◄ Carnets Geol. 13 (L02) ►
![]()
Contents
[1. Introduction] [2.Geological background and locality] [3. Results]
[4. Discussion] [Bibliographic references]
Department of Geology, University of Tartu, Ravila 14A, EE-50411 Tartu (Estonia)
Department of Geology, The College of Wooster, Wooster, OH 44691 (USA)
Manuscript online since April 24, 2013
[Editor and copy editor: Christian C.
]
![]()
Abundant Skolithos burrows are here described from a possible regressive event bed at Ohesaare cliff (Pridoli), Saaremaa, Estonia. The vertical, cylindrical burrows are identified as Skolithos rather than Trypanites because they intercept and bypass rather than cut bioclasts in the limestone matrix. The absence of encrustation on the upper bedding surface also is evidence that these traces are soft-sediment burrows rather than hardground borings. We interpret this intensive bioturbation by Skolithos-producing organisms as an indicator of a shallow water paleoenvironment with high hydrodynamic energy.
Trace fossils; Skolithos; shallow shelf; late Silurian; Baltica.
O. & M.A. (2013).- An event bed with abundant Skolithos burrows from the late Pridoli (Silurian) of Saaremaa (Estonia).- Carnets de Géologie [Notebooks on Geology], Brest, Letter 2013/02 (CG2013_L02), p. 83-87.
Un banc remarquable par ses skolithes dans le Pridoli supérieur (Silurien) de Saaremaa (Estonie).- D'abondants terriers de type Skolithos sont décrits dans une strate remarquable, bathydécroissante, identifiée dans la falaise de Ohesaare (Pridoli) à Saaremaa (Estonie). Ces terriers cylindriques et verticaux sont attribués à Skolithos plutôt qu'à Trypanites, car quand ils rencontrent de gros bioclastes dans la matrice calcaire du banc ils ont pris l'option de les contourner plutôt que de les traverser. L'absence d'encroûtement à la surface sommitale du banc est une preuve supplémentaire que ces traces correspondent à des terriers dans un sédiment meuble plutôt qu'à des perforations dans un fond durci. Nous interprétons cette intense bioturbation par les organismes produisant des skolithes comme une des caractéristiques d'un paléoenvironnement de faible bathymétrie à hydrodynamisme élevé.
Traces fossiles ; Skolithos ; environnement littoral ; Silurien inférieur ; Baltica ; Fenno-Sarmatia.
Skolithos , 1840, is an ichnogenus of vertical, mostly cylindrical burrows. They are especially common in the onshore sandy sediments of the Cambrian, but they remained important constituent of shallow water ichnofabrics from Cambrian onwards ( et al., 2010). Skolithos is generally associated with shallow water environments with high hydrodynamic energy. Several polychaete species today produce burrows similar to Skolithos ( et al., 2010).
The island of Saaremaa, Estonia, has well exposed late Silurian (Pridoli) outcrops on its southwestern coast, mostly in the form of small coastal cliffs. While the stratigraphy and lithology of these beds have been extensively studied, little attention has been paid to the ichnology of this sequence, and the Silurian of Estonia in general, apart from biogenic hard substrates ( & , 2010a) and hardgrounds ( & , 2010b).
There are two known sedimentary discontinuity surfaces in the limestones at Ohesaare cliff (, 1990; & , 2010b). The upper surface has been interpreted as a hardground, based on the occurrence of abundant encrusters and Trypanites borings. However, the nature of the lower discontinuity surface is unresolved.
The aims of this paper are: 1) to determine whether the abundant vertical shafts in this studied bed are hard-substrate Trypanites borings or soft-sediment Skolithos burrows; 2) to describe these traces in detail; and 3) to discuss the paleoecology and paleoenvironmental environmental conditions at the time of their formation.
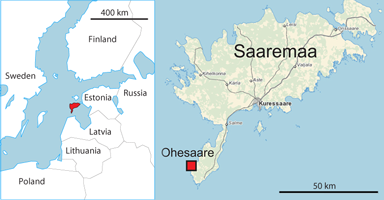
The Ohesaare cliff (58°00'02"N, 22°01'10"E) is located on the western coast of the Sõrve Peninsula of Saaremaa island (Estonia) near Ohesaare village (Fig. 1 ![]() ). The total length of the cliff is over 600 m and it is up to 4 m high (Fig. 2
). The total length of the cliff is over 600 m and it is up to 4 m high (Fig. 2 ![]() ). All together 3.5 m of bedrock are exposed, whereas the thicknesses of individual beds (lenticular and slightly wavy) are variable throughout the outcrop (,
2008).
). All together 3.5 m of bedrock are exposed, whereas the thicknesses of individual beds (lenticular and slightly wavy) are variable throughout the outcrop (,
2008).
Click on thumbnail to enlarge the
image
Figure 1: Map showing the location of Ohesaare cliff (Saaremaa, Estonia).
The section is characterized by the intercalation of thin-bedded limestones and marlstones of normal marine shelf origin (Fig. 2 ![]() ). The bed with abundant Skolithos burrows is situated in the lower middle part of the cliff (Figs. 2
). The bed with abundant Skolithos burrows is situated in the lower middle part of the cliff (Figs. 2 ![]() & 3
& 3 ![]() ).
).

Click on thumbnail to enlarge the
image
Figure 2: Section of Ohesaare cliff after (1990) and & (2010b).

Click on thumbnail to enlarge the
image
Figure 3: Ohesaare cliff. Hammer at the lower surface of the bed with Skolithos burrows.
During the late Silurian the Baltica paleocontinent was located in equatorial latitudes ( et al., 2004). An epicontinental basin covered the area of what is now Estonia. The Baltic Silurian basin was characterized by a wide range of tropical environments and diverse biotas (, 2008). & (1977) recognized five main facies belts in that basin: tidal flat/lagoonal, shoal, open shelf, transitional (basin slope), and a basin depression. The first three facies belts formed a carbonate platform ( & , 1997). The rocks of the Ohesaare cliff were formed in shoal and open shelf conditions (, 2008; & , 2010b).
Figured specimens are deposited at the Geological Museum, Museum of Natural History, University of Tartu (TUG).
Burrows occur in a 2.0 to 4.0 cm thick packstone (Fig. 4 ![]() ). The bioclasts in the packstone are moderately sorted, usually 0.25-0.50 mm in size, but larger bioclasts up to 7.0 mm in size are also common. The packstone layer with burrows is overlain by a thin marlstone interlayer. There is no indication of the layer with burrows being mixed by bioturbation before Skolithos colonization. The cylindrical burrows, 2.0 to 4.0 cm long, usually completely penetrate the layer. The diameters of individual burrows are almost constant.
). The bioclasts in the packstone are moderately sorted, usually 0.25-0.50 mm in size, but larger bioclasts up to 7.0 mm in size are also common. The packstone layer with burrows is overlain by a thin marlstone interlayer. There is no indication of the layer with burrows being mixed by bioturbation before Skolithos colonization. The cylindrical burrows, 2.0 to 4.0 cm long, usually completely penetrate the layer. The diameters of individual burrows are almost constant.

Click on thumbnail to enlarge the
image
Figure 4: Apertures of Skolithos burrows (arrows) on the surface of the regressive event bed.
The burrow apertures are mostly cylindrical, but some apertures are slightly funnel-shaped with the maximum diameter at the bedding plane surface (Fig. 5 ![]() ). There are no successive conical laminae around the aperture of burrows with slightly funnel-shaped apertures.
). There are no successive conical laminae around the aperture of burrows with slightly funnel-shaped apertures.

Click on thumbnail to enlarge the
image
Figure 5: Detail figure of funnel-shaped apertures of Skolithos burrows (arrow).
The apertures are usually circular, sometimes slightly oval, with maximum diameter 1.6 to 3.0 mm (n=17, mean 2.03 mm, σ = 0.35). The maximal diameter of burrows in longitudinal section at about mid-length is 1.4 to 2.1 mm (n=14, mean 1.74 mm, σ = 0.26). The burrows are straight to slightly curved, perpendicular to the surface (79.2 %, n=38) or slightly skewed (20.8 %, n=10) (Fig. 6 ![]() ). The boundaries between the burrows and the surrounding matrix are not sharp (Fig. 6
). The boundaries between the burrows and the surrounding matrix are not sharp (Fig. 6 ![]() ).
).

Click on thumbnail to enlarge the
image
Figure 6: Skolithos burrows in the longitudinal section. Note that boundaries between the burrows and the surrounding matrix are not sharp.
The burrows are filled with bluish grey marl with abundant small bioclasts (resembling brachiopod and echinoderm debris). The lithology of the burrow infilling is similar to the upper thin marl interlayer. The infilling of burrows does not show any backfilling structures. There is no concentric lamination of the burrow infillings. The burrows are densely packed in large numbers (Fig. 4 ![]() ). There are up to 11 burrows per 4 cm2 of top surface. Some burrows are deflected by bioclasts in the matrix (Fig. 7
). There are up to 11 burrows per 4 cm2 of top surface. Some burrows are deflected by bioclasts in the matrix (Fig. 7 ![]() ). Rarely, some burrows intersect others.
). Rarely, some burrows intersect others.

Click on thumbnail to enlarge the
image
Figure 7: Longitudinal section of Skolithos burrow deflected by a bioclast in the matrix (arrow).
No encrustation was discovered on the upper bedding surface of the bed with abundant Skolithos burrows, nor any other indication that this was a carbonate hardground.
The presence of uncut bioclasts intersecting the burrows indicates that they were excavated into soft sediment and are not borings (Fig. 7 ![]() ). The unlithified nature of the layer with burrows is also supported by the lack of encrustation. Abundant encrusters of the same age have been described from a hardground located in the upper part of the Ohesaare cliff ( & ,
2010b). The oval apertures of some burrows can be interpreted as result of post-depositional sediment compression. Alternatively, it could represent the original cross sectional shape of the burrow apertures if the diameter of the burrowing organism was close to the minimum diameter of the oval. It is difficult to interpret the rare joined burrows. The ecology of burrowing organisms is not known in detail, so the joined burrows may have been simultaneously inhabited. The maximal diameter of the burrowing organism was likely close to the average diameter of burrows in longitudinal section 1.4 to 2.1 mm (n=14, mean 1.74 mm, σ = 0.26). The length of the burrowing organism cannot be deduced from the burrow morphology as animals with very different length can produce burrows of similar diameter. However, the burrowing animals were likely able to complete retract into the burrow, so their maximal length was likely less than 4 cm.
). The unlithified nature of the layer with burrows is also supported by the lack of encrustation. Abundant encrusters of the same age have been described from a hardground located in the upper part of the Ohesaare cliff ( & ,
2010b). The oval apertures of some burrows can be interpreted as result of post-depositional sediment compression. Alternatively, it could represent the original cross sectional shape of the burrow apertures if the diameter of the burrowing organism was close to the minimum diameter of the oval. It is difficult to interpret the rare joined burrows. The ecology of burrowing organisms is not known in detail, so the joined burrows may have been simultaneously inhabited. The maximal diameter of the burrowing organism was likely close to the average diameter of burrows in longitudinal section 1.4 to 2.1 mm (n=14, mean 1.74 mm, σ = 0.26). The length of the burrowing organism cannot be deduced from the burrow morphology as animals with very different length can produce burrows of similar diameter. However, the burrowing animals were likely able to complete retract into the burrow, so their maximal length was likely less than 4 cm.
The elongate cylindrical shape of the burrows likely reflects the worm-like body plan of the burrowing organism. Similar burrows are made by polychaete worms in modern seas ( & , 1988; et al., 2010). Skolithos burrows have traditionally been interpreted as domichnia, based on modern analogues (, 1967; & , 1988; & , 1989). We also interpret Skolithos burrows of the Ohesaare cliff as domichnia of possible polychaete worms or worm-like organisms.
A burrowing nereidid polychaete Alitta virens () typically produces burrows similar to Arenicolites and Skolithos ( et al., 2010). The Ohesaare Skolithos burrows are similar to vertical burrows made during the pioneer digging behavior of Alitta virens. In an experimental study, et al. (2010) observed that within 24 hours Alitta virens made numerous vertical burrows penetrating the full depth of sediment in the tank. Thus, it is possible that the Ohesaare Skolithos burrows may represent a similar behavior in a freshly deposited bed of carbonate sand.
et al. (2008) found that low densities of suspension-feeding animals are 1–10 individuals/m2. In case of high densities of interface-feeding (i.e., switching between deposit and suspension feeding) Corophium (a marginal-marine arthropod) commonly exceed 40,000 individuals/ m2, and Euzonus, a deposit-feeding opheliid polychaete is reported to reside in populations exceeding 5,000 individuals/m2 ( et al., 2008). The maximum density of 11 burrows per 4 cm2 in the Ohesaare event bed roughly corresponds to high population densities of modern burrowing organisms. However, it is not clear whether all the burrows were populated with the burrowers at the same time, so the actual population density may have been lower. It is difficult to determine whether the bed with Skolithos was colonized once or several times. However, the relatively moderate variation in diameter of burrows could indicate multiple inhabiting organisms of similar size and age. Thus, most likely the burrowed bed was colonized by single generation of Skolithos trace makers. The lack of other types of traces in the bed with Skolithos also supports a single fortuitous colonization event. The apparent absent of Skolithos burrows in other packstone layers could indicate a different water depth and hydrodynamic regime. Alternatively, Skolithos-makers may have been pioneer colonizers, and in other packstone beds the burrows could have been destroyed by later bioturbation.
Skolithos is usually associated with high-energy environments ( et al., 2010). However, Skolithos burrows have rarely been described from carbonates (Brian , 2013, pers. comm.). The bed in the Ohesaare cliff with Skolithos burrows has been interpreted by (1990) as a regressive event bed (shoal) within generally deeper shelf sediments (open shelf). It is possible to interpret the intensive bioturbation degree by Skolithos burrows as an indicator of a shallow water environment. Presumably during a regressive event a carbonate sand bed was deposited in waters of high hydrodynamic energy. Shortly after deposition, the bed was intensely burrowed by Skolithos trace makers. Alternatively, these burrows could have been made in soft sediment that had ceased movement and that the Skolithos burrows do not indicate a regressive event.
O.V. is indebted for financial support to the Grant Program (Paleontological Society), Estonian Science Foundation grant ETF9064, and the target-financed project (from the Estonian Ministry of Education and Science) SF0180051s08 (Ordovician and Silurian climate changes, as documented from the biotic changes and depositional environments in the Baltoscandian Palaeobasin). M.W. thanks the Faculty Development Fund at The College of Wooster. We are grateful to Brian and an anonymous reviewer for the constructive reviews.
P.R.; M.G., L.A. & B.R. (2010).- Skolithos pipe rock and associated ichnofabrics from the southern Rocky Mountains, Canada: colonization trends and environmental controls in an early Cambrian sand-sheet complex.- Lethaia, Oslo, vol. 43, p. 507-528.
M.K., S.G., S. & L. (2008).- How fast do marine invertebrates burrow?- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 270, p. 280–286.
L.G., O.A. & D. (2010).- Ecosystem engineering by bioturbating polychaetes in event bed microcosms.- Palaios, Lawrence, vol. 25, p. 46–58.
O. (2008).- The Silurian System in Estonia. In: O., L., P. & T. (eds.), The seventh Baltic stratigraphical conference. Abstracts and field guide.- Geological Society of Estonia, Tallinn, p. 46.
M.J., R.A. & P.M. (2004).- The Silurian Period. In: F.M., J.G. & A.G. (eds.), A geologic time scale 2004.- Cambridge University Press, p. 188-201.
H. (1990).- Locality 7:4 Ohesaare cliff. In: D. & H. (eds). Field meeting, Estonia 1990. An excursion guidebook.- Institute of Geology, Estonian Academy of Sciences, Tallinn, p. 175–178.
H. & R. (1977).- Model of facies and sedimentology for Paleobaltic epicontinental basin. In: D.L. (ed.), Facies and fauna Silurian of Baltica.- Institute of Geology AN ESSR, Tallinn, p. 89-121. [in Russian]
S.G. & B. (1988).- Ichnology of the Pleistocene Ironshore Formation, Grand Cayman Island, British West Indies.- Journal of Paleontology, Tulsa, vol. 62, p. 495-505.
A. & A. (1997).- Geology and mineral resources of Estonia.- Estonian Academy Publishers, Tallinn, 436 p.
A. (1967).- Bathymetry of trace fossils.- Marine Geology, Amsterdam, vol. 5 (5–6), p. 413–428.
O. & M.A. (2010a).- Occurrence of giant borings of Osprioneides kampto in the lower Silurian (Sheinwoodian) stromatoporoids of Saaremaa, Estonia.- Ichnos, Philadelphia, vol. 17, p. 166-171.
O. & M.A. (2010b).- Microconchid-dominated hardground association from the late Pridoli (Silurian) of Saaremaa, Estonia.- Palaeontologia Electronica, San Francisco, vol. 13, n° 2, 9A, 12 p.
S.M. & S.G. (1989).- Ichnology and paleoecology of offshore siliciclastic deposits in the Cardium Formation (Turonian, Alberta, Canada).- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 74, p. 217-239.