![]()
Contents
[1. Introduction] [2. History overview]
[3. Genus, species and author]
[4. A case study]
[5. Implications for the future and final recommendations] [Bibliographic references]
CNRS-UMR 6538, Université de Bretagne Occidentale, 6 Avenue Le Gorgeu, CS 93837, 29238 Brest
Cedex 3 (France)
Manuscript online since December 25, 2012
[Editor and copy editor: Christian C. ;
language editor: Simon F. ]
![]()
Since the 19th century, about 1800 brachiopod genera or subgenera have been erected for the period covering the Silurian to the Devonian. In recent years, there has been a pronounced increase in the number of genera containing a single species. Without questioning the technological advances, one can, however, question the value and usefulness of monospecific genera both for systematics and palaeogeographical studies. Some editorial policy regarding erection of new monospecific genera and/or the evaluation of calls for projects are now needed.
Monospecific genera; brachiopods; emendation; systematics; methodology.
R. (2012).- Inflation in the number of monospecific genera in brachiopod: progress or regression? A trivial approach to a real problem.- Carnets de Géologie [Notebooks on Geology], Brest, Letter 2012/04 (CG2012_L04), p. 285-292.
Inflation du nombre de genres monospécifiques chez les brachiopodes : progrès ou régression ? Une approche prosaïque d'un problème réel.- Depuis le XIXe siècle, environ 1800 genres ou sous-genres de brachiopodes ont été créés pour la période couvrant le Silurien et le Dévonien. On constate, depuis quelques années, une inflation considérable du nombre de genres n'incluant qu'une seule et unique espèce. Sans remettre en cause les progrès technologiques, on peut cependant s'interroger sur la pertinence de la méthodologie adoptée par nombre d'auteurs et sur l'utilité in fine des genres monospécifiques, tant sur le plan de la systématique que sur celui des disciplines annexes. Certaines politiques éditoriales ou encore l'évaluation des appels à projets ne sont peut-être pas étrangères à ce constat et mériteraient d'être revues.
Genres monospécifiques ; brachiopodes ; émendation ; systématique ; méthodologie.
While building a database on Silurian to Devonian brachiopods, I was intrigued to find that the names of some authors were encountered very frequently. A closer examination of these data shows that of the 388 authors who erected 1,865 brachiopod genera from late Ordovician to early Carboniferous, 35 (9%) authors erected 918 (58%) genera. These authors, often called "splitters," are hereinafter referred to as "prolific authors". The data relating to genera were extracted from the Treatise on Invertebrate Paleontology, part H, Brachiopoda, revised (, 2000, 2002, 2006; , 2007). Data on species were collected on databases available on the Web, namely in the Index to Organic Names (ION), which also includes the Zoological Records database, and ZipcodeZoo. Subgenera were treated at generic rank. In this paper the historical development and recent dramatic increase in the number of monospecific genera is analyzed.
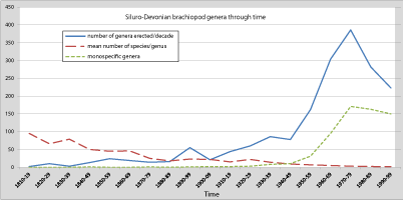
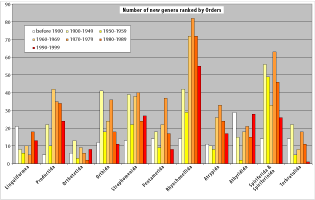
During the 19th and first half of the 20th centuries, the erection of new genera was steady and moderate, but increased during the second half of the 20th century, reaching a peak during the 1970s (Fig. 1 ![]() ). Activity was not uniform across different brachiopod groups, some have intervals when many genera were erected, and others intervals when few or no genera were erected (e.g., lingulids, athyrides, Fig. 2
). Activity was not uniform across different brachiopod groups, some have intervals when many genera were erected, and others intervals when few or no genera were erected (e.g., lingulids, athyrides, Fig. 2 ![]() ). This relates, to some extent, to the publication of major monographs, the retirement of older authors, the arrival of new researchers, the development of new research areas, or a revision of a group for stratigraphic purposes, but this is insufficient to account for the general trend within the data. Whatever the activity within a group, be it irregular, or regular and intense (e.g., for spiriferids and rhynchonellids, in which many new genera were erected in recent times), we always observe a similar trend, that is a progression towards a greater number of monospecific genera.
). This relates, to some extent, to the publication of major monographs, the retirement of older authors, the arrival of new researchers, the development of new research areas, or a revision of a group for stratigraphic purposes, but this is insufficient to account for the general trend within the data. Whatever the activity within a group, be it irregular, or regular and intense (e.g., for spiriferids and rhynchonellids, in which many new genera were erected in recent times), we always observe a similar trend, that is a progression towards a greater number of monospecific genera.
Click on thumbnail to enlarge the image.
Figure 1: Evolution by decade of the number of new genera (in blue), mean number of species per genus (in red) and number of monospecific genera (in green) for Silurian-Devonian brachiopods.
After a slight decrease in the erection of new genera during the mid-20th century, probably due to World War II (Fig. 1 ![]() ), the increase in publication activity in the
1950s is related to the increasing number of authors erecting new genera. The number of authors was relatively constant (from 22 to 31 per decade) in the first half of the 20th century, but reached 41 per decade in the 1950s. This trend accelerated in the 1960s with an increase up to the 1990s, from which time the number of active authors per decade has stabilized, remaining essentially unchanged until now (between 92 and 107 from 1970 to present). One might assume that technological progress has an effect on the increase in average number of genera erected by an author. However, this does not explain why only a small number of authors is involved in this inflation, and secondly this argument would also be applicable for the period from the 19th century to the first half of the 20th century which was also facing many technological advances and progress in the biological sciences, without resulting in an explosion in the erection of new genera. Therefore, the recent inflation of new genera is not only related to an increase in the number of active authors but also to a change in the taxonomic approach. One can notice that a similar trend was observed in the late 19th century, a period also characterized by a relatively stable number of authors per decade. However this increase was moderate, with no sharp contrast to previous decades.
), the increase in publication activity in the
1950s is related to the increasing number of authors erecting new genera. The number of authors was relatively constant (from 22 to 31 per decade) in the first half of the 20th century, but reached 41 per decade in the 1950s. This trend accelerated in the 1960s with an increase up to the 1990s, from which time the number of active authors per decade has stabilized, remaining essentially unchanged until now (between 92 and 107 from 1970 to present). One might assume that technological progress has an effect on the increase in average number of genera erected by an author. However, this does not explain why only a small number of authors is involved in this inflation, and secondly this argument would also be applicable for the period from the 19th century to the first half of the 20th century which was also facing many technological advances and progress in the biological sciences, without resulting in an explosion in the erection of new genera. Therefore, the recent inflation of new genera is not only related to an increase in the number of active authors but also to a change in the taxonomic approach. One can notice that a similar trend was observed in the late 19th century, a period also characterized by a relatively stable number of authors per decade. However this increase was moderate, with no sharp contrast to previous decades.
Click on thumbnail to enlarge the image.
Figure 2: Number of new Silurian-Devonian brachiopod genera ranked by decades and by Orders (former "Inarticulate" orders aggregated at the Subphylum level).
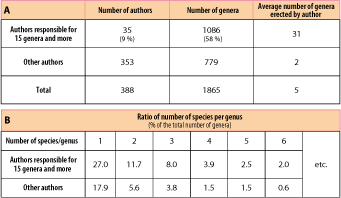
During the last two centuries, nearly 400 authors have erected more than 1,800 genera for the time interval from the late Ordovician to the early Carboniferous; each author has erected an average of about 5 genera. However less than 10% of these authors, here called "prolific authors", have created over 1,000 new genera (about 60% of the total), with an average of 31 genera per author (30 if we consider only publications as first author). Putting these prolific authors aside of the remainder, the average falls to two genera per author (Fig. 3A ![]() ). Further analysis of the structure of genera erected by prolific authors shows that this increase in productivity is accompanied by an increase in the number of monospecific genera. The numbers of monospecific genera erected by these authors markedly exceeds those erected by other authors, especially from the 1950s onwards. Furthermore, the frequency distribution curve of the number of species per genus shows a very rapid decline of genera including two, three or more species (Fig. 3B
). Further analysis of the structure of genera erected by prolific authors shows that this increase in productivity is accompanied by an increase in the number of monospecific genera. The numbers of monospecific genera erected by these authors markedly exceeds those erected by other authors, especially from the 1950s onwards. Furthermore, the frequency distribution curve of the number of species per genus shows a very rapid decline of genera including two, three or more species (Fig. 3B ![]() ). It may be noted that the upward trend in productivity is a relatively recent phenomenon since only seven prolific authors published their results before the 1950s.
). It may be noted that the upward trend in productivity is a relatively recent phenomenon since only seven prolific authors published their results before the 1950s.
Click on thumbnail to enlarge the image.
Figure 3: Table showing the ratio of "normal" versus "prolific" authors, the difference in their respective production (A) and the ratio of species per genus in the two categories (B). Data include genera present in uppermost Ordovician to lowermost Carboniferous.
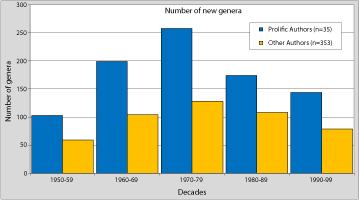
If we consider the change in the number of new genera erected in recent decades, we can see that the difference of productivity between prolific authors and others, which is particularly well expressed in the 1960s and the 1970s, remains constant with a ratio of about 50%, regardless of how many genera were created by decade (Fig. 4 ![]() ). A closer analysis of these data shows that the difference between the two types of authors is based primarily on the processing of monospecific genera (and genera with few species). Prolific authors have created more than 42% of monospecific genera. In contrast this ratio is reversed in favour of normal authors for the multi-specific genera (Fig. 5
). A closer analysis of these data shows that the difference between the two types of authors is based primarily on the processing of monospecific genera (and genera with few species). Prolific authors have created more than 42% of monospecific genera. In contrast this ratio is reversed in favour of normal authors for the multi-specific genera (Fig. 5 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 4: Comparison of the number of Silurian-Devonian brachiopod genera erected by "prolific" and other authors for each decade since 1950.
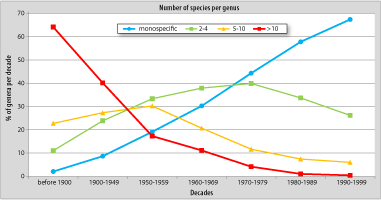
In order to examine the changes in the number of species per genus during the second half of the last century, we can arbitrarily classify genera into three categories:
and compare the evolution of these categories in both groups of authors.
The first category shows a significant and steady increase: the percentage of monospecific genera created by prolific authors in the 1990s is four times higher than in the 1950s (Fig. 6A ![]() ). This trend is also true for other authors, but the ratio is here by only 1 to 2. The gap between the curves representing the prolific authors and other authors is increasing steadily. Regarding genera comprising between 2 and 5 species (Fig. 6B
). This trend is also true for other authors, but the ratio is here by only 1 to 2. The gap between the curves representing the prolific authors and other authors is increasing steadily. Regarding genera comprising between 2 and 5 species (Fig. 6B ![]() ), the evolution is parallel in both cases, but with a regular decrease and an overall impression of stability. Finally, for the genera with more than five species, we observe, as might be expected, a severe drop particularly well marked among the prolific authors. Contrary to what is observed in monospecific genera, the gap between the curves of both groups of authors decreases regularly to finally disappear when the prolific authors' curve passes under that of normal authors in the 1980s (Fig. 6C
), the evolution is parallel in both cases, but with a regular decrease and an overall impression of stability. Finally, for the genera with more than five species, we observe, as might be expected, a severe drop particularly well marked among the prolific authors. Contrary to what is observed in monospecific genera, the gap between the curves of both groups of authors decreases regularly to finally disappear when the prolific authors' curve passes under that of normal authors in the 1980s (Fig. 6C ![]() ). The predominance of genera including few species is not surprising per se:
(1978) had previously demonstrated that, at least in invertebrates, "monotypic taxa, regardless of group or rank, are generally far more common than any other single type", which results in what he termed hollow curves in the frequency distribution of taxonomic units
(e.g., in our Fig. 5
). The predominance of genera including few species is not surprising per se:
(1978) had previously demonstrated that, at least in invertebrates, "monotypic taxa, regardless of group or rank, are generally far more common than any other single type", which results in what he termed hollow curves in the frequency distribution of taxonomic units
(e.g., in our Fig. 5 ![]() ). However, no explanation has been advanced so far to explain such a phenomenon. Concerning Paleozoic brachiopods the problem lies in a particularly high proportion of monospecific genera due to the hyperactivity of a relatively small cohort of authors. It can therefore be suspected that the human factor has taken precedence over the biological factors.
). However, no explanation has been advanced so far to explain such a phenomenon. Concerning Paleozoic brachiopods the problem lies in a particularly high proportion of monospecific genera due to the hyperactivity of a relatively small cohort of authors. It can therefore be suspected that the human factor has taken precedence over the biological factors.
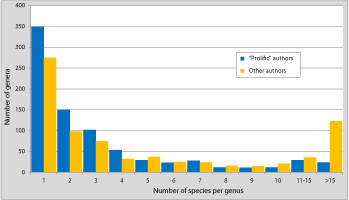
Click on thumbnail to enlarge the image.
Figure 5: Distribution of the number of species per genus in "normal" and "prolific" authors for Silurian-Devonian brachiopods since 1950.
The increasing number of monospecific genera is not only realized at the expense of genera with a (perhaps too) large number of species, but also at the expense of genera with a "reasonable" number of species (2 to 5, which is comparable to the standards observed in the biology of present organisms) whose numbers had steadily increased until the 1970s and have since steadily declined. Curiously, when publishing, prolific authors tend to opt to publish jointly: this represents 49% of the genera with multiple authors, to be compared with 31% of associations between prolific authors and normal authors, and 20% of normal authors associated with each other.

Click on thumbnail to enlarge the image.
Figure 6: Evolution of the structure of Silurian-Devonian genera by decade in the second half of the 20th century (in percent of total number of genera erected by decade): monospecific genera (A), genera including 2 to 5 species (B) and genera including more than 5 species (C). Note the contrast between monospecific genera and those with more than 5 species and the gap between "prolific" (dark blue) and "normal" (light blue) authors.
To illustrate this point, five authors, representing different periods mentioned above, were chosen for a case study: for the 19th century, for the first half of the 20th century, , and for the modern period. was also selected as the absolute record holder of productivity for Silurian-Devonian brachiopods, with the erection of 200 genera (197 as first author). In , multi-specific genera largely dominate (54 genera with 10 species or more) against only two monospecific genera (Fig. 7A ![]() ). In the different categories are roughly in balance (27 genera including 1-3 species against 15 genera with 10 species or more). With we enter a new period where things are reversed with a dominance of monospecific genera (65 monospecific genera, 71 genera with 2 or 3 species and only 18 genera with 10 species or more). This is also true for the other two authors: 49 genera with three or fewer species, 6 genera with 10 or more species for and 3 genera with 63 species or less, 5 genera with 10 species or more for . It may be noted that the profile of is rather similar to that of , if we disregard the difference concerning monospecific genera. We can also notice that the profile of is almost exactly opposite to that of , illustrating the contrast between the 19th century and the modern period. These trends are even more apparent if we consider the data in terms of percentages of new genera erected by each author (Fig. 7B
). In the different categories are roughly in balance (27 genera including 1-3 species against 15 genera with 10 species or more). With we enter a new period where things are reversed with a dominance of monospecific genera (65 monospecific genera, 71 genera with 2 or 3 species and only 18 genera with 10 species or more). This is also true for the other two authors: 49 genera with three or fewer species, 6 genera with 10 or more species for and 3 genera with 63 species or less, 5 genera with 10 species or more for . It may be noted that the profile of is rather similar to that of , if we disregard the difference concerning monospecific genera. We can also notice that the profile of is almost exactly opposite to that of , illustrating the contrast between the 19th century and the modern period. These trends are even more apparent if we consider the data in terms of percentages of new genera erected by each author (Fig. 7B ![]() ).
).
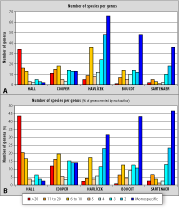
Click on thumbnail to enlarge the image.
Figure 7: A case study applied to five representative authors chosen among the "prolific" tribe. Histograms for each author represent the absolute number of genera including 1, 2, 3 species, etc. (A) and their percentage in the production of each author (B). Note the contrast between an author of the 19th century () and a modern one ().
If we generalize this study to the entire community of authors who have erected Silurian-Devonian genera (from the 19th century until today), we get very similar results (Fig. 8 ![]() ). The opposite evolution between monospecific genera and genera with 10 species or more is clear on this diagram. The genera with few species (mainly monospecific genera) show a dramatic increase while the proportion of genera with several species declined steadily. Historically, the number of genera with more than 10 species has regularly decreased since the 19th century. Genera containing from 5 to 10 species started to decline severely from the 1950s. Then, during the 1970s, it is genera comprising 2 to 4 species which have begun to decline. In the 1970s, genera with more than 20 species virtually disappeared. A sort of balance between the different categories was achieved in the 1950s, typified by the profile of (see above). In the absence of any change in the current methodology, a virtual extension in the future of these curves shows that the almost complete disappearance of the multispecific genera would occur around 2030. Note that although the number of new genera tends to decrease since the 1980s, resulting in a de facto reduction in the number of monospecific genera, the latter decreases considerably less rapidly (Fig. 1
). The opposite evolution between monospecific genera and genera with 10 species or more is clear on this diagram. The genera with few species (mainly monospecific genera) show a dramatic increase while the proportion of genera with several species declined steadily. Historically, the number of genera with more than 10 species has regularly decreased since the 19th century. Genera containing from 5 to 10 species started to decline severely from the 1950s. Then, during the 1970s, it is genera comprising 2 to 4 species which have begun to decline. In the 1970s, genera with more than 20 species virtually disappeared. A sort of balance between the different categories was achieved in the 1950s, typified by the profile of (see above). In the absence of any change in the current methodology, a virtual extension in the future of these curves shows that the almost complete disappearance of the multispecific genera would occur around 2030. Note that although the number of new genera tends to decrease since the 1980s, resulting in a de facto reduction in the number of monospecific genera, the latter decreases considerably less rapidly (Fig. 1 ![]() ) so that their proportion continues to rise.
) so that their proportion continues to rise.
Click on thumbnail to enlarge the image.
Figure 8: A generalization of the previous figure to the entire community of authors active since the 19th century showing the opposite evolution of monospecific and multispecific Silurian-Devonian brachiopod genera. The percentage is calculated on the total number of genera erected during each decade.
What might be the consequences of an uncontrolled increase in the number of monospecific genera? Two types of consequences are clear: for systematics and for palaeogeographical reconstructions.
For systematics, if this trend is confirmed, it could lead to an untenable and ludicrous situation in which each genus would include a single species. What would be left would be a redundancy at the taxonomic level since the status of the genus would gradually become that of the species as it is understood today. We can already feel the effects of this trend in some groups. This would lead to a loss of interest in binomial nomenclature: to maintain its usefulness, all taxonomic levels should be incremented by one degree in the future. Somewhat caricaturing the situation, we could say that it is a return to the 19th (not to mention the 18th) century! A possible, if not likely, explanation for this situation is the gradual shift of the criteria used for defining genera. These criteria are more and more restrictive: there is a shift in the hierarchy of the characters used for generic diagnosis and many of them should remain restricted to the species level. By doing so, we reduce more and more the definition of the genera (i.e., their margins of variation). This is a distortion of the spirit that governs (or should we say "governed"?) the basis of the Linnaean classification. In the latter, the species constitutes the basic unit of the classification and the genus is a taxon used to gather species for the classification or any other purpose. This is becoming less and less true among the brachiopods. The question here is not to condemn the technological advances that reveal, more clearly than in the past, detailed morphological features useful for understanding evolution. But one should avoid being overwhelmed by this influx of new discoveries by returning them to their respective places in order to maintain an efficient working tool.
With regard to palaeogeographical reconstructions, it is clear that monospecific genera are almost necessarily endemic. Therefore these genera are useless for comparisons and correlations and the multiplication of monospecific genera may become a serious obstacle to palaeogeographical studies. Incidentally, this is perhaps an indication of the malfunction of systematic methods. Of course, palaeogeography has nothing to do with systematic classification, but must instead adapt to changes in the latter. Nonetheless we should keep in mind that an excess of monospecific genera makes this adaptation impossible or at least unlikely. This would require that the palaeogeographical analyzes become based on the Family taxon, with the results we know. One might argue that the bursting of "classic" genera is the evidence that the brachiopods as a whole are endemic: experience shows that this is wrong (although this depends on the period and many other elements) and their relative success in dating and correlation, particularly in the Paleozoic, shows that this is not the case.
The progress of knowledge and more detailed analytical tools necessarily lead to a better understanding, but any blind excess and indiscriminate use of data may be hazardous. It seems that our approach suffers from a systematic methodological problem that can lead us to dead ends in the short or medium term in the future. The message that may be retained from this is that authors (especially the younger generation) should keep in mind the following:
It is clear that this inflation of monospecific genera is not exclusive to Silurian-Devonian brachiopods and other groups and other periods are concerned, revealing a general trend. Finally, and without wanting to offend anyone, the previous recommendations should apply to the entire editorial process, from the author to the editor, not to mention the experts examining the calls for projects, the latter being often reluctant to accept a proposal which does not include the word "new", a supposedly sign of excellence, leaving the question open: progress or regression?
The author is grateful to Prof. S.F. , University of West Indies at Mona, Jamaica, for his linguistic support that helped to improve the original manuscript and C.C. (Marseille) for his useful comments.
Index to Organic Names (ION) (last accessed Dec 17, 2012). Online at: http://organismnames.com (edited by Thomson Reuters, also including the Zoological Records database).
W.D.L., H.G., C., O., A., F.C. & P.K. (1999).- International Code of Zoological Nomenclature, 4th ed., International Trust for Zoological Nomenclature, 306 p.
C.C. (last accessed Dec 17, 2012).- Brachiopoda in the Antarctic Benthos. Online at: http://paleopolis.rediris.es/BrachNet/Antarctic-brachiopoda
R.L. (Ed.) (2000).- Treatise on Invertebrate Paleontology. Part H (Revised), vol. 2-3.- Geological Society of America & Paleontological Institute, Boulder, Colorado & Lawrence, Kansas. XXX + 919 p.
R.L. (Ed.) (2002).- Treatise on Invertebrate Paleontology. Part H (Revised), vol. 4.- Geological Society of America & Paleontological Institute, Boulder, Colorado & Lawrence, Kansas. XXXIX + 768 p.
R.L. (Ed.) (2006).- Treatise on Invertebrate Paleontology. Part H (Revised), vol. 5.- Geological Society of America & Paleontological Institute, Boulder, Colorado & Lawrence, Kansas. XLVI + 631 p.
P.A. (Ed.) (2007).- Treatise on Invertebrate Paleontology. Part H (Revised), vol. 6.- Geological Society of America & Paleontological Institute, Boulder, Colorado & Lawrence, Kansas. L + 905 p.
J.J. (1978).- A kinetic model of Phanerozoic taxonomic diversity. I. Analysis of marine orders.- Paleobiology, vol. 4 (n°3), p. 223-251.
ZipcodeZoo (last accessed Dec 17, 2012). Online at: http://zipcodezoo.com