◄ Carnets Geol. 13 (M01) ►
![]()
Sommaire
[1. Introduction] [2. Situation géographique et contexte géologique]
[3. Lithologie et biostratigraphie des gisements] [4. Systématique]
[5. Discussion] [6. Conclusion] [Références bibliographiques]
et ... [Planches]
5b, rue de Camargue, La Cigalière, 05000 Gap (France)
Manuscrit en ligne depuis le 21 Janvier 2013
[Éditeur : Bruno ]
![]()
Les espčces Hemidiadema rugosum , 1846, et H. neocomiense (, 1869), petits échinides réguliers mal connus et sources de nombreuses confusions – de part leur rareté et l'insuffisance de leur description originale par leur auteur respectif –, sont révisées afin de clarifier leurs différences et leur statut taxonomique. Cette révision s'appuie sur l'examen de six exemplaires d'H. rugosum (dont l'holotype, non figuré par son auteur à l'origine) tous provenant du locus typicus et du stratum typicum ("Grčs ferrugineux" du Clansayésien [Aptien supérieur sensu gallico] de Grandpré, Ardennes) et sur un unique spécimen d'H. neocomiense provenant du stratum typicum ("Calcaire à Spatangues" de l'Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum)) de Ville-sur-Saulx (Meuse), région proche du locus typicus (Auxerre, Yonne)). Pour cette derničre espčce, un néotype est désigné en remplacement de l'holotype non retrouvé. Des remarques sur la paléoécologie et les répartitions géographique et stratigraphique de ces deux espčces sont précisées. La révision de ces deux espčces donne l'occasion de proposer une diagnose émendée du genre Hemidiadema , 1846, Glyphocyphidae peu connu.
Échinides ; Glyphocyphidae ; Hemidiadema ; Crétacé inférieur ; Hauterivien ; Aptien ; Bassin de Paris ; France.
A. (2013).- Hemidiadema rugosum , 1846, et H. neocomiense (, 1869) (Euechinoidea, Camarodonta, Glyphocyphidae) du Crétacé inférieur du Bassin parisien (France).- Carnets de Géologie [Notebooks on Geology], Brest, Mémoire 2013/01 (CG2013_M01), p. 1-57.
Revision of the species Hemidiadema rugosum , 1846, and H. neocomiense (, 1869) (Euechinoidea, Camarodonta, Glyphocyphidae) from the Lower Cretaceous of the Basin of Paris (France).- Hemidiadema rugosum , 1846, and H. neocomiense (, 1869) are small and poorly known "regular" echinoid species. They were the source of much confusion due to their rarity and the inadequacy of their original descriptions. Both species are revised here to clarify their differences and taxonomic status. This revision is based on six specimens of H. rugosum (including the holotype, not shown by its original author), all from the locus typicus and stratum typicum (Clansayesian [Upper Aptian sensu gallico] "Grčs ferrugineux" from Grandpré, Ardennes) and on a single specimen of H. neocomiense from the stratum typicum (Lower Hauterivian (? Lyticoceras nodosoplicatum Zone) "Calcaire à Spatangues" from Ville-sur-Saulx (Meuse), a locality near to the locus typicus (Auxerre, Yonne)). For this latter species, a neotype is designated to replace the holotype that is lost. Remarks on the paleoecology and stratigraphic and geographic distributions of these two species are given. The revision of these two species provides an opportunity to propose an emended diagnosis of the genus Hemidiadema , 1846, a poorly known member of the Glyphocyphidae.
Echinoids; Glyphocyphidae; Hemidiadema; Lower Cretaceous; Hauterivian; Aptian; Paris Basin; France.
(1846, p. 351) n'a donné qu'une diagnose trčs succincte du genre Hemidiadema qu'il propose ("Ce genre diffčre des Diadčmes par un seul caractčre, c'est que les aires ambulacraires ne sont composées que d'une seule rangée de tubercules"). De męme son espčce-type – Hemidiadema rugosum – n'est que trčs sommairement décrite ("Trčs petite espčce. Les tubercules ambulacraires sont aussi gros et męme plus gros que les tubercules
interambulacraires". Cette description est établie à partir de l'étude d'un exemplaire unique indiqué comme appartenant à la collection Paul .
(1862-1867, p. 533) puis
(1892b, p. 71) rappellent que cet échantillon – ayant servi à définir le genre et son espčce-type –, n'appartient pas à la collection mais en réalité à la collection . Aucune figuration de ce spécimen n'est donnée par , ni désignation explicite de celui-ci en tant que type précisée. Toutefois, la mention "Nov. Genus" inscrite sur le support cartonné oů l'échinide était collé (Fig.
1 ![]() )
prouve bien que cet échantillon est celui étudié par pour définir son genre et son espčce-type. Ce spécimen, qui doit ainsi ętre considéré comme un holotype par monotypie, est conservé dans les collections paléontologiques de la Faculté des Sciences de Lyon, sous le numéro d'inventaire EM 13095 (collections de l'École nationale supérieure des Mines de Paris).
)
prouve bien que cet échantillon est celui étudié par pour définir son genre et son espčce-type. Ce spécimen, qui doit ainsi ętre considéré comme un holotype par monotypie, est conservé dans les collections paléontologiques de la Faculté des Sciences de Lyon, sous le numéro d'inventaire EM 13095 (collections de l'École nationale supérieure des Mines de Paris).
Il faut attendre dans la Paléontologie Française
(1864, p. 543,
Pl. 1128, figs. 16-22) pour avoir une description détaillée et une figuration de cet holotype (Fig. 2 ![]() ), puis ensuite
(1892b, p. 70,
Pl. IV, figs. 5-6) pour avoir des remarques descriptives complémentaires et la figuration d'un autre individu
(Fig. 3
), puis ensuite
(1892b, p. 70,
Pl. IV, figs. 5-6) pour avoir des remarques descriptives complémentaires et la figuration d'un autre individu
(Fig. 3 ![]() ). Une diagnose détaillée du genre est donnée pour la premičre fois par
(1883, p. 103), reprise et complétée par
(1892b, p. 68-70). Trčs récemment cet holotype a fait l'objet d'une brčve étude ( & ,
2010, p. 21-24) publiée au moment męme de la soumission du manuscrit du présent article (décembre 2010). Le travail présenté par ces auteurs est dépourvu de toute rigueur
scientifique et rédactionnelle et souffre de graves omissions (absences d'une description littérale pertinente rendant compte des caractčres diagnosiques de l'espčce étudiée, de comparaison avec d'autres espčces voisines, de conclusion, de références bibliographiques
; synonymie loin d'ętre exhaustive ; omission des études historiques majeures concernant ce taxon).
). Une diagnose détaillée du genre est donnée pour la premičre fois par
(1883, p. 103), reprise et complétée par
(1892b, p. 68-70). Trčs récemment cet holotype a fait l'objet d'une brčve étude ( & ,
2010, p. 21-24) publiée au moment męme de la soumission du manuscrit du présent article (décembre 2010). Le travail présenté par ces auteurs est dépourvu de toute rigueur
scientifique et rédactionnelle et souffre de graves omissions (absences d'une description littérale pertinente rendant compte des caractčres diagnosiques de l'espčce étudiée, de comparaison avec d'autres espčces voisines, de conclusion, de références bibliographiques
; synonymie loin d'ętre exhaustive ; omission des études historiques majeures concernant ce taxon).

Click on thumbnail to enlarge the
image
[Cliquer sur la miniature pour agrandir l'image]
Figure 1 : Support cartonné écrit de la main d' sur lequel était collé l'holotype d'Hemidiadema rugosum , 1846, et authentifiant le statut holotypique de ce spécimen. On y lit : "Hemidiadema rugosa Nov. Genus" aprčs correction de rugosa par rugosum sans doute par ; "Grčs verts Grandpré (Ard)." ("Ard" pour Ardennes ; "PD" désignant les initiales de Paul ).

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour
agrandir l'image]
Figure 2 : Premičre figuration originale de l'holotype par monotypie d'Hemidiadema rugosum , 1846, par en 1864 (Paléontologie Française, pl. 1128, figs. 16-22 ; coll. , EM 13095) ; fig. 16 : vue latérale ; fig. 17 : vue apicale ; fig. 18 : vue orale ; fig. 19 : aire ambulacraire ; fig. 20 : aire interambulacraire ; fig. 21 : plaques ambulacraires et interambulacraires ; fig. 22 : disque apical.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour
agrandir l'image]
Figure 3 : Figuration originale d'un autre individu d'Hemidiadema rugosum , 1846, par (1892b, pl. IV, figs. 5-6 ; coll. , MNHN.F R62314) ; fig. 5 : vue apicale ; fig. 6 : plaques ambulacraires et interambulacraires.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature
pour agrandir l'image]
Figure 4 : Figuration originale de l'holotype par monotypie d'Hemidiadema neocomiense (, 1869) (, 1869, pl. 13, figs. 5-10 ; coll. ) ; fig. 5 : vue latérale ; fig. 6 : vue apicale ; fig. 7 : vue orale ; fig. 8 : aire ambulacraire ; fig. 9 : plaques interambulacraires ; fig. 10 : disque apical.
L'espčce Hemidiadema neocomiense (, 1869) a été incomplčtement décrite et figurée par son auteur (, 1869, p. 245,
Pl. 13, figs. 5-10) (Fig. 4 ![]() ). Dans son étude, ne précise pas explicitement si la description qu'il en donne est établie à partir d'un unique exemplaire. Toutefois, il ne donne les mesures et la figuration que d'un seul individu, et précise cette espčce trčs rare. Il semble donc que n'est étudié qu'un unique exemplaire – celui-là męme figuré –, et par conséquent celui-ci doit ętre considéré également comme un holotype par monotypie. Cette espčce n'a jamais refait l'objet d'études depuis, ni de figurations. Des recherches, effectuées au sein des collections paléontologiques de la Faculté des Sciences de Lyon et du Musée d'Histoire Naturelle d'Auxerre, n'ont pas permis de retrouver ce spécimen type de la collection qui doit ętre considéré comme perdu.
). Dans son étude, ne précise pas explicitement si la description qu'il en donne est établie à partir d'un unique exemplaire. Toutefois, il ne donne les mesures et la figuration que d'un seul individu, et précise cette espčce trčs rare. Il semble donc que n'est étudié qu'un unique exemplaire – celui-là męme figuré –, et par conséquent celui-ci doit ętre considéré également comme un holotype par monotypie. Cette espčce n'a jamais refait l'objet d'études depuis, ni de figurations. Des recherches, effectuées au sein des collections paléontologiques de la Faculté des Sciences de Lyon et du Musée d'Histoire Naturelle d'Auxerre, n'ont pas permis de retrouver ce spécimen type de la collection qui doit ętre considéré comme perdu.
Du fait de leur rareté, de leur petitesse et du peu d'études faites à leur sujet, Hemidiadema rugosum et H. neocomiense restent des espčces trčs mal connues. Les descriptions et figurations historiques lacunaires ont conduit à des confusions et des erreurs d'attribution (générique et spécifique), en particulier lors de ces derničres décennies (voir synonymie).
Le présent travail propose, pour chacune de ces deux espčces, une ré-étude complčte et détaillée à partir des rares spécimens retrouvés provenant de leur stratum typicum respectif : "Grčs ferrugineux" du Clansayésien (Aptien supérieur sensu gallico, voir et al., 2011) de Grandpré (Ardennes) pour H. rugosum et "Calcaire à Spatangues" de l'Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum) de Ville-sur-Saulx (Meuse) pour H. neocomiense. Un néotype est désigné pour l'espčce H. neocomiense, en remplacement de l'holotype non retrouvé. L'étude détaillée de ces différents spécimens permet également de proposer une diagnose émendée du genre Hemidiadema , 1846.
La présente étude servira ainsi de base par la suite pour aborder une révision des Hemidiadema du Crétacé de diverses autres localités (Sud-Est de la France, Espagne, Maroc et Tunisie), études en cours de préparation pour certaines d'entre-elles (, in prep. ; et al., in prep.).
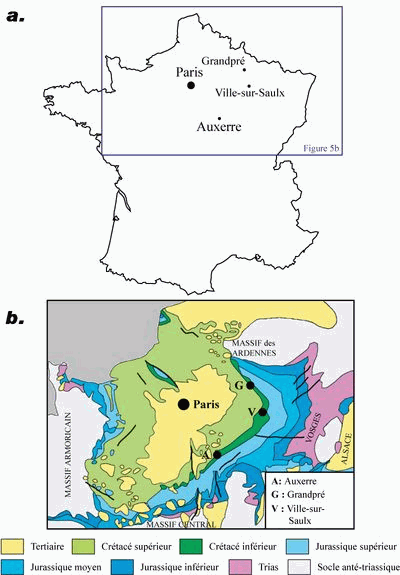
Les spécimens des deux espčces étudiées proviennent de la bordure orientale du vaste bassin sédimentaire de Paris (Fig.
5b ![]() ).
).
Les H. rugosum proviennent du locus typicus, la petite ville de Grandpré dans le département des Ardennes. Bien que les "Grčs ferrugineux" fussent observés en diverses localités de cette région, un seul affleurement de l'époque semble avoir fourni des H. rugosum : une miničre située au niveau des Bois des Loges, au Nord-Est immédiat de Grandpré.
L'holotype de l'espčce H. neocomiense provient des "environs" d'Auxerre dans le département de l'Yonne, n'en ayant pas donné de localisation plus précise. Le spécimen que nous décrivons dans ce travail provient de la localité de Ville-sur-Saulx dans le département de la Meuse (localité située à 150 km au Nord-Est d'Auxerre) (Fig.
5a ![]() ). L'individu a été récolté par P. dans l'une des nombreuses carričres souterraines situées au Nord-Est de Ville-sur-Saulx à son époque.
). L'individu a été récolté par P. dans l'une des nombreuses carričres souterraines situées au Nord-Est de Ville-sur-Saulx à son époque.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 5 : a. Situation géographique des localités citées : Grandpré : locus typicus de l'holotype d'Hemidiadema rugosum , 1846 ; Auxerre : locus typicus de l'holotype d'Hemidiadema neocomiense (, 1869) ; Ville-sur-Saulx : locus typicus du néotype d'Hemidiadema neocomiense (, 1869) désigné dans le présent travail. b. Carte géologique simplifiée du Bassin de Paris (d'aprčs la synthčse géologique du Bassin de Paris (1980) et la carte géologique de la France au 1/1 000 000čme (6čme édition, 1996)).
La biozonation ammonitique suit & (reporters) et al. (2009) et , & (reporters) et al. (2011).
Dans les Ardennes, les premiers dépôts crétacés discordants sur le
Jurassique terminal correspondent à la formation dite des "Grčs ferrugineux", plus localement connue aux environs de Grandpré sous le nom de "Minerai du Bois des Loges" ou "Miničre de Grandpré". Cette formation, discordante sur les "Calcaires compacts et marnes blanchâtres à Ostrea virgula" du Kimméridgien supérieur, a fait l'objet de nombreuses observations et discussions ( & ,
1842, p. 44-49 et 346-354
; & , 1873, p. 86-94
; de , 1868, p. 284 ; ,
1875, p. 57
; 1878, p. 240-246 ; ,
1925, p. 445-446 ; et plus récemment ,
1986, et , 1991). Épaisse d'environ 1 à 2 m, cette formation correspond à des grčs ferrugineux tendre à nombreux grains de silice, de limonite et de glauconie, inclus dans un ciment carbonaté jaunâtre (,
1986, p. 104-105). C'est de ce niveau que provient l'holotype sommairement décrit par (coll. , EM 13095) puis figuré par dans la Paléontologie Française, ainsi que l'exemplaire décrit et figuré par en
1892 (coll. , MNHN.F
R62314) et les quatre autres exemplaires de la collection (MNHN.F A20547, MNHN.F A33782, MNH.F A33783, MNHN.F A33784) étudiés dans ce travail. Vient par dessus la formation dite des "Sables verts" contenant des nodules phosphatés à sa base et des argiles gris-vert (,
1986, p. 104-105). Ces différentes formations ont pu ętre précisément datées par les ammonites. Les grčs ferrugineux contiennent Hypacanthoplites milletianus (d', 1841), H. milletioides , 1961, et H. peroni
(, 1905) (,
1875, p. 57
; 1878, p. 243, et com. pers. F. ), une association attribuée
au Clansayésien (Aptien supérieur sensu gallico). Les "Sables verts" qui surmontent ce minerai débutent par un lit de nodules phosphatés qui a fournit Leymeriella tardefurcata (d', 1841) et L. regularis (, 1780), une association de l'Albien inférieur basal (Zone à Leymeriella tardefurcata) (comm. pers. F. ). Les affleurements ponctuels (miničres) du Bois des Loges n'existant plus depuis déjà l'époque de , nous nous référons aux observations de &
(1842, p. 350 et 354),
(1878, p. 240) complétées des observations récentes de
(1986, p. 104-105) faites plus au Sud pour proposer une coupe détaillée de l'Apto-Albien de cette localité
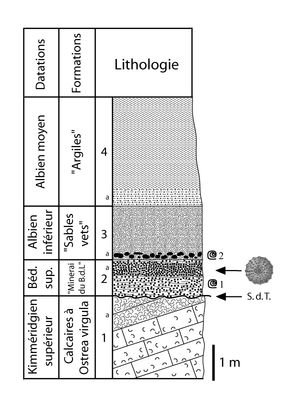
(Fig. 6 ![]() ).
).
L'holotype de l'espčce H. neocomiense et le néotype que nous désignons dans le présent travail proviennent tous deux de la męme formation : le "Calcaire à Spatangues" des anciens auteurs. Sur la bordure orientale du Bassin parisien les dépôts du Crétacé inférieur, en discontinuité sur les calcaires érodés du Jurassique terminal (Portlandien) aprčs la régression purbeckienne, ont fait l'objet de nombreuses études stratigraphiques et paléontologiques (, 1857 ; , 1916 ; , 1925 ; , 1933 ; , 1954, 1962 ; , 1976 ; , 1984 ; et al., 1987 ; et al., 2006). Le Crétacé inférieur y débute par des dépôts argilo-détritiques de milieux fluvio-deltaďques à marins littoraux (facičs "Wealdien" rapporté au Valanginien). Vient ensuite le "Calcaire à Spatangues", série épaisse de quelques mčtres seulement, représentant le maximum de transgression. Cette formation a livré à divers endroits des ammonites, en particulier à sa base (Acanthodiscus radiatus (, 1789), Leopoldia leopoldina (d', 1841), Breistrofferella castellanensis (d', 1840), Lyticoceras cryptoceras (d', 1840), Olcostephanus (Crespedites) carteroni (d', 1841), Neocomites neocomiensis (d', 1841), Saynella sp., Cymatoceras pseudoelegans (d', 1841), Cymatoceras neocomiensis (d', 1840), Eucymatoceras plicatus (, 1835), (, 1852, p. 475 ; , 1916, p. 32 ; , 1925, p. 121 ; et al., 1996, p. 31-32 ; , 2002). Acanthodiscus radiatus, Breistrofferella castellanensis et Leopoldia leopoldina caractérisent l'Hauterivien inférieur basal (Zone à Acanthodiscus radiatus, Horizon à Breistrofferella castellanensis), Lyticoceras cryptoceras la partie supérieure de l'Hauterivien inférieur (Zone à Lyticoceras nodosoplicatum). Le "Calcaire à Spatangues" appartient donc incontestablement à l'Hauterivien inférieur.
Cette formation est représentée par des calcaires détritiques à sa base et des calcaires marneux, organisés en bancs grossiers décimétriques à surface irréguličre ou en débit à miches, admettant des inter-bancs marneux de plus en plus fréquents à son sommet. L'un des fossiles le plus caractéristique est un échinide spatangide, Toxaster retusus (, 1816). Par-dessus repose la formation dite des "Argiles ostréennes", série marneuse dans laquelle (1916, p. 33) y distingue une partie inférieure avec Eucymatoceras plicatus (, 1835) et une partie supérieure avec Toxaster ricordeaui (, 1857). Longtemps rattachée au Barrémien inférieur, la datation précise de cette formation reste incertaine. Sa base pourrait ętre encore d'âge Hauterivien (supérieur ?) d'aprčs (1984, p. 406). Récemment et al. (2006) ont démontré par l'étude des kystes de dinoflagellés que la totalité des "Argiles ostréennes" correspondrait au passage Barrémien inférieur/Barrémien supérieur.
L'étude des nombreuses carričres souterraines aux environs de Ville-sur-Saulx a permis à
(1852, p. 467-469) et
(1925,
Pl. I) de suivre la succession lithologique du Crétacé inférieur de ce secteur avec précisions. Les affleurements n'étant plus actuellement visibles, nous reproduisons ici la coupe levée par dans sa thčse
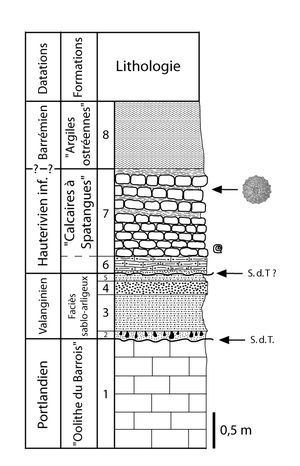
(Fig. 7 ![]() ).
).
Un exemplaire d'H. neocomiense – étiqueté comme provenant de l'Hauterivien de Ville-sur-Saulx (Meuse) – a été récemment retrouvé dans la collection P. (propriété de l'École nationale supérieure de Géologie de Nancy, mais en dépôt au Muséum-Aquarium de Nancy). P. a vraisemblablement récolté ce spécimen dans l'une des nombreuses carričres situées au Nord-Est de Ville-sur-Saulx, en pleine exploitation à son époque. La gangue qui obstrue l'aire péristomiale du spécimen correspond à une marne trčs finement sableuse, gris-claire, tendre, ce qui laisse à penser qu'il provient des intercalations marneuses de la partie supérieure du "Calcaire à Spatangues".
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Figure 6 : Coupe lithologique de l'Apto-Albien discordant sur le Jurassique terminal au Bois des Loges, prčs de Grandpré
[lithologie dessinée d'aprčs les observations de & (1842, p. 350, 354), (1878, p. 240) et
(1986, p. 104-105) ; informations biostratigraphiques d'aprčs
(1986, p.
104-105) et (com. pers.)].
1 : Calcaires compacts blanchâtres ŕ Ostrea virgula, avec lumachelle au sommet (a) ; 2 : Argiles sableuses riches en grains de silice, de limonite et de glauconie devenant plus compactes dans sa partie supérieure (a : "Grčs ferrugineux") ; 3 : Marnes verdâtres glauconieuses, trčs sableuses avec nodules phosphatés ŕ la base (a) ; 4 : Argile gris-vert ŕ vert, sableuse ŕ sa base
(a) ; S.d.T : Surface de transgression ;
@ 1 : Hypacanthoplites milletianus, H. milletioides et H.
peroni ;
@ 2 : Leymeriella tardefurcata et L. regularis.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Figure 7 : Coupe lithologique du Crétacé inférieur discordant sur le Jurassique terminal relevée ŕ l'entrée d'une des carričres de Ville-sur-Saulx (lithologie redessinée d'aprčs la figure I de la thčse de , 1925).
1 : Calcaires dolomitisés du Portlandien ; 2 : Lit d'ocre avec Teredo sp. ; 3 : Marnes sableuses avec oolithe ferrugineuse ; 4 : Grčs ferrugineux compact ; 5 : Marnes sableuses fossilifčres ; 6 : Calcaires oolithiques tendres, contenant ŕ la base des ossements de reptiles et de poissons ; 7 : Calcaires jaunes rognoneux ŕ
Toxaster retusus avec intercalations marneuses ŕ Exogyra aquila dans la partie supérieure ; 8 : Argiles ŕ
Liostrea leymeri ; S.d.T : Surface de transgression ;
@ : Acanthodiscus radiatus, Neocomites houdardi (ammonites trouvées ŕ Ville-sur-Saulx d'aprčs
, 1933).
La systématique suit (editor) (2005) et & (2010).
Classe Echinoidea , 1778
Sous-classe Euechinoidea , 1860
Infra-classe Acroechinoidea , 1981
Echinacea , 1876
Ordre Camarodonta , 1912
Infra-ordre Temnopleuridea & , 2010
Famille Glyphocyphidae , 1889
Genre Hemidiadema in & , 1846
Espčce-type. Hemidiadema rugosum , 1846.
Autres espčces associées. H. intermedius (, 1864), H. neocomiense (, 1869) et H. guebhardi , 1920 [nomen dubium] (= ? H. neocomiense (, 1869)).
Diagnose émendée. La diagnose émendée proposée est basée sur l'étude détaillée des différents individus d'H. rugosum et H. neocomiense étudiés dans le présent travail, complétée par quelques observations faites sur d'autres individus du Crétacé inférieur en cours d'études (, in prep. ; et al., in prep.).
* La terminologie utilisée pour décrire les fossettes suturales suit (1933, p. 113-114).
Remarques descriptives complémentaires concernant la structure ambulacraire :
1) L'aspect et la disposition des tubercules au niveau des ambulacres méritent une attention particuličre, comme l'ont déjà fait remarquer & (1909-1925, p. 192-193), (1933, p. 83, Fig. 32 C) et (1933, p. 54). Cette structure ambulacraire est propre au genre Hemidiadema et constitue ainsi l'un des caractčres diagnosiques essentiels du genre.
2) La composition des majeures ambulacraires n'est pas clairement identifiable sur les échantillons étudiés dans ce travail. L'absence de zones usées à la surface des tests ne permet pas d'avoir accčs à la forme des
assules, au nombre de demi-plaques et à la disposition de celles-ci au sein de chaque
majeure
ambulacraire. (1892b, p. 71) indique cependant qu'un "[…] échantillon de la collection de M.
permet de suivre distinctement les sutures des assules élémentaires […]. Chaque majeur est constituée, au dessous de l'ambitus, par trois primaires, puis, à l'ambitus, par deux primaires et une demi-plaque aborale
; au dessus de l'ambitus, les majeures sont simplement bisociées." Ces remarques n'ont pu ętre vérifiées qu'en partie. L'observation des ambulacres nous permet effectivement d'affirmer que les majeures sont trisociées et deviennent bisociées dans la partie supérieure de la zone supra-ambitale, en dessous le disque apical
(Fig. 10A ![]() ). L'association de deux primaires et d'une demi-plaque aborale n'a pas pu ętre observée.
). L'association de deux primaires et d'une demi-plaque aborale n'a pas pu ętre observée.
Affinités. Le genre Hemidiadema se rapproche du genre Glyphocyphus d' & , 1853. Ces deux genres se distinguent essentiellement et facilement par leur tuberculation et l'allure générale des aires ambulacraires. Chez Glyphocyphus, contrairement à ce que nous venons de préciser dans la diagnose d'Hemidiadema, les aires ambulacraires sont symétriques (demi-ambulacres identiques), garnies de tubercules primaires organisés en deux colonnes (arrangement bisérié). En outre, les tubercules ambulacraires et interambulacraires chez Glyphocyphus sont plus petits et plus nombreux sur chaque série et sont reliés entre eux par deux petites côtes verticales. Les zones miliaires adradiales, interradiales et perradiales sont nettement et plus réguličrement développées et recouvertes de nombreux et denses granules.
Le genre Hemidiadema, de part son ornementation générale, se rapproche du genre Zeuglopleurus , 1889 (= Nannoglyphus , 1978). Toutefois, l'examen du disque apical permet de les distinguer. Monocyclique chez Hemidiadema, le disque apical est hémicyclique (plaques ocellaires I et V insertes) chez Zeuglopleurus (, 1986 ; & , 1996).
Remarques. Comme l'a déjà trčs clairement fait remarquer (1908, p. 334-335) l'insuffisance de la description qu'a proposée de son genre et de son espčce-type (1846, p. 351) a entraîné de trčs nombreuses confusions et erreurs d'interprétation ayant conduit de nombreux échinologues contemporains de l'auteur à rejeter – à tort – le genre Hemidiadema proposé par .
(1858, p. 57) définissait les Hemidiadema comme "des Hemicidaris avec cette seule différence que les tubercules de la base des aires ambulacraires (semitubercules), au lieu de former deux rangs distincts se serrent de maničre à alterner et à ne former parfois qu'une seule rangée". Cette observation l'a conduit à rattacher au genre Hemidiadema quelques espčces jurassiques : Cidaris serialis , 1852, Pedina bakeri , 1854, Hemicidaris stramonium , 1852, et H. gagnebini , 1858. Cette conception de est erronée. Si une disposition alterne en une seule rangée des tubercules ambulacraires s'observe chez ces espčces, il n'en est pas le cas des autres caractčres diagnosiques du genre Hemidiadema. Ces espčces sont donc à rattacher au genre Hemicidaris , 1838. De męme, comme l'ont déjà signalé & (1993, p. 263-264), Hemidiadema morinieum & , 1872, du Portlandien de Boulogne-sur-Mer et Hemidiadema mortensensi , 1947, du Jurassique du Sud de l'Éthiopie sont également des Hemicidaris.
(1862-1867, p. 533) ne considérant pas l'arrangement unisérié des tubercules ambulacraires primaires observés chez l'espčce rugosum d' suffisant pour ętre retenue comme un caractčre d'ordre générique, a rejeté le genre Hemidiadema qu'il rattache au genre Glyphocyphus ("[…] C'est en 1853 que Jules en a fait le type du genre Glyphocyphus que les auteurs ont adopté depuis, et qui nous parait comprendre également le genre Hemidiadema . L'exemplaire unique qui, dans l'origine, a servi à établir le genre Hemidiadema, nous a été communiqué par M. , et nous avons retenu, en l'étudiant dans ses détails, que ce type curieux offre une étroite ressemblance avec les Glyphocyphus, et n'en diffčre réellement que par ses ambulacres pourvus d'une seule rangée de tubercules au lieu de deux. Il nous paraît que ce caractčre n'a pas, au point de vue zoologique, l'importance que lui a attribué ").
(1883, p. 103) est le premier à saisir la conception originale du genre et ainsi à distinguer clairement les Hemidiadema à la fois des Hemicidaris et des Glyphocyphus. Il est également le premier à énoncer une diagnose détaillée du genre. (1892b, p. 68-70), (1908, p. 334-335) et & (1925) adoptent les idées de .
(1889, p. 50) suit la conception de et considčre le genre Hemidiadema comme un sous-genre du genre Hemicidaris , 1838.
(1933, p. 115) considčre le genre Hemidiadema comme un sous-genre du genre Glyphocyphus, la distinction des deux genres ne se faisant que sur des "particularités dans la répartition des fossettes et des tubercules".
(1937, p. 64) suit l'opinion de et considčre le genre Hemidiadema comme un synonyme de Glyphocyphus.
(1943, p. 27), & (1966, p. U417) et & (1993, p. 263-264) retiennent la disposition unisériée des tubercules primaires ambulacraires comme l'un des caractčres génériques permettant de distinguer et de séparer les Hemidiadema des Glyphocyphus.
& (2010, p. 22-23) précisent : "il [l'holotype] nous fait examiner [...] les différences entre les genres Hemidiadema et Glyphocyphus". Une série de comparaisons de rapports de mesures entre radiatus (sous-entendu Glyphocyphus), différentes espčces d'Hemidiadema (non spécifiquement nommées) et H. rugosum les conduit à la conclusion suivante : "radiatus a donc une structure ambulacraire et interambulacraire différente des autres individus étudiés dont le type de rugosum". Cette conclusion peu claire sous-entend que ces auteurs distinguent Hemidiadema et Glyphocyphus comme deux genres distincts.
L'espčce Nannoglyphus wherlli , 1978, ne peut pas ętre rattachée au genre Hemidiadema comme l'ont fait et al. (2004). En effet, chez Hemidiadema le disque apical est monocyclique. Or chez N. wherlii celui-ci est hémicyclique (plaques ocellaires I et V insertes), ce qui rattache cette espčce au genre Zeuglopeurus , 1899, et à la famille des Zeuglopeuridae , 1986 (, 1986 ; & , 1996).
Répartitions géographique et stratigraphique. En France le genre Hemidiadema est connu du Valanginien inférieur jusqu'au Cénomanien (, in prep. ; & , in prep.). Des représentants y ont été cités et décrits dans de nombreuses localités mésogéennes : l'Aptien d'Espagne (, 1928 ; , 1991, 2003 ; et al., 2007 ; & , 2006 ; , 2008), le Barrémien et le Gargasien (Formation de Tadhart d'Imi n'Tanout) du Maroc (, 1959 ; et al., 1986, 1988 ; et al., 2004), dans l'Aptien de Tunisie (-, 1971 ; , 1996), dans l'Urgonien (Aptien) de Serbie (, 1977, 1998, 2000) et dans le Maestrichtien supérieur de Turquie ( & , 2003, p. 29). Pour cette derničre citation, il s'agit trčs probablement d'un représentant du genre Zeuglopleurus et non d'un Hemidiadema cf. intermedius comme le mentionnent les auteurs. Hemidiadema sp. a également été citée dans l'Aptien (Mayao Formation) du Xizang (Tibet) ( et al., 1981, p. 314 ; & , 1983, p. 147). Dans une révision des échinides de cette région par l'un de ces auteurs (, 1992), Hemidiadema sp. n'y est toutefois plus citée. La présence du genre dans cette région de Chine est donc trčs douteuse.
Hemidiadema rugosum , 1846
in & (1846)
(Fig. 8 ![]() ; Pls. 1
; Pls. 1 ![]() - 2
- 2 ![]() - 3
- 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() - 9
- 9 ![]() ;
;
Pl. 13 ![]() ,
figs. 1, 2, 4, 6, 8, 10 ; Pl. 14
,
figs. 1, 2, 4, 6, 8, 10 ; Pl. 14 ![]() ,
figs. 1-5
; Pl. 15
,
figs. 1-5
; Pl. 15 ![]() )
)
? 1842 – Diadema : & , p. 367.
1846 – Hemidiadema rugosum : & , p. 351.
1847 – Hemidiadema rugosum : & , p. 363.
1850 – Hemidiadema rugosum : d', p. 142.
1857 – Hemidiadema rugosum : , p. 245.
1858 – Hemidiadema rugosum : , p. 58.
1862 – Hemidiadema rugosum : & , p. 496.
1864 – Glyphocyphus rugosus : , p. 533, 543, Pl. 1128, figs. 16-22.
1868 – Hemidiadema rugosum : de , p. 286.
1870 – Hemidiadema rugosum : , p. 121.
1875 – Glyphocyphus rugosus : , p. 57.
1878 – Glyphocyphus rugosus : , p. 242, 244.
1883 – Hemidiadema rugosum : , p. 103.
1889 – Glyphocyphus rugosum : , p. 59 et 89.
1892a – Hemidiadema rugosum : , p. XXXVII.
1892b – Hemidiadema rugosum : , p. 69, Figs. 14, 70, 97, Pl. IV, figs. 5-6.
? 1901 – Hemidiadema rugosum : , p. 429.
1908 – Hemidiadema rugosum : , p. 334.
? 1912 – Hemidiadema cf. rugosum : , p. 239, 273.
? 1913 – Hemidiadema rugosum (Glyphocyphus rugosus) : , p. 307, 315, 362.
pars 1916 – Hemidiadema rugosum : , p. 79, 91 (p. 79, l'exemplaire cité de l'Albien de Clérey = Hemidiadema sp.)
? 1920 – Hemidiadema rugosum : , p. 3, 17, 20.
1925 – Hemidiadema rugosum : , p. 277, 279, 291.
1925 – Hemidiadema rugosum : & , p. 193.
? 1928 – Hemidiadema rugosum : , p. 149.
1933 – Glyphocyphus rugosus : , p. 83, Fig. 22c.
1933 – Glyphocyphus rugosus : , p. 54, 55.
? 1938 – Hemidiadema rugosum : , p. 49.
1943 – Hemidiadema rugosum : , p. 21, Fig. 18e, p. 22, Fig. 20c, Fig. 21c, p. 28, Fig. 24a, b, c (upper figure), d.
1947 – Hemidiadema rugosum : , p. 117.
? 1959 – Hemidiadema rugosum : , p. 95, Pl. XI, figs. 9-14.
1966 – Hemidiadema rugosum : & , p. U417, Fig. 312/1h.
? 1971 – Hemidiadema rugosum : -., p. 5, 12, 14, 15, 16.
? 1977 – Hemidiadema rugosum : , p. 185, 187, 195, 199, Pl. II, figs. 6-7.
? 1986 – Hemidiadema rugosum : et al., p. 398.
? 1988 – Hemidiadema rugosum : et al., p. 150.
non 1991 – Hemidiadema rugosum : , p. 213, Pl. 11, figs. 4-6. = Hemidiadema nov. sp. **
? 1996 – Hemidiadema rugosum : , p. 181, 185, Pl. I, figs. 1, 1a, 1b.
? 1998 – Hemidiadema rugosum : , p. 128.
? 2000 – Hemidiadema rugosum : , p. 78.
non 2003 – Hemidiadema rugosum : , p. 76, Pl. 2, figs. 14-16. = Hemidiadema nov. sp. **
non 2004 – Hemidiadema rugosum : et al., p. 600, Figs. 2/7a-c, Fig. 6 = Hemidiadema sp.
2006 – Hemidiadema rugosum : & , p. 39, 70, Fig. 22.
non 2007 – Hemidiadema rugosum : et al., photo n° 25. = Hemidiadema nov. sp. **
non 2008 – Glyphocyphus rugosum : et al., p. 159. = Hemidiadema nov. sp. **
non 2008 – Hemidiadema rugosa : , p. 6, 21, photos 35-36. = Hemidiadema nov. sp. **
2010 – Hemidiadema rugosum : & , p. 221-224, 5 Figs. (non numérotées).
2012 – Hemidiadema rugosum : et al., p. 2-3, Fig. 3.1.
** nouvelles espčces en cours de description ( et al., in prep.).
Matériel type. Hemidiadema rugosum , 1846, p. 47, des "Grčs ferrugineux" du Clansayésien (Aptien supérieur sensu gallico) de Grandpré (Ardennes, France) : Holotype par monotypie (EM 13095, coll. ).
Matériel connu.
EM 13095 (coll. ), holotype par monotypie (non figuré).
MNHN.F R62314 (coll. ), figuré in , 1892b, p. 70, Fig. 14, Pl. IV, figs. 5-6.
MNHN.F A20547, MNHN.F A33782, MNHN.F A33783 et MNHN.F A33784 (coll. ).
Ces six individus proviennent tous du Clansayésien (Aptien supérieur sensu gallico) de Grandpré (Ardennes, France), locus typicus et stratum typicum de l'espčce.
Spécimens étudiés et figurés.
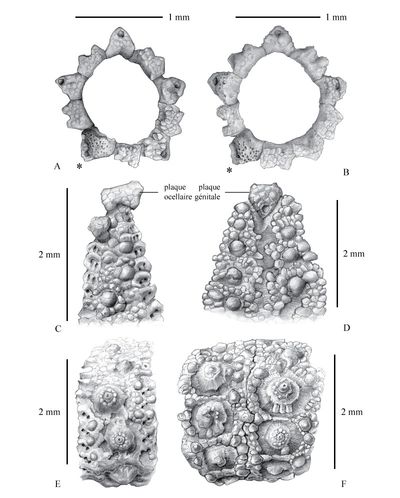
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
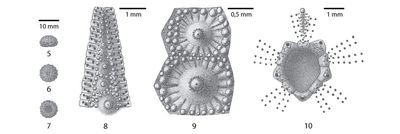
Figure 8 : Dessins d'observation d'Hemidiadema rugosum , 1846, réalisés sous binoculaire ŕ la chambre claire ; figs. A et B : disques apicaux ; fig. C : partie supra-ambitale de l'ambulacre 1 ; fig. D : partie supra-ambitale de l'interambulacre 1 ; fig. E : ambulacre V en vue ambitale ; fig. F : interambulacre 1 en vue ambitale ; * : madréporite ; A, C-F : MNHN.F A20547 ; B : MNHN.F 33783 [dessins : C. , MNHN Paris].
Mesures.
| Échantillons | Test | Ambulacres | Inter-ambulacres | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Nb. pla. ass. | Largeur (en mm) |
Nb tub. infra-A. | Nb tub. supra-A. | Nb tub. | Largeur (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| EM 13095 holotype | circa 9,04 | - | 26 / 27 | 1,84 | 6 | 6 | 12 | 3,10 | 2,09 | - |
| MNHN.F R62314 | 10,23 | 5,21 | - | 2,18 | 6 | 6 | 12 à 13 | 3,70 | 2,84 | 3,28 |
| MNHN.F A20547 | 7,88 | - | - | 1,51 | 5 | 5 | 10 à 11 | 2,52 | 1,85 | - |
| MNHN.F A33782 | circa 7,54 | - | 26 | 1,57 | 5 | 6 | 11 à 12 | 2,48 | 1,98 | - |
| MNHN.F A33783 | 6,90 | 3,62 | 25 | 1,54 | 5 | 5 à 6 | 10 à 11 | 2,38 | 1,97 | 2,97 |
| MNHN.F A33784 | 8,50 | - | 26 | 1,86 | 6 | 6 | 12 à 13 | 2,95 | 1,80 | - |
Diagnose émendée.
Description. La description ci-dessous est établie à partir de l'étude de l'holotype, complétée et affinée par celle des autres spécimens.
Morphologie générale. Test de petite taille (diamčtre généralement inférieur à 1 cm), à face inférieur plane, nettement enfoncée dans la partie centrale péristomiale et à surface supérieure faiblement bombée.
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Cônes lisses nettement crénelés en leur sommet et supportant un mamelon perforé. Aires scrobiculaires étroites, enfoncées par rapport à la base du cône et des zones miliaires, ornées de trčs courtes côtes rayonnantes plus ou moins marquées, généralement observables qu'au niveau des tubercules ambitaux. À l'aplomb des cônes se développent généralement deux à trois côtes verticales et parallčles plus saillantes, séparées de profondes dépressions (fossettes suturales scrobiculaires) entaillant la base des cônes
(Pl. 3 ![]() ,
figs. 6-8). Les aires scrobiculaires sont bordées de granules de taille irréguličre, trčs serrés les uns aux autres. Ces granules scrobiculaires, peu distinguables des granules miliaires qui les côtoient, sont disposés trčs irréguličrement et ne forment pas nettement un cercle autour de chaque aire scrobiculaire. En dessous de l'ambitus certains de ces granules sont différenciés en tubercules secondaires mamelonnés, perforés et parfois scrobiculés
(Pl. 3
,
figs. 6-8). Les aires scrobiculaires sont bordées de granules de taille irréguličre, trčs serrés les uns aux autres. Ces granules scrobiculaires, peu distinguables des granules miliaires qui les côtoient, sont disposés trčs irréguličrement et ne forment pas nettement un cercle autour de chaque aire scrobiculaire. En dessous de l'ambitus certains de ces granules sont différenciés en tubercules secondaires mamelonnés, perforés et parfois scrobiculés
(Pl. 3 ![]() ,
figs. 6-8
; Pl. 15
,
figs. 6-8
; Pl. 15 ![]() ). Ils sont particuličrement développés aux angles adradiaux et interradiaux des majeures. L'étroit scrobicule de certains de ces tubercules secondaires est parfois entouré d'un cercle de granules microscopiques.
). Ils sont particuličrement développés aux angles adradiaux et interradiaux des majeures. L'étroit scrobicule de certains de ces tubercules secondaires est parfois entouré d'un cercle de granules microscopiques.
Interambulacres. Aires interambulacraires composées d'un arrangement bisérié de 10 à 13 tubercules sur chaque série. Tubercules confluents, en particulier au niveau et en dessous de l'ambitus. Les aires scrobiculaires de deux tubercules consécutifs d'une męme série sont ainsi fréquemment directement en contact, sans intercalation de granules, seulement séparées par une sorte d'étroite petite muraille saillante ou de quelques granules alignés de trčs faible dimension. Dans la région ambitale et infra-ambitale, les zones miliaires interradiales et adradiales sont étroites, garnis de petits granules serrés, de taille variable, accolés aux granules scrobiculaires desquels ils sont difficilement distinguables
(Fig. 8F ![]() ). En dessus de l'ambitus, la zone miliaire interradiale s'élargit progressivement. Celle-ci est recouverte de granules et généralement de dépressions (fossettes suturales angulaires) plus ou moins développées, sculptant profondément la surface du test
(Fig. 8D
). En dessus de l'ambitus, la zone miliaire interradiale s'élargit progressivement. Celle-ci est recouverte de granules et généralement de dépressions (fossettes suturales angulaires) plus ou moins développées, sculptant profondément la surface du test
(Fig. 8D ![]() ; Pl. 8
; Pl. 8 ![]() , fig. 2).
, fig. 2).
Ambulacres. Aires ambulacraires droites, relativement larges (largeur A / largeur IA = 0,68 à 0,72) trisociées.
Zones interporifčres. Zones interporifčres ornées d'un arrangement unisérié de 10 à 12 tubercules sur chaque série. Le tubercule le plus développé est situé au niveau de l'ambitus ou légčrement en dessus. De l'ambitus en se dirigeant vers le péristome, 5 à 6 tubercules ornent la partie infra-ambitale. Ces tubercules, tangents à faiblement confluents, sont souvent disposés presque parfaitement linéairement les uns en dessus les autres. Ils occupent toute la largeur de la zone interporifčre et sont pour la plupart en partie tronqués sur leur côté adradial, l'aire scrobiculaire ou la base du cône étant en contact direct avec la zone porifčre
(Pl. 8 ![]() , fig. 3). Du côté opposé, le scrobicule est intčgre, bordé de granules scrobiculaires mamelonnés de taille irréguličre. De part de cette disposition des tubercules, il n'y a pas de zones miliaires adradiales ni perradiales. En dessus de l'ambitus
(Pl. 8
, fig. 3). Du côté opposé, le scrobicule est intčgre, bordé de granules scrobiculaires mamelonnés de taille irréguličre. De part de cette disposition des tubercules, il n'y a pas de zones miliaires adradiales ni perradiales. En dessus de l'ambitus
(Pl. 8 ![]() , fig. 1), les ambulacres montrent 5 à 6 tubercules dont la taille diminue assez rapidement en se dirigeant vers l'apex. Du fait de cette réduction de taille les tubercules, ainsi plus espacés, montrent une disposition nettement alterne. Les espaces situés entre les tubercules sont occupés par des granules miliaires et des dépressions liées au développement des
fossettes suturales angulaires au niveau du perradius.
Zones porifčres. Zones porifčres étroites, garnies de pores subcirculaires à ovales de petite taille, disposés en paires obliques. Le pourtour des pores est relevé, formant ainsi une sorte de cręte saillante délimitant un péripode, en particulier en dessous de l'ambitus. Un granule se développe souvent sur l'angle adradial supérieur des assules, au dessus du pore externe.
, fig. 1), les ambulacres montrent 5 à 6 tubercules dont la taille diminue assez rapidement en se dirigeant vers l'apex. Du fait de cette réduction de taille les tubercules, ainsi plus espacés, montrent une disposition nettement alterne. Les espaces situés entre les tubercules sont occupés par des granules miliaires et des dépressions liées au développement des
fossettes suturales angulaires au niveau du perradius.
Zones porifčres. Zones porifčres étroites, garnies de pores subcirculaires à ovales de petite taille, disposés en paires obliques. Le pourtour des pores est relevé, formant ainsi une sorte de cręte saillante délimitant un péripode, en particulier en dessous de l'ambitus. Un granule se développe souvent sur l'angle adradial supérieur des assules, au dessus du pore externe.
Fossettes suturales. Les fossettes suturales sont généralement bien développées. On distingue des fossettes scrobiculaires et des fossettes angulaires. Ces fossettes sont généralement nettement développées en dessus de l'ambitus, aussi bien au niveau des ambulacres
(Pl. 8 ![]() , fig. 1) qu'au niveau des interambulacres
(Pl. 8
, fig. 1) qu'au niveau des interambulacres
(Pl. 8 ![]() , fig. 2) oů elles sont les plus développées. Les fossettes suturales scrobiculaires, au nombre de trois, se développent sur la bordure adorale des majeures, sous les cônes dont elles entament la base et l'aire scrobiculaire. Elles sont séparées par des côtes verticales, étroites et parfois trčs saillantes
(Pl. 3
, fig. 2) oů elles sont les plus développées. Les fossettes suturales scrobiculaires, au nombre de trois, se développent sur la bordure adorale des majeures, sous les cônes dont elles entament la base et l'aire scrobiculaire. Elles sont séparées par des côtes verticales, étroites et parfois trčs saillantes
(Pl. 3 ![]() ,
figs. 6-7). Les fossettes angulaires, bien observables au niveau des interambulacres, plus irréguličrement visibles dans les ambulacres, correspondent à des dépressions se développant aux angles de la bordure adorale des majeures, en particulier du côté de l'interradius. La fossette suturale scrobiculaire la plus externe (du côté interradial du cône) tend parfois à se connecter et à fusionner avec la fossette suturale angulaire interradiale adjacente de la męme majeure, formant ainsi une large dépression s'ouvrant sur l'interradius. Les larges sutures "scrobiculo-angulaires" de l'ensemble des
majeures supra-ambitales se rejoignent et se connectent au niveau de l'interradius, formant alors à la surface du test une dépression interradiale parfois trčs large
(Pl. 8
,
figs. 6-7). Les fossettes angulaires, bien observables au niveau des interambulacres, plus irréguličrement visibles dans les ambulacres, correspondent à des dépressions se développant aux angles de la bordure adorale des majeures, en particulier du côté de l'interradius. La fossette suturale scrobiculaire la plus externe (du côté interradial du cône) tend parfois à se connecter et à fusionner avec la fossette suturale angulaire interradiale adjacente de la męme majeure, formant ainsi une large dépression s'ouvrant sur l'interradius. Les larges sutures "scrobiculo-angulaires" de l'ensemble des
majeures supra-ambitales se rejoignent et se connectent au niveau de l'interradius, formant alors à la surface du test une dépression interradiale parfois trčs large
(Pl. 8 ![]() , fig. 2). Au niveau et en dessous de l'ambitus, les fossettes suturales angulaires sont nettement moins développées, voir inexistantes. Les fossettes scrobiculaires interradiales restent toujours nettement distinctes, sans connexion et fusion avec les fossettes angulaires réduites. Au niveau des ambulacres, en dessus de l'ambitus, les fossettes scrobiculaires sont trčs étroites et les fossettes angulaires trčs irréguličrement développées. Chez certains exemplaires, des phénomčnes de fusion de sutures s'y observent, formant de larges et profondes dépressions perradiales
(Pl. 8
, fig. 2). Au niveau et en dessous de l'ambitus, les fossettes suturales angulaires sont nettement moins développées, voir inexistantes. Les fossettes scrobiculaires interradiales restent toujours nettement distinctes, sans connexion et fusion avec les fossettes angulaires réduites. Au niveau des ambulacres, en dessus de l'ambitus, les fossettes scrobiculaires sont trčs étroites et les fossettes angulaires trčs irréguličrement développées. Chez certains exemplaires, des phénomčnes de fusion de sutures s'y observent, formant de larges et profondes dépressions perradiales
(Pl. 8 ![]() , fig. 1).
, fig. 1).
Disque apical (Fig. 8A-B ![]() ; Pl. 14
; Pl. 14 ![]() ,
figs. 1-5). Disque apical monocyclique, circulaire à subcirculaire, uniformément large, à surface bombée.
Plaques génitales. Les plaques génitales 1, 2, 3 et 4 sont aussi hautes que larges, de taille et de forme identiques, la madréporite étant un peu plus grande et bombée. La plaque génitale 5 est un peu plus large, de forme plus triangulaire et son pourtour interne plus arqué. Gonopores circulaires de grande taille, nettement excentrés vers la bordure externe.
Plaques ocellaires. Les cinq plaques ocellaires sont de forme et de taille sensiblement identiques, avec un petit pore s'ouvrant latéralement à leur extrémité. La surface de l'ensemble des dix plaques du disque apical est irréguličre, granuleuse par le développement de nombreuses petites verrues.
,
figs. 1-5). Disque apical monocyclique, circulaire à subcirculaire, uniformément large, à surface bombée.
Plaques génitales. Les plaques génitales 1, 2, 3 et 4 sont aussi hautes que larges, de taille et de forme identiques, la madréporite étant un peu plus grande et bombée. La plaque génitale 5 est un peu plus large, de forme plus triangulaire et son pourtour interne plus arqué. Gonopores circulaires de grande taille, nettement excentrés vers la bordure externe.
Plaques ocellaires. Les cinq plaques ocellaires sont de forme et de taille sensiblement identiques, avec un petit pore s'ouvrant latéralement à leur extrémité. La surface de l'ensemble des dix plaques du disque apical est irréguličre, granuleuse par le développement de nombreuses petites verrues.
Périprocte. Aire périproctale assez peu étendue (Ø aire périproctale / Ø test = 0,24 à 0,29), circulaire à subcirculaire.
Péristome. Aire péristomiale circulaire, moyennement large (Ø aire péristomiale / Ø test = 0,32 à 0,38), assez profondément enfoncée par rapport à la surface du test.
Variation intraspécifique. Elle se traduit essentiellement par le développement plus ou moins important des fossettes suturales en dessus l'ambitus au détriment des granules miliaires.
Quelques remarques sur les figurations originales de
(Fig. 2 ![]() ). Comme l'a déjà fait remarquer
(1892b, p. 72), les figurations de présentent certaines erreurs et lacunes d'observation. Sur les figures 19 et 21, les granules (scrobiculaires et miliaires) et les tubercules secondaires sont totalement omis. Sur la figure 20 et 21, la largeur des zones miliaires adradiales est un peu exagérée, la taille des granules scrobiculaires et miliaires est trop homogčne, les granules mamelonnés et ceux différenciés en tubercules secondaires n'ont pas été représentés. Sur la figure 21 les majeures ambulacraires ont été par erreur toutes représentées comme étant 4-sociées, alors qu'elles ne sont que trisociées en réalité.
). Comme l'a déjà fait remarquer
(1892b, p. 72), les figurations de présentent certaines erreurs et lacunes d'observation. Sur les figures 19 et 21, les granules (scrobiculaires et miliaires) et les tubercules secondaires sont totalement omis. Sur la figure 20 et 21, la largeur des zones miliaires adradiales est un peu exagérée, la taille des granules scrobiculaires et miliaires est trop homogčne, les granules mamelonnés et ceux différenciés en tubercules secondaires n'ont pas été représentés. Sur la figure 21 les majeures ambulacraires ont été par erreur toutes représentées comme étant 4-sociées, alors qu'elles ne sont que trisociées en réalité.
Quelques remarques sur les figurations originales de (Fig. 3 ![]() ). Sur la figure 5, la face apicale de l'échantillon a été dessinée dans ses moindres détails (tuberculation, granulation, disque apical). Il s'agit en grande partie d'une interprétation du dessinateur, car sur cet échantillon (coll. , MNHN.F R62314) la face apicale est de conservation trčs médiocre du fait des trčs nombreuses impressions de petits lithoclastes qui ne permettent pas d'observer l'ornementation de la surface du test avec détails, en particulier le disque apical. Sur la figure 6, les tubercules ambulacraires sont en réalité pas aussi parfaitement alignés verticalement les uns en dessus des autres.
). Sur la figure 5, la face apicale de l'échantillon a été dessinée dans ses moindres détails (tuberculation, granulation, disque apical). Il s'agit en grande partie d'une interprétation du dessinateur, car sur cet échantillon (coll. , MNHN.F R62314) la face apicale est de conservation trčs médiocre du fait des trčs nombreuses impressions de petits lithoclastes qui ne permettent pas d'observer l'ornementation de la surface du test avec détails, en particulier le disque apical. Sur la figure 6, les tubercules ambulacraires sont en réalité pas aussi parfaitement alignés verticalement les uns en dessus des autres.
Hemidiadema neocomiense (, 1869)
(Figs. 9 ![]() - 10
- 10 ![]() ; Pls. 10
; Pls. 10 ![]() - 11
- 11 ![]() - 12
- 12 ![]() ;
;
Pl. 13 ![]() ,
figs. 3, 5, 7, 9, 11 ; Pl.
14
,
figs. 3, 5, 7, 9, 11 ; Pl.
14 ![]() , fig. 6
; Pl. 16
, fig. 6
; Pl. 16 ![]() )
)
1869 – Glyphocyphus neocomiensis , p. 245, Pl. 13, figs. 5-10.
1883 – Glyphocyphus neocomiensis : , p. 103.
1908 – Hemidiadema neocomiense : , p. 334.
? 1920 – Hemidiadema guebhardi : , p. 3, Fig. 3. [nomen dubium]
? 1923 – Hemidiadema guebhardi : , p. 484. [nomen dubium]
1925 – Hemidiadema neocomiensis : , p. 146.
1925 – Hemidiadema neocomiense : & , p. 193.
1943 – Hemidiadema neocomiense : , p. 21, Fig. 18f, p. 28.
1959 – Hemidiadema neocomiense : , p. 96.
? 1965 – Hemidiadema guebhardi : , p. 60. [nomen dubium]
1966 – Hemidiadema neocomiense : & , p. U417, Fig. 312/1d-g.
1971 – Hemidiadema neocomiensis : -, p. 103.
1976 – Hemidiadema neocomiense : , tabl. 8.
1976 – Hemidiadema neocomiense : & , p. 47.
1993 – Hemidiadema intermedium : & , p. 263.
2000 – Hemidiadema neocomiense : , p. 5, 9.
Matériel type. Glyphocyphus neocomiensis , 1869, p. 245, Pl. 13, figs. 5-10 du Néocomien (Couches à Echinospatagus cordiformis) [= " Calcaire à Spatangues" de l'Hauterivien inférieur] des environs d'Auxerre (Yonne, France) : Holotype par monotypie (coll. ).
Remarque préliminaire. Malgré des recherches, l'holotype n'a pas été retrouvé au sein de la collection conservée à la Faculté des Sciences de Lyon (collections de l'École nationale supérieure des Mines de Paris) et Musée d'Histoire Naturelle d'Auxerre. Nous devons considérer comme définitivement perdu ce spécimen.
Matériel connu et étudié. Malgré de longues recherches dans différentes collections paléontologiques (MNHN de Paris, UPMC de Paris, MHN d'Auxerre) oů du matériel paléontologique de l'Hauterivien du Bassin parisien est conservé, aucun exemplaire d'H. neocomiense n'a été trouvé parmi l'abondant matériel échinologique. L'unique exemplaire trouvé l'a été dans la collection échinologique de Paul déposée au Muséum-Aquarium de Nancy. Ce constat souligne l'extręme rareté de cette espčce.
Néotype. Coll. n° 384 de l'ENSGN/MAN. Le choix de cet individu a un double intéręt. D'une part il provient de la męme région géographique (Ville-sur-Saulx (Meuse) en bordure orientale du bassin de Paris) et surtout de la męme formation ("Calcaire à Spatangues", Hauterivien inférieur) que l'holotype. D'autre part cet échantillon est complet, non déformé, d'une remarquable conservation et sa taille et ses caractéristiques sont conformes à la description de .
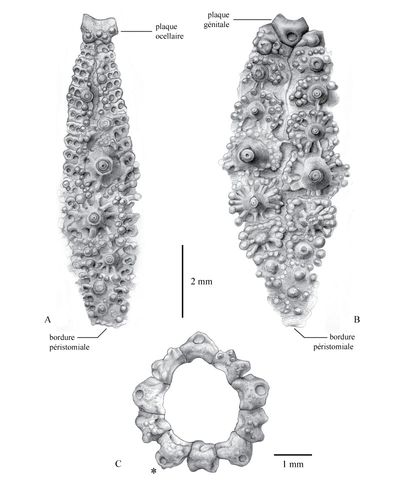
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Figure 9 : Dessins d'observation du néotype d'Hemidiadema neocomiense (, 1869) (ENSGN/MAN ; coll. n° 384) réalisés sous binoculaire ŕ la chambre claire ; fig. A : ambulacre V ; fig. B : interambulacre 5 ; fig. C : disque apical ; * : Madréporite [dessins : C. , MNHN Paris].
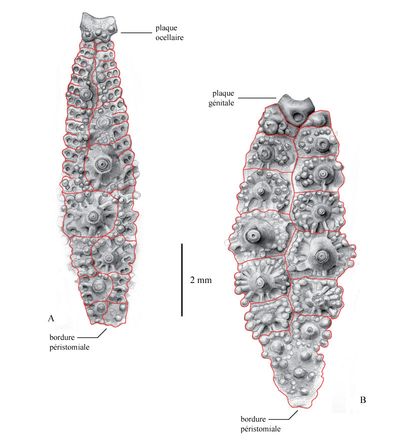
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Figure 10 : Dessins d'interprétation des contours des plaques (en rouge) du néotype d'Hemidiadema neocomiense (, 1869) (ENSGN/MAN ; coll. n° 384) ; fig. A : ambulacre V ; fig. B : interambulacre 5 [dessins : C. , MNHN Paris ; contour des plaques : A. ].
Mesures.
| Échantillons | Test | Ambulacres | Inter-ambulacres | Périprocte | Péristome | |||||
| Diamčtre (en mm) |
Hauteur (en mm) |
Largeur (en mm) |
Nb. pla. ass. | Nb tub. infra-A. | Nb tub. supra-A. | Nb tub. | Largeur (en mm) |
Diamčtre (en mm) |
Diamčtre (en mm) |
|
| Coll. n° 384 | 8,48 | 5,14 | 1,90 | 24 / 25 | 5 | 3 à 4 | 9 | 3,10 | 2,12 | 2,95 |
Diagnose émendée.
Description du spécimen.
Morphologie générale. Test de petite taille, à face inférieur plane, enfoncée dans la région centrale péristomiale et à surface supérieure moyennement bombée, s'aplanissant à son apex.
Tubercules. Tubercules scrobiculés, crénelés, mamelonnés et perforés. Cônes lisses, moyennement développés, crénelés à leur sommet et supportant un net mamelon possédant un pore bien développé. Aires scrobiculaires généralement étendues, ornées de côtes rayonnantes épaisses et bien marquées, fusionnant à la base du cône. À l'aplomb des cônes, deux à trois côtes parallčles verticales plus étroites et plus saillantes s'observent, séparant deux à trois fossettes suturales. Chaque côte rayonnante se termine par un granule grossier. L'ensemble de ces granules, toujours bien distincts et généralement réguličrement espacés les uns des autres, s'organise en un cercle autour du scrobicule. Ces cercles de granules scrobiculaires sont surtout nets au niveau et en dessous de l'ambitus. Ils deviennent plus incomplets en dessus l'ambitus, les granules scrobiculaires devenant plus éparses et espacés, et ainsi irréguličrement disposés autour des scrobicules. Ces granules srobiculaires sont généralement surmontés d'un petit mamelon. En dessous de l'ambitus la taille de ces granules scrobiculaires tend à augmenter aux angles des majeurs et sont plus nettement mamelonnés, sans toutefois se développer en de véritables tubercules secondaires.
Interambulacres (Figs. 9B ![]() - 10B
- 10B ![]() ; Pl. 12
; Pl. 12 ![]() ,
figs. 1, 2, 5, 6, 9,
10). Aires interambulacraires composées d'un arrangement bisérié de 9 tubercules sur chaque série. Tubercules souvent confluents, les aires scrobiculaires de deux tubercules consécutifs d'une męme série sont en contact direct par atténuation ou disparition totale des granules scrobulaires. La taille des tubercules, maximale à l'ambitus, diminue progressivement en en se dirigeant vers l'apex et le péristome. Zone miliaire interradiale inexistante en dessous de l'ambitus, les deux séries de tubercules étant séparées par un étroit mais assez profond sillon interradial, celui-ci se prolongeant en dessus l'ambitus jusqu'à l'apex. En dessus de l'ambitus, une étroite zone miliaire interradiale se développe. De trčs rares granules miliaires s'y observent, la plupart accolés aux granules scrobiculaires. Sa partie médiane est entaillée par une dépression zigzagante, résultat de la fusion de fossettes angulaires avec le sillon interradial préexistant séparant les deux séries de tubercules. Zones miliaires adradiales inexistantes sur toute la hauteur du test, les granules scrobiculaires étant directement en contact avec la zone porifčre ou espacés d'un trčs étroit espace lisse.
,
figs. 1, 2, 5, 6, 9,
10). Aires interambulacraires composées d'un arrangement bisérié de 9 tubercules sur chaque série. Tubercules souvent confluents, les aires scrobiculaires de deux tubercules consécutifs d'une męme série sont en contact direct par atténuation ou disparition totale des granules scrobulaires. La taille des tubercules, maximale à l'ambitus, diminue progressivement en en se dirigeant vers l'apex et le péristome. Zone miliaire interradiale inexistante en dessous de l'ambitus, les deux séries de tubercules étant séparées par un étroit mais assez profond sillon interradial, celui-ci se prolongeant en dessus l'ambitus jusqu'à l'apex. En dessus de l'ambitus, une étroite zone miliaire interradiale se développe. De trčs rares granules miliaires s'y observent, la plupart accolés aux granules scrobiculaires. Sa partie médiane est entaillée par une dépression zigzagante, résultat de la fusion de fossettes angulaires avec le sillon interradial préexistant séparant les deux séries de tubercules. Zones miliaires adradiales inexistantes sur toute la hauteur du test, les granules scrobiculaires étant directement en contact avec la zone porifčre ou espacés d'un trčs étroit espace lisse.
Ambulacres (Figs. 9A ![]() - 10A
- 10A ![]() ; Pl. 12
; Pl. 12 ![]() ,
figs. 3, 4, 7, 8, 11,
12). Aires ambulacraires droites, relativement larges (largeur A / largeur IA = 0,65).
Zones interporifčres. Zones interporifčres ornées d'un arrangement unisérié de 8 à 9 tubercules sur chaque série, à disposition alterne ou parfois presque tous disposés du męme côté, linéairement les uns en dessous des autres, selon l'ambulacre considéré. Tubercules ambitaux les plus gros au niveau de l'ambitus, leur taille diminuant progressivement en se dirigeant vers le péristome. Dans les zones ambitales
(Pl. 12
,
figs. 3, 4, 7, 8, 11,
12). Aires ambulacraires droites, relativement larges (largeur A / largeur IA = 0,65).
Zones interporifčres. Zones interporifčres ornées d'un arrangement unisérié de 8 à 9 tubercules sur chaque série, à disposition alterne ou parfois presque tous disposés du męme côté, linéairement les uns en dessous des autres, selon l'ambulacre considéré. Tubercules ambitaux les plus gros au niveau de l'ambitus, leur taille diminuant progressivement en se dirigeant vers le péristome. Dans les zones ambitales
(Pl. 12 ![]() ,
figs. 7-8) et
infra-ambitales (Pl. 12
,
figs. 7-8) et
infra-ambitales (Pl. 12 ![]() ,
figs. 11-12) les tubercules ambitaux et sous-ambitaux occupent toute la largeur de la zone inter-porifčre. Ils sont asymétriques, leur côté adradial est tronqué d'une partie des granules scrobiculaires et de l'aire scrobiculaire, celle-ci ou la base du cône étant en contact direct avec la zone porifčre. Dans la zone
supra-ambitale (Pl. 12
,
figs. 11-12) les tubercules ambitaux et sous-ambitaux occupent toute la largeur de la zone inter-porifčre. Ils sont asymétriques, leur côté adradial est tronqué d'une partie des granules scrobiculaires et de l'aire scrobiculaire, celle-ci ou la base du cône étant en contact direct avec la zone porifčre. Dans la zone
supra-ambitale (Pl. 12 ![]() ,
figs. 3-4) la taille des tubercules diminue depuis l'ambitus en se dirigeant vers l'apex. Ces tubercules se réduisent à un cône surmonté d'un mamelon, nettement accolé à la zone porifčre, entouré de quelques granules épars, l'aire scrobiculaire étant trčs réduite à inexistante. La zone miliaire perradiale se réduit à un étroit et profond sillon, résultant en partie du développement de fossettes suturales angulaires qui atteignent le perradius et y fusionnent.
Zones porifčres. Zones porifčres assez larges dont les pores circulaires à sub-circulaires sont disposés en paire oblique. Surface des assules en dessus l'ambitus généralement plane, recouverte de nombreuses verrues microscopiques donnant un aspect finement granuleux, plus bombée et irréguličre en dessous de l'ambitus avec localement développement d'un pore adradial à l'angle supérieur.
,
figs. 3-4) la taille des tubercules diminue depuis l'ambitus en se dirigeant vers l'apex. Ces tubercules se réduisent à un cône surmonté d'un mamelon, nettement accolé à la zone porifčre, entouré de quelques granules épars, l'aire scrobiculaire étant trčs réduite à inexistante. La zone miliaire perradiale se réduit à un étroit et profond sillon, résultant en partie du développement de fossettes suturales angulaires qui atteignent le perradius et y fusionnent.
Zones porifčres. Zones porifčres assez larges dont les pores circulaires à sub-circulaires sont disposés en paire oblique. Surface des assules en dessus l'ambitus généralement plane, recouverte de nombreuses verrues microscopiques donnant un aspect finement granuleux, plus bombée et irréguličre en dessous de l'ambitus avec localement développement d'un pore adradial à l'angle supérieur.
Fossettes suturales (Pls. 12 ![]() & 16
& 16 ![]() ). Les fossettes suturales sont nettement développées sur la bordure adorale des majeures. On y distingue des fossettes scrobiculaires et des fossettes angulaires. Les fossettes scrobiculaires se développent en dessous les cônes, au nombre de trois généralement. Elles sont séparées par des côtes verticales et parallčles, généralement plus prononcées que les côtes rayonnantes en dessus l'ambitus
(Pl. 16
). Les fossettes suturales sont nettement développées sur la bordure adorale des majeures. On y distingue des fossettes scrobiculaires et des fossettes angulaires. Les fossettes scrobiculaires se développent en dessous les cônes, au nombre de trois généralement. Elles sont séparées par des côtes verticales et parallčles, généralement plus prononcées que les côtes rayonnantes en dessus l'ambitus
(Pl. 16 ![]() , fig. 1). En dessous l'ambitus, ces côtes sont plus étroites et saillantes. Les fossettes suturales angulaires sont particuličrement bien développées en dessus de l'ambitus. Au niveau des aires inter-ambulacraires, elles correspondent à de profondes dépressions se développant à l'angle de la bordure adorale des majeures, surtout du côté interradial. Elles atteignent et viennent fusionner au niveau de l'interradius avec le sillon interradial qu'elles élargissent en une profonde dépression s'étendant depuis l'ambitus jusqu'à l'apex
(Pl. 12
, fig. 1). En dessous l'ambitus, ces côtes sont plus étroites et saillantes. Les fossettes suturales angulaires sont particuličrement bien développées en dessus de l'ambitus. Au niveau des aires inter-ambulacraires, elles correspondent à de profondes dépressions se développant à l'angle de la bordure adorale des majeures, surtout du côté interradial. Elles atteignent et viennent fusionner au niveau de l'interradius avec le sillon interradial qu'elles élargissent en une profonde dépression s'étendant depuis l'ambitus jusqu'à l'apex
(Pl. 12 ![]() ,
figs. 1, 2, 5, 6). Certaines sutures scrobiculaires les plus internes (du côté interradial) se fondent dans cette dépression, par élargissement et fusion avec les sutures angulaires adjacentes. En dessous de l'ambitus, cette dépression se poursuit sous la forme d'un sillon médian un peu plus étroit, séparant les deux séries de tubercules
(Pl. 12
,
figs. 1, 2, 5, 6). Certaines sutures scrobiculaires les plus internes (du côté interradial) se fondent dans cette dépression, par élargissement et fusion avec les sutures angulaires adjacentes. En dessous de l'ambitus, cette dépression se poursuit sous la forme d'un sillon médian un peu plus étroit, séparant les deux séries de tubercules
(Pl. 12 ![]() ,
figs. 9-10). Au niveau des ambulacres, depuis l'ambitus jusqu'à l'apex, s'observe également une nette et profonde dépression perradiale, résultant en partie de la connexion de fossettes angulaires
(Fig. 9A
,
figs. 9-10). Au niveau des ambulacres, depuis l'ambitus jusqu'à l'apex, s'observe également une nette et profonde dépression perradiale, résultant en partie de la connexion de fossettes angulaires
(Fig. 9A ![]() ; Pl. 12
; Pl. 12 ![]() ,
figs. 3, 4, 7, 8).
,
figs. 3, 4, 7, 8).
Disque apical (Fig. 9C ![]() ; Pl. 14
; Pl. 14 ![]() ,
fig. 6). Monocyclique, massif, un peu sur-épaissi sur le pourtour interne, à surface irréguličre.
Plaques génitales. Plaques génitales 1, 2, 3 et 4 sont de taille et de forme identiques, plus hautes que larges. Gonopores circulaires, de grande taille, trčs excentrés vers la bordure externe. Un granule se développe au dessus de chaque gonopore, parfois un peu excentré. Plaque génitale 5 un peu plus large et de forme plus triangulaire. Pas de granule au dessus du gonopore de celle-ci.
Plaques ocellaires. Plaques ocellaires II, III et IV de taille et de formes analogues, un peu plus hautes que larges, pore circulaire trčs excentré, presque accolé à la bordure externe. Plaques ocellaires I et V moins hautes et plus larges. Quelques granules s'y observent. La surface du disque est recouverte de nombreuses verrues microscopiques, non visibles à l'œil nu.
,
fig. 6). Monocyclique, massif, un peu sur-épaissi sur le pourtour interne, à surface irréguličre.
Plaques génitales. Plaques génitales 1, 2, 3 et 4 sont de taille et de forme identiques, plus hautes que larges. Gonopores circulaires, de grande taille, trčs excentrés vers la bordure externe. Un granule se développe au dessus de chaque gonopore, parfois un peu excentré. Plaque génitale 5 un peu plus large et de forme plus triangulaire. Pas de granule au dessus du gonopore de celle-ci.
Plaques ocellaires. Plaques ocellaires II, III et IV de taille et de formes analogues, un peu plus hautes que larges, pore circulaire trčs excentré, presque accolé à la bordure externe. Plaques ocellaires I et V moins hautes et plus larges. Quelques granules s'y observent. La surface du disque est recouverte de nombreuses verrues microscopiques, non visibles à l'œil nu.
Périprocte. Aire périproctale subcirculaire, moyennement large (Ø aire périproctale / Ø test = 0,3), un peu allongé dans l'axe antéro-postérieur.
Péristome. Aire péristomiale circulaire, assez peu étendue (Ø aire péristomiale / Ø test = 0,25), assez profondément enfoncé par rapport à la surface du test.
Quelques remarques sur les figurations originales de
(Fig. 4 ![]() ). La figure 9 –
présentant deux tubercules interambulacraires successifs d'une męme série, l'inférieur semblant ętre le tubercule ambital – montre trčs exactement l'allure rayonnée des aires scrobiculaires par le développement de côtes rayonnantes, chacune portant à son extrémité un granule dont l'ensemble dessine un cercle autour de l'aire scrobiculaire. Les fossettes scrobiculaires séparées par des cotes verticales parallčles plus saillantes se développant en dessous les cônes sur la bordure aborale de la majeure, par contre, ne sont pas représentées. Ce caractčre est également omis dans la description littérale. Sur la figure 8, montrant la partie supra-ambitale d'un ambulacre, les tubercules sont trop situés dans la partie médiane de la zone interporifčre et le sillon interradial séparant les deux demi-ambulacres n'est pas représenté.
). La figure 9 –
présentant deux tubercules interambulacraires successifs d'une męme série, l'inférieur semblant ętre le tubercule ambital – montre trčs exactement l'allure rayonnée des aires scrobiculaires par le développement de côtes rayonnantes, chacune portant à son extrémité un granule dont l'ensemble dessine un cercle autour de l'aire scrobiculaire. Les fossettes scrobiculaires séparées par des cotes verticales parallčles plus saillantes se développant en dessous les cônes sur la bordure aborale de la majeure, par contre, ne sont pas représentées. Ce caractčre est également omis dans la description littérale. Sur la figure 8, montrant la partie supra-ambitale d'un ambulacre, les tubercules sont trop situés dans la partie médiane de la zone interporifčre et le sillon interradial séparant les deux demi-ambulacres n'est pas représenté.
Hemidiadema neocomiense (, 1869) se distingue nettement d'H. rugosum par :
Hemidiadema neocomiense (, 1869) se distingue d'H. intermedius (, 1864) – trčs rare espčce albo-cénomanienne incomplčtement décrite et par conséquent mal connue – par au moins la disposition des tubercules ambulacraires en dessous l'ambitus. Presque parfaitement disposés verticalement les uns en dessus les autres chez H. neocomiense, les tubercules ambulacraires sont moins larges et montrent un arrangement unisériée nettement alterne en dessous l'ambitus chez H. intermedius [d'aprčs la description littérale et les figurations qu'en donne (1862-1867, p. 540-543, Pl. 1128, figs. 6-15), les trois exemplaires décrits et figurés dans la Paléontologie Française par n'ayant malheureusement pas été retrouvés au sein des collections de l'Université Pierre et Marie Curie à Paris (collections de la Sorbonne) et n'ayant donc pas pu ętre étudiés en tant que matériel comparatif dans le cadre de ce travail].
Hemidiadema rugosum , 1846, de part les caractéristiques que nous venons d'énoncer, se différencie trčs facilement des autres espčces actuellement connues (H. neocomiense, H. intermedius) et des autres espčces en cours d'étude ( et al., in prep. ; , in prep.).
Hemidiadema rugosum , 1846, est connue en France qu'avec certitude dans le Clansayésien (Aptien supérieur sensu gallico) de Grandpré (Ardennes) ainsi que dans le Sud-Est de la France, dans les "Grčs glauconieux à Discoides decoratus" de l'Aptien supérieur (Zone à Acanthohoplites nolani, Sous-Zone à Diadochoceras subnodosocostatum) du Teil, prčs de Montélimar (Ardčche) (, in prep.). Cette espčce avait déjà été citée dans cette męme formation dans le Gard, prčs d'Uzčs (, 1901, p. 429).
L'espčce a également été citée dans le Gargasien du Sud-Est de la France (, 1913, p. 307, 315, 362), l'Albien des Alpes Maritimes et de l'Aube (, 1916, p. 79 ; 1920, p. 17). Il ne s'agit toutefois que de citations dont il n'est pas possible d'en vérifier l'exactitude. Trčs récemment deux Hemidiadema ont été découverts dans l'Albien de l'Aube par l'Association Géologique Auboise (déterminés comme des Polydiadema rhodani , 1840, par Claude in (Coord.), 2010, p. 208, Fig. 138 E, F1 et F2). Si ces découvertes confirment bien la présence du genre Hemidiadema dans l'Albien du Bassin de Paris, elles ne confirment pas la citation de (1916, p. 79) car ces deux individus – en cours d'étude ( & , in prep.) – n'appartiennent pas à l'espčce rugosum (tout comme probablement l'exemplaire cité par ).
En dehors de la France H. rugosum a été citée à de nombreuses reprises : dans le Bédoulien (Aptien inférieur sensu gallico) d'Espagne (, 1928, p. 149 ; , 1991, 2003 ; et al., 2007 ; & , 2006 ; et al., 2008 ; , 2008), dans le Gargasien (Aptien moyen sensu gallico) du Maroc (, 1959 ; et al., 1986, 1988 ; et al., 2004), dans l'Aptien de Tunisie (-, 1971 ; , 1996) et dans l'Aptien de Serbie (, 1977). La plupart de ces déterminations et figurations semblent erronées ou douteuses et restent à ętre confirmées par une ré-étude des spécimens cités par ces auteurs. De nombreux spécimens provenant de différentes localités aptiennes d'Espagne (dont ceux décrits et figurés par (1991, p. 213, Pl. 11, figs. 4-6 ; 2003, p. 76, Pl. 2, figs. 14-16)) ont récemment fait l'objet d'une ré-étude préliminaire ( et al., 2012) qui est actuellement approfondie ( et al., in prep.). Ces spécimens, initialement attribués à H. rugosum, n'appartiennent pas à l'espčce décrite par mais constituent de nouveaux taxons. H. rugosum n'est actuellement connu avec certitude en Espagne que par un unique spécimen provenant de l'Aptien de la région d'Alicante ( & , 2006, p. 39, 70, Fig. 22 ; et al., 2012, p. 2-3, Fig. 3.1 ; et al., in prep.).
Hemidiadema neocomiense (, 1869) n'est connu qu'en France. Hormis l'Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum) de la bordure orientale du Bassin de Paris (Yonne, Meuse) d'oů proviennent l'holotype et le néotype, l'espčce a été rencontrée dans d'autres niveaux stratigraphiques du Sud-Est de la France, en particulier dans le Barrémien inférieur ( & , 1976, p. 47 ; , 1976, tabl. 8 ; , 2000, p. 5, 9). L'espčce semble y apparaître dčs le Valanginien inférieur et se rencontre vraisemblablement jusque dans le Bédoulien de cette męme région (, in prep.).
La terminologie utilisée pour définir les relations paléoécologiques des espčces en fonction de la nature (texture, granulométrie, …) du substrat suit (1965).
Hemidiadema rugosum , 1846 : Le "Minerai de fer du Bois des Loges" prčs de Grandpré (Ardennes) formé de trčs nombreux lithoclastes roulés de silice, de limonite et de glauconie emballés dans un ciment argileux correspond à d'anciens substrats oů la fraction de sable grossier est nettement dominante. Il en est de męme pour l'exemplaire provenant des "Grčs glauconieux à Discoides decoratus" de l'Aptien supérieur (Zone à Acanthohoplites nolani, Sous-Zone à Diadochoceras subnodosocostatum) du Teil prčs de Montélimar (Ardčche, France) (, in prep.) et de l'exemplaire figuré par & (2006, p. 39, 70, Fig. 22 ; et al., 2012, p. 3, Fig. 3.1) provenant de l'Aptien d'Alicante en Espagne. H. rugosum apparaît ainsi une espčce typiquement gravellicole.
Hemidiadema neocomiense (, 1869) : La gangue qui peut ętre observée dans l'aire péristomiale du néotype est une marne argileuse trčs finement sableuse, ne montrant pas de lithoclastes et bioclastes. Les spécimens d'H. neocomiense trouvés dans le Barrémien inférieur du Sud-Est de la France proviennent également de niveaux marneux oů la fraction détritique est inexistante à trčs faible (vasičres circalitorales de bordure de plate-forme externe). H. neocomiense fréquentait ainsi des milieux à dominante vaseuse. Elle paraît ainsi une espčce vasicole.
Les caractčres spécifiques et le statut taxonomique des espčces Hemidiadema rugosum , 1846, et H. neocomiense (, 1869) ont été précisés à partir de l'étude détaillée d'individus provenant des stratum typicum respectifs de ces deux espčces. Les espčces rugosum et neocomiense sont deux espčces distinctes appartenant au genre Hemidiadema , 1846, aisément reconnaissables par leur ornementation et leur position stratigraphique. Elles possčdent de plus des préférences paléoécologiques différentes. La révision de ces deux espčces, dont l'espčce-type du genre – H. rugosum – a également permis de clarifier le statut taxonomique du genre Hemidiadema , 1846, longtemps rattaché aux genres Glyphocyphus d' & , 1853, ou Hemicidaris , 1840, selon les auteurs. Une diagnose émendée complčte du genre a pu ętre établie.
La présente étude pourra ainsi servir de travail de base pour la révision des Hemidiadema du Crétacé d'autres localités mésogéennes (Sud-Est de la France, Espagne, Maroc et Tunisie), la révision de certaines d'entre elles étant en cours (, in prep. ; et al., in prep. ; & , in prep.). H. intermedium (, 1864) et H. guebhardi , 1920 [nomen dubium] (= ? H. neocomiensis (, 1869), les deux autres espčces décrites rattachées au genre Hemidiadema provenant du Crétacé inférieur du Sud-Est de la France sont également trčs incomplčtement connues et leur type respectif non actuellement retrouvé. Une révision devra donc s'imposer pour préciser la validité taxonomique de ces deux espčces et en clarifier les caractčres spécifiques (, in prep.). Enfin, des spécimens d'Hemididema ont été cités en dehors de la France – tous rattachés à l'espčce rugosum d', – mais sans étude approfondie des spécimens citées et/ou figurés. Les déterminations avancées dans ces travaux sont douteuses voir erronées, car basée sur les descriptions historiques lacunaires et sans étude comparative détaillée. Un examen préliminaire d'une partie de ce matériel – en particulier de l'Aptien d'Espagne ( et al., 2012) – démontre la non appartenance de ces spécimens à l'espčce d' et l'identification de plusieurs nouveaux taxons ( et al., in prep.).
Tous les dessins scientifiques (Figs. 8 ![]() - 9
- 9 ![]() - 10
- 10 ![]() ) ont été réalisés par Charlčne
(Illustratrice scientifique, Muséum national d'Histoire naturelle de Paris, France).
) ont été réalisés par Charlčne
(Illustratrice scientifique, Muséum national d'Histoire naturelle de Paris, France).
L'auteur tient à remercier vivement toutes les personnes qui ont rendu possible l'accčs, la recherche (fructueuse ou infructueuse) et l'emprunt pour étude de certains échinides : Abel (Faculté des Sciences de Lyon, France), Sylvain (Muséum national d'Histoire naturelle de Paris, France), Sandrine Derson (Muséum-Aquarium de Nancy, France), Gilles (Musée d'Histoire naturelle d'Auxerre, France), Isabelle (Université Pierre et Marie Curie, Paris, France), Sebastián Calzada (Musée géologique du Séminaire de Barcelone, Espagne), Joaquín (Caudete, Espagne), Claude et Claude (Association géologique auboise). L'auteur remercie également vivement Francis pour ses précieuses informations biostratigraphiques concernant des différentes formations apto-albiennes de Grandpré, Edwige (Université Pierre et Marie Curie, Paris, France) pour son aide technique concernant l'utilisation du MEB et Charlčne (Muséum national d'Histoire naturelle de Paris, France) pour la réalisation des dessins. L'auteur remercie enfin les rapporteurs – Andreas (Muséum d'Histoire naturelle de Vienne, Autriche), Loďc (Université Pierre et Marie Curie, Paris, France) et Jaume (Musée des Sciences naturelles de Barcelone, Espagne) – pour leurs remarques constructives ayant permis d'améliorer la qualité de ce travail.
L. & E. (1846-1847).- Catalogue raisonné des espčces, des genres et des familles d'Échinides.- Annales des Sciences naturelles, Paris, (série 3), t. VI (1846), p. 306-374 (Pls. 15-16) ; t. VII (1847), p. 129-168 ; t. VIII (1847), p. 5-35 et p. 355-380.
M. & N. (2003).- Paleontology and stratigraphy of the Upper Cretaceous of sediments in the Ilgaz region.- The Arabian Journal for Science and Engineering, Dharhan, vol. 28, n° 1A, p. 25-34.
F. (2008).- Support for a Vraconian Stage between the Albian sensu stricto and the Cenomanian (Cretaceous System).- Carnets de Géologie - Notebooks on Geology, Brest, Memoir 2008/02 (CG2008_M02), 83 p. ( 9 Pls.).
C. (1875).- Sur le Gault et les couches entre lesquelles il est compris dans le Bassin de Paris.- Annales de la Société géologique du Nord, Lille, t. II (1874-75), p. 1-61.
C. (1878).- Mémoire sur le terrain crétacé des Ardennes et des régions voisines.- Annales de la Société géologique du Nord, Lille, t. V, p. 227- 487.
K. (1937).- Revision des Seeigel aus dem norddeutschen Jura. II. Die regulären Seeigel.- Abhandlungen der Preussischen Geologischen Landesanstalt, Berlin, Heft 174, p. 1-179 (1 Pl.).
J. (1912).- Esquisse géologique du Bassin de la Seybouse et de quelques régions voisines.- Bulletin du Service de la Carte géologique de l'Algérie, Alger, (2čme série), Stratigraphie, n° 6, iv + 394 p. (5 Pls.).
A. (1852).- Statistique géologique, minéralogique, minérallurgique et paléontologique du département de la Meuse.- Imprimerie de Lallemant, Verdun, 694 p.
A. (2000, inédit).- Biodiversité des bélemnites du Barrémo-Bédoulien du Sud-Est de la France et de l'Europe téthysienne : analyse comparative, implications paléobiogéographiques et paléoenvironnementales.- Mémoire de DEA (École Doctorale), Université Claude Bernard, Lyon, 39 p. (4 Pls.).
A., O. & J.A. (2012).- El registro de especies de Hemidiadema (Echinoidea) durante el Aptiense (Cretácico Inferior) en las cuencas del Maestrazgo, Salou-Garraf y Organyà.- Geo-Temas, Salamanca, vol. 13, p. 1-4.
C., C., M., R. & G. (1996).- La géologie du département du département de l'Aube : essai de synthčse avec carte géologique 1/ 200 000.- Association Géologique Auboise, Troyes, 213 p. (21 Pls.).
C. (Coord.) (2010).- Stratotype Albien.- Muséum national d'Histoire naturelle, Paris ; Biotope, Mčze ; BRGM, Orléans, 332 p. (Patrimoine géologique 2).
G. (1923).- Réunion extraordinaire de la Société Géologique de France dans le Gard, le Vaucluse et la Drôme.- Bulletin de la Société géologique de France, Paris, (série 4), t. 23, p. 460-487.
G. (1925).- Le Néocomien de la bordure orientale du Bassin de Paris.- Imprimerie J. Coubé & Fils, Nancy, 334 p. (11 Pls.).
G. (1857).- Étude sur les échinides fossiles du département de l'Yonne. Deuxičme partie. Terrain Crétacé. I. Étage néocomien.- Bulletin de la Société des Sciences historiques et naturelles de l'Yonne, Auxerre, t. XI, p. 402-433 (Pls. 47-50).
G. (1862-1867).- Paléontologie française. Terrain Crétacé. Tome 7 : Échinides réguliers.- Masson Ed., Paris, 892 p. (1204 Pls.).
G. (1869).- Échinides nouveaux ou peu connus.- Revue et Magasin de Zoologie pure et appliquée, Paris, 2čme série, t. XXI, p. 238-248 (Pls. 12-13).
B., S. & S. (2006).- Les kystes de dinoflagellés des Argiles Ostréennes du Bassin de Paris, France.- Revue de micropaléontologie, Paris, vol. 49, n° 1, p. 11-19.
E. (1858).- Synopsis des échinides fossiles.- Ch. Reinwald éditeur, Paris, LXIII-490 p. (44 Pls.).
F. & H. (1862).- Histoire naturelle des Zoophytes échinodermes.- Paris, 626 p. (10 Pls.).
P.M. (1889).- A revision of the Genera and great Groups of the Echinoidea.- Zoological Journal of the Linnean Society, (Zoology), London, vol. XXIII, p. 1-131.
M. (1938).- Excursion dans la Haute Vallée de la Tave.- Bulletin de la Société d'Étude des Sciences naturelles de Vaucluse, Avignon, n° 3, p. 45-56.
H.B. & D.L. (1966).- Echinacea. In : R.C. (Ed.), Echinodermata 3.- Treatise on Invertebrate Paleontology, The University of Kansas and The Geological Society of America, Inc., Boulder, Part U, vol. 2, p. U367-U440.
M. (1933).- Les oursins réguliers fossiles, évolution et classification.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Grenoble, t. XVII, fasc. 2, 125 p. (9 Pls.).
S. (1965).- État de nos connaissances sur la répartition stratigraphique des échinides du Crétacé inférieur du sud-est de la France.- Annales de la Faculté des Sciences de Marseille, t. 37, p. 57-70.
A. (1933).- Sur le passage des plaques ambulacraires alternantes aux plaques opposées chez quelques Échinides mésozoďques : tentative d'explication géométrique.- Bulletin de la Société neuchâteloise des Sciences naturelles, t. 58, p. 49-56.
W. (1907-1913).- Das Mesozoicum. Kreide. Unterkreide (Palaeocretacicum).- Lethaea Geognostica, Stuttgart, Teil 2, Band 3, p. 289-398 (Pls. 9-14).
A. & A.B. (2010).- The phylogeny and classification of post-Palaeozoic echinoids.- Journal of Systematic Palaeontology, London, vol. 8, issue 2, p. 147-212.
J. (1892a).- Note sur les Échinides des sables ferrugineux (Aptien) de Grandpré (Ardennes).- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, (série 3), t. XX, fasc. 4, p. XXXVII.
J. (1892b).- Recherches sur les Échinides de l'Aptien de Grandpré.- Bulletin de la Société géologique de France, Paris, (série 3), t. XX, p. 38-100 (Pls. II-IV).
J. (1916).- Sur l'existence de l'étage Valangien et sur l'oscillation barrémienne dans l'Aube et dans l'Yonne avec observations sur les échinides de ces étages.- Mémoires de la Société académique d'agriculture, des sciences, des arts et des belles-lettres du département de l'Aube, Troyes, t. 80, p. 19-94.
J. (1920).- Note sur quelques échinides du Crétacé inférieur de la Provence.- Notes provençales, Saint-Vallier-de-Thiey, n° 11, p. 1-24 (Pls. II-III).
J. (1928).- Note sur quelques Échinides du Crétacé d'Espagne communiqués par M. le Prof. y .- Boletin de la Real Sociedad española de Historia natural, Madrid, t. 28, n° 3, p. 147-157 (1 Pl.).
J. & P. (1909-1925).- Essai de nomenclature raisonnée des Échinides.- Librairie L. Ferričre, Chaumont, 607 p. (15 Pls.).
A. de (1868).- Note sur l'existence du terrain crétacé inférieur dans le Nord du bassin parisien.- Bulletin de la Société géologique de France, Paris, (2čme série), t. XXV, p. 284-289.
D. N. (1986).- The Cretaceous echinoid Boletechinus, with notes on the phylogeny of the Glyphocyphidae and Temnopleuridae.- Bulletin of the British Museum of natural History, (Geology series), London, vol. 40, n° 2, p. 59-90.
J. & C. (2006).- Equínoidos fósiles de la provincia de Alicante.- Memorias del Mupe 2, Museo paleontológico de Elche (Grupo Cultural Paleontológico de Elche), Elche, 264 p.
A.M. (1947).- Illustrazione degli echinidi giurassici della Somalia.- Atti della Accademia nazionale dei Lincei (Classe di scienze fisiche, matematiche e naturali), Roma, anno CCCXLIV, (ser. VIII), vol. I, sezione IIa, fasc. 5, p. 99-136 (1 Pl.).
F. (1984).- Signification biostratigraphique et paléoécologique des associations de Foraminifčres de L'Hauterivien-Barrémien du Sud-Est du Bassin de Paris (France).- Benthos 83, Pau, 2nd International Symposium of Benthic Foraminifera (Pau, April 1983), p. 401-408 (1 Pl.).
M., A. & F. (2004).- Succession des peuplements d'échinides du Crétacé inférieur dans les Haut Atlas Atlantique (Maroc) : révision systématique et intéręt stratigraphique.- Géobios, Lyon, vol. 37, p. 595-617.
J.-P. (1976).- Les calcaires urgoniens de Provence (Valanginien-Aptien inférieur). Stratigraphie, paléontologie, les paléoenvironnements et leur évolution.- Thčse Doctorat čs Sciences, Université d'Aix-Marseille II, 445 p.
J.-P. & S. (1976).- Les échinides du Crétacé inférieur provençal. Biostratigraphie et paléoécologie.- Géologie Méditerranéenne, Marseille, t. III, n° 1, p. 45-56.
P.L. (1986).- Un contact Jurassique-Crétacé à la limite des départements de la Meuse et des Ardennes.- Bulletin de l'Académie et Société lorraines des Sciences, Nancy, vol. 25, n° 3, p. 104-108.
A. & E. (1873).- Statistique agronomique de l'arrondissement de Vouziers, département des Ardennes.- Eugčne Jolly libraire-éditeur, 424 p.
R. (1976).- Continental sedimentation, soil genesis and marine transgression in the basal beds of the Cretaceous in the east of Paris Basin.- Sedimentology, Oxford, vol. 23, p. 235-253.
J. (1977).- Cretaceous echinoids of Stara Planina.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Beograd, t. LXI, p.181-202 (4 Pls.).
J. (1996).- Cretaceous Echinoidea from Djebel Djerissa, Central Tunisia.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Beograd, t. 60, n° 1, p. 169-194.
J. (1998).- Cidarida fossilized with spines from Barremian-Aptian rocks of Stara Planina, Eastern Serbia.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Beograd, t. 62, p. 193-211.
J. (2000).- Mesozoic Echinodermata of Stara Planina, Eastern Serbia.- Geološki anali Balkanskoga poluostrva [Annales géologiques de la Péninsule balkanique], Beograd, t. 63 (1999), p.71-89 (3 Pls.).
J.A., O., P., T., M., J., C., R., A., R. & S. (2008).- Bioestratigrafía integrada del Aptiense del Macizo del Garraf (Cadenas Costero-Catalanas, NE de España). Datos preminilares. In : , L. & J.C. (eds.) (2008).- Libro del resúmenes, XXIV Jornadas de la Sociedad Española de Paleontología. Museo del Jurássico de Asturias (MUJA), Colunga, 15-18 de octubre de 2008, p. 158-159.
T. (1943).- A monograph of the Echinoidea. Vol. 3,2 : Camarodonta 1 (Orthopsidae, Glyphocyphidae, Temnopleuridae, and Toxopneustidae).- C.A. Reitzel, Copenhagen, 546 p (56 Pls.).
M., B. & G. (2011).- The Aptian Stage: Back to fundamentals.- Episodes, Journal of International Geosciences, Bangalore, vol. 34, n° 3, p. 148-156.
A. d' (1850).- Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson Ed., Paris, vol. 2, 428 p.
E. (1901).- L'Aptien des envions d'Uzčs (Gard).- Bulletin de la Société géologique de France, Paris, (4čme série), t. I, p. 428-429.
M.-L. (1959).- Contribution à l'étude des Échinides fossiles du Maroc (Jurassique et Crétacé).- Notes et Mémoires du service géologique du Maroc, Rabat, n° 146, 183 p. (Pls. I-XVII).
J. (1965).- Recherches qualitatives sur les biocénoses marines des substrats meubles dragables de la région marseillaise.- Thčse, Marseille, 160 p.
F.-J. (1857).- Traité de paléontologie, Seconde édition.- Paris, t. 4, 768 p.
A. (1883).- Classification méthodique et Genera des Échinides fossiles et vivants.- Thčse doctorale, Alphonse Jourdan, Alger, 131 p. (1 Pl.).
P., B., F. & O. (1987).- Le golfe du Crétacé inférieur sur le Sud-Est du Bassin parisien : milieux (échinides, foraminifčres) et évolution de la transgression. In : Transgressions et régressions au Crétacé, France et régions voisines.- Mémoires géologiques de l'Université de Dijon, t. 11, p. 15-29.
S. (2002).- Les Ammonites de l'Hauterivien de l'Aube : systématique et évolution.- Association Géologique Auboise, Bulletin annuel, Troyes, n° 23, décembre 2002, p. 37-52 (1 Pl.).
S. & J. (reporters), R., M., C., A., S.N., J., O., J.M., Z. & J. (2009).- Report on the 3rd International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Vienna, Austria, 15th April 2008).- Cretaceous Research, London, vol. 30, n° 2, p. 496-502.
S., P.F. & J.A. (reporters), M.B., R., Y., M., C., V.I., A., B., V., H., Z., E.J., D., S., T.N., L.G., J.L., I.A., P. & O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Dijon, France, 30th August 2010).- Cretaceous Research, London, vol. 32, n° 6, p. 786-793.
M., L. & M. (2004).- The echinoderms of the Rügen White Chalk (Maastrichtian, Germany). In : T. & J.H. (eds).- Echinoderms: München.- Proceedings of the 11th International Echinoderm Conference, Munich, Germany, 6-10 October 2003, A.A. Balkema Publishers, p. 495-501.
M.-L. (1971).- Précisions stratigraphiques sur les Échinides du Crétacé inférieur de Tunisie.- Notes du Service géologique de Tunisie, Tunis, n° 34, p. 5-17.
J., J., A., K. & J.-P. (1986).– Le Crétacé inférieur sur le versant nord du haut-Atlas (région d'Izmi n'Tanout et Amizmiz) : données biostratigraphiques et évolutions sédimentaires.- Revue de la Faculté des Sciences de Marrakech, Section Sciences de la Terre, PICG-UNESCO n° 183, Marrakech, 1985, numéro spécial n° 2, p. 393-441.
J., J. & B. (1988).- Lithostratigraphy, biostratigraphy and sedimentary dynamics of the Lower Cretaceous deposits on the northern side of the wester High Atlas (Marocco).- Cretaceous Research, London, vol. 9, p. 141-158.
A.E. (2008).- Unterkreide-Lokalitäten im Maestrazgo/Nordost-Spanien.- Fundstellen, www.leitfossil.de, 47 p.
F. (1933).- Sur quelques formes de Céphalopodes de l'Hauterivien de l'Yonne et des régions voisines.- Travaux du Laboratoire de géologie de la Faculté des Sciences de Lyon, fasc. XXII, 22 p. (4 Pls.).
O. (1991).- Découverte de brushite dans les nodules phosphatés de Grandpré.- Bulletin des Académie et Société Lorraines des Sciences, Nancy, vol. 30, n° 2, p. 71-78.
M., J.A. & i J. (2007).– Fauna equínida del Maestrat, Cretaci inferior.- Asociación Paleontológica de Onda, 1 poster.
F.C. & A. (1842).- Statistique minéralogique et géologique du département des Ardennes.- Trécourt imprimeur, Mézičres, 554 p. (5 Pls.).
A.B. (editor) (2005 ; dernier accès le 30 Déc. 2012).- The Echinoid Directory.- En ligne à : http://www.nhm.ac.uk/research-curation/projects/echinoid-directory/
A.B. & C.W. (1993).- British Cretaceous echinoids. Part 3, Stirodonta 2 (Hemicidaroid, Arbacioida and Phymosomatoida, part 1).- Monograph of the Palaeontographical Society, London, Publication n° 593, part of Vol. 147 for 1993, p. 199-267 (Pls. 73-92).
A.B. & C.W. (1996).- British Cretaceous Echinoids. Part 4. Stirodonta 3 (Phymosomatidae, Pseudodiadematidae) and Camarodonta.- Monograph of the Palaeontographical Society, London, Publication n° 602, part of Vol. 150 for 1996, p. 268-341 (Pls. 93-114).
V. (1954).- Le Crétacé inférieur de l'Est du bassin de Paris d'aprčs de nouvelles données.- Bulletin de la Société géologique de France, Paris, (série 6), t. IV, fasc. 7-9, p. 597-608.
V. (1962).- Histoire géologique de la Haute-Marne et des régions voisines.- Thčse Faculté des Sciences de Paris. 223 p.
Dong-li & Bing-gao (1983).- Aspects of the Marine Cretaceous of China.- Cretaceous Research, London, vol. 4, p. 145-158.
A. & P. (2010).- Échinides nouveaux, peu ou mal connus.- Annales de la Société d'Histoire naturelle du Boulonnais, Boulogne-sur-Mer, t. IX, fasc. 3, p. 1-24.
D.A. (1908).- Révision des échinides fossiles de l'Yonne. 2čme partie (Crétacé).- Bulletin de la Société des Sciences historiques et naturelles de l'Yonne, Auxerre, t. LXI, p. 183-385.
M.P. (1991, inédit).- Revisión de los equínidos del Cretácico inferior y medio español.- Colección Tesis Doctorales, Universidad Complutense Madrid, Facultad de Ciencias geológicas, Madrid, nş 250/93, t. I y II, 714 p. (33 Pls.).
M.P. (2003).- Los equínidos regulares de la provincia de Castellón.- Nemus, Revista de l'Ateneu de Natura, Castelló de la Plana, n° 3, p. 37-91 (4 Pls.).
T. (1870).- Monograph of British fossil Echinodermata from the Cretaceous formations, vol. 1, part. III (on the Diademadae).- Palaeontographical Society Monograph, London, vol. XXIII (1869), p. 113-136 (Pls. XXII, XXIX, XXIXa, XXIXb).
Sheng-qui (1992).- Cretaceous echinoid fauna of Xizang (Tibet).- Bulletin of Nanjing Institut of Geology and Paleontology Academia Sinica, n° 13, p. 96-137 (13 Pls.).
Bing-gao, Dong-li, Sheng-qui & Chun-hui (1981).- New observations on the Cretaceous of Baingoin and Xainza areas, northern Xizang.- Acta Stratigraphica Sinica [Journal of Stratigraphy], Beijing, vol. 5, n° 4, p. 313-315.
Toutes les photographies ont été prises par l'auteur avec un Canon EOS 350D muni d'un objectif 18-55 mm, associé à trois bagues-allonges Kepcor 31 mm, 21 mm et 13 mm. Les échinides ont été éclairés en utilisant des ampoules "day light".
En vues apicale et orale, l'astérisque "*" indique l'interambulacre 2 supportant à son extrémité apicale la madréporite (plaque génitale 2).
Les photographies au MEB ont été réalisées par l'auteur sans métallisation des échantillons.
All photographs were taken by the author using a Canon EOS 350D equipped with a 18-55 mm lens, together with Kepcor 31 mm, 21 mm and 13 mm extension rings. The echinoids were illuminated using "day light" bulbs.
In apical and oral views, the asterik "*" indicates the interambulacrum 2 bearing the madreporite on his apical end (genital plate 2).
The SEM-images were made by the author without metal coating of the samples.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 1 - Plate 1
Hemidiadema rugosum , 1846 – Holotype / Holotype
EM 13095 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Premičres figurations photographiques de l'holotype / First photographic representation of the holotype.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 2 / Ambital view centered on interambulacrum 2.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 3 / Ambital view centered on interambulacrum 3.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 4 / Ambital view centered on interambulacrum 4.
Fig. 5 : Vue supra-ambitale centrée sur l'interambulacre 2 / Supra-ambital view centered on interambulacrum 2.
Fig. 6 : Vue générale de la face orale / General oral view.
Fig. 7 : Vue ambitale centrée sur l'ambulacre III / Ambital view centered on ambulacrum III.
Fig. 8 : Vue ambitale centrée sur l'ambulacre V / Ambital view centered on ambulacrum V.
Fig. 9 : Vue ambitale centrée sur l'ambulacre II / Ambital view centered on ambulacrum II.
Fig. 10 : Vue supra-ambitale centrée sur l'ambulacre III / Supra-ambital view centered on ambulacrum III.
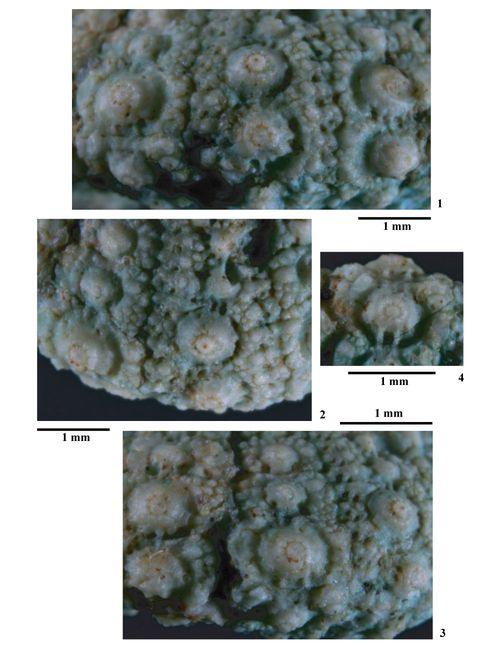
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 2 - Plate 2
Hemidiadema rugosum , 1846 – Holotype / Holotype
EM 13095 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Premičres figurations photographiques de l'holotype / First photographic representations of the holotype.
Détails montrant les caractéristiques des tubercules primaires : mamelons perforés, cônes crénelés, scrobicules étroits et profonds / Details showing the characteristics of primary tubercles : perforate mamelons, crenulate platforms, deep and narrow areoles.
Fig. 1 : Détails de l'ambulacre III en vue ambitale / Details of the ambulacrum III in ambital view.
Fig. 2 : Détails de l'ambulacre III en vue supra-ambitale / Details of the ambulacrum III in supra-ambital view.
Fig. 3 : Détails de l'interambulacre 2 en vue ambitale / Details of the interambulacrum 2 in ambital view.
Fig. 4 : Détails d'un tubercule primaire interambulacraire supra-ambital / Details of a supra-ambital interambulacral primary tubercle.
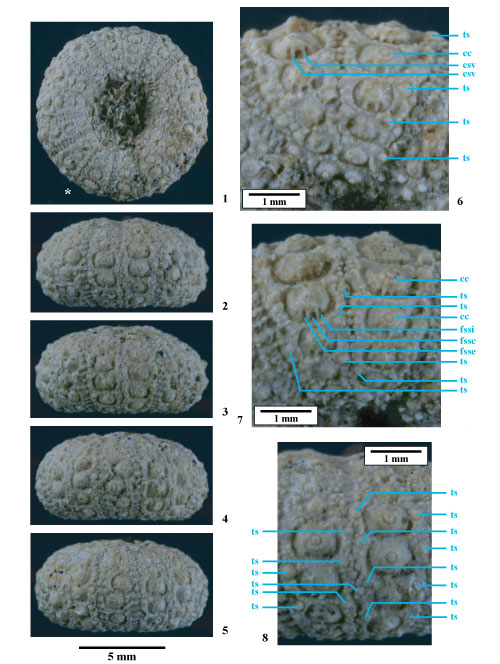
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Planche 3 - Plate 3
Hemidiadema rugosum , 1846
MNHN.F R62314 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Premičres figurations photographiques du spécimen figuré (dessiné) par (1892b, Pl. 4, figs. 5-6) / First photographic representations of the specimen figured (drawn) by (1892b, Pl. 4, figs. 5-6).
Fig. 1 : Vue générale de la face orale / General oral view.
Fig. 2 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 4 / Ambital view centered on interambulacrum 4.
Fig. 4 : Vue ambitale centrée sur l'ambulacre I / Ambital view centered on ambulacrum I.
Fig. 5 : Vue ambitale centrée sur l'ambulacre V / Ambital view centered on ambulacrum V.
Figs. 6-7 : Détails de l'interambulacre 4 en vue infra-ambitale (sous deux angles légčrement différents) / Details of the interambulacrum 4 in infra-ambital view (from two slighty different angles).
Fig. 8 : Details de l'interambulacre 5 en vue ambitale / Details of the interambulacrum 5 in ambital view.
Nomenclature / Nomenclature :
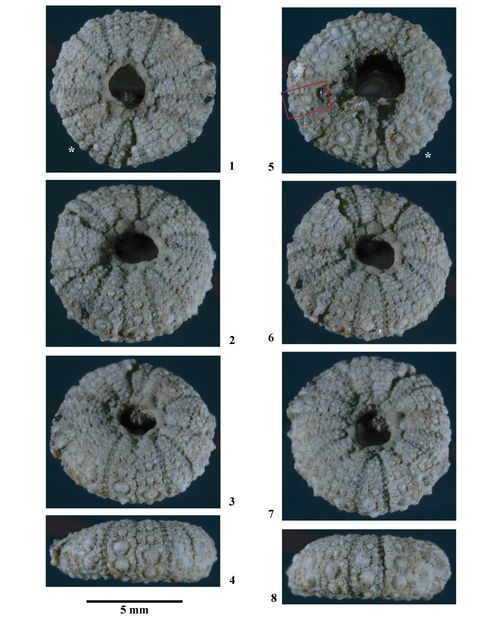
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 4 - Plate 4
Hemidiadema rugosum , 1846
MNHN.F A20547 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Individu avec fossettes suturales moyennement développées / Individu with moderately developed suturals pits.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue supra-ambitale centrée sur l'interambulacre 4 / Supra-ambital view centered on interambulacrum 4.
Fig. 3 : Vue supra-ambitale centrée sur l'interambulacre 5 / Supra-ambital view centered on interambulacrum 5.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
Fig. 5 : Vue générale de la face orale / General oral view.
Fig. 6 : Vue supra-ambitale centrée sur l'ambulacre V / Supra-ambital view centered on ambulacrum V.
Fig. 7 : Vue supra-ambitale centrée sur l'ambulacre I / Supra-ambital view centered on ambulacrum I.
Fig. 8 : Vue ambitale centrée sur l'ambulacre I / Ambital view centered on ambulacrum I.
Encadré rouge : voir Pl. 15 ![]() / Red frame : see Pl. 15
/ Red frame : see Pl. 15 ![]()

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 5 - Plate 5
Hemidiadema rugosum , 1846
MNHN.F A20547 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Individu avec fossettes suturales moyennement développées / Individual with moderately developed sutural pits.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue ambitale centrée sur l'ambulacre V / Ambital view centered on ambulacrum V.
Fig. 3 : Vue ambitale centrée sur l'ambulacre I / Ambital view centered on ambulacrum I.
Fig. 4 : Vue ambitale centrée sur l'interambulacre 1 / Ambital view centered on interambulacrum 1.
Fig. 5 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
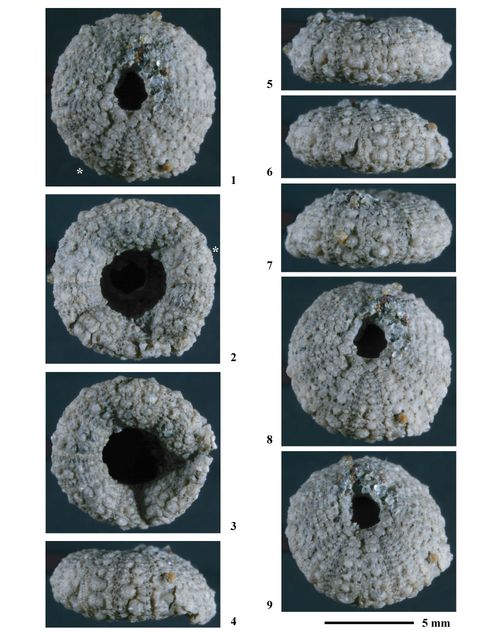
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 6 - Plate 6
Hemidiadema rugosum , 1846
MNHN.F A33784 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Individu avec fossettes suturales moyennement développées / Individual with moderately developed sutural pits.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue générale de la face orale / General oral view.
Fig. 3 : Vue infra-ambitale centrée sur l'interambulacre 2 / Infra-ambital view centered on interambulacrum 2.
Fig. 4 : Vue ambitale centrée sur l'ambulacre III / Ambital view centered on ambulacrum III.
Fig. 5 : Vue ambitale centrée sur l'interambulacre 1 / Ambital view centered on interambulacrum 1.
Fig. 6 : Vue ambitale centrée sur l'interambulacre 2 / Ambital view centered on interambulacrum 2.
Fig. 7 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
Fig. 8 : Vue supra-ambitale centrée sur l'ambulacre III / Supra-ambital view centered on ambulacrum III.
Fig. 9 : Vue supra-ambitale centrée sur l'interambulacre 2 / Supra-ambital view centered on interambulacrum 2.
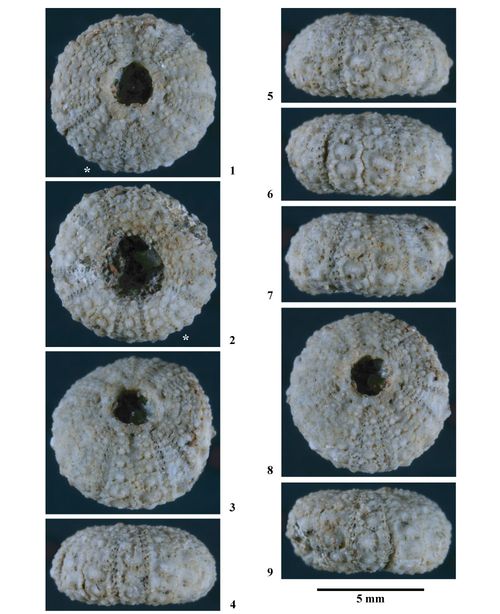
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 7 - Plate 7
Hemidiadema rugosum , 1846
MNHN.F A33782 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Individu avec fossettes suturales fortement développées et fusionnées / Individual with strongly developed and merged sutural pits.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue générale de la face orale / General oral view.
Fig. 3 : Vue supra-ambitale centrée sur l'interambulacre 4 / Supra-ambital view centered on interambulacrum 4.
Fig. 4 : Vue ambitale centrée sur l'ambulacre I / Ambital view centered on ambulacrum I.
Fig. 5 : Vue ambitale centrée sur l'interambulacre 1 / Ambital view centered on interambulacrum 1.
Fig. 6 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
Fig. 7 : Vue ambitale centrée sur l'interambulacre 4 / Ambital view centered on interambulacrum 4.
Fig. 8 : Vue supra-ambitale centrée sur l'ambulacre V / Supra-ambital view centered on ambulacrum V.
Fig. 9 : Vue ambitale centrée sur l'ambulacre II / Ambital view centered on ambulacrum II.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 8 - Plate 8
Hemidiadema rugosum , 1846
MNHN.F A33782 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Détails de l'ambulacre V et de l'interambulacre 5 et nomenclature descriptive utilisée / Details of the ambulacrum V and the interambulacrum 5 and descriptive nomenclature used.
Fig. 1 : Détails de l'ambulacre V en vue supra-ambitale / Details of the ambulacrum V in supra-ambital view.
Fig. 2 : Détails de l'interambulacre 5 en vue supra-ambitale / Details of the interambulacrum 5 in supra-ambital view.
Fig. 3 : Détails de l'ambulacre II en vue supra-ambitale / Details of the ambulacrum II in supra-ambital view.
Fig. 4 : Détails de l'interambulacre 5 en vue ambitale / Details of the interambulacrum 5 in ambital view.
Nomenclature / Nomenclature :
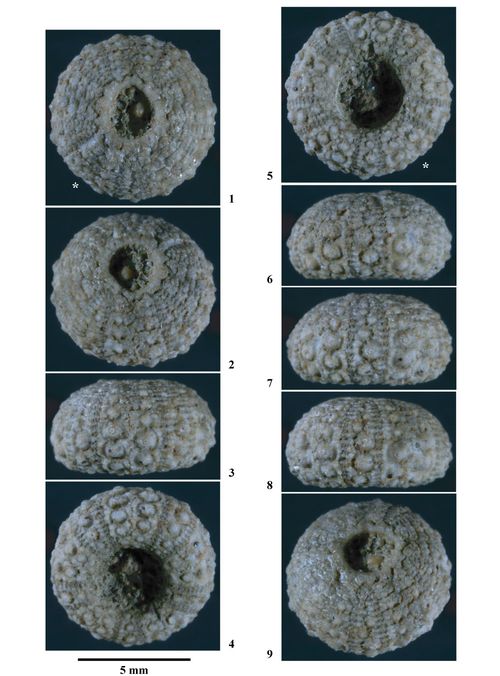
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 9 - Plate 9
Hemidiadema rugosum , 1846
MNHN.F A33783 ; Coll.
Clansayésien (Aptien supérieur sensu gallico), Grandpré, Ardennes, France / Clansayesian (Upper Aptian sensu gallico), Grandpré, Ardennes, France.
Individu avec fossettes suturales moyennement développées / Individual with moderately developed sutural pits.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue supra-ambitale centrée sur l'interambulacre 5 / Supra-ambital view centered on interambulacrum 5.
Fig. 3 : Vue ambitale centrée sur l'interambulacre 3 / Ambital view centered on interambulacrum 3.
Fig. 4 : Vue infra-ambitale centrée sur l'interambulacre 3 / Infra-ambital view centered on interambulacrum 3.
Fig. 5 : Vue générale de la face orale / General oral view.
Fig. 6 : Vue ambitale centrée sur l'ambulacre II / Ambital view centered on ambulacrum II.
Fig. 7 : Vue ambitale centrée sur l'ambulacre III / Ambital view centered on ambulacrum III.
Fig. 8 : Vue ambitale centrée sur l'ambulacre IV / Ambital view centered on ambulacrum IV.
Fig. 9 : Vue supra-ambitale centrée sur l'ambulacre IV / Supra-ambital view centered on ambulacrum IV.
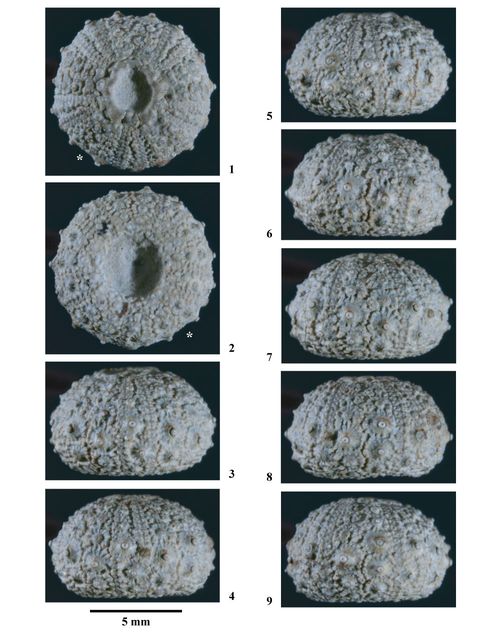
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 10 - Plate 10
Hemidiadema neocomiense (, 1869) – Néotype / Neotype
ENSGN / MAN ; Coll. n° 384
Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum), Ville-sur-Saulx, Meuse, France / Lower Hauterivian (? Lyticoceras nodosoplicatum Zone), Ville-sur-Saulx, Meuse, France.
Fig. 1 : Vue générale de la face apicale / General apical view.
Fig. 2 : Vue générale de la face orale / General oral view.
Fig. 3 : Vue ambitale centrée sur l'ambulacre I / Ambital view centered on ambulacrum I.
Fig. 4 : Vue ambitale centrée sur l'ambulacre II / Ambital view centered on ambulacrum II.
Fig. 5 : Vue ambitale centrée sur l'ambulacre V / Ambital view centered on ambulacrum V.
Fig. 6 : Vue ambitale centrée sur l'interambulacre 5 / Ambital view centered on interambulacrum 5.
Fig. 7 : Vue ambitale centrée sur l'interambulacre 1 / Ambital view centered on interambulacrum 1.
Fig. 8 : Vue ambitale centrée sur l'interambulacre 2 / Ambital view centered on interambulacrum 2.
Fig. 9 : Vue ambitale centrée sur l'interambulacre 4 / Ambital view centered on interambulacrum 4.
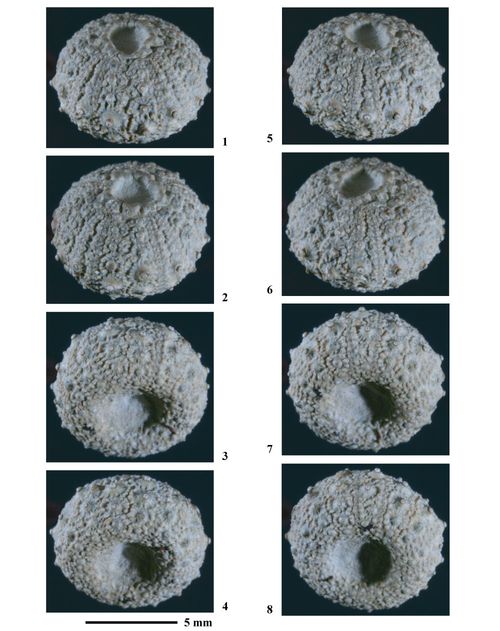
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 11 - Plate 11
Hemidiadema neocomiense (, 1869) – Néotype / Neotype
ENSGN / MAN ; Coll. n° 384
Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum), Ville-sur-Saulx, Meuse, France / Lower Hauterivian (? Lyticoceras nodosoplicatum Zone), Ville-sur-Saulx, Meuse, France.
Fig. 1 : Vue supra-ambitale centrée sur l'interambulacre 5 / Supra-ambital view centered on interambulacrum 5.
Fig. 2 : Vue supra-ambitale centrée sur l'interambulacre 4 / Supra-ambital view centered on interambulacrum 4.
Fig. 3 : Vue infra-ambitale centrée sur l'interambulacre 1 / Infra-ambital view centered on interambulacrum 1.
Fig. 4 : Vue infra-ambitale centrée sur l'interambulacre 2 / Infra-ambital view centered on interambulacrum 2.
Fig. 5 : Vue supra-ambitale centrée sur l'ambulacre V / Supra-ambital view centered on ambulacrum V.
Fig. 6 : Vue supra-ambitale centrée sur l'ambulacre I / Supra-ambital view centered on ambulacrum I.
Fig. 7 : Vue infra-ambitale centrée sur l'ambulacre I / Infra-ambital view centered on ambulacrum I.
Fig. 8 : Vue infra-ambitale centrée sur l'ambulacre V / Infra-ambital view centered on ambulacrum V.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 12 - Plate 12
Hemidiadema neocomiense (, 1869) – Néotype / Neotype
ENSGN / MAN ; Coll. n° 384
Hauterivien inférieur (? Zone à Lyticoceras nodosoplicatum), Ville-sur-Saulx, Meuse, France / Lower Hauterivian (? Lyticoceras nodosoplicatum Zone), Ville-sur-Saulx, Meuse, France.
Détails de quelques ambulacres et interambulacres / Details of some ambulacra and interambulacra.
Fig. 1 : Interambulacre 5 en vue supra-ambitale / Interambulacrum 5 in supra-ambital view.
Fig. 2 : Interambulacre 4 en vue supra-ambitale / Interambulacrum 4 in supra-ambital view.
Fig. 3 : Ambulacre I en vue supra-ambitale / Ambulacrum I in supra-ambital view.
Fig. 4 : Ambulacre V en vue supra-ambitale / Ambulacrum V in supra-ambital view.
Fig. 5 : Interambulacre 5 en vue ambitale / Interambulacrum 5 in ambital view.
Fig. 6 : Interambulacre 1 en vue ambitale / Interambulacrum 1 in ambital view.
Fig. 7 : Ambulacre II en vue ambitale / Ambulacrum II in ambital view.
Fig. 8 : Ambulacre V en vue ambitale / Ambulacrum V in ambital view.
Fig. 9 : Interambulacre 5 en vue infra-ambitale / Interambulacrum 5 in infra-ambital view.
Fig. 10 : Interambulacre 4 en vue infra-ambitale / Interambulacrum 4 in infra-ambital view.
Fig. 11 : Ambulacre II en vue infra-ambitale / Ambulacrum II in infra-ambital view.
Fig. 12 : Ambulacre I en vue infra-ambitale / Ambulacrum I in infra-ambital view.
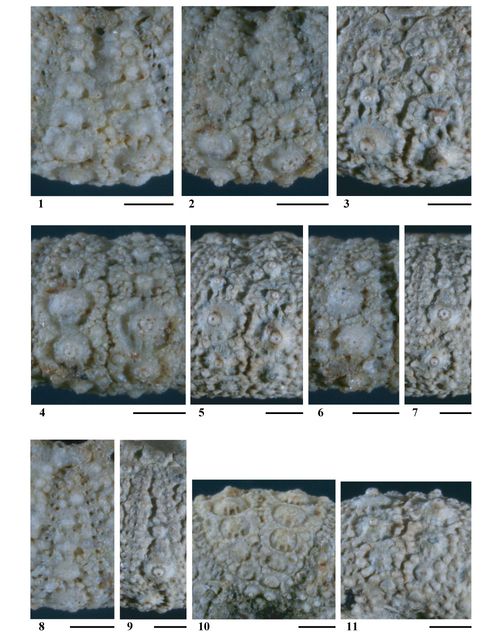
Click on thumbnail to enlarge the image
[Cliquer sur la miniature
pour agrandir l'image]
Planche 13 - Plate 13
Figs. 1-3 : Comparaison des interambulacres en vue supra-ambitale / Comparison of the interambulacra in supra-ambital view.
Figs. 4-5 : Comparaison des interambulacres en vue ambitale / Comparison of the interambulacra in ambital view.
Figs. 6-7 : Comparaison des ambulacres en vue ambitale / Comparison of the ambulacra in ambital view.
Figs. 8-9 : Comparaison des ambulacres en vue supra-ambitale / Comparison of the ambulacra in supra-ambital view.
Figs. 10-11 : Comparaison des interambulacres en vue infra-ambitale / Comparison of the interambulacra in infra-ambital view.
Barre d'échelle : 1 mm / Scale bar: 1 mm
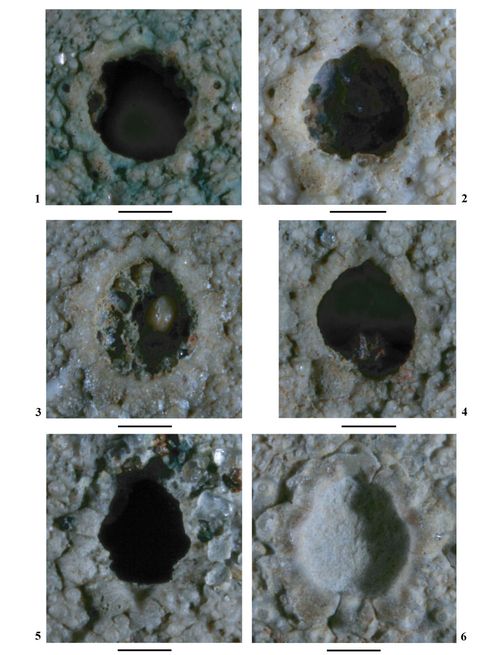
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 14 - Plate 14
Disques apicaux / Apical discs.
Fig. 1 : H. rugosum (EM 13095 ; Coll. ) ; Holotype / Holotype.
Fig. 2 : H. rugosum (MNHN.F A33782 ; Coll. ).
Fig. 3 : H. rugosum (MNHN.F A33783 ; Coll. ).
Fig. 4 : H. rugosum (MNHN.F A20547 ; Coll. ).
Fig. 5 : H. rugosum (MNHN.F A33784 ; Coll. ).
Fig. 6 : H. neocomiense (ENSGN / MAN, coll. n° 384) ; Neotype / Neotype.
Barre d'échelle : 1 mm / Scale bar: 1 mm
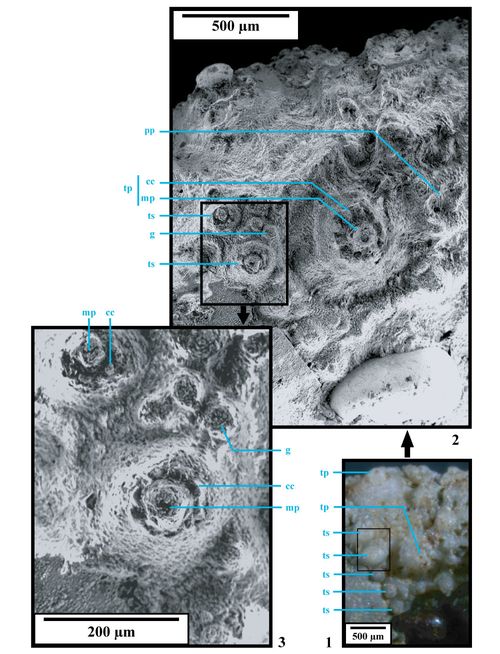
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir
l'image]
Planche 15 - Plate 15
Tubercules primaires et tubercules secondaires ambulacraires infra-ambitaux d'H. rugosum (MNHN.F A20547 ; Coll. ) observés au MEB / SEM-pictures of infra-ambital ambulacral primary and secondary tubercles of H. rugosum (MNHN.F A20547 ; Coll. ).
Fig. 1 : Ambulacre IV en vue supra-ambitale (voir encadré rouge sur la
Pl. 4 ![]() , fig. 5) / Ambulacrum
IV in supra-ambital view (see red frame on
Pl. 4
, fig. 5) / Ambulacrum
IV in supra-ambital view (see red frame on
Pl. 4 ![]() , fig. 5).
, fig. 5).
Fig. 2 : Męme ambulacre observé au MEB / SEM-picture of the same ambulacrum.
Fig. 3 : Détails sur les tubercules secondaires crénelés et perforés / Details on perforated and crenulate secondary tubercles.
Nomenclature / Nomenclature :

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Planche 16 - Plate 16
Plaques interambulacraires de la zone ambitale du néotype d'H. neocomiense (ENSGN / MAN, coll. n° 384) observées au MEB / SEM-pictures of interambulacral plates in ambital view of the neotype of H. neocomiense (ENSGN / MAN, coll. n° 384).
Nomenclature / Nomenclature :