◄ Carnets Geol. 13 (M02) ►
![]()
Sommaire
[Introduction] [Contexte géodynamique et géologique ...]
[Présentation du gisement ...] [Description paléontologique ...]
[Conclusions] [Références bibliographiques]
et ... [Planches]
La Guilletière, 38700 Sarcenas (France)
Laboratoire de Paléontologie de l'ISE-M, Université Montpellier II
Adresse personnelle : 46 rue Rouget de Lisle, 30000 Nîmes (France)
Manuscrit en ligne depuis le 14 Mai 2013
[Éditeur : Bruno ]
![]()
Une lentille de calcaire marneux induré à la base de la série tertiaire des Grandes Platières (massif de Platé, Haute-Savoie) a livré la plus importante collection de restes de vertébrés actuellement connue dans le Paléogène des Alpes occidentales. Les mammifères ongulés sont représentés par 35 dents attribuées à 9 taxons (4 périssodactyles et 5 artiodactyles). Cette faune paraît contemporaine de celle de La Débruge (niveau MP 18), datée d'environ -35,5 MA. Le nouveau site, dénommé gisement des Perdrix, est situé stratigraphiquement à 20-35 m en-dessous de celui dans lequel et al. (1991) avaient mis au jour quelques restes de mammifères qu'ils ont attribués au niveau MP 20. La nouvelle faune est donc plus ancienne d'environ 1,5 MA. Sa composition et la nature palustre du dépôt suggèrent que les animaux concernés vivaient dans un environnement varié comportant des espaces boisés à proximité de zones humides marécageuses.
Alpes externes ; Éocène ; Vertébrés fossiles ; périssodactyles ; artiodactyles.
M. & J.A. (2013).- Mammifères et reptiles priaboniens du massif de Platé (Haute-Savoie, France).- Carnets de Géologie [Notebooks on Geology], Brest, Mémoire 2013/02 (CG2013_M02), p. 105-129.
Priabonian mammals and reptiles from the Platé Massif (Savoy, France).- A lens of marly limestone at the bottom of the Tertiary series of the Grandes Platières (Platé Massif, Savoy) has yielded the most important collection of fossil vertebrates so far known from the Paleogene of Western Alps. Ungulate mammals are represented by 35 teeth to be referred to 9 taxa (4 perissodactyls and 5 artiodactyls). This new fauna appears contemporaneous with that of La Débruge (MP 18 level), dated at about 35.5 My old. The new locality, called "gisement des Perdrix", is stratigraphically located 20-35 m underneath the other one in which et al. (1991) had found mammals to be attributed to the MP 20 level. So the new fauna is about 1.5 My older than this latter. Its faunal composition and the swampy nature of the deposit suggest that the occurring mammals were living in variegated surroundings, with woody areas close to wet marshy ones.
External Alps; Eocene; fossil vertebrates; perissodactyls; artiodactyls.
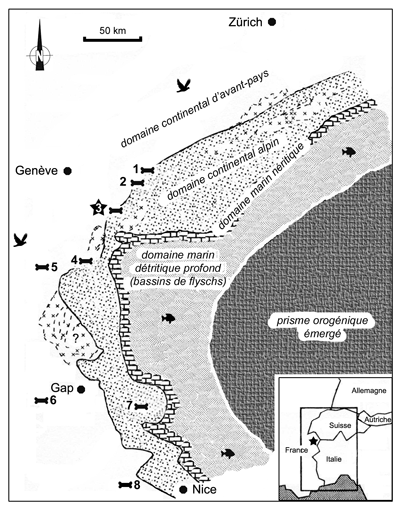
Les restes de vertébrés éocènes dans les Alpes sont particulièrement rares : moins d'une dizaine de sites fossilifères échelonnés entre la Bavière et les Alpes de Haute Provence sont connus (Fig.
1 ![]() ). En outre, la plupart d'entre eux n'ont livré que des spécimens isolés appartenant à un seul taxon. Cinq gisements seulement ont livré un matériel plus
conséquent : la Croix du Lauzanier au nord de l'Argentera
(, 1965), le Jabron dans l'Arc de Castellane ( et al.,
1973), la Montagne du Charbon dans les Bauges
( et al., 1984), Eygalayes dans les Baronnies drômoises ( &
, 1993) et enfin celui des Grandes Platières ( et al.,
1991).
). En outre, la plupart d'entre eux n'ont livré que des spécimens isolés appartenant à un seul taxon. Cinq gisements seulement ont livré un matériel plus
conséquent : la Croix du Lauzanier au nord de l'Argentera
(, 1965), le Jabron dans l'Arc de Castellane ( et al.,
1973), la Montagne du Charbon dans les Bauges
( et al., 1984), Eygalayes dans les Baronnies drômoises ( &
, 1993) et enfin celui des Grandes Platières ( et al.,
1991).
Cliquer
sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 1 : Cadre paléogéographique des gisements alpins à mammifères éocènes (modifié d'après , 1997). 1 : Haute Corde-Essets (Morcles) ; 2 : Saint Maurice (Aiguilles Rouges) ; 3 : Grandes Platières (Platé) ; 4 : Montagne du Charbon (Bauges) ; 5 : Les Échelles (Chartreuse) ; 6 : Eygalayes (Baronnies) ; 7 : Lauzanier (Mercantour) ; 8 : Jabron (Arc de Castellane).
C'est dans ce dernier site que de nouvelles découvertes ont été réalisées par l'un de nous lors d'une séance de photos pour un ouvrage de vulgarisation géologique du Pays du Mont Blanc (, 2002). Ce nouveau gisement a fourni la plus importante collection de dents de vertébrés actuellement connue dans le Paléogène des Alpes occidentales.
Dans les Alpes externes, la transition entre le Mésozoïque et le Cénozoïque est marquée par une importante rupture géodynamique : les séries marines de marge passive du Crétacé supérieur cèdent la place à des dépôts tertiaires de bassin d'avant-pays en réponse à l'initiation de la flexuration de la lithosphère continentale européenne engendrée par la collision de la plaque apulienne avec la marge passive européenne ( & 1992 ; et al., 1999).
La limite de mégaséquence séparant le cycle sédimentaire secondaire du cycle tertiaire est représentée par une discontinuité majeure ("basal unconformity" : et al., 1991 ; "forebulge unconformity" : & 1995) traduisant la présence d'un bombement cratonique en réponse à la formation du prisme orogénique des Alpes internes. Dans la zone delphino-helvétique, la mise en place du bombement cratonique s'accompagne du développement d'accidents tectoniques ( & 1975 ; 1995) dont certains réactivent des paléofailles héritées du rifting ( & 1996). Avec les érosions liées à l'émersion engendrant des vallées incisées, ces accidents engendrent une topographie anté-Tertiaire complexe (, 1998).
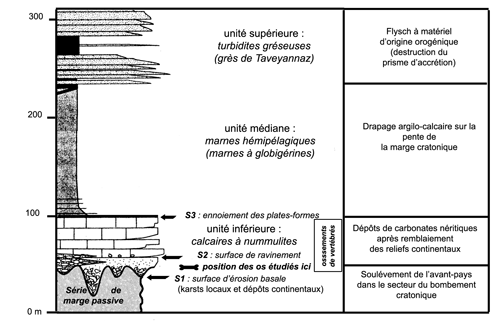
Avant l'installation de la plate-forme
nummulitique, premier terme de la trilogie de bassin sous alimenté (1997, et Fig. 2 ![]() ), les creux de cette surface topographique seront comblés
par des sédiments variés indiquant des environnements terrestres (paléosols
à Microcodium, karst à cuirasse ferrifère), fluviatiles (cônes alluviaux à
stromatolites), lacustres (oncolites), voire même marins peu profonds
(calcaires gréseux à grandes nummulites). Constitués en majeure partie par
des conglomérats dont les éléments proviennent de la destruction de roches crétacées,
voire jurassiques et plus anciennes localement (bordure sud du Pelvoux :
et al., 1987 ; ,
1999), les dépôts clastiques
infra-nummulitiques interprétés comme des dépôts de bas niveau marin
terminal (, 1997) sont très mal datés si ce n'est par la présence
de galets les plus jeunes (calcaires à grandes nummulites). Dans ce contexte,
la découverte d'ossements de vertébrés, en particulier de dents de mammifères,
dans le massif de Platé (Haute Savoie) fournit un précieux jalon de datation
des dépôts comblant la morphologie du substratum anté-Tertiaire.
), les creux de cette surface topographique seront comblés
par des sédiments variés indiquant des environnements terrestres (paléosols
à Microcodium, karst à cuirasse ferrifère), fluviatiles (cônes alluviaux à
stromatolites), lacustres (oncolites), voire même marins peu profonds
(calcaires gréseux à grandes nummulites). Constitués en majeure partie par
des conglomérats dont les éléments proviennent de la destruction de roches crétacées,
voire jurassiques et plus anciennes localement (bordure sud du Pelvoux :
et al., 1987 ; ,
1999), les dépôts clastiques
infra-nummulitiques interprétés comme des dépôts de bas niveau marin
terminal (, 1997) sont très mal datés si ce n'est par la présence
de galets les plus jeunes (calcaires à grandes nummulites). Dans ce contexte,
la découverte d'ossements de vertébrés, en particulier de dents de mammifères,
dans le massif de Platé (Haute Savoie) fournit un précieux jalon de datation
des dépôts comblant la morphologie du substratum anté-Tertiaire.
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 2 : Place et extension verticale des niveaux à ossements dans la trilogie des faciès du Tertiaire alpin (adapté d'après , 1997).
La première découverte connue de restes osseux aux Grandes Platières daterait de 1979 mais il est amusant de noter à ce sujet que la quasi-totalité des restes de végétaux signalés dans les coupes de & (1975) correspondent à des restes osseux de vertébrés.
Il faudra cependant attendre 1991 pour que, à la suite des prospections effectuées par , paraisse une première publication révélant la présence, dans le sommet des Couches à Cérithes, de trois espèces de périssodactyles appartenant au genre Palaeotherium (et al., 1991).
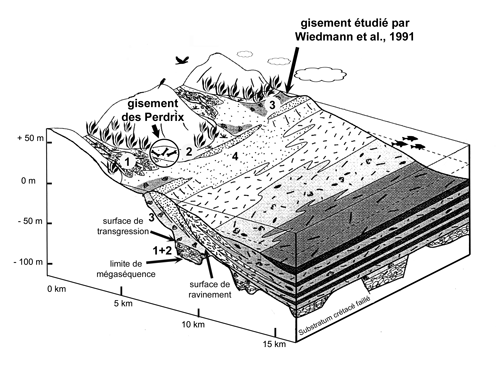
Le nouveau gisement, objet de la présente étude,
se situe au nord-est de la gare sommitale de la télécabine des Grandes Platières,
entre les télésièges du Diamant Noir et celui des Perdrix
(Fig. 3 ![]() ). Nous le dénommerons
"gisement des Perdrix", dénomination empruntée au télésiège voisin et
faisant référence à la présence de Lagopèdes, oiseaux galliformes encore
appelés "perdrix des neiges".
). Nous le dénommerons
"gisement des Perdrix", dénomination empruntée au télésiège voisin et
faisant référence à la présence de Lagopèdes, oiseaux galliformes encore
appelés "perdrix des neiges".

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 3 : Position stratigraphique du gisement des Perdrix d'après photographie. Coordonnées du point central : 5 095 378 m N 323 572 m E (altitude 2 360 m) ; (UTM transverse Mercator, fuseau 32 T).
Ce nouveau gisement s'est vite avéré beaucoup
plus riche en restes osseux que les niveaux conglomératiques des couches à cérithes
exploités par et al. (1991). Malgré cette
abondance relative et la présence de pièces plus complètes, les conditions de
récoltes restent délicates, les fossiles de vertébrés se présentant dans
une gangue de marne très indurée, souvent schistosée, déterminant un "éclatement"
des spécimens les plus importants (Fig. 4 ![]() ).
).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 4 : Trois exemples d'ossements de vertébrés
du niveau fossilifère des Perdrix avant extraction.
A : fragment de mandibule droite d'un
Artiodactyla indet. (V5598) ;
B : maxillaire gauche de Palaeotherium (V5599) ;
C : fragment de maxillaire d'alligatoridé
(V5593).
Le gisement des Perdrix a en partie disparu durant l'été 2005 suite à des travaux d'aménagement d'une piste de ski pour la station de Flaine (Fig.
5 ![]() ). Malgré une information préliminaire faite auprès de la Direction régionale de l'Environnement de Rhône-Alpes et auprès du Bureau de Recherche géologique et minière, ces travaux ont été
autorisés ! En conséquence, cet aménagement a irrémédiablement détruit environ un tiers de ce site paléontologique exceptionnel, unique dans les Alpes par sa richesse en vertébrés tertiaires. Il reste encore beaucoup à faire en France pour que la notion de patrimoine géologique devienne une réalité.
). Malgré une information préliminaire faite auprès de la Direction régionale de l'Environnement de Rhône-Alpes et auprès du Bureau de Recherche géologique et minière, ces travaux ont été
autorisés ! En conséquence, cet aménagement a irrémédiablement détruit environ un tiers de ce site paléontologique exceptionnel, unique dans les Alpes par sa richesse en vertébrés tertiaires. Il reste encore beaucoup à faire en France pour que la notion de patrimoine géologique devienne une réalité.

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 5 : Destruction partielle du gisement des Perdrix durant l'année 2005 (état au 28/08/2005).
Les ossements du gisement des Perdrix se situent entre +7m et +20 m par rapport à la surface d'érosion de la base des dépôts du Paléogène (limite de mégaséquence ; surface S1 en Fig. 2 ![]() ) recoupant le toit des calcaires lités à
Globotruncanita du sommet du Crétacé supérieur. Ce niveau est donc sous-jacent à la surface de transgression des couches à cérithes, soit environ 20 à 35 m plus bas que les niveaux prospectés par et al.
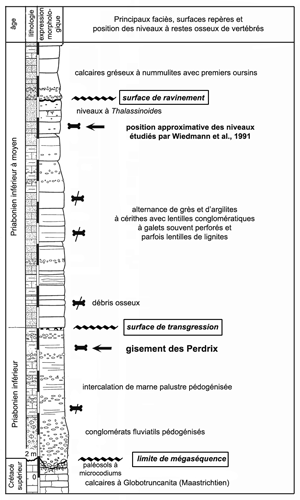
(1991). Une coupe simplifiée du gisement est donnée en
Figure 6
) recoupant le toit des calcaires lités à
Globotruncanita du sommet du Crétacé supérieur. Ce niveau est donc sous-jacent à la surface de transgression des couches à cérithes, soit environ 20 à 35 m plus bas que les niveaux prospectés par et al.
(1991). Une coupe simplifiée du gisement est donnée en
Figure 6 ![]() .
.
Selon la nomenclature lithostratigraphique élaborée dans les nappes helvétiques des Diablerets et du Wildhorn (, 1994), le gisement des Perdrix se rattache à la Formation du Sanetsch, peu au-dessus de la base du Membre des Diablerets.
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 6 : Situation stratigraphique du gisement des Perdrix et extension verticale des restes osseux (adapté d'après , 1998).
Sur un peu plus d'un hectare, sont disséminés
plusieurs dizaines d'ossements dont des dents, des fragments de mâchoires et
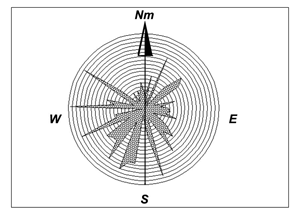
des débris d'os longs. Les mesures d'orientation portant sur les débris osseux
suffisamment allongés révèlent une assez grande dispersion avec cependant un
maximum de concentration dans le quadrant SW, ce qui suggère un apport en
provenance du NW ou du SE (Fig. 7 ![]() ). Compte tenu des données paléogéographiques
locales suggérant la présence de terres émergées en direction du NW (,
1943 ; & ,
1975 ; ,
1998), ces restes
osseux sont très vraisemblablement originaires du secteur du
Haut-Giffre.
). Compte tenu des données paléogéographiques
locales suggérant la présence de terres émergées en direction du NW (,
1943 ; & ,
1975 ; ,
1998), ces restes
osseux sont très vraisemblablement originaires du secteur du
Haut-Giffre.
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 7 : Répartition des orientations des fragments des os longs fondée sur 57 mesures prises dans le gisement des Perdrix.
Le plus ancien matériel récolté est représenté
par une dent trouvée à la cote +5m par rapport à la limite de mégaséquence,
au toit immédiat de la corniche conglomératique ("Assise conglomératique"
sensu & ,
1975) représentant des cônes alluviaux partiellement pédogénisés à débris
de Microcodium et à matériel crétacé et plus rarement
tertiaire (galets
à grandes nummulites, calcaires lacustres pédogénisés : Fig. 8 ![]() ).
).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 8 : Faciès du toit de la corniche basale conglomératique remaniant des calcaires lacustres pédogénisés (1) et de rares restes osseux très dispersés (flèches) [vue en coupe verticale].
Mais la majorité du matériel du gisement des
Perdrix provient d'un niveau de marne indurée situé à environ +10 à +15m
au-dessus de la discontinuité basale et plus proche de la surface de
transgression marquant la base des couches à cérithes (Fig. 9 ![]() ).
).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 9 : Position stratigraphique du principal niveau à ossements du gisement des Perdrix.
Ce niveau présente une extension latérale
limitée (Fig. 10 ![]() ), disposition stratonomique particulière qui explique
probablement la découverte tardive de ce gisement qui avait jusqu'à présent échappé à la sagacité des nombreux géologues ayant
arpentés les dépôts tertiaires des Grandes Platières depuis près de
deux siècles (, 1826).
), disposition stratonomique particulière qui explique
probablement la découverte tardive de ce gisement qui avait jusqu'à présent échappé à la sagacité des nombreux géologues ayant
arpentés les dépôts tertiaires des Grandes Platières depuis près de
deux siècles (, 1826).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 10 : Disparition latérale du niveau à ossements du gisement des Perdrix (en jaune). 1 : calcaires du Crétacé supérieur ; 2 : conglomérat pédogénisé ; 3 : couches à cérithes ; 4 : calcaires gréseux à nummulites (barre tidale) ; 5 : calcaire de plate-forme à mélobésiées. FD : faille décrochante.
À l'exception des restes osseux dispersés, ce
faciès est pratiquement dépourvu de macrofossiles. Cependant, on note çà et là
des concentrations décimétriques de petits gastéropodes et de petites huîtres
évoquant des apports brutaux et localisés de haute énergie dans un milieu de
décantation boueuse (Fig. 11 ![]() ). Ces caractéristiques faciologiques et sédimentologiques
évoquent un milieu palustre ou un dépôt de "wash over" ou de
"crevasse splay" dans un environnement d'estran péri-estuarien
("mud flat").
). Ces caractéristiques faciologiques et sédimentologiques
évoquent un milieu palustre ou un dépôt de "wash over" ou de
"crevasse splay" dans un environnement d'estran péri-estuarien
("mud flat").

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 11 : Faciès des intercalations microconglomératiques lenticulaires au sein des niveaux à vertébrés. A : calcaire gréseux à malacofaune silicifiée. B : microconglomérat à galets et prisme d'inocérames (fragments blancs) remaniés du Crétacé supérieur.
Les restes osseux du gisement des Perdrix proviennent donc d'un environnement de dépôt très différent de celui des couches à cérithes des récoltes de et al. (1991).
Bien que le matériel osseux soit le plus
souvent réduit à l'état de débris, on ne relève pas de traces importantes
de roulage ; de nombreuses pièces se rencontrent aussi brisées sur place dans
le sédiment et l'os est parfois plus ou moins dissous tout en laissant les
dents dans leur position relative naturelle (Fig. 4 ![]() ).
).
Le mauvais état de conservation de certaines dents est lié en partie aux possibilités d'exploitation du gisement. Étant en présence d'une assise de marne compacte épaisse de plusieurs décimètres, seules les pièces apparaissant en surface par dissolution différentielle ont pu faire l'objet d'une extraction à l'aide d'une broche et d'une masse. De plus, cette récupération a été souvent rendue délicate par un écaillage suivant des plans de clivage schisteux.
En laboratoire, le dégagement des pièces a été effectué par attaque acide à l'aide d'acide formique dilué à 5 %. Elles ont été consolidées par imprégnation d'une dissolution de résine acrylique (Paraloïd 72) pour éviter un possible délitement.
La collection du nouveau gisement comprend quelques restes de reptiles et surtout 35 dents de mammifères ongulés se rattachant à 9 taxons, 4 espèces de périssodactyles (2 Palaeotherium et 2 Plagiolophus) et 5 espèces d'artiodactyles (2 Anoplotherium, 1 Cebochoerus, 1 Choeropotamus et 1 Artiodactyla indet.).
Ce matériel est déposé au Muséum d'Histoire Naturelle de Genève (collection , MHNG GEPI V5565-V5618). Les mesures sont données en millimètres.
Les sites de conservation des spécimens cités dans le texte sont abrégés de la façon suivante :
Classe Sauropsida , 1864
Ordre Crocodylia , 1789
Famille Alligatoridae , 1844
Genre Diplocynodon , 1847
cf. Diplocynodon sp.
Pl. 1 ![]() ,
figs. 1-5
,
figs. 1-5
Matériel :
Description : La dent V5580, un peu ébréchée à l'apex et à la base, mesure 19,5 mm de hauteur. Sa section, presque circulaire sur toute la hauteur, est de 11,5x11,0 à la base. La couronne est légèrement dissymétrique, un peu recourbée vers l'arrière et vers l'intérieur. La surface de l'émail est quasiment lisse, ornée seulement par une crête sagittale, mésiale et distale, fine, aiguë et non dentelée.
Le fragment de maxillaire V5593 de petite taille comporte une dent visible dans son alvéole, associée à une autre trouvée à proximité et qui provient à l'évidence du même spécimen. Ces 2 dents sont beaucoup plus petites que V5580; la dent libre, très pointue, mesure 6 mm de hauteur pour une section de 2,9x2,2 ; l'émail est lisse et présente, comme V5580, une fine crête mésiale et distale, non dentelée.
Les deux morceaux de plaques dermiques V5585, V5592 sont creusés de cupules arrondies de forme irrégulière mais à bords nets, plus grandes sur V5585 que sur V5592.
La vertèbre caudale est procle et mesure 15 mm de long.
Discussion : Dans le contexte régional d'effondrement de la diversité générique des crocodiliens à l'Éocène supérieur, l'attribution de la grande dent non ziphodonte au genre Diplocynodon apparaît comme probable. L'ornementation des plaques dermiques n'est au surplus pas très différente de celle des spécimens de Sainte-Neboule attribués à ce genre par (1978). Quant au maxillaire, il ne paraît pas impossible qu'il appartienne lui aussi au même taxon, sa très petite taille et les particularités de ses dents seraient à mettre alors au compte de la juvénilité.
La médiocrité de tous ces restes impose cependant de s'en tenir à une détermination prudente de "cf. Diplocynodon".
Classe Mammalia , 1758
Ordre Perissodactyla , 1848
Famille Palaeotheriidae , 1850
Genre Palaeotherium , 1804
Palaeotherium muehlbergi thaleri , 1985
Fig. 12 ![]() ;
Tableaux 1 - 2 ; Pl. 2
;
Tableaux 1 - 2 ; Pl. 2 ![]() ,
figs. 1-5
,
figs. 1-5
Matériel :
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
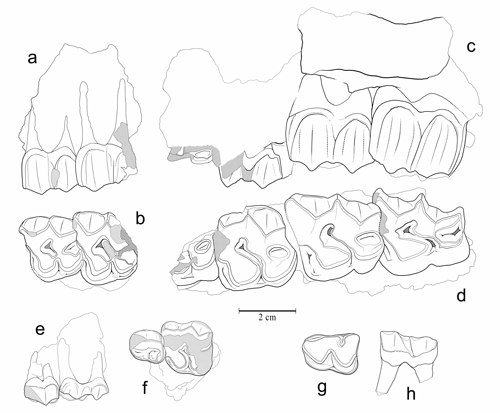
Figure 12 : Palaeotherium muehlbergi thaleri. a-b : MHNG GEPI V5573, P3/-P4/ droites (inversées) (a : vue labiale ; b : vue occlusale). c-d : MHNG GEPI V5599, (P4/)-M1/-M3/ gauches (c : vue labiale ; d : vue occlusale). e-f : MHNG GEPI V5588, P1/-DP2/ droites (inversées) (e : vue labiale ; f : vue occlusale). g-h : MHNG GEPI V5584, P/4 gauche, (g : vue occlusale ; h : vue labiale).
Description : Le maxillaire V5573 offre une paroi osseuse externe très haute, légèrement concave, sans trace de foramen infra-orbitaire ni de départ de l'arcade zygomatique ; il est pourvu de deux dents complètement molarisées ; la morphologie osseuse et la présence sur ces deux dents d'une courte liaison entre protocône et hypocône et de cingulums internes épais, montre qu'il s'agit bien de prémolaires ; l'absence de retrait de l'angle antéro-interne de la 1ère dent montre que celle-ci n'est pas une P2/. Les plans de l'ectolophe des deux dents sont un peu convexes avec des côtes bien marquées au niveau des cuspides ; quoique assez usées, les cuspides labiales sont très allongées dans le plan vertical, peu incurvées, mais fortement inclinées, si bien que l'indice d'hypsodontie (, 2004) reste faible, env. 0,57. Les styles sont peu proéminents, mais le mésostyle est complet, même sur P3/. Le protocône de P4/ présente une section anguleuse, triangulaire et l'hypocône des 2 dents est un peu en retrait surtout sur P3/.
Le maxillaire V5599, bien que très dégradé, a conservé une partie de la tubérosité et de la base de l'orbite. Le plancher orbitaire est élevé, situé à 22 mm au-dessus du rebord alvéolaire. On voit également la base écrasée de l'arcade zygomatique, qui semble avoir été très épaisse (jusqu'à 30 mm à l'angle de l'orbite). Le bord antérieur du zygoma est redressé presque à la verticale, et se termine par une zone très fortement rétrécie à sa racine antérieure, en avant de l'orbite.
Les dents jugales sont déjà assez abrasées, mais l'usure n'est encore que modérée sur l'hypocône de M3/. L'indice d'hypsodontie de cette dent peut être estimé à 0,61. Elle présente un ectolophe fortement courbe dans le plan frontal avec des styles proéminents et un bourrelet cingulaire labial continu, renflé à la base du mésostyle ; les cuspides labiales sont soulignées par des côtes émoussées bien marquées. La dent est par ailleurs assez peu allongée transversalement, rétrécie au niveau du lobe postérieur avec un bord distal échancré et concave. M2/ est également peu transverse, son ectolophe est très allongé et moins recourbé que celui de M3/ et les styles sont nettement moins puissants. La section du protocône est plus anguleuse que sur M3/, presque triangulaire. L'ectolophe de M1/ est assez détérioré ; on voit cependant qu'elle devait être moins allongée et possédait des styles encore plus faibles. De P4/ il ne subsiste que l'angle postéro-interne ; on peut seulement observer que protocône et hypocône étaient bien séparés. La seule donnée ostéologique observable sur le spécimen est le départ du zygoma qui laisse supposer que le bord antérieur de l'orbite devait être aligné sur le milieu de M2/.
Les molaires du spécimen V5566 sont très dégradées et fragmentaires ; elles semblent avoir eu des dimensions un peu plus faibles que celles du maxillaire précédent (Tableaux 1 - 2). Une partie de l'ectolophe de M1/ est conservée ; il est très incliné, ce qui dénote une faible hauteur de couronne; le cingulum labial est net, il s'atténue vers le milieu de l'ectolophe dont le renflement à ce niveau suggère la présence d'un fort mésostyle. Le protocône, visible sur les 3 molaires est massif, allongé transversalement avec une facette d'usure triangulaire.
La P1/ du spécimen V5588 est trapézoïdale, rétrécie à l'avant, son ectolophe est très abrasé ; un large cingulum antéro-interne et postérieur délimite une cuvette centrale ; il est accidenté par deux renflements, qui forment deux petites cuspides, antéro-interne et postéro-interne. DP2/ est très brachyodonte avec un émail mince ; elle est entièrement molariforme, son mésostyle semble avoir été bien développé, le cingulum interne est continu mais peu proéminent.
| Matériel | SUP | L | l | D | d |
| V5588 | P1/ | 12,9 | 11,4 | ||
| DP2/ | 18,0 | 19,0 | 22,8 | 19,4 | |
| V5573 | P3/ | 20,7 | 26,2 | 27,9 | 22,7 |
| P4/ | - | 28,7 | 30,8 | - | |
| V5599, V5565 |
M1/ | - - |
- 28,9 |
- - |
30,6 ~28,5 |
| V5599, V5566 |
M2/ | 32,0 - |
32,3 ≥30,0 |
37,8 (≥31,0) |
32,4 (29,5) |
| M3/ | 31,9 28,3 |
33,5 ≥29,5 |
40,3 (≥35,4) |
33,3 (30,0) |
Tableau 1 : Mesures (en mm) des dents jugales supérieures de Palaeotherium muehlbergi thaleri. L : longueur de l'ectolophe ; l : largeur maximale ; D : grand diamètre du paracône à l'hypocône ; d : mesure du diamètre perpendiculaire à D (entre parenthèses : mesures estimées d'après le contour au collet).
| Matériel | INF | L | l1 | l2 |
| V5596 | P/3 | 21,0 | 12,4 | 12,9 |
| V5584 | P/4 | 20,0 | 13,0 | 14,9 |
| V5583 | /P | 18,7 | 12,7 | 13,6 |
Tableau 2 : Mesures (en mm) des dents jugales inférieures de Palaeotherium muehlbergi thaleri. L : longueur ; l1 : largeur du trigonide ; l2 : largeur du talonide.
Les dents jugales inférieures sont des prémolaires. La P/4 V5584 présente un croissant du trigonide assez anguleux avec l'indice d'une crête externe sur le protoconide ; les cingulums labial et lingual sont hauts mais peu en relief. La dent V5596 est interprétée comme une P/3 à cause de ses dimensions un peu plus faibles; le bord externe du protoconide est oblique, mais il n'y a pas de crête postéro-externe à ce niveau. La dent V5583, très usée et ébréchée, n'apporte pas de données complémentaires.
Discussion : La détermination spécifique est fondée principalement sur les dents maxillaires. Tous ces spécimens appartiennent visiblement à un même taxon. Les dimensions des dents jugales supérieures, la section caractéristique de leur protocône et la molarisation des prémolaires, ne peuvent s'accorder qu'avec l'espèce P. muehlbergi.
Les quelques caractères osseux observables confirment pleinement cette attribution. La tubérosité maxillaire très haute, l'arcade zygomatique très large et fortement rétrécie à l'avant sont en effet typiques de cette espèce (, 1992).
Au niveau subspécifique, notion introduite chez ce genre sur une base biochronologique (, 1968 ; , 1985), les prémolaires se distinguent de P. m. praecursum par la présence de costulations nettes sur les lobes de l'ectolophe, par une meilleure séparation entre protocône et hypocône et par la liaison entre métaconule et hypocône. De la sous-espèce nominale, elles se distinguent par leur largeur nettement moins grande, par des mésostyles moins proéminents sur les prémolaires et les deux premières molaires, et surtout par un surplomb lingual de la couronne des prémolaires assez accentué (, 1985).
Les dimensions de ces dents dépassent légèrement les valeurs observées chez P. m. thaleri, mais l'écart ne semble pas significatif compte tenu du petit nombre de spécimens connus pour ce taxon et parce que leur intégration à celui-ci ne génèrerait que des coefficients de variation modérés, inférieurs ou égaux à 6.
Sous-genre Franzenitherium , 1992
Palaeotherium (Franzenitherium) duvali , 1853
Matériel :
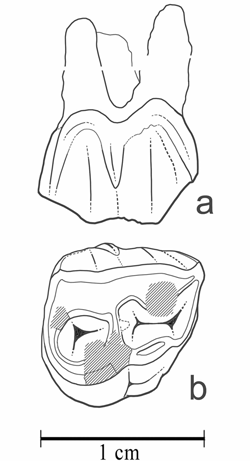
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 13 : Palaeotherium (Franzenitherium) duvali. MHNG GEPI V5597, P2/ sup. droite (a : vue labiale ; b : vue occlusale).
Description : La présence de ce taxon est attestée par une prémolaire supérieure de très petite taille (L : 9,6 ; l : 10,0 ; D : 11,8 ; d : 9,8), assez abrasée, remarquable par le retrait important de l'angle antéro-interne qui lui donne une forme quasiment triangulaire et révèle qu'il s'agit d'une P2/. L'ectolophe comporte deux cuspides rapprochées et soulignées sur la face labiale par une convexité bien marquée. Le cingulum labial est faible et donne naissance à un mésostyle bien caractérisé quoique peu saillant et court, qui ne dépassait probablement pas le tiers de la hauteur de la couronne. Le protolophe est bas, peu différenciable d'un cingulum antérieur, sans trace de paraconule ; il aboutit à un protocône allongé et raccordé à l'hypocône, qui est la cuspide interne dominante et qui se rattache à l'ectolophe par un métalophe transversal étroit. Le cingulum interne est pratiquement inexistant, développé seulement en face du sillon intercuspidien. Le surplomb lingual de l'hypocône est assez marqué. Le fragment de molaire de Palaeotherium V5601 offre des dimensions compatibles avec le même taxon.
Discussion : Les dimensions de cette dent, nettement inférieures à celles de P. siderolithicum, ne sont compatibles qu'avec les trois plus petites espèces de Palaeotherium. Les dents de P. eocaenum d'Egerkingen, interprétées comme des P2/, présentent à la fois un contour moins triangulaire, un hypocône moins développé et une absence totale de mésostyle. La P2/ du Guépelle MNHN EBP-78 est même dépourvue d'hypocône avec un protocône central et un métaconule rudimentaire. L'éventuelle attribution de la dent V5597 à cette espèce, d'ailleurs cantonnée à l'Éocène moyen (MP 13 et 14), peut donc être écartée. La P2/ est bien représentée dans les collections chez P. lautricense ; elle est généralement encore plus petite que le spécimen en question et moins triangulaire ; l'ectolophe est dépourvu de côtes externes et de mésostyle ; enfin, protocône et hypocône sont mieux séparés.
Au contraire le spécimen correspond dans toutes ses caractéristiques à la P2/ du lectotype de P. duvali de Montmartre (, 1968, Pl. 2, fig. 1). Cette détermination concorde en outre avec les indications biochronologiques fournies par le reste du matériel ; l'espèce est connue depuis (MP 17), voire même jusqu'au niveau MP19 (Mormont-Eclépens? d'après & , 2000).
La variabilité intraspécifique de cette dent n'est que médiocrement documentée ; on notera cependant que le développement du mésostyle relativement important pour une P2/, laisserait présager une forme plus progressive que duvali priscum d', dont le type, une P3/, n'en exhibe qu'une courte amorce. Au contraire, la liaison protocône - hypocône indiquant une molarisation un peu moins avancée que sur le maxillaire de Mähringen figuré par (1968, Pl. 2, fig. 2) serait en faveur d'un âge un peu plus ancien que ce gisement rapproché du MP 19.
La détermination du spécimen présente un intérêt paléogéographique. En effet, P. duvali, qui ne se trouve en concurrence avec siderolithicum que sur un territoire relativement limité (, 1968, carte p. 159) présente un domaine de répartition plus nordique que celui-ci, et était connu jusqu'ici seulement dans l'Île de Wight, le Bassin parisien, l'Allemagne du Sud et la Suisse ; notre site alpin apparaît donc comme son gisement le plus méridional.
Genre Plagiolophus , 1847
Plagiolophus sp. 1
Fig. 14 ![]() ;
Tableau 3 ; Pl. 3
;
Tableau 3 ; Pl. 3 ![]() ,
figs. 2-3
,
figs. 2-3
Matériel :
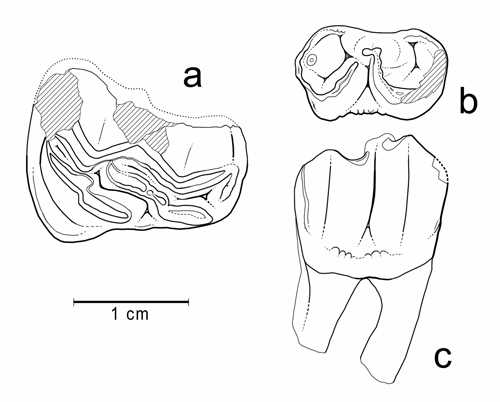
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
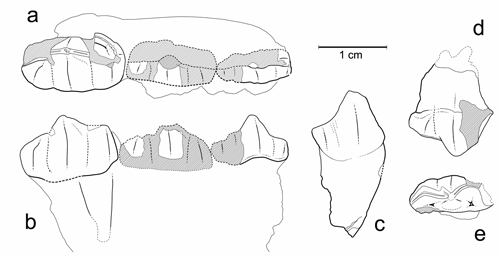
Figure 14 : Plagiolophus sp. 1. a : MHNG GEPI V5570 (pars), M3/ gauche (vue occlusale). b-c : MHNG GEPI V5572, P/4 droite (b : vue occlusale, c : vue labiale).
Description : Le maxillaire V5570 est complètement désagrégé, mais les dents ont conservé leur ordre et il paraît indiscutable qu'il s'agisse d'un unique individu. Les dents sont elles-mêmes incomplètes, surtout P3/ et P4/ dont il ne subsiste que l'angle antéro-interne, et elles ont perdu leur ectolophe sauf la M3/ où il est partiellement présent. Par suite, peu de mesures sont possibles (Tableau 3), mais la longueur M1/-M3/ peut cependant être estimée à environ 49 mm.
D'après ce que l'on peut voir de P2/ et P3/, leur protocône est prolongé distalement par une surface d'usure qui évoque un hypocône bas ou un cingulum postérieur relevé. Le cingulum interne des prémolaires semble avoir été large, mais interrompu sur le protocône. Les molaires semblent avoir eu du cément qui ne comblait cependant pas les vallées médianes. M3/ n'est pas très allongée, mais fortement rétrécie à l'arrière et son bord disto-labial n'est pas prolongé obliquement ; les cuspides internes sont très allongées et parallèles à l'ectolophe ; avec un degré d'usure de 2,5 le rapport Pa-Pr/l est de 30,2 (, 2004).
La P/4 V5572 mesure : L : 12,9 ; l1 : 7,7 ; l2 : 8,1 ; elle est peu usée (U = 1) et son taux d'hypsodontie I'H (Hprotoconide / lmax) peut être évalué à 1,56. Le talonide est un peu plus bas que le trigonide mais à peine raccourci ; on note la présence d'un petit hypoconulide bas séparé de l'hypolophide. La molaire inférieure V5602 (L : 13,9 ; l1 : 6,9 ; l2 : 7,1) est un germe un peu érodé au collet ; de ce fait on ne peut pas évaluer son coefficient d'hypsodontie ; sa largeur est très faible mais sans doute un peu minorée.
Quant à la canine V5571, d'ailleurs de petite taille (8x6 mm), elle n'apporte aucun élément de détermination spécifique en raison des variations importantes, notamment d'origine sexuelle, de ce type de dent (, 2004).
| Matériel | SUP | L | l | D | d |
| V5570 | P2/ | - | 9,0 | 9,3 | - |
| P3/ | - | - | - | - | |
| P4/ | - | - | - | - | |
| M1/ | (≥14,2) | - | - | (≥14,0) | |
| M2/ | - | - | (≥17,6) | - | |
| M3/ | 19,0 | 14,9 | 20,0 | ~18,2 |
Tableau 3 : Mesures (en mm) des dents jugales supérieures de Plagiolophus sp. 1.
Discussion : La détermination spécifique du maxillaire est malaisée, notamment en raison de son mauvais état de conservation. Le faible allongement de M3/ suggère un rapprochement avec P. annectens, mais l'alignement parasagittal des cuspides est plutôt en faveur de minor. En outre, les dimensions des dents et l'écartement relatif entre paracône et protocône (Pa-Pr/l) sont intermédiaires entre P. annectens de Fons et P. minor de Célas ou de La Débruge (, 2004).
Pour ce qui est de la P/4, son hypsodontie paraît compatible avec les deux espèces, mais ses dimensions et surtout sa longueur, ainsi que le faible développement de l'entoconide s'accorderaient mieux avec P. annectens. Même en tenant compte de l'érosion du collet, la M/1-2 apparaît au contraire trop étroite pour cette espèce. On peut présumer que ces divers spécimens appartiennent à un même taxon ; tous présentent le même caractère ambigu qui interdit de trancher entre P. annectens et P. minor.
Plagiolophus sp. 2
Pl. 3 ![]() ,
fig. 4
,
fig. 4
Matériel : MHNG GEPI V5606 : M3/ droite.
Description : La M3/ est une dent de grande taille (L : 26,5 ; l : >18,2 ; D : 29,3 ; d : 21,0), très usée (U = 3) et dont l'ectolophe est érodé par suite d'exposition aux intempéries en surface. Sa forme générale est rectangulaire et allongée, mais le bord distal de la dent n'est pas prolongé obliquement vers le collet. Le protolophe apparaît rectiligne, sans débordement du paraconule vers l'arrière. L'hypocône est légèrement moins volumineux que le protocône et un peu en retrait. Le métalophe est soudé à la paroi antéro-interne du métacône. A cause de l'abrasion, il est impossible d'évaluer le coefficient d'hypsodontie, le degré d'allongement des cuspides, ni le resserrement de celles-ci dans le plan frontal (distance Pa-Pr), tous caractères essentiels pour la détermination spécifique d'un Plagiolophus.
Discussion : Ses dimensions rendent la dent compatible avec plusieurs espèces de grande taille. Cependant, le diamètre D est nettement plus grand que chez ministri ou oweni et la largeur nettement plus faible que chez major ou fraasi. D'autre part, le paraconule déborde distalement le protolophe chez major, les M3/ de fraasi sont peu allongées et celles connues chez oweni ont un bord distal très oblique (, 2004).
Finalement, si la présence d'une seconde espèce de Plagiolophus dans le gisement est indéniable, le spécimen en question est néanmoins insuffisamment caractérisé pour permettre une détermination spécifique.
Ordre Artiodactyla , 1841
Famille Anoplotheriidae , 1821
Genre Anoplotherium , 1804
Anoplotherium laurillardi , 1851
Fig. 15 ![]() ;
Tableau 4 ; Pl. 3
;
Tableau 4 ; Pl. 3 ![]() ,
figs. 6-8
,
figs. 6-8
Matériel :
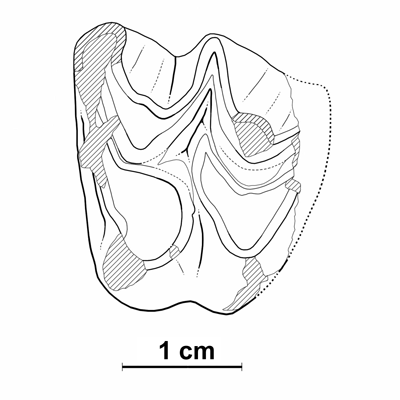
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 15 : Anoplotherium laurillardi. MHNG GEPI V5578, M2/ gauche (vue occlusale).
Description : La M2 V5578, assez fortement usée, est ébréchée au niveau du métastyle et la perte de longueur est estimée à 2 mm env. (L estimée : ?21,0 ; l : 23,3) Le faible retrait de l'hypocône montre qu'il ne s'agit pas d'une M3/, et la grande largeur relative de la dent est en faveur d'une M2/ plutôt que d'une M1/. La M1/ V5587 est extrêmement usée et abîmée du côté mésial ; son attribution au genre est fondée sur le gonflement du mésostyle ; l'aspect plutôt carré de la dent indique une M1/.
| Matériel | SUP | L | l |
| V5587 | M1/ | ≥22,0 | ≥23,0 |
| V5578 | M2/ | ≥21,0 | 23,3 |
| V5577 | M/ | - | ~23,0 |
Tableau 4 : Mesures (en mm) des dents jugales supérieures de Anoplotherium laurillardi.
Discussion : La morphologie de ces molaires est caractéristique du genre Anoplotherium et seule l'espèce A. laurillardi paraît correspondre à leurs mesures ( & , 2003).
Anoplotherium sp.
Pl. 3 ![]() ,
fig. 5
,
fig. 5
Matériel :
Description : Les deux molaires sont en très mauvais état ; V5568 est usée à l'extrême et la morphologie de la face occlusale a totalement disparu ; V5609, un peu moins abrasée, a perdu son trigonide et le talonide est un peu écrasé du côté interne. Il s'agit de dents de grande taille (V5568 : L : 43,1 ; l1 :17,5 ; l2 :16,3 ; l3 (hypoconulide) : 10,0. V5609 : L estimée : 40 à 42 ; l3 :9,9). Elles sont dépourvues de tout cingulum labial et lingual et l'orientation des plans d'usure montre que les cuspides internes, métaconide et entoconide, étaient proéminentes. V5608, trouvée à proximité et dans l'alignement de V5609, appartenait probablement au même individu. C'est une prémolaire allongée (mais un peu écrasée). Son trigonide est très ouvert du côté interne et le talonide, plus court, n'est pourvu que d'un petit entoconide bas. Ses dimensions (L : 19,5 ; l : 9,2) sont compatibles avec celles de la M3.
Discussion : Les dimensions de ces dents montrent qu'une deuxième espèce d'Anoplotherium, plus grande que laurillardi existe dans le gisement. Elles correspondent à celles des grandes espèces du genre, A. commune et A. latipes, mais leur état de conservation ne permet pas une détermination spécifique.
Famille Cebochoeridae , 1883
Genre Cebochoerus , 1852
Cebochoerus aff. fontensis , 1978
Fig. 16 ![]() ;
Tableau 5 ; Pl. 4
;
Tableau 5 ; Pl. 4 ![]() ,
figs. 1-2
,
figs. 1-2
Matériel :
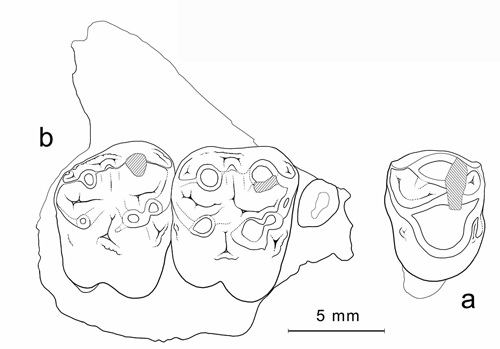
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 16 : Cebochoerus aff. fontensis. a : MHNG GEPI V5591, M2/-M3/ droites (vue occlusale). b : MHNG GEPI V5586, P4/ gauche (inversée) (vue occlusale).
Description : Cette petite forme est caractérisée par des dents aux cuspides bunodontes, très globuleuses. Leurs facettes d'usure sont tout à fait horizontales. La P4/ comporte 2 cuspides principales. Le paracône, dépourvu de cingulum labial, est cependant limité par des bords mésial et distal nets et un peu soulevés. Du côté interne, il est enserré entre les deux crêtes émanant du protocône, qui est très abrasé et légèrement déporté vers l'avant ; la crête antérieure basse (protolophe) présente une étroite facette de dentine dénudée qui signe la présence d'un faible paraconule ; la crête postérieure rejoint le bord disto-labial de la dent. Il n'y a pas de cingulum lingual.
Le maxillaire V5591 comporte l'arrière de la tubérosité maxillaire et la naissance de l'arcade zygomatique qui est fortement divergente. Les 2 molaires sont moins usées que la P4/ V5586, ce qui laisse supposer qu'elles appartenaient à un autre individu. Outre les 4 tubercules principaux, M2/ possède un paraconule bien développé ; les cuspides internes sont très inclinées ; les cingulums sont limités au versant externe du métacône, à l'avant du protocône et à l'ouverture du medivallum. Les racines ne paraissent pas très divergentes et la racine linguale, quoique creusée par un profond sillon, n'est pas dédoublée. M3/, qui présente la même structure, est légèrement plus petite que la M2/ et plus trapézoïdale avec un lobe postérieur moins large ; elle ne possède pas de conule central.
| Matériel | SUP | L | l |
| V5586 | P4/ | 5,2 | 6,8 |
| V5591 | M2/ | 6,4 | 8,3 |
| M3/ | 5,9 | 7,7 |
Tableau 5 : Mesures (en mm) des dents jugales supérieures de Cebochoerus aff. fontensis.
Discussion : On est en présence d'un Cebochridae de petite taille, nettement plus petit que les espèces Cebochoerus lacustris, helveticus ou minor (voir par exemple , 1986, text-fig. 57). L'appartenance au genre Acotherulum est écartée à cause de la forte bunodontie, de la présence d'un paraconule relativement gros et de la faible divergence des racines linguales qui ne sont pas dédoublées. On a donc affaire à un Cebochoerus (et plus précisément au sous-genre Gervachoerus), en raison de la petite taille, de la forte bunodontie et de l'absence d'un conule central sur M3/. C'est finalement de C. fontensis que cette forme se rapproche le plus. Cependant les cuspides linguales sont moins inclinées qu'à Fons, donc moins brachyodontes et les dents plus allongées transversalement (, 1986, text-fig. 56), signes d'un stade évolutif un peu plus avancé.
Famille Choeropotamidae , 1845
Genre Choeropotamus , 1821
Choeropotamus sp.
Figs. 17 ![]() -
18
-
18 ![]() -
19
-
19 ![]() ;
Tableaux 6 - 7 -
8 ; Pl. 4
;
Tableaux 6 - 7 -
8 ; Pl. 4 ![]() ,
figs. 3-8
,
figs. 3-8
Matériel :
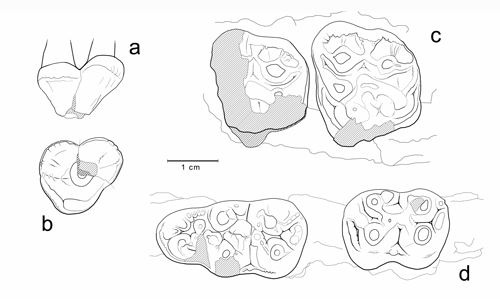
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 17 : Choeropotamus sp. a-b : MHNG GEPI V5590, P3/ sup. gauche (a : vue labiale, b : vue occlusale). c : MHNG GEPI V5567, M2/-M3/ gauches (vue occlusale). d : MHNG GEPI V5575, M/2-M/3 droites (vue occlusale).
Description : L'ensemble de ce matériel est homogène et correspond visiblement à un seul taxon de grande taille. Les molaires supérieures et inférieures ont une morphologie en tous points conforme à celles des grands Choeropotamus avec des tubercules bulbeux et bas ; les cuspides labiales des inférieures ne sont que vaguement prolongées par des crêtes en croissant.
La prémolaire supérieure interprétée comme une P3/ se présente, après restauration, comme une dent triangulaire à sommet antérieur, élargie à l'arrière ; elle possède trois racines (les deux postérieures sont séparées sur toute leur longueur) ; la couronne est un peu plus haute que celle des molaires ; la cuspide principale (paracône) présente une faible crête postérieure bien marquée ; le cingulum interne est bas, peu proéminent mais renflé à l'angle postéro-interne de la dent en forme de cuspide accessoire. Quant à la P2/, elle est biradiculée, à peine élargie postérieurement et elle offre une structure similaire mais simplifiée.
La P/4 est allongée, biradiculée, sa cuspide centrale (protoconide) est haute avec des crêtes sagittales émoussées, la postérieure dédoublée ; le cingulum circulaire est très faible, légèrement renflé à sa jonction avec les crêtes de la cuspide.
La branche horizontale de la mandibule est partiellement conservée sur le spécimen V5594 qui est un individu âgé. Elle ne semble pas avoir été très haute, probablement pas plus de 37 mm sous M/3. Le bord antérieur de la branche montante est oblique et reporté nettement en arrière de cette dent (>15 mm). Ces caractéristiques sont conformes à celles de la mandibule de Mormoiron figurée par (1859, Pl. 31, fig. 5) ; la position plus reculée de la branche montante pourrait être liée au grand âge de l'individu.
| Matériel | SUP | L | l1 | l2 |
| V5589 | P2/ | ~19,5 | 9,0 | 10,4 |
| V5590 | P3/ | ~16,0 | 9,9 | 15,0 |
| V5567 | M2/ | ~19,8 | ~20,5 | (19,5) |
| M3/ | 20,6 | ~24,9 | 20,3 |
Tableau 6 : Mesures (en mm) des dents jugales supérieures de Choeropotamus sp. l1 : largeur du lobe antérieur (paracône - protocône) ; l2 : largeur du lobe postérieur (métacône - hypocône).
| Matériel | INF | L | l1 | l2 | l3 |
| V5576 | P/4 | 16,4 | 10,6 | 11,8 | |
| M/1 | (19,0) | 12,4 | (12,8) | ||
| V5567 | M/2 | 20,4 | 15,5 | 14,9 | |
| M/3 | 29,4 | 15.6 | 14,7 | 10,8 | |
| V5569 | M/3 | 28,5 | 17,2 | 15,9 | 11,8 |
| V5594 | M/3 | - | 16,3 | 15,5 | 11,9 |
Tableau 7 : Mesures (en mm) des dents jugales inférieures de Choeropotamus sp. l1 : largeur du trigonide ; l2 : largeur du talonide ; l3 : largeur de l'hypoconulide.
| Réf. bibliographiques | Gisements | Espèces | SUP |
| , 1917, Pl. 16-1 | Euzet | C. depereti | 1.21 |
| , 1859, Pl. 31-1 | La Débruge | C. affinis | 1,00 |
| , 1859, Pl. 32-1 | Gypse | C. parisiensis | 0,86 |
| (moulage coll. UMII) | 0,94 | ||
| V5567 | Perdrix | 1,13 | |
| INF | |||
| Roc de Santa | C. sudrei | 1,28 | |
| , 1917, Pl. 16-3 | Euzet | C. depereti | 1,39 |
| , 1917, Pl. 16-5 | 1,37 | ||
| , 1917, Pl.16-6 | 1,31 | ||
| , 1859, Pl. 31-6 | La Débruge | C. affinis | 1,53 |
| V5575 | Perdrix | C. sp. | 1,27 |
Tableau 8 : Rapports de surface entre M3 et M2 chez les diverses espèces de Choeropotamus.
Discussion : La détermination spécifique de ce
matériel des Grandes Platières est difficile car les Choeropotamus sont des animaux toujours rares et les normes de
variations des deux grandes espèces de l'Éocène supérieur sont mal documentées.
Les molaires présentent des mesures intermédiaires entre C. affinis de
la Débruge et C. parisiensis du Gypse parisien et de St-Capraise ; la M3/ par exemple est plus proche de
parisiensis de Paris, les M/3 se tiennent à la limite supérieure de longueur
d'affinis, elles sont plus petites que la M/3 de St-Capraise (Fig.
18 ![]() ). Ce critère de différenciation reste donc un peu ambigu.
). Ce critère de différenciation reste donc un peu ambigu.
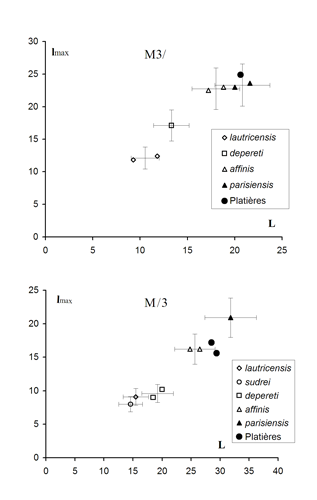
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 18 :
Comparaison des mesures de M3/ et M/3 chez les diverses espèces de Choeropotamus.
Les points représentent les spécimens connus chez les différents taxons. Les limites de variation figurées sont conjecturales. Elles sont destinées à mettre en évidence quels seraient les recouvrements entre ces taxons, dans les limites ±2s autour des valeurs connues si lon part de lhypothèse quun coefficient de variation de 7 représenterait une valeur moyenne de variabilité chez une espèce donnée.
M3/ :
C. lautricensis : Robiac (FSL, FMO) ; C. depereti : Euzet
(, 1917, Pl. 16-1) ;
C. affinis : la Débruge (, 1859,
Pl. 31, FMO 4325) ; C. parisiensis : Gypse de Paris (FMO), St-Capraise (FPO SCE-1) ;
Choeropotamus sp. des Perdrix : MHNG GEPI V5567.
M/3 : C. lautricensis : Robiac (FSL 4862) ; C. sudrei : Roc de Santa (IPS RS-439) ;
C. depereti : Euzet (, 1917, Pl.
16-5 & 16-6) ; C. affinis : la Débruge
(, 1859, Pl. 31, FMO 4326) ;
C. parisiensis : St-Capraise (FPO SCE-89) ; Choeropotamus sp. des Perdrix : MHNG GEPI V5575, MHNG GEPI V5569.
Deux autres caractères semblent conférer au
matériel savoyard un aspect un peu archaïque. Tout d'abord les rapports de
surface entre les M3 et les M2 supérieures et inférieures paraissent
s'inverser au cours du temps chez les Choeropotamus : au maxillaire, la M3/ est nettement plus grande que la M2/ au niveau d'Euzet, elle devient plus petite que cette dent à la Débruge et à Montmartre. Au contraire, la surface relative des M/3 semble s'accroître, par suite notamment d'un élargissement de l'hypoconulide. Pour ces deux critères, les molaires des Grandes Platières offrent des proportions archaïques
(Fig. 19 ![]() ).
).
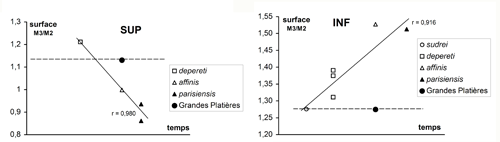
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 19 : Rapports de surface des M3 par rapport aux M2 chez Choeropotamus. Mesures de surface réelle des faces occlusales (plus grand contour).
En second lieu, la P3/ (V5590) présente après reconstitution une morphologie intermédiaire entre les P3/ d'Euzet (, 1917, Pl. 16, fig. 1) et du Gypse parisien (, 1859, Pl. 32, fig. 1).
Finalement le matériel en question présente une association de caractères assez originale, mais à cause de son état fragmentaire on s'est abstenu de toute attribution spécifique.
Artiodactyla indet.
Fig. 20 ![]() ; Tableau 9 ; Pl. 4
; Tableau 9 ; Pl. 4 ![]() ,
figs. 9-12
,
figs. 9-12
Matériel :
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 20 : Artiodactyla indet. du gisement des Perdrix. a-b : MHNG GEPI V5598, P/2-P/4 droites (a : vue occlusale ; b : vue labiale). c : MHNG GEPI V5582, /C droite (vue labiale). d-e : MHNG GEPI V5595, P2/ gauche (d : vue labiale ; e : vue occlusale).
Description : On présume que ces pièces,
malheureusement très dégradées, se rapportent à un seul et même taxon de
taille relativement grande (Tableau 9). La canine V5582 est légèrement prémolariforme,
pourvue d'une cuspide principale pointue et d'un talonide très court et bas, séparé
de celle-ci par un sillon mousse vertical, et dont le faible développement écarte
l'éventualité qu'il s'agisse d'une prémolaire. L'incisive V5581 est carénée
antéro-postérieurement avec une pointe arrondie, et dépourvue de cingulum
labial ; la présence d'un indice de talon distal délimité par une concavité
verticale permet de la rapprocher du spécimen précédent. Les 3 prémolaires
V5598 ont été particulièrement érodées du côté lingual par exposition aux
intempéries. La figure 20d-e ![]() rend compte des rares structures qui demeurent
observables. P/4 présente un aspect trilobé ; le protoconide est bunodonte et
pointu ; le paraconide, brisé au sommet, est un peu plus bas, allongé
transversalement et prolongé du côté interne par un cingulum proéminent ; du
lobe distal, il ne subsiste que la paroi labiale renflée. P/3 offre le même
aspect général, mais son versant interne est écaillé et l'on ne peut juger
de sa largeur que dans la région du collet. P/2 encore plus simple, semble
avoir été très étroite avec une unique cuspide centrale pointue. La dent
V5595 interprétée comme une P2/ est monoradiculée ; sa cuspide principale
(paracône) est prolongée par une crête distale arrondie jusqu'à une large
facette d'usure oblique ; elle présente des crêtes accessoires antérieures
labiale et linguale qui enserrent un lobe antérieur bas légèrement
crescentiforme. La dent V5604, qui est probablement une DP3/, devait être étroite
dans sa partie antérieure ; la cuspide centrale (paracône), basse, est prolongée
distalement par un talon élargi pourvu sans doute de 2 cônes bas alignés
transversalement.
rend compte des rares structures qui demeurent
observables. P/4 présente un aspect trilobé ; le protoconide est bunodonte et
pointu ; le paraconide, brisé au sommet, est un peu plus bas, allongé
transversalement et prolongé du côté interne par un cingulum proéminent ; du
lobe distal, il ne subsiste que la paroi labiale renflée. P/3 offre le même
aspect général, mais son versant interne est écaillé et l'on ne peut juger
de sa largeur que dans la région du collet. P/2 encore plus simple, semble
avoir été très étroite avec une unique cuspide centrale pointue. La dent
V5595 interprétée comme une P2/ est monoradiculée ; sa cuspide principale
(paracône) est prolongée par une crête distale arrondie jusqu'à une large
facette d'usure oblique ; elle présente des crêtes accessoires antérieures
labiale et linguale qui enserrent un lobe antérieur bas légèrement
crescentiforme. La dent V5604, qui est probablement une DP3/, devait être étroite
dans sa partie antérieure ; la cuspide centrale (paracône), basse, est prolongée
distalement par un talon élargi pourvu sans doute de 2 cônes bas alignés
transversalement.
| Matériel | L | l | |
| V5595 | P2/ | 12,4 | 6,1 |
| V5582 | /C | 10,8 | 6,6 |
| V5598 | P/2 | 11,1 | 4,8 |
| P/3 | 12,5 | 6,8 | |
| P/4 | 15,0 | 8,3 |
Tableau 9 : Mesures (en mm) des dents jugales de Artiodactyla indet.
Discussion : Ce matériel ne peut pas être rapporté aux autres grands artiodactyles du gisement. Les prémolaires de Choeropotamus sont beaucoup plus globuleuses, celles de Anoplotherium nettement plus grandes et moins étroites. Parmi les autres genres susceptibles d'être rencontrés, Catodontherium possède une P/3 très aplatie et une P/4 au contraire raccourcie (ce genre n'est en outre pas connu au-delà de Robiac). Chez Dichodon, les prémolaires inférieures sont extrêmement étroites, leur protoconide moins haut et la P/4 possède un métaconide qui ne semble pas apparaître sur V5598. Une certaine ressemblance avec un Mixtotherium pourrait être évoquée, mais les espèces de ce genre sont plus petites et le lobe central de P/4 est également dédoublé en protoconide et métaconide. Chez Dacrytherium, en dépit d'une certaine ressemblance morphologique de la canine, les prémolaires inférieures P/3 et P/4 ont un paraconide moins développé et plus en retrait et un lobe postérieur plus bas et plus court ; la plus grande espèce, D. ovinum que l'on connaît à Fons et Euzet, est en outre beaucoup plus petite (, 1978). Une espèce du genre Elomeryx, définie à la Débruge sous le nom de Hyopotamus crispus par (1859, Pl. 12, fig. 7) d'après 2 molaires supérieures, présenterait des dimensions compatibles avec le matériel des Grandes Platières, mais ses prémolaires, connues par d'autres gisements (, 1991), sont plus simples et moins allongées et la canine est dépourvue de talonide. Finalement, à défaut de pièces mieux conservées et plus caractéristiques, le statut de ce matériel demeure indéterminé et ne fournit aucune indication biochronologique.
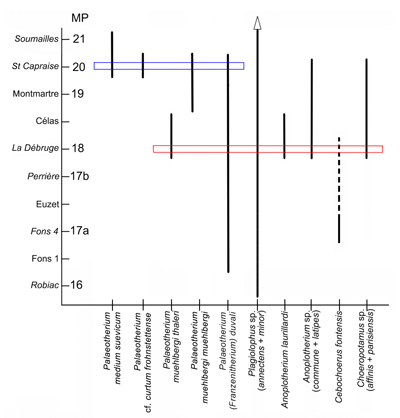
La composition faunique conduit à une estimation assez précise de la situation biostratigraphique du gisement des Perdrix
(Fig. 21 ![]() ).
).
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 21 : Répartition biochronologique des mammifères des gisements du massif de Platé. Les gisements-repères des niveaux paléomammalogiques (MP) sont indiqués en italique. La flèche surplombant Plagiolophus rappelle que la lignée minor se prolonge au-delà de Soumailles. Les pointillés indiquent l'extension supposée de l'espèce Cebochoerus fontensis. La zone bleue correspond à la faune décrite par et al. (1991) ; la zone rouge à celle du gisement des Perdrix.
La présence de deux Anoplotherium fournit une limite inférieure à la fourchette de datation, puisque ce genre n'est pas connu avant la Débruge, niveau-repère MP 18 dans l'échelle biochronologique des mammifères paléogènes (, 1987). Le très gros Choeropotamus va dans le même sens car il y a un hiatus important de stature par rapport à l'espèce d'Euzet, C. depereti.
On peut dégager aussi avec une bonne sécurité la limite biostratigraphique supérieure du gisement. En effet, Anoplotherium laurillardi n'est pas connu ultérieurement à la Débruge. De même, il est probable que la lignée de Cebochoerus fontensis ne dépasse pas ce niveau (, 1986 : text-fig. 62).
Les Palaeotheriidés confirment cette datation. La sous-espèce thaleri de la lignée de Palaeotherium muehlbergi est caractéristique de la Débruge et des gisements un peu plus récents des Grès de Célas (, 1985). Dans ces derniers, on connaît également un Plagiolophus apparemment intermédiaire entre P. annectens, connu jusqu'au niveau de Perrière, et P. minor, qui en est un probable descendant à la Débruge (, 2004).
Finalement, il semble donc que la situation biostratigraphique du nouveau gisement des Perdrix soit assez proche du niveau de la Débruge, ce qui correspondrait à un âge absolu d'environ -35,5 millions d'années ( et al., 1997 ; et al., 2004). Il est intéressant de faire un rapprochement avec le gisement décrit dans cette même série par et al. (1991) situé stratigraphiquement quelques dizaines de mètres plus haut et que ces derniers auteurs ont rapproché du niveau de Frohnstetten daté d'environ -34 millions d'années. Ces deux gisements seraient ainsi séparés par un intervalle de temps d'environ 1,5 millions d'années.
Mis à part les 5 fragments osseux rapportés à un crocodile, on compte 35 fragments de mammifères identifiables, dont on peut évaluer qu'ils représentent un nombre minimum de 19 individus se répartissant entre 8 palaeothériidés et 11 artiodactyles. Cependant, cette composition faunique ne reflète certainement que très imparfaitement la thanatocnose compte tenu des conditions d'exploitation du gisement. Ainsi, l'absence totale de micromammifères n'est probablement pas significative. Les espèces représentées fournissent toutefois quelques éléments d'appréciation du milieu dans lequel ont pu évoluer ces animaux.
Chez les palaeothériidés, la plupart des espèces de Palaeotherium sont réputées avoir fréquenté des milieux relativement fermés, boisés, voire forestiers (, 1992, p. 210) ; ce serait notamment le cas des espèces du groupe muehlbergi (, 1968, p. 158). Par contre, il est bien établi que les Plagiolophus occupaient dès l'Éocène supérieur des biotopes nettement plus ouverts (, 2004).
Pour ce qui est des anoplothériidés, la morphologie de leur squelette post-crânien et spécialement la configuration particulière des pattes, ont donné lieu, parallèlement à l'étude des dents, à diverses conjectures concernant le mode de vie de ces artiodactyles. Plusieurs auteurs, après (1835), ont envisagé qu'ils aient pu fréquenter des zones humides. (1983) est arrivé à la conclusion que l'un d'entre eux, Diplobune minor, avait développé une aptitude à se déplacer sur des sols ou des supports irréguliers, comme des roseaux ou des branchages couchés. La patte avant de Anoplotherium laurillardi présentant une morphologie tout à fait similaire ( & , 2003), on peut avancer l'hypothèse que cette espèce ait été inféodée elle aussi à des biotopes particuliers comme des roselières ou des plages boisées en bordure de rivières ou de points d'eau.
Quant au genre Choeropotamus, il est assez mal documenté. Son squelette post-crânien notamment n'est pas connu. La dentition bunodonte polytuberculée de type omnivore permet cependant de supposer que ces animaux partageaient les adaptations préférentielles des suiformes, à savoir des marécages boisés.
L'environnement humide, sinon aquatique, qui a présidé à la concentration de la thanatocnose d'origine est encore confirmé par la présence du crocodilien d'eau douce Diplocynodon.
On peut donc finalement supposer, à partir de
la documentation fossile dont nous disposons, reflet certainement très
incomplet de la thanatocnose, que la faune de vertébrés des Grandes Platières
vivait, avant le transport de ses restes sur le lieu du gisement des Perdrix,
dans un environnement assez varié où se juxtaposaient des zones boisées avec
des clairières ouvertes, à proximité de cours d'eau bordés de zones marécageuses
à roselières (Fig. 22 ![]() ).
).
Cette hypothèse est en bonne concordance avec les données sédimentologiques du gisement où la faune s'est ensuite trouvée déposée.
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Figure 22 : Environnements de dépôt des deux types de gisement à mammifères des Grandes Platières (adapté d'après et al., 1998). 1 & 2 : dépôts conglomératiques et palustres. 3 : couches à cérithes (niveaux saumâtres à influences marines sporadiques. 4 : grès calcaire à nummulites (avec figures tidales).
Les auteurs expriment leur reconnaissance envers les rapporteurs, Laureline et Damien (Paléojura, Porrentruy) ainsi que Loïc (Muséum d'Histoire naturelle de Bâle), dont les remarques constructives ont permis l'amélioration de cette publication.
P.A., S.L. & H.D. (1991).- The inception and early evolution of the North Alpine Foreland Basin.- Basin Research, London, vol. 3, p. 143-163.
E. (1978).- La poche à phosphate de Ste-Néboule (Lot) et sa faune de Vertébrés du Ludien supérieur. 4 - Crocodiliens.- Palaeovertebrata, Montpellier, vol. 8, p. 191-199.
L.W. (1943).- La Nappe de Morcles entre Arve et Rhône.- Matériaux pour la Carte Géologique de la Suisse, Berne, (nouvelle série), 79e livraison, 146 p.
P., G., P. & M. (1973).- Manifestation de mouvements tectoniques anté-oligocènes dans la partie sud de l'arc de Castellane : les formations conglomératiques du bassin tertiaire de Jabron (Var).- Comptes Rendus des séances de l'Académie des Sciences, Paris, (série D), vol. 276, p. 2361-2364.
S.L. & P.A. (1995).- Recognition of forebulge unconformities associated with early stage foreland basin development: example from the North Alpine Foreland Basin.- American Association of Petroleum Geologists, Bulletin, Boulder, vol. 79, p. 1495-1514.
G. (1835-1836).- Recherches sur les ossemens fossiles où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruits les espèces.- D'Ocagne, Paris, Texte (1835): 687 p., Atlas (1836): 161 Pls.
M. (2002).- Le Pays du Mont-Blanc - 9 itinéraires à travers paysages et roches du Mont-Blanc et du Haut-Faucigny.- Gap éditeur, La Ravoire, 239 p.
C. (1917).- Monographie de la faune de mammifères fossiles du Ludien inférieur d'Euzet-les-Bains (Gard).- Annales de l'Université de Lyon, (Nouvelle Série), I. Sciences, Médecine, fasc. 40, 228 p.
G., B. & S. (1997).- Sur l'âge numérique des faunes de mammifères du Paléogène d'Europe occidentale, en particulier celles de l'Éocène inférieur et moyen. In : Actes du Congrès BiochroM'97.- Mémoires et Travaux de l'École Pratique des Hautes Études, Montpellier, vol. 21, p. 443-460.
M., W.H. & N.J. (1999).- Tertiary foreland sedimentation in the Southern Subalpine Chains, SE France: a geodynamic appraisal.- Basin Research, London, vol. 11, p. 315-336.
J.L. (1968).- Revision der Gattung Palaeotherium , 1804 (Palaeotheriidae, Perissodactyla, Mammalia).- Inaugural-Dissertation, Albert-Ludwigs Universität zu Freiburg im Breisgau, 181 p.
P. (1859).- Zoologie et Paléontologie françaises.- Arthus Bertrand éditeur, Paris, (2ème édition), 544 p.
S. (1999).- Controls on sedimentation in distal margin palaeovalleys in the Early Tertiary Alpine foreland basin, south-eastern France.- Sedimentology, Oxford, vol. 46, p. 357-384.
M. (1991).- Revision der europäischen Species der Gattung Elomeryx 1894 (Anthracotheriidae, Artiodactyla, Mammalia) - Odontologische Untersuchungen.- Palaeontographica, Stuttgart, Abteil A, vol. 220, p. 1-101.
R., M., B. & M. (1984).- Données nouvelles sur les mammifères et les nummulites de l'Éocène supérieur subalpin (synclinaux du Charbon et d'Entrevernes, Bauges, Savoie et Haute-Savoie).- Géobios, Lyon, vol. 17, p. 221-234.
J.J. (1986).- Mammals from the Bartonian (middle/late Eocene) of the Hampshire Basin, southern England.- Bulletin of the British Museum (Natural History), London, (Geology), vol. 39, p. 191-478.
J.J. & M. (2000).- The Eocene mammal faunas of Mormont, Switzerland. Systematic revision and resolution of dating problems.- Schweizerische Paläontologische Abhandlungen, Bâle, vol. 120, 143 p.
A., P., J.-L. & M. (1987).- Les caractères du détritisme paléogène aux abords du massif du Pelvoux (Alpes externes méridionales).- Géologie Alpine, Grenoble, mémoire hors série, n° 13, p. 319-328.
J.C. & P.A. (1996).- Importance of inherited rift margin structures in the early North Alpine Foreland Basin, Switzerland.- Basin Research, London, n° 8, p. 425-442.
H.P., J.R., H., F.M., J.J., S., J., J., U., A. & B. (2004).- The Paleogene Period. In : J.G., J.G. & A.G. (eds.), A geologic time scale.- Cambridge University Press, p. 384-408.
U. (1994).- DieWildstrubel-, die Hohgant-, und Sanetsch-Formation: drei neue lithostratigraphische Einhelten des Eoceans der helvetischen Decken.- Eclogae geologicae Helvetiae, Bâle, vol. 87, p. 789-809.
U. (1995).- Stratigraphie, Fazies und Palaeogeographie des Eoceans der helvetischen Decken der Westschweiz (Diablerets- und Wildhorn-Decke).- Eclogae geologicae Helvetiae, Bâle, vol. 88, p. 115-134.
L.A. (1826).- Lettre du Prof. L.A. au Prof. G. , sur les filons granitiques et porphyriques de Valorsine et sur le gisement des couches coquillières de Sales, des Fizs et de Platet.- Bibliothèque de l'Université des Sciences, Belles-Lettres & Arts, Genève, vol. 33, p. 62-92.
B. & J.-L (1975).- Précisions nouvelles sur le Tertiaire du massif de Platé (Haute-Savoie).- Géologie Alpine, Grenoble, t. 51, p. 83-127.
J.A. (1985).- Nouveaux gisements de Mammifères et Reptiles dans les Grès de Célas (Éocène sup. du Gard). Étude des Palaeothériidés (Perissodactyla, Mammalia).- Palaeontographica A, Stuttgart, B. 189, p. 171-225.
J.A. (1992).- Observations sur l'anatomie crânienne du genre Palaeotherium (Perissodactyla, Mammalia); mise en évidence d'un nouveau sous-genre, Franzenitherium.- Palaeovertebrata, Montpellier, t. 21, p. 103-224.
J.A. (2004).- Le genre Plagiolophus (Palaeotheriidae, Perissodactyla, Mammalia): révision systématique, morphologie et histologie dentaires, anatomie crânienne, essai d'interprétation fonctionnelle.- Palaeovertebrata, Montpellier, t. 33, p. 17-281.
J.A. & F. (2003).- Mammifères fossiles des Grès de Célas (Éocène supérieur du Gard): Découvertes récentes.- Bulletin de la Société d'Étude des Sciences Naturelles de Nîmes et du Gard, Nîmes, vol. 64, p. 18-30.
N. (1987).- European Reference Levels and Correlation Tables, International Symposium on Mammalian Biostratigraphy and Paleoecology of the European Paleogene, Mainz.- Münchner Geowissenschaftliche Abhandlungen, München, Reihe A, p. 13-31.
H.D. (1997).- Tectonostratigraphic model for underfilled peripheral foreland basins: an Alpine perspective.- Geological Society of America, Bulletin, Boulder, vol. 109, p. 324-346.
H.D. & P.A. (1992).- Vertical versus horizontal motions in the Alpine orogenic wedge: stratigraphic response in the foreland basin.- Basin Research, London, vol. 4, p. 215-232.
H.D., Z.R. & M.E. (1998).- Carbonate sedimentation during early foreland subsidence: the Eocene succession of the French Alps. In : V.P. & T.P. (eds.), Carbonate ramps.- Geological Society, London, Special Publications, n° 149, p. 205-227.
C. (1965).- Présence de Palaeotherium et de Pulmonés dans l'Éocène continental du Lauzanier (couverture sédimentaire de l'Argentera, Basses-Alpes).- Travaux du Laboratoire de Géologie, Université de Grenoble, t. 41, p. 229-246.
J. (1978).- Les Artiodactyles de l'Éocène moyen et supérieur d'Europe occidentale (Systématique et évolution).- Mémoires et Travaux de l'École Pratique des Hautes Études, Montpellier, vol. 7, 229 p.
J. (1983).- Interprétation de la denture et description des éléments du squelette appendiculaire de l'espèce Diplobune minor ( 1877) : apports à la connaissance des Anoplotheriinae , 1850. In : E., J.-M. & E. (éds), Communications données à l'occasion du 150e anniversaire de la mort de Georges .- Actes du Symposium paléontologique G. , Montbéliard, p. 439-458.
J. & B. (1993).- First discovery of an Homacodontinae (Artiodactyla, Dichobunidae) in the Middle Eocene of Western Europe: Eygalayodon montenati new genus, new species. Considerations on the evolution of primitive Artiodactyls.- Kaupia, Darmstädter Beiträge zur Naturgeschichte, Darmstadt, B. 3, p. 157-164.
F. (1998).- Sédimentologie et analyse séquentielle du Paléogène des Chaînes subalpines (Domaine helvétique, Haute-Savoie, France).- Terre et Environnement, (Section Sciences de la Terre), Université de Genève, n° 11, 189 p.
M., J.L. & J. (1991).- Sur l'âge des Couches à Cérithes ou Couches des Diablerets de l'Éocène alpin.- Eclogae geologicae Helvetiae, Bâle, vol. 84, p. 893-919.

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Planche 1 :
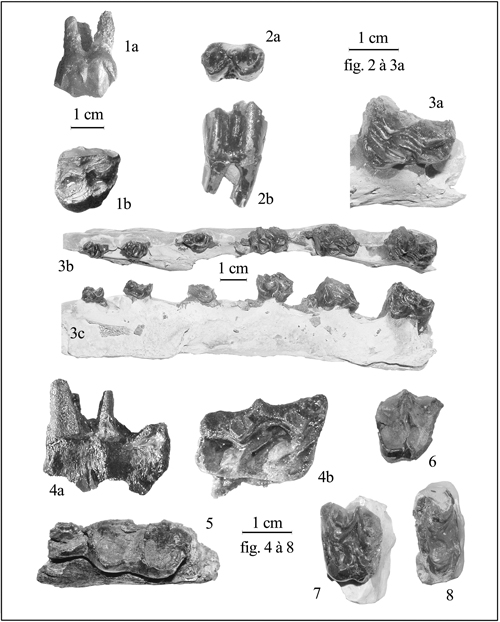
figs. 1-5 :
cf. Diplocynodon sp.
1 : MHNG GEPI V5580 : dent.
2 : MHNG GEPI V5585 : fragment de plaque
dermique.
3 : MHNG GEPI V5592 : fragment de plaque
dermique.
4 : MHNG GEPI V5593 : fragment de maxillaire
(4a, vue externe ; 4b, vue interne).
5 : MHNG GEPI V5605 : vertèbre caudale (5a,
vue antérieure ; 5b, vue dorsale).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
Planche 2 :
figs. 1-5 :
Palaeotherium muehlbergi thaleri ,
1985
1 : MHNG GEPI V5599 : maxillaire gauche avec
(P4/)-M1/-M3/ (1a, vue labiale ; 1b, vue occlusale).
2 : MHNG GEPI V5573 : fragment de maxillaire
droit avec P3/-P4/ (2a, vue labiale ; 2b, vue occlusale).
3 : MHNG GEPI V5588 : fragment de maxillaire
droit avec P1/-DP2/ (3a, vue labiale ; 3b, vue occlusale).
4 : MHNG GEPI V5583 : prémolaire inférieure
droite (vue occlusale).
5 : MHNG GEPI V5584 : P/4 gauche (5a, vue
labiale ; 5b, vue occlusale).
Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
fig. 1 : Palaeotherium (Franzenitherium) duvali , 1853
MHNG GEPI V5597 : P2/ droite (1a, vue labiale ; 1b, vue occlusale).
figs. 2-3 : Plagiolophus sp. 1
2 : MHNG GEPI V5572 : P/4 droite (2a, vue labiale ; 2b, vue
occlusale).
3 : MHNG GEPI V5570 : maxillaire gauche désagrégé avec (P2/)-(M3/)
(3a, M3/ en vue occlusale ; 3b, vue occlusale ; 3c, vue linguale).
fig. 4 : Plagiolophus sp. 2
MHNG GEPI V5606 : M3/ droite (4a, vue labiale ; 4b, vue occlusale).
fig. 5 : Anoplotherium sp.
MHNG GEPI V5568 : M/3 droite (vue occlusale).
figs. 6-8 : Anoplotherium laurillardi ,
1851
6 : MHNG GEPI V5587 : M1/ droite (vue occlusale).
7 : MHNG GEPI V5578 : M2/ gauche (vue occlusale).
8 : MHNG GEPI V5577 : lobe antérieur d'une M/ gauche (vue occlusale).

Cliquer sur la miniature pour agrandir l'image
[Click on thumbnail to enlarge the image]
figs. 1-2 : Cebochoerus aff. fontensis ,
1978
1 : MHNG GEPI V5591 : fragment de maxillaire droit avec M2/-M3/ (vue
occlusale).
2 : MHNG GEPI V5586 : P4/ gauche (vue occlusale).
figs. 3-8 : Choeropotamus sp.
3 : MHNG GEPI V5594 : fragment de mandibule gauche avec (M/3) (vue
latérale) [note : suite à la déformation du spécimen, la face occlusale
est vue obliquement]
4 : MHNG GEPI V5567 : fragment de maxillaire gauche avec (M2/)-M3/ (vue
occlusale).
5 : MHNG GEPI V5589-V5590 : P2/? et P3/? (un seul individu ?) (vue
occlusale).
6 : MHNG GEPI V5576 : P/4 et M/1 droites (vue labiale).
7 : MHNG GEPI V5569 : germe de M/3 gauche (vue occlusale).
8 : MHNG GEPI V5575 : mandibule droite avec M/2-M/3 (8a, vue occlusale ;
8b, vue labiale).
figs. 9-12 : Artiodactyla indet.
9 : MHNG GEPI V5598 : fragment de mandibule droite avec P/2-P/4 (9a, vue
occlusale ; 9b, vue labiale).
10 : MHNG GEPI V5595 : P2/ gauche (10a, vue labiale ; 10b, vue occlusale).
11 : MHNG GEPI V5582 : /C droite (vue labiale).
12 : MHNG GEPI V5581 : incisive (I/3 droite?) (vue labiale).