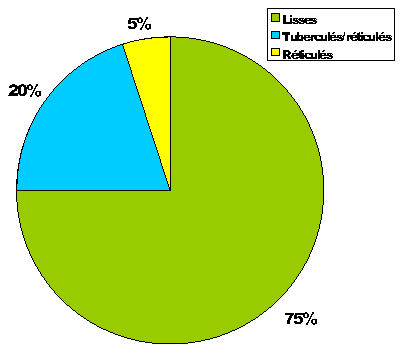
Dans l'échantillon étudié, cette espèce est nettement polymorphe
(Fig. 4 ![]() ) avec des spécimens lisses (75 %), tuberculés (tubercules creux, ou
"nodes") et réticulés (20 %) et seulement réticulés (5 %). Ce polymorphisme suggère une saisonnalité marquée avec une salinité variable ( et alii,
1988a). La dominance des morphes lisses bien calcifiés indique un milieu avec un équilibre entre le
Ca++ et le Mg++, les morphes réticulés caractérisant un milieu plus riche en
Mg++. La présence de formes tuberculées-réticulées suggère des apports saisonniers organo-siliceux continentaux ( et alii,
1988b ; ,
1985) et une salinité pouvant temporairement atteindre des valeurs inférieures ou égales à 5 psu (,
1990, 1993). Chez le genre actuel proche Cyprideis , 1857 ( & ,
1990), la présence de tubercules a été depuis assez longtemps utilisée comme un marqueur (direct ou indirect) de faible salinité (,
1975 ; van , 1996,
2000, …).
) avec des spécimens lisses (75 %), tuberculés (tubercules creux, ou
"nodes") et réticulés (20 %) et seulement réticulés (5 %). Ce polymorphisme suggère une saisonnalité marquée avec une salinité variable ( et alii,
1988a). La dominance des morphes lisses bien calcifiés indique un milieu avec un équilibre entre le
Ca++ et le Mg++, les morphes réticulés caractérisant un milieu plus riche en
Mg++. La présence de formes tuberculées-réticulées suggère des apports saisonniers organo-siliceux continentaux ( et alii,
1988b ; ,
1985) et une salinité pouvant temporairement atteindre des valeurs inférieures ou égales à 5 psu (,
1990, 1993). Chez le genre actuel proche Cyprideis , 1857 ( & ,
1990), la présence de tubercules a été depuis assez longtemps utilisée comme un marqueur (direct ou indirect) de faible salinité (,
1975 ; van , 1996,
2000, …).
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 4 : Proportions des divers morphes de Neocyprideis meguerchiensis nov. sp.
Figure 4: Proportions of various morphs of Neocyprideis meguerchiensis nov. sp.
Le genre Perissocytheridea, selon & 1979) et et alii (1996), est typiquement lagunaire. Dans les baies actuelles du Golfe du Mexique, ce genre disparaît pratiquement totalement lorsque la salinité approche de 30 psu. Selon (1977), en Floride ce genre est oligo- à polyhalin, plutôt mesohalin (5-18 psu).
Selon (1971), le genre Hemicyprideis est un genre euryhalin présent dans des lagons tropicaux à subtropicaux, abondant dans des eaux de salinité comprise entre 0 et 18 psu, mais aussi pouvant vivre dans des milieux hyperhalins et dans des eaux polyhalines et euhalines.
Le genre Neocyprideis est connu depuis le Crétacé supérieur (Cénomanien-Turonien) de la marge N de la Téthys (Europe du Sud) et du Moyen-Orient ( et alii, 1990 ; , 2002). Pendant le Crétacé supérieur (Sénonien, Maastrichtien), il reste cantonné à l'Europe du Sud. Sa présence en Amérique du Sud (Argentine) est peu probable, les espèces attribuées à ce genre appartenant certainement au genre Fossocytheridea & , 1964 (comm. perso. E. ).
Nous restreignons l'attribution à ce genre au concept de Neocyprideis s.s., contrairement à & (2006), qui mettent en synonymie les genres Miocyprideis , 1960, et Bishopina , & , 1976.
Ce genre a réellement commencé à se diversifier à partir du Paléocène supérieur en Europe de l'Ouest (, 1972), puis surtout dans l'Éocène et l'Oligocène ( et alii, 1990 ; , 1990). Sur le continent africain jusqu'à présent n'étaient connues que des présences ponctuelles dans l'Éocène de Somalie ( & , 1996) et l'Éocène inférieur du Sénégal (, 1989). La découverte en Afrique du Nord d'une espèce du genre Neocyprideis, corrobore en partie la théorie de (1989), supposant "un flux migratoire avien Nord/Sud". Cet auteur suggérait aussi des "étapes" migratoires probables en Mauritanie et au Maroc.
Pendant le Paléogène, le genre Neocyprideis est aussi présent en Oman ( et alii, 1991 ; & , 1991 ; et alli, 1992), au Pakistan (, 2000), en Inde (, 1988 ; , 1996 ; & , 2002) et dans le Bassin de Tarim en Chine ( et alii, 1995). Selon (2005), le genre Neocyprideis serait toujours existant, dans l'Actuel d'Indonésie.
L'apparition des formes tuberculées ("nodées") date du Sénonien ( et alii, 1999), et surtout à partir du Maastrichtien en Europe méridionale (Neocyprideis murciensis & , 1971, non Neocyprideis durocortoriensis , 1956, sensu & , 2006). Dans le Paléogène, les espèces ayant des morphes tuberculés sont connues jusqu'à l'Oligocène, en Europe et en Algérie. Citons par exemple :
Le genre Hemicyprideis était jusqu'à présent uniquement connu en Europe et en Chine ( & , 2007), surtout dans l'Oligocène et le Miocène (, 1978). En Algérie il a été trouvé dans le Miocène ( & , 1997).
Le genre Perissocytheridea est un genre d'origine gondwanienne. Sur les continents africain ( et alii, 1996 ; , 1976) et sud-américain & , 1978 ; et alii, 2000), en Grčce (, 1988), et au Moyen-Orient (, 2002), il est connu dans le Crétacé supérieur, depuis le Cénomanien. Il est possible que ce genre soit présent dans le Cénozoïque européen. En effet certaines espèces attribuées au genre Cladarocythere , 1972 (famille des Limnocytheridae), pourraient en fait appartenir au genre Perissocytheridea telles que Cladarocythere fredericensis (, 1956) du Lutétien du Bassin de Paris et Cladarocythere sp. 1 , 1985, du Thanétien supérieur de l'Aude, S de la France (voir illustrations in et alli, 1985).
Dans le Crétacé supérieur, des espèces attribuables à ce genre ont été décrites sous différentes appellations génériques : Neocythere , 1956 ( & , 1986, Turonien-Coniacien d'Israël), Afrocytheridea , 1975 et alli, 2010, Santonien du Brésil), Soudanella , 1961 , 2009, Coniacien-Santonien du Brésil), … La présence dans l'Éocène d'Algérie est la plus jeune occurrence sur le continent africain. En Amérique ce genre est toujours présent ( & , 1979).
Les holotypes et paratypes sont déposés dans les collection du Départment de Géologie de l'Université de Lisbonne (EJPCH = holotypes; EJPCP = paratypes).
Classe Ostracoda , 1806
Sous-classe Podocopa .. , 1894
Ordre Podocopida ,
Sous-ordre Podocopina , 1866
Super-famille Cytheroidea , 1850
Famille Cytherideidae , 1925
Sous-famille Cytherideinae , 1925
Genre Neocyprideis , 1956
Neocyprideis meguerchiensis nov. sp.
(Pl. 1 ![]() ,
figs. 1-8)
,
figs. 1-8)
Origine du nom : De la localité type, l'Oued Meguerchi.
Holotype : Une valve gauche, femelle (Pl. 1 ![]() , fig. 1), EJPCH 2011-01.
, fig. 1), EJPCH 2011-01.
Paratypes : 7 valves, EJPCP 2011-01, 01-07.
Matériel : Environ 200 valves.
Localité-type : Éocène inférieur (Sparnacien-Ilerdien), Oued Meguerchi, Atlas saharien central, Algérie.
Diagnose : Espèce attribuée au genre Neocyprideis, seulement identifiable par ses morphotypes tuberculés. Les tubercules, de grande taille sont au nombre de 6, et portent des petites papilles.
Description sommaire : Les individus non tuberculés ne sont pas différenciables de la plupart des autres espèces tertiaires du genre. À noter une extrémité antérieure lisse et fortement comprimée, un sulcus subvertical antéro-dorsal bien marqué en arrière du tubercule oculaire. Le dimorphisme sexuel est prononcé, les femelles possédant une cavité incubatrice postérieure marquée en vue latérale par une extrémité postérieure relevée.
Les morphotypes tuberculés sont aussi réticulés. Les tubercules, creux ("nodes"), sont généralement au nombre de 6, de grande taille, et ornés de petites papilles. Les 2 plus gros sont situés en position postéro-ventrale et en arrière du sulcus.
La charnière est mérodonte-entomodonte, l'élément antérieur étant nettement plus long que le postérieur
(Pl. 1 ![]() ,
figs. 7-8). La zone marginale est étroite, sans vestibule. Les empreintes musculaires n'ont pas été observées.
,
figs. 7-8). La zone marginale est étroite, sans vestibule. Les empreintes musculaires n'ont pas été observées.
Dimensions :
Holotype : L = 0,920 mm; h = 0,540 mm
Paratypes : L = 0,660-1,090 mm; h = 0,380-0,620 mm
Remarques : Les individus non tuberculés sont très difficilement différenciables des autres espèces du Paléogène. Seules les morphes tuberculés sont diagnostiques. L'espèce la plus proche est Neocyprideis parallela (, 1902) de l'Oligocène d'Allemagne (, 1973). Cette espèce possède aussi de gros tubercules avec des papilles mais a aussi un gros tubercule postéro-dorsal, absent chez Neocyprideis meguerchiensis nov. sp., et de petits tubercules lisses en position antérieure et postérieure. De plus l'extrémité antérieure semble moins comprimée. Dans l'Éocène moyen terminal de l'Atlas Saharien, Algérie, (1999) illustre 3 espèces, Neocyprideis sp. 1, Neocyprideis sp. 2 et Neocyprideis sp. 3 qui sont certainement des morphotypes d'une seule espèce, mais sans types tuberculés. Cette espèce diffère de Neocyprideis meguerchiensis nov. sp. par son extrémité postérieure régulièrement arrondie.
Genre Hemicyprideis & , 1970
Hemicyprideis sp.
(Pl. 2  , fig. 8)
, fig. 8)
Matériel : 2 valves.
Remarques : Cette espèce de grande taille et d'aspect quasiment lisse, finement ponctuée et avec un faible sulcus antéro-dorsal, diffère de l'espèce la plus proche, Hemicyprideis clivosa , 1978, de l'Oligocène supérieur d'Allemagne, par ses bords ventral et dorsal parallèles. Hemicyprideis montosa ( & , 1889), du Sannoisien d'Europe du NW, a un contour proche mais est très fortement ponctué (, 1971, 1977).
Dimensions : L = 1,250 mm; h = 0,640 mm
Famille Cytheridae , 1850
Sous-famille Cytherinae , 1850
Tribu Perissocythereidini van den , 1963
Genre Perissocytheridea , 1938
Perissocytheridea algeriensis nov. sp.
(Pl. 2 ,
figs. 1-6)
Origine du nom : En provenance d'Algérie.
Holotype : Une valve gauche femelle (Pl. 2 , fig. 1), EJPCH 2011-02
Paratypes : 5 valves, EJPCP 2011-02,08-12.
Matériel : 15 valves.
Localité-type : Éocène inférieur (Sparnacien-Ilerdien), Oued Meguerchi, Atlas saharien central, Algérie.
Diagnose : Espèce attribuée au genre Perissocytheridea dont les femelles ont une carapace subtriangulaire lisse ou faiblement réticulée. Les mâles sont nettement plus allongés. Le bord ventral est caché par le renflement des valves.
Description sommaire : Les femelles sont de forme sub-triangulaire avec le bord dorsal rectiligne incliné vers l'arrière. Les valves sont lisses à faiblement réticulées et renflées ventralement. Les bords antérieur et postérieurs sont lisses et comprimés. On note la présence de deux sulci antéro-dorsaux en arrière du tubercule oculaire, le plus postérieur étant souvent très faible.
Les mâles sont nettement plus allongés que les femelles dans le tiers postérieur et leur carapace est
sub-rectangulaire. Le sulcus postérieur est plus distinct. Certains individus montrent un renflement postéro-dorsal et un faible tubercule musculaire
(Pl. 2 , fig. 6).
La charnière est entomodonte/mérodonte (Pl. 2 ,
figs. 3-4). Les empreintes musculaires n'ont pas pu être observées. La zone marginale est étroite, sans vestibule.
Dimensions :
Holotype : L = 0,640 mm; h = 0,340 mm
Paratypes : Femelles : L = 0,580 - 0,640 mm; h = 0,330-0,350 mm
Mâles : L = 0,640 - 0,680 mm; h = 0,310-0,330 mm
Remarques : L'espèce morphologiquement la plus proche est Perissocytheridea carrenoae & , 2008, du Miocène de Bolivie. Elle en diffère essentiellement par l'absence de troncation postérieure de la côte ventrale chez les individus femelles. Les individus mâles sont aussi très ressemblants à ceux de Perissocytheridea informalis , 1978 (in & , 1978) du Maastrichtien d'Argentine.
Famille Limnocytheridae ,
1938
Sous-famille Limnocytherinae , 1938
Genre Limnocythere , 1866
Limnocythere sp.
(Pl. 2 , fig. 7)
Matériel : Une seule valve.
Remarques : Cette espèce de Limnocythere possède 2 fortes épines coniques en arrière de la mi-longueur et une extrémité antérieure très comprimée.
Dimensions : L = 0,610 mm; h = 0,410 mm.
Nous tenons à remercier tout particulièrement notre collègue M.C. (Lisbonne, Portugal) pour les photos M.E.B, P. (Talence, France) pour ses judicieux commentaires, ainsi que feu E. (Commodoro Rivadavia, Argentine, décédé le 18 Mai 2011) pour les données concernant l'Argentine. Nous remercions aussi les deux rapporteurs, M.C. (Lisboa, Portugal) et J. (Bilbao, Espagne), pour les corrections apportées à notre texte. Ce travail a été présenté pour la première fois lors de la 22ème Réunion des Ostracodologistes de Langue Française (ROLF 23) organisée à Bruxelles en 2008 par J.-G. .
C.M. (2009, inédit).- Taxonomia, distrubuição estratigráfica e paleoecologia de ostracodes do Coniaciano ao Mioceno da Bacia de Santos, margem continental Sul do Brasil.- Thèse Doctorat, Universidade de Brasília-UnB, Brasilia, 108 p.
B. (1996).- Nouvelles espèces d'ostracodes du Turonien supérieur (?)-Coniacien (?)-Santonien de la région de Boulmane, Moyen Atlas, Maroc. Systématique, biostratigraphie et paléoécologie, paléobiogéographie des associations. In : S., de I. & J.-P. (éds.), Géologie de l'Afrique et de l'Atlantique Sud.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, Mémoire 16, p. 483-509.
V. & C. (1992).- Les ostracodes de la région Forcalquier-Manosque (bassin continental d'Apt; Haute-Provence).- Revue de Micropaléontologie, Paris, vol. 35, n° 2, p. 91-115.
J.-F. (1988).- Données préliminaires sur les associations d'ostracodes du Cénomanien de Grèce (Zone sub-pélagonienne, Argolide). Implications paléobiogéographiques et géodynamiques.- Geobios, Lyon, n° 21, fasc. 4, p. 435-463.
M.A.A. (2002).- Middle Cretaceous (Aptian - Early Turonian) Ostracoda from Sinai, Egypt.- Neue Paläontologische Abhandlungen, Dresden, Band 5, 123 p.
M.A.A. & P. (1996).- Middle Eocene ostracoda from Northern Somalia.- Courier Forschungstintut Senckenberg, Frankfurt am Main, n° 192, 139 p.
A. (1996).- Atlas of Paleogene ostracodes of Rajasthan basins.- Paleontographica Indica, Dehra Dun, n° 4, 157 p.
M. & B. (2006).- Les Marnes d'Auzas (Maastrichtien supérieur sous-pyrénéen). Stratigraphie et paléoenvironnements, associations d'ostracodes.- Revista Española de Micropaleontologia, Madrid, vol. 38, n° 2-3, p. 309-320.
G. (1989).- Les ostracodes saumâtres des lignites de l'Éocène inférieur au Sénégal; point de convergence des flux migratoires.- Courier Forschungsintitut Senckenberg, Frankfurt am Main, n° 113, p. 89-95.
P., J.-P., D., H. & I. (1988a).- The paleoecology of limnic ostracodes, a review of some major topics.- Palaeogeography, Palaeoecology, Palaeoclimatology, Amsterdam, special issue, vol. 62, n° 1-4, p. 413-461
P., P. & J.-P. (1988b).- Le polymorphisme induit par l'environnement : rôle du rythme saisonnier.- Strata, Toulouse, vol. 4, n° 1, p. 1-12.
G. & M.-D. (1997).- Ostracodes miocènes d'Algérie (Systématique, biostratigraphie, distribution palinspatique).- Mémoires du Muséum d'Histoire Naturelle de Lyon, Lyon, n° 1, 131 p.
J.-P. & P. (1990).- Phylogenetical affinities of Cytherissa with other Cytherideinae (Vernoniella, Fabanella, Neocyprideis, Cyprideis). A paleontological approach. In : D.L., P. & J.-P. (eds.), Cytherissa (Ostracoda) - The Drosophila of Paleolimnology.- Bulletin de l'Institut de Géologie du Bassin d'aquitaine, Talence, n° 47-48, p. 83-96.
J.-P., P. & R. (1990).- A review of the paleobiogeography and paleoecology of the closest group of Cytherissa: from the Mesozoic Fabanella and Vernoniella to the Cenozoïc Cyprideis. In : D.L., P. & J.-P. (eds.), Cytherissa (Ostracoda) - The Drosophila of Paleolimnology.- Bulletin de l'Institut de Géologie du Bassin d'Aquitaine, Talence, n° 47-48, p. 119-134.
J.-P., Y. & V.A. (1996).- Ostracodes limniques et lagunaires dans le Crétacé supérieur du Mali (Afrique de l'Ouest) : systématique, paléoécologie et affinités paléobiogéographiques.- Revue de Micropaléontologie, Paris, vol. 39, n° 3, p. 211-222.
A. (1947).- Observations géologiques sur l'Atlas Sud-Oranais.- Bulletin de la Société d'Histoire Naturelle d'Afrique du Nord, Alger, t. 33, p. 63-68.
B. (1962).- Nouveaux ostracodes de l'Éocène et de l'Oligocène de l'Aquitaine méridionale.- Actes de la Société Linnéenne de Bordeaux, Bordeaux, t. 100, p. 127-211.
O., C. & Y. (1985).- Paléogène. In : H.J. (éd.), Atlas des Ostracodes de France.- Bulletin des Centres de Recherche Exploration-Production Elf-Aquitaine, Pau, Mémoire 9, p. 257-311.
N. (1999).- Ostracodes de l'Éocène inférieur-moyen de l'Atlas Saharien (Algérie) : interprétation environnementale et paléogéographique.- Geobios, Lyon, vol. 32, n°3, p. 459-481.
J. (1952).- La chaîne de Djurdjura. Monographie régionale.- 19ème Congrès Géologique International, Alger, p. 1-48.
E. & R.F. (1979).- Zoogeography of Holocene Cytheracean ostracodes in the bays of Texas.- Journal of Paleontology, Tulsa, vol. 53, n° 4, p. 841-919.
C. (1981).- Ostracodes sparnaciens du Bassin de Paris (France).- Revue de Micropaléontologie, Paris, vol. 24, n° 1, p. 51-66.
C., de C. & J. (1991).- Ostracodes paléogènes du Dhofar (Oman). Intérêt stratigraphique et paléogéographique.- Revue de Micropaléontologie, Paris, vol. 34, n° 4, p. 297-311.
D. van (1996).- Cyprideis torosa (Ostracoda) revisited. Of salinity, nodes and shell size. In : M.C. (ed.), Proceedings of the 2nd European Ostracodologists Meeting.- British Micropalaeontological Society, London, p. 191-194.
D. van (2000).- Variable noding in Cyprideis torosa (Ostracoda: Crustacea): an overview, experimental resumes and a modem from catastrophe theory. In : .. & K. (eds.), Evolutionary Biology and Ecology of Ostracoda.- Hydrobiologia, Amsterdam, vol. 419, p. 131-139.
A. & A. (1986).- Late Turonian-Early Coniacian ostracodes from the Zihor Formation, southern Israel.- Revista Española de Micropaleontologia, Madrid, vol. 7, n° 3, p. 447-466.
Youtang & Yunxian (2007).- Fossil Ostracoda of China vol. 2. Cythreacea and Cytherellidae.- China Scientific Books, Beijing, 798 p.
M.C. (1971).- A palaeoecological study of the ostracod Hemicyprideis montosa ( & ) from the Sannoisian of NW Europe. In : H.J. (éd.), Paléoécologie des Ostracodes.- Bulletin du Centre de Recherches Pau - SNPA, supplément au vol. 5, p. 523-543.
M.C. (1972).- The Sannoisian and some other Upper Palaeogene ostracoda from north-west Europe.- Palaeontology, London, vol. 15, part 12, p. 267-325.
M.C. (1977).- Ostracods assemblages and depositional environments of the Headon, Osborne and Bembridge Beds (Uppper Eocene) of the Hampshire Basin.- Palaeontology, London, vol. 20, part 2, p. 405-445.
M.C. (1990).- The ecology and evolution of the Palaeogene ostracod Neocyprideis.- Courier Forschungstintut Senckenberg, Frankfurt am Main, n° 123, p. 217-228.
M.C. (1993).- Ostracoda as palaeoenvironmental indicators: examples from the Tertiary and Early Cretaceous.- In : D.G. (ed.), Applied Micropaleontology.- Kluwer Academic Publishers, Dordrecht, p. 41-67.
M.C., S.S.J., A. & A.M. (1992).- Tertiary ostracods of North Africa and the Middle East. In : M.D. (ed.), Micropalaeontology and Hydrocarbon Exploration in the Middle East.- Chapman & Hall, London, p. 371-401.
M.C. & A. (1991).- Lower Eocene ostracods from the Rusayl Shale Formation of Oman.- Journal of Micropalaeontology, London, vol. 10, n° 2, p. 227-233.
D. (1977).- Ecology and zoogeography of recent brackish-water ostracoda (Crustacea) from south-west Florida. In : H. & . (eds.), Aspects of Ecology and Zoogeography of Recent and Fossil Ostracoda.- Dr W Junk b.v. Publishers, The Hague, p. 207-221.
S.C. & M.L. (2002).- Ostracodes from the intertrappean beds (Early Paleocene) of the east coast India.- Paleontological Research, Tokyo, vol. 6, n° 2, p. 191-210.
H. (1973).- Ostracoden aus dem Sannois und jüngeren Schichten des Mainzer Beckens, 3 : Ehemalige "Cytheridea"-Arten und -Verwandte.- Sonderdruck aus dem Notizblatt des Heissischen Landesamtes füt Bodenfosrchung zu Wiesbaden, Band 101, p. 188-201.
H. (1978).- Neue Hemicyprideis-Arten (Ostracoda; Tertiär) aus dem Mainzer Becken und seinen Rendgebieten.- Senckenbergiana Lethaea, Frankfurt am Main, vol. 59, n° 1-3, p. 71-91.
H. & E. (1970).- Ostracoden aus dem Sannois und jüngeren Schichten des Mainzer Beckens, 2 : Hemicyprideis n.g.- Senckenbergiana Lethaea, Frankfurt am Main, vol. 51, n° 1, p. 1-47.
F. (1993, inédit).- Charophytes du Tertiaire continental de l'Algérie. Systématique et biostratigraphie.- Thèse, Université d'Oran, 173 p.
F. & M. (1999).- Nouvelles charophytes de l'Éocène continental de l'Algérie. Géologie Méditerranéenne, Marseille, vol. 26, n° 1-2, p. 29-45.
F., M., M. & M. (1997).- L'apport des charophytes à la stratigraphie des formations continentales paléogènes de l'Algérie.- Geobios, Villeurbanne, vol. 30, n° 2, p. 171-177.
M. (1996).- Eocene ostracods of Hungary. Systematical part 1 (Cytheracea 1).- Annales de l'Université des Sciences de Budapest, Section Géologie, Budapest, t. 31, p. 27-74.
A. (1993).- Micropaleontological (ostracoda and foraminifera) investigation of Tertiary sequence of Darende Basin.- Geological Bulletin of Turkey, Ankara, vol. 36, p. 13-36 (en turc).
D.D. & J.C. (2008).- Perissocytheridea carrenoae sp. nov. (Crustacea, Ostracoda) and associated calcareous microfauna from Yecua Formation (Miocene), Bolivia).- Revista Brasileira de Paleontologia, Porto Alegre, vol. 11, n° 3, p. 179-186.
E.K., C.T. & G. (2010).- New ostracode species from the Upper Cretaceous of the Santos basin, Brazil.- Revista Brasileira de Paleontologia, Porto Alegre, vol. 3, n° 3, p. 175-180.
J. (1986).- Les charophytes du Paléogène et du Miocène inférieur d'Europe occidentale. Biostratigraphie des formations continentales.- Cahiers de Paléontologie, Paris, n° 22, 227 p.
Q.A. (2000).- Some species of the genus Neocyprideis in the early Tertiary of Pakistan.- Journal of Micropalaeontology, London, vol. 19, n° 1, p. 1-7.
P. (1988).- Palaeobiogeography and palaeoecology of the Tertiary ostracods on northwestern India with paleooceanographic remarks. In : ., . & K. (eds.), Evolutionaly Biology of Ostracoda.- Elsevier/Kodansha, Amsterdam-Tokyo, p. 619-636.
Y. (1972).- Thanétien supérieur et Ilerdien inférieur des Petites Pyrénées, du Plantaurel et des Chaînons audois.- Travaux du Laboratoire de Géologie-Pétrologie de l'Université Paul Sabatier, Toulouse, 376 p.
Y., M., C. & M. (1989).- Caractérisation de l'Ilerdien continental dans le domaine ouest-méditerranéen.- Comptes Rendus de l'Académie des Sciences, Paris, n° 308, p. 689-695.
M., P., J.-P. & A. (1999).- On the Upper Cretaceous age and affinities of Neocyprideis lusitanicus (Ostracoda).- Ciencias da Terra, Lisbonne, vol. 13, p. 87-91.
R. & R.C. (2006). Recent marine ostracoda from the Solomon Islands. Part 2. Cytheroidea, Bythocytheridae, Cytherideidae, Krithidae, Neocytherideidae, Cytheruridae.- Revista Española de Micropaleontologia, Madrid, vol. 38, n° 1, p. 169-189.
M. (1985, inédit).- Causalité des variations morphologiques de la carapace chez les ostracodes : essai d'interprétation sur des populations actuelles et fossiles.- Thèse de Doctorat de 3ème Cycle, Université de Bordeaux I, 285 p.
M.A. & E.A. (1978).- Microfossiles calcáreos no-marinos del Cretacico superior en Zampal, Provincia de Mendoza, Argentina.- Ameghiniana, Buenos Aires, tomo 15, n° 1-2, p. 111-135.
B. (1975).- To the problem of Cyprideis torosa (, 1850). In : F.M., L.S. & R.F. (eds.), Biology and Paleobiology of Ostracoda.- Bulletin of American Paleontologists, New-York, vol. 65, n° 282, p. 205-216.
M.C., E.A.M., A.C. & P. (2000).- Stratigraphy and biogeographic affinities of the late Aptian-Campanian ostracods of the Potiguar and Sergipe basins in northeastern Brazil.- Cretaceous Research, London, vol. 21, n° 2-3, p. 407-455.
K. (2005).- On an extant species of the genus Neocyprideis from Java (Indonesia), with the description of the appendages (Crustacea, Ostracoda).- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Biologie, Bruxelles, n°75, p. 89-95.
Hengren, Xianting & Shupan (1995).- Late Cretaceous-Early Tertiary ostracod fauna from western Tarim Basin, S Xinjiang, China.- Science Press, Beijing, 173 p. (en chinois).
Europe :
Neocyprideis apostolescui (, 1957) : Éocène
moyen et supérieur du Bassin de Paris ( et alii,
1985 ; , 1990).
Neocyprideis colwellensis (, 1857) : Éocène supérieur de Grande-Bretagne (,
1977, 1990).
Neocyprideis durocortoriensis , 1956 : Paléocène supérieur-Éocène inférieur (Sparnacien et Thanétien supérieur) du Bassin de Paris (,
1981 ; et alii,
1985 ; , 1990).
Neocyprideis grandinatus , 1972 : Paléocène supérieur-Éocène inférieur (Thanétien-supérieur / Ilerdien inférieur) du S de la France (,
1972 ; et alii,
1985 ; et alii,
1989).
Neocyprideis moyesi , 1962 : Oligocène supérieur (Chattien) d'Aquitaine, SW France (,
1962).
Neocyprideis parallela (, 1905) : Oligocène d'Allemagne ( & ,
1970).
Neocyprideis revestensis & ,
1992, Oligocène de Haute-Provence, SE de la France ( & ,
1992).
Neocyprideis scapha , 1971 : Oligocène supérieur (Chattien) d'Aquitaine, SW de la France.
Neocyprideis sp. : Oligocène de Haute-Provence, SE de la France ( & ,
1992)
Neocyprideis vacherensis & ,
1992, Oligocène de Haute-Provence, SE de la France ( & ,
1992).
Neocyprideis williamsoniana (, 1852) : Oligocène (Rupélien), de Belgique, du Bassin de Paris et de Grande-Bretagne (,
1972, 1990 ; et alii,
1985). Aussi mentionné par
(1993) dans le Lutétien de Turquie et par
(1996) dans l'Éocène supérieur de Hongrie.
Chine :
Neocyprideis galba (, 1956) : Éocène inférieur du Tarim ( et alii,
1995).
Neocyprideis minuta , 1995 (in et alii,
1995) : Éocène inférieur du Tarim.
Inde :
Neocyprideis bhupendri ( & , 1968) : Éocène inférieur du Rajasthan (,
1988 ; , 1996).
Neocyprideis raoi (, 1978) : Paléocène inférieur de Madya Pradesh (in & ,
2002).
Oman :
Neocyprideis rusaylensis & ,
1991 : Éocène inférieur.
Miocyprideis sp. : Éocène (Bartonien) in et alii (1991); = Neocyprideis selon et alii (1992).
Cyprideis ? sp. : Oligocène in et alii (1991); = Neocyprideis selon et alii (1992).
Pakistan :
Neocyprideis formosa , 2000 : Éocène terminal (,
2000).
Neocyprideis regularis , 2000 : Éocène supérieur.
Neocyprideis simplex , 2000 : Éocène
inférieur.
Neocyprideis sp. A, , 2000 : Paléocène.
Sénégal :
Neocyprideis priani , 1989 : Éocène inférieur (, 1989).
Somalie :
Neocyprideis armata armata & ,
1996 : Éocène moyen.
Neocyprideis armata isoarcuata & ,
1996 : Éocène moyen.
Neocyprideis dacquei & ,
1996 : Éocène moyen.
Neocyprideis rotundata & ,
1996 : Éocčne moyen.
Algérie :
Neocyprideis meguerchiensis nov. sp. : Éocène inférieur.
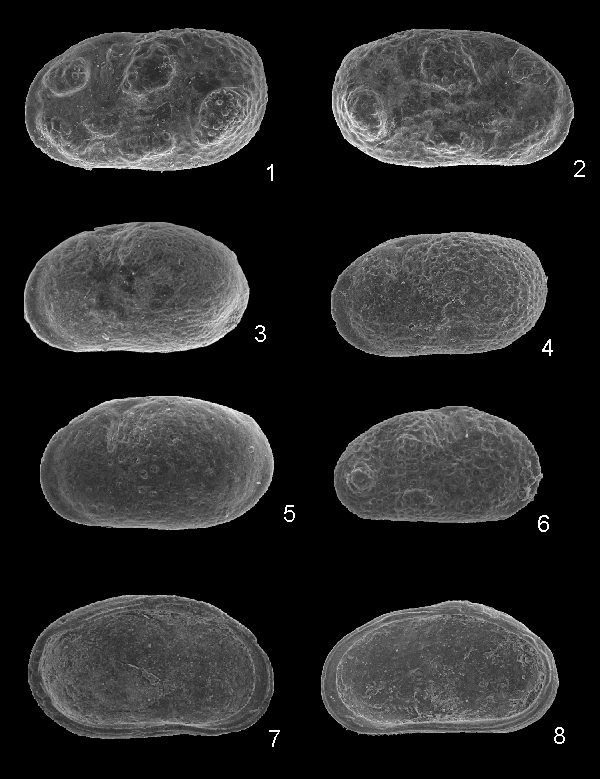
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Neocyprideis meguerchiensis nov. sp.
1. Valve gauche, femelle tuberculée-réticulée, holotype (EJPCH 2011-01) (L= 0,920 mm)
2. Valve droite, femelle, tuberculée-réticulée, paratype (EJPCH 011-01, 01) (L = 0,916 mm)
3. Valve gauche, mâle, lisse, paratype (EJPCP 2011-01, 02) (L = 0,990 mm)
4. Valve gauche, femelle, réticulée (EJPCP 2011-01, 03) (L = 1,09 mm)
5. Valve gauche, mâle, lisse, paratype (EJPCP 2011-01, 04) (L = 1,05 mm)
6. Valve droite, juvénile, tuberculée-réticulée, paratype (EJPCP 2011-01, 05) (L = 0,660 mm)
7. Valve gauche, mâle, vue interne, paratype (EJPCP 2011-01, 06) (L = 1,00 mm)
8. Valve droite, femelle, vue interne, paratype (EJPCP 2011-01, 07) (L = 0,920 mm)
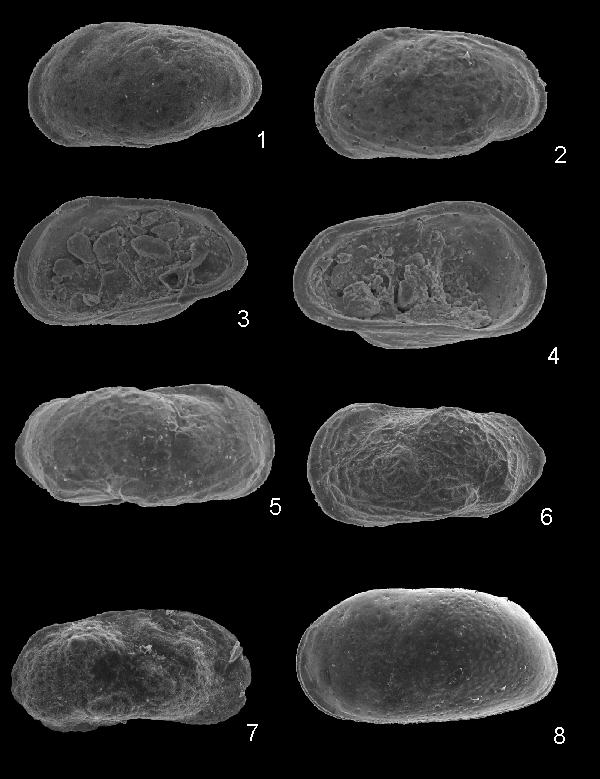
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Perissocytheridea algeriensis nov. sp.
1. Valve gauche, femelle, holotype (EJPCH 2011-02 (L = 0,640mm)
2. Valve gauche, femelle, paratype (EJPCH 2011-02, 08) (L = 0,640 mm)
3. Valve droite, femelle, vue interne, paratype (EJPCH 2011-02, 09 (L = 0,580 mm)
4. Valve gauche, femelle, vue interne, paratype (EJPCP 2011-02, 10 (L = 0,600 mm)
5. Valve droite, mâle, paratype (EJPCP 2011-02, 11) (L = 0,680 mm)
6. Valve gauche, mâle, tuberculée, paratype (EJPCP 2011-02, 12) (L = 0,640 mm)
Limnocythere sp.
7. Valve droite (L = 0,610 mm)
Hemicyprideis sp.
8. Valve gauche (L = 1,250 mm)