![]()
Contents
[1. Introduction] [2. Material]
[3. Systematic part]
[4. Discussion] [Bibliographic references]
Institute of Paleobiology, ul. Twarda 51/55, PL-00-818 Warszawa (Poland)
Manuscript online since December 31, 2011
[Editor: Christian C. ]
![]()
The genus Xenobrochus, with the type species Gryphus africanus , 1973, was erected for short-looped brachiopods of small size, rectimarginate and having a loop with anteriorly convex transverse band. A new species of Xenobrochus, X. norfolkensis sp. nov. has been identified in the material collected during the French cruises SMIB 8, NORFOLK 1 and NORFOLK 2 to the Norfolk Ridge, New Caledonia, SW Pacific. This species differs from those hitherto described in the absence of cardinal process and relatively wide outer hinge plates. The genus, represented now by nine species, has a distribution restricted to the Indian Ocean and West Pacific.
Xenobrochus; Dyscoliidae; Brachiopoda; Norfolk Ridge; South-West Pacific; new species.
M.A. (2011).- Xenobrochus norfolkensis (Brachiopoda: Dyscoliidae), a new species from the Norfolk Ridge, New Caledonia, South-West Pacific.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2011/05 (CG2011_A05), p. 203-211.
Xenobrochus norfolkensis (Brachiopoda: Dyscoliidae), une nouvelle espèce de la Ride de Norfolk, Nouvelle-Calédonie, SW Pacifique.- Le genre Xenobrochus (espèce-type Gryphus africanus , 1973) a été établi pour des brachiopodes de petite taille, rectimarginés et ayant une boucle courte avec une bande transverse à convexité antérieure. Une nouvelle espèce, Xenobrochus norfolkensis sp. nov., a été identifiée au sein du matériel récolté au cours des campagnes françaises SMIB 8, NORFOLK 1 et NORFOLK 2 dans la Ride de Norfolk (Nouvelle-Calédonie, SW Pacifique). Cette espèce diffère des précédentes par l'absence du processus cardinal et par des plaques cardinales relativement larges. Le genre comporte maintenant neuf espèces ; sa distribution géographique est restreinte à l'Océan Indien et à l'Ouest de l'Océan Pacifique.
Xenobrochus ; Dyscoliidae ; Brachiopoda ; Ride de Norfolk ; SW Pacifique ; nouvelle espèce.
The genus Xenobrochus, with the type species Gryphus africanus , 1973, was erected by (1981) for short-looped brachiopods having a small size, rectimarginate anterior commissure, and a loop with anteriorly convex transverse band. So far this genus is represented by eight species (, 2007; , 2008). Particular species differ from each other mostly in the presence or absence of the cardinal process, the size of the outer hinge plates, and the presence of a median fold in the transverse band.
The aim of this paper is to describe a new Xenobrochus species, previously erroneously attributed to X. australis , 1981 (see , 2009). However, examination of the type material of X. australis kept at the Muséum National d'Histoire Naturelle de Paris (MNHN-BRA-78-14a) excluded the present material from that species.
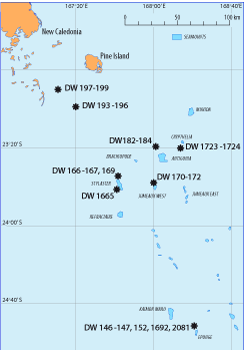
The material examined here was collected during three cruises to the Norfolk Ridge south of New Caledonia, SW Pacific (Fig. 1 ![]() ). The major part of the material (272 specimens) was dredged during the cruise SMIB 8. This cruise was carried out within the research program "Substances Marines d'Intérêt Biologique" (SMIB) organized by the Institut de Recherche pour le Développement (IRD, formerly ORSTOM) and Muséum National d'Histoire
Naturelle de Paris (MNHN). The aim of this program was to collect deepwater invertebrates for experimentation. The cruise SMIB 8 was carried out from 26 January to 3 February 1993 on board RV Alis
( de & ,
1996; et alii,
2008). A minor part of the material was dredged during the cruises NORFOLK 1 and NORFOLK 2 in 2001 and 2003, respectively (see also ,
2009).
). The major part of the material (272 specimens) was dredged during the cruise SMIB 8. This cruise was carried out within the research program "Substances Marines d'Intérêt Biologique" (SMIB) organized by the Institut de Recherche pour le Développement (IRD, formerly ORSTOM) and Muséum National d'Histoire
Naturelle de Paris (MNHN). The aim of this program was to collect deepwater invertebrates for experimentation. The cruise SMIB 8 was carried out from 26 January to 3 February 1993 on board RV Alis
( de & ,
1996; et alii,
2008). A minor part of the material was dredged during the cruises NORFOLK 1 and NORFOLK 2 in 2001 and 2003, respectively (see also ,
2009).
The total number of specimens is 307 (210 articulated specimens and 97 separate valves), all housed in the Muséum National d'Histoire Naturelle de Paris, under the inventory numbers NMHN BRA-3211-3221.
Click on thumbnail to enlarge the image.
Figure 1: Map of the Caledonian part of the Norfolk Ridge with seamounts indicated; stations with brachiopods are indicated by a star. Map from the "programme ZoNéCo", simplified.
| Station | Location | Depth | Specimens |
| SMIB 8 | |||
| Seamount Eponge | |||
| DW 146 | 24°55.20'S, 168°21.73'E | 514-522 m | 89 articulated specimens, 17 ventral valves, 6 dorsal valves |
| DW 147 | 24°54.90'S, 168°21.85'E | 508-532 m | |
| DW 152 | 24°54.35'S, 168°22.23'E | 514-530 m | one articulated specimen |
| Seamount Stylaster | |||
| DW 166 | 23°37.83'S, 167°42.69'E | 433-450 m | 51 articulated specimens, 26 ventral valves, one dorsal valve |
| DW 167 | 23°38.13'S, 167°43.16'E | 430-452 m | 3 articulated specimens, one ventral valve |
| DW 169 | 23°37.73'S, 167°42.57'E | 447-450 m | 6 articulated specimens, one dorsal valve |
| Seamount Jumeaux West | |||
| DW 170 | 23°41.23'S, 168°00.56'E | 241-244 m | 3 articulated specimens |
| DW 171 | 23°40.54'S, 168°00.26'E | 233-250 m | |
| DW 172 | 23°40.50'S, 168°01.50'E | 280-290 m | |
| Seamount Antigonia | |||
| DW 182 | 23°19.28'S, 168°04.82'E | 314-330 m | 6 articulated specimens |
| DW 183 | 23°18.27'S, 168°04.95'E | 330-367 m | |
| DW 184 | 23°18.32'S, 168°04.84'E | 305-320 m | |
| Slope of the Isle of Pines | |||
| DW 193 | 22°58.71'S, 167°20.06'E | 500-508 m | 24 articulated specimens, 29 ventral valves, 7 dorsal valves |
| DW 194 | 22°59.63'S, 167°22.50'E | 491 m | |
| DW 195 | 22°58.93'S, 167°20.23'E | 508-514 m | |
| DW 196 | 22°58.52'S, 167°20.92'E | 530-558 m | |
| DW 197 | 22°51.27'S, 167°12.54'E | 414-436 m | one articulated specimen |
| DW 198 | 22°51.59'S, 167°12.44'E | 414-430 m | |
| DW 199 | 22°51.63'S, 167°12.22'E | 408-410 m | |
| NORFOLK 1 | |||
| Seamount Stylaster | |||
| DW 1665 | 23°43'S, 167°43'E | 923 m | 3 articulated specimens, one dorsal valve |
| Seamount Eponge | |||
| DW 1692 | 24°56'S, 168°21'E, | 507-967 m | 2 articulated specimens |
| Seamount Crypthelia | |||
| DW 1723 | 23°18'S, 168°15'E, | 266-267 m | one articulated specimen, 3 ventral valves |
| DW 1724 | 23°17'S, 168°14'E | 200-291 m | 19 articulated specimens, 4 ventral valves |
| NORFOLK 2 | |||
| Seamount Eponge | |||
| DW 2081 | 25°54.40'S, 168°21.64'E | 500-505 m | one articulated specimen, one dorsal valve |
Table 1: Data and specimens of Xenobrochus per station.
Phylum Brachiopoda , 1805
Subphylum Rhynchonelliformea et alii, 1996
Order Terebratulida , 1883
Suborder Terebratulidina , 1883
Superfamily Dyscolioidea et , 1891
Family Dyscoliidae et , 1891
Subfamily Aenigmathyridinae , 1983
Genus Xenobrochus , 1981
Type species: Gryphus africanus , 1973, by original designation of (1981, p. 19).
Emended diagnosis: Small to medium, oval, ventribiconvex, smooth anterior commissure rectimarginate; beak long, erect, labiate, foramen large to small, submesothyrid to permesothyrid, symphytium visible; pedicle collar short, teeth large; cardinal process variable, absent or broad, semielliptical; hinge plates narrow, poorly defined or absent; loop narrow, rounded, transverse band convex anteriorly.
Remarks: Because of discrepancies between the diagnoses of the genus Xenobrochus by (1981) and by (2006), as well as the description of new material, the diagnosis needs emendation. In the original diagnosis (1981) described the foramen as submesothyrid to permesothyrid. Surprisingly, in the diagnosis given in the Treatise (, 2006, p. 2140) a foramen is described only as permesothyrid, although most species of Xenobrochus have a mesothyrid foramen. Also the size of a foramen is variable, from large to small (see X. rotundus , 2008). Similarly, outer hinge plates can vary from relatively wide (see , 2009: fig. 6 C, D) to rudimentary or absent (see , 2008: fig. 6 F, H). The main controversy concerns a cardinal process. According to the diagnoses (, 1981; , 2006) it should be broad and semielliptical in all species. However, the cardinal process is not mentioned in the description of X. africanus (see , 1973; , 1986; , 1997) and the species X. translucidus has no trace of a cardinal process (, 1920, p. 320).
Xenobrochus norfolkensis sp. nov.
Synonymy: 2009 Xenobrochus australis ; , p. 11, fig. 6A-D.
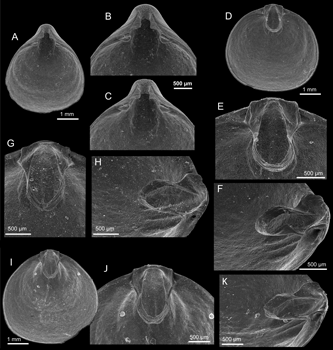
Holotype: The specimen in Fig. 2 E-H ![]() (NMHN BRA-3211).
(NMHN BRA-3211).
Paratypes: the specimens in Fig. 2 C, D, I-L ![]() ; Fig. 3 A-K
; Fig. 3 A-K ![]() (NMHN BRA-3212-3217).
(NMHN BRA-3212-3217).
Type locality: Norfolk Ridge, seamount Eponge, stn DW 146-147, 24°55'E, 168°22'E, 508-532 m.
Etymology: Refers to the type locality, Norfolk Ridge.
Diagnosis: Small Xenobrochus with elongate oval outline, rectimarginate; beak erect, foramen medium, mesothyrid, symphytium visible; no cardinal process, outer hinge plates triangular, loop rounded with a wide, anteriorly convex transverse band.
Material examined: The material examined comprises 271 specimens collected during the cruise SMIB 8, 33 specimens collected during the cruise NORFOLK 1, and 2 specimens collected during the cruise NORFOLK 2. In a few cases the material collected during the cruise SMIB 8 was joined in one sample from two or more neighbouring stations. For details see Table 1. All specimens are dry with many empty shells.
Depth range: 193-967 m.
| Station | Length | Width | Thickness | Figure |
| DW 146-147 (paratype) | 5.5 | 4.4 | 2.9 | Fig. 2 I-L |
| DW 146-147 (holotype) | 5.3 | 4.7 | 3.1 | Fig. 2 E-H |
| DW 152 | 5.0 | 3.9 | 2.7 | - |
| DW 167 | 4.4 | 3.6 | 2.4 | - |
| DW 167 | 3.9 | 3.2 | 2.1 | - |
| DW 167 | 2.5 | 2.1 | 1.2 | - |
Table 2: Measurements (in mm) of Xenobrochus norfolkensis sp. nov.
Description: Shell small (maximum observed length 5.5 mm), thin, elongate oval to rounded pentagonal in outline, biconvex with the ventral valve slightly more convex. Shell surface smooth, ornamented only by growth lines. Beak erect, beak ridges rounded. Foramen of medium size, circular, mesothyrid. Deltidial plates conjunct forming a visible symphytium. Lateral commissures straight, anterior commissure rectimarginate.
Ventral valve interior with pedicle collar relatively wide, excavate anteriorly. Teeth small, hooked-shape with weak swollen bases (Fig. 3 B-C ![]() ). Dorsal valve interior with short but high inner socket ridges and deep dental sockets. No cardinal process; the surface between socket ridges rough. Outer hinge plates triangular, relatively wide. Crural processes short, blunt, directed ventrally. Loop short with a broad, rounded transverse band convex anteriorly; a weak median fold on the transverse band observed in some specimens. Lophophore zygolophous. Muscle scars oval, well defined on both valves.
). Dorsal valve interior with short but high inner socket ridges and deep dental sockets. No cardinal process; the surface between socket ridges rough. Outer hinge plates triangular, relatively wide. Crural processes short, blunt, directed ventrally. Loop short with a broad, rounded transverse band convex anteriorly; a weak median fold on the transverse band observed in some specimens. Lophophore zygolophous. Muscle scars oval, well defined on both valves.
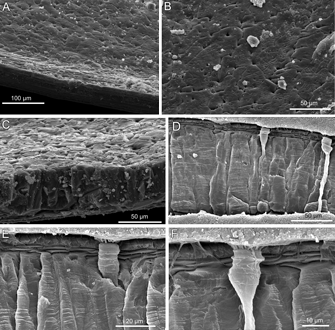
Ultrastructural analysis was carried out on the transverse section as well as on the inner surface of both valves. The shell is composed of three layers (Fig. 4). The microgranular primary and fibrous secondary layers are very thin, 4.5-6 µm and 3.5-4.6 µm, respectively. The fibres are 12.5-26 µm wide and 1.7-2.9 µm thick. The tertiary layer, 125-130 µm thick, consists of large prisms. The prisms observed on the internal surface are interlocking irregularly showing discrete character (Fig. 4 A-C ![]() ). The punctae are wide. The maximum shell thickness in X. norfolkensis is 137 µm.
). The punctae are wide. The maximum shell thickness in X. norfolkensis is 137 µm.

Click on thumbnail to enlarge the image.
Figure 2: Xenobrochus norfolkensis sp. nov., SEM micrographs, cruise SMIB 8, Norfolk Ridge.
A - B: dorsal view of articulated specimen, and enlargement of the posterior part to show details of the beak; st. DW 166, 433-450 m, NMHN BRA-3218.
C - D: interior of ventral valve, and enlargement of posterior part to show small teeth; paratype, st. DW 146-147, NMHN BRA NMHN BRA-3213.
E - H: ventral, dorsal, and lateral views of articulated specimen (G: enlargement of posterior part); holotype, st. DW 146-147, 508-532 m, NMHN BRA-3211.
I - L: ventral, dorsal, and lateral views of articulated specimen (K: enlargement of posterior part); paratype, st. DW 146-147, 508-532 m, NMHN BRA-3212.
Click on thumbnail to enlarge the image.
Figure 3: Xenobrochus norfolkensis sp. nov., SEM micrographs, SMIB 8, Norfolk Ridge.
A - C: interior of ventral valve, enlargement of posterior part (B) and tilted view (C) to show small teeth and weak swollen bases; paratype, st. DW 166, 433-450 m, NMHN BRA-3214.
D - F: interior of dorsal valve, enlargement (E) of posterior part to show details of cardinalia and loop, and side view (F); paratype, st. DW 166, 433-450 m, NMHN BRA-3215.
G - H: interior and lateral views of dorsal valve; paratype, st. DW 146-147, 508-532 m, NMHN BRA-3216.
I - K:, interior of dorsal valve, enlargement (J) of posterior part and side view (K) to show cardinalia and loop; paratype, st. DW 146-147, 508-532 m, NMHN BRA-3217.
Click on thumbnail to enlarge the image.
Figure 4: Xenobrochus norfolkensis sp. nov., SEM micrographs, SMIB 8, Norfolk Ridge.
A: interior of dorsal valve showing prisms of the tertiary layer and fibres of the secondary layer on the margin; st. DW 146-147, 508-532 m, NMHN BRA-3219.
B: inner surface, visible discrete units of prisms interlocking irregularly; ventral valve, st. DW 166, 433-450 m, NMHN BRA-3220.
C: transverse section of the shell, visible prisms of tertiary layer (top); ventral valve, st. DW 166, 433-450 m, NMHN BRA-3220
D: transverse section of the shell showing the relationship between the primary, secondary and tertiary layers, with two punctae; ventral valve, st. DW 166, 433-450 m, NMHN BRA-3221.
E - F: sections showing details of boundaries between the layers and caeca penetrated a shell with a radiating brush; ventral valve, st. DW 166, 433-450 m, NMHN BRA-3221.
Remarks: This species was first investigated with rather limited material (35 specimens) collected during the cruises NORFOLK 1 and NORFOLK 2 (, 2009). The additional material now available for investigation is much richer (272 specimens) and was collected earlier from the same region during the cruise SMIB 8.
Initially, based on the absence of the cardinal process, the material collected during NORFOLK 1 and NORFOLK 2 was assigned by me to Xenobrochus africanus (, 1973), a species already reported from the New Caledonia region (, 1997). Nevertherless, I transferred it to X. australis , 1981, following the suggestions of one of the reviewers (, 2009). However, examination of the type material of this species allows excluding the Norfolk brachiopods from X. australis which is much larger (maximum length 15 mm), has a prominent cardinal process, very long crura and a distinct median angulation on the transverse band. None of those characters is observed in the studied material.
The presence of the cardinal process in such species as Xenobrochus indianensis (, 1973), X. agulhasensis (, 1939), X. anomalus , 1981, X. naudei , 1994, X. rotundus , 2008, as well as in as-yet undescribed material from the New Zealand region, clearly distinguish those species from the material from the Norfolk Ridge (, 1973, 1981, 1983; , 1986, 1994a, 1994b; , 2008). X. indianensis has incurved beak and partly concealed symphytium (, 1973, 1981; , 1997), while in X. norfolkensis sp. nov. the symphytium is wholly visible. Additionally, the pedicle collar in X. anomalus is tubular and extravagantly developed (, 1981; , 1994b). In turn, X. rotundus can be distinguished from the studied specimens by its rounded outline, a small foramen and absent outer hinge plates (, 2008).
Only two species of Xenobrochus, X. africanus (, 1973) and X. translucidus (, 1920) have no cardinal process like X. norfolkensis. In size X. norfolkensis is closest to X. africanus, but they differ in hinge plate width and loop outline; in X. africanus the hinge plates are rudimentary and the loop has a characteristic angular boundary between the descending branches and the transverse band (see , 1973, 1981, 1983; , 1986). In turn, X. translucidus is larger than X. norfolkensis and has a partly concealed symphytium and narrower hinge plates (, 1920; , 1983).
The microstructure of Xenobrochus has here been investigated for the first time. Although the shell of terebratulides is usually built of two layers, many short-looped brachiopods including Xenobrochus have a three-layered shell ( & , 1974; , 2007). The prismatic tertiary layer in X. norfolkensis is very thick when compared with the primary and secondary layers and the fibrous secondary layer is exceptionally thin, being even thinner than the primary layer.
Rare traces of gastropod predation activity are also observed on the investigated specimens. Of 307 specimens 25 (~8%) bear drill holes.
The family Dyscoliidae is characterized by a very short loop with a horizontal or anteriorly convex transverse band. Of 11 members of the family five genera are living today (, 2006). While the genera Dyscolia et , 1890, and Abyssothyris , 1927, also have their representatives in the fossil record, three other genera, Goniobrochus , 1983, Acrobelesia , 1983, and Xenobrochus , 1981, are known only from modern waters. In addition, Dyscolia and Abyssothyris have a worldwide distribution, being recorded from all oceans (, 2007). But Goniobrochus and Acrobelesia are monospecific and restricted to small regions; Goniobrochus to the south-east of Mar del Plata, Argentina (, 1983), while Acrobelesia to the Gulf of Gascogne and the Canary Islands, north-eastern Atlantic (, 1976; , 1983; , 1983; & , 2005).
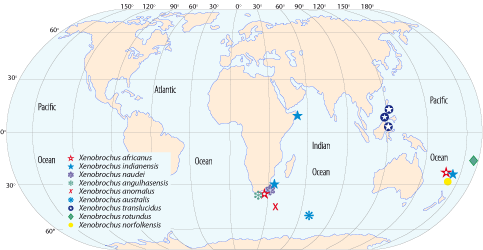
The genus Xenobrochus, including the species described herein, is now represented by nine species and is restricted to the Indian Ocean and West Pacific (Fig. 5 ![]() ) with a wide depth range from 65 to 1272 m (,
2007; , 2008, this paper). Apart from X. africanus and X. indianensis that are recorded from the western Indian Ocean and south-western Pacific (,
1973; , 1997; ,
2010), other Xenobrochus species seem to have restricted distributions: X. agulhasensis and X. naudei in the Indian Ocean off southern Africa (,
1986, 1994a). X. anomalus and X. australis are also found in the Indian Ocean; the former south-east of Marion Island (,
1981; , 1994b), the latter west of Heard Island (,
1981). In the Pacific Ocean three species are recorded. X. translucidus occurs off the Philippines and Borneo (,
1920; & ,
1937; ,
1983) and X. rotundus in the region of Fiji and Wallis and Futuna Islands (,
2008), while X. norfolkensis can be regarded as endemic to the Norfolk Ridge.
) with a wide depth range from 65 to 1272 m (,
2007; , 2008, this paper). Apart from X. africanus and X. indianensis that are recorded from the western Indian Ocean and south-western Pacific (,
1973; , 1997; ,
2010), other Xenobrochus species seem to have restricted distributions: X. agulhasensis and X. naudei in the Indian Ocean off southern Africa (,
1986, 1994a). X. anomalus and X. australis are also found in the Indian Ocean; the former south-east of Marion Island (,
1981; , 1994b), the latter west of Heard Island (,
1981). In the Pacific Ocean three species are recorded. X. translucidus occurs off the Philippines and Borneo (,
1920; & ,
1937; ,
1983) and X. rotundus in the region of Fiji and Wallis and Futuna Islands (,
2008), while X. norfolkensis can be regarded as endemic to the Norfolk Ridge.
Click on thumbnail to enlarge the image.
Figure 5: Geographical distribution of the species of Xenobrochus. Data from (2007), (2008) and this paper.
I am very grateful to Drs B. de (IRD, Nouméa), B.L. (University of Glasgow, Glasgow) and A. (Institute of Paleobiology, Warszawa) for the opportunity to study the material collected by them. Dr P. (MNHN, Paris) made possible to examine the type material of X. australis housed at the Muséum National d'Histoire Naturelle, Paris. Drs D.E. and J.H. (both University of Otago, Dunedin) are thanked for the opportunity to study the brachiopod collections housed at the University of Otago, New Zealand. The visit at the University of Otago was partially financed by the grant No. N N307 129837 from the Ministry of Science and Higher Education (Poland). I am grateful to B.L. for reading the manuscript, making useful comments and improving English. Thanks go also to Prof. F. (University of Oviedo, Oviedo) and Dr C.C. (BrachNet, Marseille) for critically reviewing the manuscript and offering suggestions for its improvement. The SEM micrographs were taken in the SEM laboratory of the Institute of Paleobiology (Warszawa) using a Philips XL-20 scanning microscope.
F. & C.C. (2005).- Brachiopoda. In: F., C.C., C. & J.M. (eds.), Lophophorata, Phoronida, Brachiopoda.- Fauna Ibérica, Madrid, vol. 27, p. 57-177.
M.A. (2007).- Recent brachiopods from the Austral Islands, French Polynesia, South-Central Pacific.- Zoosystema, Paris, vol. 29, n° 3, p. 491-502.
M.A. (2008).- New data on the Recent brachiopods from the Fiji and Wallis and Futuna islands, South-West Pacific.- Zoosystema, Paris, vol. 30, n° 2, p. 419–461.
M.A. (2009).- Recent Brachiopoda from the Norfolk Ridge, New Caledonia, with description of four new species.- Zootaxa, Auckland, vol. 2235, p. 1–39.
M.A. (2010).- Biodiversity of shallow-water brachiopods from New Caledonia, SW Pacific, with description of a new species.- Scientia Marina, Barcelona, vol. 73, p. 643-657.
P., V., P. & P. (2008).- A quater-century of deep-sea malacological exploration in the South and West Pacific: Where do we stand? How far to go? In: V., R.H. & P. (eds.), Tropical Deep-Sea Benthos 25.- Mémoires du Muséum National d'Histoire Naturelle, Paris, vol. 196, p. 9-40.
G.A. (1973).- New Brachiopoda from the Indian Ocean.- Smithsonian Contributions to Paleobiology, Washington, vol. 16, 43 p.
G.A. (1981).- Brachiopoda from the Southern Indian Ocean (Recent).- Smithsonian Contributions to Paleobiology, Washington, vol. 43, 93 p.
G.A. (1983).- The Terebratulacea (Brachiopoda), Triassic to Recent: a study of the brachidia (loops).- Smithsonian Contributions to Paleobiology, Washington, vol. 50, 445 p.
J.-L. (1976).- Sur quelques Brachiopodes actuels (Océan Atlantique, Méditerranée, Kerguelen).- Bulletin du Muséum National d'Histoire Naturelle de Paris, (Série 3, Zoologie, n° 350), vol. 243, p. 1-13.
W.H. (1920).- Annoted list of the Recent Brachiopoda in the collection of the United States National Museum, with description of thirty-three new forms.- Proceedings of the United States National Museum, Washington, vol. 57, p. 261-377.
P. & D.P. (1890).- Diagnoses de nouveaux Brachiopodes.- Journal de Conchyliologie, Paris, vol. 38, p. 70-74.
J.-G. (1939).- Die Brachiopodem des zoologischen Museums zu Berlin.- Sitzungsberichte der Gesellschaft naturforschender Freunde zu Berlin, Jarhgang 1938, p. 221-268.
N. (1986).- The South African Museum's Meiring Naude cruises. Part 16. Brachiopoda from the 1975-1979 cruises.- Annals of the South African Museum, Kaapstad, vol. 97, p. 97-140.
N. (1994a).- The environment, biogeography, and origin of the southern African Recent brachiopod fauna.- Journal of Paleontology, Lawrence, vol. 68, p. 776-786.
N. (1994b).- The biogeographic relationships of the brachiopod fauna from Marion and Prince Edward Islands.- South African Journal of Antarctic Research, Pretoria, vol. 24, p. 67-74.
J.W. & G.. (1937).- The Brachiopoda of the Siboga Expedition.- Siboga-Expeditie, Leiden, vol. 27, p. 1-20.
B. (1997).- Brachiopodes récoltés dans les eaux de la Nouvelle-Calédonie et des îles Loyauté, Matthew et Chesterfield. In:A. (éd.), Résultats des campagnes MUSORSTOM, vol. 18.- Mémoires du Muséum National d'Histoire Naturelle, Paris, vol. 176, p. 411-471.
D.E. (2006).- Dyscolioidea.- In: R.L. (ed.), Treatise on Invertebrate Paleontology. Part H, Revised, Brachiopoda.- Geological Society of America and University of Kansas, Boulder, Colorado, and Lawrence, Kansas, vol. 5, p. 2136-2144.
A. (1983).- Brachiopoda collected by CANCAP I-III expeditions to the south-east North Atlantic, 1976-1978. Zoologische Mededelingen, Leiden, vol. 57, p. 165-189.
A. (2007).- Geographic distribution of extant articulated brachiopods. In: P.A. (ed.), Treatise on Invertebrate Paleontology. Part H, Revised, Brachiopoda.- Geological Society of America and University of Kansas, Boulder, Colorado, and Lawrence, Kansas, vol. 6, p. 3082-3115.
D.I. & A. (1974).- Shell structure of terebratulid brachiopods.- Palaeontology, London, vol. 17, p. 179-202.
de B. & C. (1996).- Les campagnes d'échantillonnage du benthos bathyal en Nouvelle-Calédonie, en 1993 et 1994 (BATHUS 1 à 4, SMIB 8 et HALIPRO 1). In: A. (éd.), Résultats des Campagnes MUSORSTOM, vol. 15.- Mémoires du Muséum National d'Histoire Naturelle, Paris, vol. 168, p. 33-53.
J.A. (1927).- Brachiopod morphology and genera (Recent and Tertiary).- New Zealand Board of Science and Art, Manual, Wellington, n° 7, 338 p.
|
Note éditoriale : La disposition 6 de l'article 8 du Code
International de Nomenclature Zoologique (4ème édition, 1999, en
ligne) intitulé "Travaux produits après 1999 par un procédé
sans support papier" stipule que "Un travail produit après 1999
par un procédé sans support papier doit contenir, pour être accepté
comme publié au regard du Code, une déclaration selon laquelle des
copies (sous la même forme que le travail publié) ont été déposées
dans au moins cinq grandes bibliothèques publiques ; ces bibliothèques
doivent être énumérées dans le travail lui-même." Afin de
répondre positivement à cette requête, nous, éditeurs des
"Carnets de Géologie", rappelons que l'édition dévédérom de
notre revue (qui restitue les publications dans leur version originale :
PDF, et accessoirement HTML) est disponible : - en France à la Bibliothèque Nationale de France (Paris), au CADIST - Bibliothèque Interuniversitaire Jussieu (Paris) et au SCD Université de la Méditerranée (Aix-Marseille II), - en Belgique à la Bibliothèque de l'Université Libre de Bruxelles (Bruxelles), - en Allemagne à la UB Regensburg (Regensburg), - en Argentine à la 'Biblioteca' de l'Instituto Superior de Correlación Geologica - INSUGEO (San Miguel de Tucumán), - au Royaume-Uni au Natural History Museum of London (Londres), - aux É.U.A. à la Amherst College Library (Amherst - MA), aux Auburn University Libraries (Auburn - AL) et à la Indiana University Geology Library (Bloomington - IN). Il ne s'agit pas là d'une liste exhaustive puisque de par le monde ce sont plus de 100 bibliothèques de recherche qui reçoivent gratuitement une copie de notre édition dévédérom. De plus pour garantir une permanence de l'archivage, ces publications sont déposées dans des sites institutionnels (CG chez HAL - hyper articles en ligne -, I-Revues de l'INIST, etc.). |