![]()
Contents
[1. Introduction] [2. Outline stratigraphy]
[3. Systematic description]
[4. Ammonite biostratigraphy
...] [5. Current definition and biozonation ...]
[6. Biozonation in study area]
[7. Conclusions] [Bibliographic references]
Geology Department, Faculty of Sciences, Birjand University, PO Box 79, Birjand (Iran)
Manuscript online since December 31, 2011
[Editors: Bruno & Christian C. ]
![]()
The Aralar Mountains located in northern Spain exposes a 983-m-thick succession of sediments of Early Aptian age. The lithological succession evolves from lutites, marls, and calcarenites of the Errenaga Formation to rudist micritic limestones of the Sarastarri Formation, and finally marls, lutites, and sandstones of the Lareo Formation. Based on ammonite assemblage faunas, the Deshayesites oglanlensis, D. weissi, D. deshayesi, and Dufrenoyia furcata biozones have been identified. A transition between the deshayesi and furcata zones with the co-occurrence of the ammonite genera Deshayesites and Dufrenoyia is described in the Aralar succession and is currently unique. The ammonites are described here and correlations are made with other Tethyan regions.
Ammonite; Aptian; biozonation; Basque Country; Spain; Tethys.
S.N. (2011).- The Early Aptian (Early Cretaceous) ammonites from the Aralar Mountains, Basque-Cantabrian Basin, Northern Spain.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2011/02 (CG2011_M02), p. 163-201.
Les ammonites de l'Aptien inférieur (Crétacé inférieur) des Monts Aralar, Bassin basco-cantabrique, Espagne septentrionale.- Le massif d'Aralar, situé dans le nord de l'Espagne, montre sur 983 m d'épaisseur une succession de sédiments d'âge Aptien inférieur. La succession lithologique débute par les lutites, puis les marnes et les calcarénites de la Formation Errenaga, auxquelles succèdent les calcaires micritiques à rudistes de la Formation Sarastarri, et enfin les marnes, puis les lutites et les grès de la Formation Lareo. Basées sur des assemblages d'ammonites, les biozones à Deshayesites oglanlensis, D. weissi, D. deshayesi et Dufrenoyia furcata ont été identifiées. La transition entre la Zone à deshayesi et celle à furcata observée ici est un cas unique dans l'état actuel des connaissances, notamment en raison de la cooccurrence d'ammonites des genres Deshayesites et Dufrenoyia. Ces ammonites sont décrites et les corrélations avec d'autres régions du domaine téthysien sont présentées.
Ammonite ; Aptien ; biozonation ; Pays Basque ; Espagne ; Téthys.
The Basque-Cantabrian Basin is located in north east Spain (Fig. 1 ![]() ). It is a peri-cratonic rift basin related to the opening of the Bay of Biscay ( et alii,
1974). The opening was dated in the
Early Aptian ( et alii, 1979; et alii,
2002), and more recently in the
Early Barremian ( et alii, 2004). Four main subsidence pulses have been detected in the Basque-Cantabrian region during the Aptian and Albian ( et alii,
2005). The very thick succession of sediments in the Aralar Mountains span in age from the Barremian-Aptian boundary to the top of the Early Aptian and has provided an important ammonite record which underpins the geochemical results and interpretations of OAE-1a obtained in this succession ( et alii,
2009).
). It is a peri-cratonic rift basin related to the opening of the Bay of Biscay ( et alii,
1974). The opening was dated in the
Early Aptian ( et alii, 1979; et alii,
2002), and more recently in the
Early Barremian ( et alii, 2004). Four main subsidence pulses have been detected in the Basque-Cantabrian region during the Aptian and Albian ( et alii,
2005). The very thick succession of sediments in the Aralar Mountains span in age from the Barremian-Aptian boundary to the top of the Early Aptian and has provided an important ammonite record which underpins the geochemical results and interpretations of OAE-1a obtained in this succession ( et alii,
2009).
The Aptian succession in the Aralar Mountains (Fig. 1 ![]() ) has been described most recently by et alii
(2009). In this region, the succession consists of the predominantly
siliciclastic sedimentation of the Errenaga and Lareo formations with a major intercalated limestone, the Sarastarri Limestone rich in rudists, orbitolinids and corals ( & ,
1997; et alii,
2005, 2007; et alii,
2009)
(Fig. 2
) has been described most recently by et alii
(2009). In this region, the succession consists of the predominantly
siliciclastic sedimentation of the Errenaga and Lareo formations with a major intercalated limestone, the Sarastarri Limestone rich in rudists, orbitolinids and corals ( & ,
1997; et alii,
2005, 2007; et alii,
2009)
(Fig. 2 ![]() ).
).
Click on thumbnail to enlarge the image.
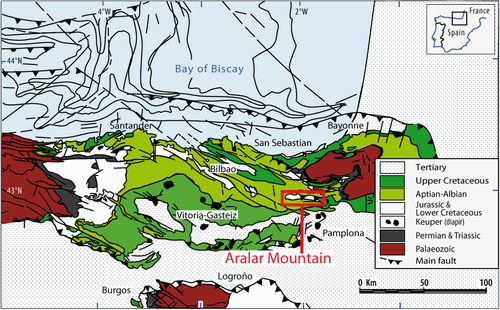
Figure 1: Location of the Aralar Mountain in the western part of the Basque-Cantabrian Basin in the northern margin of Iberia and south of the Bay of Biscay (modified after et alii, 2009).
The outcrop of these sediments extends from Iribas in the east to Ataun in the west
(Fig. 2 ![]() ) and they have been described in detail by et alii
(2009). Since these two formations were reported to contain ammonites in previous studies, a careful search was made by the team led by Professor of the Universidad del Pais Vasco, Bilbao, Spain, which resulted in the gathering of a large amount of new material. Although most of the specimens are crushed, they have permitted the biozonation of these
formations based on the ammonite occurrences. The present paper, therefore, presents a systematic description of these Early Aptian ammonite faunas together with their biozonation.
) and they have been described in detail by et alii
(2009). Since these two formations were reported to contain ammonites in previous studies, a careful search was made by the team led by Professor of the Universidad del Pais Vasco, Bilbao, Spain, which resulted in the gathering of a large amount of new material. Although most of the specimens are crushed, they have permitted the biozonation of these
formations based on the ammonite occurrences. The present paper, therefore, presents a systematic description of these Early Aptian ammonite faunas together with their biozonation.
Apart from some records of the presence of Deshayesites deshayesi in localized areas (e.g., Aralar: et alii, 1972; & , 1975), the only important description of Early Aptian ammonites was made in the Cuchía (Santander) region in the northwestern part of the basin ( et alii, 1979). These authors identified several species of Prodeshayesites, Deshayesites, Cheloniceras and Vectisites corresponding to the four zones of the Early Aptian in England ( et alii, 1998) (Table 1).
Click on thumbnail to enlarge the image.
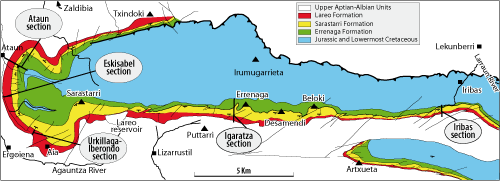
Figure 2: Geological map of the Aralar Mountain showing the Errenaga, Sarastarri and Lareo formations, and sections of Iribas, Igaratza, Urkillaga-Iberondo, Eskisabel and Ataun (modified after et alii, 2009).
(1982) in Lérida, (2007) and et alii (2009) in Barcelona, and et alii (1997) in the Betic Cordillera describe Early Aptian Iberian ammonite occurrences outside of the present area. Although ammonite occurrences are relatively few in the Aralar succession and restricted to certain horizons, they are sufficient to permit a more precise ammonite biozonation to be made of the Early Aptian in this region than has been possible hitherto.
| Age | Substage | , 1961c; , 1991; et alii, 1998 (SE England) |
, 1973; ,
1983 (North Germany / England) |
,
1984; , 1997; et alii, 1999, 2000 (France) |
et alii,
2003; et alii, 2009 (Mediterranean Region) |
et alii,
2000 (Caucasus) |
|
| Aptian | Late | Cheloniceras (Epicheloniceras) martinioides |
Epicheloniceras tschernyschewi | Middle | C. (Epicheloniceras) martinioides | C. (Epicheloniceras) subnodosocostatum/ C. (Epicheloniceras) martini |
C. (Epicheloniceras) subnodosocostatum |
| Tropaeum drewi | |||||||
| Early | Tropaeum bowerbanki | Tropaeum bowerbanki | Early | T. bowerbanki / D. furcata | Dufrenoyia furcata | Dufrenoyia furcata | |
| Deshayesites deshayesi | Deshayesites deshayesi | Deshayesites deshayesi | Deshayesites deshayesi | Deshayesites deshayesi | |||
| Deshayesites forbesi | Deshayesites weissi | Deshayesites weissi | |||||
| Prodeshayesites fissicostatus | Prodeshayesites fissicostatus | Deshayesites tuarkyricus | Deshayesites oglanlensis | D. weissi-procheloniceras albrechti-austria |
|||
| Barrem. | Late | Parancyloceras bidentatum & Parancyloceras scalare |
Pesudocrioceras waagenoides | Pesudocrioceras waagenoides | Colchidites securiformis | ||
| Martelites sarasini | Martelites sarasini | ||||||
Table 1: Correlation chart of the Early Aptian biozones in the Tethyan and Boreal Realm.
The top of the non-marine Wealden series, which rests upon an
Upper Jurassic succession, is overlain by marls with ammonites including Valdedorsella sp. of latest Barremian age. These sediments in turn underlie an ammonite and orbitolinid-rich marine Early Aptian succession. The detailed stratigraphical succession is described by et alii
(2009) and is shown in
Figs. 3 ![]() - 4
- 4 ![]() .
.
a. Errenaga Formation
The Errenaga Formation comprises shales, marls, siltstones, sandstones and marly limestones. There are significant thickness variations in an E-W transect, from Iribas (69 m) to Igaratza (200 m) and Ataun (482 m)
(Fig. 3 ![]() ). There is no lateral continuity of exposure; thus correlation of the three sections has been based on the lithologic similarity of subunits, controlled sedimentological changes, vertical arrangement of facies associations, TOC curve trends of selected intervals and ammonite occurrences et alii,
2009).
). There is no lateral continuity of exposure; thus correlation of the three sections has been based on the lithologic similarity of subunits, controlled sedimentological changes, vertical arrangement of facies associations, TOC curve trends of selected intervals and ammonite occurrences et alii,
2009).
Various fossiliferous horizons containing ammonites of the Deshayesites oglanlensis, Deshayesites weissi, Deshayesites deshayesi zones and Deshayesites deshayesi - Dufrenoyia furcata transition zones of the Early Aptian have been identified and correlated
(Fig. 4 ![]() ).
).
b. Sarastarri Formation
The Sarastarri Formation consists of intercalation of limestones, marls, and marly limestones that extend at least 21.5 km from east (Iribas section) to west (Ataun section) and thicken progressively westward, from Iribas (53 m), to Igaratza (150 m) and Ataun (178 m)
(Fig. 3 ![]() ). The limestones stand as a prominent mappable feature in the field and thus provides an important marker in the determination of the tectonic structures of the Sierra de Aralar. Although no ammonites have been found in this Formation, it is dated as Early Aptian deshayesi-furcata zones transition on account of the faunas in the uppermost Errenaga Formation below and the Lareo Formation above. The Sarastarri limestones indicate a stepped drowning in two different phases in the Deshayesites deshayesi-Dufrenoyia furcata transition Zone et alii,
2007). In the lower part of the immediately overlying Lareo Formation at Ataun, there is a specimen of Dufrenoyia sp. already of basal furcata Zone age.
). The limestones stand as a prominent mappable feature in the field and thus provides an important marker in the determination of the tectonic structures of the Sierra de Aralar. Although no ammonites have been found in this Formation, it is dated as Early Aptian deshayesi-furcata zones transition on account of the faunas in the uppermost Errenaga Formation below and the Lareo Formation above. The Sarastarri limestones indicate a stepped drowning in two different phases in the Deshayesites deshayesi-Dufrenoyia furcata transition Zone et alii,
2007). In the lower part of the immediately overlying Lareo Formation at Ataun, there is a specimen of Dufrenoyia sp. already of basal furcata Zone age.
Click on thumbnail to enlarge the image.
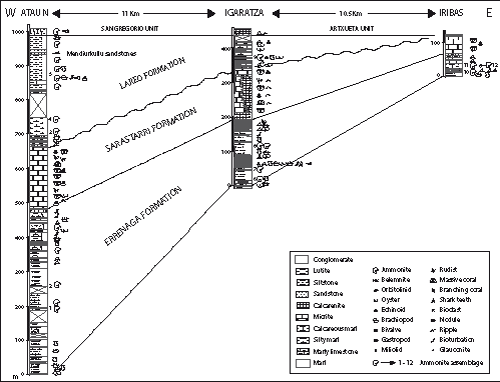
Figure 3: Lithostratigraphic correlation chart of the Lower Aptian sediments of the Aralar, sections of Iribas, Igaratza and Ataun, with indication of the units and ammonite assemblages described in the text.
At Igaratza, assemblage 9 in the basal Lareo Formation also contains Deshayesites, and specimen LEIZ-1 is a Dufrenoyia cf. lurensis . Thus, the Sarastarri Limestone marks a pause in terrigenous sedimentation in the deshayesi-furcata zones transitional interval. At Iribas, the Sarastarri Limestone is overlain directly by the Late Aptian San Gregorio-Artxueta Units
(Fig. 3 ![]() ). The Formation terminates in an areally extensive 20 m thick micritic limestone unit containing corals and rudists (Gorrin-txabola). It is capped by an erosional surface overlain by the Lareo Formation.
). The Formation terminates in an areally extensive 20 m thick micritic limestone unit containing corals and rudists (Gorrin-txabola). It is capped by an erosional surface overlain by the Lareo Formation.
c. Lareo Formation
The Lareo Formation is dated with ammonites initially as still within the Deshayesites deshayesi-Dufrenoyia furcata transitional Zone and subsequently in the D. furcata Zone. This unit shows a variety of facies and it wedges out from west to east ( et alii,
2009). The Lareo Formation is overlain by the
Upper Aptian San Gregorio Unit which cuts down to the Sarastarri Limestone at Iribas
(Fig. 3 ![]() ).
).
d. Upper Aptian formations
The units overlying the Early Aptian succession comprise the
Upper Aptian San Gregorio Formation in the west, made up of marly limestones and marls, and the Artxueta Unit in the east, made up of shallow-water Urgonian limestones ( & ,
1975; ,
1997)
(Fig. 3 ![]() ). They make up a broad carbonate inner ramp (Artxueta) grading downdip to the correspondent outer ramp (San Gregorio).
). They make up a broad carbonate inner ramp (Artxueta) grading downdip to the correspondent outer ramp (San Gregorio).
Click on thumbnail to enlarge the image.
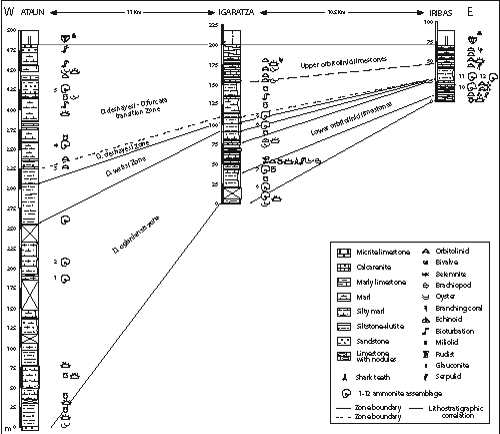
Figure 4: Stratigraphic correlation of the Errenaga Formation from Iribas to Igaratza and Ataun sections. Note the east to west thickening of the unit.
The present study stems from the systematic sampling of the Lower Aptian succession in the Aralar region. Every genus and species which have been found are described briefly, and their occurrence and distribution in the stratigraphical column of the study area is discussed where appropriate. There are some genera or species new to the basin. The classification follows that given in the Treatise on Invertebrate Palaeontology, part L ( et alii, 1996).
Shell measurements (in millimeters) are given in the following order: Diameter (D), whorl height (WH), ratio of whorl height to diameter (WH/D), whorl thickness (WT), ratio of whorl thickness to diameter (WT/D), umbilical width (U), ratio of umbilical width to diameter (U/D), primary ribs in the umbilical area (PR) and all ribs (primary and secondary) on the ventral side (SR). Because some specimens are incomplete and not all dimensions can be measured, the ratio of whorl thickness to whorl height (WT/WH) is also given.
The following abbreviations are given for localities. ATAN= Ataneta locality, IRIBAS section (Errenaga Formation), AIZMUSU= Aizmusu locality, 2km east of the URKILLAGA-IBERONDO section, Ataun area (Lareo Formation), ER= IGARatZA section (Errenaga Formation). (material collected loose from surface debris], ERG= IGARatZA section (Errenaga Formation). (fauna collected in situ), IRI= IRIBAS section (Errenaga Formation), IMA= ATAUN section (Errenaga Formation), NAZ= Nazca Maizagi locality, 2km east of the URKILLAGA-IBERONDO section and 0.5km north of the Aizmusu locality, Ataun area (Lareo Formation), SANGRE= ESKISABEL section, Ataun area (Lareo Formation), URKI= URKILLAGA-IBERONDO section, Ataun area (Lareo Formation), URG = Urgoxo locality, IRIBAS section (Errenaga Formation).
The palaeontological material is housed in the Departamento de Estratigrafía y Paleontología, Universidad del País Vasco, Bilbao (UPV/EHU) and the assemblage and specimen numbers apply to that institution.
Suborder Ammonitina ,
1889
Superfamily Haplocerataceae , 1884
Family Oppeliidae , 1890
Subfamily Aconeceratinae , 1923
Genus Aconeceras , 1903
Type species: Ammonites nisus d', 1841
Generic characters: Involute oxycone with flattened or gently convex sides that narrow above to a hollow microscopically serrated carina or keel. Ventro-lateral shoulders are distinct or broadly rounded; umbilicus with angular rim and low steep wall; flanks almost smooth or bearing sickle-shaped, forwardly inclined striae or faint flattened ribs. Suture line with narrow trifid lateral lobes and tall lateral saddle and many secondary elements declining in regular series to the umbilicus.
Discussion: The separation of some genera of the Aconeceratinae is based on the presence of ribs and denticulation of the keel. However, it seems that this division is not useful in practice ( et alii, 1996). Thus Aconeceras, Sanmartinoceras, Sinzovia, Theganoceras and Gyaloceras are grouped as subgenera of Aconeceras. The subgenus Aconeceras s.s. is known principally by immature (incomplete) shells or nuclei that lack the mouth-borders. Therefore, it is not clear whether lappets and rostrum comparable to those of the subgenus Sanmartinoceras occur, of if the edge of its peristome follows the growth-lines as in Falciferella.
Occurrence: The genus is recorded from Europe, north east Greenland, Algeria, South Africa, Madagascar, Australia, Argentina, Nepal and Antarctica (subgenus Theganoceras) ( et alii, 1996) and Iran (, 2002).
Subgenus Aconeceras (Aconeceras) ,
1903
Aconeceras cf. nisoides (, 1893)
cf. 1893 Oppelia nisoides (), p. 155, pls. iv-v, fig. 10a-c; text-figs. 3, 5.
cf. 1924 Oppelia nisoides (): , p. 311, pl. 26, figs. 4a-b; text-fig. B9.
cf. 1958 Sinzova nisoides (): , p.
129-130.
cf. 1961a Aconeceras nisoides (): , p. 125-128, pl. 26, fig. 3-5; text-fig. 41a-c.
cf. 1995 Aconeceras nisoides (): , taf. 8, fig. 4.
Lectotype: (1893, pls. iv-v, fig. 10a-c) from the Lower Aptian of the Haute-Marne (collection École des Mines de Paris).
Material: NAZ-6, 7; URKI-1, 2, 3, 4, 5, 6, 8, 9, 10, URKI-C-1
Description: Involute oxycone, whorls high and compressed, sub-rectangular, nearly convex sides, ventral margin with a low keel in middle part, test nearly smooth with trace of fine ribs.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| NAZ-7 | 21 | 11.5 | 0.54 | 5 | 0.23 | 0 | 0.43 | |
| URKI-1 | 15 | 7 | 0.46 | 3 | 0.2 | 3 | 0.2 | 0.43 |
| URKI-2 | 12.5 | 7 | 0.56 | 3 | 0.24 | 2.1 | 0.17 | 0.43 |
| Lectotype | 27 | 13.7 | 0.51 | 3.78 | 0.14 | 5.94 | 0.22 | 0.27 |
Discussion: Suture line and rib pattern are diagnostic for the species. A. nisoides differs from A. nisus by possessing a wider umbilicus and more feeble ornamentation on whorl flanks.
Occurrence: England (, 1924; , 1961a), France (, 1893; , 1938), Germany (, 1907) and Russia (, 1958). (1961a, p. 126) also mentioned that the species is reported from Sweden and Zululand.
Distribution: Lower part Nazca Maizagi locality, 2km east of the Urkillaga-Iberondo section and
lower part of Urkillaga-Iberondo section, Lareo Formation (Fig.
6 ![]() ).
).
Aconeceras haugi , 1893
1893 Oppelia haugi , p. 156, pl. 4-6, fig. 11a-c.
1961a Aconeceras cf. haugi (): , p. 128, text-fig. 40g-h.
1973 Aconeceras haugi (): , p. 42, pl. 2, fig. 4.
1982 Aconeceras haugi (): , p. 21-22, pl. 1, fig. 14a-b.
1995 Aconeceras haugi (): , pl. 2, fig. 4.
Type by monotypy, , 1893, p. 145, pls. 4-6, fig. 11a-c from the Aptian of St. Dizier, France (collection École des Mines de Paris).
Material: AIMUSU-2, 3; URKI-11
Description: High whorled, involute, oxycones, whorl section sub-fastigiate, ventro-lateral shoulder rounded but distinct, sides parallel, flattened, keel distinct.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| AIZMUSU-2 | 6 | 3 | 2.8 | 0.5 | ||||
| AIZMUSU-3 | 12 | 5.5 | 0.458 | 3 | 0.25 | 3 | 0.25 | 0.545 |
Discussion: A. haugi differs from A. nisus by possessing ventro-lateral shoulders. According to (1982), Sanmartinoceras groenlandicum (see , 1961b, p. 131, text-fig. 42) is distinguished from Aconeceras haugi by more pronounced costae, a high-serrated keel and a spiral depression.
Occurrence: France, Venezuela (, 1982) and England (, 1965).
Distribution: In Aizmusu locality, 2km east of the Urkillaga-Iberondo section, Ataun area, Lareo Formation and Urkillaga-Iberondo section, Ataun area, Lareo Formation
(Fig.
6 ![]() ).
).
Subgenus Aconeceras (Theganoceras) ,
1926
A. (Theganoceras) sp.
Material: IMA-7, 9
Description: Two crushed specimens are to hand. Ribs fine, falcate, smooth around the umbilicus and more distinct near the venter, Venter with a low keel.
Discussion: (1926) recognised the genus Theganoceras as a distinct genus, but (1961b) regarded it as subgenus of Sanmartinoceras as does et alii (1996). Theganoceras is separated from Sanmartinoceras by its denser and finer ribs.
Occurrence: England, Germany, South Africa (Zululand), and Antarctica et alii, 1996).
Distribution: In Ataun section, Errenaga Formation (ammonite assemblage 1,
Figs. 3 ![]() - 4
- 4 ![]() ).
).
Superfamily Desmocerataceae ,
1895
Family Desmoceratidae , 1895
Subfamily Pseudosaynellinae , 1961a
Genus Pseudosaynella , 1923
Pseudosaynella sp.
Material: ERG-5-4, ER-22
Description: Discoidal, involute, with flattened sides and sharpened venter. Umbilicus narrow with distinct rim, shell smooth or with nearly simple sigmoidal rib pattern.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ER-22 | 28 | 15 | 0.53 | 3 | 0.11 | 4.5 | 0.16 | 0.2 |
Discussion: Rib pattern is the character that could be used for diagnosis. The costation is very like Cleoniceras & , 1895. However the later genus belongs to an Albian assemblage fauna, while Pseudosaynella is an Aptian form. Because of compression the specimens look like Aconeceras, but their rib pattern is distinct. Specimens can be compared with P. cf. raresulcata (d', 1841) (, 1961a, p. 171).
Distribution: In Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Suborder Ancyloceratina , 1966
Superfamily Ancylocerataceae , 1871
Family Ancyloceratidae , 1871
Subfamily Helicancylinae , 1894
Toxoceratoides ? sp.
Material: SANGRE-22, 31
Description: Specimens are fragments of crushed shafts, more or less curved; shaft with spherical to sub-rectangular whorl section; ribs straight and single, one row of tubercles on each side of venter.
Discussion: Curved shaft and two rows of tubercles are characteristic for identification. Tuberculation is not clear in the specimen. The specimens are not complete, and show similarity to Tonohamites in its curved shaft.
Distribution: San Georgio section, Lareo Formation (Fig.
6 ![]() ). &
(2002) described the genus from
Late Aptian-Early Albian as well.
). &
(2002) described the genus from
Late Aptian-Early Albian as well.
Family Hemihoplitidae , 1924
Genus Hemihoplites , 1924
Hemihoplites sp.
Material: SANGRE-27, SANGRE-34
Description: Shell evolute, sides compressed, umbilicus deep and umbilical wall vertical. Ribs straight or slightly curved, simple or branching; primary ribs mostly single, starting at the umbilical margin, secondary ribs intercalated between primaries starting in the lower part of the flank.
Discussion: The rib pattern and whorl section are the characteristics for genus and species identification. The San Georgio specimens show similarity to H. feraudianus (d', 1841). This species is similar to H. varicostatus , 1989, in its growth and rib pattern, but (2002) believes the latter species is characterised by a different range of morphological variation in spite of overlap with that of H. feraudianus. This reasoning does not make any sense. (1990) suggested that probably H. feraudianus and H. soulueri ( in , 1878) represent morphotypes variation of a single species.
Distribution: Eskisabel section, Ataun area, Lareo Formation (Fig.
6 ![]() ).
).
Superfamily Douvilleiceratacea & ,
1897
Family Douvilleiceratidae & ,
1897
Subfamily Roloboceratinae , 1961b
Genus Roloboceras , 1954
Type species: Ammonites hambrovi , 1845, Early Aptian (forbesi Zone) SE England.
Generic characters: whorl shape semi-rounded, blunt ribs with tubercles around the umbilicus, umbilicus deep, suture line simplified with deep ventral lobe, body chamber half whorl in length, mouth border plain with a shallow umbilical sinus.
Discussion: (1961b) recognised the genus based on its whorl shape and tuberculation on the flanks.
Occurrence: Southern England, France, Spain (, 1961a; et alii, 2009).
Roloboceras cf. hambrovi (, 1845)
cf. 1845 Ammonites Hambrovi , p. 354, pl. xiii, fig. 4.
cf. 1906 Ammonites Hambrovi : , p.
162-163.
cf. 1930 Cheloniceras hambrovi (): , p. 444-445.
cf. 1954 Roloboceras hambrovi (): , p. 114.
cf. 1961b Roloboceras hambrovi (): , p. 179-182, pl. 29, figs. 5-6; pl. 30, figs. 7-8; pl. 30, fig. 3a-b; pl. 32, fig. 5a-b; text-fig. 54, 55a-b, 57a-b.
cf. 2006 Roloboceras hambrovi (): et alii, pl. 11, fig. 2.
cf. 2008 Roloboceras hambrovi (): et alii, pl. 6, fig. 2.
cf. 2009 Roloboceras hambrovi (): et alii, fig. 10E.
cf. 2009 Roloboceras hambrovi (): et alii, fig. I, B-D, fig. II, A
Lectotype: British Geological Survey Museum, collection 2295, the larger of the two specimens figured by , from Atherfield Clay Series, Lower Lobster Bed.
Material: ERG-3-8, 3-12
Description: Two crushed specimens are to hand. Whorl section sub-rounded to sub-rectangular, venter rounded, primary ribs straight, single, thick, between each primary there are thinner ribs. Tubercles around the umbilicus are prominent.
Discussion: The specimens show similarity to R. horridum (, 1938), but this species has more depressed adolescent whorls and tubercles that are more prominent still.
Occurrence: England (, 1954, 1961c; , 1930), France (, 1913; et alii, 2008), Caucasus (, 1933), Russia (, 1906), USA (, 1949).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Roloboceras cf. saxbyi , 1961b
cf. 1961b Roloboceras saxbyi , p. 188-189, pl. 30, figs. 1a-b, 2a-b; text-fig. 57c-d.
Holotype: Natural History Museum, London, 46590, Atherfield Clay Series, Lower Lobster Bed, Atherfield, Isle of Wight.
Material: SANGRE-11, 13, 28, 30.
Description: Whorl section sub-rectangular to oval, ribs uniform, single or bifurcate from umbilical edge, lateral tubercles not very distinct.
Discussion: (1921, p. 313, quoted in , 1961b, p. 189) believed that R. saxbyi was comparable to Cheloniceras gottschei (, 1902). Detailed examination shows they differ in ornamentation and suture line and Cheloniceras gottschei shows sharper ribs and a venter more flattened venter. R. perli also shows similarity to R. saxbyi but differs from the latter species in its more close and uniform ribbing and less complete tuberculation (, 1961b).
Occurrence: England (, 1961b)
Distribution: Eskisabel section, Ataun area, Lareo Formation.
Superfamily Deshayesitaceae ,
1949
Family Deshayesitidae , 1949
The family is restricted to the latest Barremian to the latest Early Aptian substages. During this interval, an increase in the involution of the whorls, flatting of the venter, appearance of tubercles and a reducing curvature and weakening of the ribs at the venter can be followed in this family. Some genera with their short time ranges and distinct characters are used in Lower Cretaceous biozonation on a global scale.
Deshayesitidae is accepted as a family (, 1955; , 1957; , 1958). et alii (1957) considered that the family comprised three genera, Deshayesites , 1914, Dufrenoyia ex & , 1915, and Burckhardites , 1949. (1964) split the family into two subfamilies Deshayesitinae and Matheroceratinae. Subsequently new genera were proposed for the subfamily. et alii (1996) followed 's subfamily suggestion at systematic level, including genera Turkmeniceras , 1962, Prodeshayesites , 1961b, Deshayesites , 1914, Neodeshayesites , 1964, Dufrenoyia ex & , 1915, Burckhardites , 1949, and Kuntziella , 1962.
& (1999) added a new genus Obsoleticeras & , 1999, and used Paradeshayesites (1967) instead of Prodeshayesites , 1961b, and indicated that Neodeshayesites, Burckhardtites and Kuntziella are possibly assigned to the family. & (2005) have assigned Neodeshayesites, a genus restricted to south and central America, to the subfamily Acanthohoplitinae, but there are problems with this approach which will need addressing. Here the taxonomic classification in et alii (1996) is followed, but, the three genera Turkmeniceras, Deshayesites and Dufrenoyia are common in all classifications. However the suprageneric classification has been under discussion for some long time and is still unsatisfactory.
Subfamily Deshayesitinae ,
1949
Genus Deshayesites , 1914
Type species: Ammonites deshayesi in d', 1841.
Generic characters: Discoidal, sides and venter flattened or gently convex, ventro-lateral and umbilical borders poorly defined. Involution varies from one-fifth to a little more than one-third the diameter. Sculpture of sigmoidal ribs, which tend to lose elevation at mid-flank, divisible into primaries, commencing at umbilicus, and secondaries, which are intercalated or branch from primaries on whorl side. On venter ribs are of uniform relief and are bent forwards in an arc: some species have a smooth siphonal band for a short at period young stage. Mouth-border plain, inclined forwards, sinuous, with shallow embayment at umbilicus, well-marked on test especially near aperture, where it stands out as a sheaf of hairlines. Suture line with trifid, fairly deep first lateral lobe and simplify auxiliary elements with only a gentle retraction towards umbilicus. Body chamber about half a whorl in length.
Discussion: The genus is derived directly from the latest Barremian genus Turkmeniceras , 1962. (1914) introduced Deshayesites with A. deshayesi d' as type species. Originally Deshayesites comprised species which are now distributed among Deshayesites, Prodeshayesites and Dufrenoyia (, 1964). Later (1967) proposed Paradeshayesites , which is similar to Prodeshayesites. (1995) considered 'Prodeshayesites' as a subjective synonym of Deshayesites. 'Prodeshayesites' is limited to east Greenland, England and North Germany. In both East Greenland and eastern England it apparently precedes true Deshayesites (, 1961a, 1964; & , 1999). However, there is evidence in both Greenland and England that the earliest Aptian with Deshayesites oglanlensis is missing in terms of marine sediments ( et alii, in prep.). The taxonomic status of Prodeshayesites is questionable; (1995) considered it to be a synonym of Deshayesites and stressed the affinity of 'Prodeshayesites' with some Turkmenian species of Deshayesites. & (1999, 2004) proposed a new genus Obsoleticeras which is similar to Paradeshayesites or Prodeshayesites. It is suggested that Deshayesites be retained and all other related genera grouped in its synonymy, or possibly regarded as sub genera. The use of Paradeshayesites or Prodeshayesites is one of publication priority. Obsoleticeras cannot be separated from Deshayesites. Parahoplitoides (1922, p. 111) is an objective synonym of Deshayesites having the same type species and was proposed in ignorance of 's genus. Reference in Canadian literature to species of Deshayesites (, 1932; , 1937) is based on of the Albian genus Subarcthoplites , 1954. (1965) believed that the Colombian species described by (1938) as Deshayesites stutzeri, D. nodosus, D. rotundus and D. colombianus are most closely related to Dufrenoyia than to Deshayesites.
Occurrence: The genus is characteristic of the earlier Early Aptian and reported from many places Europe, Russia, Africa, Arctic, America and Iran.
Deshayesites cf. tuarkyricus , 1983
nom. nud. cf. 1979 Deshayesites tuarkyricus , , tab. 2, fig. 2.
cf. 1983 Deshayesites tuarkyricus sp. nov., , p. 132, tab. 1, figs. 1-4, tab. 2, fig. 4.
cf. 1999a Deshayesites tuarkyricus : et alii, fig. 6c-d.
cf. 1999 Deshayesites tuarkyricus : & , pl. 3, fig. a-c.
cf. 1999 Paradeshayesites tuarkyricus (): & , p. 527.
cf. 2000 Deshayesites sp. gr. tuarkyricus : et alii, pl. 1, fig. 3.
cf. 2004 Deshayesites cf. tuarkyricus : , p. 123, fig. 4F.
cf. 2004 Paradeshayesites tuarkyricus (): & , p. 212, pl. 7, fig. 7; text-figs. 19-20.
Holotype: St. Petersburg Museum, N° 1/ 9442, from Lausan, Taurkyr, Turkmenistan.
Material: IRI-1-35, 36, 44, 83, 85, 86, 102.
Description: Whorl section sub-rectangular, venter flat or broadly convex, coiling being more open in the last whorl. Ornament consists of ribs, dense and fine, primary ribs originate from the umbilical edge and are more coarser and stronger in the upper third of the whorl flanks, secondary ribs start from mid flank; every primary rib is accompanied by two secondary ribs. Thirty three primary ribs and seventy secondary ribs are present at thirty five mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| IRI-83 | 28.5 | 11 | 0.38 | 0 | 9 | 0.31 | 0 | 20 | ||
| IRI-86 | 34.8 | 12.5 | 0.35 | 0 | 6 | 0.17 | 0 | 33 | 70 | |
| Holotype | 90.8 | 46 | 0.5 | 24.3 | 0.26 | 17 | 0.18 | 0.52 |
Discussion: Fine ribbing and the coiling pattern are the characters used for recognition. Identification is not easy because specimens show similarity to Turkmeniceras and D. oglanlensis. In fact the species could be a transition form between Turkmeniceras and Deshayesites. D. tuarkyricus differs from D. oglanlensis by its more dense rib pattern. It is supposed that D. tuarkyricus has been reported form Turkmenistan only and is an endemic and local species, but based on et alii (2000), et alii (2009) and this study, it is possible that D. tuarkyricus is present outside of the Kopet Dagh basin. Even if the latter specimens are not D. tuarkyricus, they accompany D. oglanlensis and are of the D. tuarkyricus Zone age.
Occurrence: Turkmenistan, Iran and Spain.
Distribution: Iribas section, Errenaga Formation (Fig.
8 ![]() ).
).
Deshayesites cf. oglanlensis , 1983
num. nud. cf. 1979 Deshayesites oglanlensis : , pl. 1, fig. 5.
cf. 1983 Deshayesites oglanlensis sp. nov.: , p. 136, pl. 1, figs. 5-9; text-figs. 5-6.
cf. 1995 Deshayesites oglanlensis : , p. 74, pl. 9, fig. 1.
cf. 1997 Deshayesites oglanlensis : et alii, fig. 7e.
cf. 1999 Deshayesites oglanlensis : , p. 441, fig. 4a-b.
cf. 1999b Deshayesites oglanlensis : et alii, pl. 1, figs. 2-4.
cf. 2000 Deshayesites oglanlensis : et alii, pl. 1, figs. 1-2; pl. 2, fig. 4.
cf. 2004 Deshayesites oglanlensis : , p. 123-124, fig. 4G-H.
cf. 2004 Paradeshayesites oglanlensis (); & , p. 214, pl. 7, fig. 4; text-figs. 22-23.
Holotype: St. Petersburg Museum, N° 12/ 9442. from Bolshoi Balkhan, Turkmenistan, figured by .
Material: IRI-13, 19, 54, 55, 56, 57, 65, 76, 87.
Description: Some specimens are compressed; whorl section sub-rectangular, venter rounded and narrow, flanks a little flattened, umbilicus is around one quarter of the diameter. Ribs fine and dense, palmate, fasciculate, ribs stem from well-marked peri-umbilical bulla, their relief is weaker in the middle part of the sides. Regularly every two primary ribs commence from one bullae, bifurcate or trifurcate, secondaries start from mid flank. Twenty seven primary ribs and sixty one secondary ribs are present at thirty mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| IRI-11 | 29 | 12.5 | 0.43 | 0 | 3.5 | 0.12 | 0 | |||
| IRI-13 | 36 | 15 | 0.42 | 0 | 12 | 0.33 | 0 | |||
| IRI-76 | 17 | 5 | 10 | 0.29 | ||||||
| IRI-57 | 19 | 5 | 0.26 | |||||||
| IRI-103 | 30 | 12.5 | 0.42 | 0 | 7 | 0.23 | 0 | 27 | 61 | |
| Holotype | 37.5 | 17 | 0.46 | 8.4 | 0.22 | 10.6 | 0.28 | 0.49 | 22 | 92 |
Discussion: Fine, dense ribs and peri-umbilical bullae are characteristic features of this species. (1995) reported the density of ribs in Deshayesites oglanlensis as being more important in the separation of this species from Deshayesites tuarkyricus. (1999) believed that the specimens described as Deshayesites oglanlensis by (1995) are more closely related to Deshayesites planicostatus (, 1991). The Spanish material studied here looks like Deshayesites callidiscus , 1961c. Review of the description of D. oglanlensis and D. callidiscus from the published papers (for example , 1964; & , 2004) show they are morphologically very similar. In my view they might be conspecific, despite the difference in age, or D. callidiscus is a descent of D. oglanlensis and they are phylogenetically related to each other. However, this view needs further study.
Occurrence: Earliest Aptian in France (, 1995) and recorded in an assemblage ammonite fauna of forbesi or weissi Zone age. Deshayesites oglanlensis is a characteristic form of the D. tuarkyricus Zone in south Spain ( et alii, 1997), Romania (, 1999), Turkmenistan (, 1979, 1983) and Iran (, 2004).
Distribution: Iribas section, Errenaga Formation (Fig.
8 ![]() ).
).
Deshayesites cf. antiquus , 1983
num. nud. cf. 1979 Deshayesites antiquus : , tab. 1, fig. 4.
cf. 1983 Deshayesites antiqnuus sp. nov. p. 138-139, tab. 2, figs. 5-6; tab. 3, fig. 8.
cf. 1999 Deshayesites antiquus : & , pl. 2, fig. e.
cf. 1999 Deshayesites antiquus : et alii, p. 178, pl. 16, figs. 1-3; pl. 18, fig. 1.
cf. 1999a Deshayesites antiquus : et alii, pl. 1, fig. 1.
cf. 2000 Deshayesites antiquus : et alii, p. 129, pl. 3, figs. 2-4.
cf. 2010 Deshayesites antiquus : et alii, p. 292, fig. 11A.
Holotype: St. Petersburg Museum, N° 18/9442. from Bolshoi Balkhan, Turkmenistan, Lower Aptian, Deshayesites tuarkyricus Zone.
Material: IRI-61, 62, 90, 95, 116, 117.
Description: Whorl section sub-rectangular to oval, flanks parallel, Primary ribs nearly sharp, originating at the upper part of the umbilical wall and cross the flank in a light sigmoidal pattern, secondary ribs branch from primaries in mid flanks.
Discussion: The species appears in the oglanlensis Zone and can be distinguished by its density of ribs which is less than in other species which appear in this zone such as D. tuarkyricus, D. oglanlensis and D. weissiformis.
Occurrence: Turkmenistan, France and Spain.
Distribution: Iribas section, Errenaga Formation.
Deshayesites cf. gracilis , 1964
cf. 1964 Deshayesites cf. gracilis sp. nov.: , p. 324-325, pl. 47, fig. 10.
cf. 1999 Deshayesites gracilis : , p. 444, fig. 3C.
Holotype: Natural History Museum, London, C3034, Atherfield Clay Series, Crackers, Atherfield, Isle of Wight.
Material: ER-76, 79, 108, 173, 183; ERG-3-9, 3-10.
Description: Specimens are crushed and incomplete, umbilical wall vertical; Ornament consists of ribs; narrow primary ribs start from middle part of the umbilical wall, sigmoidal, secondaries intercalated between primaries, free or attached, starting from lower third of the whorl flank, some fusing with adjacent primaries.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ER-173 | 30 | 12.5 | 0.41 | 0 | 8 | 0.26 | 0 | |
| ERG-34-9 | 46.5 | 18.5 | 0.40 | 0 | 16 | 0.34 | 0 | |
| Holotype | 41 | 18.45 | 0.45 | 12.3 | 0.3 | 0 | 0.66 |
Discussion: (1964) based the species on an incomplete specimen. The species differs from D. forbesi var. koeneni , 1964 by its deep umbilical wall and steep-sided ribs. D. multicostatus , 1963, shows similarity to D. gracilis, but the former species has stronger sigmoidal ribs and a flattened ventral area.
Occurrence: England (, 1964), Romania (, 1999).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).

Click on thumbnail to enlarge the image.
Figure 5: Early Aptian ammonites of northern Spain. A.- Aconeceras cf. nisoides (, 1893), Lareo Formation, NAZ-7; B-C.- Aconeceras haugi , 1893, Lareo Formation, AIZMUSU 2, Lareo Formation; D.- Aconeceras (Theganoceras) sp., Errenaga Formation, IMA-7; E.- Pseudosaynella sp., Errenaga Formation, ER-22; F.- Hemihoplites sp., Errenaga Formation, Sangre-34; G.- Roloboceras cf. hambrovi (, 1845), Errenaga Formation, ERG-3-8; H.- Roloboceras cf. saxbyi , 1961b, Errenaga Formation, Sangre-11; I.- Deshayesites cf. tuarkyricus , 1983, Errenaga Formation, IRI-83; J.- Deshayesites cf. oglanlensis , 1983, Errenaga Formation, IRI-57; K.- Deshayesites cf. antiquus , 1983, Errenaga Formation, IRI-90; L.- Deshayesites cf. gracilis , 1964, Errenaga Formation, ER-76.
Click on thumbnail to enlarge the image.
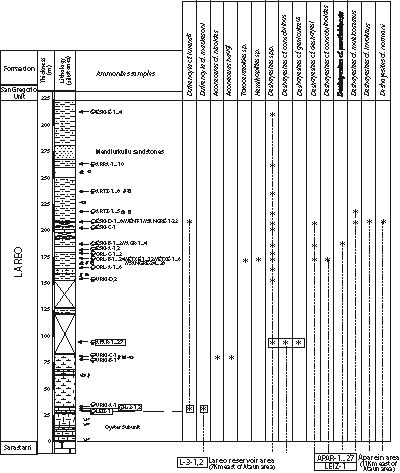
Figure 6: Ammonite range chart of the deshayesi-furcata transitional interval Zone in the composite Lareo Formation succession.
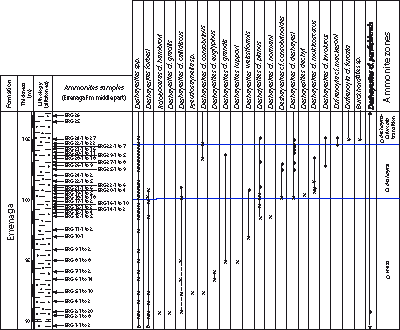
Click on thumbnail to enlarge the image.
Figure 7: Ammonite range chart of the weissi, deshayesi, deshayesi-furcata transitional interval zones in the Errenaga Formation of the Igaratza section.
Click on thumbnail to enlarge the image.
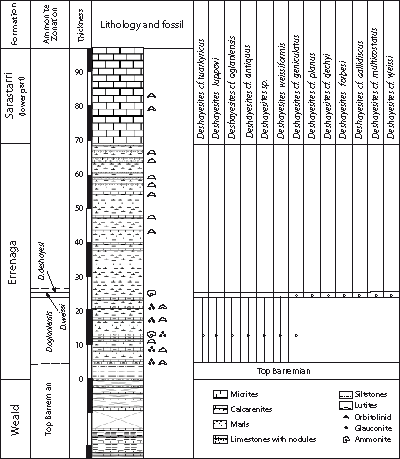
Figure 8: Ammonite range chart and biozonation for the lower part of the Errenaga Formation of the Iribas section.
Deshayesites cf. bodei (von , 1902)
cf. 1902 Hoplitidae bodei (von ), p. 221, pl. 9, fig. 1a-c.
cf. 1924 Parahoplitoides bodei (von ): , p. 433.
cf. 1960 Deshayesites bodei (von ): & , tab. 1, fig. 4.
cf. 1964 Prodeshayesites bodei (von ): , p. 358-359, pl. 58, figs. 4a-b, 5a-b; pl. 59, fig. 3; text-fig. 127a-b.
cf. 1967 Deshayesites bodei (von ): , tab.
12-13.
cf. 1971 Deshayesites bodei (von ): , pl. 30, fig. 3.
cf. 1973 Deshayesites bodei (von ): , pl. 91.
cf. 1979 Prodeshayesites bodei (von ): et alii, p. 147, pl. 2, figs. 1a-b, 2a-b.
cf. 1995 Deshayesites bodei (von ): , taf. 3, figs. 3-4; taf. 5, fig. 4.
cf. 1999 Prodeshayesites cf. bodei (von ): & , p. 87-88, fig. 3a-d.
cf. 2004 Deshayesites bodei (von ): & , pl. 2, fig. 1.
Lectotype: The example figured by von (1902), pl. 9, fig. 1a-c; text-fig. 127a-b, from the Lower Aptian of Timmern, Brunswick (selected by , 1964).
Material: ER-9.
Description: One crushed specimen. Whorl section sub-rectangular, flanks parallel, venter nearly curved and broad. Ribs sharp, primary ribs originate from upper part of umbilical wall and cross the flank in a sigmoidal curve; secondary ribs branch from primaries near to the umbilical margin.
Discussion: D. bodei is similar to D. fissicostatus, but differs from the latter by a wider umbilicus and coarser ribs. (1964, p. 359) indicated that at some stages of growth, the two species are very similar, but believes that since D. bodei is used as subzonal index it is better to keep the two as distinct. However, the presence of D. bodei confirms the fissicostatus Zone.
Occurrence: Spain ( et alii, 1979), England (, 1924; , 1964), Germany (, 1967), Russia ( & , 1960; , 1973).
Distribution: Igaratza section, Errenaga Formation.
Deshayesites weissiformis , 1983
1983 Deshayesites weissiformis sp. nov.: , 34, tab. 2, figs. 1-3; tab. 3, fig. 7; text-figs. 3-4.
1995 Deshayesites weissiformis : , p. 74, pl. 5, fig. 2.
1999 Deshayesites weissiformis : , p. 440-441, fig. 3A-E.
1999a Deshayesites weissiformis : et alii, p. 278, pl. 6, figs. 2-3.
2004 Deshayesites cf. weissiformis : , p. 124, fig.
4.
2004 Paradeshayesites weissiformis (): & , p. 212-214, pl. 7, fig. 3; text-fig. 21.
Holotype: St. Petersburg Museum, N° 7/9442, from Taurkyr, Turkmenistan.
Material: ER-2, 13, 14, 28, 55, 129?, ERG-10-1, 20-3, IRI-3, 4, 11, 15, 20, 21. 24, 32, 42, 45, 46, 59, 77, 81, 101, 102, 103, 104, 105, 109, 110, 114, 115, 107, 108, ATAN-11.
Description: Some specimens crushed and incomplete. Whorl section sub-rectangular, flanks parallel, flat and broad venter, umbilical area is between one third and one fourth of the diameter. Ornament consists of ribs; primary ribs start from the upper part of the umbilical wall with tubercles around the umbilical margin, bifurcating in the lower quarter of the flanks. Secondary ribs are of same thickness as the primaries; some intercalating between primary rib branches, starting near half way up the flank, free or attached to primaries. All ribs are curved forward on the upper part of the flank, but are straight at the ventral area. Twenty eight primary ribs and fifty eight secondary ribs are present at a diameter thirty seven mm.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| IRI-4 | 37 | 14.5 | 0.39 | 5 | 0.13 | 5 | 0.13 | 0.34 | 28 | 58 |
| IRI-110 | 29 | 12.5 | 0.43 | 0 | 4.5 | 0.15 | 0 | |||
| IRI-13 | 36.5 | 11 | 0.30 | 0 | 9 | 0.24 | 0 | |||
| IRI-59 | 17 | 8 | 0.47 | 0 | 3.5 | 0.21 | 0 | 24 | ||
| IRI-110 | 36 | 16 | 0.44 | 0 | 0 | 0 | 48 | |||
| ATAN-26 | 35 | 14.5 | 0.41 | 0 | 7 | 0.2 | 0 | |||
| ERG-10-1 | 52 | 19 | 0.36 | 0 | 16.5 | 0.31 | 0 | |||
| Holotype | 110.7 | 51.6 | 0.46 | 32.7 | 0.29 | 26.9 | 0.24 | 0.63 | 24 | 94 |
Discussion: the species is identified by its fine and dense ribs. Shells are similar to D. oglanlensis and sometimes can not be easily separated at the same dimensions as D. weissiformis and D. oglanlensis. However the number of ribs in D. weissiformis is less than D. oglanlensis at the same diameter.
Occurrence: Turkmenistan (, 1983), France (, 1995; et alii, 1999a), Romania (, 1999) and Iran (, 2004).
Distribution: Ataneta locality, Iribas section, Igaratza section, Iribas section, Errenaga Formation
(Figs.
7 ![]() - 8
- 8 ![]() ).
).
Deshayesites cf. consobrinus (d', 1841)
cf. 1841 Deshayesites consobrinus (d'), p. 147, pl. 47, figs. 1-3.
cf. 1964 Deshayesites consobrinus (d'): , text-figs. 123-124.
cf. 1973 Deshayesites consobrinus (d'): , tab. 88, 89, 90.
cf. 1979 Deshayesites consobrinus (d'): , p. 159, pl. 2, figs. 3-4.
cf. 1999 Deshayesites consobrinus (d'): & , pl. 3, figs. g-i.
cf. 1999 Deshayesites consobrinus (d'): et alii, pl. 19, figs. 1, 4.
cf. 2000 Deshayesites consobrinus (d'): et alii, pl. 5, fig. 1.
cf. 2004 Deshayesites consobrinus (d'): & , p. 202-203, pl. 4, figs. 1-3.
Lectotype: Laboratoire de Paléontologie, Muséum National d'Histoire naturelle de Paris, d' collection 5597a, Lower Aptian, La Bédoule (Bouches-du-Rhône, France) (selected , 1964).
Material: ERG-5-8, 28-2, 33-15, APAR-16.
Description: Whorls sub-rectangular, flanks parallel to convex, umbilical wall nearly vertical. Ribs narrow, sharp, sigmoidal; primary ribs originate from umbilical wall, secondary ribs, attached or free, single, occurring between a pair of primaries and appear at one-third up the whorl flank: rarely more than one secondary between each pair of primaries in the last whorl.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ERG-5-8 | 24.5 | 8.5 | 0.34 | 0 | 7 | 0.28 | 0 | 20 | ||
| Holotype | 40 | 0 | 10 | 0.25 | 14 | 0.35 |
Discussion: D. consobrinus differs from D. deshayesi in having less sigmoidal ribs and less overlapping whorls. et alii (2000) revised collected specimens from its stratotype area that agreed with lectotype. (1999, p. 452) assigned a few specimens to D. bogdanovae sp. nov. & (2004, p. 209) described a new species D. kemperi ( & , 2004) derived from D. consobrinus. The figured specimens indicate that both new species are very similar to D. consobrinus and might be just intra-specific variations.
Occurrence: England (, 1964), France ( et alii, 2000), Russia (, 1973) and Turkmenistan ( & , 1999).
Distribution: Igaratza section, Errenaga Formation (Fig.
6 ![]() ).
).
Deshayesites forbesi , 1961c
1845 Ammonites Deshayesii : , p. 353, pl. 13, fig. 2.
1947 Deshayesites deshayesi (d'): , p. 172-173, fig. 14a-b.
1961c Deshayesites forbesi sp. nov.: , p. 593, pl. 81, fig. 2a-b.
1964 Deshayesites forbesi : , p. 314-317, pl. 47, figs. 1-7; text-fig. 104g, 106i, 110a.
1999 Deshayesites forbesi : , p. 449, fig. 6F-H.
2009 Deshayesites forbesi : et alii, fig. 9E.
2010 Deshayesites forbesi : et alii, fig. 11C.
Holotype: British Geological Survey Museum, 30918, Atherfield Clay Series, Crackers, Atherfield, Isle of Wight.
Material: ER-50, 51, 65, 70, 125, 127, 135?, 137?, 138, 139, 140, 142, 143, 144, 145, 146, 147, 149, 153, 154, 156, 158, 161, 168, 169, 170, 171, 175, 177, 178, 180, 181, 185, 186, ERG-1-1, 2-1, 2-3, 2-4, 2-5, 2-7, 2-8, 3-1, 3-2, 3-5, 3-14, 3-15, 3-16, ERG-4-1, 5-1, 5-2, 5-3, 5-6, 5-9, 12-1, 17-1, 18-1, 20-2, ATAN-25, IMA 5, 6, 8, 10, 12.
Description: Whorls sub-rectangular, flanks parallel to convex, umbilical wall nearly oblique; umbilical width is between one fifth and one quarter of the diameter. Ornamentation consist of ribs; primary ribs sharp, nearly sigmoidal; secondary ribs bifurcating, appear at mid flank, rarely some primary ribs are single in later growth stages.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ER-50 | 11.5 | 8 | 0.69 | |||||||
| ER-65 | 31 | 13.5 | 0.43 | 0 | 8 | 0.25 | 0 | |||
| ER-137 | 17 | 7 | 0.41 | 3 | 0.18 | 5 | 0.29 | 0.42 | 22 | 48 |
| ER-140 | 19 | 8 | 0.42 | 0 | 5 | 0.26 | 0 | |||
| ER-144 | 23 | 9 | 0.39 | 0 | 5 | 0.22 | 0 | |||
| ER-145 | 18 | 8.5 | 0.47 | 0 | 4 | 0.22 | 0 | |||
| ER-147 | 23 | 11 | 0.47 | 0 | 5 | 0.22 | 0 | |||
| ER-156 | 19.5 | 11.5 | 0.58 | 0 | 4 | 0.21 | 0 | |||
| ER-168 | 26 | 11.5 | 0.44 | 0 | 4.5 | 0.17 | 0 | |||
| ER-171 | 25 | 11 | 0.44 | 0 | 5 | 0.2 | 0 | |||
| ER-181 | 20.5 | 10 | 0.48 | 0 | 4.5 | 0.22 | 0 | |||
| ERG-5-1 | 37 | 15 | 0.40 | 0 | 12 | 0.32 | 0 | |||
| Holotype | 72 | 27.3 | 0.38 | 24.4 | 0.33 | 19.4 | 0.26 | 0.33 |
Discussion: The diagnostic feature for the species is density of its ribbing. D. forbesi shows similarity to D. deshayesi in rib pattern, but the latter species has strong and more uniform ribs and also a vertical umbilical wall.
Occurrence: England (, 1961c), Romania (, 1999).
Distribution: Ataneta locality, Iribas section, Igaratza section, Errenaga Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. euglyphus , 1964
cf. 1964 Deshayesites euglyphus sp. nov.; , p. 336-337, pl. 52, figs. 1-4; pl. LVI, fig. 1a-b.
cf. 1979 Deshayesites euglyphus : et alii, p. 148, pl. 2, fig. 4a-c.
cf. 1999 Deshayesites euglyphus : , pl. 2, fig. 7.
cf. 2004 Deshayesites cf. euglyphus : , p. 125. fig. 4I.
cf. 2010 Deshayesites cf. euglyphus : et alii, fig. 11D.
Holotype: Reading University, Geology Dept., N° 6958, from Atherfield Clay Series, Atherfield, Isle of Wight, UK.
Material: ER-1, 3, 4, 5, 6, 16, 17, 40, 62, 73, 85, 103, ERG-6-8, 6-10, 6-13, 7-1.
Description: Early whorls cannot be seen because specimens are incomplete. Ornament of ribs; primary ribs start from upper part of the umbilical wall, secondary ribs attach to primaries or are free, bifurcating and originate in the lower third of the flank to mid flank, ribs more flattened and projected forward on the venter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ER-17 | 10.5 | 6.5 | 0.61 | |||||
| Holotype | 66 | 29 | 0.44 | 19.8 | 0.33 | 18.4 | 0.28 | 0.68 |
Discussion: The pattern of rounded and flattened ribs in the ventral area is a characteristic feature for species identification. According to (1964) there are some similarities between D. euglyphus and D. kiliani , 1930. The Spanish assemblage is of the D. euglyphus morphotype.
Occurrence: England (, 1964), Spain ( et alii, 1979) and Iran (, 2004).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Deshayesites cf. punfieldensis , 1930
cf. 1930 Deshayesites punfieldensis , p. 431-432, pl. 16, fig. 3a-b.
cf. 1964 Deshayesites punfieldensis : , p. 338-341, pl. 46, figs. 1-3.
Holotype: British Geological Survey Museum, 30915, from the Lower Lobster Bed, Atherfield, Isle of Wight (F.W. Simms University of the Basque Country).
Material: ER-77, 81, 89, 101, ERG-3-9, 3-10.
Description: Whorls sub-rectangular, sides flattened. Ornament of sharp ribs; primary ribs curved from mid-flanks, commencing from the upper part of umbilical wall; secondary ribs start from the upper third of the flank, intercalated between primaries, mostly free or attached to the primaries.
Discussion: The rib pattern is a character for identification. There is similarity between D. vectensis , 1930, and D. punfieldensis, but the latter species has a wider umbilicus, is more coarsely ribbed and is less flattened on the venter. Hitherto, it seems, the species has not been recorded outside of England.
Occurrence: England (, 1964)
Distribution: Igaratza section, Errenaga Formation (Fig.
6 ![]() ).
).
Deshayesites luppovi , 1983
1952 Deshayesites aff. dechyi : , 203, tab. 7, fig. 1.
nom. nud. 1971 Deshayesites luppovi sp. nov.; , 22.
1983 Deshayesites luppovi sp. nov.: , p. 139. tab. 3, figs. 1-6.
1997 Deshayesites cf. luppovi : et alii, fig. 7f-g.
1999 Deshayesites luppovi : , p. 447, fig. 5f-g.
1999 Deshayesites luppovi : , pl. 1, fig. 10.
1999 Deshayesites luppovi : & , pl. 3, fig. f.
1999 Deshayesites luppovi : et alii, p. 178-179, pl. 16, figs. 4-5.
1999a Deshayesites luppovi : et alii, pl. 1, fig. 8.
2000 Deshayesites luppovi : et alii, pl. 2, figs. 1-2; pl. 4, fig. 2.
2004 Deshayesites luppovi : , p. 125, figs. 5A-C, L.
2004 Deshayesites luppovi : & , p. 207-208, pl. 2, figs. 8-9; text-fig. 14.
2010 Deshayesites luppovi : et alii, fig. 11B.
Holotype: St. Petersburg Museum, N° 23/ 9442, from Bolshoi Balkhan, Turkmenistan.
Material: ER-20, 24, 43, 128, ERG-8-5, IRI-5, 7, 8, 9, 10, 16, 17,22, 23, 25, 26, 30, 43, 47, 49, 51, 52, 60, 63, 66, 67, 73, 74, 79, 80, 88, 89, 94, 96, 97, 100, 106, 112, 118.
Description: Most specimens are crushed and incomplete. Whorl section sub-rectangular, whorl height is greater than whorl width, thickest in the lower part of the whorl flank, compressed. Umbilical wall vertical, umbilical width is about one quarter of the diameter. Ornament consists of sigmoidal ribs, strongly curved backwards on the lower third of the flanks; primary ribs start from the upper part of the umbilical wall, thickest in the lower and upper part of flanks; secondaries at first are irregular and in mode of intercalation between primaries, either in ones or twos, starting from the lower third of the flank, some fusing with adjacent primaries. With further growth, ribbing becomes more regular. Twenty two primary ribs in 37 mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| IRI-49 | 27.5 | 11 | 0.4 | 0 | 4.5 | 0.16 | ||||
| IRI-63 | 37.5 | 16 | 0.42 | 0 | 9 | 0.24 | 23 | |||
| IRI-67 | 26.5 | 12.5 | 0.47 | 0 | 6.5 | 0.24 | 22 | |||
| IRI-100 | 22.5 | 10 | 0.44 | 0 | 6 | 0.26 | ||||
| Holotype | 157 | 71 | 0.45 | 40 | 0.25 | 38 | 0.24 | 0.56 | 34 | 93 |
Discussion: The Spanish specimens are comparable with the holotype figures in the rib pattern. S shape rib pattern is a good character for identification. Some specimens resemble Deshayesites forbesi and there is not much difference between Deshayesites forbesi and Deshayesites luppovi expect in their suture lines and this feature cannot be seen in the studied specimens. It seems that the rib pattern in Deshayesites forbesi is more regular than Deshayesites luppovi. Deshayesites forbesi differs from Deshayesites pappi , 1991, by the secondary ribs, which start in the upper part of the whorl flanks in the latter species.
Occurrence: Lower Aptian (weissi Zone) in north Caucasus and in Turkmenistan (, 1971, 1983), Romania (, 1999) and Iran (, 2004).
Distribution: Igaratza section, Iribas section, Errenaga Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. weissi & , 1881
cf. 1881 Hoplites weissi & , p. 179, pl. 46, fig. 1; pl. 47, fig. 1.
cf. 1902 Hoplites (Deshayesites) weissi & : von , p. 207, pl. 45, fig. 1.
cf. 1960 Deshayesites weissi ( & ): & , p. 310, pl. 1, fig. 1.
cf. 1971 Deshayesites weissi ( & ): , p. 22.
cf. 1977 Deshayesites weissi ( & ): , p. 47, pl. 1, figs. 1-4; pl. 4, fig. 6.
cf. 1999 Deshayesites weissi ( & ): , p. 439, fig. 2a-c.
cf. 1999 Deshayesites weissi ( & ): , pl. 4, figs. b-c.
cf. 2000 Deshayesites sp. gr. weissi ( & ): et alii, pl. 6, fig. 1.
cf. 2004 Deshayesites weissi ( & ): , p. 127, fig. 4L.
Syntypes: Both the specimens figured by & (1881) are lost; one was in the collection in the Königlichen Geologischen Landesanstalt, Berlin, and the other was in the Geologischen Reichsanstalt, Vienna. A neotype should be designated from appropriate German material.
Material: ATAN-2, 10, 20.
Description: Whorl height greater than whorl thickness, subparallel flanks. Ribs start from umbilical wall, S shaped, bifurcating in the lower third to mid flank; some secondary ribs are free and unattached to the primary ribs, projected forward in ventral area; secondary ribs are about twice in number to the primaries. In small specimens, ribs are stronger and more dense.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ATAN-20 | 29 | 13.5 | 0.46 | 0 | 7 | 0.24 | 0 | |||
| Holotype | 157 | 71 | 0.45 | 40 | 0.25 | 38 | 0.24 | 0.56 | 34 | 93 |
Discussion: Curved compact (dense) ribs are characteristic for specific separation. However their density is less than that of D. tuarkyricus and D. weissiformis. The rib pattern of D. weissi is similar in some cases to D. tuarkyricus at small diameter, but at larger diameter the ribs are coarser in D. weissi and differ from D. tuarkyricus. & (2004) believe D. weissi is a transition form between D. tuarkyricus and D. weissiformis.

Click on thumbnail to enlarge the image.
Figure 9 : Early Aptian ammonites of northern Spain. A-B.- Deshayesites cf. bodei (von ) 1902, Errenaga Formation, ER-9; C.- Deshayesites weissiformis , 1983, Errenaga Formation, IRI-4; D.- Deshayesites cf. consobrinus (d', 1841), Errenaga Formation, ERG-5-8; E.- Deshayesites forbesi , 1961c, Errenaga Formation, ER-137; F-G.- Deshayesites cf. euglyphus , 1964, Errenaga Formation, ER-17; H.- Deshayesites cf. punfieldensis , 1930, Errenaga Formation, ERG-3-9; I.- Deshayesites luppovi , 1983, Errenaga Formation, IRI-94; J.- Deshayesites cf. weissi & , 1881, Errenaga Formation, ATAN-20; K.- Deshayesites spp., Errenaga Formation, IRI-53.
Occurrence: Germany (von , 1902), Russia ( & , 1960), Turkmenistan (, 1977), Romania (, 1999).
Distribution: Ataneta locality, Iribas section, Errenaga Formation (ammonite assemblage 11,
Figs. 3 ![]() - 4
- 4 ![]() ).
).
Deshayesites dechyi , 1907
1907 Parahoplites dechyi; , p. 171, pl. 9, figs. 1-5.
1952 Deshayesites dechyi : , p. 204, pl. 7, figs. 2-4.
1960 Deshayesites dechyi : & , p. 310, pl. 1, fig. 6.
1977 Deshayesites dechyi : , p. 50, pl. 2, figs. 1-5.
1979 Deshayesites dechyi :
et alii, p. 5, pl. 1, figs. 1-5; pl. 2, figs. 1-3; text-fig. 2.
1999 Deshayesites dechyi : , pl. 2, figs. 3-4.
1999 Deshayesites dechyi : & , pl. 4, fig. h.
2004 Deshayesites dechyi : , p. 129, fig. 4M-N.
2004 Deshayesites dechyi : & , p. 203-204, pl. 3, figs. 6-8.
Lectotype: K7593 Geological Museum of the Geological Institute of Hungary, Budapest (selected by from collection pl. 9, fig. 3), Lower Aptian, Lavaschi, Daghestan.
Material: ER-36?, 58, 59, 63, 64, 83, 88, 91, 92, 113, ERG-19-4, 20-1, ATAN-15.
Description: Whorl section sub-rectangular, semi-involute; umbilical area is about one quarter of the diameter. Ribs sigmoidal, maximum elevation and strength at lower third of sides, one or two secondary ribs are intercalated with primaries; in last half whorl ribs are more regular. Twenty two primary ribs and forty four secondary ribs are present 21 mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ER-58 | 33 | 12.5 | 0.38 | 0 | 8 | 0.24 | ||||
| ER-113 | 26 | 13.5 | 0.51 | 0 | 7 | 0.26 | ||||
| ATAN-15 | 21 | 10 | 0.47 | 3.5 | 0.16 | 0 | 22 | 44 | ||
| Holotype | 122 | 48.5 | 0.4 | 30.5 | 0.25 | 34 | 0.27 | 0.62 |
Discussion: Rib pattern and suture line are characteristic for species identification. Deshayesites dechyi is similar to D. forbesi in rib pattern, but the latter is more evolute and has less strong ribs. D. dechyi differs from D. consobrinoides , 1909, by a less convex venter and from Deshayesites consobrinus by having irregularly branching ribs ( et alii, 1979). In whorl section D. dechyi is similar to Prodeshayesites bodei. The main difference between these two species lies in the suture line. The suture of D. dechyi is characterised by narrow and high elements. Moreover the rib pattern of D. dechyi is denser than D. bodei.
Occurrence: Early Aptian northwest and north Caucasus (dechyi-deshayesi Zone) ( & , 1960; et alii, 1979), Caspian region (weissi-deshayesi Zone) (, 1977) and Kopet Dagh (, 2004).
Distribution: Ataneta locality, Iribas section, Igaratza section, Errenaga Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. callidiscus , 1961c
cf. 1961c Deshayesites callidiscus sp. nov.: , p. 594, 507, 609, pl. 80, fig. 10.
cf. 1964 Deshayesites callidiscus , p. 327-328, pl. 49, figs. 3a-b, 4a-b; pl. 51, fig. 3; pl. 53, fig. 2a-b; text-fig. 114a.
cf. 1971 Deshayesites callidiscus : , pl. 22, fig. 4.
cf. 1979 Deshayesites callidiscus : et alii, p. 149, pl. 3, fig. 4a-b.
cf. 1999 Deshayesites callidiscus : & , pl. 6, figs. e-f.
cf. 2000 Deshayesites callidiscus : et alii, pl. 6, fig. 3.
cf. 2002 Deshayesites callidiscus : & , pl. 1, fig. 1.
cf. 2004 Paradeshayesites callidiscus (): & , p. 211-212, pl. 3, fig. 1; pl. 8, figs. 1-2; pl. 10, fig. 1; text-fig. 18.
Holotype: Natural History Museum, London, 48836, Atherfield Clay Series, Crackers, Atherfield, Isle of Wight.
Material: ER-11, 21, 25, 27, 29, 32, 34, 37, 42, 44, 47, 52, 53, 61, 62, 93, 100, 126, 133, ERG-3-13, 6-7, 8-1, 8-2, 18-7, 18-16, 21-3, 21-4, ATAN-7, 8, 16, 19, 31.
Description: Whorl section sub-rectangular, flanks parallel to convex, umbilical wall nearly vertical, umbilical width is between one fifth and one quarter of the diameter. Ribs low, flat topped in the upper part and around the venter, primary ribs originate from just above the umbilical wall, secondary ribs, mostly single, originate at the mid flank, rarely more than one secondary rib between each pair of primaries in the last whorl. Twenty six primary ribs at umbilical margin and forty eight primary and secondary ribs in ventral margin at 28 mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ER-21 | 88 | 41 | 0.46 | 16 | 0.18 | 24.5 | 0.27 | 0.39 | ||
| ER-47 | 27 | 12 | 0.44 | 0 | 0 | 0 | ||||
| ER-93 | 37.5 | 18 | 0.48 | 0 | 7.5 | 0.2 | 0 | |||
| ERG-8-1 | 29.5 | 12 | 0.40 | 0 | 5.5 | 0.19 | 0 | |||
| ERG-21-3 | 25 | 10 | 0.4 | 0 | 6 | 0.24 | 0 | 18 | ||
| ERG-21-4 | 27.8 | 11 | 0.39 | 0 | 5 | 0.18 | 0 | 26 | 48 | |
| Holotype | 110 | 53.9 | 0.48 | 29.7 | 0.27 | 25.3 | 0.23 | 0.55 |
Discussion: Flattened ribs in the upper part of the flank is a character for the species. D. callidiscus differs from D. kiliani , 1930, in its stronger, denser and more sigmoidal ribs. The whorl section is also more rectangular. D. callidiscus shows affinity with D. grandis (, 1930), but the latter species has denser ribs and degeneration of ribbing does not extend to umbilical margin. D. callidiscus shows similarity to D. topleyi (, 1930), but the former species has less sharp ribs.
Occurrence: Spain ( et alii, 1979), England (, 1964), France ( et alii, 2000), Russia ( & , 2002) and Turkmenistan ( & , 1999).
Distribution: Ataneta locality, Iribas section, Igaratza section, Errenaga Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. normani , 1964
cf. 1930 Deshayesites sp. nov.: , p. 431, pl. 17, fig. 5.
cf. 1961c Deshayesites sp. nov.: , p. 507, 609.
cf. 1964 Deshayesites normani sp. nov.: , p. 344-347, pl. 50, fig. 7; pl. 54, fig. 1; pl. 55, figs. 2-4; pl. 56, fig. 3; pl. 57, figs. 7-8; text-fig. 121.
cf. 1999 Deshayesites normani : , p. 450-451, fig. 7C-D.
cf. 2000 Deshayesites sp. gr. spathi / normani : et alii, pl. 5, fig. 2.
Holotype: Museum, Cambridge, SM B27036, Atherfield Clay series, Crackers, Atherfield, Isle of Wight.
Material: ERG-12-4, SANGRE-23.
Description: Whorl section sub-rectangular, whorl height is nearly equal to whorl width, umbilical wall vertical; sides flattened, venter nearly convex. Ornament consists of ribs, primary ribs thick, sigmoidal, projected backwards on the mid flanks, starting from umbilical wall, secondaries bifurcate from mid flank. On the venter side ribs are nearly straight.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-12-4 | 30 | 13.5 | 0.45 | 0 | 8 | 0.26 | 0 | |
| ERG-30-1 | 28.5 | 12.5 | 0.43 | 0 | 5.5 | 0.19 | 0 | |
| Holotype | 173 | 62.2 | 0.36 | 0 | 0 | 0 |
Discussion: D. normani shows similarity in rib pattern to D. spathi , 1961c, but differs in its more rectangular venter, fewer secondary ribs and absence of smoothness in middle of the sides.
Occurrence: England (, 1964), Romania (, 1999).
Distribution: Igaratza section, Errenaga Formation, Eskisabel section, Ataun area, Lareo Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. saxbyi , 1964
cf. 1930 Deshayesites consobrinoides (): , p. 427.
cf. 1961c Deshayesites sp. nov.: , p. 507, 609.
cf. 1964 Deshayesites saxbyi sp. nov.: , p. 349-350, pl. 53 fig. 1a-c; pl. 56, fig. 6a-b.
Holotype: Natural History Museum, London, 46587, Atherfield Clay Series, Crackers, Atherfield, Isle of Wight ( collection).
Material: ER-46, 54, 74, 118, 172, ERG-18-2, 25-3, 26-3, 28-2, SANGRE-13, 28, 30.
Description: Umbilical wall nearly vertical, sides parallel. Ribs strong, primary ribs sigmoidal, originating from just above the umbilical wall, secondary ribs in one or twos, mostly attached but some free originating at the lower or mid flank.
Discussion: D. saxbyi differs from D. latilobatus (, 1909), in its less flattened sides and ribbing which is sharper than D. latilobatus. D. saxbyi might show similarity with D. consobrinoides , 1909, but the latter species has more regular ribbing which rarely bifurcates.
Occurrence: England (, 1964).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Deshayesites cf. involutus , 1930
cf. 1930 Deshayesites involutus , p. 432.
cf. 1961c Deshayesites involutus : , p. 609.
cf. 1964 Deshayesites involutus : , p. 310, pl. 45, figs. 1a-c, 4a-b; text-fig. 107.
cf. 1999 Deshayesites cf. involutus : , p. 454, fig. 8d.
cf. 2004 Deshayesites cf. involutus : , p. 129-130, fig. 6A-C.
Holotype: British Geological Survey, N° 30919, from Hythe beds, Hythe, Kent, UK.
Material: ERG-26-2, 26-8, 34-13, 34-8, SANGRE-2.
Description: Umbilical wall vertical with rounded shoulders, umbilical area is around one third of diameter. Ribs weakly sigmoidal in shape, primaries smooth at mid-flank and strong near the umbilical margin and at the ventral area, costate near the umbilical wall. Bifurcate or trifurcate, secondary ribs irregular, start in the mid flank.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ERG-34-8 | 41 | 15.5 | 0.37 | 0 | 11.5 | 0.28 | 0 | 24 | ||
| Holotype | 122 | 48.5 | 0.4 | 30.5 | 0.25 | 34 | 0.28 |
Discussion: Rib pattern is comparable to figured specimens of Deshayesites involutus and differs from Deshayesites grandis in finer ribbing. The incomplete specimens are insufficient to permit precise identification.
Occurrence: England (, 1930; , 1964), Romania (, 1999) and Iran (, 2004).
Distribution: Igaratza section, Errenaga Formation (Figs.
6 ![]() -
7
-
7 ![]() ).
).
Deshayesites cf. planus , 1961c
cf. 1961c Deshayesites planus , p. 609.
cf. 1964 Deshayesites planus : , p. 323, pl. 57, fig. 5; text-figs. 112a-b, e.
cf. 1971 Deshayesites planus : , p. 22.
cf. 1977 Deshayesites planus : , p. 52, pl. 3, figs. 1-5; pl. 4, figs. 7-8; text-fig. 4a-b.
cf. 1999 Deshayesites planus : , p. 445, fig. 5c-e.
cf. 1999 Deshayesites planus : & , pl. 4, figs. g, i.
cf. 2004 Deshayesites cf. planus :
, p. 130, fig. 5J-K.
Holotype: British Geological Survey, ZM 1667 ( collection), from Atherfield Clay Series, Atherfield, Isle of Wight, UK.
Material: ER-8, 12, 31, 41, 48, 60, 84, 94, ERG-12-2, 16-1, 18-5, 19-1, 22-3, 22-4, 22-5, 28-7, 34-1, 34-3, 34-4, 34-6, ATAN-24, 26, 28.
Description: Whorl section sub-rectangular, thickest in the lower one third of flanks; umbilical area is one third of diameter. Ribs narrow; primary ribs originate from upper part of umbilical wall and cross the flank in a sigmoidal curve, maximum elevation at lower third of flank, minimum elevation in middle of flank. Tubercles are apparent around the umbilical margin as a result of thickening of the ribs. One or two secondary ribs are intercalated between each pair of primaries.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ER-94 | 17 | 7.5 | 0.44 | 0 | 4.5 | 0.26 | 18 | |||
| ERG-28-7 | 26 | 11 | 0.42 | 0 | 6 | 0.23 | ||||
| Holotype | 33 | 14.7 | 0.46 | 11 | 0.33 | 7.9 | 0.24 | 0.74 | 44 |
Discussion: Ribs are strong at the umbilical margin with low elevation at mid-flank and with evolute coiling are features characteristic of this species. Measured dimensions similar to the holotype.
Occurrence: England (, 1964), Romania (, 1999), Turkmenistan (, 1977) and Iran (, 2004).
Distribution: Ataneta locality, Iribas section Igaratza section, Errenaga Formation
(Fig.
7 ![]() ).
).
Deshayesites cf. multicostatus , 1935
cf. 1935 Deshayesites multicostatus , p. 31, pl. 1, fig. 1a-c.
cf. 1961c Deshayesites multicostatus : , p. 508, 523, 569, 570, 609.
cf. 1964 Deshayesites multicostatus : , p. 304-305, pl. 43, figs. 5a-b, 6.
cf. 1973 Deshayesites multicostatus : , p. 130, pl. 84, fig. 1.
cf. 1999 Deshayesites multicostatus : , p. 441, fig. 3b-d.
cf. 2004 Deshayesites cf. multicostatus : , p. 132, fig. 5M.
Holotype: Natural History Museum, London, C36366 ( collection), from the Sutterby, Marl of Sutterby Lincolnshire, UK.
Material: ER-45, ERG-23-1, 23-2, 28-5, ATAN-3, 4, SANGRE-1, 2, 18.
Description: Whorls sub-rectangular, sides flattened, venter convex. Ornament consisting of slightly rounded ribs; primary ribs commence from umbilical wall, are sigmoidal, at first and straighter in the last whorl of adult specimens. Secondary ribs single or in pairs, interposed between primaries mostly free in early whorls and attach in last half whorl.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-23-1 | 31.5 | 13.5 | 0.42 | 4 | 0.12 | 0 | 0.29 | |
| Holotype | 50 | 24 | 0.48 | 0 | 13 | 0.26 | 0 |
Discussion: Dense ribs in ventral area and small umbilicus are characteristic of D. multicostatus. It is distinguished from D. deshayesi var. strigosus by its sharper and denser ribbing and more stronger curved ribs. The Spanish specimens are comparable to 's figures.
Occurrence: England (, 1961c, 1964), Russia (, 1973), Romania (, 1999) and Iran (, 2004).
Distribution: Ataneta locality, Iribas section, Igaratza section, Errenaga Formation, Eskisabel section, Ataun area, Lareo Formation
(Figs.
6 ![]() -
7
-
7 ![]() ).
).
Deshayesites cf. deshayesi in d', 1841
cf. 1841 Ammonites deshayesi in d', p. 288, pl. 85, figs. 3-4.
cf. 1899 Hoplites deshayesi : , p. 108.
cf. 1914 Hoplites (Deshayesites) deshayesi , p. 100-103, pl. 6, figs. 81-83; pl. 7, figs. 100-101.
cf. 1960 Deshayesites deshayesi (d'): & , p. 309, pl. 1, figs. 2, 5.
cf. 1961c Deshayesites deshayesi (d'): , p. 508, 523, 538, 593, 609.
cf. 1964 Deshayesites deshayesi (d'): , p. 295, pl. 43, fig. 3; pl. 47, fig. 9; pl. 51, fig. 6 (see for extensive synonymy).
cf. 1971 Deshayesites deshayesi (d'): , pl. 3, fig. 6; pl. 4, figs. 1-2.
cf. 1971 Deshayesites deshayesi (d'): , pl. 29, fig. 7.
cf. 1973 Deshayesites deshayesi (d'): , p. 120, pl. 76, fig. 1.
cf. 1977 Deshayesites deshayesi (d'): , p. 55, pl. 3, fig. 6; pl. 4, figs. 1-2.
cf. 1979 Deshayesites deshayesi (d'): , pl. 2, fig. 6.
cf. 2004 Deshayesites deshayesi (d'): , p. 130-131, fig. 5D-E & 5H-I.
cf. 2004 Deshayesites deshayesi (d'): & , p. 80, pl. 1, figs. 11, 13.
cf. 2010 Deshayesites deshayesi (d'): et alii, fig. 11, E-G, I; fig. IV, I-L.
Lectotype: Muséum National d'Histoire Naturelle de Paris, N° 5579c. Selected by (1961c) from d''s surviving syntypes. Argiles à Plicatules of Bailly-aux-Forges, Paris Basin, France.
Material: ERG-18-19, 25-5, 25-7, 27-1, 27-2, 28-4, 28-6, 30-1, 30-9, 30-11, 31-1, 31-4, 31-5, 33-1, 31-7, 31-9, 31-12, 32-1, 32-2, 33-5, 34-10, SANGRE-2, 3, 14, ORL-B-1, 2, 21.
Description: Umbilical wall nearly vertical, umbilical width is between one third and one quarter of the diameter. Whorls sub-rectangular, flanks parallel to convex, venter sub-truncate. Ribs narrow, sigmoidal; primary ribs originate from just above the umbilical wall, secondary ribs commence forward of the primaries at the mid flank, rarely more than one secondary between each pair of primaries in the last whorl. Twenty four primary ribs at umbilical margin and fifty primary and secondary ribs in ventral margin at 33 mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH | PR | SR |
| ERG-33-1 | 44.5 | 20 | 0.45 | 0 | 12 | 0.26 | 0 | |||
| ERG-31-7 | 20 | 9 | 0.45 | 0 | 6 | 0.3 | ||||
| ERG-31-12 | 17.5 | 8 | 0.45 | 0 | 8 | 0.45 | 0 | 20 | 41 | |
| ERG-33-1 | 33 | 16 | 0.48 | 0 | 0 | 0 | 24 | 50 | ||
| SANGRE-3 | 52 | 21 | 0.40 | 0 | 11 | 0.21 | 0 | 24 | 48 | |
| Lectotype | 32 | 14.72 | 0.46 | 9.28 | 0.29 | 10.56 | 0.33 | 0.63 |
Discussion: Because of the similarity in ontogeny in the early stage of growth in Deshayesites and Dufrenoyia, small specimens might show similarity in their flattened venter and effacing ribs in the ventral area. However the presence of Deshayesites in these beds suggests a correct assignation to Deshayesites for these samples. Some specimens are weathered and rib numbers cannot be counted. The Aralar material is comparable to specimens of D. deshayesi in the Natural History Museum, London, numbers C4041, C71943-5, C71447-52.
Occurrence: England (, 1964); Russia ( & , 1960; , 1973); Turkmenistan (, 1977, 1991) and Iran (, 2004).
Distribution: Igaratza section, Errenaga Formation, Eskisabel section, Ataun area, Lareo Formation
(Figs.
6 ![]() -
7
-
7 ![]() ).
).
Deshayesites cf. consobrinoides , 1909
cf. 1909 Parahoplites consobrinoides , p. 3-4.
cf. 1947 Deshayesites consobrinoides d': , p. 170, figs. 18, 14b.
cf. 1961c Deshayesites consobrinoides d': , p. 508, 523, 609.
cf. 1964 Deshayesites consobrinoides : , p. 302, pl. 44, figs. 5-6; pl. 52, fig. 2; text-fig. 106j-n (see for extensive synonyms).
cf. 1971 Deshayesites consobrinoides : , pl. 3, fig. 3.
cf. 1973 Deshayesites consobrinoides : , p. 123, pl. 77, figs. 1-5.
cf. 1979 Deshayesites consobrinoides :
et alii, pl. 3, fig. 3.
cf. 1999 Deshayesites consobrinoides : , pl. 2, fig. 1.
cf. 1999 Deshayesites consobrinoides : & , pl. 6, figs. c-d.
cf. 2000 Deshayesites consobrinoides :
et alii, pl. 3, fig. 1.
cf. 2004 Deshayesites consobrinoides : & , pl. 1, figs. 4-6.
cf. 2004 Deshayesites cf. consobrinoides : , p. 131-132, figs. 4O, P, 5F, G.
Lectotype: Museum, St. Petersburg (one of the specimens collected by , 1898), N° 17727, from the Lower Aptian of Saratov, Russia.
Material: ERG-18-17, 25-3, 26-1, ORL-B-7, 12, 15.
Description: three crushed specimens are to hand. Sides nearly parallel; ribs sharp and strong, sigmoidal near ventral area, primary ribs start from upper part of umbilical wall, secondary ribs are intercalated between primaries.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-18-7 | 28 | 11.5 | 0.41071 | 0 | 7 | 0.25 | 0 | |
| Holotype | 31 | 13 | 0.42 | 9.9 | 0.32 | 9.6 | 0.31 | 0.76 |
Discussion: Deshayesites consobrinoides is characterised by the presence of smooth external sides in the early stages of ontogeny and is very similar to Deshayesites deshayesi, but possesses coarser ribbing. Another feature is that Deshayesites consobrinoides is more strongly evolute than average examples of Deshayesites deshayesi (, 1964).

Click on thumbnail to enlarge the image.
Figure 10: A.- Deshayesites dechyi , 1907, Errenaga Formation, ER-58; B.- Deshayesites cf. callidiscus , 1961c, Errenaga Formation, ATAN-16; C.- Deshayesites cf. normani , 1964, Errenaga Formation, ERG, 12-4; D.- Deshayesites cf. saxbyi , 1964, Errenaga Formation, ER-118; E.- Deshayesites cf. involutus , 1930, Errenaga Formation, ERG-34-8; F.- Deshayesites cf. planus , 1961c, Errenaga Formation, ER-94; G.- Deshayesites cf. multicostatus , 1935, Lareo Formation, SANGRE-2; H.- Deshayesites cf. deshayesi in d', 1841, Errenaga Formation, ERG-31-12; I.- Deshayesites cf. consobrinoides , 1909, Errenaga Formation, ERG-18-7.
Occurrence: Europe (, 1964), Russia (, 1973), Caucasus ( et alii, 1979), Turkmenistan (, 1999) and Iran (, 2004).
Distribution: Igaratza section, Errenaga Formation (Figs.
6 ![]() -
7
-
7 ![]() ).
).
Deshayesites cf. geniculatus , 1964
cf. 1964 Deshayesites cf. geniculatus sp. nov. ; p. 307, pl. 46, fig. 5; pl. 51, fig. 9.
Holotype: British Geological Survey Museum, ZM 1937, Ferruginous Sands, Scaphites Beds, west of Whale Chine, Atherfield, Isle of Wight.
Material: ATAN-29, APAR-18.
Description: Whorl section sub-rectangular, whorl height is more than whorl width, thickest in the upper part of the flank; umbilical wall vertical. Ornament consists of ribs, thick, recurved backwards on the mid flank. Primary ribs start from upper part of the umbilical wall; secondaries are irregular and intercalated between primaries, start from mid to upper one third part of the flank, some fusing with adjacent primaries.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ATAN-29 | 12 | 14 | 1.16 | |||||
| Holotype | 37 | 11.8 | 0.31 | 10.3 | 0.27 | 10.7 | 0.28 | 0.87 |
Discussion: Occasional bifurcation might be a character for the species. D. geniculatus differs from D. wiltshirei , 1964, in its thick and curved ribbing. (1930) described the species as a transition form between D. punfieldensis and D. vectensis , 1930.
Occurrence: England (, 1964).
Distribution: Ataneta locality, Iribas section, Errenaga Formation
(Fig.
6 ![]() ).
).
Deshayesites cf. grandis , 1930
cf. 1930 Deshayesites grandis sp. nov.: , p. 427-429, pl. xvii, fig. 2a-b.
cf. 1961c Deshayesites grandis : , p. 508-510.
cf. 1964 Deshayesites grandis : , p. 308-309, pl. 43, figs, 1a-b; pl. 44, figs. 1-3; pl. 51, fig. 7a-b; text-fig. 110c.
cf. 2010 Deshayesites grandis : et alii, fig. 11J, fig. V, B.
Holotype: British Geological Survey Museum, Geological Society collection 2300, from Atherfield (probably from Bed IV or V), said evidently from the Scaphites Beds (group V) of Atherfield (F. W. collection).
Material: ER-136, ERG-8-3.
Description: Two crushed specimens. Ornament consist of ribs; primary ribs start from upper part of the umbilical wall, feebly curved; secondaries are irregular and intercalated between primaries, start from mid to upper one third part of the flank, most of them attached to primaries; all primary and secondary ribs are flattened and rounded in upper part of the flank.
Discussion: The only character for identification is rib pattern in the studied specimens. (1930) stated that the species is closer to D weissi than to D. consobrinus. The species is common in the type locality and a sub-zone is named on the occurrence of this species. believed that the species appears in the upper part of the deshayesi Zone and even occurs in the early Late Aptian. On the other hand, (1964) mentioned that D. grandis is limited to the deshayesi Zone and its record in the early Late Aptian is due to the confusing of large examples of Dufrenoyia with D. grandis by .
Occurrence: England (, 1930; , 1964).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Deshayesites cf. wiltshirei , 1964
cf. 1964 Deshayesites wiltshirei sp. nov.: , p. 305-306, pl. 53, fig. 4; pl 46, figs. 8-9; pl. 51, fig. 10.
cf. 1999 Deshayesites cf. wiltshirei : , p. 352, fig. 6.
Holotype: Museum, Cambridge, B 27096, Ferruginous Sands, Atherfield, Isle of Wight (T. collection).
Material: ER-77, 81, 89, 101.
Description: Whorls sub-rectangular, sides flattened. Ornament consisting of primary and secondary ribs; primary ribs curved in mid part of the flanks, commence from upper part of umbilical wall; secondary ribs start from upper one third part of the flank, intercalated between primaries mostly free at early whorls and some attach at last half whorl.
Discussion: The rib pattern is a character for identification. There is similarity between D. wiltshirei and D. punfieldensis, but the later species has a wider umbilicus, coarser ribs which are less flattened on the venter. Hitherto, the definitive species has not been recorded outside of England.
Occurrence: England (, 1964); Spain.
Distribution: Igaratza section, Errenaga Formation.
Deshayesites spp.
A large number of crushed and juvenile specimens have been collected. Some of these are perhaps determinable at species level. The only importance of this material is that they are present in the deshayesi-furcata transition Zone discussed below.
Genus Burckhardtites , 1949
Type species: Neocomites nazasensis , 1925
Generic characters: Discoidal, compressed, whorl height and width increasing during ontogeny. Sides and venter flattened with ventro-lateral shoulders. Ornament consists of primary sigmoidal ribs on the whorl flanks; secondary ribs are intercalated or branch from primaries. Ribbing is retrorsive on the lower part of the flank, convex forward near mid flanks and again convex backward on upper third of the flanks. On venter ribs are of uniform relief and nearly straight. Suture line seems to be of parahoplitid type.
Discussion: Fine and irregular ribs are characteristic for the genus. On the body chamber ribbing becomes very sharp. (1949) introduced the genus for all forms of Neocomites nazasensis from the Aptian Rio Nazas, Mexico. However Neocomites disappeared in Hauterivian and it is believed Burckhardtites is more related to Dufrenoyia, than it is to Neocomites ( et alii, 1996; & , 2008). Burckhardtites is probably derived from Paradeshayesites ( & , 2004). Burckhardtites differs from Dufrenoyia by lack of clear tubercles on the ventro-lateral bend, dense ribbing and sharp double S shaped ribs on the flanks.
Occurrence: Mexico (, 1949; & , 2008), Turkmenistan (, 2004).
Burckhardtites sp.
Material: ERG-34-7, 34-22
Description: Whorl section sub-rectangular, venter flat. Ornament consists of ribs, dense, fine and curved on flanks and nearly straight on venter; primary ribs originate from the umbilical margin, some single. Secondary ribs start from one-third of the lower flank or at the mid flank, intercalated and mostly attached to primary ribs. Twenty four primary ribs are present at thirty three mm diameter.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-34-7 | 34 | 15 | 0.44 | 0 | 12 | 0.35 | 0 | |
| ERG-34-22 | 32 | 12 | 0.37 | 0 | 10.5 | 0.32 | 0 |
Discussion: The specimens occur in a deshayesi Zone assemblage. The rib patterns and number are interesting and similar to early deshayesitids. The specimens can be compared to 's figures of Burckhardtites gregoriensis (, 1949, pl. 11, figs. 3-4) and it is suggested that this is the specific identity of the Aralar specimens. This species is recorded also from Turkmenistan ( & , 2004).
For a long time it has been assumed that Burckhardtites is limited to America, but the recent record from Turkmenistan shows that its distribution is more widespread within the Aptian Tethyan belt. The present record might allow us to determine the migration pattern of this genus from west to east of the Tethys.
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Genus Dufrenoyia ex & , 1915
Type species: Ammonites furcatus in , 1836.
Generic characters: Semi involute, whorl section sub-rectangular to oval, with flattened and angular venter, umbilicus relatively wide; costation consists of ribs; primary ribs rise in the umbilicus margin, with light tuberculation, secondary ribs are intercalated, starting at the lower third or middle flank, most are free; ribs cross the venter isometrically, weak or disappearing on venter and forming denticulation on ventro-lateral shoulders. Suture line consists of four lobes, a bifid ventral lobe, an entire umbilical, inner lateral and dorsal lobe.
Discussion: The genus formerly considered to be a parahoplitid is now assigned to Deshayesitidae proposed by (1949). Interruption of ribs in the ventral area is good character for genus level identification. Ribs are thicker than Deshayesites in same diameter and are more regular.
The genus differs from Deshayesites in its flat venter, thicker more regular ribs and tubercles on ventro-lateral shoulder. The genus Kuntziella , 1962, previously known as a subgenus of Deshayesites, shows similarity in some stage of growth with Dufrenoyia, but with higher whorls, flatter sides and without marginal tubercles differs from later genus in the juvenile stage. The Early Albian homoeomorph genus Neodeshayesites (recently assigned to the subfamily Acanthohoplitinae by & (2005) in its early stage of growth resembles Dufrenoyia but in its later stage of growth the venter is flattened with ribbing continuous across the venter with slight to moderate tuberculation at the ventro-lateral margins' Stenhoplites , 1923, is an objective synonym.
Occurrence: Europe, Russia, Turkmenistan, Iran, Japan, USA, Mexico, Venezuela and Colombia (, 1949; , 1949; & , 1960; , 1964; , 1979; et alii, 1996; , 2004; et alii, 2010).
Dufrenoyia cf. furcata (, 1836)
cf. 1836 Ammonites furcatus in , p. 339, pl. xiv, fig. 17.
cf. 1902 Hoplites furcatus : von , p. 202.
cf. 1915 Parahoplites (Duferonyia) furcatus : & , p. 34.
cf. 1923 Stenhoplites furcatus (): , p. 147.
cf. 1925 Dufrenoya furcata (): , p. 17, pl. 10, figs. 12-13.
cf. 1940 Dufrenoya furcata (): , p. 1021.
cf. 1949 Dufrenoya furcata (): , p. 120-121.
cf. 1960 Dufrenoya furcata (): & , tab. 1, fig. 7.
cf. 1964 Dufrenoyia furcata (): , p. 378-382, pl. 62, figs. 2-3; pl. 63, fig. 1; pl. 65, fig. 1a-b; text-figs. 134d, 135-136 (see this reference for full synonyms before 1964).
cf. 1971 Dufrenoyia furcata (): , tab. 51, fig. 10.
cf. 1979 Dufrenoyia furcata (): , tab. 51, fig. 10.
cf. 1982 Dufrenoyia furcata (): , pl. 1, fig. 7.
cf. 1995 Dufrenoyia furcata (): , taf. 2, fig. 2.
cf. 1999 Dufrenoyia furcata (): & , pl. 8, figs. d-e.
cf. 2004 Dufrenoyia furcata (): & , p. 219, pl. 12, figs. 1-3; text-fig. 28.
cf. 2010 Dufrenoyia furcata (): & , p. 129, fig. 1, A-P2, fig. 2, A1-X, AA-AM.
cf. 2010 Dufrenoyia furcata (): et alii, fig. 11, K, M; fig. VI, D-E; fig. VIII, D-E; fig. IX, A.
Holotype: British Geological Survey Museum, Geological society collection, Hythe Beds, Hythe, Kent (W.H. collection).
Material: ERG-34-14.
Description: Shell discoidal, whorl section is sub-rectangular to oval, truncated, forming angular ventro-lateral margins. Ornament consists of ribs; primary ribs rise in the umbilicus margin with light tuberculation; secondary ribs intercalate starting at mid- to upper part of the flank, most are free. Ribs cross the venter isometrically, feebly flattened in the upper part of the flank and ventral area, with feeble tuberculation on the ventral shoulders.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-34-14 | 13.5 | 11 | 0.81 | |||||
| Holotype | 50 | 21 | 0.42 | 17 | 0.34 | 17.5 | 0.35 | 0.80 |
Discussion: The flat venter and flattened ribs in the upper part of the flanks and ventral side are characteristic feature. Dufrenoyia furcata was confused with Dufrenoyia dufrenoyi d' by some authors; full discussion is found in (1964). D. furcata differs from the latter species by possessing less overlapping whorls and coarser, sharp and broader ribs. D. furcata differs from D. lurensis by its more involute whorls and less strong ribs ( & , 2004). The species is well known in Aptian successions in Tethys and a characteristic faunal element for the latest Early Aptian. It has been proposed as a zonal index on account of its geographically widespread occurrence (, 1979, 1983; & , 1994; , 1995; et alii, 1993, 1995, 2003).
Occurrence: England (, 1964), Germany (, 1995), Russia ( & , 1960), Turkmenistan (, 1979, 1999), USA (, 1940; , 1949).
Distribution: Igaratza section, Errenaga Formation (Fig.
7 ![]() ).
).
Dufrenoyia cf. mackesoni , 1964
cf. 1964 Dufrenoyia mackesoni sp. nov.: , p. 397-398, pl. 52, fig. 4a-b; pl. 62, fig. 5a-b; pl. 65, fig. 4a-b; text-figs. 140d, 144.
Holotype: British Geological Survey Museum, 108189, Hythe Beds, Shepway Cross Quarry, Lympne, Kent ( collection).
Material: ERG-33-4, 33-5, 33-7, 34-11, SANGRE-17?
Description: Shell discoid, transverse section is sub-rectangular. Sculpture consists of ribs of narrow and moderate relief, curved; primary ribs start at the umbilical margin; intercalated secondary ribs start from mid-flank, most are free, the ribs form feeble denticulation on the ventral-lateral margin.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| ERG-33-7 | 42.5 | 18 | 0.42 | 0 | 8 | 0.18 | 0 | |
| ERG-34-11 | 42.5 | 18 | 0.42 | 0 | 8 | 0.18 | 0 | |
| Holotype | 64 | 26.24 | 0.41 | 17.9 | 0.2796 | 17.9 | 0.27 | 0.68 |

Click on thumbnail to enlarge the image.
Figure 11: A.- Deshayesites cf. geniculatus , 1964, Errenaga Formation, ATAN-29; B.- Deshayesites cf. grandis , 1930, Errenaga Formation, ER-136; C.- Deshayesites cf. wiltshirei , 1964, Errenaga Formation, ER-77; D.- Burckhardtites sp., Errenaga Formation, ERG-34-7; E-F.- Dufrenoyia cf. furcata (, 1836), Errenaga Formation, ERG-34-14; G.- Dufrenoyia cf. mackesoni , 1964, Errenaga Formation, ERG-34-11; H.- Dufrenoyia cf. lurensis (, 1888), Errenaga Formation, SANGRE-6; I.-J. Toxoceratoides ? sp., Lareo formation, SANGRE-22.
Discussion: The species differs from D. predufrenoy , 1964, by its weaker ribbing. There are similarities also between the D. mackesoni , 1964, and D. stenzeli , 1949, the latter species possessing straighter ribs and a higher umbilical wall.
Occurrence: England (, 1964).
Distribution: Igaratza section, Errenaga Formation (Fig.
6 ![]() ).
).
Dufrenoyia cf. lurensis (, 1888)
cf. 1888 Hoplites lurensis sp. nov.: , p. 681, pl. 20, fig. 2a-b.
cf. 1889 Hoplites furcatus Sowerby var. lurensis: , p. 209.
cf. 1907 Hoplites (Parahoplites) lurensis ; , p. 354, 357.
cf. 1913 Parahoplites lurensis : , pl. 7, fig. 8a-b; pl. 10, fig. 5.
cf. 1925 Dufrenoyia lurensis (): , p. 17.
cf. 1930 Dufrenoyia lurensis (): , p. 436, pl. 15, fig. 4.
cf. 1940 Dufrenoyia lurensis (): , p. 1022.
cf. 1949 Dufrenoyia lurensis (): , p. 121, 124-125, p. 8, figs. 7-8 & 12-13.
cf. 1964 Dufrenoyia lurensis (): , p. 382-383, pl. 63, fig. 2; pl. 64, figs. 3-4; text-figs. 137-140.
Holotype: From the Upper Aptian (Gargasian) of the Carniol (Basses Alpes), France (Sorbonne collection).
Material: SANGRE-6, 7
Description: Whorl section is sub-rectangular, becoming narrower towards the venter. Ornament consists of nearly straight ribs. Primary ribs rise in the upper part of umbilical wall, with light tuberculation. Secondary rib intercalation is rare. Ribbing commences at the lower part of the flank and cross the venter isometrically with ventro-lateral clavi, Shell flattened in the upper part of the flank and across the ventral area.
Measurements:
| Sample Number | D | WH | WH/D | WT | WT/D | U | U/D | WT/WH |
| SANGRE-6 | 62 | 20 | 0.32 | 14.5 | 0.23 | 14.8 | 0.23 | 0.72 |
| SANGRE-7 | 43 | 16 | 0.37 | 0 | 7 | 0.16 | 0 | |
| Holotype | 42 | 15.9 | 0.37 | 15.9 | 0.37 | 15.1 | 0.35 | 1 |
Discussion: The form is similar to D. furcata, but differs from the latter in its more evolute whorls and coarser, less flexuous ribbing and fewer secondary ribs.
Occurrence: England (, 1930; , 1964), Germany (, 1907), France (, 1889), Mexico (, 1925) and USA (, 1940; , 1949).
Distribution: Eskisable section, Ataun area, Lareo Formation (Fig. 6 ![]() ).
).
Ammonites provide one of the most precise biostratigraphical tools for correlating marine Lower Cretaceous sediments. For much of Early Cretaceous time there was a separation into Tethyan and Boreal Realms, with distinct endemic ammonite faunas, which sometimes makes long-distance correlation difficult. However, some ammonite genera are of wide geographical distribution and this helps to resolve some of the correlation problems.
Because the Early Cretaceous ammonite successions of Western Europe have been studied in more detail than those in other areas of the world, the stage stratotypes are situated in this region and the ammonite zones proposed there are normally accepted as the "standard" biozonation. The Aptian Stage boundaries and index faunas in Europe have been discussed most recently by (1996) and by & in et alii (2009). Work by the Lower Cretaceous Cephalopod Team of the SCS has improved the standard biozonation for the Mediterranean area of the Tethyan Realm ( et alii, 1990, 1993, 1995, 2000, 2003; et alii, 1999, et alii, 2006, 2009) (Table 1). In this section of the paper, the current division and biozonation of the Early Aptian sub-Stage will be discussed and an attempt is made to correlate successions throughout the whole of the northern Mediterranean Tethyan belt, stretching eastward to Iran.
The Early Aptian ammonite zonation used here is that refined for the northern region of the Tethyan province ( et alii, 2003; et alii, 2006, 2009) (Table 1) and is relevant to the Spanish succession in Spain. This scheme embraces some of the Early Aptian deshayesitid zones first established in Mangyschlak (, 1971; & , 1994; , 1968, 1980). A different ammonite zonation has been proposed by (1961c) and et alii (1998) for the succession in the Lower Greensand Group of southern and eastern England. This region belongs to a more Boreal province, but his scheme is applicable more widely in northern Europe and Greenland (e.g., , 1999) with some local modifications (e.g., , 1976). Both schemes for the Early Aptian reflect the occurrence of ammonite genera and species, particularly deshayesitids that are inter-provincial in their geographical range (e.g., , 1981; , 1988; , 1995, 1999; et alii, 1971; et alii, 1997; , 2004). The correlation of 's scheme (, 1961c; et alii, 1998) with that of the Mediterranean Tethys has been discussed by & in et alii (2009) and is shown in Table 2.
(1961c) proposed a biozonation for the Early Aptian of England. In ascending order, the earliest Zone is that of Prodeshayesites fissicostatus divided into an earlier Subzone of P. bodei and a later P. obsoletus Subzone. These two subzones have been reported also from East Greenland ( & , 1999). (1995) considered that the species of Prodeshayesites are similar to species of Deshayesites reported from the Deshayesites tuarkyricus Zone of Turkmenistan by (1971, 1983). He considered Prodeshayesites to be a synonym of Deshayesites and this view is accepted here. Moreover it seems that the genera Paradeshayesites and Obsoleticeras of & (1999) are subjective synonyms of Deshayesites. & (1994) correlated the Deshayesites tuarkyricus Zone (now Deshayesites oglanlensis Zone) in Turkmenistan with the fissicostatus Zone in England and the tenuicostatus Zone in Germany. (2002) considered the geographical occurrence of D. tuarkyricus to be restricted to Mangyschlak (Turkmenistan) and Transcaspia. Therefore he suggested that the more geographically widespread species D. oglanlensis to be a more suitable index fossil for this Zone and this has been accepted ( et alii, 2003; et alii, 2006). Recently, D. tuarkyricus has been recorded in the Errenaga Formation in the Aralar Mountains ( et alii, 2009).
(1961c) introduced a Zone of D. forbesi and divided it into four subzones; D. fittoni, D. kiliani, D. callidiscus and D. annelidus in ascending order. The Deshayesites weissi Zone was proposed for the roughly equivalent time period in the rest of Europe and the Mediterranean Tethyan belt. The weissi Zone has been identified in north Germany, Romania, the Balkans, Tuarkyr and Kopet Dagh ( & , 1994). (1999) believed the forbesi and weissi zones are only partially coeval and that the weissi Zone commences with the callidiscus Subzone of continuing to the base of the grandis Subzone of the deshayesi Zone of .
The author (in et alii, 2006) considered that D. weissi is a nomen dubium and suggested, either to find a type from specimen of the collection in Berlin (Germany) or to replace D. weissi with Deshayesites planus in the Standard zonation. These two species are found together in England as well as the rest of Europe in sediments of this age. This suggestion is under review.
The D. deshayesi Zone of (1961c, 1964) and et alii (1998) is accepted by most authors (e.g., , 1971, 1979; & , 1994; & , 1999; & , 1960; & , 1979; , 1991; , 1995, 1999; , 1983; , 1996). (1961c), et alii (1998) suggested two subzones; an earlier Cheloniceras (Cheloniceras) parinodum , 1961b, and a later D. grandis. The co-occurrence of the first appearance of D. deshayesi and Cheloniceras (Cheloniceras) could be useful in the recognition of the weissi and deshayesi zones boundary. & (1999) considered the deshayesi Zone in Transcaspia could be correlated with upper part of the English deshayesi Zone.
Dufrenoyia is a widely distributed genus and D. furcata is the index fossil for the latest part of the Lower Aptian in the Mediterranean region. (1961a) suggested Tropaeum bowerbanki as the index species for the latest zone of the Lower Aptian in southern England, but Dufrenoyia furcata also occurs in that zone and Tropaeum is facies restricted.
The vertical distribution of ammonites in the Errenaga and Lareo formations is shown in the range charts
(Figs. 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() ). A representative sample of the ammonite fauna from the Aralar sections is illustrated in
Figs. 5
). A representative sample of the ammonite fauna from the Aralar sections is illustrated in
Figs. 5 ![]() - 9
- 9 ![]() - 10
- 10 ![]() - 11
- 11 ![]() .
.
In the Errenaga Formation of Iribas section typical fauna of the oglanlensis Zone has been collected in-situ (assemblage 10,
Fig. 3 ![]() ). The assemblage ammonite fauna (IRI-2 to 134) includes: D. cf. oglanlensis ,
1983, D. cf. tuarkyricus ,
1983, D. weissiformis ,
1983, D. luppovi ,
1983, D. dechyi ,
1907, and D. cf. antiquus ,
1983.
). The assemblage ammonite fauna (IRI-2 to 134) includes: D. cf. oglanlensis ,
1983, D. cf. tuarkyricus ,
1983, D. weissiformis ,
1983, D. luppovi ,
1983, D. dechyi ,
1907, and D. cf. antiquus ,
1983.
The lowest occurrence of ammonites in the Errenaga Formation is in the Igaratza section at approximately one m above the base of the unit (assemblage 6,
Fig. 3 ![]() ) and consists of an indeterminate Deshayesites. A zonal age cannot be determined, but the basal Errenaga Formation at Igaratza is Early Aptian and there is no evidence of the presence of Barremian sediments. The interval corresponds probably to the prolific fauna of D. oglanlensis Zone age found at Iribas.
) and consists of an indeterminate Deshayesites. A zonal age cannot be determined, but the basal Errenaga Formation at Igaratza is Early Aptian and there is no evidence of the presence of Barremian sediments. The interval corresponds probably to the prolific fauna of D. oglanlensis Zone age found at Iribas.
No ammonites have been found in the lower part of the Errenaga Formation in the Ataun sections but an oglanlensis Zone age is probable on sedimentary subunit correlation evidence
(Fig. 3 ![]() ). Higher in the lower part of the Formation, assemblage 1
(Figs. 3
). Higher in the lower part of the Formation, assemblage 1
(Figs. 3 ![]() - 4
- 4 ![]() ) at 185.0 m above the base of the section, contains a poorly preserved fauna (IMA-A-1 to 14) with D. cf. weissiformis and is probably of oglanlensis Zone age.
) at 185.0 m above the base of the section, contains a poorly preserved fauna (IMA-A-1 to 14) with D. cf. weissiformis and is probably of oglanlensis Zone age.
In the Aralar district, an oglanlensis Zone fauna is present at the lower part of the Errenaga Formation. The genus Procheloniceras , 1923, occurs in the oglanlensis Zone throughout southern Europe. Although absent in the Aralar district, it has been recorded in sediments of this age in the Betic Cordillera of southern Spain ( et alii, 1997). This is of significance when considering the fauna of the succeeding D. weissi Zone at Aralar, discussed next.
The best assemblage of ammonites is to be found in the Iribas and Igaratza sections
(Fig. 7 ![]() ). At Iribas, a precise boundary between oglanlensis and weissi Zone faunas occurs at 23.85 m above the base of the unit (assemblage 11,
Figs. 3
). At Iribas, a precise boundary between oglanlensis and weissi Zone faunas occurs at 23.85 m above the base of the unit (assemblage 11,
Figs. 3 ![]() - 4
- 4 ![]() ). The weissi Zone material in this interval (ATAN-2 to 32, URG-1 & 2) contains D. cf. weissi & ,
1881, D. weissiformis ,
1983, D. cf. planus ,
1961c, D. cf. callidiscus ,
1961c, D. dechyi ,
1907, D. cf. geniculatus , 1964, D. cf. multicostatus ,
1935, and D. forbesi ,
1961c. A laterally equivalent outcrop situated 1 km West has yielded Roloboceras cf. regale ,
1961c, and Aconeceras (Aconeceras) cf. nisoides (,
1893), indicating a weissi Zone age (assemblage 12,
Figs. 3
). The weissi Zone material in this interval (ATAN-2 to 32, URG-1 & 2) contains D. cf. weissi & ,
1881, D. weissiformis ,
1983, D. cf. planus ,
1961c, D. cf. callidiscus ,
1961c, D. dechyi ,
1907, D. cf. geniculatus , 1964, D. cf. multicostatus ,
1935, and D. forbesi ,
1961c. A laterally equivalent outcrop situated 1 km West has yielded Roloboceras cf. regale ,
1961c, and Aconeceras (Aconeceras) cf. nisoides (,
1893), indicating a weissi Zone age (assemblage 12,
Figs. 3 ![]() - 4
- 4 ![]() ).
).
Igaratza has provided the most important ammonite occurrence series within the weissi Zone
(Fig. 7 ![]() ). The fauna includes Deshayesites cf. bodei (von ,
1902), D. weissiformis ,
1983, D. cf. euglyphus , 1964, D. cf. planus ,
1961c, D. cf. callidiscus ,
1961c, D. luppovi ,
1983, D. dechyi ,
1907, D. forbesi ,
1961c, D. cf. multicostatus ,
1935, D. cf. saxbyi , 1964, D. cf. gracilis ,
1930, D. cf. punfieldensis ,
1930, D. cf. consobrinus
(d', 1841), D. cf. grandis ,
1930, D. cf. normani , 1964, Pseudosaynella sp. and Roloboceras cf. hambrovi (,
1845); a typical weissi Zone fauna.
). The fauna includes Deshayesites cf. bodei (von ,
1902), D. weissiformis ,
1983, D. cf. euglyphus , 1964, D. cf. planus ,
1961c, D. cf. callidiscus ,
1961c, D. luppovi ,
1983, D. dechyi ,
1907, D. forbesi ,
1961c, D. cf. multicostatus ,
1935, D. cf. saxbyi , 1964, D. cf. gracilis ,
1930, D. cf. punfieldensis ,
1930, D. cf. consobrinus
(d', 1841), D. cf. grandis ,
1930, D. cf. normani , 1964, Pseudosaynella sp. and Roloboceras cf. hambrovi (,
1845); a typical weissi Zone fauna.
At a height of 256.0 m above the base at Ataun, IMA-1 to 17 (assemblage 2, Figs. 3 ![]() - 4
- 4 ![]() ) is of definite weissi Zone age and includes D. forbesi, Aconeceras (Aconeceras) cf. nisoides, Aconeceras (Theganoceras) sp.
) is of definite weissi Zone age and includes D. forbesi, Aconeceras (Aconeceras) cf. nisoides, Aconeceras (Theganoceras) sp.
In the Igaratza section, within a thickness of 20 m, the ammonite occurrences indicate successively the first appearance of the D. weissi Zone fauna, a typical D. weissi Zone assemblage, the D. weissi-D. deshayesi
zones boundary and a relatively thin development of the D. deshayesi Zone. The distribution of the ammonites is shown in
Fig.
7 ![]() .
.
The view that the weissi and forbesi zones are coeval ( & , 1994) is supported by the occurrences within the weissi Zone fauna in the Aralar area. This fauna includes: D. cf. forbesi , D. cf. planus , D. cf. callidiscus , D. luppovi , D. dechyi together with species of Roloboceras including R. cf. hambrovi (), and R. cf. regale . The occurrences of ammonite horizons in the Errenaga Formation are few and widely separated and thus, there are gaps in our knowledge of the ammonite succession in the Aralar area. However, in England, the genus Roloboceras is replaced by Cheloniceras (Cheloniceras) at the base of the deshayesi Zone ( et alii, 1998). Based on above intrepretation the Aralar weissi Zone could be equivalent of the D. callidiscus Subzone only.
A precise sediment boundary between the weissi and succeeding deshayesi zones is only recognisable at Igaratza
(Fig.
7 ![]() ) where assemblage ERG-17 is of weissi Zone age and ERG-18, 0.29 m above at 100.13 m above the base of the section is of deshayesi Zone age. At Ataun, the boundary between sediments of the weissi and deshayesi zones cannot be given precision in the lack of ammonite evidence.
) where assemblage ERG-17 is of weissi Zone age and ERG-18, 0.29 m above at 100.13 m above the base of the section is of deshayesi Zone age. At Ataun, the boundary between sediments of the weissi and deshayesi zones cannot be given precision in the lack of ammonite evidence.
At Iribas, it is possible that the deshayesi Zone is represented in 2 metres of sediment upon equally condensed weissi Zone sediments if differences in subsidence and correlations of lithological subunits between the Iribas and Igaratza sections are taken into account
(Fig. 4 ![]() ).
).
In the well-documented section at Igaratza (Fig.
7 ![]() ), D. cf. deshayesi appears as a rarity at 100.13 m (ERG-18-19) above the base of the section and then more commonly up to 104.53 m above the base. This interval of 4.4 m is classified only with the deshayesi Zone. The ammonites from this Zone at Igaratza (ERG-18-19 to ERG-28-10) include: D. cf. deshayesi in d', 1841, D. cf. callidiscus ,
1961c, D. cf. consobrinus (d', 1841), D. cf. consobrinoides ,
1909, D. cf. planus ,
1961c, D. dechyi ,
1907, D. cf. multicostatus ,
1935, D. weissiformis ,
1983, D. cf. involutus ,
1930, and in the lower part of the succession include elements that have survived from the underlying weissi Zone.
), D. cf. deshayesi appears as a rarity at 100.13 m (ERG-18-19) above the base of the section and then more commonly up to 104.53 m above the base. This interval of 4.4 m is classified only with the deshayesi Zone. The ammonites from this Zone at Igaratza (ERG-18-19 to ERG-28-10) include: D. cf. deshayesi in d', 1841, D. cf. callidiscus ,
1961c, D. cf. consobrinus (d', 1841), D. cf. consobrinoides ,
1909, D. cf. planus ,
1961c, D. dechyi ,
1907, D. cf. multicostatus ,
1935, D. weissiformis ,
1983, D. cf. involutus ,
1930, and in the lower part of the succession include elements that have survived from the underlying weissi Zone.
The sediments of the D. deshayesi Zone typically are thinly represented in the upper part of the Errenaga Formation
(Fig. 4 ![]() ). I take the base of the Zone at the first appearance of D. deshayesi, but it is difficult determination of specimens in the upper part of the Errenaga Formation and in the Lareo Formation above the Sarastarri Limestone due to the co-occurrence of Deshayesites and Dufrenoyia
(Figs. 6
). I take the base of the Zone at the first appearance of D. deshayesi, but it is difficult determination of specimens in the upper part of the Errenaga Formation and in the Lareo Formation above the Sarastarri Limestone due to the co-occurrence of Deshayesites and Dufrenoyia
(Figs. 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() ).
Elsewhere in Europe, sediments of the deshayesi Zone are characterised by species of Deshayesites only and the transitions to Dufrenoyia are absent. Sediments of the succeeding Dufrenoyia furcata Zone yield Dufrenoyia only, Deshayesites being absent
(e.g., , 1960; et alii,
1998). This suggests that the earliest part of the furcata Zone with its transitional fauna is not represented by sediments over much of the European region although detailed collecting in the Vocontian Basin (France) and the Cau Section (SE Spain) ( et alii,
1997) may show a similar transitional interval. Thus, the Aralar succession in Europe is currently unique and reflects the extremely rapid accumulation of sediments during this relatively brief period of time.
).
Elsewhere in Europe, sediments of the deshayesi Zone are characterised by species of Deshayesites only and the transitions to Dufrenoyia are absent. Sediments of the succeeding Dufrenoyia furcata Zone yield Dufrenoyia only, Deshayesites being absent
(e.g., , 1960; et alii,
1998). This suggests that the earliest part of the furcata Zone with its transitional fauna is not represented by sediments over much of the European region although detailed collecting in the Vocontian Basin (France) and the Cau Section (SE Spain) ( et alii,
1997) may show a similar transitional interval. Thus, the Aralar succession in Europe is currently unique and reflects the extremely rapid accumulation of sediments during this relatively brief period of time.
The ammonite fauna in the Igaratza section includes Dufrenoyia mackesoni , 1964, which first appears in the succession at 104.53 m above the base, where it is associated with species of Deshayesites. A similar association of ammonites occurs a little higher in the section (ERG-34)
(Fig. 4 ![]() ) and in the Lareo Formation throughout the Aralar region. Such an association is unique in the known sections in Europe spanning the deshayesi-furcata zones boundary. Elsewhere in Europe, there is a distinct changeover in the ammonite fauna at the base of furcata Zone sediments with Dufrenoyia replacing Deshayesites. The rapid sedimentation in the Aralar region reflecting the Biscay marginal subsidence has preserved the transition between the two zones. Dufrenoyia is clearly related to Deshayesites, but hitherto, the transitional forms have not been found.
) and in the Lareo Formation throughout the Aralar region. Such an association is unique in the known sections in Europe spanning the deshayesi-furcata zones boundary. Elsewhere in Europe, there is a distinct changeover in the ammonite fauna at the base of furcata Zone sediments with Dufrenoyia replacing Deshayesites. The rapid sedimentation in the Aralar region reflecting the Biscay marginal subsidence has preserved the transition between the two zones. Dufrenoyia is clearly related to Deshayesites, but hitherto, the transitional forms have not been found.
| Sub-Stage | Standard biozonation | Proposed Biozonation (Aralar) |
Assemblage fauna | ||
| SE England (, 1961c; , 1991) |
Mediterranean Region ( et alii, 2003; et alii, 2009) |
||||
| Lower Aptian | Deshayesites deshayesi | Deshayesites deshayesi | Deshayesites cf. deshayesi - Dufrenoyia cf. furcata transition |
Deshayesites and Dufrenoyia coexistence | Aconeceras cf. nisoides, Aconeceras haugi, Burckhardtites sp., Deshayesites cf. consobrinus,D. cf. consobrinoides, D. cf. deshayesi, cf. geniculatus, D. cf. involutus, D. cf. multicostatus, D. cf. normani, D. cf. punfieldensis, D. spp., Dufrenoyia cf. mackesoni, Dufrenoyia cf. furcata, Hemihoplites sp., and Toxoceratoides sp. |
| Deshayesites forbesi | Deshayesites weissi |
Deshayesites cf. deshayesi | Aconeceras haugi, Aconeceras cf. nisoides, Deshayesites cf. callidiscus, D. cf. consobrinus, D. cf. consobrinoides, D. dechyi, D. cf. deshayesi, D. forbesi, D. cf. involutus, D. cf. multicostatus, D. cf. planus, D. weissiformis, D. spp., and Roloboceras cf. hambrovi | ||
| Deshayesites cf. weissi | Aconeceras (Aconeceras) cf. nisoides, Deshayesites cf. bodei, D. cf. callidiscus, D. cf. consobrinus, D. dechyi, D. cf. euglyphus, D. forbesi, D. cf. geniculatus, D. cf. gracilis, D. cf. grandis, D. luppovi, D. cf. multicostatus, D. cf. normani, D. cf. planus, D. cf. punfieldensis, D. cf. weissi, D. weissiformis, D. cf. saxbyi, Pseudosaynella sp., and Roloboceras cf. regale | ||||
| Prodeshayesites fissicostatus | Deshayesites oglanlensis |
Deshayesites cf. oglanlensis | Deshayesites cf. oglanlensis, D. cf. tuarkyricus, D. weissiformis, D. luppovi, D. dechyi, and D. cf. antiquus, Pseudosaynella sp., Aconeceras (Theganoceras) sp., and Roloboceras cf. hambrovi | ||
Table 2: Comparison of the Early Aptian ammonite biozonation in SE England, Mediterranean Region and Aralar (N Spain).
The Aralar succession provides this transitional interval in a very thick sediment succession. Within the Errenaga succession at Igaratza, Dufrenoyia furcata (,
1836), the zonal index of the furcata Zone, appears in ERG-34 at 104.82 m above the base of the section associated with Deshayesites cf. deshayesi in d', 1841, D. cf. involutus ,
1930, and Burkhardites sp. among others listed in
Fig.
7 ![]() . The Errenaga Formation was deposited in an open shelf environment with a local
basinward polarity from east to west. Towards the top of the Errenaga Formation in the Deshayesites deshayesi-Dufrenoyia furcata zones transition, a new regressive episode is deduced from the emplacement of shallow-water carbonate platform facies of the Sarastarri Formation
(Fig. 4
. The Errenaga Formation was deposited in an open shelf environment with a local
basinward polarity from east to west. Towards the top of the Errenaga Formation in the Deshayesites deshayesi-Dufrenoyia furcata zones transition, a new regressive episode is deduced from the emplacement of shallow-water carbonate platform facies of the Sarastarri Formation
(Fig. 4 ![]() ).
).
The co-occurrence of Deshayesites and Dufrenoyia in this transitional interval might eventually be recognised as a distinct Subzone, to be indexed when more extensive ammonite material is available. At the top of the Errenaga Formation at Igaratza Dufrenoyia is rare and does not become more common until the Lareo Formation above.
The lower boundary of the furcata Zone is taken at the base of the Lareo Formation. This would probably be the boundary in other areas of more incomplete ammonite record. Strictly, its base should be taken at its first appearance in the Igaratza section in sample ERG-33 at 104.53 m. Elsewhere in Europe and Iran, Deshayesites does not occur in the furcata Zone ammonite fauna (e.g., , 1961c; et alii, 1998; , 2004). The succession in the Vocontian Basin in France (e.g., et alii, 1997), Switzerland ( et alii, 2001) and Spain ( et alii, 1997; et alii, 2001) for this interval is not controlled by ammonites as precisely as in Aralar ( et alii, 2009).The co-occurrence of Deshayesites and Dufrenoyia in this transitional interval might eventually be recognised as a distinct Subzone, to be indexed when more extensive ammonite material is available.
The first Deshayesitidae (Turkmeniceras) appeared in the Late Barremian of the restricted region of the Kopet Dagh and Turkmenistan, while its descendant, Deshayesites, spread to the Caucasus, Europe and even Greenland.
The Lower Aptian ammonite biozonation based on species of Deshayesites and its descendent genus Dufrenoyia in north Spain discussed here is widely applicable throughout the Mediterranean Tethyan belt from northern Spain eastward to the Kopet Dagh region of Iran and Turkmenistan. The assemblage fauna in the D. oglanlensis Zone of the Aralar Mountains resembles that of France and the Kopet Dagh. Moreover this Tethyan Zone can be correlated with the Prodeshayesites fissicostatus Zone of northern Europe proposed by (1961c). The next ascending ammonite biozone is based on the stratigraphic distribution of Deshayesites weissi and Deshayesites planus. The weissi Zone has long been established, however D. planus shows a greater geographical dispersion than D. weissi. The next ascending Zone is based on the widespread occurrence of D. deshayesi and is a long established zonal index species for the later Early Aptian. It is capable of widespread recognition in the European shelf seas as well as in the Tethyan belt. The successor of Deshayesites, Dufrenoyia also has a cosmopolitan distribution. The genus is more widely distributed than Deshayesites, especially to the west, where it occurs also in North America and the northern part of South America where Deshayesites is absent or recorded only questionably. Deshayesites species in north Spain show the closest similarity to those species recorded from Turkmenistan and Iran (Kopet Dagh), France, Romania and Caucasus. Among twenty identified species from north Spain, twelve species are common with Iran and Turkmenistan (Kopet Dagh), and nearly the same number of species with England, whereas seven species are shared with France and Romania (Table 3). This indicates good marine connections permitting the widespread distribution of these species and enabling the palebiogeographic interpretation of the Mediterranean Province.
The Lower Aptian in Aralar is subdivided into the Errenaga, Sarastarri and Lareo formations. Representative species of the following genera: Aconeceras, Pseudosaynella, Toxoceratoides? sp., Hemihoplites, Roloboceras, Deshayesites, Dufrenoyia are identified and described for first time from study area. Based on ammonite assemblage fauna D. oglanlensis, D. weissi, D. deshayesi biozones and an important D. deshayesi-D. furcata transitional interval in which Deshayesites and Dufrenoyia co-exist is suggested. The latter is currently unique to the Aralar succession and has important implications for the current global biostratigraphical correlations which need thorough revision.
The Deshayesites species of the Aralar Mountains resembles that of France, Romania, Caucasus, Kopet Dagh and England. It is capable of widespread recognition in the Tethyan belt as well as in the European shelf seas. This indicates good marine connections in above areas and could use in the paleobiogeographic interpretation of the Mediterranean Province.
| Deshayesites species | Spain | France | Romania | Caucasus | Turkmenistan | Iran (Kopet Dagh) |
England |
| D. bodei | x (cf.) | x | --- | ||||
| D. callidiscus | x (cf.) | x | x | --- | x | ||
| D. consobrinoides | x (cf.) | x | x | x (cf.) | x | ||
| D. dechyi | x (cf.) | ? | --- | x | x | x (cf.) | --- |
| D. deshayesi d' | x (cf.) | x | x | x | x | x | |
| D. euglyphus | x (cf.) | x | --- | ? | --- | x (cf.) | x |
| D. forbesi | x (cf.) | x | ? | --- | --- | x | |
| D. geniculatus | x (cf.) | --- | x | ||||
| D. gracilis | x (cf.) | x | --- | x | |||
| D. grandis | x (cf.) | ? | --- | --- | --- | --- | x |
| D. involutus | x (cf.) | ? | x | --- | --- | x (cf.) | x |
| D. luppovi | x (cf.) | x | x | ? | x | x | --- |
| D. multicostatus | x (cf.) | x? | x | x | --- | x (cf.) | x |
| D. oglanlensis | x (cf.) | x | x | ? | x | x | --- |
| D. planus | x (cf.) | ? | x | --- | x | x (cf.) | x |
| D. punfieldensis | x (cf.) | ? | --- | --- | --- | --- | x |
| D. saxbyi | x (cf.) | --- | x | ||||
| D. tuarkyricus | x (cf.) | --- | --- | --- | x | x (cf.) | --- |
| D. weissi & | x (cf.) | x | x | x | x | x | ? |
| D. weissiformis | x (cf.) | x | x | ? | x | x (cf.) | --- |
Table 3: Comparison of Deshayesites species of the north Spain and other areas in the Mediterranean Province.
I thank Dr Hugh (Natural History Museum London) for his valuable comments during my research on these ammonites and the manuscript. I thank Prof. and Dr (both from Pais Vasco University, Bilbao, Spain), for help in sampling, stratigraphy and drawing of first drafts of some figures. I would wish to thank Phil of the Natural History Museum, London, for the excellent photographs of difficult material. I am indebted to Dr H. (Natural History Museum, London), Dr O. (Natural History Museum, Budapest) and P. (Université de Provence, Aix-Marseille) who acted as referees, for their useful comments. I also thank Dr. Christian C. for his help to edit the manuscript. This research has been supported by the University of the Basque Country Project BTE2003-04823, and the Spanish Science-Education Ministry Project CGL2006-12518 and is greatly appreciated.
R., M., J. & J.M. (1997).- Biostratigraphic events at the Barremian / Aptian boundary in the Betic Cordillera, southern Spain.- Cretaceous Research, London, vol. 18, n° 3, p. 309-329.
M.B. (2002).- Hemihoplitid ammonoids from the Austral Basin of Argentina and Chile.- Abhandlungen Geologisthen Bundesantalt, Wien, Band 57, p. 491-500.
F. & B. (2004).- Les ammonites aptiennes des argiles à Plicatules de la bordure orientale du bassin de Paris.- Association Géologique Auboise, Bulletin annuel, Sainte-Savine, n° 24-25, p. 76-80.
D.J. (1899).- Über die Kreidefossilien des Kaukasus.- Beiträge zur Paläontologie und Geologie von Österreich-Ungarns und des Orients, Wien, vol. XII, p. 53-159.
W.J. (1947).- The geology of the country around Weymouth, Swanage, Corfe and Lulworth (explanation of sheets 341, 342, 343, with small portions of Sheets 327, 328, 329).- Memoir of Geological Survey of the Great Britain - England and Wales, London, 386 p.
W.J., B. & W.C. (1957).- Mesozoic Ammonoidea. In: R.C. (Ed.), Mollusca.- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, Vol. 4, p. L80-L437.
E. (1999).- The Deshayesites , 1914 (Ammonoidea) representatives in Romania, a link between the West-European and Caspian assemblages of this genus. In: F. & F. (eds.), Advancing Research on Living and Fossil Cephalopods.- Kluwer Academic/Plenum Press, New York, p. 437-462.
E.J. & I.A. (2002).- Новая стратиграфическая схема нижнего апта Среднего Поволжья [New stratigraphical scheme of the Lower Aptian of Middle Povolzhie].- Stratigrafiya, Geologicheskaya Korrelyatsiya (Stratigraphy and Geological Correlations), Moscow, vol. 10, n° 6, p. 100-123 [in Russian].
R. & F.J.-M.R. (2008).- Lower Aptian (Lower Cretaceous) ammonites from the basal strata of the La Peña Formation of Nuevo León State, Northeast Mexico: biochronostratigraphic implications.- Revista mexicana de Ciencias geológicas, Mexico, vol. 25, n° 1, p. 145-157.
T.N. (1971).- Lower Aptian and boundary beds of west and south Turkmenia (Stratigraphy, ammonites).- Avtorefera Dissertatii na soiskanie uchenoi stepenni kandidata geologo-mineralogicheskih nauk, 30 p. [in Russian].
T.N. (1977).- On some Deshayesites of western Turkmenia.- Ezhegodnik Vsesojuznogo Paleontologicheskogo Obshchestva (Akademiya Nauk SSSR), Leningrad, vol. 19, p. 46-70 [in Russian].
T.N. (1979).- Аммониты семейства Deshayesitidae Туркмении [Ammonites of the Family Deshayesitidae of Turkmenia]. In: T.N. (ed.), Plankton I organicheskii mir pelagiali v istorii Zemli [Planktonic and organic world of the pelagic zone in the history of the earth].- Trudy XIX sessii Vsesoyuznogo Paleontologicheskogo Obshchestva [Proceeding of the 19th session of the All-Union Paleontological Society], Nauka, Leningrad, p. 152-170 [in Russian].
T.N. (1983).- Зона Deshayesites tuarkyricus - нижняя зона апта Туркмении [Deshayesites tuarkyricus Zone - The lower zone of the Aptian in Turkmenia].- Ezhegodnik Vsesojuznogo Paleontologicheskogo Obshchestva (Akademiya Nauk SSSR), Leningrad, vol. 26, p. 128-147 [in Russian].
T.N. (1991).- Новые виды аммонитов из нижнего апта Туркмении [New Lower Aptian ammonite species of Turkmenia].- Ezhegodnik Vsesojuznogo Paleontologicheskogo Obshchestva (Akademiya Nauk SSSR), Leningrad, vol. 34, p. 77-98 [in Russian].
T.N. (1999).- The Lower Aptian of the Mangyshlak Mountains. Stratigraphy and Geological Correlation.- Stratigrafiya, Geologicheskaya Korrelyatsiya (Stratigraphy and Geological Correlations), Leningrad, vol. 7, p. 343-355 [in Russian].
T.N., I.V. & M.Z. (1979).- Некоторые раннеаптские дегезиты Центрального Дагестан [Some Early Aptian Deshayesitidae from central Daghestan].- Geologica Balcanica, Sofia, vol. 9, n° 3, p. 3-12 [in Russian, with English abstract].
T.N. & I.A. (1999).- Происхождение и развитие семейства Deshayesitidae (Ammonoidea) [Origin and evolution of the Family Deshayesitidae (Ammonoidea)].- Paleontological Journal, Moscow, vol. 33, n° 5, p. 522-529.
T.N. & I.A. (2004).- Origin, evolution and stratigraphic significance of the Superfamily Deshayesitaceae .- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, (Science de la Terre), vol. 74, p. 189-243.
T.N. & V. (1999).- Substantiation of the Barremian/Aptian boundary. In: P. & P.J. (eds.), Proceedings 4th International Workshop of the Lower Cretaceous Cephalopod Team (IGCP-Project 362).- Scripta Geologica, Leiden, Special Issue n° 3, p. 45-81.
T. & S.Z. (1994).- On development of the Aptian Ammonite zonal standard for the Mediterranean Region.- Géologie Alpine, Grenoble, Mém. H.S., n° 20, p. 51-59.
C. (1925).- Faunas del Aptiano de Nazas (Durango).- Boletín del Instituto Geológico de México, n° 45, 71 p.
R. (1984).- Crétacé inférieur : 1.3.1. Ammonites.- In: S. (éd.), Synthèse Géologique du Sud-Est de la France. I : Stratigraphie et paléogéographie.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, n° 125, p. 292-294.
R. (1954).- Falciferella, a new genus of Gault ammonites, with a review of the family Aconeceratidae in the British Cretaceous.- Proceedings of the Geologists' Association, London, vol. 65, n° 3, p. 262-277.
R. (1957).- The Cretaceous ammonite genus Leymeriella, with a systematic account of its British occurrences.- Palaeontology, Oxford, vol. 1, p. 29-59.
R. (1960).- The Ammonoidea of the Lower Greensand. Part 1.- Monograph of the Palaeontographical Society, London, vol. 113, n° 490, xxxvi + 44 p.
R. (1961a).- The Ammonoidea of the Lower Greensand. Part 2.- Monograph of the Palaeontographical Society, London, vol. 114, n° 493, p. 45-118.
R. (1961b).- The Ammonoidea of the Lower Greensand. Part 3.- Monograph of the Palaeontographical Society, London, vol. 115, n° 495, p. 119-216.
R. (1961c).- The stratigraphical palaeontology of the Lower Greensand.- Palaeontology, London, vol. 3, part 4, p. 487-621.
R. (1964).- The Ammonoidea of the Lower Greensand. Part 5.- Monograph of the Palaeontographical Society, London, vol. 117 (1963), n° 502, p. 289-398.
R. (1965).- The Ammonoidea of the Lower Greensand. Part 6.- Monograph of the Palaeontographical Society, London, vol. 118, n° 506, p. 399-546.
R., H.M. & M.I. (1998).- Observations on the lithostratigraphy and ammonite succession of the Aptian (Lower Cretaceous) Lower Greensand of Chale Bay, Isle of Wight, UK.- Cretaceous Research, London, vol. 19, n° 4, p. 511-535.
J.M., M., G.A. de & R. (2001).- Biostratigraphy of the Aptian - Middle Cenomanian platform to basement domain in the Prebetic Zone of Alicante, SE Spain: Biostratigraphy of the AptianMiddle Cenomanian platform to basin domain in the Prebetic Zone of Alicante, SE Spain: calibration between shallow water benthonic and pelagic scales.- Cretaceous Research, London, vol. 22, n° 2, p. 145-156.
F., A.V. & T.N. (1999a).- The Aptian stratigraphy of the southern Tuarkyr (NW. Turkmenistan, Central Asia).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 105, n° 3, p. 377-396.
F., P. & R. (1999b).- The appearance of the genus Deshayesites (, 1914, Ammonoidea) in the lowermost Aptian (Lower Cretaceous) of La Bédoule (SE France).- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 105, n° 2, p. 267-276.
M. (1962).- Atlas des fossiles de Madagascar (Ammonites).- Service Géologique - Ministère de l'Industrie et des Mines, Tananarive, fasc. IX (Aptien), 64 p., pls. CCXV-CCXL.
M., A., B. & J. (1979).- Faunes d'ammonites de l'Aptien de la région de Santander (Espagne).- Annales de Paléontologie, Invertébrés, Paris, vol. 65, fasc. 2, p. 139-156.
G. (1990).- Données nouvelles sur l'espèce-index Hemihoplites feraudianus (d', 1841) (Ammonoidea, Ancyloceratina).- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 310, p. 661-666.
G. (1995).- About some significant ammonites from the Lower Aptian (Bedoulian) of the Angle-Barrême area (Southeast France).- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 65-101.
G. (1997).- Biostratigraphie haute résolution du Barrémien supérieur du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), t. 325, p. 689-694.
H. (1890).- Sur la classification des Cératites de la Craie.- Bulletin de la Société géologique de France, Paris, (3ème Série), t. XVIII, p. 275-292.
V.V. & T.N. (1979).- Zonengliederung der Unteren Kreide der südlichen UdSSR nach Ammoniten und Foraminiferen.- Aspekte der Kreide Europas, Stuttgart, (IUGS Series A), n° 6, p. 107-116.
V.V. & M.P. (1960).- Атлас нижнемеловой фауны Северного Кавказа и Крыма [Atlas of Lower Cretaceous fauna of the northern Caucasus and Crimea].- Trudy Vsesoyuznogo Nauchno-Issledovatelskogo Geologicheskogo Instituta, Moscow, 702 p. [in Russian].
C.H., M. & B. (1972).- La Sierra d'Aralar (Pyrénées basques espagnoles) : stratigraphie, structure.- Mémoire synthétique, Thèses de Doctorat de 3ème Cycle, Institut des Sciences de la Terre, Université de Dijon, 240 p. ronéotées, 9 pls.
E. (1996).- The Aptian stage.- Bulletin de l'Institut royal des Sciences naturelles de Belgique, Bruxelles, (Science de la Terre), vol. 66, p. 31-43.
M. & P. (1975).- Un exemple d'interrelation entre socle, paléo-géographie et structure dans l'arc pyrénéen basque : La sierra de Aralar.- Revue de Géographie physique et de Géologie dynamique, Paris, vol. XVII, fasc. 5, p. 497-512.
E. (1845).- Catalogue of the Lower Greensand fossils in the Museum of the Geological Society, with notices of species new to Britain.- Quarterly Journal of the Geological Society, London, vol. 1, p. 237-250 & 345-355.
R. & J.A. (2010).- Dufrenoyia furcata (, 1836), ammonite del Aptiense inferior (Cretácico Inferior) del Perelló (Tarragona). In: M., I., J.M., M., R. & V. (coords.), VIII Encuentro de Jóvenes Investigadores en Paleontología, Volumen de Actas.- Cidaris, Elche, n° 30, p. 129-137.
J., M.A., A. & P.A. (2005).- Pulsating subsidence during a rift episode: stratigraphic and tectonic consequences (Aptian-Albian, Northern Spain).- Terra Nova, Paris, vol. 17, n° 6, p. 517-525.
J., H.G., N., M.I. & P.A. (2009).- The Early Aptian of Aralar (North Spain): Stratigraphy, sedimentology, ammonite biozonation, and OAE1.- Cretaceous Research, London, vol. 30, n° 2, p. 434-464.
T. (1871).- Arrangement of the families of Mollusks.- Smithsonian Miscellaneous Collections, Washington, vol. X, Smithsonian Institution Publication, n° 227, xvi + 49 p.
A.E. (1973).- Палеонтологическое обоснование стратиграфического расчленения меловых отложений Поволжья. Верхний мел [Paleontological evidence for the stratigraphic subdivision of the Cretaceous in Povolzhie, Lower Cretaceous].- Nedra, Moscow, 324 p. [in Russian].
R., R. & G. (2000).- Les Deshayesites de la Zone à D. tuarkyricus dans l'Aptien inférieur de Cassis-La Bédoule (SE France).- Géologie méditerranéenne, Marseille, t. XXV, n° 3-4 (1998), p. 125-147.
J.M. (1991).- Ammonite scales for the Cretaceous System.- Cretaceous Research, London, vol. 12, n° 3, p. 259-291.
P.J. & L. (reporters), E., R., M., G., M., E., J., I., J.L., L., P.F., J., J.M., J.-P., G., Z. & J. (1990).- Preliminary ammonite zonation for the Lower Cretaceous of Mediterranean region (Lower Cretaceous Cephalopod team, Working Group of IGCP, Project 262).- Géologie Alpine, Grenoble, t. 66, p. 123-127.
P.J. & M. (reporters), M.B., E., T.N., L., L., F., G., M., L., H., P.F., J., J.M., J.-P., S.Z. & Z. (1993).- Ammonites zonation for the Lower Cretaceous of the Mediterranean region; basis for the stratigraphic correlation within IGCP-Project 262.- Revista Española de Paleontología, Madrid, vol. 8, n° 1, p. 117-120.
P.J. & F. (reporters), E., M., G., E., M., P., M., G., A., L., G., P.F., P., J., J.M. & Z. (1995).- Report on the 3rd International Workshop on the standard Lower Cretaceous ammonite zonation of the Mediterranean region.- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 213-215.
P.J. & P.F. (reporters), M.B., T.N., I., P., C., M., J., A., N., O. & Z. (2000).- Report on the 5th International Workshop of the Lower Cretaceous Cephalopod team (Vienna, 5 September 2000).- Cretaceous Research, London, vol. 21, n° 6, p. 857-860.
P.J. & S. (reporters), M.B., P., M., F., R., M., C., J., A., I., N., P.F., P., Z., J. & M.G.E. (2003).- Report on the 1st International Workshop of the IUGS Lower Cretaceous Ammonite Working Group, the ' Group' (Lyon, 11 July 2002).- Cretaceous Research, London, vol. 24, n° 1, p. 89-94.
W.E. (1949).- Geology of the Sierra de los Muertos area, Mexico.- The Geological Society of America, Bulletin, Boulder, vol. 60, p. 89-176.
A. (1889).- Genesis of the Arietidae.- Smithsonian Contributions of Knowledge, Washington, vol. XXVI, Smithsonian Institution Publication, n° 673, p. xi + 239.
A. (1903).- Pseudoceratites of the Cretaceous- Monographs of the United States Geological Survey, Washington, 351 p.
H., K. & A. (1997).- Kreide-Ammoniten aus dem iranischen Teil des Koppeh-Dagh (NE Iran).- Zitteliana, München, vol. 21, p. 159-190.
C. (1907).- Études paléontologiques et stratigraphiques sur la partie moyenne des terrains crétacés dans les Alpes françaises et les régions voisines.- Annales de l'Université de Grenoble, vol. 19, n° 2, p. 221-534.
P.A. (1914).- Описание коллекции головоногих из меловых отложений Дагестана со списком форм других классов и стратиграфическим очерком [Description d'une collection de céphalopodes des terrains crétacés du Daghestan].- Izvestiia Tomskogo Teckhnologicheskoga Instituta, t. 32, n° 4, 127 p. [in Russian].
S.R.A. & A. G. (1999).- Deshayesitid ammonites from the Lower Aptian (Lower Cretaceous) of North-East Greenland.- Scripta Geologica, Leiden, Special Issue 3, p. 89-95.
E. (1967).- Die älteste Ammoniten Fauna im Aptium Nordwest-Deutschlands.- Paläontologische Zeitschrift, Stuttgart, vol. 41, p. 119-131.
E. (1971).- Zur Gliderung und Abgrenzung des norddeutschen Aptium mit Ammoniten.- Geologisches Jahrbuch, Hannover, Band 89, p. 359-390.
E. (1973).- Die Unterkreide im Untergrund der Gehrdener Berge und in der Deistermulde.- Bericht der naturhistorischer Gesellschaft zu Hannover, Bericht 117, p. 29-54.
E. (1976).- Geologischer Führer durch die Grafschaft Bentheim und die angrenzenden Gebiete, mit einem Abriss der emsländischen Unterkreide.- Das Bentheimer Land, Nordhorn, n° 64, 206 p.
E. (1995).- Die Entfaltung der Ammoniten und die Meeresverbindungen im borealen Unter- und Mittel-Apt.- Geologisches Jahrbuch, Hanover, (Reihe A), Heft 141, p. 171-199.
W. (1888).- Sur quelques fossiles du Crétacé inférieur de la Provence.- Bulletin de la Société Géologique de France, Paris, (3ème série), t. XVI, p. 663-691, pls. XVII-XXXI.
W. (1889).- Description géologique de la montagne de Lure.- Thèse, Masson édit., Paris, 458 p.
W. (1907-1913).- Unterkreide (Palaeocretacicum). In: F. (ed.), Lethaea Geognostica.- Handbuch der Erdgeschichte, 2, Das Mesozoicum, 3, Kreide Lief 1, 1907, p. 1-168, Lief 2, 1910, p. 169-287, Lief 3, 1913, p. 289-398.
W. & P. (1915).- I. La faune de l'Aptien inférieur des environs de Montélimar (Drôme) (Carrière de l'Homme d'Armes). In: W. (ed.), Contribution à l'étude des faunes paléocrétacées du Sud-Est de la France.- Mémoires pour servir à l'Explication de la Carte Géologique détaillée de la France, Paris, n° 14, 221 p., 9 pls.
A. von (1902).- Die Ammonitiden des Norddeutschen Neocom (Valanginien, Hauterivien, Barrêmien und Aptien).- Abhandlungen der Königlich Preußischen Geologischen Landesanstalt und Bergakademie, (N.F.), n° 24, 451 p.
E.V. (1988).- Distribution globale des ammonites éocrétacées du Caucase. In: J. & J. (eds.), Cephalopods - Present and Past, Proceedings of the Second International Cephalopod Symposium.- Schweizerburt'sche verlagbuchhandlung, Stuttgart, p. 453-468.
E.V., M. & I. (2000).- Steps in evolution of the Early Cretaceous ammonites of Georgia.- Trudy Akademiya Nauk Gruzinskoy SSR, Tiblisi, (Novaya Serya), Vyp. 115, p. 7685 [in Russian].
D. (1997, unpublished).- El Aptiense del Sur de la Sierra de Aralar.- Trabajo de investigación del Programa de Doctorado, Universidad del País Vasco, 19 p.
D. & J. (1997).- El surco de Aia-Zaldibia (Aptiense inferior, Aralar, Gipuzkoa).- Geogaceta, Madrid, vol. 22, p. 105-108.
I.P. (1952).- Нижнемеловые отложения Северо-Западного Кавказа и их фауна [Lower Cretaceous deposits of the northwestern Caucasus and their fossils].- Trudy Vsesojuznogo Neftjanogo Nauchno-Issledovatel'skogo Geologo-Razvedochnogo Instituta, Moscow, vol. 65, 270 p. [in Russian].
R. (1982).- Ammonoideos cretácicos del Prepirineo de la provincia de Lleida.- Publicaciones del Instituto Geológico de la Universidad de Barcelona, vol. 17, 197 p.
F., J.-G. & G. (1997).- Un exemple de spéciation lié à l'eustatisme : l'apparition précoce de Schackoina cabri (foraminifère planctonique mésogéen).- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Planètes), vol. 325, n° 3, p. 225-230.
F.H. (1932).- Problem of the Lower Cretaceous of the Canadian interior.- Transactions of the Royal Society of Canada, Ottawa, vol. XXVI, section IV, p. 157-175.
L. (1995).- Aptian biostratigraphy of northeastern Tunisia. A stratigraphical approach of Aptian and Albian of Tunisia.- Memorie Descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 109-120.
L. (1999).- L'Aptien et L'Albien de Tunisie. Biostratigraphie à partir des ammonites.- Bulletin de la Société géologique de France, Paris, vol. 170, n° 3, p. 303-309.
I.A. (1958).- Дегезитиды из нижнемеловых отложений Дагестана и Центрального Предкавказья [Deshayesitids from the Lower Cretaceous of Daghestan and Central Fore-Caucasus].- Materialy kosnovam Paleontologii, vol. 2, p. 21-29 [in Russian].
M.I., L.M., P.A. & J. (2005).- El surco calcarenítico intraplataforma de Desamendi: origen y evolución (Aptiense inferior, Aralar-Navarra).- Geo-Temas, Madrid vol. 8, p. 73-76.
M.I., P.A. & J. (2007).- Pulsos de inundación marina en la terminación de una plataforma carbonatada (Aptiense Inferior de Aralar, Cuenca Vasco-Cantábrica).- Geogaceta, Madrid, vol. 41, p. 127-130.
L., E., J.R. & G. (1974).- Continental margins of Galicia-Portugal and Bay of Biscay.- In: C.A. & C.L. (eds.), The geology of continental margins.- Springer-Verlag, New York, p. 323-342.
L., D.G., O. & P. (1979).- Rifting and subsidence of the northern continental margin of the Bay of Biscay.- Initial Reports of the Deep Sea Drilling Project, College Station, vol. 48, p. 1025-1059.
J.A. (2007).- Biostratigrafía del Aptiense del Macizo del Garraf (NE de la Península Ibérica).- Geogaceta, Madrid, vol. 41, p. 131-134.
J.A., M., T., R., G., R. & A. (2009).- Biostratigraphic characterization by means of ammonoids of the Lower Aptian Oceanic Anoxic Event (OAE 1a) in the eastern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, London, vol. 30, n° 4, p. 864-872.
J.A., M., T., R., G., F.J.-M.R., A. & R. (2010).- Lower Aptian ammonite biostratigraphy in the Maestrat Basin (Eastern Iberian Chain, Eastern Spain). A Tethyan transgressive record enhanced by synrift subsidence.- Geologica Acta, Barcelona, vol. 8, n° 3, p. 281-299.
M. & V. (1881).- Über Ammonitiden aus den Hilsbildungen Norddeutschlands.- Palaeontographica, Cassel, vol. 27, n° 3-6, p. 129-203.
A. d' (1841).- La Paléontologie française. Description des mollusques et rayonnés fossiles - Terrains Crétacés. Tome Premier. Céphalopodes.- Masson, Paris, 662 p.
K. (1907).- Beschreibung der während der Forschungsreisen M. v. im Kaukasus gesammelten Versteinerungen. In: M. von (ed.), Kaukasus. Reisen und Forschungen im kaukasischen Hochgebirge.- Band III, Dietrich Reimer (Ernst Vohsen), Berlin, p. 141-174.
C.F. & G. (1895).- Sur la faune du Callovien inférieur (Chanasien) de Savoie.- Mémoires de l'Académie royale des Sciences, Belles-lettres et Arts de Savoie, Chambéry, (Série IV), t. VI, p. 35-211, 11 pls.
C.F. & G. (1897).- Fossili albiani d'Escragnolles, del Nizzardo e della Liguria occidentale.- Paleontographia Italica, Pisa, vol. 2, p. 53-112.
S.N. (2002, unpublished).- Lower Cretaceous (Upper Barremian-Lower Albian) Ammonite faunas of the Kopet Dagh Basin, NE Iran.- PhD thesis, University College London, 337 p.
S.N. (2004).- The ammonite family Deshayestidae in the Kopet Dagh Basin, north-east Iran.- Cretaceous Research, London, vol. 25, n° 1, p. 115-136.
P.F. (1981).- Early Cretaceous ammonite biostratigraphy and biogeography. In: M.R. & J.R. (eds.), The Ammonoidea: The evolution, classification, mode of life and geological usefulness of a major group.- Systematics Association Special Volume Series, London, vol. 18, p. 499-529.
P.F. (1983).- The Valanginian to Aptian stages - current definitions and outstanding problems. In: Symposium Kreide.- Zitteliana, München, Heft 10, p. 493-500.
P.F. & P.J. (reporters), M.B., E., M., S.R. A, J., E., H.G., P., M.R.A., M. & Z. (1999).- Report on the 4th International Workshop of the Lower Cretaceous Cephalopod team (IGPC-Project 362).- Scripta Geologica, Leiden, Special Issue 3, p. 313.
S. & P.J. (reporters), M.B., P., F., E.Y., M., G., Y., J. J.L., A., V., F.A., I., N., P., J., J.M., Z., J., H., B. & I. (2006).- Report on the second international meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Neuchâtel, Switzerland, 8 September 2005).- Cretaceous Research, London, vol. 27, n° 5, p. 712-715.
S. & J. (reporters), R., M., C., A., N., J., O., J.M., Z. & J. (2009).- Report on the 3rd International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Vienna, Austria, 15th April 2008).- Cretaceous Research, London, vol. 30, n° 2, p. 496-502.
O. (1982).- The Cretaceous ammonites of Venezuela.- Birkhäuser Verlag, Basel, 132 p.
A.C. & M.B. (1989).- Hemihoplitid ammonoids from the Lower Cretaceous of Southern Patagonia.- Palaeontology, Oxford, vol. 32, n° 2, p. 447-462, Pls. 51-52
L. (1938).- Amonitas del Cretácico inferior de la Cordillera Oriental.- Estudios Geológicos y Paleontológicos sobre la Cordillera Oriental de Colombia, Parte 2, Ministerio de industria y trabajo. Departmento de minas y petróleos, Bogotá, p. 7-78.
E. & L.G. (2005).- Albian ammonite faunas from Peru: the Genus Neodeshayesites , 1964.- Journal of Paleontology, Tulsa, vol. 79, n° 3, p. 611-618.
F. (1938).- Les ammonites jurassiques et crétacées. Essai de genera.- Masson, Paris, 554 p.
P., F. & R. (2000).- The stratigraphic position of Ammonites consobrinus d' (Deshayesitidae, Ammonoidea) in the Lower Aptian of Cassis-La Bédoule (SE France).- Géologie méditerranéenne, Marseille, t. XXV, n° 3-4 (1998), p. 159-165.
P., G., M., G. & R. (2008).- The Douvilleiceratidae (Ammonoidea) of the Lower Aptian historical stratotype area at Cassis-La Bédoule (SE France).- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2008/03 (CG2008_M03), 60 p.
P., R. & G. (1999).- The 'Pseudocrioceras interval' and adjacent beds at La Bédoule (SE France): implications to highest Barremian/lowest Aptian biostratigraphy.- In: P.F. & P.J. (eds.), Proceedings 4th International Workshop Cephalopod Team (IGCP-Project 362)- Scripta Geologica, Leiden, Special Issue 3, p. 159-213.
P., M., R., G. & G. (2006).- The Deshayesitidae , 1949 (Ammonoidea) of the Aptian stratotype region at Cassis-la Bédoule (SE France).- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2006/01 (CG2006_M01), 46 p.
I. (1933).- Les ammonites aptiennes de la Géorgie occidentale.- Bulletin de l'Institut géologique de Géorgie (1932), Tbilisi, vol. 1, n° 3, p. 165-273.
C. (1893).- Étude sur les Oppelia du groupe du nisus et les Sonneratia du groupe du bicurvatus et du raresulcatus.- Bulletin de la Société géologique de France, Paris, (3e série), t. XXI, p. 149-164.
I.G. (1958).- Lower Cretaceous deposits of the central regions ofthe Russian Platform. In: O.V. (ed.), Mezozoiskie i tretichnye otlozhenija tsentralnykh oblastei Russkoi Platformy.- Gostoptekhizdat, Leningrad, p. 31-184 [in Russian].
G. (1940).- Cephalopods from the Cretaceous Trinity Group of the South-Central United States.- The University of Texas, Publication, Austin, vol. 3945, p. 969-1107.
K., A. & F. (1971).- Stratigraphy of the Cretaceous rocks southeast of Esfahan.- Geological Survey of Iran, Report, Tehran, n° 20, p. 540.
N.I. (1996).- Biostratigraphic chart for the Lower Cretaceous of the central and eastern European (Russian) Platform.- Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg, vol. 77 (Proceedings of the 4th International Cretaceous Symposium, Hamburg 1992), p. 89-93.
J.C., S.P. & W. (2004).- Pyrenean orogeny and plate kinematics.- Journal of Geophysical Research, Washington, vol. 109 (B08104), 18 p.
I. (1906).- Die Beschreibung einiger Douvilleiceras-Arten aus dem oberen Neokom Russlands.- Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu Sankt-Petersburg, (ser. 2), vol. 44, p. 157-198.
I. (1909).- Beiträge zur Kenntniss des sudrussischen Aptien und Albian. Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu Sankt-Petersburg, (ser. 2), vol. 47, 48 p.
R. (1971).- El Jurásico marino de la Sierra de Aralar (Cuenca Cantábrica Oriental): los problemas post-kimméricos.- Cuadernos de Geología Ibérica, Madrid, vol. 2, p. 509-532.
J. de C. (1836).- Appendix A, Descriptive notes respecting the shells figured in plates 11-23. In: W.H. (ed.), Observations on some of the strata between the Chalk and the Oxford Oolite in the South−East of England.- Transactions of the Geological Society of London, (Series 2), vol. IV, p. 103-390.
L.F. (1921).- On Cretaceous Cephalopoda from Zululand.- Annals of the South African Museum, Cape Town, vol. 12, n° 7, p. 217-321.
L.F. (1922).- On Cretaceous Ammonoidea from Angola collected by Professor J.W. , D.Sc., F.R.S.- Transactions of the Royal Society of Edinburgh, vol. 53, p. 91-160.
L.F. (1923).- A monograph of the Ammonoidea of the Gault, pt. 1.- Palaeontolographical Society (1921), London, 72 p.
L.F. (1924).- On the ammonites of the Speeton Clay and the subdivisions of the Neocomian.- Geological Magazine, Cambridge, vol. 61, n° 716, p. 73-89.
L.F. (1930).- XLVI. On some Ammonoidea from the Lower Greensand.- Annals and Magazine of Natural History, Oxford, (Series 10), vol. 5, n° 29, p. 417-464.
A. (1949).- Lower Cretaceous stratigraphy in southeastern Arizona.- Geological Society of America, Memoir, Boulder, vol. 38, 170 p.
E. (1907).- Über ein norddeutsches Äquivalent der Clansayes-Fauna Südfrankreichs und der Schweiz.- Zentralblatt für Mineralogie, Geologie und Paläontologie, Stuttgart, vol. XX, p. 266-270.
A., M. & M. (2001).- The Aptian, Albian and Cenomanian of Roter Sattel, Romandes Prealps, Switzerland: a high resolution record of oceanographic changes.- Cretaceous Research, London, vol. 22, n° 2, p. 173-199.
O. & N. (2002).- Heteromorph ammonites from the Tata Limestone Formation (Aptian-Lower Albian), Hungary.- Palaeontology, Oxford, vol. 45, p. 1137-1149.
H.H. (1935).- The rocks below the Red Chalk of Lincolnshire and their cephalopod faunas.- Quarterly Journal of the Geological Society, London, vol. 91, p. 1-46.
I., J.P. & L. (2002).- The syn-rift sedimentary cover of North Biscay Margin (Bay of Biscay): from new reflection seismic data.- Bulletin de la Société géologique de France, Paris, vol. 173, n° 6, p. 515-522.
S.Z. (1968).- About the Zone Acanthohoplites prodromus in the Aptian-Albian boundary beds of Turkmenia.- Izvestiya Akademii Nauk Turkmenskoi, Achgabat, (seriya Physico Technicheskogo, Chemicheskogo i Geologicheskogo), N° 2, p. 100-109 [in Russian].
S.Z. (1980).- К вопросы о зоне Acanthohoplites prodromus в верхнем апте Туркмении [On the Zone Acanthohoplites prodromus of the Upper Aptian of Turkmenia].- Izvestiya Akademii Nauk SSSR, Moscow, (seriya Geologiskaya), N° 7, p. 142-144 [in Russian].
P.S. (1937).- An Aptian horizon in the Cretaceous of the Lower Mackenzie Valley.- Journal of Paleontology, Tulsa, vol. 11, p. 69-72.
F.W. (1926).- The Cretaceous Ammonoidea of Eastern Australia.- Memoirs of the Queensland Museum, vol. 8, n° 3, p. 195-242.
J. (1966).- Stammesgeschichte und System der posttriadischen Ammonoideen. Ein Überblick.- Neues Jahrbuch für Geologie und Paläeontologie, Abhandlungen, Stuttgart, vol. 1, n° 125, p. 49-79; vol. 2, n° 127, p. 13-81.
C.W. (1955).- LXIX. Notes on Cretaceous ammonites.-II. The phylogeny of the Desmocerataceae and the Hoplitaceae.- Annals and Magazine of Natural History, Oxford, (Series 12), vol. 8, n° 92, p. 561-575.
C.W., J.H. & M.K. (eds., 1996).- Cretaceous Ammonoidea. In: R.L. (ed.), Mollusca (revised).- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, Vol. 4, 362 p.
K.A. von (1884).- Cephalopoda.- In: K.A. von (ed.), Handbuch der Palaeontologie.- R. Oldenbourg, München & Leipzig, vol. 1, n° 2, Edition 3, p. 329-522.
K.A. von (1895).- Grundzüge der Paläeontologie.- R. Oldenbourg, München & Leipzig, viii + 971 p.