![]()
Contents
[1. Introduction] [2. Lithology and stratigraphy]
[3. Distribution of belemnites in the ABSS] [4. Comparison to more facies]
[5. Comparison to other areas] [6. Systematic descriptions]
[7. Belemnite zonation for the latest Hauterivian to earliest Late Barremian]
[8. Conclusion] [Bibliographic references]
and ... [Plates]
Geertekerkhof 14bis, 3511XC Utrecht (The Netherlands)
5b, rue de Camargue, La Cigalière, 0500 Gap (France)
Doornburg 56, 1081JX Amsterdam (The Netherlands)
Manuscript online since December 10, 2012
[Editor: Michel ; copy editor: Christian C. ; language editor: John ]
![]()
This paper deals with the distribution of belemnites in the latest Hauterivian to early Bedoulian of the Angles Barremian Stratotype Section (ABSS). The distribution of the belemnites in the ABSS is not uniform, mainly due to the inaccessibility of certain beds. To cover this, and to compare the distribution with more proximal settings, we investigated a section to the north of Le Bourguet. The latest Hauterivian sediments mainly yield Hibolithes ex gr. subfusiformis besides some Duvalia ex gr. dilatata. The earliest Barremian sediments deliver a richer association that yields the last Hibolithes spp. The first typical Barremian belemnites occur just one bed above the Hauterivian-Barremian boundary based on ammonites. This belemnite association (BaBA1) consists of Duvalia ex gr. silesiaca-gagrica, Duvalia pontica and several species of Hibolithes. At the boundary between the Nicklesia pulchella and the Kotetishvilia compressissima zones the diversity increases and the first classical Barremian belemnites occur. These were formerly attributed to Mesohibolites (BaBA2). These species are herein attributed to a new genus Shvetsovia. Together with the duvaliids from BaBA1 they were first described from Abkhasia by (1913). The latest Early Barremian (BaBA3) and the earliest Late Barremian (BaBA4) show well diversified belemnite associations, with many classical species, dominated by few genera closely resembling the true Mesohibolites. Eventually, these belemnite associations are compared to more proximal sections within the Vocontian Basin, and areas outside the Vocontian Basin (chiefly Hungary and Georgia). Some differences in the frequency and abundance of several species in these different palaeogeographical settings are believed to indicate differences in natural habitat. Duvalia ex gr. grasiana appears to be more abundant in more distal sections, while juvenile Mesohibolitidae, Conohibolites and Curtohibolites appear to be more abundant in the more proximal environments. Finally, a biozonation is presented and defined based on the distribution of the belemnites in the ABSS. This biozonation appears applicable in the more proximal sections, although some biozones are diachronous. The first Mesohibolites occur in the Upper Barremian Imerites giraudi Zone. In the Barremian-Bedoulian boundary sediments, as defined in the ABSS, Neohibolites first occurs, but the latter is only dominant in the marly sediments above the "calcareous Bedoulian". In the Late Barremian-early Bedoulian seven main belemnite associations can be distinguished, viz. BaBA5, BaBA6, BaBA7, BdBA1, BdBA2, BdBA3 and BdBA4. The following new species and genera are described: Hibolithes keleptrishvilii sp. nov. (latest Hauterivian), Duvalia vermeuleni sp. nov. (Early Barremian), Curtohibolites (?) bourguetensis sp. nov. (Early Barremian), and Shvetsovia gen. nov. (late Early-early Late Barremian). Besides, the Late Barremian yields the new species Mesohibolites anglesensis. Moreover, eleven species are described in open-nomenclature.
Angles; Hauterivian; Barremian stratotype; Bedoulian; Aptian; Belemnites; Bourguet; stratigraphy.
N.M.M., A. & W. (2012).- Mediterranean Neocomian belemnites, part 4: belemnites of the Barremian stratotype section.- Carnets de Géologie [Notebooks on Geology], Brest, Memoir 2012/02 (CG2012_M02), p. 201-274.
Bélemnites néocomiennes des régions méditerranéennes, 4�me partie: bélemnites de la coupe stratotypique du Barrémien.- Cette étude aborde la distribution des bélemnites de l'Hauterivien terminal au B�doulien inférieur de la Coupe Stratotypique du Barrémien d'Angles (ABSS). La distribution des bélemnites sur l'ABSS n'est pas uniforme, principalement due à l'inaccessibilité de certains bancs. Pour y rem�dier et pouvoir comparer leur distribution avec celle qui est observ�e en des domaines moins profonds, nous avons étudié d'autres localités comme les environs du Bourguet. Les sédiments de l'Hauterivien terminal ont fourni principalement Hibolithes ex gr. subfusiformis et quelques Duvalia ex gr. dilatata. Les sédiments du Barrémien basal ont livré une association plus riche comportant les dernières Hibolithes spp. Les premières bélemnites barrémiennes apparaissent juste un banc au dessus de la limite Hauterivien-Barrémien fondée sur les ammonites. Cette association de bélemnites (BaBA1) se compose de Duvalia ex gr. silesiaca-gagrica, Duvalia pontica et de plusieurs espèces d'Hibolithes. À la limite des zones à Nicklesia pulchella et Kotetishvilia compressissima, la diversité augmente et apparaissent alors les bélemnites "classiques" du Barrémien. Anciennement attribuées au genre Mesohibolites (BaBA2), ces espèces sont à présent rattachées au nouveau genre Shvetsovia. En association avec les duvaliid�s de BaBA1 elles ont été initialement décrites dans l'Abkhazie par (1913). Le Barrémien inférieur somminal (BaBA3) et la partie basale du Barrémien supérieur (BaBA4) montrent des associations de bélemnites diversifiées, avec de nombreuses espèces classiques. En particulier les niveaux du Barrémien inférieur terminal et du Barrémien supérieur basal montrent une association riche et diversifiée, dominée par quelques genres ressemblant étroitement aux vrais Mesohibolites. Ces associations de bélemnites sont ensuite comparées à celles de plusieurs coupes plus proximales dans le Bassin vocontien et à des zones situées en dehors du Bassin vocontien (principalement la Hongrie et la Géorgie). Certaines différences dans la fréquence et l'abondance de plusieurs espèces dans ces diverses coupes sont censées refl�ter des différences dans leur habitat naturel. Duvalia ex gr. grasiana semble être plus abondante dans les coupes les plus distales, tandis que les jeunes Mesohibolitidae, Conohibolites et Curtohibolites semblent être plus abondants dans les environnements plus proximaux. Les premiers Mesohibolites ont �t� récoltés dans le Barrémien supérieur (zone à Imerites giraudi) et les derniers dans le partie sommitale du "calcaire Bédoulien". Les premiers Neohibolites ont �t� récoltés dans la "zone non caractérisée" (Barrémien-B�doulien) de l'ABSS, mais ce genre est plus fréquent dans les dépôts du "Bédoulien marneux". Les dépôts du Barrémien supérieur-B�doulien inférieur se composent de sept associations de bélemnites, viz. BaBA5, BaBA6, BaBA7, BdBA1, BdBA2, BdBA3 et BdBA4. Pour conclure, une biozonation est présentée, définie par la distribution des bélemnites dans l'ABSS. Cette biozonation semble applicable dans les coupes les plus proximales, bien que certaines biozones puissent �tre diachrones. Les nouvelles espèces et les nouveaux genres suivants sont décrits: Hibolithes keleptrishvilii sp. nov. (Hauterivien terminal), Duvalia vermeuleni sp. nov. (Barrémien inférieur), Curtohibolites (?) bourguetensis sp. nov. (Barrémien inférieur), Shvetsovia gen. nov. (Barrémien) et Mesohibolites anglesensis sp. nov. (Barrémien supérieur). Enfin onze taxons ont été laissés en nomenclature ouverte.
Angles ; Hauterivien ; Barrémien stratotype ; B�doulien ; Aptien ; bélemnites ; Bourguet ; stratigraphie.
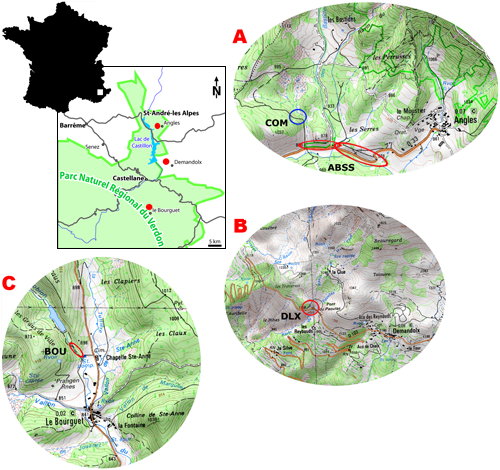
We present information about the distribution of belemnites in the Angles Barremian Stratotype section (ABSS). This section is exposed along the road to the hamlet of Angles,
Alpes-de-Haute-Provence (Fig. 1 ![]() ), and encompasses latest
Hauterivian to
early Bedoulian sediments. The section starts with beds that are exposed to the right of a small road. Depending on the conditions the first two beds are often covered by loose sediments (Pl. 18
), and encompasses latest
Hauterivian to
early Bedoulian sediments. The section starts with beds that are exposed to the right of a small road. Depending on the conditions the first two beds are often covered by loose sediments (Pl. 18 ![]() , fig. A). To the left of this road, few beds are visible, but these are generally not well exposed.
, fig. A). To the left of this road, few beds are visible, but these are generally not well exposed.
The exposures along the road to Angles are selected as a stratotype section for the Barremian (,
1965, p. 103; 1967). Former biostratigraphic studies, chiefly ammonites (,
1980a, 1996, 1998a,
1998b,
2002, 2005; ,
1995, 1998; et al.,
2008), give us the opportunity to present our data in a well-established stratigraphic framework. We will deal with the latest Hauterivian to
Bedoulian belemnites from the section. Furthermore, supplementary information regarding the distribution of these belemnites was obtained by comparing with other sections. Additional stratigraphical information came from the so-called "coupe complémentaire (ou coupe C)" (,
1965, p. 105, fig. 2). The latter section will be named by its acronym COM throughout the text, as it is often
referred to as the Combe Lambert section (, 1995,
1998; et al.,
2009). Actually this area is known as "Les Hermas" on the cadastral maps. Additional information about the palaeogeographical distribution of the Early Barremian belemnites came from a more proximal section near Le Bourguet,
Var (Fig. 1 ![]() ; to the south of Castellane), and
in between these sections, section DLX (Fig. 1
; to the south of Castellane), and
in between these sections, section DLX (Fig. 1 ![]() ; to the west of the hamlet of Demandolx).
; to the west of the hamlet of Demandolx).
Click on thumbnail to enlarge the
image
[Cliquer sur la miniature pour agrandir l'image]
Figure 1: Geographical position of the sections investigated in the surroundings of Castellane.
A) The Barremian Stratotype section, ABSS, is indicated by red ovals (Coordinates: 06°32'32.6"E
- 43°56'28.8"N to 06°33'02"E - 43°56'24.8"N) and
the section COM indicated by a blue circle (Coordinates: 06°32'33"E - 43°56'39"N).
The
Barremian-Bedoulian boundary is positioned roughly just ahead of the bend in the road.
B) Section DLX (circled) to the west of the hamlet of Demandolx (Coordinates:
06°33'59"E - 43°52'24"N). C) The section BOU to the west of chapelle Ste-Anne, north of Le Bourguet,
Var (Coordinates: 06°31'02"E - 43°47'20"N).
� www.geoportail.fr
& IGN - Institut G�ographique National, 73 avenue de Paris, F-94165 Saint-Mand�
Cedex (France)
Except for the unpublished work of (2000), papers that relate to the distribution of belemnites in the Barremian of the western Mediterranean Tethys are absent. However, some papers deal with single finds or faunal lists (, 1896, 1913; , 1989; , 2000). Guided by the fact that we collected over 700 belemnites from the ABSS, and are able to put them into a suitable stratigraphic framework, this work contributes towards a better understanding of the stratigraphic distribution of the Barremian and the early Bedoulian belemnites. Moreover, as we are able to compare this distribution pattern with other regions, a first attempt at a proper biostratigraphic zonation based on belemnites seems possible.
In the exposures along the road to the hamlet of Angles calcareous sedimentary rocks prevail, though certain intervals show dark coloured marls, sometimes with thin, undulating, very marly calcareous intercalations. Due to tectonically induced pressure, parts of the marly beds are diminished, as evidenced by tectonically disturbed and partially dissolved belemnites. Owing to minor faults, and some slopes consisting of fallen-down rock waste, minor parts of the Angles section were and still are less well exposed (see ,
1965, p. 110-111; ,
1980a; & ,
1986). Especially the beds of latest Hauterivian age are momentarily affected by rock waste. However, the lithological development of the missing parts in the section herein depicted can be seen in
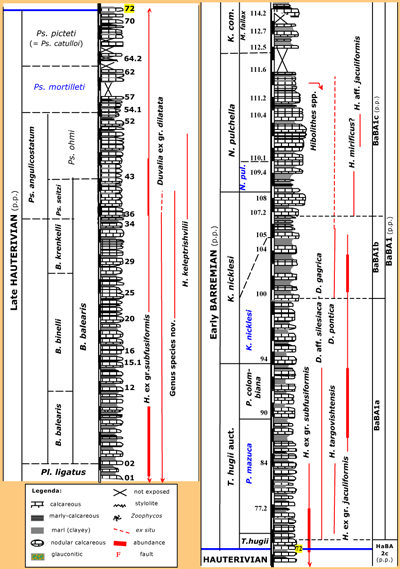
(2005, p. 31 and 38). The latest Hauterivian to earliest Barremian part of the section (Fig. 2 ![]() ) is characterized by more or less
regular alternating calcareous and marly beds. At the moment the Late Hauterivian ammonite zonation is still debatable and several different proposals exist. For example, compare the papers of
(1995),
(2002, 2004, 2005), and et al.
(2003).
) is characterized by more or less
regular alternating calcareous and marly beds. At the moment the Late Hauterivian ammonite zonation is still debatable and several different proposals exist. For example, compare the papers of
(1995),
(2002, 2004, 2005), and et al.
(2003).
The Early Barremian Nicklesia pulchella and Kotetishvilia compressissima
zones (Fig. 2 ![]() ; Pl. 18
; Pl. 18 ![]() , fig. B) mainly yield rather thick calcareous beds, but sometimes more marly levels occur. Currently, the base of the Barremian is set at ANG072, while the base of the Upper Barremian is documented by the first occurrence of Heinzia sayni (ANG147.2; Pl. 19
, fig. B) mainly yield rather thick calcareous beds, but sometimes more marly levels occur. Currently, the base of the Barremian is set at ANG072, while the base of the Upper Barremian is documented by the first occurrence of Heinzia sayni (ANG147.2; Pl. 19 ![]() , fig. A; cf. ,
2002, p. 45) (Fig. 3
, fig. A; cf. ,
2002, p. 45) (Fig. 3 ![]() ). Conditionally, based on lithological grounds, this boundary was placed at ANG137 (,
1965, p. 110-111), because the interval between the beds ANG137 to ANG152 was relatively devoid of ammonites. Still some parts of the studied interval, notably the
Barremian-Bedoulian boundary interval, are relative devoid of characteristic ammonites. Also, belemnites appear to be distributed unevenly. Especially, the earliest Barremian, up to and including the base of the K. compressissima Zone yielded few belemnites only. In the late Early Barremian (Coronites darsi Zone) thick calcareous beds occur, alternated with relatively thick marly beds. In these marly sets, thin
irregular marly to calcareous beds sometimes occur. The latter, especially the thinnest beds, often become visible as whitish, weathered layers with a peculiar powder-like appearance. From the top of the C. darsi Zone on, the lithology is dominated by calcareous beds, often separated by very small marly levels that are sometimes barely visible, while a thicker marl level only exists very sporadically. Herein, the Holcodiscus uhligi Zone sensu
(2002) marks the top of the Lower Barremian (Pl. 19
). Conditionally, based on lithological grounds, this boundary was placed at ANG137 (,
1965, p. 110-111), because the interval between the beds ANG137 to ANG152 was relatively devoid of ammonites. Still some parts of the studied interval, notably the
Barremian-Bedoulian boundary interval, are relative devoid of characteristic ammonites. Also, belemnites appear to be distributed unevenly. Especially, the earliest Barremian, up to and including the base of the K. compressissima Zone yielded few belemnites only. In the late Early Barremian (Coronites darsi Zone) thick calcareous beds occur, alternated with relatively thick marly beds. In these marly sets, thin
irregular marly to calcareous beds sometimes occur. The latter, especially the thinnest beds, often become visible as whitish, weathered layers with a peculiar powder-like appearance. From the top of the C. darsi Zone on, the lithology is dominated by calcareous beds, often separated by very small marly levels that are sometimes barely visible, while a thicker marl level only exists very sporadically. Herein, the Holcodiscus uhligi Zone sensu
(2002) marks the top of the Lower Barremian (Pl. 19 ![]() , fig. A). It is not untill the Imerites giraudi Zone that marly sediments become again more pronounced but calcareous sediments remain the dominant lithology (Fig. 3
, fig. A). It is not untill the Imerites giraudi Zone that marly sediments become again more pronounced but calcareous sediments remain the dominant lithology (Fig. 3 ![]() ). It should be noted that the name-bearing ammonite of the I. giraudi Zone was challenged by &
(2009, p. 96) and &
(2010a, p. 19, 22) in favour of Imerites cristatus (). However, eventually they concluded, guided by the acceptation these species being distinct, that only a small adjustment of the base of the name-bearing ammonite zone would be necessary ( & ,
2010b, p. 107).
). It should be noted that the name-bearing ammonite of the I. giraudi Zone was challenged by &
(2009, p. 96) and &
(2010a, p. 19, 22) in favour of Imerites cristatus (). However, eventually they concluded, guided by the acceptation these species being distinct, that only a small adjustment of the base of the name-bearing ammonite zone would be necessary ( & ,
2010b, p. 107).
Along the road to Angles, the upper part of the section, i.e., the
"calcareous Bedoulian" is disturbed by a minor fault (between the beds ANG216 and ANG223; Pl. 4 ![]() , fig. A) while the topmost part is covered by vegetation. Contrary, these beds are quite well exposed in the section COM (,
1965, p. 105; , 1995,
Pl. 10; 1998, p. 218). The last bed of the "calcareous Bedoulian" is thought to be a hardground (,
1965, p. 108; , 1967), but that could not be confirmed in the field
, fig. A) while the topmost part is covered by vegetation. Contrary, these beds are quite well exposed in the section COM (,
1965, p. 105; , 1995,
Pl. 10; 1998, p. 218). The last bed of the "calcareous Bedoulian" is thought to be a hardground (,
1965, p. 108; , 1967), but that could not be confirmed in the field
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 2: The stratigraphical distribution of belemnites in the latest Hauterivian to earliest Barremian (ANG001 to ANG114.3). See (2005, p. 31 and 38) for non-exposed parts. Scale bar graduations = 1 m. Ammonite zones or subzones in blue are not (yet) accepted in et al. (2011). Abbreviations: B. = Balearites, D. = Duvalia, H. = Hibolithes, N. pul. = Nicklesia pulchella, K. = Kotetishvilia, K. com. = Kotetishvilia compressissima, M. = Metahoplites, P. = Psilotissotia, p.p. = pro parte, Pl. = Plesiospitidiscus, Ps. = Pseudothurmannia, and T. = Taveraidiscus.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
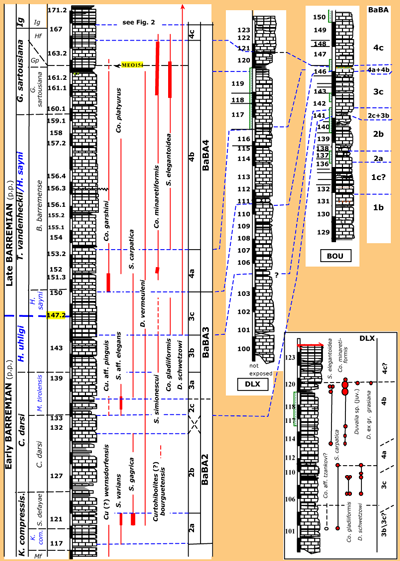
Figure 3: The late
Early to early Late Barremian (ANG115 to ANG171.3) in the ABSS. From left to right are indicated:
(sub-) stages, ammonite zones, ammonite subzones, bed-numbers, lithology (for
legend explanation see Fig. 2 ![]() ), the FO and LO of the belemnite species, and the belemnite associations. Scale bar graduations = 1 m [note difference in seize between distal (ABSS) and proximal sections (DLX, BOU - see Fig. 5
), the FO and LO of the belemnite species, and the belemnite associations. Scale bar graduations = 1 m [note difference in seize between distal (ABSS) and proximal sections (DLX, BOU - see Fig. 5 ![]() for
details)]. Ammonite zones and/or subzones in blue are not (yet) accepted in the latest publication of the KWG ( et al.,
2011). Abbreviations: Co. = Conohibolites, K. com. = Kotetishvilia compressissima, Cu. = Curtohibolites, D. = Duvalia, Gp = Gerhardtia provincialis, Hf = Hemihoplites feraudianus, Ig = Imerites giraudi, MEO = Méouilles, K. com. and K. compressis. = Kotetishvilia compressissima, Mf = Metahoplites fallauxi, and S. = Shvetsovia.
for
details)]. Ammonite zones and/or subzones in blue are not (yet) accepted in the latest publication of the KWG ( et al.,
2011). Abbreviations: Co. = Conohibolites, K. com. = Kotetishvilia compressissima, Cu. = Curtohibolites, D. = Duvalia, Gp = Gerhardtia provincialis, Hf = Hemihoplites feraudianus, Ig = Imerites giraudi, MEO = Méouilles, K. com. and K. compressis. = Kotetishvilia compressissima, Mf = Metahoplites fallauxi, and S. = Shvetsovia.
As stated before, it is in the I. giraudi Zone that marly sediments become more pronounced. However, part of the apparent absence of readily visible marly levels might have been caused by tectonic pressure.
Especially, the marly beds between ANG165.1 and ANG169.2 show signs of shearing. In the early Late Barremian a compressional fault system in the top of the He. sayni Zone is accompanied by some stylolites. It causes some fluctuations in the thickness of individual beds. This system is well visible along the road around the indication of
hectometer 6 (Pls. 22-23). However, the absence of more marly levels might better be explained by a certain amount of condensation or bypassing. Some beds, notably the top of bed ANG169.1, show abundant corroded belemnites that could suggest some amount of condensation. Also, in the Gerhardtia sartousiana Zone (top of ANG161.2; Fig. 3 ![]() ) and in the marls of
Bedoulian age (on top of COM137; Fig. 4
) and in the marls of
Bedoulian age (on top of COM137; Fig. 4 ![]() ), levels with pitted and worn belemnites occur. Moreover, the beds ANG161.2 top, ANG161.3/4-162 (Pl. 20-23
), levels with pitted and worn belemnites occur. Moreover, the beds ANG161.2 top, ANG161.3/4-162 (Pl. 20-23 ![]() ), ANG171.2-172, and the top of bed ANG179.2 delivered some sponges (?
Elasmostoma) attached to ammonites. Besides, the beds ANG161.2 (top part), ANG162 (,
2002, p. 47), ANG165.1, ANG165.2, ANG167, and ANG170.1 (G. sartousiana Zone to the base of the I. giraudi Zone) do show traces of glauconite, confined to the surface of ammonite-remains. However, et al.
(2002) most probably overrated the amount of time which is presented by these, most probably local, condensations (see also et al.,
2009, p. 1257-1259).
), ANG171.2-172, and the top of bed ANG179.2 delivered some sponges (?
Elasmostoma) attached to ammonites. Besides, the beds ANG161.2 (top part), ANG162 (,
2002, p. 47), ANG165.1, ANG165.2, ANG167, and ANG170.1 (G. sartousiana Zone to the base of the I. giraudi Zone) do show traces of glauconite, confined to the surface of ammonite-remains. However, et al.
(2002) most probably overrated the amount of time which is presented by these, most probably local, condensations (see also et al.,
2009, p. 1257-1259).
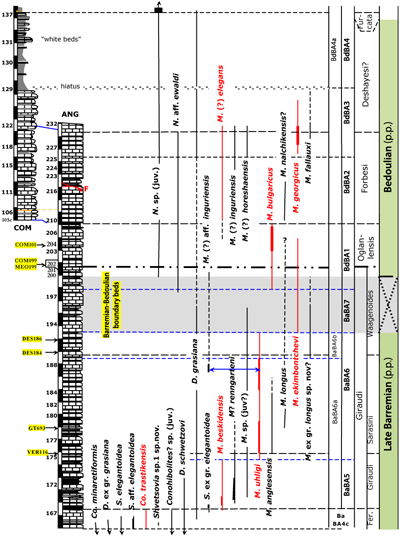
Fig. 4 ![]() depicts the Late
Barremian-early Bedoulian of the ABSS. The ammonite zonation is adopted and modified after
(1995, 1998) and
(2005). These modifications are based on bed-by-bed correlations with various sections in the neighbourhood. The data are extracted from the work of
(1995, 1998), viz. the sections of Descouère (DES), Grande-Terre (GT), Méouilles (MEO), and Moriez (MOR). Especially, the Hemihoplites feraudianus Subzone appears to be ill represented in the ABSS (Fig. 3
depicts the Late
Barremian-early Bedoulian of the ABSS. The ammonite zonation is adopted and modified after
(1995, 1998) and
(2005). These modifications are based on bed-by-bed correlations with various sections in the neighbourhood. The data are extracted from the work of
(1995, 1998), viz. the sections of Descouère (DES), Grande-Terre (GT), Méouilles (MEO), and Moriez (MOR). Especially, the Hemihoplites feraudianus Subzone appears to be ill represented in the ABSS (Fig. 3 ![]() ), but see et al.,
2009. Herein, ammonites occur (i.e., Spinocrioceras) that are known from the lower Parancyloceras bidentatum Zone of Germany (,
1973; & ,
1995; & ,
2009). The H. feraudianus Subzone marks the transition from the overall calcareous latest Early to early Late Barremian, to the late Late Barremian with more marked marly beds.
), but see et al.,
2009. Herein, ammonites occur (i.e., Spinocrioceras) that are known from the lower Parancyloceras bidentatum Zone of Germany (,
1973; & ,
1995; & ,
2009). The H. feraudianus Subzone marks the transition from the overall calcareous latest Early to early Late Barremian, to the late Late Barremian with more marked marly beds.
In the upper part of the Barremian of the ABSS, the top of the Martelites sarasini Subzone, age-indicative ammonites are rare or absent. However, the close Descouère section (,
1998, p. 242: beds DES184 and DES186) yielded few correlative ammonites, notably Pseudocrioceras and some Kutatisites. This interval correlates to the beds ANG189 to ANG191. Particularly, in the area of the Bedoulian stratotype, a comparable ammonite association ( et al.,
2000; et al.,
2000, 2006), is known from the top of the former M. sarasini Zone (now a Subzone; cf. et al.,
2011). Its upper limit reaches just above the " Horizon" (sensu et al.,
2000). In the Bedoulian type-area near Marseille, et al.
(1997) placed the
Barremian-Bedoulian boundary at the first occurrence of Deshayesites (see et al.,
2000). In the ABSS the first ammonite that characterizes the
Bedoulian is apparently found in bed ANG200 [cf. & ,
1995, p. 107 - Prodeshayesites
? sp. ind. (= deshayesitidae)]. The thick beds below this level and in the Combe Lambert section have not yielded any significant ammonites to date. This interval is known as the "non-characteristic interval", and ever since
(1965) the base of this level (bed ANG197) is used as the boundary between the Barremian and
Bedoulian (Pl. 4 ![]() , fig. B).
, fig. B).
et al. (2006) concluded that only few species of dinoflagellates appeared isochronous between the ABSS (Tethyan Realm) and the Austral Realm. Among those species Pseudoceratium retusum var. securigerum first occur in bed ANG199 of the ABSS. Also, the Boreal-Atlantic Prodeshayesites fissicostatus Zone yields this species. This suggests it as a good indicator of strata of early Bedoulian age. Recently, & (2010, p. 30) placed the Barremian-Bedoulian boundary even lower, i.e., at bed ANG192 without further explanation.
The index ammonite for the early Bedoulian, Paradeshayesites oglanlensis (), is not known from the ABSS. It apparently does occur in the nearby Méouilles section in bed MEO199 [see ,
1998 (=
, 1995,
Pl. 2, fig. 1a, 1b), p. 231, Pl. 6, fig. 3a, 3b (erroneously indicated from bed
MEO208)]. However, according to &
(2004, p. 214) this species is more similar to Paradeshayesites weissiformis. Nevertheless, this still would indicate the same age. The bed MEO199 correlates with the middle part of bed ANG202 (Fig. 4 ![]() ). In bed ANG210, Deshayesites ex gr. spathi-normanni
occurs: it is an ammonite characteristic for the Deshayesites forbesi Zone (former Deshayesites weissi Zone) (,
1998, p. 221). It appears that in general, index or indicative ammonite species for the
Barremian-Bedoulian boundary and the early Bedoulian interval are absent in the ABSS. This is believed to be the result of palaeogeographical bias, of shallow versus deeper taxa (,
1995; et al., 2006, p. 3-4).
). In bed ANG210, Deshayesites ex gr. spathi-normanni
occurs: it is an ammonite characteristic for the Deshayesites forbesi Zone (former Deshayesites weissi Zone) (,
1998, p. 221). It appears that in general, index or indicative ammonite species for the
Barremian-Bedoulian boundary and the early Bedoulian interval are absent in the ABSS. This is believed to be the result of palaeogeographical bias, of shallow versus deeper taxa (,
1995; et al., 2006, p. 3-4).
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 4: Distribution of belemnites in the latest Late Barremian to
early Bedoulian. See Fig. 2 ![]() for
legend explanation. Abbreviations: Co. = Conohibolites, D. = Duvalia, F = fault, Fer. = Hemihoplites feraudianus, M. = Mesohibolites, N. = Neohibolites, S. = Shvetsovia, ANG = Angles, COM = Combe Lambert, DES = Descoure, GT = Grande Terre, MEO = Méouilles, and VER = Vergons-1 (cf. ,
1995, 1998). Scale bar graduations = 1 m.
for
legend explanation. Abbreviations: Co. = Conohibolites, D. = Duvalia, F = fault, Fer. = Hemihoplites feraudianus, M. = Mesohibolites, N. = Neohibolites, S. = Shvetsovia, ANG = Angles, COM = Combe Lambert, DES = Descoure, GT = Grande Terre, MEO = Méouilles, and VER = Vergons-1 (cf. ,
1995, 1998). Scale bar graduations = 1 m.
Roughly, the Early to early Late Barremian can be divided into four main belemnite associations, i.e., BaBA1, BaBA2, and BaBA3 and BaBA4 (the latter two are partially BaBA2 and BaBA3 sensu & ,
2005). The calcareous Late Barremian to
Bedoulian can be divided into six main belemnite associations: BaBA5, BaBA6, BaBA7, BdBA1, BdBA2, and BdBA3
(Figs. 2 ![]() - 3
- 3 ![]() - 4
- 4 ![]() ).
).
The latest Hauterivian did not deliver abundant belemnites but the general picture is comparable to other areas in and outside the Vocontian Basin. Based on ammonites, the Hauterivian-Barremian boundary is fixed at ANG072 in the ABSS (, 2002). So far, the first genuine Barremian belemnites occur in the marly level above ANG073. This earliest Barremian belemnite association (BaBA1) yields species first described from Gagra (or Gagry) in Abkhazia by (1913) and to some extent recorded from the ABSS by (2000, "faune à Duvalia"). The earliest Barremian, up to and including part of the N. pulchella Zone, yields belemnites that are closely related to Hauterivian taxa. The typical Hauterivian H. ex gr. subfusiformis last occurs in the Taveraidiscus hugii Zone, together with H. targovishtensis. It is succeeded by H. ex gr. jaculiformis. The last occurrence of this species is recorded in the Kotetishvilia nicklesi Zone. However, closely related species occur in the N. pulchella Zone. The latter interval yields ? Hibolithes mirificus , 1965, and strongly flattened fusiform species, either related to H. mirificus, the jaculiformis-group or to Shvetsovia (a new genus, see palaeontological part). Also, in Hungary this interval yielded peculiar, yet rather indefinite, taxa like Curtohibolites bakalovi (, 1965) and H. mirificus. These taxa appear to represent a transitional fauna between the earliest Barremian BaBA1 and BaBA2.
Subsequently, there is a belemnite association (BaBA2) that is also characterized by belemnites first described from Gagra. Most characteristic is the S. varians-gagrica group, accompanied by a new species related to Duvalia grasiana (, 1841) and Curtohibolites (?) wernsdorfensis , 1963. The first elements of BaBA2 occur in ANG115. The beds of the K. compressissima Acme Subzone yield a relative abundant fauna of belemnites dominated by the S. gagrica-varians group. The first genuine Curtohibolites , 1963, occurs in the upper part of the C. darsi Zone (BaBA2).
This association is followed by BaBA3 and BaBA4, or "mid" Barremian association sensu & (2004, 2005). Species of the Conohibolites gladiiformis - Shvetsovia carpatica complex characterize BaBA3. Also, Shvetsovia simionescui and common D. ex gr. grasiana occur in this interval. BaBA4 is packed with a group of belemnites preceding the true Mesohibolites, of which Conohibolites ex gr. minaretiformis (, 1913) and Shvetsovia elegantoidea (, 1965) are the most common in the ABSS. These belemnites appear to be especially abundant in the upper part of the Toxancyloceras vandenheckii Zone (Barrancyloceras barremense Subzone; Pl. 21, fig. B) and disappear in the H. feraudianus Subzone. Also, the last Conohibolites [Co. trastikensis (, 1965)] disappears in the H. feraudianus Subzone. S. elegantoidea is succeeded by Shvetsovia ex gr. elegantoidea (, 1965). The latter disappears in the lower part of the I. giraudi Zone (ANG168-169.1).
The first Mesohibolites [Mesohibolites beskidensis (, 1883)] occurs in the I. giraudi Zone. Species that show intermediate characteristics between Shvetsovia and Mesohibolites also appear here. This species is herein refered to as Mesohibolites (?) renngarteni , 1939. It is quite abundant in the I. giraudi Subzone and disappears in the lower part of the M. sarasini Subzone. In the lower and topmost part of the M. sarasinii Subzone Mesohibolites uhligi (, 1913) becomes abundant, as does a new species of belemnite, viz. "Mesohibolites minaret " sensu in et al., 2002 (= Mesohibolites anglesensis sp. nov., see palaeontological part). In the marly calcareous bed ANG171.1 a small belemnite that is very similar to Neohibolites first occurs. This belemnite is either a juvenile specimen of Mesohibolites, related to Neohibolites, or related to the Georgian "Hibolites" ex gr. bsibiensis , 1938. The latest Barremian is characterized by robust belemnites related to Mesohibolites bulgaricus (, 1965) and Mesohibolites ekimbontchevi , 1965. Duvalia grasiana occurs throughout the I. giraudi Zone. However, this species appears to be absent, or very rare, in the Barremian-Bedoulian boundary beds. It is again relative common in the uppermost part of the "calcareous Bedoulian".
Mesohibolites bulgaricus , 1965, is also characteristic for the lowermost beds of the Bedoulian, where it occurs in abundance. In addition, Mesohibolites georgicus , 1973, Mesohibolites fallauxi (, 1883), Mesohibolites (?) elegans (, 1913), Mesohibolites (?) ex gr. horeshaensis-inguriensis (, 1938), Neohibolites aff. ewaldi (, 1861), and Neohibolites occur in these sediments. Like in Romania ( & , 2006, p. 533), Neohibolites appears earlier as compared to the Bedoulian deshayesitidae. However, it is nearly impossible to separate between juvenile Mesohibolites and Neohibolites, at least for part of the material (see palaeontological part).
Stratigraphical data from the Barremian interval concerning the distribution of ammonites are mainly based on the relative distal basin section of Angles and the platform deposits in the Bedoulian type-area (la Bédoule-Cassis). To the north and northwest of Angles, and to the north of the Montagne de Lure, more inward basin facies are developed in typical subpelagic marl-limestone deposits (, 1989, p. 32-35). Therein, frequent and abundant disturbances in the sedimentary development occur. As a result, the latest Hauterivian, the Barremian and the Bedoulian sedimentation is rather complex in the Vocontian Basin. These large and small-scaled gravity-driven deposits ( & , 1979; , 1989; & , 2003) result in areas with chaotic lithology, and other areas with sedimentary gaps on various scales.
(2000, p. 5-6) depicted a section near Montclus (cf. Palluel section and other sections figured by , 1989, p. 38-41), that show these synsedimentary gliding phenomena ("coulée boueuse") clearly. In the Montclus section, the base of the Barremian is characterized by the "Gagra-fauna", situated above a chaotic marly interval with glauconite, which yields reworked calcareous blocks, some with ammonites, pointing to the Hauterivian-Barremian boundary sediments. Abundant brachiopods, echinoids and belemnites occur in the marly sedimentary rocks. Among these, species characteristic for the "mid" Barremian occur (Curtohibolites, Conohibolites & , 2005, and Shvetsovia gen. nov.). This level is succeeded by a calcareous succession with the ammonites Camereiceras limentinus () and Ezeiceras heberti (det. J. ), followed by an interval where characteristic latest Barremian to early Bedoulian Mesohibolites occur.
Despite being rare in the ABSS, the genus Curtohibolites is rather common in the more proximal palaeogeographical settings, with condensed and glauconitic calcareous and marly beds. Curtohibolites is also known from distal deposits near Montclus, Gard (, 2000), and Serre de Bleyton, Dr�me (, 2010). However, there gravity-flows are frequent ( & , 1979; , 2005; et al., 2010). Therefore, we assume these belemnites most probably to be transported into the deeper parts of the Vocontian Basin.
To the south of Angles, more proximal facies are developed with lesser depositional. Sediments are developed in typical condensed, glauconitic facies. In these successions belemnites may occur abundantly. We investigated some outcrops to the north of Le Bourguet. Here, latest Hauterivian to earliest Late Barremian rocks are well-exposed below an Albian cover of marls and calcareous nodular sandstones. Especially the glauconitic latest Early Barremian sediments often yield abundant belemnites (Fig. 5 ![]() ). We aimed to get information about the distribution of belemnites in the interval which we could not sample well in the
ABSS, the boundary between the N. pulchella and the K. compressissima
zones, and to compare the general distribution of belemnites between the two areas (Fig. 3
). We aimed to get information about the distribution of belemnites in the interval which we could not sample well in the
ABSS, the boundary between the N. pulchella and the K. compressissima
zones, and to compare the general distribution of belemnites between the two areas (Fig. 3 ![]() ).
).
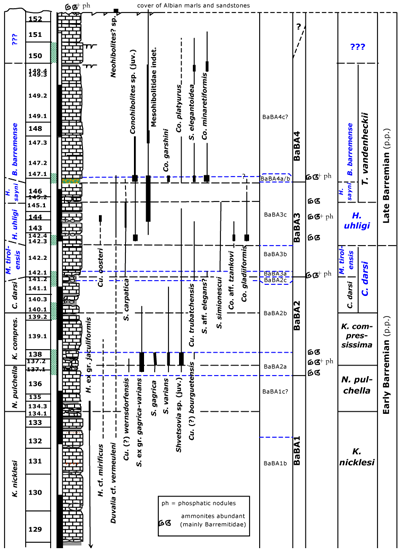
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 5: Distribution of belemnites in the section (BOU) to the west of chapelle Ste-Anne (N of Le Bourguet). Ammonite zonation in black, after
(1980b,
2002, and pers. com.). Ammonite zones in blue, based on the mutual distribution of belemnite species in the ABSS. Scale bar for sections graduations = 1 m. Abbreviations: D. = Duvalia, Co. = Conohibolites, Cu. = Curtohibolites, H. = Hibolithes, S. = Shvetsovia. See Fig. 2 ![]() for
legend explanation.
for
legend explanation.
The section BOU is exposed to the west of the chapelle Sainte-Anne (Fig. 1 ![]() ; 06°31'02"E - 43°47'20"N). Along the
eastern side of a water-reservoir, typical thick bedded Hauterivian sediments that are exposed yielded few H. ex gr. subfusiformis. Some glauconitic beds are intercalated. Eventually, bed thicknesses diminish and more and more beds with glauconite occur. In general, these beds are often delimited by intensively bioturbated horizons (Thalassinoides-burrows). As a result, these levels are irregular and with nodular calcareous remains are abundant. Consequently, the transition to the thin intercalating marl beds is often obscured. Near the top of the section a pronounced glauconitic marly level exists with abundant belemnites. Fossils appear to be reworked in part and phosphatic ammonite remains occur. This bed is captured by a glauconitic calcareous bed with abundant belemnites, and followed by few calcareous beds with glauconitic spots. Eventually, these beds are truncated by Albian marls and sandstones, separated by a reworked level with phosphatic ammonites and rolled belemnites. In some beds rather deep burrows filled with glauconitic sediment, can be noted. Brachiopods, corals, but especially cephalopods are abundant in these deposits. This will partially be the result of condensation but as some species and genera are more respectively less abundant in these more proximal facies as compared to the
ABSS, part of it will reflect natural habitat. In these more proximal facies, juvenile species and belemnites related to the S. gagrica-varians group are especially abundant.
; 06°31'02"E - 43°47'20"N). Along the
eastern side of a water-reservoir, typical thick bedded Hauterivian sediments that are exposed yielded few H. ex gr. subfusiformis. Some glauconitic beds are intercalated. Eventually, bed thicknesses diminish and more and more beds with glauconite occur. In general, these beds are often delimited by intensively bioturbated horizons (Thalassinoides-burrows). As a result, these levels are irregular and with nodular calcareous remains are abundant. Consequently, the transition to the thin intercalating marl beds is often obscured. Near the top of the section a pronounced glauconitic marly level exists with abundant belemnites. Fossils appear to be reworked in part and phosphatic ammonite remains occur. This bed is captured by a glauconitic calcareous bed with abundant belemnites, and followed by few calcareous beds with glauconitic spots. Eventually, these beds are truncated by Albian marls and sandstones, separated by a reworked level with phosphatic ammonites and rolled belemnites. In some beds rather deep burrows filled with glauconitic sediment, can be noted. Brachiopods, corals, but especially cephalopods are abundant in these deposits. This will partially be the result of condensation but as some species and genera are more respectively less abundant in these more proximal facies as compared to the
ABSS, part of it will reflect natural habitat. In these more proximal facies, juvenile species and belemnites related to the S. gagrica-varians group are especially abundant.
In the "Holcodiscus-beds" of an exposure (approx. 05°46'44"E - 44°07'16"N) NW to the classical section at Combe-Petite (Montagne de Lure; & , 1966, p. 416-417), with platform margin deposits, a comparable picture can be observed (abundance of the S. gagrica-varians group). In contrast, in the ABSS this group is clearly less abundant. On the other hand, D. ex gr. grasiana is much more abundant in the deeper facies of the ABSS, but the latter species is very rare in the more proximal facies.
As noted before, due to the near absence of belemnites hitherto in the N. pulchella Zone and in the lower part of the K. compressissima Zone the exact position of the change in the belemnite faunal association is difficult to fix in the ABSS. To better delimit this change, we collected belemnites in palaeogeographically more proximal sections (BOU) and in another to the ENE of Clos (or Clot) de Barral (CBL; 06°38'07.4"E - 43°45'28.8"N). In the latter, Cu. (?) wernsdorfensis associated with H. mirificus occurs in the nodular bed CBL134d (corresponding to bed BOU137.1). This represents the transition between the N. pulchella Zone and the K. compressissima Zone. In BOU the glauconitic top part of the calcareous bed BOU136 yields the first Cu. (?) wernsdorfensis associated with the first specimens of the S. gagrica-varians group. The marly bed CBL134d-135 (= BOU137.1-2) yielded the first specimens of the S. gagrica-varians group, above beds with abundant N. pulchella, and occurring up to CBL140 (= BOU140.1). The latter bed yields, among others, the ammonite Subtorcapella defayae. While the beds in between yielded a rich association of ammonites that includes, among others: Kotetishvilia compressissima, Nicklesia didayana, Holcodiscus caillaudianus and Heinzia communis (cf. , 1980a, 1980b, 1998b, 2002, p. 50).
To the south of the ABSS, at Demandolx (DLX: 06°33'59"E - 43°52'24"N), in an abandoned quarry, the boundary beds between the Early and Late Barremian are exposed (Fig. 3 ![]() ). The top part of the section is disturbed by faults and heavily disturbed, and is succeeded by Early
Bedoulian marls. This section is of interest because of the more proximal position with respect to the ABSS. The relative few belemnites that could be recovered show belemnite associations intermediate between the ABSS and the more proximal facies (BOU, CBL). This is indicated by the occurrence of Co. tzankovi, relative frequent occurrence of D. schwetzowi, and also Co. gladiiformis is relative common.
There and by, the beds DLX116c to DLX119 contained traces of glauconite. However, most abundant appears to be Co. minaretiformis. Apparently, this species was common in all the investigated palaeogeographical settings.
). The top part of the section is disturbed by faults and heavily disturbed, and is succeeded by Early
Bedoulian marls. This section is of interest because of the more proximal position with respect to the ABSS. The relative few belemnites that could be recovered show belemnite associations intermediate between the ABSS and the more proximal facies (BOU, CBL). This is indicated by the occurrence of Co. tzankovi, relative frequent occurrence of D. schwetzowi, and also Co. gladiiformis is relative common.
There and by, the beds DLX116c to DLX119 contained traces of glauconite. However, most abundant appears to be Co. minaretiformis. Apparently, this species was common in all the investigated palaeogeographical settings.
The latest Hauterivian did not deliver abundant belemnites but the general picture is comparable to other areas in and outside the Vocontian Basin. A rather species poor association dominated by H. ex gr. subfusiformis (, 1829), D. ex gr. dilatata (, 1827) and some poorly known, undescribed species. D. ex gr. dilatata is so far not known from any sediments younger than the latest Hauterivian (e.g. Río Argos, Spain; , 1997, p. 7). However, more illustrative from a stratigraphical point of view, is the absence of H. subfusiformis, and the genera Pseudobelus and Pseudoduvalia (HaBA2c sensu , 2009, p. 16). It should be noted that the ranges of H. ex gr. subfusiformis and H. subfusiformis s.s. are mixed up in the scheme presented therein.
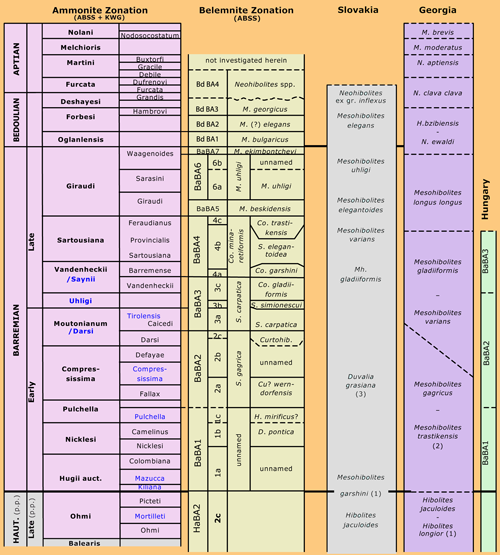
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 6: Comparing belemnite zonations and correlations of ABSS and BOU, with Hungary ( & , 2005), Georgia (, 1998) and Slovakia ( et al., 1994). Abbreviations: Co. = Conohibolithes, Cu. = Curtohibolites, D. = Duvalia, H. = Hibolithes, p.p. = pro parte, and S. = Shvetsovia. (1) = Hibolithes keleptrishvilii sp. nov., (2) = Curtohibolites (?) wernsdorfensis, (3) = Duvalia vermeuleni sp. nov.
The earliest Barremian belemnite fauna described by
(1913) and herein
referred to as the "Gagra-fauna" occurs throughout the Mediterranean Tethys. Most characteristic species (D. pontica and D. gagrica-silesiaca group) occur in the southeast of Spain, both in deep water deposits of the Río Argos, as well in more proximal facies of the Tornajo Mountain (pers. obs.). While the smaller species D. aff. silesiaca appears to be more frequent in the deeper deposits, D. pontica is especially abundant in the more proximal facies. In general, these
duvaliids are accompanied by H. ex gr. jaculiformis, apart from other rarer species. This association is also known from Azerbaijan (,
1972), Bulgaria (,
1964, 1965,
1979), Crimea (,
1966, 1974), Georgia (,
1913; et al.,
2002), Hungaria ( & ,
2004, 2005), and the Western Carpathians ( et al.,
1994). Also, the association dominated by species of the S. gagrica-varians group occurs throughout the same regions. &
(2005, p. 66) showed the boundary between BaBA1 and BaBA2 to roughly coincide with the younger part of the K. compressissima Zone in Hungary (Fig. 6 ![]() ). This could best be concluded from their section B ( & ,
2009), herein D. ex gr. grasiana (=
Duvalia vermeuleni sp.
nov., see systematic description) that first occurs in bed 124 together with abundant Holcodiscus fallax (). Moreover, because Hibolithes
? sp. is at least partially conspecific with Shvetsovia gagrica, this boundary would even be lower in that section. Note that the D. grasiana figured by et al.
(1994) is in our opinion D. vermeuleni sp. nov., the oldest Mesohibolites garshini figured is most probably H. keleptrishvilii sp. nov., and Mesohibolites varians is S. carpatica. In the belemnite zonal scheme of Georgia (,
1998, p. 443), the Mesohibolites trastikensis appears to be our Cu. (?) wernsdorfensis. Thereandby, the M. gargicus - M. trastikensis zone yields Duvalia pontica and D. gagrica (Fig. 6
). This could best be concluded from their section B ( & ,
2009), herein D. ex gr. grasiana (=
Duvalia vermeuleni sp.
nov., see systematic description) that first occurs in bed 124 together with abundant Holcodiscus fallax (). Moreover, because Hibolithes
? sp. is at least partially conspecific with Shvetsovia gagrica, this boundary would even be lower in that section. Note that the D. grasiana figured by et al.
(1994) is in our opinion D. vermeuleni sp. nov., the oldest Mesohibolites garshini figured is most probably H. keleptrishvilii sp. nov., and Mesohibolites varians is S. carpatica. In the belemnite zonal scheme of Georgia (,
1998, p. 443), the Mesohibolites trastikensis appears to be our Cu. (?) wernsdorfensis. Thereandby, the M. gargicus - M. trastikensis zone yields Duvalia pontica and D. gagrica (Fig. 6 ![]() ).
).
Stratigraphical data regarding belemnites of the "mid" Barremian are less well-known in detail in the Caucasus but Shvetsovia-Conohibolites associations do occur in Georgia: , 1939; , 1973; et al., 2002; & , 2005; & , 2005 (Abkhazia), et al., 2002 (Megreliya), & , 1975; et al., 2002 (Kutaisi), and , 2005 (South-Ossetia).
& (2005) described a well-diversified earliest to "mid" Barremian belemnite association from the Gerecse Mountains (Hungary). These belemnites were collected together with abundant ammonites, adding biostratigraphic details to these belemnite associations, which represent a mix of Curtohibolites, Conohibolites, and Shvetsovia species. Despite that belemnite taxa from the Curtohibolites - Shvetsovia and Conohibolites associations are briefly mentioned from Bulgaria (, 1963, 1965, 1970a), Czech Republic (, 1978a: Outer Western Carpathians), Georgia, Hungary, and Slovakia ( et al., 1994: Central Western Carpathians), faunal elements from the belemnite association characterized by the true Mesohibolites are most commonly known throughout the Mediterranean Tethys (, 1939; , 1952; , 1965, 1970a; , 1968, 1973; , 1972; , 1973; , 1990; et al., 1994; et al., 2002; & , 2005). Late Barremian to Bedoulian belemnites are well known from the Caucasus. Unfortunately, publications with detailed stratigraphical information on belemnites are scarce. In general, papers dealing with ammonite stratigraphy also deal with other groups of fossils, including belemnites: , 1970; , 1971, 2000; & , 1975; et al., 1988; & , 2003.
During the times we collected, named and described the specimens mentioned herein it became apparent that several nominal species were not defined accurately enough from a stratigraphical point of view. As a result several "Lazarus species" occurred in our material. For instance, some of the species described by (1883b) originate from the upper part of the Hradiště-Schichten. This formation is of Barremian to Bedoulian age (cf. , 1978b, p. 118). Subsequent researchers attributed Late Barremian to Bedoulian ages to these species, without much further detail. From our investigations it appears that, morphologically similar species occur in restricted and stratigraphically well-separated intervals. As we can virtually rule out the influence of facies in the ABSS, the inaccurate origin of the type material results in ambivalent time-slice. Hence, we sometimes use the affiliation "aff.".
Throughout the text, the following abbreviations are used: "+" = invalid: a nomen dubium (nom. dub.), nomen nudum (nom. nud.), or nomen nullum (nom. null.) (= secondary typing error), "pt" = partim, LT = lectotype, HT = holotype, MT = by monotypy, and OD = original designation.
Materials will be stored in the collection of NCB-Naturalis (Leiden, The Netherlands). The inventory numbers are: RGM361471-361499, RGM361511-361598, RGM361800-361999, RGM543000-543034, RGM582933-583148, RGM583421-583835 (ABSS: ANG, and Combe Lambert: COM), RGM583149-583420 (chapelle Ste-Anne, north of Le Bourguet: BOU), RGM583836-583840 (Méouilles: MEO), and RGM583841-583877 (Demandolx: DLX).
We have arranged this chapter in several sections in relation to the stratigraphical distribution of the belemnite associations because we think it is practical and improves clarity.
Family Mesohibolitidae , 1983
Genus Hibolithes de , 1808
Hibolithes keleptrishvilii sp. nov.
Pl. 1 ![]() ,
figs. 22-23
,
figs. 22-23
? 1994 Mesohibolites garshini ; et al., p. 82-83, Pl. 26, figs. 3-4.
2002 Hibolites longior ; et al., p. 64-65, Pl. VI, figs. 1-2, 3-4 (not the synonymy).
? 2004 Hibolithes jaculiformis brevicostatus [sic!] ; & , p. 32 (bed 208 only).
pt 2009 Hibolithes jaculiformis ; , p. 16.
Derivation of name: Named after the belemnitologist Shalva (Tbilisi, Georgia).
Holotype: RGM 361817.
Stratum: Beds ANG042-043 to ANG050-051.
Material: Five specimens, all from ABSS. The holotype is the most complete one.
Description: The rostrum is characterized by a dorso-ventrally flattened apical part and a lateral compressed alveolar area. The rostrum is more or less spindle-shaped in ventral view with the widest part situated in the apical region. The alveolar groove is relative long, well-developed, approximately reaching halfway the rostrum. The alveolus reaches about 1/3 of the length of the rostrum. In lateral view the rostrum is more or less parallel, tapering towards a centrally placed apex in the apical part.
Remarks: The rostrum figured by et al. (1994) is probably a gerontic specimen. The specimens from Hungary are believed to originate from the Plesiospitidiscus ligatus Zone, but no ammonites were found that would rule out a stratigraphically younger level.
Phylogeny: From a morphological point of view it is probably descendant from H. subfusiformis s.s.
Stratigraphic occurrence: Latest Hauterivian (Pseudothurmannia ohmi Zone). The specimens from Georgia are from the Pseudothurmannia angulicostata Zone. The latter is (partially) equivalent to the P. ohmi Zone.
Belemnite association: HaBA2c (FO and LO).
Geographical occurrence: France (Vocontian Basin), Georgia (Kutaisi-area), Hungaria (probably) and Slovakia (probably).
Hibolithes ex gr. subfusiformis (, 1829)
Pl. 1 ![]() ,
figs. 1-2, 3-4 (juv.)
,
figs. 1-2, 3-4 (juv.)
Material: Several tens of nearly complete to partially preserved specimens.
Stratum: Beds ANG001-001a to ANG084-085.
Phylogeny: Most probably descendant from Hibolithes related to the H. longior-group.
Stratigraphic occurrence: Earliest Hauterivian (Acanthodiscus radiatus Zone) to earliest Barremian (T. hugii Zone).
Belemnite association: HaBA2c (common) to BaBA1a (LO).
Geographical occurrence: Circum Mediterranean Tethys.
Family Duvaliidae , 1914
Genus Duvalia , 1878
Duvalia ex gr. dilatata (, 1827)
Pl. 2 ![]() ,
figs. 3-4
,
figs. 3-4
Material: Seven specimens.
Stratum: Beds ANG007-008 to ANG015.1 and one loose incomplete specimen from the beds ANG036-040 (all Balearites balearis Zone).
Specimens included: D. dilatata dilatata (, 1827) and D. dilatata binervioides , 1965.
Specimens excluded: D. maioriana , 1965 (originally binomial dilatata majoriana) and D. variegata (, 1829). The first occurs in the Early/Late Hauterivian boundary beds while the latter occurs in the latest Valanginian and earliest Hauterivian and precedes D. ex gr. dilatata subsp.
Remarks: The two subspecies might represent a dimorphic pair.
Phylogeny: Most probably descendant from D. variegata - "vaunagensis" group.
Stratigraphic occurrence: Hauterivian (Crioceratites loryi Zone to Pseudothurmannia picteti Zone).
Belemnite association: HaBA2c (LO).
Geographical occurrence: Circum Mediterranean Tethys.
Family, Genus, Species unknown
Pl. 1 ![]() ,
figs. 24-25, 26-27
,
figs. 24-25, 26-27
Material: Six specimens.
Stratum: Beds ANG020-021 to ANG041-042.
Description: A small, spindle-like, laterally compressed (duvaloid?) specimen with conspicuous pseudobeloid morphology, with feeble ventro-lateral incisions.
Remarks: The alveolar area is not preserved, but the general appearance is like a juvenile Hibolithes. However, the latter taxa are always round to subrounded in cross-section. Due to the well-marked, but rather shallow, ventro-lateral incisions, it can not be attributed to any known genus. Pseudobelus-type of taxa are always characterized by (deep) mid-lateral incisions. These incisions relate to the so-called double-lines, and might indicate strengthening of the mantle muscle attachment. However, Pseudobelus is believed to be extinct by this time (cf. , 2009, Fig. 15).
Phylogeny: These rather small species might relate to dilatatoid Duvalia or to Pseudobelus.
Stratigraphic occurrence: Latest Hauterivian (Balearites balearis Zone).
Belemnite association: HaBA2c (FO and LO).
Geographical occurrence: Vocontian Basin (France).
Most characteristic are the belemnites that relate to H. jaculiformis and to D. aff. silesiaca-gagrica.
Family Mesohibolitidae , 1983
Genus Hibolithes de , 1808
Hibolithes ex gr. jaculiformis , 1913
Pl. 1 ![]() ,
figs. 5-6, 7-8, 9, 10-11, 12-13, 14-15, 16-17
,
figs. 5-6, 7-8, 9, 10-11, 12-13, 14-15, 16-17
1913 Hibolites jaculiformis var. brevisulcatus , p. 52-53, 68, Pl. III, fig. 4a-c, 4h-i.
1913 Hibolites jaculiformis n. sp. , p. 52-53, 68, Pl. II, figs. 5-6; Pl. III, figs. 4e-g, 4k-l, 11-12.
1913 Hibolites jaculiformis var. inflata , p. 52-53, 68, Pl. III, figs. 4d, 4j, 13-14.
? 1994 Hibolites jaculoides depressirostris ; et al., Pl. 25, figs. 1-2.
1994 Hibolites longior ; et al., Pl. 25, figs. 7-8.
? 2002 Vaunagites pistilliformis (); et al., Pl. V, fig. 6.
pt 2004 Hibolithes gr. jaculiformis ; & , Pl. III, fig. 6, non figs. 24-25.
+ 2004 Hibolithes jaculiformis brevicostatus [sic!] ; & , p. 32.
2004 Hibolithes gr. subfusiformis (); & , Pl. III, fig. 10.
? 2005 Hibolithes targovishtensis ; & , p. 68, Pl. III, figs. 7-12; Pl. V, figs. 12-13.
2005 ? Hibolithes jaculiformis ; & , p. 65, Pl. III, figs. 3-4.
? 2005 Hibolithes ? sp. & , Pl. III, figs. 18-19.
Material: Twenty-eight (ABSS) and fifteen (BOU) rostra in various stages of preservation.
Stratum: Beds ANG087 to ANG105.
Remarks: No type indicated by (1913).
Phylogeny: Most probably descendant from H. ex gr. subfusiformis.
Stratigraphic occurrence: Earliest Barremian (T. hugii Zone to K. nicklesi Zone).
Belemnite association: BaBA1a (FO) and BaBA1b (LO).
Geographical occurrence: Circum Mediterranean Tethys.
Hibolithes aff. jaculiformis , 1913
Pl. 1 ![]() ,
figs. 20-21
,
figs. 20-21
Material: One complete and four partially preserved rostra.
Stratum: Beds ANG110.2 to ANG110.4.
Description: The depicted specimen shows a medium sized, rather robust, dorso-ventrally well compressed rostrum. Herein, it differs from the generally more slender H. ex gr. jaculiformis, and the larger less compressed H. jaculiformis brevisulcatus.
Stratigraphic occurrence: Earliest Barremian (upper part of N. pulchella Zone).
Belemnite association: BaBA1c (FO and LO).
Geographical occurrence: France (Vocontian Basin).
? Hibolithes mirificus , 1965
* 1965 Hibolites mirificus sp. nov. , p. 151-152, Pl. I, figs. 1a-c [HT], 2-4.
pt 2005 Hibolithes mirificus ; & , p. 65, 67-68, Pl. III, figs. 5-6; Pl. V, fig. 14 (cum synonymy).
Material: Seven largely incomplete, hence the question-mark, specimens of different ontogeny.
Stratum: Beds ANG107.1-2 to ANG109.3-4.
Description: Parts of well-rounded to compressed guards, which occasionally show the (typical) pointed to mucronate apex preserved.
Remarks: The specimens mentioned by & (2005) from the beds 12, 399 and 410 do most probably not belong to the nominal species.
Stratigraphic occurrence: Earliest Barremian (N. pulchella Subzone).
Belemnite association: BaBA1c (FO and LO).
Geographical occurrence: Bulgaria, France, Hungary, and Slovakia.
Hibolithes targovishtensis , 1979
Pl. 1 ![]() ,
figs. 18-19
,
figs. 18-19
pt 1913 Hibolithes jaculiformis var. inflata , Pl. III, fig. 6.
* 1979 Hibolites targovishtensis sp. nov. , p. 37-39, Pl. I, figs. 1 [HT by OD], 2-7.
+ 1979 Hibolites targovichtensis sp. nov. , p. 37 (lapsus calami).
? 2005 Hibolithes targovishtensis ; & , p. 68, Pl. III, figs. 7-12; Pl. V, figs. 12-13.
Material: Eleven specimens.
Stratum: Beds ANG074-075 to ANG084-085.
Description: A small to medium sized, slightly dorso-ventrally compressed hibolitoid rostrum, with a well-developed alveolar groove and a short alveolus.
Remarks: Certain resemblance to immature H. ex gr. jaculiformis exists. In the ABSS in many, but not all of the specimens Acrothoracia-burrows occur.
Phylogeny: Most probably descendant from H. ex gr. subfusiformis.
Stratigraphic occurrence: Earliest Barremian (T. hugii Zone).
Belemnite association: BaBA1a (FO and LO).
Geographical occurrence: Bulgaria, France, Georgia (Abkhasia), and possibly Ukraine (Crimea).
Family Duvaliidae , 1914
Genus Duvalia , 1878
Duvalia ex gr. gagrica-silesiaca
Material: Fifteen specimens (ABSS) and see below for separate species descriptions.
Stratum: Beds ANG073 to ANG093-094.
Description: The species are well characterized by a bend, generally well visible, in the ventral part of the rostrum solidum. However, two different types of bending occur. In D. gagrica this bend is not accompanied by a strong constriction of the alveolar area, in contrast to D. aff. silesiaca, herein, it is like in D. dilatata binervioides. Moreover, it is smaller, more leaf-like in outer morphology (like in D. dilatata binervioides). Duvalia gagrica is more robust like D. dilatata dilatata, with a more elongated rostrum solidum. The alveolar opening shows a peculiar almost cross-like shape. Herein, in contrast to D. pontica, the lateral expulsions are shifted towards the dorsal side, while in D. pontica these are more or less situated in the middle part of the lateral side. However, as can be observed on the material depicted by (1913), in more mature specimens these expulsions become comparable.
Remarks: It is well possible that D. gagrica and D. aff. silesiaca are ontogenetical stages, or a dimorphic pair. Therefore, we have not introduced a new name yet for D. aff. silesiaca.
Phylogeny: Most probably descendant from D. ex gr. dilatata. It is succeeded by D. pontica, which is characterized by a more expanded alveolar part and a straight, laterally well flattened rostrum solidum with more or less parallel dorsal and ventral sides.
Stratigraphic occurrence: Earliest Barremian (T. hugii Zone).
Belemnite association: BaBA1a (FO and LO).
Geographical occurrence: Circum Mediterranean Tethys.
Duvalia gagrica , 1913
Pl. 2 ![]() ,
figs. 7-8 (?), 15-16 (?), 17-18, 27-28
,
figs. 7-8 (?), 15-16 (?), 17-18, 27-28
* 1913 Duvalia gagrica , p. 47, 67, Pl. II, fig. 4 [HT by MT].
2004 Duvalia gagrica ; & , p. 45, Pl. II, figs. 2-5; Pl. III, fig. 29 (cum synonymy).
Material: Five specimens.
Stratum: Beds ANG083 to ANG093-094.
Stratigraphic occurrence: Earliest Barremian (T. hugii Zone).
Belemnite association: BaBA1a (FO and LO).
Geographical occurrence: Circum Mediterranean Tethys.
Duvalia aff. silesiaca , 1902
Pl. 2 ![]() ,
figs. 9-10, 13-14, 19-20, 21-22, 23-24, 25-26
,
figs. 9-10, 13-14, 19-20, 21-22, 23-24, 25-26
1902 Belemnites (Duvalia) dilatatus forma silesiaca , p. 64, Pl. I, figs. 5 [LT], 6.
pt 1994 Duvalia binervia (); et al., Pl. 28, figs. 8-9; Pl. 30, figs. 7-9.
2004 Duvalia silesiaca ; & , p. 46, Pl. III, fig. 26 (cum synonymy).
Material: Ten specimens (ABSS), and two specimens from CBL.
Stratum: Beds ANG073-074 to ANG092-093.
Remarks: Lectotype indicated by (1978a). Specimens are virtually indistinguishable from Duvalia binervia (, 1829), except for the taxa that accompany this species. D. binervia occurs in post earliest Late Valanginian sediments and up to and including the earliest Hauterivian. As the age of the type stratum of Duvalia silesiaca is unclear (probably from Hauterivian), we prefer to use the affiliation "aff." for the species herein indicated.
Phylogeny: Most probably descendant from D. ex gr. dilatata.
Stratigraphic occurrence: Earliest Barremian (T. hugii Zone).
Belemnite association: BaBA1a (FO and LO).
Geographical occurrence: Circum Mediterranean Tethys.
Duvalia pontica , 1913
Pl. 2 ![]() ,
figs. 1-2, 5-6, 11-12; Pl. 5
,
figs. 1-2, 5-6, 11-12; Pl. 5 ![]() ,
figs. 15-16
,
figs. 15-16
* 1913 Duvalia pontica n. sp. , p. 46-47, 67, Pl. II, fig. 7 [HT by MT].
1965 ? Polygonalia pontica (); , p. 142.
1997 Duvalia sp. B; , p. 25-26, Pl. 4 ![]() ,
figs. 3, 6.
,
figs. 3, 6.
2002 Pseudoduvalia pontica (); et al., p. 165, Pl. XXIV, fig. 1 (cum synonymy).
Material: Seven specimens (ABSS) and one from CBL.
Stratum: Beds ANG100-101 to ANG103-104 and loose specimens from the beds ANG104 to 106, and the beds around ANG110.
Description: A robust, elongated, dilatatoid specimen, with a clear long groove and more or less parallel sides. In the alveolar area the outline of the rostrum is rhomboidal. Immature specimens are smaller but otherwise comparable to mature ones.
Phylogeny: Descendant from D. ex gr. silesiaca-gagrica.
Stratigraphic occurrence: Earliest Barremian (K. nicklesi to N. pulchella Zone).
Belemnite association: BaBA1b (FO) and BaBA1c (LO).
Geographical occurrence: Circum (Mediterranean) Tethyan.
Most characteristic are the species that relate to the new genus herein established, and the species of the genus Curtohibolites.
Family Mesohibolitidae , 1983
Genus Shvetsovia gen. nov.
= pars Mesohibolites auct. Pl.
= "Mesohibolites" sensu & , 2004
= "Mesohibolites" sensu & , 2005
Derivation of name: Named after the Russian geologist Mikhail Sergeevich (Михаил Сергеевич ).
Genotype: Hibolites gagricus , 1913.
Differential diagnosis: Elongated to very elongated fusiform (spindle-like) species, both from lateral as from dorsal or ventral view, with moderate to strong dorso-ventral flattening (subrounded to strong ellipsoidal cross-sections), except for the most apical part which has a more or less round cross-section. The length of the alveolar groove is variable, but generally it is short to very short while the alveolus is shallow to very shallow. Immature Conohibolites show a comparable morphology but can be separated by their dorso-ventrally compressed heart-shaped cross-sections. In Conohibolites the dorsal or ventral view is conical to cylindro-conical, but the lateral view is always conical with the apex clearly shifted towards the ventral side. The alveolar slit is more or less comparable to the text-fig. 2c depicted by
(1970a, p. 9). However, contrary to the slight curving depicted, it appears to be running almost straight from the apical line towards the ventral side, to the point where the alveolar groove ends abruptly. Very juvenile to very immature rostra show round cross-sections (Pl. 4 ![]() , fig. 22) but in Conohibolites these are ventro-dorsally compressed and more elongated (Pl. 6
, fig. 22) but in Conohibolites these are ventro-dorsally compressed and more elongated (Pl. 6 ![]() ,
figs. 13-14; Pl. 9
,
figs. 13-14; Pl. 9 ![]() ,
figs. 29-30).
,
figs. 29-30).
Remarks: Immature specimens of Conohibolites gladiiformis do show a more or less comparable elongated fusiform morphology, as do some Co. minaretiformis. In contrast to fusiform shaped Hibolithes, the spindle-shape in Shvetsovia is in some species, the result of broadening ("shoulder-like" developments) of the area in which the apical part is defined as part of the rostrum solidum. If present, it appears to result in a much more angular shaped rostrum as compared to the smoothly curved fusiform morphology. In the genus Hibolithes the alveolar area appears to be much more constricted as compared to Shvetsovia, giving the latter a more robust appearance. Moreover, in Hibolithes, the constriction between the rostrum solidum and the rostrum cavum often results in the exfoliation of the alveolar area, unlike in Shvetsovia.
Phylogeny: Most probably descendant from H. ex gr. jaculiformis.
Stratigraphic occurrence: Early Barremian (K. compressissima Zone) to early Late Barremian (I. giraudi Subzone).
Belemnite association: BaBA2a (FO) and lowermost part of BaBA5 (LO).
Geographical occurrence: Circum Mediterranean Tethys.
Shvetsovia gagrica (, 1913)
Pl. 3 ![]() ,
figs. 7-8 (ex gr.), 9-10, 13-14 (ex gr.), 19-20, 21-22, 25-26
,
figs. 7-8 (ex gr.), 9-10, 13-14 (ex gr.), 19-20, 21-22, 25-26
1913 Hibolites gagricus n. sp. , p. 57-58, 69, Pl. IV, fig. 1a-c.
2002 Mesohibolites gagricus (); et al., p. 76-77, Pl. VIII, fig. 1 (cum synonymy).
? 2005 Hibolithes ? sp.; & , p. 68, Pl. III, figs. 18-19.
non 2005 "Mesohibolites" ? gagricus (); & , p. 79 (= Co. ex gr. minaretiformis).
Material: Twenty specimens (ABSS) and twentyone specimens (BOU).
Stratum: Beds ANG120-121 to ANG131-132.
Description: A slender elongated, more or less fusiform guard with a relatively long alveolar groove and a relatively short alveolus. The alveolar opening is rounded to laterally compressed, while the rostrum solidum shows rounded to dorso-ventrally compressed cross-sections. More mature specimens show the development of "shoulders" (see remarks of the genus description) and the elongation of the apical area.
Remarks: A lectotype has been chosen from among the specimens that were not figured from 's collection by (1970a, p. 30-31). Subsequently (1972) and et al. (2002) invalidly indicated the most mature figured specimen to be the holotype.
Stratigraphic occurrence: S. defayae Subzone to C. darsi Subzone (ABSS), apparently being most abundant in the S. defayae Subzone. In BOU it is found in the topmost part of the N. pulchella Zone up to C. darsi Subzone. In BOU it is most abundant in the lower part of the K. compressissima Zone. Incomplete and certain phylogenetic stages in the development can be difficult to separate from varians (Pl. 3 ![]() ,
figs. 7-8, 13-14).
,
figs. 7-8, 13-14).
Belemnite association: BaBA2a (FO and common) and BabA2b (LO).
Geographical occurrence: Azerbaijan, Bulgaria, France, Georgia, Hungaria, and Russia (Kabardino-Balkariya ASR).
Shvetsovia varians (, 1913)
Pl. 3 ![]() ,
figs. 11-12, 15-16, 17-18, 23-24, 27-28, 29-30
,
figs. 11-12, 15-16, 17-18, 23-24, 27-28, 29-30
pt 1913 Hibolites varians n. sp. , p. 56-57, 69, Pl. IV, fig. 3a-d, 3e-f.
? pt 2002 Mesohibolites varians (); et al., p. 80-81, Pl. IX, figs. 1, 3 (?).
2005 Conohibolites ? varians (); & , p. 80-81, Pl. IV, figs. 16-17; Pl. V, figs. 2-3 (cum synonymy).
Material: Thirteen specimens (ABSS) and thirteen specimens (BOU).
Stratum: Beds ANG117-118 to ANG122-123.
Description: A smaller, much more spindle-like species (sensu stricto) as compared to S. gagrica. Mature (? or sensu lato) morphs show the development of an extremely long apical area comparable to the development in the conically shaped Co. gladiiformis.
Remarks: Based on stratigraphical data & (2005) provisionally regarded only the sensu stricto morph as the nominal one. Based on the belemnites we collected from the ABSS and BOU, 's original interpretation appears correct, but mature incomplete specimens are difficult to separate, hence they were regarded as species of the S. gagrica-varians group. et al. (2002) designated a lectotype (, 1913: Pl. IV, fig. 3e-f), as no type was originally indicated.
Stratigraphic occurrence: K. compressissima Zone (ABSS and BOU).
Belemnite association: BaBA2a (FO and common) and BabA2b (LO).
Geographical occurrence: Azerbaijan, France (Vocontian Basin), Georgia, Hungary (Gerecse Mts.), Ukraine (Crimea).
Shvetsovia aff. elegans (, 1913)
Pl. 4 ![]() ,
figs. 1-2, 3-4, 5-6, 7-8, 16-17,
23-24 (?), 27, 32-33; Pl. 8
,
figs. 1-2, 3-4, 5-6, 7-8, 16-17,
23-24 (?), 27, 32-33; Pl. 8 ![]() ,
figs. 1-2 (?)
,
figs. 1-2 (?)
pt 1841 Belemnites platyurus , Pl. XI, fig. 3.
non 1913 Hibolites elegans n. sp. , p. 60-61, 70, Pl. V, figs. 3a-b, 3c-f, 8c.
2005 ? "Mesohibolites" ? aff. elegans () sp. nov.; & , p. 78, Pl. IV, figs. 20-25; Pl. V, figs. 10-11 (pars cum synonymy).
Material: Twelve specimens (ABSS) and three specimens from BOU.
Stratum: Beds ANG133-134 to ANG138-139 and BOU141.2 to BOU142.1.
Description: A more or less subfusiform hibolitoid rostrum, however with a more or less feeble to very distinct (Pl. 4 ![]() ,
figs. 7-8) flattening of the ventral side of the rostrum solidum, except for the apical part. The latter has a much more rounded outline. The length of the alveolar groove is variable extending relatively far beyond the initiation of the alveolus. Lateral lines are often well visible.
,
figs. 7-8) flattening of the ventral side of the rostrum solidum, except for the apical part. The latter has a much more rounded outline. The length of the alveolar groove is variable extending relatively far beyond the initiation of the alveolus. Lateral lines are often well visible.
Remarks: Apparently, the length of the alveolar groove is variable, both in immature as well as in more mature specimens (Pl. 4 ![]() ,
figs. 5-6, 16-17). Also, the outline of the cross-sections in the rostrum solidum varies from specimens with well flattened ventral areas, to more rounded outlines. The nominal species was originally described from
"Aptian" sediments of Abkhasia by
(1913), hence the affiliation "aff.".
,
figs. 5-6, 16-17). Also, the outline of the cross-sections in the rostrum solidum varies from specimens with well flattened ventral areas, to more rounded outlines. The nominal species was originally described from
"Aptian" sediments of Abkhasia by
(1913), hence the affiliation "aff.".
Stratigraphic occurrence: In the C. darsi Subzone (top part) to Macroscaphites tirolensis Subzone.
Belemnite association: BaBA2c (FO) and BaBA3a (LO).
Geographical occurrence: France (Vocontian Basin), Georgia (?), Hungary (Gerecse Mts.) and Slovakia (?).
Shvetsovia simionescui (, 1970)
Pl. 4 ![]() ,
figs. 15, 25-26; Pl. 8
,
figs. 15, 25-26; Pl. 8 ![]() ,
figs. 3-4 (?)
,
figs. 3-4 (?)
pt 1898 Belemnites jaculum ; , p. 52 (108)-53 (109), Pl. I, fig. 5.
1952 Mesohibolites minaret (); , p. 715, text-fig. 38.
* 1970a Hibolites simionescui sp. nov. , p. 21-22, 68, Pl. VI, fig. 4 [HT by MT].
pt 1970a Mesohibolites minaret (); , Pl. VIII, figs. 5-6.
Material: Twelve (ABSS) and seven (BOU) specimens.
Stratum: Beds ANG140 to ANG149.3-4.
Description: A rather striking species with a well compressed (ventro-dorsally) rostrum. It has a deep alveolus, a well developed alveolar groove that fades away in the apical part, giving way to a characteristic flattened area. In lateral view the rostrum tapers down towards the apex, while in ventral view, the rostrum is subcylindrical to cylindro-conical. The typical broadening ("shoulder-like" development) is well visible.
Remarks: Certain resemblance to (very) mature S. gagrica exists. However, the latter is generally of larger size with a much more extended apical area.
Stratigraphic occurrence: Ho. uhligi Zone to lower part of To. vandenheckii Zone (He. sayni Subzone).
Belemnite association: BaBA3b (FO) and BaBA3c (LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), and Romania.
Genus Curtohibolites , 1963
Within the original concept of Curtohibolites, five specimens were distinguished. Of these, two show a conical to subcylindrical outer morphology, while the other three are much more subcylindrical to pistilliform, thus with a more constricted alveolar part. The latter species include the genotype (Curtohibolites trubatchensis , 1963). The conical morphs include Curtohibolites rasgradensis , 1963, Cu. wernsdorfensis , 1963, and a new species described herein. This group of species seems to have a different stratigraphical level, they occur in older sediments, as compared to the more subcylindrical group of species, and are placed, with some reservation, in the nominal genus.
Curtohibolites (?) bourguetensis sp. nov.
Pl. 4 ![]() ,
figs. 18-19, 28-29
,
figs. 18-19, 28-29
Derivation of name: Named after the geographical locality it is found in the vicinity of.
Holotype: RGM 583366.
Type stratum: BOU137.2-138 (K. compressissima Zone).
Material: One immature specimen from the ABSS and one mature specimen from BOU (holotype).
Stratum: The specimen from the ABSS originates from the beds ANG129-131. The mature specimen originates from the marly interval with calcareous nodules between BOU137.2 and bed BOU138.
Description: A robust specimen with a short rostrum solidum. It is ventro-dorsally well compressed with an alveolus which reaches half-way the preserved part of the rostrum. The apex is shifted to the ventral side. Also, the immature specimen is well-compressed but shows a relative long alveolar groove, combined with a deep alveolus.
Remarks: Differs from Cu. (?) wernsdorfensis by the absence of two plains of symmetry. Moreover, it is much more dorso-ventrally compressed, and shows a less elongated apical area.
Stratigraphic occurrence: Topmost part of the K. compressissima Zone and lower part of the C. darsi Zone.
Belemnite association: BaBA2a (FO) and BaBA2c (LO).
Geographical occurrence: France (Vocontian Basin).
Curtohibolites aff. pinguis (, 1913)
2005 Curtohibolites aff. pinguis (); & , p. 79, Pl. IV, figs. 30-31; Pl. V, figs. 20-21.
2010 Curtohibolites aff. pinguis (); , p. 663, 665, text-fig. 2.16-17.
Material: One specimen (ABSS).
Stratum: A loose specimen from the marly beds between ANG135 to ANG137.
Description: Specimen is comparable to Cu. oosteri but for the development of the alveolar part of the rostrum. It shows a more or less sub-trapezoidal with the widest part being situated at the dorsal side.
Remarks: See Cu. oosteri.
Stratigraphic occurrence: M. tirolensis Subzone.
Belemnite association: BaBA2c (FO and LO).
Geographical occurrence: France (Vocontian Basin), and Hungary (Gerecse Mts.).
Curtohibolites (?) wernsdorfensis , 1963
Pl. 3 ![]() ,
figs. 1-2, 3-4, 5-6
,
figs. 1-2, 3-4, 5-6
1883 Belemnites aff. extinctorius [sic!] (); , p. 175, Pl. I, fig. 12.
* 1963 Curtohibolites wernsdorfensis sp. nov. , p. 216-217, Pl. II, fig. 7 [HT by MT] (cum synonymy).
1970a Curtohibolites wernsdorfensis ; , p. 24, Pl. VII, figs. 6 (= , 1963), 7.
+ 1975 Mesohibolites garschini [sic!] ; & , p. 140-141, Pl. I, fig. 2.
pt 2002 Mesohibolites garshini ; et al., Pl. IX, fig. 4 (= & , 1975).
2005 ? Conohibolites (?) aff. platyurus () sp. nov.; & , p. 75, Pl. V, figs. 8-9.
non 2010 Curtohibolites (?) wernsdorfensis ; , p. 665, text-fig. 2.7-8 (=Co. tzankovi).
Material: Five specimens (ABSS) and two from BOU.
Stratum: Beds ANG115 to ANG120-121.
Description: A species with a striking cylindro-conical morphology with round to rounded or subrounded to dorso-ventrally compressed cross-sections. The alveolus reaches about half-way the rostrum, while the clear alveolar groove reaches just beyond the extension of the alveolus.
Remarks: Specimen strongly resembles Conohibolites (but they are not cylindro-conical in both lateral as in dorsal or ventral view) or very large Curtohibolites. However, both genera appear in younger strata only, probably indicating this to be the ancestor to either one of these taxa. As the S. gagrica-varians group is most probably the ancestor of Co. gladiiformis, attribution to the generally smaller Curtohibolites appears to be most probable. Also, its outer-morphology suggests a close resemblance to the latter genus, which is sub-cylindrical to cylindro-conical. Characteristic appears to be the double plain of symmetry, both from lateral as from dorso-ventral view the rostrum appears to be near symmetrical.
Phylogeny: Doubtful, but probably descendant from H. mirificus, which from a morphological point of view appears to be most comparable.
Stratigraphic occurrence: In the upper part of the K. compressissima Zone (ABSS). In the upper part of the N. pulchella Zone and the K. compressissima Zone of BOU and CBL.
Belemnite association: Topmost part of BaBA1c (FO) and BaBA2a (LO).
Geographical occurrence: Bulgaria, Czech Republic (Veřovice (= Wernsdorf), Moravia), France (Vocontian Basin), and Hungary (Gerecse Mts.).
Family Duvaliidae , 1914
Genus Duvalia , 1878
Duvalia vermeuleni sp. nov.
Pl. 5 ![]() ,
figs. 1-2, 3-4, 7-8
,
figs. 1-2, 3-4, 7-8
pt 1841 Belemnites grasianus , p. 63.
1989 Duvalia grasiana (); & , Pl. 2 ![]() , fig. 4.
, fig. 4.
1994 Duvalia grasiana (); et al, Pl. 30, figs. 3-4 (= & , 1989), 5-6.
2005 Duvalia sp. nov. (immature?); & , Pl. III, figs. 1-2.
2005 Duvalia grasiana (); & , Pl. III, figs. 38-39; Pl. V, figs. 18-19, 25-26.
2010 Duvalia ex gr. grasiana (); , p. 667-668, Text-fig. 2.20-21.
Derivation of name: Named after Jean (Barrême, France), specialist in Barremian ammonites, who donated the holotype.
Material: Twenty-four specimens (ABSS) and four, in part ex situ, specimens (BOU).
Holotype: RGM 361561 (a gerontic specimen from ANG120; Pl. 5 ![]() ,
figs. 1-2).
,
figs. 1-2).
Paratype: RGM 583021 (ANG120-121).
Type stratum: Bed ANG120, S. defayae Subzone, K. compressissima Zone.
Differential diagnosis: Dilatatoid rostrum with a relative long alveolar groove and more or less lozenge-shaped cross-sections. Essentially it differs from typical D. grasiana by the shorter length of the alveolar groove (however, atypically long in the holotype), the less deep alveolus, and the more lozenge-shaped cross-sections. In typical D. grasiana these are always ellipsoidal. Also, D. grasiana differs by its “fatter" appearance, i.e., more latatoid as compared to the more slender, dilatatoid D. vermeuleni. Also, the apical area is more flattened in D. vermeuleni as compared to typical grasiana. It differs from D. pontica due to its parallel lateral sides, without the long pronounced expanded alveolar area. Duvalia grasiana shows a much longer alveolar groove, always reaching the apex, and it is much more latatoid, i.e., less laterally compressed. In mature D. grasiana the apical area appears to be slightly different as compared to the stratigraphically older species. In general, it is characterized by a more laterally flattened ventral part (as if it were squeezed), as compared to the dorsal side of the apical area.
Description: The dorsal and ventral sides are more or less parallel but for the most apical part. The apex is displaced to the dorsal side. As a result the dorsal side is less curved in the apical part, compared to the ventral side. The deep alveolus reaches about half-way the rostrum. The dorsal alveolar groove generally ends near the apical area. It should be noted that the holotype shows an atypical long alveolar groove. In the paratype (Pl. 7 ![]() ,
figs. 3-4) the more typical length of the alveolar groove is shown.
,
figs. 3-4) the more typical length of the alveolar groove is shown.
Ontogeny: The ontogenetical development is essentially simple. All stages are comparable but for their size.
Variations: As in most duvaliids, mature specimens develop dorsal bulges, giving way to a more
irregular outline. Some specimens are partially more latatoid, i.e., they show a thicker, more robust rostrum in the alveolar part, with clearer lozenge-shaped cross-sections, but with more flattened lateral sides in the apical area
[see Duvalia schwetzowi (, 1999)]. In the G. sartousiana Zone the more rounded latatoid cross-sections appear (Pl. 5 ![]() ,
figs. 7-8), however initially with a shorter groove as compared to typical D. grasiana (Pl. 5
,
figs. 7-8), however initially with a shorter groove as compared to typical D. grasiana (Pl. 5 ![]() ,
figs. 6, 9-10, 11-12). The latter
undoubtedly occurs in the I. giraudi Zone and in younger sediments, but specimens with squeezed ventral apical areas first occur in the H. feraudianus Subzone (ANG166.2-ANG167).
,
figs. 6, 9-10, 11-12). The latter
undoubtedly occurs in the I. giraudi Zone and in younger sediments, but specimens with squeezed ventral apical areas first occur in the H. feraudianus Subzone (ANG166.2-ANG167).
Remarks: The sometimes feeble differences (splitting?) that are noted between specimens of D. ex gr. grasiana are based on stratigraphical observations. From this stratigraphical point of view, within the investigated material from the ABSS the range of variation within all the early specimens does not exceed the observed range which exists in the later species. However, as we compare the variation of D. grasiana from Germany as depicted by (1911, Pl. VII and VIII) it seems that the population is morphologically comparable to specimens from older stratigraphical layers and even to D. silesiaca-like morphs (, 1911, Pl. VIII, fig. 11, a teratological specimen according to the author).
Phylogeny: Most probably descendant from D. pontica.
Stratigraphic occurrence: Beds ANG120 to ANG161.2-3 (S. defayae Subzone to G. sartousiana Subzone).
Belemnite association: BaBA2a (FO) and BaBA4b (LO).
Geographical distribution: France (Vocontian Basin), Hungary (Gerecse Mts.), and Slovakia.
Duvalia schwetzowi (, 1999)
Pl. 7 ![]() ,
figs. 5a-c, 13-14, 17-18
,
figs. 5a-c, 13-14, 17-18
? 1883 Duvalia Grasi [sic!] ; , Pl. I, fig. 6.
1913 Duvalia Grasi [sic!] var. B; , p. 48, 67, Pl. II, fig. 8e-g.
pt 1957 Duvalia Grasi [sic!] var. schwetzovi var. nov. , p. 53-54 (pars cum synonymy).
non 1957 Duvalia Grasi [sic!] var. schwetzovi var. nov. , Pl. II, figs. 8-9.
* 1999 Pseudoduvalia schwetzowi sp. nov. , p. 23-24, Pl. 2 ![]() , fig. 8 [HT by OD].
, fig. 8 [HT by OD].
2005 Duvalia grasiana (); & , p. 65 (section E, beds: 300/10, 300/2).
Material: Six specimens (ABSS) and four specimens from DLX.
Stratum: Beds ANG142.3-4 to ANG172.3, and beds DLX106-107 to DLX110-111.
Differential diagnosis: Dilatatoid rostrum with a relative short alveolar groove. It shows a typical quadrangular shaped outline of the alveolar region.
Description: The dorsal and ventral sides are more or less parallel but for the apical part. The apex is displaced to the dorsal side. As a result the dorsal side is less curved in the apical part, compared to the ventral side. The alveolar groove is relative short and more or less restricted to the alveolar area, the depth of the alveolar groove is comparable to the length of the alveolar groove. The alveolar opening is characterized by a typical quadrangular shaped outline. The widest parts are more or less centrally placed.
Remarks: A certain resemblance to D. pontica can be observed. However, it differs from D. pontica due to the absence of the typical bend that characterizes the rostrum solidum of the latter species. D. grasiana shows a much longer alveolar groove. It is much more latatoid, i.e., less laterally compressed, in the rostrum solidum. The new specimen of (1999) is well comparable. However, it originates from the early Bedoulian Deshayesites deshayesi Zone.
Stratigraphic occurrence: From the Ho. uhligi Zone to I. giraudi Zone. In Georgia it occurs in the early Bedoulian D. deshayesi Zone.
Belemnite association: BaBA3b (FO) and BaBA5b (LO).
Geographical distribution: France (Vocontian Basin), Georgia (Abkhasia), Hungary and probably Slovakia.
This association is characterized by the abundance of Shvetsovia and the first occurrence of Conohibolites, the last occurrence of Curtohibolites while Hibolithes is absent.
Family Mesohibolitidae , 1983
Genus Conohibolites & , 2005
Conohibolites gladiiformis (, 1883)
Pl. 4 ![]() ,
figs. 9-10, 11-12; Pl. 8
,
figs. 9-10, 11-12; Pl. 8 ![]() ,
figs. 13-14, 15-16, 19-20, 25-26, 30-31
,
figs. 13-14, 15-16, 19-20, 25-26, 30-31
* 1883 Belemnites gladiiformis n. sp. , p. 176, Pl. I, fig. 2 [HT by MT].
non 1978a Mesohibolites cf. gladiiformis (); , p. 14-16, Pl. II, fig. 5 (= M. beskidensis).
? 2002 Mesohibolites garshini ; et al., Pl. VIII, fig. 6.
? 2005 Conohibolites garshini (); & , Pl. III, figs. 36-37 (imm.).
? 2005 Conohibolites gladiiformis (); & , Pl. IV, figs. 5-6.
pt 2005 Conohibolites gladiiformis (); & , p. 74, Pl. IV, figs. 7-11; Pl. V, fig. 22 (cum synonymy).
2010 Conohibolites cf. gladiiformis (); , p. 662-663, Text-fig. 2.24.
2010 Conohibolites ex gr. gladiiformis (); , p. 663, Text-fig. 2.9-10.
2010 Conohibolites sp. A; , p. 663, Text-fig. 2.22-23 (cum synonymy).
Material: Nine (ABSS), five (DLX), and thirty-three (BOU) specimens.
Stratum: Beds ANG144 to ANG149.4, and BOU142.3 to 146.1.
Description: Elongated ventro-dorsally compressed rostrum. It tapers down from the alveolar opening towards the apex. Both from lateral view as from a ventral view it is conical to cylindro-conical. Mature to gerontic specimens (Conohibolites sp. A sensu , 2010, text-fig. 2.22-23) appear very robust while immature specimens are clearly much more slender (hence they are comparable to S. carpatica) and elongated. The alveolar opening is more or less rounded in immature specimens and ventro-dorsally compressed in mature to gerontic specimens. Juvenile to immature specimens are well flattened and spindle-like in ventral view.
Remarks: The species is relative rare in the ABSS. However, it is common in the proximal facies of BOU. Especially, mature to gerontic specimens occur quite frequently in these more proximal deposits.
Stratigraphic occurrence: H. uhligi Zone to To. vandenheckii Zone (He. sayni Subzone or lower part of B. barremense Subzone).
Belemnite association: BaBA3c (FO and LO).
Geographical occurrence: Bulgaria, Czech Republic, France (Vocontian Basin), Georgia, Hungary (Gerecse Mts.), and Slovakia.
Conohibolites ex gr. gladiiformis (, 1883)
2005 Conohibolites gr. gladiiiformis () ? n. sp.; & , p. 74-75, Pl. IV, figs. 14-15; Pl. V, figs. 23-24.
Material: One very incomplete specimen from the ABSS.
Stratum: Bed ANG142.2-143.1 (possibly ex situ).
Description: A very compressed and elongated specimen with a very long alveolar groove and a short alveolus.
Remarks: Due to the extreme ventro-dorsal compression of the rostrum solidum and the long alveolar groove, small parts can easily be identified, except for the apical most parts. The partially preserved specimen from the ABSS appears to be even more flattened as compared to the specimen from Hungary.
Stratigraphic occurrence: Ho. uhligi Zone.
Belemnite association: BaBA3b (single occurrence).
Geographical occurrence: France (Vocontian Basin) and Hungary (Gerecse Mts.).
Conohibolites tzankovi (, 1965)
Pl. 7 ![]() ,
figs. 13-14, 27-30; Pl. 8
,
figs. 13-14, 27-30; Pl. 8 ![]() ,
figs. 36-38; Pl. 11
,
figs. 36-38; Pl. 11 ![]() ,
figs. 17-18a
,
figs. 17-18a
1898 Belemnites minaret ; , Pl. I, fig. 3.
* 1965 Mesohibolites tzankovi n. sp. , p. 158-159, Pl. I, fig. 7 [HT by OD]; Pl. IV, fig. 8; Pl. VII, figs. 2-4.
? 1970a Mesohibolites platyurus (); , Pl. XVIII, fig. 5; Pl. XIX, fig. 4.
1970a Mesohibolites tzankovi ; , p. 37-38, Pl. X, figs. 1-4; Pl. XXXII, fig. 14.
2010 Curtohibolites (?) wernsdorfensis ; , p. 665, Text-fig. 2.7-8.
Material: Six specimens (BOU), and one from DLX.
Stratum: BOU142.3-BOU144.
Description: A typical conical to cylindro-conical rostrum with a deep alveolus and consequently a relative short rostrum solidum with a more or less pointed to elongate apex. Cross-sections are typically heart-shaped, but are more subrounded to heart-shaped in the alveolar part. In lateral view, the dorsal side tapers much more rapidly to the apical area, as compared to the ventral side. The latter is more or less straight, except for the apical most part in which a feeble incurvation towards the apex can be observed. The apex is elongated. From ventral view, the apical area is more or less minaret-like shaped. Overall, the rostrum is dorso-ventrally compressed.
Remarks: In Co. platyurus the rostrum solidum is larger giving it a more elongated appearance and the alveolar opening is well-rounded to feebly dorso-ventrally flattened. In the nominal species the extension of the apical area appears to be much larger as compared to the specimens described herein. Also the rostrum solidum appears to be larger in the specimens herein described.
Phylogeny: Most probably descendant from Co. gladiiformis.
Stratigraphic occurrence: Latest Early Barremian to earliest Late Barremian (M. tirolensis Subzone (?) - Ho. uhligi Zone).
Belemnite association: BaBA3c (FO and LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin; apparently absent from the more distal facies in the ABSS), and Romania.
Genus Curtohibolites , 1963
Curtohibolites oosteri , 1963
Pl. 4 ![]() ,
figs. 20-21, 30-31
,
figs. 20-21, 30-31
1857 Belemnites Orbignyanus ; , Pl. I, figs. 9-10, 11, 12-15.
* 1963 Curtohibolites oosteri sp. nov. , p. 215-216, Pl. I, figs. 2 [HT]-3 (pars cum synonymy).
1964 Curtohibolites oosteri ; , p. 138, 139, 146.
pt 1963 Curtohibolites orignyanus (); , p. 214-215, Pl. I, fig. 1 (not the synonymy).
? 1997 Conobelus orbignyanus (); , p. 149, Pl. 59, fig. 7.
Material: Seven specimens (BOU).
Stratum: From the beds BOU142.4 (top part) to BOU144.
Description: A small sized, clavate to pistilliform specimen, with a more or less mucronate apex, a relative long alveolar groove and a shallow alveolus. The rostrum solidum is slightly compressed, ventro-dorsally. The alveolar cross-section in one specimen is comparable to the one shown by (1963, Pl. I, fig. 2d), that is more or less sub-trapezoidal.
Remarks: Herein, the widest part is situated in the ventral area, in contrast to Cu. aff. pinguis in which it is situated to the dorsal side. At the moment it is not known whether or not this difference is part of the variety of Cu. oosteri.
Stratigraphic occurrence: From the M. tirolensis Subzone.
Belemnite association: BaBA3c (FO and LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Suisse, and possibly Ukraine (Crimea).
Genus Shvetsovia gen. nov. (for description see above)
Shvetsovia carpatica (, 1883)
Pl. 4 ![]() ,
figs. 13-14; Pl. 8
,
figs. 13-14; Pl. 8 ![]() ,
figs. 5-6 (?)
,
figs. 5-6 (?)
* 1883 Belemnites carpaticus n. sp. , p. 177, Pl. I, fig. 1 [HT by MT].
2005 Hibolithes? carpaticus (); & , p. 68-69, Pl. III, figs. 20-27; Pl. V, figs. 15, 17 (cum synonymy).
Material: Eleven specimens (ABSS), two (DLX), and fifteen specimens from BOU.
Stratum: Beds ANG136.1-2 to ANG151.3-152.
Description: Very elongated, ventro-dorsally compressed rostrum, except for the alveolar part. The alveolus is shallow, with a short alveolar groove. Sometimes cross-sections in the apical part are more rounded. The alveolar area is more or less laterally compressed. In ventro-dorsal view, immature specimens are more or less spindle-shaped.
Remarks: Rather typical taxon with a very elongated rostrum. Differentiation between incomplete specimens of S. carpatica and Co. gladiiformis is sometimes difficult, but for their mature to gerontic stages.
Stratigraphic occurrence: In the upper part of the C. darsi Zone (M. tirolensis Subzone) to To. vandenheckii Zone (B. barremense Subzone).
Belemnite association: BaBA3a (FO) to BaBA4a (LO).
Geographical occurrence: Czech Republic, France (Vocontian Basin), Georgia, and Hungary (Gerecse Mts.).
This association is characterized by the abundance of Conohibolites and Shvetsovia, while Curtohibolites and Hibolithes are absent, and Mesohibolites is not yet developed.
Family Mesohibolitidae , 1983
Genus Conohibolites & , 2005
Conohibolites garshini (, 1965)
Pl. 7 ![]() ,
figs. 9-10, 11-12, 17-18, 19-20; Pl. 8
,
figs. 9-10, 11-12, 17-18, 19-20; Pl. 8 ![]() ,
figs. 17-18
,
figs. 17-18
+ 1964 Mesohibolites garschini sp. nov. , p. 139, 145 (nom. nud.).
* 1965 Mesohibolites garshini sp. nov. , p. 157, Pl. III, fig. 4 [HT by OD]-6.
1970a Mesohibolites garshini ; , p. 36-37, Pl. XVI, figs. 5-7 (= , 1965); Pl. XXXII, fig. 15.
non 1975 Mesohibolites garschini [sic!] ; & , Pl. I, fig. 2 (= Cu. (?) wernsdorfensis).
non 1994 Mesohibolites garshini ; et al, Pl. 26, figs. 3-4 (= & , 1989: Pl. 2, fig. 3), 5 (= Co. ex gr. minaretiformis).
non 2005 Conohibolites garshini (); & , Pl. III, figs. 36-37.
pt 2005 Conohibolites garshini (); & , p. 72, Pl. IV, figs. 3-4.
pt 2005 "Mesohibolites" sp. A (aff. minaretiformis ); & , Pl. IV, figs. 26-27.
Material: Fifteen specimens (ABSS) and seven (BOU).
Stratum: Beds ANG150 to ANG163.1b.
Description: Cylindro- to subcylindro-conical specimen with a relative robust appearance. The ventral side is more or less flattened, with a well developed alveolar groove reaching more or less halfway to the rostrum. The alveolus occupies approximately 1/3 of the length of the rostrum. The apical area is rather short with a ventrally shifted apex. In lateral view, the rostrum typically tapers down to the apex in its dorsal side, but the ventral side is more or less straight, except for its apical-most part. Due to the flattened ventral area, however with a well-developed alveolar groove, the outline of the cross-sections in the alveolar part and the rostrum solidum, except for the apical most part, a more or less lozenge-shaped.
Remarks: The ventro-lateral tapering aspect is clearly not as pronounced as in Co. platyurus and Co. tzankovi, while the apical end is clearly less extended.
Stratigraphic occurrence: Top part of the He. sayni Zone to the Gerhardtia provincialis Subzone.
Belemnite association: BaBA4a (FO and common) and BaBA4b (LO).
Geographical occurrence: Bulgaria, France, Georgia, and Hungary.
Conohibolites ex gr. minaretiformis (, 1913)
The specimens that belong to this group of belemnites are quite variable. From a morphological point of view they are close to several species that occurred previously or occur simultaneously, viz. Conohibolites garshini, Co. gladiiformis, Co. platyurus, and Shvetsovia carpatica. Also, they bear some resemblance to certain kinds of true Mesohibolites exists (e.g. Mesohibolites beskidensis). In particular, individual incomplete specimens can often not be distinguished with certainty. Several of these closely resembling species show their last occurrence approximately at the beginning of the initiation of this group of belemnites, or disappear short after the rise of this group. For the moment it is not clear whether or not a new genus should be introduced. Therefore they are for the moment included within the genus Conohibolites, which they most resemble morphologically.
Conohibolites minaretiformis (, 1913)
Pl. 6 ![]() , figs, 1-2, 3-4, 5-6, 7-8, 9-10, 11-12, 17-18, 19-20, 21-22, 25-26, 27-28;
, figs, 1-2, 3-4, 5-6, 7-8, 9-10, 11-12, 17-18, 19-20, 21-22, 25-26, 27-28;
Pl. 7 ![]() ,
figs. 1-2, 3-4, 5-6; Pl. 8
,
figs. 1-2, 3-4, 5-6; Pl. 8 ![]() ,
figs. 21-22, 11-12, 28-29, 32-33, 34-35;
,
figs. 21-22, 11-12, 28-29, 32-33, 34-35;
Pl. 9 ![]() ,
figs. 5-6, 13-14, 15-16, 26-27
,
figs. 5-6, 13-14, 15-16, 26-27
pt 1883 Belemnites minaret ; , Pl. I, fig. 9, non fig. 8 (= S. elegantoidea).
1898 Belemnites beskidensis ; , p. 107 (51)-108 (52), Pl. I, fig. 4.
? 1898 Belemnites sp. , Pl. I, fig. 7.
1913 Hibolites minaretiformis n. sp. , p. 54-55, 68-69, Pl. IV, fig. 5a-d, e-f [LT].
non 1915 Belemnites (Aulacobelus) minaretiformis ; & , p. 255, 269 (= Adiakritobelus sp.).
1919 Mesohibolites minaretiformis ; , p. 45.
1970 Mesohibolites beskidensis ; , p. 105-106, Pl. XX, fig. 4.
1970a Mesohibolites minaretiformis (); , p. 26-27, Pl. X, figs. 5-7.
? 1995 Mesohibolites beskidensis ; & , p. 106 (probably from the To. vandenheckii Zone).
2002 Mesohibolites minareticus ; et al., Pl. XI, fig. 4.
pt 2005 "Mesohibolites" sp. A (aff. minaretiformis ); & , p. 78, Pl. IV, figs. 28-29.
pt 2005 "Mesohibolites" ? gagricus (); & , p. 79.
Material: About seventy specimens (ABSS) and about twenty specimens from BOU. Besides several dozens of incomplete specimens (BOU) that might partially belong to S. elegantoidea.
Stratum: ANG151.2 to ANG166.1.
Description: An elongated, dorso-ventrally compressed, rostrum with a pointed apex and a shallow alveolar cavity. Two clear ontogenetical stages can be observed. In juvenile to immature specimen, the whole rostrum is dorso-ventrally flattened. In ventral view the rostrum is elongated spindle-like, with a very pointed apical part, a short alveolar groove and a shallow alveolar cavity. In immature specimen the first hint of a feeble ventro-lateral ridge occurs in the alveolar part that is not always well-visible, but well-defined by touch. This ridge disappears in gerontic specimens. Mature to gerontic specimens have a cylindro-conical outline. In lateral view, this outline is askew. The ventral side is more or less straight in the alveolar part and part of the rostrum solidum. However, the apical part is characterized by a curvation towards the central to slightly ventral shifted apex. This curvation is slightly stronger in the dorsal side, and increases in gerontic specimens. However, the outlines of the rostrum cavum and part of the rostrum solidum remain more or less parallel for about half of the total length of the rostrum, like in Co. garshini. However, the latter has a much more robust appearance as compared to Co. minaretiformis. Moreover, Co. garshini has a minaret-like apical part, while the stout morph of Co. ex gr. minaretiformis has a blunt apex. The alveolar groove is well-defined and in mature specimens it runs onto the rostrum solidum and disappears gradually. The alveolar cavity is short, approximately one-fifth to one-sixth of the total length of the rostrum. In mature to gerontic specimens the alveolar opening can be rounded to faintly laterally compressed, while in less mature specimens the alveolar opening is more or less dorso-ventrally flattened.
In Conohibolites the alveolar slit has a more or less a straight appearance, with a sudden curvation near the apical line, comparable to the Mesohibolites-type depicted by (1970a, text-fig. 2c). Shvetsovia shows an alveolar slit which is different. Its appearance is more or less comparable to something in between the text-fig. 2b (Hibolithes) and 2f (Duvalia; without the "dentation") as depicted by (1970a). However, it is not clear if the morphology of the alveolar slit is of generic taxonomical significance.
Remarks: (1970a) selected a rather blunt, more robust morph as the LT. In our material this morph (Pl. 6 ![]() ,
figs. 1-2, 3-4) is rare and restricted to the lower beds of the occurrence of Co. ex gr. minaretiformis. The specimen described as M. beskidensis () by
(1970), appears to originate from the G. sartousiana Zone and not from beds with Pulchellia and Heinzia (cf. ,
2002, p. 182).
,
figs. 1-2, 3-4) is rare and restricted to the lower beds of the occurrence of Co. ex gr. minaretiformis. The specimen described as M. beskidensis () by
(1970), appears to originate from the G. sartousiana Zone and not from beds with Pulchellia and Heinzia (cf. ,
2002, p. 182).
Stratigraphic occurrence: B. barremense Subzone to H. feraudianus Subzone.
Belemnite association: BaBA4a (FO, common in topmost part) and BaBA4c (LO).
Geographical occurrence: Bulgaria, Czech Republic, France (Vocontian Basin), Georgia, and Hungaria.
Conohibolites platyurus (, 1841)
Pl. 7 ![]() ,
figs. 7-8, 15-16, 21-22 (?), 23-24
,
figs. 7-8, 15-16, 21-22 (?), 23-24
pt 1841 Belemnites platyurus , p. 73-74, Pl. XI, fig. 1, 4 [LT].
non 1841 Belemnites platyurus , Pl. XI, fig. 2 (= juvenile Mesohibolitidae), non fig. 3 (= S. aff. elegans).
? 1952 Belemnites (Hibolites?) escragnollensis n. sp. , p. 150-154, text-figs. 60-63, Pl. V, fig. 69.
pt 1970a Mesohibolites platyurus (); , p. 27-28, Pl. XIX, figs. 5-7; Pl. XXXII, fig. 13.
? 2002 Mesohibolites platyurus (); et al., p. 79-80, Pl. VIII, fig. 5.
2005 Conohibolites escragnollensis (); & , p. 70, 72, Pl. IV, figs. 1-2; Pl. V, figs. 6-7 (cum synonymy).
Material: Eight specimens (ABSS) and five specimens (BOU).
Stratum: ANG151.3 to ANG157.2.
Description: A rather robust, typical conical to cylindro-conical rostrum with a deep alveolus and a rather short rostrum solidum with a very elongated apex. Cross-sections are typically heart-shaped, but change to well rounded outlines in the alveolar part.
Remarks: In Co. tzankovi the rostrum solidum is smaller and the apical tapering is more abrupt as compared to the nominal species. (1841) indicated in his description that fig. 1 is typical. However, (1970a) indicated fig. 4 to be the LT and subsequently et al. (2002) indicated fig. 1 to be the LT.
A certain resemblance to large (mature to gerontic) Co. gladiiformis or Co. garshini exists. However, the latter species show dorso-ventrally flattened outlines in the alveolar part of their rostrum. Moreover, the apical area is much more extended and dorso-ventrally flattened in Co. gladiiformis but much less in Co. garshini. The latter shows a minaret-like apical part, more or less comparable to S. simionescui, and has a much larger rostrum solidum.
Phylogeny: Probably descendant from Co. garshini.
Stratigraphic occurrence: Earliest Late Barremian B. barremense Subzone.
Belemnite association: BaBA4a (FO) and BaBA4b (LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Georgia (?) and Hungaria.
Conohibolites cf. trastikensis (, 1965)
Pl. 9 ![]() ,
figs. 17-18, 19-20
,
figs. 17-18, 19-20
+ 1964 Mesohibolites trastikensis sp. nov. , p. 139, 146 (nom. nud.).
+ 1964 Mesohibolites trastickensis sp. nov. , p. 144 (nom. nud.).
* 1965 Mesohibolites trastikensis sp. nov. , p. 157-158, Pl. III, figs. 1 [HT by OD], 2, 3.
+ 1965 Mesohibolites trastickensis sp. nov. , p. 168 (lapsis calami).
non 1975 Mesohibolites trastikensis ; & , pp. 138-140, Pl. I, fig. 1.
non 2002 Mesohibolites trastikensis ; et al., p. 77-78, Pl. VIII, figs. 2, 3 (= & , 1975).
Material: Eight incomplete to badly preserved (partially dissolved) specimens.
Stratum: ANG165.2 to ANG167.
Description: A robust, typical conical to cylindro-conical, tapering rostrum with a near minaret-like apex. Cross-sections are subrounded, slightly dorso-ventrally depressed. The alveolus reaches about a third to halfway the total length of the rostrum, as does the alveolar groove.
Remarks: In Co. tzankovi the rostrum solidum is smaller and the apex tapers much more as compared to the nominal species. In Co. platyurus the rostrum has a well-rounded outline in the alveolar opening, and the apical area typically tapers into an extended, elongated pointed apex.
Phylogeny: Probably descendant from Co. garshini.
Stratigraphic occurrence: H. feraudianus Subzone.
Belemnite association: BaBA4c (FO and LO).
Geographical occurrence: Bulgaria and France (Vocontian Basin).
Conohibolites sp. (teratological?)
Pl. 7 ![]() ,
figs. 25-26
,
figs. 25-26
Material: One specimen.
Stratum: ANG166.2 to ANG167.
Description: A relative short conical belemnite, which differs from all other Mesohibolitidae mentioned herein by a well-defined laterally compression of the rostrum solidum. The alveolar groove is very short and the alveolus is likewise shallow.
Remarks: As this kind of outer-morphology is unknown from any Mesohibolitidae, except for teratological specimens, it is believed to be a misshapen specimen too.
Stratigraphic occurrence: H. feraudianus Subzone.
Belemnite association: BaBA4c (single occurrence).
Geographical occurrence: France (Vocontian Basin).
Genus Shvetsovia gen. nov. (for description see above)
Shvetsovia elegantoidea (, 1965)
Pl. 6 ![]() ,
figs. 15-16, 23-24; Pl. 8
,
figs. 15-16, 23-24; Pl. 8 ![]() ,
figs. 7-8, 9-10, 23-24, 26-27;
,
figs. 7-8, 9-10, 23-24, 26-27;
Pl. 9 ![]() ,
figs. 7-8, 11-12, 21-22, 25-26; Pl. 16
,
figs. 7-8, 11-12, 21-22, 25-26; Pl. 16 ![]() ,
figs. 1-2, 3-4, 5-6
,
figs. 1-2, 3-4, 5-6
pt 1883 Belemnites minaret ; , Pl. I, fig. 8, non fig. 9 (= Co. minaretiformis).
+ 1964 Mesohibolites elegantoides sp. nov. , p. 139, 144, 146 (nom. nud.).
* 1965 Mesohibolites elegantoides sp. nov. , p. 155-156, Pl. IV, figs. 1 [HT by OD], 3-4.
1970 Mesohibolites elegans ; , p. 106, Pl. XX, fig. 2.
? 1973 Hibolites jaculum ; , Pl. I, figs. 12-13.
? 1994 Mesohibolites elegantoides ; et al, p. 82, Pl. 27, figs. 3-4.
non 2002 Mesohibolites elegantoides ; et al., p. 100-101, Pl. XIII, figs. 4-5.
Material: Forty specimens (ABSS) and over fifty from BOU.
Stratum: ANG153.2 to ANG166.2-167.
Description: Both in ventral as well in lateral view the rostra have a more or less fusiform outline. Mature to gerontic specimens do show a less well pronounced fusiform outline in lateral view. The maximal outline of the cross-sections is situated towards the apical part. The alveolar part shows a well-rounded outline. The alveolus is rather short, about one-fifth of the total length of the rostrum. The alveolar groove reaches onto the rostrum solidum, occupying about one-third of the total length of the rostrum. At the transition from the rostrum cavum towards the rostrum solidum, the outline of the rostrum changes to ventro-dorsally flattened cross-sections. This flattening is maintained throughout the larger part of the rostrum solidum, except for the apical most part. The apex is pointed and orientated slightly to the ventral side. This relates to the more pronounced incurvation of the dorsal side as compared to the ventral side in lateral view.
Remarks: Some fragmentary specimens from the marly bed between ANG157.1-2 and in younger strata can not be grouped with certainty within the nominal species. These could equally well belong to immature specimens of Co. ex gr. minaretiformis. They differ from M. (?) renngarteni , 1939, by a clearly more ventro-dorsally compressed rostrum. M. (?) renngarteni is even more spindle-like, has a longer alveolar groove, and a rounded to slightly laterally compressed alveolar part of the rostrum. Moreover, the apex is almost mucronate.
Stratigraphic occurrence: In the upper part of the B. barremense Subzone and disappears in the H. feraudianus Subzone. The material from Bulgaria originates from the Barremian, except for some material from the Bedoulian of Opaka. The latter most probably relates to M. (?) elegans (, 1913).
Belemnite association: BaBA4b (FO) and BaBA4c (LO).
Geographical occurrence: Bulgaria, Czech Republic (?), France (Vocontian Basin), Georgia, and Slovakia (?).
Shvetsovia aff. elegantoidea (, 1965)
Pl. 9 ![]() ,
figs. 3-4
,
figs. 3-4
Material: Eleven specimens, varying from near complete to partially preserved specimens.
Stratum: Beds ANG165.1 to ANG166.2-167.
Remarks: Comparable to S. elegantoidea but with a clearly less-well developed alveolar groove, a more robust appearance and a more ventro-dorsally compressed rostrum solidum. Also, the alveolar part is dorso-ventrally compressed.
Stratigraphic occurrence: In the "middle" to lower upper part of the H. feraudianus Subzone.
Belemnite association: BaBA4c (FO and LO).
Geographical occurrence: France (Vocontian Basin).
Shvetsovia ex gr. elegantoidea (, 1965)
Pl. 9 ![]() ,
figs. 31-32, 33-34
,
figs. 31-32, 33-34
Material: Three specimens, one immature is complete while the other two are more mature but incomplete.
Stratum: Beds ANG167-168 to ANG168-169.1.
Remarks: Well-flattened, rather robust, spindle-shaped rostrum with a fairly long alveolar groove relative to the depth of the alveolus. The latter is very shallow and also ventro-dorsally compressed. The apical part is pointed.
Stratigraphic occurrence: The upper part of the H. feraudianus Subzone to the lowermost part of the I. giraudi Zone.
Belemnite association: BaBA4c (FO in topmost part) and BaBA5a (LO and common in lowermost part).
Geographical occurrence: France (Vocontian Basin).
Shvetsovia sp. 1 nov. ?
Pl. 9 ![]() ,
figs. 1-2
,
figs. 1-2
? + 2003 Hibolites irregularis : & , p. 40 (nom. null.).
Material: Three specimens, of which is only one almost completely preserved.
Stratum: Beds ANG165.2-166.1 to ANG166.1.
Remarks: Elongated spindle-like rostrum, with a relative long alveolar groove in relation to the depth of the alveolus. The latter is very shallow and laterally compressed. The rostrum becomes more rounded to slightly ventro-dorsally compressed towards the rostrum solidum. The apical part is pointed and elongated. The general morphology is reminiscent of S. carpatica. However, the latter is restricted to the boundary beds between the Early and Late Barremian. Moreover, it is much more spindle-like, with a much better expressed "point-of-widest-section", i.e., the rostrum solidum is almost straight.
Stratigraphic occurrence: In the “middle" part of the H. feraudianus Subzone.
Belemnite association: BaBA4c (FO and LO).
Geographical occurrence: France (Vocontian Basin) and Georgia (?).
Family Duvaliidae , 1914
Genus Duvalia , 1878
Duvalia grasiana (, 1841)
Pl. 5 ![]() ,
figs. 6, 9-10, 11-12, 19-20
,
figs. 6, 9-10, 11-12, 19-20
1841 Belemnites Grasianus , p. 63 (pars), Pl. 7 ![]() ,
figs. 1, 2, 3, 4-5 (?).
,
figs. 1, 2, 3, 4-5 (?).
1842 Belemnites pseudo-dilatatus Nob. , p. 45 (nomen oblitum; nom. dub.).
non 1994 Duvalia grasiana (); et al., Pl. 30, figs. 3-4, 5-6 (= D. vermeuleni sp. nov.).
2002 Duvalia grasiana (); et al., p. 158-160, Pl. XXII, figs. 4-6 (pars cum synonymy).
non 2005 Duvalia grasiana (); & , Pl. III, figs. 38-39; Pl. V, figs. 18-19, 25-26 (= D. vermeuleni sp. nov.).
2008 Duvalia grasiana (); , p. 607, 615, 617, 619, Text-fig. 3K-L.
Material: Twenty-five specimens.
Stratum: Beds ANG166.2-167 to COM137 (top part of bed).
Description: Latatoid rostrum with a long alveolar groove and a deep alveolus. The cross-sections are ellipsoidal and laterally flattened. The dorsal and ventral sides are more or less parallel but for the apical part. The apex is placed to the dorsal side. As a result the dorsal side is less curved in the apical part, compared to the ventral side. The dorsal alveolar groove generally ends near the apical area. The deep alveolus reaches about half-way the rostrum.
Remarks: In the G. sartousiana Zone specimens intermediate between D. vermeuleni and D. grasiana occur. An immature specimen (see Pl. 5 ![]() ,
figs. 19-20) is very reminiscent of D. silesiaca.
,
figs. 19-20) is very reminiscent of D. silesiaca.
Phylogeny: Descendant from Duvalia vermeuleni sp. nov.
Belemnite association: BaBA4c (FO) to BdBA4 (LO).
Stratigraphical distribution: They first occur in the H. feraudianus Subzone, but are most common in the I. giraudi and M. sarsini subzones of the I. giraudi Zone. The apparent absence, or at least relative rarity, of this species in the Barremian-Bedoulian boundary beds is striking. The last specimens occur in the condensed (?) bed on top of COM137 (Aptian; Dufrenoya furcata Zone or Epicheloniceras debile Subzone).
Geographical distribution: Circum Mediterranean Tethys, Mexico, eastern Africa, and in the Boreal-Atlantic Provence (northern Germany).
These associations are characterized by the occurrence of Mesohibolites (FO in BaBA5), the absence of Conohibolites, while Shvetsovia last occurs in the lowermost part of BaBA5. The FO of Neohibolites is in BdBA1. Especially in BaBA7, the diversity among the belemnites is rather low.
Family Mesohibolitidae , 1983
Genus Mesohibolites ,
1919
emend. & ,
2005 nomen conservandum
Type species: Hibolites uhligi , 1913 (see & , 2005, p. 59, 63).
Remarks: The genus is characterized by mature specimens with cylindro-conical outlines, both in lateral as well as in ventral view. In these species the immature specimens show a spindle-like outline in ventral view. However, there are some exceptions, viz. the specimens around Mesohibolites renngarteni , 1939, Mesohibolites elegans (, 1913), and the Georgian belemnites related to "Hibolithes" ex gr. horeshaensis-inguriensis , 1938. Therefore, these specimens are attributed with some doubt to the nominal genus. Especially, the latter two probably belong to Neohibolites, or to a new genus. They can almost certainly not belong to Hibolithes de , 1808. The latter genus was already extinct in the Tethyan Realm and only endemic, dwarfed specimens (Hibolithes minutus , 1935) survived in the Atlantic-Boreal Province, up to the Barremian-Bedoulian boundary beds (also see juvenile Mesohibolites).
Generally, Mesohibolites differs from Shvetsovia by the absence of spindle-like lateral views in mature specimens (but see above for exceptions). In general, Conohibolites is shorter with a less elongated apical area. Moreover, the latter shows much more ventro-dorsally compressed cross-sections, generally throughout its rostrum. The development of the apical area appears to be somewhat different. In general, in Conohibolites it is characterized by a sudden kink in the outline of the rostrum solidum, giving way to the apical area. In Mesohibolites the apical area appears to be developed in a much more gradual way, in relation to the rostrum solidum. The alveolus is displaced to the dorsal side in Mesohibolites, while being centrally placed in Shvetsovia and Conohibolites. Juvenile specimens of Shvetsovia are rather comparable in morphology, but tend to be more clavate-like, while juvenile Conohibolites tend to be more elongated and even more dorso-ventrally flattened.
The occurrence of small, probably juvenile specimens - at least herein interpreted as being such - with the first occurrence of the genus Mesohibolites is interesting. Probably these specimens point to a relation between the genera Mesohibolites and Neohibolites, the latter being related to the stratigraphically older genus by neoteny.
Range: Late Barremian - early Bedoulian (I. giraudi Zone to D. deshayesi Zone).
Phylogeny: Most probably descendant from Co. ex gr. minaretiformis.
Belemnite association: BaBA5 (FO) and BdBA3 (LO).
Occurrence: Circum Mediterranean Tethys.
Mesohibolites anglesensis sp. nov.
Pl. 10 ![]() ,
figs. 11-12, 15-16; Pl. 13
,
figs. 11-12, 15-16; Pl. 13 ![]() ,
figs. 27-28
,
figs. 27-28
? +1968 Mesohibolites warians [sic!] ; , p. 635.
? 1975 Mesohibolites minaret (); & , p. 136.
? 1994 Mesohibolites ex gr. minaret (); et al., p. 83, Pl. 26, figs. 6-7 (gerontic?).
pt 2002 Mesohibolites minaret (); et al., Pl. XI, fig. 6 (not the synonymy).
Derivation of name: Named after the hamlet of Angles.
Material: Thirteen specimens.
Holotype: RGM 583631 (ANG179.3; Pl. 10 ![]() ,
figs. 11-12).
,
figs. 11-12).
Paratype: RGM 583617 (ANG178.2-179; Pl. 10 ![]() ,
figs. 15-16).
,
figs. 15-16).
Type stratum: Martelites sarasini Subzone.
Stratum: Beds ANG175.2-176.1 to ANG186-187.
Description: A medium sized, tapering down to the apex, belemnite. In both ventral as well as in lateral view it is almost conical. The rostrum is dorso-ventrally compressed, except for its apical-most part. This part shows round cross-sections. The alveolar groove reaches approximately half-way the rostrum, while the alveolar cavity is slightly less deep. The apex is very pointed. The alveolar side is almost straight, with only a slight curvation to the apex. As a consequence, the apex is shifted towards the ventral side.
Remarks: Immature specimens of M. beskidensis are cylindro-conical and more slender. Also M. longus is cylindro-conical but much more slender, while M. georgicus is more robust, i.e., with a less elongated apical area.
Phylogeny: Probably descendant from Co. ex gr. minaretiformis.
Stratigraphic occurrence: Topmost part of the I. giraudi Subzone to M. sarasini Subzone.
Belemnite association: BaBA6a (FO and LO).
Geographical occurrence: Georgia (Abkhazia), France (Vocontian Basin), and Slovakia (?).
Mesohibolites beskidensis (, 1883)
Pl. 10 ![]() ,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 13-14, 17-18, 19-20
,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 13-14, 17-18, 19-20
+ 1883a Belemnites Beskidensis n. sp. , p. 87 (nom. nud.).
1883b Belemnites Beskidensis n. sp. , p. 177-178, Pl. I, fig. 3 [LT].
non 1898 Belemnites beskidensis ; , p. 107 (51)-108 (52), Pl. I, fig. 4 (= Co. minaretiformis).
non 1939 Mesohibolites beskidensis ; , p. 16-17, Pl. III, figs. 2-4 (= M. georgicus).
non 1952 Mesohibolites beskidensis ; , p. 77, Pl. IV, figs. 2-3 (=? Co. minaretiformis).
non 1960 Mesohibolites beskidensis ; , p. 359, Pl. II, fig. 6 (= M. georgicus).
? pt 1970a Mesohibolites beskidensis (); , p. 28-29, Pl. XII, fig. 8 (from Golyamo Novo, Targovishko - Barremian), non figs. 5-7 (from Opaka - Bedoulian).
non 1973 Mesohibolites beskidensis ; , p. 151, Pl. XLVIII, fig. 2a-c (= M. ekimbontchevi).
non 1973 Mesohibolites beskidensis (); , p. 33-34, Pl. 3, figs. 4-6; Pl. 10, figs. 1-2.
non 1976 Mesohibolites sp. ex gr. M. beskidensis (); & , p. 140 (6), 146 (12) (= Adiakritobelus).
1978a Mesohibolites cf. gladiiformis (); , p. 14-16, Pl. II, fig. 5.
1978a Mesohibolites beskidensis (); , p. 19, Pl. II, fig. 2 (= , 1883b: Pl. I, fig. 3).
non 2002 Mesohibolites beskidensis (); et al., p. 96-97, Pl. XII, figs. 4-5 (= M. georgicus).
Material: Twenty-six specimens.
Stratum: Beds ANG168 (in top of bed) to ANG174.2-175.1.
Description: Rather large elongated rostrum with cylindro-conical outlines, both in ventral as well as in lateral view. However, the ventral outline is more spindle-like in more mature specimens. The alveolar groove is long, approximately up to the point in which the rostrum has its maximum outline, and the alveolar cavity occupies approximately 1/3 of the total length of the rostrum. The rostrum cavum is laterally compressed, being more pronounced in the more mature specimens. The rostrum solidum is dorso-ventrally compressed, except for its apical most part. The latter has round cross-sections. The apex is pointed and displaced slightly to the ventral side.
Remarks: LT (, 1883b, Pl. I, fig. 3) selected by (1970a). The other figured specimen (fig. 7) was attributed to Mh. uhligi by (1913), which was subsequently followed by other researchers.
Phylogeny: Most probably descendant from Co. ex gr. minaretiformis.
Stratigraphic occurrence: Restricted to the I. giraudi Subzone (BaBA5), and being most abundant in the I. giraudi-Horizon. The specimen described by originates from the upper part of the Hradiště-Schichten. This formation is of Barremian to Bedoulian age (cf. , 1978b, p. 118).
Belemnite association: BaBA5 (FO, common, and LO).
Geographical occurrence: Bulgaria, Czech Republic, France (Vocontian Basin). It appears to be restricted to the western part of the Mediterranean Tethys.
Mesohibolites bulgaricus , 1965
Pl. 13 ![]() ,
figs. 3-4 (?), 5-6, 11-12, 13-14, 23-24
,
figs. 3-4 (?), 5-6, 11-12, 13-14, 23-24
* 1965 Mesohibolites longus bulgaricus n. sp. , p. 160-161, Pl. V, figs. 1 [HT by OD]-2; Pl. VII, fig. 1.
pt 1973 Mesohibolites uhligi ; , p. 151, Pl. XLIX, figs. 9-10.
non 1994 Mesohibolites longus bulgaricus ; et al., p. 83, Pl. 27, figs. 1-2 [= M. longus ()].
2002 Mesohibolites longus bulgaricus ; et al., p. 91-92, Pl. XI, fig. 2 (cum synonymy).
Material: Twenty-nine specimens.
Stratum: ANG198 to ANG209.
Description: Elongated to moderately robust cylindro-conical rostrum, with weak dorso-ventrally compressed rostrum solidum and rounded rostrum cavum. Maximum outline, but almost not spindle-like, near end of alveolar groove. The alveolus occupies approximately half the length of the alveolar groove. It is clearly less compressed as M. uhligi and more robust as compared to M. longus.
Remarks: As the specimen appears throughout the Mediterranean Tethys there seems to be no reason to distinguish between geographical species or subspecies. In our material, a quite slender elongated specimen occurs that is much like M. longus (Pl. 13 ![]() ,
figs. 3-4).
,
figs. 3-4).
Phylogeny: Probably descendant of M. ekimbontchevi.
Stratigraphic occurrence: Barremian-Bedoulian boundary sediments to early Bedoulian (Pa. oglanlensis Zone).
Belemnite association: BdBA1 (FO, common and LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Georgia, and Ukraine (Crimea).
Mesohibolites ekimbontchevi , 1965
Pl. 13 ![]() ,
figs. 21-22; Pl. 15
,
figs. 21-22; Pl. 15 ![]() ,
figs. 1-2, 3-4, 13-14
,
figs. 1-2, 3-4, 13-14
+ 1964 Mesohibolites ekimbontchevi sp. nov. , p. 140, 146 (nom. nud.).
* 1965 Mesohibolites ekimbontchevi sp. nov. , p. 159-160, Pl. VI, figs. 1 [HT by OD]-2; Pl. VII, fig. 5.
1970a Mesohibolites ekimbontchevi ; , p. 38-39, Pl. XX, figs. 1-5.
1973 Mesohibolites beskidensis ; , p. 151, Pl. XLVIII, fig. 2a-c.
? 1973 Mesohibolites ekimbontchevi ; , p. 42-43, Pl. 4, figs. 12-14; Pl. 11, figs. 5-6.
non 1989 Mesohibolites ekimbontchevi ; , p. 28, text-fig. d (teratological; =? M. uhligi).
1998 Neohibolites ekimbontchevi ; & , Pl. I, fig. 8.
2002 Mesohibolites ekimbontchevi ; et al.., p. 98-99, Pl. XIII, fig. 1.
Material: Eleven specimens.
Stratum: ANG194 to ANG205 (i.e., the Barremian-Bedoulian boundary beds).
Description: A rather robust cylindro-conical (lateral view) to slightly spindle-shaped (ventral view), dorso-ventrally compressed rostrum. The pointed apex is centrally placed. The alveolar groove is well-pronounced, while the alveolar cavity is rather shallow.
Remarks: In all the mature to gerontic specimens (Pl. 15 ![]() ,
figs. 1-2, 13-14) the alveolar part of the rostrum is not preserved. The other specimen is probably a teratological one (Pl. 15
,
figs. 1-2, 13-14) the alveolar part of the rostrum is not preserved. The other specimen is probably a teratological one (Pl. 15 ![]() ,
figs. 3-4). Overall, the specimens are much like mature to gerontic M. uhligi (Pl. 14
,
figs. 3-4). Overall, the specimens are much like mature to gerontic M. uhligi (Pl. 14 ![]() ,
figs. 1-2) but are characterized by a longer apical part of the rostrum. Also, the dorso-ventral compression is less, as compared to M. uhligi.
,
figs. 1-2) but are characterized by a longer apical part of the rostrum. Also, the dorso-ventral compression is less, as compared to M. uhligi.
Phylogeny: Probably descendant from M. uhligi.
Stratigraphic occurrence: Latest Barremian (topmost Pseudocrioceras waagenoides Subzone).
Belemnite association: BdBA1 (FO and LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Georgia, Romania, and Ukraine (Crimea).
Mesohibolites fallauxi (, 1883)
Pl. 16 ![]() ,
figs. 11-12, 15-16, 25-26
,
figs. 11-12, 15-16, 25-26
+ 1883a Belemnites Fallauxi n. sp. , p. 187 (nom. nud.).
1883b Belemnites Fallauxi n. sp. , p. 177 (53), Pl. I, figs. 4 [LT], 13 (teratological), 14 (?).
? pt 1970a Mesohibolites minaret (); , Pl. VIII, figs. 1-4, non figs. 5-6.
? 1970a Mesohibolites ex gr. minaret (); , Pl. IX, figs. 1-6.
1973 Mesohibolites fallauxi ; , p. 152, Pl. XLVIII, figs. 8-9.
2002 Mesohibolites fallauxi (); et al., p. 95-96, Pl. XII, fig. 1 (cum synonymy).
Material: Four specimens.
Stratum: ANG225-226 (= ANG116-117) to ANG231, and one loose specimen from the topmost part of the calcareous section to the east, off the road to Angles. The latter beds correspond to COM122-129 (= ANG231-238).
Description: The specimen is quite robust and characterized by a dorso-ventrally well-flattened rostrum solidum, and a dorso-ventrally depressed rostrum cavum. The alveolar groove is well-visible, broad and relatively long, approximately reaching half-way the rostrum. The alveolus is rather deep. The maximum outline is more or less in the middle part of the rostrum, but mature specimens tend to become almost cylindro-conical. However, the general outline is more spindle-like than cylindro-conical.
Remarks: LT (, 1883b: Pl. I, fig. 4) selected by , 1970a. The specimens herein named S. ex gr. elegantoidea are morphologically comparable but show a very shallow alveolus, a less pronounced alveolar groove, and a less robust rostrum solidum.
Phylogeny: Probably descendant from M. georgicus.
Stratigraphic occurrence: Top part of D. forbesi Zone to lower part of D. deshayesi? Zone.
Belemnite association: BdBA3 (FO and LO).
Geographical occurrence: Bulgaria, Czech Republic, France (Vocontian Basin), Georgia, Russia (Kabardino-Balkaria ASR), and Ukraine (Crimea).
Mesohibolites georgicus (, 1973)
Pl. 16 ![]() ,
figs. 17-18a, 19-20, 23-24;
,
figs. 17-18a, 19-20, 23-24;
Pl. 17 ![]() ,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 11-12
,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 11-12
? 1952 Mesohibolites semicanaliculatus (); , Pl. VIII, figs. 1-2; Pl. IX, figs. 1-2.
? 1960 Mesohibolites uhligi ; , p. 359, Pl. II, fig. 4.
pt 1973 Mesohibolites uhligi ; , p. 151, Pl. XLVIII, fig. 1a-c.
1973 Mesohibolites uhligi georgicus , p. 32-33, Pl. 3, figs. 1-3; Pl. 9, figs. 13-14 (cum synonymy).
2002 Mesohibolites uhligi georgicus ; et al., p. 85-86, Pl. IX, fig. 6; Pl. X, fig. 1; Pl. XIV, fig. 6 (pars cum synonymy).
Material: Thirty-two specimens.
Stratum: Beds ANG225-226 to COM127 (= ANG236).
Description: Rather robust, cylindro-conical, slightly dorso-ventrally depressed specimen with a pointed to blunt apex. The latter is more or less centrally placed, or shifted slightly to the ventral side. Also, the alveolar area is slightly dorso-ventrally depressed. The alveolar groove is rather pronounced, occupying approximately 1/3 of the length of the rostrum, or less. In general, the depth of the alveolus is comparable to the length of the alveolar groove.
Remarks: The specimen shown by (1973) is indicated to be the HT by et al (2002). However, this is not the original designation, nor the only specimen available. In fact a LT has to be indicated, as originally no type was indicated. As the specimen appears throughout the Mediterranean Tethys there seems to be no reason to distinguish between geographical species or subspecies.
Phylogeny: Probably descendant from the specimens herein named M. nalchikensis?
Stratigraphic occurrence: In the ABSS restricted to the D. forbesi and D. deshayesi zones.
Belemnite association: BdBA3 (FO, common, and LO).
Geographical occurrence: Bulgaria, Georgia, France (Vocontian Basin), Russia (Kabardino-Balkaria ASR), Slovakia, and Ukraine (Crimea).
Mesohibolites longus (, 1913)
Pl. 13 ![]() ,
figs. 1-2, 9-10, 19-20
,
figs. 1-2, 9-10, 19-20
1913 Hibolites longus n. sp. , pp. 59-60, 70, Pl. V, figs. 1a-b [LT], 1c-f, 8b.
1931 Hibolites longus (); , p. 127-128, Pl. I, figs. 2-3.
1994 Mesohibolites longus bulgaricus ; et al., p. 83, Pl. 27, figs. 1-2.
2000 Mesohibolites sp.; , p. 177-178, Text-fig. 1a-b.
2002 Mesohibolites longus longus (); et al., p. 90-91, Pl. X, fig. 5; Pl. XI, fig. 1 (cum synonymy).
Material: Ten specimens.
Stratum: ANG178.2-179.1 to ANG181-182.
Description: Cylindro-conical, elongated rostrum with a relative long alveolar groove, but a relative short alveolar cavity. In ventral view specimens often show a spindle-like outline. Overall, the rostrum has rounded to slightly dorso-ventrally compressed cross-sections, except for its alveolar part. Herein, often a feeble lateral compression occurs. The apical part is elongated (stretched) and shows a pointed apex.
Remarks: Lectotype chosen by
(1972). The specimen appears throughout the Mediterranean Tethys thus there seems to be no reason to distinguish between geographical species or subspecies. The specimens of
(2000) originated from the beds 45 and 48, i.e., the boundary beds of the M. sarasini and the Pc. waagenoides
subzones. It is quite well comparable to the specimen figured by us as M. ex gr. longus (Pl. 14 ![]() ,
figs. 3-4) but appears to be much less dorso-ventrally compressed.
,
figs. 3-4) but appears to be much less dorso-ventrally compressed.
Phylogeny: Probably descendant from M. anglesensis.
Stratigraphic occurrence: M. sarasini Subzone (Upper Barremian).
Belemnite association: BaBA6a (FO and LO).
Geographical occurrence: Azerbaijan, Bulgaria, France (Vocontian Basin), Georgia, and Slovakia.
Mesohibolites ex gr. longus (, 1913)
Pl. 13 ![]() ,
figs. 15-16; Pl. 14
,
figs. 15-16; Pl. 14 ![]() ,
figs. 3-4
,
figs. 3-4
pt 1913 Hibolites longus n. sp. , Pl. V, fig. 1e-f.
Material: Five specimens.
Stratum: ANG189 to ANG196.2-197.
Description: Cylindro-conical specimens with an elongated rostrum. Mature and immature specimens are characterized by a very strong flattening (dorso-ventral compression) of the rostrum. The apex is shifted to the ventral side. The alveolar groove is quite long and fades away approximately halfway the rostrum. The alveolus is quite shallow, approximately 1/5 of the length of the rostrum.
Remarks: Much more dorso-ventrally compressed rostrum as compared to M. longus, and a stronger elongation as compared to M. uhligi. Much more elongated as compared to M. bulgaricus. It is probably a new species.
Phylogeny: Probably descendant from M. longus.
Stratigraphic occurrence: Pc. waagenoides Subzone (latest Barremian).
Belemnite association: BaBA6b (FO) and BaBA7 (LO).
Geographical occurrence: France (Vocontian Basin) and Georgia (Abkhasia).
Mesohibolites nalchikensis? , 1939
Pl. 15 ![]() ,
figs. 5-6
,
figs. 5-6
+ 1937 Mesohibolites cf. nalcikensis sp. nov. (); , p. 40 (nom. nud.).
* 1939 Mesohibolites nalčikensis sp. nov. , p. 18-19, Pl. IV, fig. 3 [HT by OD], 4.
2002 Mesohibolites nalchikensis ; et al., p. 86-87, Pl. X, fig. 3 (cum synonymy).
Material: Five specimens and one loose specimen.
Stratum: ANG210 to ANG218-219 (= 222-223).
Description: Relative small, cylindro-conical (lateral view) to conical (ventral view) specimens with slightly, laterally depressed rostrum cavum. The alveolar groove is relative long, while the alveolar cavity is rather shallow. In both mature (not figured) and immature specimens, the guard tapers down to the apex. The rostrum solidum is dorso-ventrally depressed well on to the apical area.
Remarks: The specimens are attributed with doubt to the nominal species. Except for one mature specimen all others are probably immature. They are best comparable to (1939: Pl. IV, fig. 4).
Phylogeny: Probably descendant from M. bulgaricus.
Stratigraphic occurrence: early Aptian, the D. forbesi Zone.
Belemnite association: BdBA2 (FO and LO).
Geographical occurrence: France (?, Vocontian Basin), Georgia, and Russia (Kabardino-Balkaria ASR).
Mesohibolites (?) renngarteni , 1939
Pl. 11 ![]() ,
figs. 1-2, 3-4, 5-6, 11-12, 13-14, 15-16
,
figs. 1-2, 3-4, 5-6, 11-12, 13-14, 15-16
*1939 Mesohibolites renngarteni sp. nov. , p. 18, Pl. IV, figs. 1, 2 [HT by OD].
+ 1952 Mesohibolites rengarteni ; , p. 78, Pl. VI, figs. 1-2.
non 1960 Mesohibolites renngarteni ; , p. 359, Pl. II, fig. 5 (= M. renngarteni caucasicus ).
1970 Hibolites subfusiformis inflata ; , p. 105, Pl. XX, fig. 1 (from the I. giraudi Zone).
? pt 1970a Hibolites krimholzi sp. nov. , p. 18-19, 67-68, Pl. I, figs. 1 (?), 3, non Pl. I, fig. 2.
non 1972 Mesohibolites renngarteni ; , p. 148-149, text-fig. 50, Pl. VII, fig. 3 (=M. renngarteni caucasicus ).
non 1973 Neohibolites renngarteni ; , Pl. I, figs. G-I (= M. (?) elegans).
2000 "Hibolithes" nov. sp. 1 , Pl. 3 ![]() , fig. 2a-b (from the I. giraudi Zone).
, fig. 2a-b (from the I. giraudi Zone).
non 2000 "Mesohibolites" renngarteni renngarteni ; , Pl. 3 ![]() , fig. 1a-b (=
M. (?) elegans).
, fig. 1a-b (=
M. (?) elegans).
2002 Mesohibolites renngarteni renngarteni ; et al., p. 87-88, Pl. X, fig. 2 (cum synonymy).
Material: Twenty-one specimens.
Stratum: Beds ANG169.1-2 to ANG176.1-2 (and a loose specimen from ANG176.2-179).
Description: This species shows a spindle-like development, both from ventral as well from lateral view. However, the spindle-like development is best seen in ventro-dorsal view. The alveolar groove is well-developed, running towards the point of maximal width of the rostrum solidum, but generally not reaching this point. The alveolus is rather short, reaching approximately halfway the length of the alveolar groove. The alveolar part is rounded or shows a slight lateral compression. However, the rostrum solidum is slightly dorso-ventrally flattened and culminates in a centrally placed pointed, almost mucronate apex. This dorso-ventral flattening is not as pronounced as in S. elegantoidea.
Remarks: Differs from more or less comparable species like S. elegantoidea by a less ventro-dorsally compressed rostrum. According to (1939) typical M. renngarteni appear to have a rostrum-solidum with well-rounded cross-sections. In our material these specimens occur too. As the specimen appears throughout the Mediterranean Tethys there seems to be no reason to distinguish between geographical species or subspecies.
Phylogeny: Unknown, either descendant from S. ex gr. elegantoidea or M. beskidensis.
Stratigraphic occurrence: The Late Barremian I. giraudi Zone and the base of the M. sarasini Zone.
Belemnite association: BaBA5 (FO and common) and probably BaBA6a (LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Georgia, and Russia (Kabardino-Balkaria ASR).
Mesohibolites uhligi (, 1913)
Pl. 11 ![]() ,
figs. 7-8, 9-10, 19-20; Pl. 13
,
figs. 7-8, 9-10, 19-20; Pl. 13 ![]() ,
figs. 17-18;
,
figs. 17-18;
Pl. 14 ![]() ,
figs. 1-2, 3-4, 5-6, 9-10, 11-12, 13-14, 15-16; Pl. 15
,
figs. 1-2, 3-4, 5-6, 9-10, 11-12, 13-14, 15-16; Pl. 15 ![]() ,
figs. 11-12
,
figs. 11-12
? 1883b Belemnites beskidensis n. sp. , Pl. I, fig. 7.
1913 Hibolites Uhligi n. sp. , p. 55-56, 69, Pl. IV, fig. 6a-c, d-e [LT], f-n; Pl. V, fig. 8a.
non 1970 Mesohibolites uhligi ; , p. 105, Pl. XX, fig. 3 (= M. georgicus).
non 1973 Mesohibolites uhligi ; , p. 151, Pl. XLVIII, fig. 1a-c (= M. georgicus).
non 1973 Mesohibolites uhligi ; , p. 151, Pl. XLIX, figs. 9-10 (= M. bulgaricus).
1973 Mesohibolites uhligi (); , Pl. I, figs. P-R.
? 1994 Mesohibolites uhligi (); et al., p. 83, Pl. 26, figs. 8-9.
2002 Mesohibolites uhligi uhligi (); et al., p. 83-84, Pl. IX, fig. 5; Pl. XIV, figs. 4, 7 (pars cum synonymy).
Material: Forty specimens.
Stratum: Beds ANG174.2-175.1 to ANG193-194.
Description: Rather robust, spindle-shaped rostrum, both in lateral as well as in ventral view. In general, the alveolar opening is round but sometimes very slightly laterally compressed. The rostrum solidum is clearly dorso-ventrally compressed. The alveolar groove is well-visible and fades away approximately half-way the rostrum. The maximal outline of the rostrum is situated slightly below this point, but is situated approximately half-way along the rostrum. The alveolus occupies approximately half the length of the groove, or less, i.e., being relative shallow. The apex is pointed and centrally placed, and sometimes near mucronate.
Remarks: As the specimen appears throughout the Mediterranean Tethys there seems to be no reason to distinguish between geographical species or subspecies. LT (,
1913:
Pl. IV, fig. 6d-e) selected by et
al. (2002). In the upper part of its range, specimens occur that show a much more laterally compressed alveolar part of the rostrum (Pl. 14 ![]() , 17-18; Pl. 15
, 17-18; Pl. 15 ![]() ,
figs. 7-8, 19-20). They appear intermediate between M. uhligi and M. (?) aff. inguriensis. These specimens appear to be comparable to
(1989, text-fig. e - Mesohibolites schaoriensis ).
,
figs. 7-8, 19-20). They appear intermediate between M. uhligi and M. (?) aff. inguriensis. These specimens appear to be comparable to
(1989, text-fig. e - Mesohibolites schaoriensis ).
Phylogeny: Probably descendant from M. beskidensis.
Stratigraphic occurrence: The M. sarasini Subzone to Pc. waagenoides Subzone.
Belemnite association: BaBA6 (FO, common, and LO).
Geographical occurrence: Azerbaijan, Bulgaria, Czech Republic (?), Georgia, France (Vocontian Basin), and Slovakia.
? Mesohibolites sp. (juvenile)
Pl. 12 ![]() ,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 11-12, 13-14, 15-16, 17, 18-19, 20-21, 22-23, 26-27, 36-37
,
figs. 1-2, 3-4, 5-6, 7-8, 9-10, 11-12, 13-14, 15-16, 17, 18-19, 20-21, 22-23, 26-27, 36-37
Material: Forty specimens.
Stratum: Beds ANG171.1 to ANG196.1-2.
Description: Small (< 3 cm), generally clavate, Neohibolites-like belemnites with often a well-visible, but short alveolar groove. However, also specimens in which no alveolar groove can be seen occur. Often, lateral lines are clearly visible. In general, the apical part is more or less obtuse. The cross-sections are generally rounded, sometimes almost square-like and sometimes slightly dorso-ventrally depressed. Especially in the smallest specimens the cross-sections are near circular. Generally, the widest cross-sections are situated near the apical area. In the alveolar area, the specimens are either slightly laterally compressed or round to near round. Especially in the very small specimens (elongated morphs) there is almost no point in which a "widest cross-section" can be differentiated.
Remarks: These small belemnites do belong to the Mesohibolitidae. The only genus that occurs in the Tethys in sediments of this age is Mesohibolites. However, from a morphological point of view, these specimens do either relate to Neohibolites or to Hibolithes. There are several species in literature that can be compared to the above mentioned specimens, not only regarding the morphology but also their stratigraphical occurrences. In the Boreal-Atlantic Province occurs the small Hibolithes minutus , 1935. However, this species relates to Hibolithes jaculoides , 1937 ( & , 1980). It is regarded as endemic to the Barremian of east England and northwest Germany, disappearing in the Barremian-Bedoulian boundary beds, and, at least in Germany, never occurs together with Neohibolites ewaldi ( & , 2009, p. 284), but see & (1999, p. 283, 284).
From the Barremian-Bedoulian of the eastern Mediterranean Tethys, (1938) described the new species of belemnites "Hibolites bsibiensis, - horeshaensis, and - inguriensis", and included some small specimens depicted by (1913, Pl. V, fig. 6). The other specimen (, 1913, Pl. V, fig. 7) was later included in the synonymy of the new species "Hibolites lashensis" by (1999). Previously however, (1957, p. 49) included this specimen in his newly species "Neohibolites patara". Already (1913, p. 71) included these belemnites in his "jeunes rostres des Bedoulien" [sic]. (1973, p. 106 and chart between p. 86-87) and & (1975, p. 133) refer to some of these specimens from latest Barremian sediments (the I. giraudi Zone). However, overall, these species are mentioned from the Bedoulian ( et al., 2002, p. 71-75).
Stratigraphic occurrence: In the I. giraudi Zone (Latest Barremian).
Belemnite association: BaBA5 (FO) and BaBA7 (LO).
Geographical occurrence: France (Vocontian Basin), Georgia, and Russia (Kabardino-Balkaria ASR).
Mesohibolites (?) elegans (, 1913)
Pl. 15 ![]() ,
figs. 9-10, 15-16, 17-18
,
figs. 9-10, 15-16, 17-18
1913 Hibolites elegans n. sp. , p. 60-61, 70, Pl. V, figs. 3a-b [LT], 3c-f, 4g-h, 8c.
non 1939 Mesohibolites elegans ; , p. 20, Pl. IV, figs. 7-8 (= S. elegantoidea).
non 1949 Mesohibolites elegans (); , p. 261, Pl. LXXXI, figs. 5-6 (= S. elegantoidea).
non 1949 Mesohibolites elegans (); , Pl. LXXXI, fig. 7 (= , 1939: Pl. IV, fig. 8).
1960 Mesohibolites elegans ; , p. 360, Pl. II, fig. 2.
non 1970 Mesohibolites elegans ; , p. 106, Pl. XX, fig. 2 (from the I. giraudi Zone = M. (?) renngarteni).
1970a Mesohibolites elegans (); , p. 33-34, Pl. XI, figs. 1-7; Pl. XXXII, fig. 6.
? 1970a Neohibolites sp. , Pl. XXI, fig. 6.
1972 Mesohibolites elegans (); , p. 145-146, Pl. V, figs. 3-4.
1973 Mesohibolites elegans ; , p. 152, Pl. XLIX, fig. 2.
non 1973 Mesohibolites elegans (); , pp. 44-45, Pl. 5 ![]() ,
figs. 4-5; Pl. 10
,
figs. 4-5; Pl. 10 ![]() ,
figs. 7-8 (= S. elegantoidea).
,
figs. 7-8 (= S. elegantoidea).
? 1989 Mesohibolites (hibolites) elegans ; , p. 28, text-fig. f.
1994 Mesohibolites elegans (); et al., p. 81-82, Pl. 27, figs. 7-8.
2002 Mesohibolites elegans (); et al., p. 99-100, Pl. XIII, figs. 2-3 (pars cum synonymy).
2008 Mesohibolites elegans (); , p. 607, 615, 616, 617, 619, text-fig. 3I-J.
Material: Nine specimens, and (?) three specimens.
Stratum: ANG210 to ANG231-232, and (?) from ANG206 to ANG209.
Description: The specimens are spindle-shaped in both ventral as well as in lateral view, but more pronounced in ventral view. The maximum cross-section is shifted towards the apex and cross-sections are well to slightly dorso-ventrally compressed in the apical part while being laterally compressed in the alveolar part. Between the initiation of the alveolus and the point in which the thickest outline is reached, cross-sections change from laterally compressed to almost round to dorso-ventrally compressed. This area is rather wide in this species. The groove reaches approximately halfway the rostrum. It fades away before the point of maximum thickness. The alveolus is very shallow, being approximately 1/3 of the length of the alveolar groove. The apex is placed centrally and very pointed.
Remarks: LT (, 1913: Pl. V, fig. 3a-b) selected by , 1972. In the beds ANG206 to ANG209 specimens occur that do not show any sign of an alveolar groove but are otherwise comparable. They occur in slightly older beds as compared to the nominal species. Moreover, some specimens are difficult to separate from M. (?) ex gr. horeshaensis-inguriensis.
Phylogeny: Probably descendant of the specimens herein called M. (?) aff. inguriensis.
Stratigraphic occurrence: Bedoulian D. forbesi Zone to base of D. deshayesi? Zone.
Belemnite association: BdBA2 (FO) and BdBA3 (LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), Georgia, Russia (Kabardino-Balkaria ASR), Slovakia, Ukraine (Crimea), and northern Africa (Egypte).
Mesohibolites (?) horeshaensis (, 1938)
Pl. 16 ![]() ,
figs. 13-14
,
figs. 13-14
1938 Hibolites horeshaensis n. sp. , p. 154, 171, 187, Pl. II, fig. 4.
1952 Hibolites horeshaensis ; , p. 110-111, Pl. X, figs. 1-3.
+ 1952 Hibolites horeshiensis ; , p. 118.
1995 Parahibolites horeshaensis (); , p. 97.
2002 Hibolites horeshaensis ; et al., p. 73-74, Pl. VII, fig. 3.
Material: Six specimens.
Stratum: ANG225-226 to ANG231-232.
Description: Spindle-like, both from ventral as well as from lateral view, relative slim specimens with a long alveolar groove and a very shallow alveolus. The alveolar area is laterally compressed, while the apical most part is dorso-ventrally depressed. In between, the outlines of the cross-sections change, as in M. (?) elegans, from laterally compressed to dorso-ventrally compressed. The apex is centrally placed and rather pointed.
Remarks: The specimen figured by (1938) shows a hibolitoid-like morph, but our material is much more comparable to the specimens figured by (1952). There was no type-specimen chosen from the material originally studied by (1938), so a lectotype has to be chosen from his material.
Phylogeny: Probably relates to M. (?) elegans.
Stratigraphic occurrence: Bedoulian D. forbesi Zone to base of D. deshayesi? Zone.
Belemnite association: BdBA2 (FO) and BdBA3 (LO).
Geographical occurrence: France (Vocontian Basin) and Georgia.
Mesohibolites (?) inguriensis (, 1938)
Pl. 16 ![]() ,
figs. 7-8, 9-10, 21-22
,
figs. 7-8, 9-10, 21-22
1938 Hibolites inguriensis n. sp. , p. 154, 171, Pl. II, fig. 3.
+ 1938 Hibolites ingouriensis n. sp. , p. 186-187.
1973 Hibolites inguriensis ; , p. 25-26, Pl. 2 ![]() ,
figs. 3-4.
,
figs. 3-4.
1995 Neohibolites inguriensis (); , p. 94.
2002 Hibolites inguriensis ; et al., p. 71-72, Pl. VII, fig. 6.
Material: Five specimens.
Stratum: ANG223-224 to ANG231-232.
Description: Rather elongated, almost cylindrical specimen in lateral view. In ventral view the specimen is cylindro-conical to slightly spindle-like. The alveolar part is laterally compressed, while the apical part is dorso-ventrally compressed. In between the rostrum changes from laterally compressed, to almost round and to dorso-ventrally compressed. The apex is pointed and centrally placed. The alveolar groove is long and reaches just below the point in which the specimen has its maximum thickness. The alveolus is rather shallow, reaching approximately halfway the extension of the groove, or less.
Remarks: It is well possible that M. (?) horeshaensis represents immature stages of M. (?) inguriensis. There was no type-specimen chosen from the material originally studied by (1938), so a lectotype has to be chosen from his material.
Phylogeny: Probably relates to M. (?) elegans.
Stratigraphic occurrence: Bedoulian D. forbesi Zone to base of D. deshayesi ? Zone.
Belemnite association: BdBA2 (FO) and BdBA3 (LO).
Geographical occurrence: Bulgaria, France (Vocontian Basin), and Georgia.
Mesohibolites (?) aff. inguriensis (, 1938)
Pl. 13 ![]() ,
figs. 7-8; Pl. 14
,
figs. 7-8; Pl. 14 ![]() ,
figs. 5-6, 7-8
,
figs. 5-6, 7-8
+ 1970a Neohibolites ingouriensis (); , p. 42-43, Pl. XXI, fig. 3.
2000 Neohibolites aff. clansayensis ; ,
Pl. 4, fig. 7 (herein Pl. 13 ![]() ,
figs. 7-8).
,
figs. 7-8).
Material: Five specimens.
Stratum: ANG187.1 to ANG188 (four specimens), and ANG200.
Description: The specimens are like M. uhligi regarding their ventral outline, but in lateral view they show a cylindro-conical to almost cylindrical outline comparable to M. (?) inguriensis. They appear to be more robust as compared to the nominal species.
Remarks: In the upper part of the range of M. uhligi, specimens occur that show a much more laterally compressed alveolar part of the rostrum [Pl. 14 ![]() , 17-18; Pl. 15
, 17-18; Pl. 15 ![]() ,
figs. 7-8, 19-20; cf. Mesohibolites schaoriensis sensu
(1989, text-fig.
e)]. They appear intermediate between M. uhligi and M. (?) aff. inguriensis (indicated by a blue two-headed arrow in Fig. 4
,
figs. 7-8, 19-20; cf. Mesohibolites schaoriensis sensu
(1989, text-fig.
e)]. They appear intermediate between M. uhligi and M. (?) aff. inguriensis (indicated by a blue two-headed arrow in Fig. 4 ![]() ). Also see remarks with M. uhligi.
). Also see remarks with M. uhligi.
Phylogeny: Probably descendant from M. uhligi.
Stratigraphic occurrence: Latest Barremian Pc. waagenoides Subzone.
Belemnite association: BaBA6a (topmost part, FO and common) and BdBA1 (LO).
Geographical occurrence: France (Vocontian Basin), and Georgia (?).
Genus Mucrohibolites , 1969
Type species: Mesohibolites schaoriensis , 1952.
Remarks: So far, the genus Mucrohibolites is known only from the Caucasus, i.e., Georgia and probably Azerbaijan. Apparently, it ranges from the Barremian to the Aptian, being especially abundant in the Aptian. Part of this material seems to be very similar to Neohibolites semicanaliculatus . (non , 1827) and Mesohibolites notus , 1935. This genus is hardly mentioned outside Georgia. It was originally defined by (1969, p. 179) with the monotypic species as genotype.
Genus Neohibolites , 1911
Type species: Belemnites semicanaliculatus , 1850 (non , 1827) (= Belemnites Ewaldi , 1861). Over the years, this genus was reason for much dispute because of some apparent inconsistencies regarding its type species (cf. , 1911, 1919; , 1964; , 1968; , 1970b) and its status is actually pending validation by the ICZN ( et al., 1998, p. 160). It was originally established on material from the Boreal-Atlantic Province.
Range: Latest Barremian (see remarks) - Earliest Middle Cenomanian ( et al., 1981).
Remarks: The first ammonite indicative for the Bedoulian was found in the bed ANG200. The first Neohibolites occurs in slightly older levels (ANG197-198). As a result, in the ABSS, this genus occurs in the latest Barremian Pc. waagenoides Subzone already. There and by small neohibolitoid species occur already well in the lower part of the Late Barremian I. giraudi Zone (see: juvenile ? Mesohibolites).
Neohibolites aff. ewaldi (, 1861)
Pl. 12 ![]() ,
figs. 24-25, 28-29, 30-31, 32-33, 34-35
,
figs. 24-25, 28-29, 30-31, 32-33, 34-35
non 1861 Belemnites Ewaldi , p. 34-38.
? + 1981 Mesohibolites eichwaldi [sic!] ; , p. 139 (Late Barremian).
1998 Neohibolites aff. ewaldi (); & , p. 1119, 1122, Pl. I, fig. 7.
2002 Neohibolites ewaldi (); et al., p. 111-112, Pl. XVI, figs. 3-4.
Material: Twenty specimens.
Stratum: ANG197-198 to ANG231-232.
Description: Small, dorso-ventrally compressed specimens with a short alveolar groove. Both in ventral as well as in lateral view, the specimens are spindle-like.
Remarks: The specimens are reminiscent of juvenile Neohibolites, and are well-comparable to the specimens figured by & (1998) and et al. (2002). Additionally, these sediments yielded forty-two juvenile specimens without alveolar groove preserved. These are well-comparable to N. patara , 1957. These occur in BdBA1 to BdBA4.
Stratigraphic occurrence: FO is in the uppermost part of the Pc. waagenoides Subzone (Barremian-Bedoulian boundary beds).
Belemnite association: BdBA1 (FO) to BdBA3.
Geographical occurrence: France (Vocontian Basin), Georgia, Romania, and Russia (Kabardino-Balkaria ASR).
Belemnite zonation in the ABSS
Based on the data presented above, and given the fact that we deal with belemnite-associations from the ABSS which we can compare to areas outside the Vocontian Basin, it seems justified to define certain biozones based on the (partial) range of selected belemnite species. Fig. 7 ![]() depicts the relationship between the biozones we define herein and the ammonite zones in the ABSS. For each zone and subzone the faunal elements will be given.
depicts the relationship between the biozones we define herein and the ammonite zones in the ABSS. For each zone and subzone the faunal elements will be given.
The latest Hauterivian is not zoned herein. The sediments contain belemnites that belong to HaBA2c. This interval yields the last occurrence (LO) of D. ex gr. dilatata, abundant H. ex gr. subfusiformis and is further characterized by the absence of Pseudoduvalia and Pseudobelus. We propose the following biozonation, based on the distribution of belemnites in the ABSS:
Zone unnamed [BaBA1]
Definition - The base of this biozone is defined by the first occurrence (FO) of Duvalia aff. silesiaca (ANG073-074), and the top is defined by the FO of taxa of the Shvetsovia gagrica-varians group (ANG117). Three subzones are defined herein.
Zone of Shvetsovia gagrica [BaBA2]
Definition - The base of this zone is defined by the FO of the S. gagrica-varians group (ANG117). The top is defined by the FO of S. carpatica (ANG136.1-2). Three subzones are defined herein.
Zone of Shvetsovia carpatica [BaBA3]
Definition - The base of this zone is defined by the FO of the index species (ANG136.1-2). The top is defined by the FO of Co. garshini (ANG150). This zone is
divided into three subzones.
Zone of Conohibolites minaretiformis [BaBA4]
Definition - The base of this zone is defined by the FO of Co. garshini (ANG150). The top is defined by the FO of Mesohibolites. This zone is
divided into three subzones.
Zone of Mesohibolites beskidensis [BaBA5]
Definition - The base of this zone is defined by the FO of M. beskidensis (ANG168). The top is defined by the FO of M. uhligi (ANG174.2-175.1).
Characteristic species - Besides the index species (FO and very common in the lower part), S. ex gr. elegantoidea (LO), M.
? renngarteni (FO and common), D. grasiana (common), and D. schwetzowi (rare). Conohibolites and Shvetsovia become extinct in the lowermost part.
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Figure 7: Distribution of the Barremian belemnites associations (BaBA) in the ABSS. Ammonite zonation based on et al. (2011) and additional data from (1996, 1998a, 1998b, 2002) and et al. (2011). Ammonite zones in blue are not (yet) recognized by et al. (2011).
Zone of Mesohibolites uhligi [BaBA6]
Definition - The base of this subzone is defined by the FO of the index species (ANG174.2-175.1). The top is defined by the FO of M. ex gr. longus (ANG189).
Zone of Mesohibolites ekimbontchevi [BaBA7]
Definition - The base of this zone is defined by the FO of the index species (ANG194). The top is defined by the FO of M. bulgaricus (ANG198).
Characteristic species - Besides the index species (FO), M. ex gr. longus sp.
nov., D. grasiana (very rare), and Neohibolites (FO in the uppermost part).
Zone of Mesohibolites bulgaricus [BdBA1]
Definition - The base of this zone is defined by the FO of M. bulgaricus (ANG198). The top is defined by the FO of M. (?) elegans (ANG210). D. grasiana is apparently absent.
Characteristic species - Besides the index species (FO), M. ekimbontchevi (LO), and regularly Neohibolites. In the top most part yields M. (?) elegans? (a species without an alveolar groove).
Zone of Mesohibolites (?) elegans [BdBA2]
Definition - The base of this zone is defined by the FO of M. (?) elegans (ANG210). The top is defined by the FO of M. georgicus (ANG226). Apparently, the sediments yielded no D. grasiana.
Characteristic species - Besides the index species (FO), M. nalchikensis ? (rare), M. (?) inguriensis (FO in the uppermost part), and Neohibolites.
Zone of Mesohibolites georgicus [BdBA3]
Definition - The base of this zone is defined by the FO of M. georgicus (ANG226). The top is defined by the absence of Mesohibolites - not found so far in situ in the beds ANG236 to 238 (=
COM 127-129), these beds are most probably followed by a hiatus of yet unknown scale between the so-called "Bédoulien calcaire" and "Bédoulien marneux".
Characteristic species - Besides the index species (FO, LO and common), M. fallauxi (FO and LO), M. (?) elegans (LO), M. (?) inguriensis (LO), M. (?) horeshaensis (FO and LO), few D. grasiana, and Neohibolites. Actually, M. (?) horeshaensis was first recorded in the marly bed below ANG226.
Zone of Neohibolites spp. [BdBA4]
Definition - The base of this zone is defined by the absence of Mesohibolites (beds above ANG236 to 238; = COM 127-129). The top is not defined herein.
Characteristic species - Neohibolites (common to abundant) and D. grasiana (LO; in the reworked, with pitted belemnites, or condensed level on top of COM137).
Belemnite zonation in BOU
In general the belemnite associations in BOU can be compared to the ABSS. However, the associations differ from several points of view. While Duvalia is largely absent from the more proximal areas, juvenile and/or immature specimens of Mesohibolitidae, several Curtohibolites and certain species of Conohibolites (gladiiformis, tzankovi) are much more common as compared to the ABSS. Partially due to the condensed nature of many of the beds distinguished in BOU. However, this also seems to reflect natural habitats. The belemnite associations show the condensed nature of BOU very well. BaBA2c and BaBA3a appear to be restricted to few condensed beds, while BaBA4a appears to be restricted to one glauconitic bed.
The base of BaBA1 was not investigated in BOU. However, the index species does occur in a comparable proximal section at Clos de Barral (the glauconitic marly level CBL119-120 and CBL120; cf. , 1980a, 1980b), so does D. pontica (CBL122 or CBL123), and H. ex gr. jaculiformis (CBL123 or 125). In BOU the latter species occurs in the beds BOU124 (not depicted) to BOU134.3-135. H. mirificus was not found in situ so far in BOU but loose specimens were found. In the top part of CBL134 this species was found together with Cu. (?) wernsdorfensis. The latter species is found in BOU136 and in BOU137.2. So, all the index species for the three subzones (BaBA1a-c) can be found in these proximal facies.
The FO of the S. gagrica-varians group (BaBA2) is in the glauconitic top of BOU136.1. The S. gagrica-varians group is especially abundant in the beds BOU137.1 to BOU138 (N. pulchella Zone to base K. compressissima Zone; BaBA2b). So far, BOU141.1 yields the last specimens of this group. These specimens, recorded as S. varians s.l. are difficult to separate from Co. gladiiformis, which they are probably ancestral to. Above this bed, probably in the top part of the C. darsi Subzone, the first Curtohibolites occur. In a glauconitic nodular level between BOU141.2 and BOU 142.1 few S. aff. elegans occur.
There and by this level yields the first S. carpatica. The latter belemnites point to the presence of BaBA3. The bed BOU142.1 yielded the first S. simionescui (BaBA3b). In the glauconitic bed BOU142.3 Co. gladiiformis is abundant, among the specimens large (mature to gerontic) morphs previously indicated as Conohibolites
sp. A ( & ,
2005; ,
2010) occur. In these glauconitic beds, and up to the bed 145.1, Co. gladiiformis, S. simionescui and Cu. oosteri are most common. In the glauconitic bed BOU146.3-147.1 belemnites are abundant. Also, phosphatic, green coloured (glauconitic) small Barremites and rolled belemnites occur (Fig. 5 ![]() ). This striking level is
preceded by a thin white weathered layer (ash-layer?), just above the calcareous bed BOU146.
). This striking level is
preceded by a thin white weathered layer (ash-layer?), just above the calcareous bed BOU146.
In the glauconitic bed, typical earliest Late Barremian belemnites occur, among which Co. minaretiformis is most abundant. There and by, few Duvalia and Co. platyurus occur. However, most abundant are Co. garshini and S. elegantoidea in this condensed bed. Except for Co. garshini, the other species can also be found in the few remaining beds before the section is truncated by Albian marls and calcareous sandstones. At the base of these sediments a level with rolled and broken belemnites occurs. These belemnites probably belong to Neohibolites.
It can be concluded that the belemnite associations from the distal facies (ABSS and DLX) match with those from the more proximal facies (BOU and CBL). The most striking differences exist in the near absence of Duvalia in the more proximal sections. More proximal facies show a higher abundance of certain species of Conohibolites, Curtohibolites, and juvenile to immature Mesohibolitidae. Especially, Co. gladiiformis appears to be much more abundant in the more proximal facies. Parts of these differences will be due to the condensed nature of the more proximal sections but it is believed that part reflect natural habitat and natural differences in abundance.
The zonation proposed herein is applicable beyond the Vocontian Basin. In parts of Bulgaria, Czech Republic, Georgia, Hungary, Slovakia, and southern Russia comparable species occur but in general, stratigraphical details are lacking. Although gaps occur in the ranges of some belemnite species and genera, we can in all probability make the following phylogenetical conclusions. In general, these conclusions are based on more or less comparable morphology of the rostrum.
Our thanks go to Myette (Réserve naturelle géologique de Haute-Provence, Digne-les-Bains, France) for permission to sample belemnites, Shalva (Tbilisi, Georgia) for discussions and information regarding certain belemnites, Jaap (Vinkeveen, The Netherlands) for supporting and encouraging this work, Jean (Barrême, France) for donation of some specimens and for the good company in and out of the field. We would like to express our cordial thanks to Martin (Prague, Czech Republic) and Zdenek (Ostrava, Czech Republic) for their conscientious review. We would like to thank John (Gainesville, U.S.A.) for linguistic corrections and improvements.
ZooBank Reference: urn:lsid:zoobank.org:pub:3AB8B960-941E-4A4B-B03C-B15AF1D7D6A4
R.H. (2008).- Lithostratigraphy and biostratigraphy of some Lower Cretaceous outcrops from Northern Sinai, Egypt.- Cretaceous Research, London, vol. 29, p. 603-624.
Sh.A., R.A., M.B., I.V., E.V. & M.Z. (1988).- Guidebook of field trips in Georgia November 10-16, 1988.- International field meeting in Georgia on Tethyan Lower Cretaceous stratigraphy. Platform carbonates. Project no. 262 IGCP-UNESCO stratigraphic correlation and facies interrelations of the Cretaceous in the Tethyan Realm, Tbilisi, 56 p.
A.A. [ Ак.А.] (1964).- О систематическом положении Neohibolites semicanaliculatus и роде Neohibolites / O sistematicheskom polozhenii Neohibolites semicanaliculatus i rode Neohibolites [About the systematic position of Neohibolites semicanaliculatus andabout the genus Neohibolites].- Палеонтологический Журнал, Москва/ Paleontological journal, Moscow, 1964, n° 4, p. 78-86 (Pls. X-XI).
A.A. [ Ак.А.] (1965).- Новый род семейства Belemnitidae / Novyi rod semeistva Belemnitidae [New genera in the family Belemnitidae].- Палеонтологический Журнал, Москва/ Paleontological journal, Moscow, 1965, n° 3, p. 142-144 (1 Pl.).
A.A. [ Ак.А.] (1972).- Меловые белемниты Азербайджана / Melovye belemnity Azerbaidzhana [Lower Cretaceous belemnites of Azerbaijan].- Недра, Москва, 279 p.
E. & M.C. (1998).- Barremian-Aptian boundary in the Dambovicioara area (Rumanian Carpathians).- Zentralblatt für Geologie und Paläontologie, Stuttgart, (Teil I), 1996, Heft 11/12, p. 1117-1129.
R. & M.C. (2006).- Palaeoenvironmental and palaeobiologic changes across the Barremian/Aptian boundary interval in the Tethys Realm, Mexico and Romania.- Cretaceous Research, London, vol. 27, p. 529-541.
É. (1878).- Fossiles principaux des terrains. Atlas, première partie.- Explication de la carte géologique de France, Paris, vol. 4 (176 Pls.).
D., G. & S. (2008).- Nouveaux biohorizons et propositions pour le découpage biozonal ammonitique du Barrémien supérieur du Sud-Est de la France.- Carnets de G�ologie [Notebooks on Geology], Brest, Article 2008/03 (CG2008_A03), 18 p.
H.-M. Ducrotay de (1827).- Mémoire sur les bélemnites, considérées zoologiquement et géologiquement.- F.G. Levrault, Paris, 127 p.
S., N., A., V., S., A., N.M.M., P. & K.B. (2009).- Early Cretaceous (late Berriasian to early Aptian) palaeocenaographic change along the northwestern Tethyan margin (Vocontian Trough, southeastern France): δ13C, δ18O and Sr-isotope belemnite and whole-rock records.- Cretaceous Research, London, vol. 30, p. 1247-1262.
T.N. & I.A. (2004).- Origin, evolution and stratigraphic significance of the superfamily Deshayesitaceae , 1949.- Bulletin de l'Institut royal des Sciences naturelles des Belgique, (Sciences de la Terre), Bruxelles, t. 74, p. 189-243.
R. (1965).- Le stratotype du Barrémien. Lithologie et macrofaune. In : Colloque sur le Crétacé inférieur (Lyon, septembre 1963).- Mémoires du Bureau de Recherches Géologiques et Minières, Paris, n° 34, p. 101-116.
R. & G. (1966).- Le Barrémien et ses limites dans le centre de la Montagne de Lure (Basses-Alpes).- Bulletin de la Société géologique de France, Paris, (série 7), t. VIII, p. 415-422.
R. & J. (1986).- La limite Hauterivien-Barrémien dans la région stratotypique d'Angles (Sud-Est de la France).- Compte rendus de l'Académie des Sciences, Paris, (série II), t. 302, fasc. 7, p. 457-459.
F., P. & R. (2000).- La base de l'Aptien à Cassis-La Bédoule (SE France).- Géologie Méditerranéenne, Marseille, t. XXV, n° 3/4 (1998), p. 149-157.
A. (2000, unpublished).- Biodiversité des bélemnites du Barrémo-Bédoulien du Sud-Est de la France et de l'Europe téthysienne: analyse comparative, implications paléobiogéographiques et paléoenvironnementales.- Mémoire de DEA Lyon, 39 p.
R. (2000).- Les Bélemnites du stratotype historique de l'Aptien inférieur dans la région de Cassis-La Bédoule (SE France).- Géologie Méditerranéenne, Marseille, t. XXV, fasc. 3/4 (1998), p. 177-179.
R., W.K., D.P. & Ch. (1981).- Les Bélemnites.- Cretaceous Research, London, vol. 2, n° 3/4, p. 283-286.
M., J. & J.M. (2003).- Ammonite biostratigraphy of the uppermost Hauterivian in the Betic Cordillera (SE Spain).- Géobios, Villeurbanne, n° 36, fasc. 6, p. 685-694.
G. (1989).- Fossiles du plateau d'Albion.- Les Alpes de Lumière, Aix-en-Provence, t. 99, 72 p.
G. (1995).- About some significant ammonites from the Lower Aptian (Bedoulian) of the Angles - Barrême area (South-East France).- Memorie Descrittive della Carte Geologica d'Italia, Roma, vol. LI, p. 65-101.
G. (1998).- Biostratigraphie des faunes d'Ammonites à la limite Barrémien-Aptien dans la région d'Angles-Barrême-Castellane. Étude particulière de la famille des Heteroceratina , 1922 (Ancyloceratina, Ammonoidea).- Annales du Muséum d'Histoire Naturelle, Nice, t. XII (1997), 270 p.
G., R., P., R., G., M. & J.-P. (1997).- The 'Pseudocrioceras bed' at La B�doule SE France and position of the Barremian-Aptian boundary in the historical lower Aptian stratotype.- Comptes Rendus de l'Acad�mie des Sciences, Paris, Sciences de la Terre et des Plan�tes, s�rie IIa, t. 325, n� 8, p. 593-599.
G. & P. (1995).- On the Genus Spinocrioceras, , 1973 (Ammonoidea, Ancyloceratina).- Geologisches Jahrbuch, Hannover, (Reihe A), Heft 141, p. 201-223.
M. (1952).- Deux nouvelles Hibolites du Crétacé inférieur.- Bulletin de la Société Géologique de France, Paris, (série 6), t. II, fasc. 1-3, p. 41-45.
de P. (1808).- Conchyliologie systématique, et classification méthodique des coquilles; offrant leur figures, leur arrangement générique, leurs descriptions caractérisques, leurs noms, ainsi que leur synonymie en plusieurs langues. Coquilles univalves, cloisonnées, vol. I.- L. Haussmann, Paris, 409 p.
J. (1841).- Bélemnites des terrains crétacés inférieures des environs de Castellane (Basses-Alpes), considérées géologiquement et zoologiquement, avec la description de ces terrains.- Académie des Sciences Naturelles, Paris, 80 p.
M.S. [ М.С.] (1952).- Грузинская глыба в нижнемеловое время / Gruzinskaya glyba v nizhnemelovoe vremya [The Georgian block in Early Cretaceous time].- Труды Геологического Института Академия Наук Грузинской ССР, Тбилиси, (минералого-петрографическая серия), t. VI (XI), p. 137-210.
M.S. [ М.С.] (1957).- Сопоставление нижнемеловых отложений Грузии и Крыма / Sopostavlenie nizhnemelovykh otlozhenie Gruzii i Kryma [Comparison of the Lower Cretaceous deposits of Georgia and the Crimea].- Академия наук Грузинской ССР, Москва, (геологический институт), Монографии n° 8, 84 p.
J.W. (1850).- Ueber die Grenze zwischen Neocom und Gault.- Zeitschrift der Deutschen Geologischen Gesellschaft, Berlin, Band 2, p. 440-478.
S. (1989).- Contrôle eustatique de la resédimentation calcaire en fosse vocontienne (Mésozo�que, S-E de la France).- Géotrope, Lyon, vol. 1 (1988), p. 32-51.
S. (2005).- Système turbiditiques calcaires du Tithonien et du Barrémo-Bédoulien subalpins (S-E de la France). In: Excursion du Groupe Français du Crétacé, 1-3 septembre 2005, Lyon, 13 p.
S. & J. (1979).- Mégabrèches de resédimentation, lacunes mécaniques et pseudo-"hard-grounds" sur la marge vocontienne au Barrémien et à l'Aptien inférieur (Sud-Est de la France).- Géologie Alpine, Grenoble, t. 55, p. 75-92.
J. (1967).- Parastratotypes du Crétacé inférieur.- Compte Rendu sommaire des Séances de la Société Géologique de France, Paris, 1967, n° 1, p. 29-30.
I. & N.N.M. (2009).- Integrated Lower Cretaceous biostratigraphy of the Bersek Quarry, Gerecse Mountains, Transdanubian Range, Hungary.- Cretaceous Research, London, vol. 30, p. 78-92.
G. & O. (2003).- Anatomy of ancient passive margin slope systems: Aptian gravity-driven deposition on the Vocontian palaeomargin, western Alps, south-east France.- Sedimentology, Amsterdam, vol. 50, p. 1231-1270.
N.K. [ Н.К.] (1966).- О стратиграфическом объеме глинистой толщи нижнего мела юго-западного Крыма / O stratigraficheskom ob'eme glinistoi tolshchi nizhnego mela yugo-zapadnogo Kryma [About the stratigraphical extend of the Lower Cretaceous clayey beds of SW Crimea].- Вестник Ленинградского Университета, (геологии и географии n° 4), Ленинград, n° 24, p. 129-131.
N.K. [ Н.К.] (1968).- Problems of nomenclature. Systematics of Early Cretaceous Belemnopsinae.- Paleontological Journal, Washington, vol. 2, n° 3, p. 383-384.
N.K. [Горн Н.К.] (1974).- Стратиграфия и история формирования нижнемеловых глин юго-западного Крыма / Stratigrafiya i istoriya formirovaniya nizhnemelovykh glin yugo-zapadnogo Kryma [Stratigraphy and history of the formation of Early Cretaceous clay of SW Crimea]. In: Вопросы Стратиграфии / Voprosy Stratigrafii [Stratigraphical questions], I.- Ленинградский Ордена Ленина и Ордена Трудового Красного Знамени Государственный Университет имени А.А. Жданова, Ленинград, p. 92-100.
M. & J. (2010).- Locality #6. Stratotypic section of the Barremian. In: P., F, K. et al. (Eds.), Fieldtrip 2. Four days in Jurassic and Cretaceous outcrops in Lyon and Digne-les-Bains areas.- Fieldtrip guidebook for the 8th International Symposium, Cephalopods - Present and Past September 5 to 8, 2010, Lyon, p. 30-32.
V.A. [ В.А.] (1973).- Подкласс Endocochlia внутрираковинные. Отряд Бelemnitida / Podklass Endocochlia vnutrirakovinnye. Otryad Belemnitida [Subclass Endocochlia. Order Belemnitida]. In: M.V. [ М.В.] (ed.), Методика проведения геологической практики и атлас руководящих форм. / Metodika provedeniya geologicheskoi praktiki и atlas rukovodyashchikh form [Practical geological guide and atlas of guide-fossils].- Руководство по Учебной Геологической Практике в Крыму / Rukovodstvo po Uchebnoi geologicheskoi Practike v Krymu [Manual and teaching of geological practice in the Crimea], Недра, Москва, vol. I, p. 146-157 (Pls. XLVII-LI.).
Ph.J. (1995).- Ammonite distribution around the Hauterivian-Barremian boundary along the Río Argos (Caravaca, SE Spain).- Géologie Alpine, Grenoble, Mémoire H.S., n° 20 (1994), p. 219-277.
H. & J. (1980).- Barrême-Cephalopoden aus dem kretazischen Untergrund des Stadtgebietes von Hannover (NW-Deutschland).- Paläontologische Zeitschrift, Stuttgart, Heft 54, p. 241-266.
N.M.M. (1997).- Mediterranean Neocomian belemnites, part 1: Río Argos sequence (province of Murcia, Spain): the Berriasian-Valanginian and the Hauterivian-Barremian boundaries.- Scripta Geologica, Leiden, vol. 114, p. 1-55.
N.M.M. & I. (2004).- Neocomian cephalopods from the Bersek Hill (Gerecse Mountains, Hungary), part I: Hauterivian - earliest Barremian.- Fragmenta Palaeontologica Hungarica, Budapest, vol. 22, p. 27-49.
N.M.M. & I. (2005).- Neocomian belemnites and ammonites from the Bersek-hegy (Gerecse Mountains, Hungary), part II: Barremian.- Fragmenta Palaeotologica Hungarica, Budapest, vol. 23, p. 59-86.
N.M.M. (2009).- Mediterranean Neocomian belemnites, part 3: Valanginian-Hauterivian belemnites.- Carnets de G�ologie [Notebooks on Geology], Brest, Memoir2009/01 (CG2009_M01), 44 p.
N.M.M. (2010).- Barremian invertebrates from Serre de Bleyton (Drôme, SE France): Belemnites.- Annalen des naturhistorischen Museums, Wien, (serie A), vol. 112, p. 659-672.
G.K. [ Г.К.] (1960).- Белемниты / Belemnity [Belemnites]. In: V.V. [ В.В.] & M.P. [ М.П.] (Eds.), Атлас нижнемеловой фауны северного Кавказа и Крыма / Atlas nizhnemelovoi fauny severnogo Kavkaza i Kryma [Atlas of the Lower Cretaceous fauna of the northern Caucasus and the Crim].- Труды Главное Управлние Газовой Промышленности при Совете Министров СССР, ВНИИГаз, Москва, 1960, p. 356-369 (3 Pls.).
I. [ И.] (2000).- On the Upper Barremian - Lower Aptian sequence boundaries (western Georgia).- Proceedings Georgian Academy of Sciences A. Janelidze Geological Institute, Tbilisi, (new series), vol. 115, p. 48-57.
M.V. [ М.В.] (1971).- Колхидиты и их стратиграфическое значение / Kolkhidity i ikh stratigraficheskoe znachenie [Colchidites and their stratigraphical significance].- Труды Геологический Институт Наук Академия Наук Грузии ССР, Тбилиси, (новая серия), vol. 26, 118 p. (22 Pls.).
M.V. [ М.В.] (1981).- Анцилоцератиды юга СССР и их стратиграфическое значение / Antsilotseratidy yuga SSR i ikh stratigraficheskoe znachenie [Ancyloceratidae of the southern Sovjet Union and their stratigrafical significance].- Труды Геологический Институт Наук Академия Наук Грузии ССР, Тбилиси, (новая серия), vol. 71, 196 p. (22 Pls.).
M.V. [ М.В.] (2005).- Барремский ярус. Местийско-Тианетская зона / Barremskii yarus. Mestiisko-Tianetskaya zona [Barremian beds. Mestiisko-Tianetska zone]. In: E.V., I.V., M.V. & L.P. (Eds.), Атлас раннемеловой фауны Грузии / Atlas rannemelovoe fauny Gruzii [Atlas of Early Cretaceous fauna of Georgia].- Proceedings Georgian Academy of Sciences A. Janelidze Geological Institute, Tbilisi, (new series), n° 120, p. 13-14.
M.V. & E.V. (1995).- New data on the Upper Barremian biostratigraphy of the Georgian region (Caucasus). In: Proceedings 3rd Workshop on Early Cretaceous Cephalopods 5-8 July 1994, Piobbico.- Memorie descrittive della Carta Geologica d'Italia, Roma, vol. LI, p. 103-108.
M.V. [ М.В.] & E.V. [ Э.В.] (2003).- K биостратиграфии верхнего баррема Грузии / K biostratigrafii verkhnego barrema Gruzii [Upper Barremian biostratigraphy of Georgia].- Бюллетень Московского Общества Испытателей Природы, Москва, (геологии), tom 78, n° 6, p. 36-45.
Sh.G. [ Ш.Г.] (1990, unpublished).- Стратиграфическое значение i условия существования раннемеловых белемнитид Грузии. Палеонтология и стратиграфия.- АН ГССР, геологический институт, Тбилиси, Диссертации Кандидата Геолого-минералогических Наук, 19 p.
Sh.G. (1998).- Stratigraphical significance of the Early Cretaceous Belemnitida of Georgia.- Bulletin of the Georgian Academy of Sciences, Tbilisi, vol. 157, n° 3, p. 443-446.
Sh.G. (1999).- New representatives of belemnites from the Lower Cretaceous deposits of Georgia.- Paleontological Journal, Moscow, vol. 33, n° 1, p. 19-24.
E. (1973).- Die Unterkreide in Untergrund der Gehrdener Berge und die Deister-Mulde.- Berichte der Naturhistorische Gesellschaft, Hannover, Heft 117, p. 29-54.
I.D. [ И.Д.] (1952).- Меловые белемниты Грузии / Melovye belemnity Gruzii [Cretaceous belemnites of Georgia].- Вестник Государственного музея Грузии, Тбилиси, vol. XV-A (1951), 118 p. (12 Pls.).
W. (1896).- Notice stratigraphique sur les environs de Sisteron et contribution � la connaissance des terrains secondaires du Sud-Est de la France.- Bulletin de la Société Géologique de France, Paris, (3e série), t. 23 (1895-1896), p. 659-803.
W. (1913).- Unterkreide (Paleocretacicum). In: F. (ed.), Lethaea geognostica. Teil 2. Das Mesozoikum. 3. Band Kreide. 1. Abteilung (1912).- Schweizerbart, Stuttgart, 398 p. (18 Pls.).
W. & P. (1915).- Sur quelques ammonites de l'Hauterivien de la B�gude (Basses-Alpes). Mat�riaux pour l'�tude de la faune de l'Hauterivien des environs de Moustiers-Sainte-Marie, La Palud et Ch�teauneuf-les-Moustiers (Basses-Alpes). In: W. (ed.), Contribution � l'�tude des faunes pal�ocr�tac�es du sud-est de la France, vol. 2.- M�moires pour servir � l'explication de la carte g�ologique d�taill�e de la France, Paris, 1915, p. 223-296 (Pls. 10-15).
E.V. [ Э.В.] (1970).- Стратиграфия и фауна колхидитового и смежных горизонтов западной Грузии / Stratigrafiya i fauna kolkhiditovogo i smezhnykh gorizontov zapadnoi Gruzii [Stratigraphy and fauna of the kolkhiditov and adjacent horizons in southern Georgia].- Труды Академия наук Грузинской ССР, (геологический институт), Тбилиси, (новая серия / novaja serija), n° 25, 115 p. (20 Pls.).
see
A., A. & G. (2010).- The invertebrate fauna from the Barremian of Serre de Bleyton (Drôme, SE France).- Annalen des naturhistorischen Museums, Wien, (serie A), vol. 112, p. 569-574.
G.Y. [ Г.Я.] (1939).- Нижнемеловые белемниты Кавказа / Nizhnemelоvye belemnity Kavkaza [Lower Cretaceous belemnites of the Caucasus]. In: Белемниты мезозойских отложений СССР / Belemnity mezozoiskikh oltozhenie SSSR [Belemnites from Mesozoic deposits of the USSR], part 1].- Монографии по палеонтологии СССР / Monografii po paleontologii SSSR, Ленинград, Москва / Leningrad, Moscow, tom LXVII, 54 p.
G.Y. [ Г.Я.] (1949).- Подкласс Endocochlia. Отряд Decapoda. Подотряд Belemnoidea Белемниты / Podklass Endocochlia. Otryad Decapoda. Podotryad Belemnoidea. Belemnity [Subclass Endocochlia. Order Decapoda. Suborder Belemnoidea. Belemnites]. In: L.S. [ Л.С.] et al. (Eds.), Атлас руководящих форм ископаемых фаун СССР, tom X (нижний отдел меловой системы) /Atlas rukovodyashchikh form iskopaemykh fauna SSSR, tom X (nizhnii otdel melovoi sistemy) [Atlas of the leading fossil fauna of the USSR, vol. X (Lower Cretaceous)].- Госгеолиздат, Москва, p. 253-269 (Pls. LXXVIII-LXXXIII).
G.Y. [ Г.Я.] (1997).- Белемниты / Belemnity [Belemnites]. Подкласс Endocochlia внутреннераковинные / Podklass Endocochlia vnytrennerakovinnye [Subclass Endocochlia inner-shelled]. In: V.V. [ В.В.] & T.N. [Богданова Т.Н.] (Eds.), Атлас меловой фауны юго-западного Крыма / Atlas melovoi fauny yugo-zapadnogo Kryma [Atlas of the Cretaceous fauna of the south-western Crimea].- Пангея, Санкт-Петербург, p. 146-155 (Pls. 59-60).
I. (1937).- Structure géologique de la Ratcha, du Letchkhoum et de l'Imérétie (Géorgie occidentale). In: A. (ed.), Excursion au Caucase RSS de Géorgie partie occidentale.- XVIIe Congrès Géologique International URSS 1937, Léningrad, Moscou, 84 p.
I.V. [ И.В.] & Sh.G. [ Ш.Г.] (2005).- Внутреннераковинные головоногие. Белемниты / Vnutrennerakovinnye golovonogie. Belemnity [Endocochlian cephalopods. Belemnites]. In: E.V., I.V., M.V. & L.P. (eds.), Атлас раннемеловой фауны Грузии / Atlas rannemelovoi fauny Gruzii [Atlas of Early Cretaceous fauna of Georgia].- Proceedings Georgian Academy of Sciences A. Janelidze Geological Institute, Tbilisi, (новая серия / novaja serija), n° 120, p. 447-474 (Pls. 106-109).
I.V. [ И.В.] & T.Yu. [ Т.Ю.] (1975).- О неизвестных нижнемеловых белемнитидах Грузии / O neizvestnykh nizhnemelovykh belemnitidakh Gruzii [About unfamiliar Early Cretaceous belemnites from Georgia].- Труды Академия наук Грузинской ССР, (геологический институт), Тбилиси, (новая серия / novaja serija), n° 47, p. 132-153.
I.V. [ И.В.] & L.Z. [ Л.З.] (2005).- Нижний баррем. Тагрско-Джавская зона /Nizhnii barrem. Targsko-Dzhavskaya zona [Early Barremian. Targsko-Dzhavskaya zone]. In: E.V., I.V., M.V. & L.P. (Eds.), Атлас раннемеловой фауны Грузии / Atlas rannemelovoi fauny Gruzii [Atlas of Early Cretaceous fauna of Georgia].- Proceedings Georgian Academy of Sciences A. Janelidze Geological Institute, Tbilisi, (новая серия / novaja serija), n° 120, p. 26-29.
P. & J. (2009).- Spinocrioceras sauvanae sp. nov. et ses implications sur la compréhension phylogénique du genre Spinocrioceras , 1973.- Annales du Museum d'Histoire naturelle de Nice, Nice, t. XXIV, fasc. 2, p. 93-102.
J. & Z. (1989).- Lower Cretaceous stratigraphy and palaeogeography of the Czechoslovakian Western Carpathians. In: J. (ed.), Cretaceous of the Western Tethys.- Proceedings of the 3rd International Cretaceous Symposium Tübingen 1987, Schweizerbart, Stuttgart, p. 505-523.
Z.A. [З.А.] (1935).- Белемниты мелового флиша юго-восточного Кавказа / Belemnity melovogo flisha yugo-vostochnogo Kavkaza [Cretaceous belemnites of the flysch of the south-eastern Caucasus].- Труды Нефтяного геолого-разведочного Института, Ленинград, Москва, (серия A), vol. 74, p. 3-37 (3 Pls.).
S.F. & Ch.J. (1999).- Lithological and faunal stratigraphy of the Aptian and Albian (Lower Cretaceous) of the type Speeton Clay, Speeton, north-east England.- Proceedings of the Yorkshire Geological Society, Yorkshire, vol. 52, part 3, p. 277-296.
M., B. & G. (2011).- The Aptian Stage: Back to fundamentals.- Episodes, Journal of International Geosciences, Bangalore, vol. 34, n� 3, p. 148-156.
M., J.-P., G., W., P., J.A., E. & M. (2000).- Le stratotype historique de l'Aptien inférieur (région de Cassis-La Bédoule, SE France) : synthèse stratigraphique.- Géologie Méditerranéenne, Marseille, t. XXV, n° 3/4 (1998), p. 289-298.
J. & K. (1995).- Die Bio- und Lithofazies der Unterkreide (Hauterive bis Apt) in NW-Deutschland.- Berliner geowissenschaftliche Abhandlungen, Berlin, (Reihe E), Heft 16, p. 227-253.
T.Yu. [ Т.Ю.] (1968).- Значение отношения послеальвеолы к послеборозде для нижнемеловых Белемнопсин Грузии / Znachenie otnosheniya posleal'veoly k posleborozde dlya nizhnemelovykh Belemnopsin Gruzii [Ratio of the postalveolar and apical parts of the Lower Cretaceous Belemnopseina of Georgia].- Сообщения АН ГССР / Bulletin of the Academy of Sciences of the Georgian SSR, Тбилиси, vol. 51, n° 3, p. 631-636.
T.Yu. [ Т.Ю.] (1969).- Род / Rod [Genus] Mucrohibolites , gen. nov. In: O. (ed.), I. Республиканская научная конференция молодых ученых и аспирантов Грузинской ССР, посвященная 100-летию со дня рождения В.И. / I. Respublikanskaya nauchnaya konferentsiya molodykh uchenykh i aspirantov Gruzinskoi SSR, posvyashchennaya 100-letiyu so dnya rozhdeniya V.I. [First republic scientific conference for young students and aspirants of the Georgian SSR, dedicated to the 100-anniversary of V.I. ].- Центральный Комитет ЛКСМ Грузии, Тбилиси, 1968, p. 179-181.
T.Yu. [ Т.Ю.] (1973).- Нижнемеловые белемнитиды Грузии / Nizhnemelovye belemnitidy Gruzii [Lower Cretaceous belemnites of Georgia].- Труды Академия наук Грузинской ССР, (геологический институт), Тбилиси, (новая серия / novaja serija), tom 40, 125 p.
V.M. [ В.М.] (1983).- Раннемеловые белемниты юга СССР / Rannemelovye belemnity Yuga SSSR [Early Cretaceous belemnites of the southern USSR]. In: Y.I. [ Я.И.] & K.N. [ К.Н.] (Eds.), Систематика и экология головоногих моллюсков / Sistematika i ekologiya golovonogikh mollyuskov [Taxonomy and ecology of cephalopods].- Akademiya nauk SSSR, Zoologicheskii Institut, Leningrad, p. 42-43.
W.A. (1857).- Catalogue des céphalopodes fossiles des Alpes suisses avec la description et les figures des espèces remarquables. 1ère partie. Céphalopodes acétabulifères. Décapodes. In: Pétrification remarquable des Alpes suisses.- Nouvelles Mémoires de la Société helvétique des Sciences naturelles, Genève, vol. 17, n° 2, p. 5-34 (Pls. 1-3).
A.M., H., G.R. R.A. & H. (2006).- Correlation of Barremian-Aptian (mid-Cretaceous) dinoflagellate cyst assemblages between the Tethyan and Austral realms.- Cretaceous Research, London, vol. 27, p. 792-813.
D. & E. (1976).- Stratigraphie et corrélation des terrains Néocomiens et Barrémo-Bédouliens du Couloir de Dîmbovicioara (Carpates orientales).- Dări de Seamă ale Şedinţelor, Bucuresti, t. LXII (1974-1975), 4 (stratigrafie), p. 135-160.
A.P. [ А.П.] (1914).- Юрскія и нижнемеловыя cephalopoda северной Сибири / Yurskiya i nizhnemelovyya cephalopoda severnoi Sibiri [Jurassic and Early Cretaceous cephalopods of northern Siberia].- Записки Императорское Академии Наукъ / Zapiski Imperatorskoe Akademii Nauk', St. Petersburg, (serija 8 - физико-математическое отделеніе), vol. XXI, n° 4 (1913), 68 p.
E. (1842).- Observations sur un nouveau genre de saurien fossile, le Neustosaurus Gigondarum, N., avec quelques notes géologiques sur la commune de Gigondas.- Meilhac, Paris; Clément St-Just, Avignon, p. 56.
F.V. (1829).- Histoire naturelle des bélemnites, accompagnée de la description et de la classification des espèces, que M. de Castellane a recuillies dans les Basses-Alpes de Provence.- Annales des Sciences d'Observation, Paris, t. 1, p. 271-331.
S., P.F., J. A., M.B., R., Y., M., C., V.I., A., B., V., H., Z., E.J., D., S., T.N., L.G., J.L., I.A., P. & O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Dijon, France, 30th August 2010).- Cretaceous Research, London, vol. 32, n� 6, p. 786-793.
W. (1995).- Cephalopoda dibranchiata fossiles (Coleoidea). In: F. (ed.), Fossilium Catalogus. I: Animalia.- Kugler, Amsterdam, New York, Part 133, 411 p.
W., N.M.M. & C. (1998).- Cephalopoda dibranchiata fossiles (Coleoidea) II. In: F. (ed.), Fossilium Catalogus. I: Animalia.- Backhuys, Leiden, Part 135, 512 p.
J. (1952).-Sous-classe des Dibranchiata 1836 (Coleoidea , Endocochlia ). In: J. (ed.), Traité de paléontologie. Tome II. Problèmes d'adaptation et de phylogénèse. Brachiopodes. Chétognathes. Annélides. Géphyriens. Mollusques.- Masson & Cie, Paris, p. 689-755.
P., G., R., J.-P. & M. (2000).- Les faunes d'Ammonites du Barrémien supérieur/Aptien inférieur (Bédoulien) dans la région stratotypique de Cassis-La Bédoule (SE France) : état des connaissances et propositions pour une zonation par Ammonites du Bédoulien-type.- Géologie Méditerranéenne, Marseille, t. XXV, n° 3/4 (1998), p. 167-175.
P., M., R., G. & G. (2006).- The Deshayesoitidae , 1949 (Ammonoidea) of the Aptian historical stratotype region at Cassis-La Bédoule (SE France).- Carnets de G�ologie [Notebooks on Geology], Brest, Mémoire 2006/01 (CG2006_M01), 46 p.
or see
I. (1931).- Notice sur les b�lemnites et les nautilus de l'Aptien de Koutaïs.- Bulletin du Musée de Géorgie, Tbilisi, t. VI (1929-1930), p. 125-140 (3 Pls.).
I. (1938).- Quelques céphalopodes nouveaux ou peu connus de l'Aptien de la Géorgie. In: A. & I. (Eds.), Céphalopodes nouveaux ou peu connus de l'Aptien de la Géorgie.- Bulletin de l'Institut Géologique de Géorgie, Тbilisi, vol. III, part 2, p. 129-191 (6 Pls.).
see
M.S. [ М.С.] (1913).- Нижне-меловые белемниты Абхазіи (Гагры-Сухумъ). Les Bélemnites infracrétacées de l'Abkhasie (Gagry-Sukhum).- Ежегодник по Геологии и Минералогии России [Annuaire Géologique et Minéralogique de la Russie], Pulawy, t. 15, n° 2-3, p. 43-74.
I. (1898).- Studii geologice si paleontologice din Carpatii sudici. II. Fauna neocomiana din basenul Dîmboviciórei.- Academia Româna, Publicatiunile fondului Vasilie Adamachi, Bucuresti, t. II, p. 101-167.
G.R. (1973).- Cretaceous belemnites. In: A. (ed.), Atlas of palaeobiogeography.- Elsevier, Amsterdam, p.385-401 (Pl. I).
E. (1911).- Beiträge zur Kenntnis der Cephalopoden der norddeutschen Unteren Kreide. I. Die Belemnitiden der norddeutschen Unteren Kreide. 1. Die Belemniten des norddeutschen Gaults (Aptiens und Albiens).- Geologische und Palaeontologische Abhandlungen, Jena, (neue Folge), Band X, Heft 3, p.203-272 (8Pls.).
E. (1919).- Die Systematik der Belemniten.- Elfter Jahresbericht des Niedersächsischen geologischen Vereins, Hannover, Geschäftsjahr 1918, 59 p.
M. [ М.] (1963).- Curtohibolites gen. nov. (Belemnitida) от долнокредните отложения в България / Curtohibolites gen. nov. (Belemnitida) ot dolnokredite otlozheniya v Bolgariya [Curtohibolites gen. nov. (Belemnitida) from Early Cretaceous beds in Bulgaria].- Трудове върху Геологията на България, София, (палеонтология), t. 5, p. 211-227.
M. [ М.] (1964).- Стратиграфско разпространение на долно-кредните белемнити в България / Stratigrafsko razrostranenie na dolnokredite belemniti v Bolgariya [Stratigraphic distribution of belemnites in the Lower Cretaceous of Bulgaria].- Списание на Българското Геологическо Дружество / Spisanie na Bulgarskogo Geologichesko Druzhestvo, Sofia, t. 25, n° 2, p. 137-150.
M. (1965).- Nouvelles espèces des bélemnites du Crétacé inférieur en Bulgarie.- Trudy v'rkhu Geologiyata na Bulgariya, Sofia, (série pal�ontologique), t. 7, p. 151-177 & 179-223.
M. (1970a).- Les fossiles de Bulgarie. IVa Crétacé inférieur. Belemnitida.- Éditions Académie Bulgare des Sciences, Sofia, 72 p.
M. (1970b).- Sur le problème de la nomenclature du genre Neohibolites .- Списание на Българското Геологическо Дружество / Review of the Bulgarian Geological Society, София, vol. XXXI, part 2, p. 187-195 (3 Pls.).
M. [ М.] (1979).- Hibolites targovishtensis n. sp. (Belemnitida) от барема в Североизточна България / Hibolites targovishtensis n.sp. (Belemnitida) ot barema v Severoiztochna Bolgariya [Hibolites targovishtensis n. sp. (Belemnitida) in the Barremian of northeast Bulgaria].- Българска Академия на Науките, София, (палеонтология, стратирафия и литология), t. 10, p. 37-40.
A. von (1861).- Über den Gault und besondere die Gargas-Mergel (Aptien d'.) im nordwestlichen Deutschland nächst dem Harze.- Zeitschrift der deutschen geologischen Gesellschaft, Berlin, Band XIII, p. 20-60.
H.H. (1935).- The rocks below the Red Chalk of Lincolnshire, and their cephalopod faunas.- The Quarterly Journal of the Geological Society, London, vol. 91, p. 1-46 (Pls. I-IV).
H.H. (1937).- A monograph of British Lower Cretaceous belemnites. Part 2 (for 1936).- Palaeontological Society, London, vol. 90, p. xvii-xxxii & 17-30 (Pls. VI-IX).
M.V. [ М.В.], S.G. [ Ш.Г.] & I.V. [ И.В.] (2002).- Юрские и меловые белемнитиды Грузии / Yurskie i melovye belemnitidy Gruzii [Jurassic and Cretaceous belemnitids of Georgia].- Труды Академия Наук Грузии геологический институт им. А.И. Джанелидзе / Trudy Akademiia nauk Gruzii geologicheskiĭ institut im A.I. Dzhanelidze, Tbilisi, (новая серия / novaya seriya), n° 118, 302 p.
V. (1883a).- Die Wernsdorfer Schichten und ihre Äquivalente.- Sitzungsberichte der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaft, Wien, Band 86, Abtheilung 1 (1882), p. 86-117.
V. (1883b).- Die Cephalopodenfauna der Wernsdorfer Schichten.- Denkschriften der kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Classe, Wien, Band 64, p. 127-290.
V. (1902).- Ueber die Cephalopodenfauna der Teschener und Grodischter Schichten.- Denkschriften der kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Classe, Wien, Band 72 (1901), 87 p.
Z. (1978a).- Untersuchungen an Kreide-Belemniten der schlesischen Einheit (Äussere Karpaten, Tschechoslowakei). Teil I.- Casopis Slezského Muzea, Opava, (serie A), vol. 27, n° 1, p. 1-16 (2 Pls.).
Z. (1978b).- Untersuchungen an Kreide-Belemniten der schlesischen Einheit (Äussere Karpaten, Tschechoslowakei). Teil II. Teil.- Casopis Slezského Muzea, Opava, (serie A), vol. 27, n° 2, p. 113-123.
Z., J. & D. (1994).- Early Cretaceous stratigraphy, palaeogeography and life in the Western Carpathians.- Beringeria, Würzburg, Heft 10, 169 p.
J. (1980a, unpublished).- Étude de la Famille des Pulchelliidae, revision de trois espèces types du Barr�mien du sud-est de la France.- Thèse de l'Université de Nice, Faculté des Sciences et Techniques, 92 p.
J. (1980b).- Biozonation homophyl�tique du Barrémien du Clos de Barral (Var). In: G. (ed.), Ammonites.- Ed. Serre, Nice, p. 181-184 (Pls. V-VI).
J. (1996).- Nouvelle biozonation du Barrémien basée sur la Famille des Pulchelliidae (Ammonoidea).- Géologie Alpine, Grenoble, t. 71 (1995), p. 199-211.
J. (1998a).- Biohorizons ammonitique dans le Barrémien (de la zone à Hugii à zone à Sartousiana).- Géologie Alpine, Grenoble, t. 73 (1997), p. 99-117.
J. (1998b).- Nouvelle biostratigraphie ammonitique du Barrémien (pro parte) du Sud-Est de la France.- Earth & Planetary Sciences, Paris, t. 327, p. 619-624.
J. (2002).- Étude stratigraphique et paléontologique de la famille des Pulchellidae (Ammonoidea, Ammonitina, Endomocerataceae).- Géologie Alpine, Grenoble, Mémoire H.S., n° 42, 333 p.
J. (2004).- Vers une nouvelle classification à fondement phylogénétique des ammonites hétéromorphes du Crétacé inférieur méditerranéen. Les cas des Crioceratitidae , 1871, nom. correct. , 1952, des Emericiceratidae fam. nov. et des Acrioceratidae fam. nov. (Ancylocerataceae , 1871).- Riviéra Scientifique, Nice, n° 88, p. 69-92.
J. (2005).- Boundaries, ammonite fauna and main subdivisions of the stratotype of the Barremian. In: T., A., H. et al. (eds.), The Hauterivian - Lower Aptian sequence stratigraphy from Jura Platform to Vocontian Basin : a multidisciplinary approach.- Géologie Alpine, Grenoble, (série spéciale colloques et excursions), n° 7, p. 147-173.
J. & P. (2010a).- Nouvelles données sur le genre Imerites , 1933 (Ammonitida, Ancyloceratina): réponses et précisions apportées au travail de , et , 2009.- Annales de Paléontologie, Paris, vol. 96, p. 13-23.
J. & P. (2010b).- Sur la compréhension de l'espèce-type du genre Imerites , 1933 et ses implications biostratigraphiques.- Annales du Museum d'Histoire naturelle de Nice, Nice, t. XXV, p. 105-108.
J., P., P., L. & E. (2009).- Nouvelles données sur la famille des Hemihoplitidae , 1924.- Annales du Museum d'Histoire naturelle de Nice, Nice, t. XXIV, fasc. 2, p. 65-91.
L., H., J.-P. & L. (2002).- Chemostratigraphic correlation of Barremian and lower Aptian ammonite zones and magnetic reversals.- International Journal of Earth Sciences (Geologische Rundschau), Berlin-Heidelberg, vol. 91, p. 272-279.
All specimens (x1) unless otherwise indicated. In all specimens the alveolar view is shown (left picture), while in the lateral view the ventral side is to the left unless otherwise indicated (VR - ventral side to the right; VL - ventral side to the left).
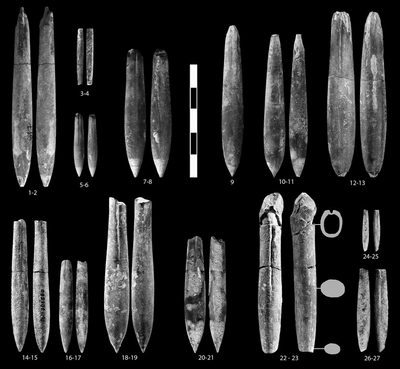
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 1. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Hibolithes ex gr. subfusiformis (,
1829), ANG006-007, B. balearis Zone, RGM 361567
figs. 3-4. Hibolithes ex gr. subfusiformis (,
1829) (juv.) ANG034-035, B. balearis Zone, RGM 582971
figs. 5-6. Hibolithes ex gr. jaculiformis ,
1913 (juv.), ANG102-103, K. nicklesi Zone, RGM 583003
figs. 7-8. ibid. (x2)
fig. 9. Hibolithes ex gr. jaculiformis ,
1913, ANG102-104, K. nicklesi Zone, RGM 361556
figs. 10-11. Hibolithes ex gr. jaculiformis ,
1913, ANG102-103, K. nicklesi Zone, RGM 583006
figs. 12-13. Hibolithes ex gr. jaculiformis ,
1913, ANG088-089, Pt. mazuca Subzone, RGM 592995
figs. 14-15. Hibolithes ex gr. jaculiformis ,
1913 (imm.), ANG087-088, Pt. mazuca Subzone, RGM 361560
(VR)
figs. 16-17. Hibolithes ex gr. jaculiformis ,
1913 (imm.), ANG100-102, K. nicklesi Zone, RGM 361820a
figs. 18-19. Hibolithes targovishtensis ,
1979, ANG084-085, Pt. mazuca Subzone, RGM 582994
figs. 20-21. Hibolithes aff. jaculiformis ,
1913, ANG110.2-3, N. pulchella Zone, RGM 361840
figs. 22-23. Hibolithes keleptrishvilii sp. nov. [HT], ANG045 P. ohmi Zone,
RGM 361817
figs. 24-25. Genus species nov., ANG021-022, B. balearis Zone, RGM 582963 (ventral side not known)
figs. 26-27. Ibid. (x2) (ventral side not known)
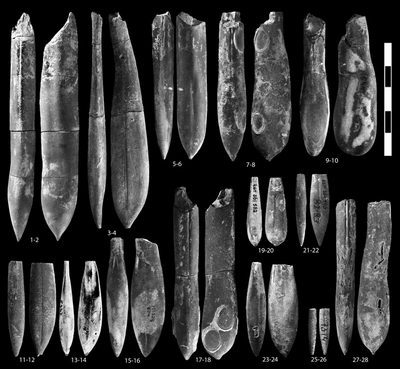
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 2. In all specimens ventral side is to the right, unless otherwise indicated.
figs. 1-2. Duvalia pontica , 1913, ANG102-103, K. nicklesi Zone, RGM 583004
figs. 3-4. Duvalia dilatata binervioides ,
1965, ANG014, B. balearis Zone, RGM 582995
figs. 5-6. Duvalia pontica , 1913
(ger.), ANG100-101, K. nicklesi Zone, RGM 583002
figs. 7-8. Duvalia gagrica ? ,
1913, ANG083-084, Pt. mazuca Subzone, RGM 582997
figs. 9-10. Duvalia silesiaca , 1902
(mat?), ANG081-082, Pt. mazuca Subzone, RGM 583019
figs. 11-12. Duvalia pontica , 1913
(imm.), ANG103-104, K. nicklesi Zone, RGM 361834
figs. 13-14. Duvalia silesiaca , 1902, ANG084-085, Pt. mazuca Subzone, RGM 582990
figs. 15-16. Duvalia gagrica ? ,
1913, ANG085-086, Pt. mazuca Subzone, RGM 361558
figs. 17-18. Duvalia gagrica , 1913, ANG083-084, Pt. mazuca Subzone, RGM 582998
figs. 19-20. Duvalia silesiaca , 1902
(imm.), ANG084-085, Pt. mazuca Subzone, RGM 361562 (VL)
figs. 21-22. Duvalia silesiaca , 1902
(imm.), ANG082-083, Pt. mazuca Subzone, RGM 361563 (VL)
figs. 23-24. Duvalia silesiaca , 1902, ANG092-093, Psilotissotia colombiana Subzone, RGM 582997
figs. 25-26. Duvalia silesiaca , 1902
(juv.), ANG073-074, T. hugii auct. Subzone, RGM 361565 (VL)
figs. 27-28. Duvalia gagrica , 1913, ANG093-094, Pt. colombiana Subzone, RGM 582998 (VL)
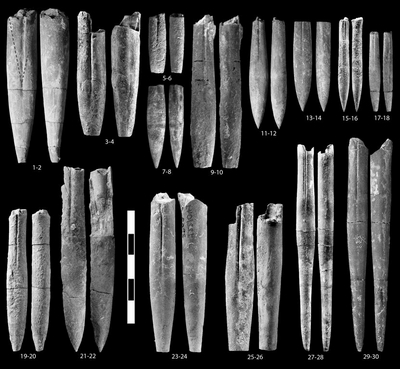
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 3. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Curtohibolites (?) wernsdorfensis (,
1963), ANG117, S. defayae Subzone, RGM 583023 (VR)
figs. 3-4. Curtohibolites (?) wernsdorfensis (,
1963), ANG120-121, S. defayae Subzone, RGM 361856 (VR)
figs. 5-6. Curtohibolites (?) wernsdorfensis ? (,
1963) (imm.), ANG120-121, S. defayae Subzone, RGM 361855 (VR)
figs. 7-8. Shvetsovia ex gr. gagrica-varians (,
1913) (imm.), ANG120-121, S. defayae Subzone, RGM 583020
figs. 9-10. Shvetsovia gagrica (, 1913), BOU137.2-138, K. compressissima Zone, RGM 583367 (VR)
figs. 11-12. Shvetsovia varians (, 1913), ANG120-121, S. defayae Subzone, RGM 361854
figs. 13-14. Shvetsovia ex gr. gagrica-varians (,
1913) (imm.), ANG121-122, S. defayae Subzone, RGM 361859
figs. 15-16. Shvetsovia varians (, 1913), ANG122, S. defayae Subzone, RGM 583031 (VR)
figs. 17-18. Shvetsovia varians (, 1913)
(juv.), BOU138-139, K. compressissima Zone, RGM 583353 (VR)
figs. 19-20. Shvetsovia gagrica (, 1913), ANG122, S. defayae Subzone, RGM 361860 (VR)
figs. 21-22. Shvetsovia gagrica (, 1913), BOU137.2, K. compressissima Zone, RGM 583369 (VR)
figs. 23-24. Shvetsovia varians (, 1913), ANG122, S. defayae Subzone, RGM 583032 (VR)
figs. 25-26. Shvetsovia gagrica (, 1913), ANG121-123, S. defayae Subzone, RGM 361861 (VR)
figs. 27-28. Shvetsovia varians (, 1913), ANG121-122, S. defayae Subzone, RGM 583034 (VR)
figs. 29-30. Shvetsovia varians (, 1913), ANG122, S. defayae Subzone, RGM 583033 (VR)
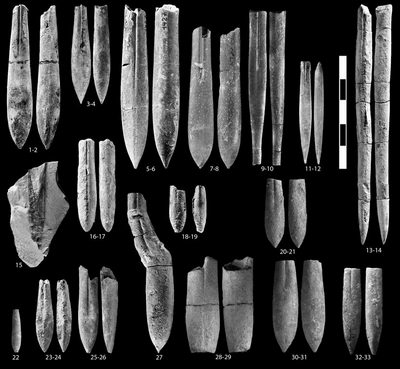
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 4. In all specimens ventral side is to the left.
figs. 1-2. Shvetsovia aff. elegans (,
1913), ANG133-134, C. darsi Zone, RGM 583046
(VR)
figs. 3-4. Shvetsovia aff. elegans (,
1913) (imm.), ANG134-135, M. tirolensis Subzone, RGM 583048
(VR)
figs. 5-6. Shvetsovia aff. elegans (,
1913), ANG135-136.1, M. tirolensis Subzone, RGM 583051
figs. 7-8. Shvetsovia aff. elegans (,
1913), ANG136.2-137, M. tirolensis Subzone, RGM 583052
(VR)
figs. 9-10. Shvetsovia gladiiformis (, 1883)
(imm./mat.), ANG144-145, Ho. uhligi Zone, RGM 361878
figs. 11-12. Shvetsovia gladiiformis (, 1883)
(juv./imm.), ANG143.5-144, Ho. uhligi Zone, RGM 361878
(VR)
figs. 13-14. Shvetsovia carpatica (, 1883), ANG138-139.1, S. defayae Subzone, RGM 361872
(VR)
fig. 15. Shvetsovia simionescui (,
1970) internal view, ANG141.1, Ho. uhligi Zone, RGM 361872
(VR)
figs. 16-17. Shvetsovia aff. elegans (,
1913) (imm.), ANG138-139, M. tirolensis Subzone, RGM 361873
figs. 18-19. Curtohibolites (?) bourguetensis sp. nov. (juv./imm.), ANG129-131, C. darsi Zone, RGM 361499
(VR)
figs. 20-21. Curtohibolites oosteri ,
1963 (imm.), BOU144 base, M. tirolensis Subzone, RGM 583290
figs. 22. Shvetsovia sp. (juv.) BOU144-145.1, M. tirolensis Subzone, RGM 583281
figs. 23-24. Shvetsovia aff. elegans? (,
1913) (imm.), ANG136.2-137, M. tirolensis Subzone, RGM 361871
(VR)
figs. 25-26. Shvetsovia simionescui (,
1970) (imm.), ANG144-145, Ho. uhligi Zone, RGM 361881
fig. 27. Shvetsovia aff. elegans (,
1913), ANG135-136.1, M. tirolensis Subzone, RGM 361867
figs. 28-29. Curtohibolites (?) bourguetensis sp. nov. [HT], BOU137.2-138, K. compressissima Zone, RGM 583366
(VR)
figs. 30-31. Curtohibolites oosteri ,
1963, BOU144 base, M. tirolensis Subzone, RGM 583289
(VR)
figs. 32-33. Shvetsovia aff. elegans (,
1913) imm., BOU141.1-142.1, C. darsi Zone, RGM 583337
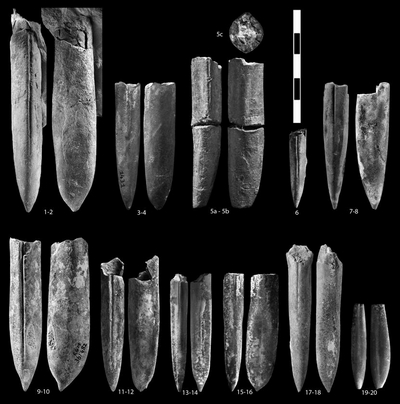
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 5. In all specimens ventral side is to the right, unless otherwise indicated.
figs. 1-2. Duvalia vermeuleni sp. nov. (HT), ANG120, S. defayae Subzone, RGM 361561
figs. 3-4. Duvalia vermeuleni sp. nov. (immature), ANG120-121, S. defayae Subzone, RGM 583021
figs. 5a-c. Duvalia schwetzowi ,
1999 (mature), ANG143.3-4, Ho. uhligi Zone, RGM 583062
fig. 6. Duvalia grasiana (, 1841) (immature), ANG171.3-172.1, I. giraudi Zone, RGM 361481
figs. 7-8. Duvalia vermeuleni sp. nov., ANG161.2-3, G. sartousiana Zone, RGM 583126
(VL)
figs. 9-10. Duvalia grasiana (, 1841), ANG169.2-170.1, I. giraudi Zone, RGM 361481
figs. 11-12. Duvalia grasiana (, 1841), ANG226-227, D. forbesi Zone, RGM 583756
figs. 13-14. Duvalia schwetzowi ,
1999, ANG151.3-152, B. barremense Subzone, RGM 583085
figs. 15-16. Duvalia pontica , 1913
(immature), ANG104-106, K. nicklesi Zone, RGM 361554
figs. 17-18. Duvalia schwetzowi ,
1999, DLX111-112; B. barremense Subzone, RGM 583855
(VL)
figs. 19-20. Duvalia grasiana (, 1841)
(juv./imm.), ANG171.1, I. giraudi Zone, RGM 583557

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 6. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Conohibolites minaretiformis (,
1913), ANG161.2-3, B. barremense Subzone, RGM 583136
figs. 3-4. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583092
figs. 5-6. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583095
(VR)
figs. 7-8. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583090
figs. 9-10. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583082
(VR)
figs. 11-12. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583219
(VR)
figs. 13-14. Conohibolites sp. juv., ANG151.3-152, B. barremense Subzone, RGM 361913
figs. 15-16. Shvetsovia elegantoidea (,
1965), ANG157.1-2, B. barremense Subzone, RGM 361492
figs. 17-18. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 361907
(VR)
figs. 19-20. Conohibolites minaretiformis (,
1913), ANG152-153.1, B. barremense Subzone, RGM 361917
(VR)
figs. 21-22. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 361566
figs. 23-24. Shvetsovia elegantoidea (,
1965), ANG153.2-154, B. barremense Subzone, RGM 361919
figs. 25-26. Conohibolites minaretiformis (,
1913), ANG156.2-3, B. barremense Subzone, RGM 583106
(VR)
figs. 27-28. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583089

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 7. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Conohibolites minaretiformis (,
1913), ANG151.3, B. barremense Subzone RGM 583081 (VR)
figs. 3-4. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583091 (VR)
figs. 5-6. Conohibolites cf. minaretiformis (,
1913) corroded, ANG161.2 top, G. sartousiana Zone, RGM 361962a (VR)
figs. 7-8. Conohibolites platyurus (,
1841), ANG152-153.1, B. barremense Subzone, RGM 583099 (VR)
figs. 9-10. Conohibolites garshini (,
1965), BOU146.3-147.1, B. barremense Subzone, RGM 583197
figs. 11-12. Conohibolites garshini (,
1965), ANG151.3-152, B. barremense Subzone, RGM 583094 (VR)
figs. 13-14. Conohibolites tzankovi (,
1965), BOU144 base, Ho. uhligi Zone, RGM 583294
figs. 15-16. Conohibolites platyurus (,
1841), ANG151.3-152, B. barremense Subzone, RGM 583096
figs. 17-18. Conohibolites garshini (,
1965), ANG151.3-152, B. barremense Subzone, RGM 583093 (VR)
figs. 19-20. Conohibolites garshini (,
1965), ANG160.1-2, B. barremense Subzone, RGM 583111 (VR)
figs. 21-22. Conohibolites platyurus (,
1841) (imm.), BOU146.3-147.1, B. barremense Subzone, RGM 583198 (VR)
figs. 23-24. Conohibolites platyurus (,
1841), ANG151.3-152, B. barremense Subzone, RGM 583097
figs. 25-26. Conohibolites sp. (teratological?), ANG166.2-167, H. feraudianus Subzone, RGM 583512 (VR)
figs. 27-30. Conohibolites tzankovi (,
1965) (imm.), BOU142.3, H. uhligi Zone, RGM 583315 (VR)
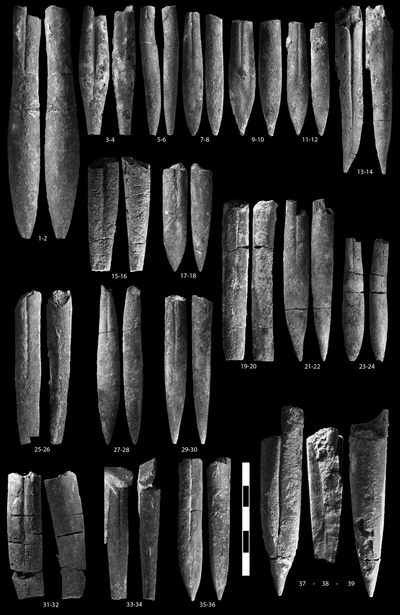
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 8. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Shvetsovia aff. elegans ? (,
1913) (gerontic?), BOU141 or BOU142, M. tirolensis? Subzone, RGM 583304 (ex situ) (VR)
figs. 3-4. Shvetsovia simionescui ? (,
1970), BOU141 or BOU142, M. tirolensis? Subzone, RGM 583335 (ex situ) (VR)
figs. 5-6. Shvetsovia carpatica ? (,
1883), BOU146.3-147.1, B. barremense Subzone, RGM 583195 (VR)
figs. 7-8. Shvetsovia elegantoidea (,
1913), BOU 146.3-147.1, B. barremense Subzone, RGM 583254
figs. 9-10. Shvetsovia sp. [? S. elegantoidea (,
1913)], BOU146.3-147.1, B. barremense Subzone, RGM 583202 (VR)
figs. 11-12. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583229 (VR)
figs. 13-14. Conohibolites gladiiformis (,
1883), BOU142.3, M. tirolensis Subzone, RGM 583322 (VR)
figs. 15-16. Conohibolites cf. gladiiformis (,
1883) imm., BOU142 or BOU143, M. tirolensis Subzone, RGM 583308 (ex situ) (VR)
figs. 17-18. Conohibolites garshini (,
1965), BOU146.3-147.1, B. barremense Subzone, RGM 583232 (VR)
figs. 19-20. Conohibolites cf. gladiiformis (,
1883), BOU142 or BOU143, M. tirolensis Subzone, RGM 583310 (ex situ)
figs. 21-22. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583223
figs. 23-24. Shvetsovia elegantoidea (,
1965), BOU146.3-147.1, B. barremense Subzone, RGM 583203
figs. 25-26. Conohibolites gladiiformis (,
1883), BOU144 or BOU145.1, Ho. uhligi Zone, RGM 583279 (ex situ) (VR)
figs. 27-28. Shvetsovia elegantoidea (,
1965), BOU146.3-147.1, B. barremense Subzone, RGM 583205
figs. 29-30. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583249 (VR)
figs. 31-32. Conohibolites gladiiformis (,
1883), BOU145.1-2, Ho. uhligi Zone, RGM 583262 (VR)
figs. 33-34. Conohibolites minaretiformis (,
1913), BOU148 base, B. barremense Subzone, RGM 583173 (VR)
figs. 35-36. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583216 (VR)
figs. 37-39. Conohibolites tzankovi (,
1965), BOU142 or BOU143, M. tirolensis Subzone, RGM 583305 (ex situ)
(figs. 39 - VR)

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 9. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Shvetsovia sp. 1 nov.?, ANG165.2-166.1, H. feraudianus Subzone, RGM 583488 (VR)
figs. 3-4. S. aff. elegantoidea (,
1965), ANG165.2 top, H. feraudianus Subzone, RGM 583478 (VR)
figs. 5-6. Conohibolites minaretiformis (,
1913), ANG161.3-4, G. sartousiana Subzone, RGM 361984 (VR)
figs. 7-8. S. elegantoidea (,
1965), ANG156.5-6, B. barremense Subzone, RGM 361921 (VR)
figs. 9-10. S. elegantoidea (,
1965), ANG166.2-167, H. feraudianus Subzone, RGM 583511 (x0.7)
figs. 11-12. S. elegantoidea (,
1965), ANG165.2-166.1, H. feraudianus Subzone, RGM 583503 (VR)
figs. 13-14. Conohibolites minaretiformis (,
1913), ANG151.3-152, B. barremense Subzone, RGM 583087 (VR)
figs. 15-16. Conohibolites minaretiformis (,
1913), ANG160.3-161, M. sartousiana Subzone, RGM 361947a (VR)
figs. 17-18. Co. trastikensis (,
1965) (imm.), ANG165.2-166, H. feraudianus Subzone, RGM 583486 (VR)
figs. 19-20. Co. trastikensis (,
1965) corroded, ANG165.2-166, H. feraudianus Subzone, RGM 583498 (VR)
figs. 21-22. S. elegantoidea (,
1965), ANG161.2-3, M. sartousiana Subzone, RGM 583132 (VR)
figs. 23-24. Co. garshini (,
1965), ANG161.3-162, M. sartousiana Subzone, RGM 583147 (VR)
figs. 25-26. S. elegantoidea (,
1965), ANG164.3-165, H. feraudianus Subzone, RGM 583457
figs. 27-28. Conohibolites minaretiformis (,
1913), BOU146.3-147.1, B. barremense Subzone, RGM 583230 (VR)
figs. 29-30. Conohibolites sp. (juv.), ANG164.3-165, H. feraudianus Subzone, RGM 583466 (VR)
figs. 31-32. S. ex gr. elegantoidea (,
1965), ANG167-168, H. feraudianus Subzone, RGM 583522 (VR) (x0.7)
figs. 33-34. S. ex gr. elegantoidea (,
1965) (imm.), ANG168-169.1, H. feraudianus Subzone, RGM 583533
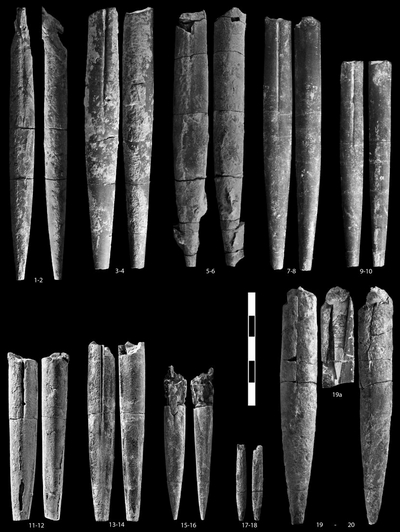
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 10. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites beskidensis (, 1883), ANG171.1, I. giraudi Subzone, RGM 583559
(VR)
figs. 3-4. Mesohibolites beskidensis (, 1883), ANG170.1, I. giraudi Subzone, RGM 583548
(VR)
figs. 5-6. Mesohibolites beskidensis (, 1883), ANG170.2, I. giraudi Subzone, RGM 583556
figs. 7-8. Mesohibolites beskidensis (, 1883), ANG170.1, I. giraudi Subzone, RGM 583549
(VR)
figs. 9-10. Mesohibolites beskidensis (,
1883), ANG170.1, I. giraudi Subzone, RGM 583550
(VR)
figs. 11-12. Mesohibolites anglesensis sp. nov. holotype, ANG178.2-179, M. sarasini Subzone, RGM 583617
(VR)
figs. 13-14. Mesohibolites beskidensis (,
1883), ANG169.2-170.1, I. giraudi Horizon, RGM 583539
figs. 15-16. Mesohibolites anglesensis sp. nov. paratype, ANG179.3 top, M. sarasini Subzone, RGM 583631
figs. 17-18. Mesohibolites beskidensis (,
1883), ANG169.2-170.1, I. giraudi Horizon, RGM 583541
figs. 19-20. Mesohibolites beskidensis (,
1883), ANG168-169.1, I. giraudi Horizon, RGM 583531
(VR)
fig. 19a. ibid. (alveolar part) (VR)
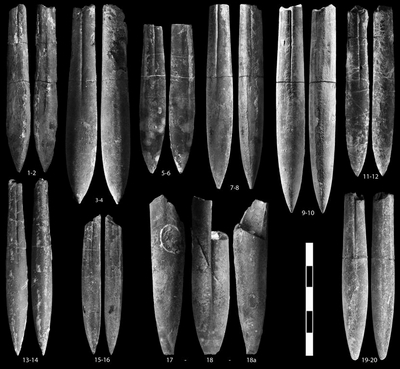
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 11. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites renngarteni ,
1939, ANG169.1-2, I. giraudi -Horizon, RGM 583536
figs. 3-4. Mesohibolites renngarteni ,
1939, ANG169.2-170.1, I. giraudi -Horizon, RGM 583543
figs. 5-6. Mesohibolites renngarteni ,
1939, ANG169.2-170.1, I. giraudi -Horizon, RGM 583544 (VR)
figs. 7-8. Mesohibolites uhligi (, 1913), ANG177.1, M. sarasini Subzone, RGM 583605
figs. 9-10. Mesohibolites uhligi (, 1913), ANG177.1, M. sarasini Subzone, RGM 583606
figs. 11-12. Mesohibolites renngarteni ,
1939, ANG172.3, I. giraudi Subzone, RGM 583571 (VR)
figs. 13-14. Mesohibolites renngarteni ? ,
1939, ANG176.1, I. giraudi Subzone, RGM 583594
figs. 15-16. Mesohibolites renngarteni ,
1939 (immature), ANG172.3, I. giraudi Subzone, RGM 583572
figs. 17-18a. Conohibolites tzankovi (,
1965), DLX101-102; C. darsi/M. uhligi Zone, RGM 583541
(fig. 18a - VR)
figs. 19-20. Mesohibolites uhligi (, 1913), ANG179.1-2, M. sarasini Subzone, RGM 583623
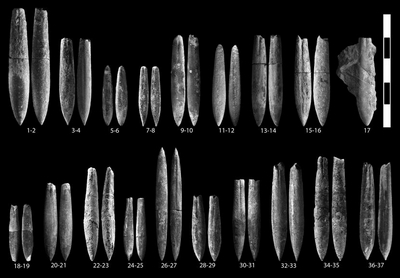
Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 12. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites ? sp. juvenile, ANG172.1 top, I. giraudi Subzone, RGM 583565
figs. 3-4. Mesohibolites ? sp. juvenile, ANG173-174.1 top, I. giraudi Subzone, RGM 583578
figs. 5-6. Mesohibolites ? sp. juvenile, ANG171.1 top, I. giraudi Subzone, RGM 583558
figs. 7-8. Mesohibolites ? sp. juvenile, ANG172.3-172, I. giraudi Subzone, RGM 583573
figs. 9-10. Mesohibolites ? sp. juvenile, ANG (167)-174, I. giraudi Subzone, RGM 583581
figs. 11-12. Mesohibolites ? sp. juvenile, ANG175.2-176, I. giraudi Subzone, RGM 583589 (VR)
figs. 13-14. Mesohibolites ? sp. juvenile, ANG178.2-179.1, M. sarasini Subzone, RGM 583620 (VR)
figs. 15-16. Mesohibolites ? sp. juvenile, ANG179.2-3, M. sarasini Subzone, RGM 583629
figs. 17. Mesohibolites ? sp. juvenile, ANG180 base, M. sarasini Subzone, RGM 583636
figs. 18-19. Mesohibolites ? sp. juvenile, COM094-095 (= ANG196.1-197), Pc. waagenoides Subzone, RGM 583803
figs. 20-21. Mesohibolites ? sp. juvenile, COM077-078 (= ANG181-182), M. sarasini Subzone, RGM 583790 (VR)
figs. 22-23. Mesohibolites ? sp. juvenile, ANG184-185, Pc. waagenoides Subzone, RGM 583647
figs. 24-25. Neohibolites aff. ewaldi (,
1861), ANG210 base, D. forbesi Zone, RGM 583744
figs. 26-27. Mesohibolites ? sp. juvenile, ANG200-201, Pc. waagenoides Subzone, RGM 583694
figs. 28-29. Neohibolites aff. ewaldi (,
1861), ANG197-198, Pc. waagenoides Subzone, RGM 583685
figs. 30-31. Neohibolites aff. ewaldi (,
1861), ANG197-198, Pc. waagenoides Subzone, RGM 583683
figs. 32-33. Neohibolites aff. ewaldi (,
1861),
COM102-103 (= ANG205-206), Pa. oglanlensis Zone, RGM 583807 (VR)
figs. 34-35. Neohibolites aff. ewaldi (,
1861), ANG228-229, D. forbesi Zone, RGM 543017
figs. 36-37. Mesohibolites ? sp. juvenile, ANG172.3-173, I. giraudi Subzone, RGM 583574

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 13. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites longus (, 1913), ANG179.3, M. sarasini Subzone, RGM 583630 (VR)
figs. 3-4. Mesohibolites bulgaricus ? ,
1965, ANG205-206, Pa. oglanlensis Zone, RGM 583730
figs. 5-6. Mesohibolites bulgaricus ,
1965, ANG202 top, Pa. oglanlensis Zone, RGM 583701
figs. 7-8. Mesohibolites (?) aff. inguriensis (,
1938), ANG200, Pc. waagenoides Subzone, RGM 361473
figs. 9-10. Mesohibolites longus (, 1913)
(imm.), ANG178.2-179.1, M. sarasini Subzone, RGM 583616 (VR)
figs. 11-12. Mesohibolites bulgaricus ,
1965, ANG207-208, Pa. oglanlensis Zone, RGM 583737 (VR)
figs. 13-14. Mesohibolites bulgaricus ,
1965 (juv.), ANG200-201, P. waagenoides Subzone, RGM 583692
figs. 15-16. Mesohibolites ex gr. longus (,
1913) (imm.), ANG196.1-2, Pc. waagenoides Subzone, RGM 583678 (VR)
figs. 17-18. Mesohibolites longus (, 1913)
(imm.), ANG175.2-176.1, I. giraudi Subzone, RGM 583586
figs. 19-20. Mesohibolites longus (, 1913), ANG179.1-2, M. sarasini Subzone, RGM 583622 (VR)
figs. 21-22. Mesohibolites ekimbontchevi ,
1965, ANG198-199, Pc. waagenoides Subzone, RGM 583688 (VR)
figs. 23-24. Mesohibolites bulgaricus ,
1965, ANG205 top, Pa. oglanlensis Zone, RGM 583715 (VR)
figs. 25-26. Mesohibolites (?) elegans ? (,
1913),
COM103-104 (= ANG206-207), Pa. oglanlensis Zone, RGM 583809 (VR)
figs. 27-28. Mesohibolites anglesensis sp. nov. (imm.), ANG178.1-2, M. sarasini Subzone, RGM 583611 (VR)

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 14. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites uhligi (, 1913), ANG183, M. sarasini Subzone, RGM 583643
(VR)
figs. 3-4. Mesohibolites ex gr. longus (,
1913), ANG189 top, Pc. waagenoides Subzone, RGM 583667
figs. 5-6. Mesohibolites (?) aff. inguriensis (,
1938), ANG187.1, Pc. waagenoides Subzone, RGM 583651
(VR)
figs. 7-8. Mesohibolites (?) aff. inguriensis (,
1938), ANG188 base, Pc. waagenoides Subzone, RGM 583661
figs. 9-10. Mesohibolites uhligi (, 1913), ANG187.2-188, Pc. waagenoides Subzone, RGM 583659
figs. 11-12. Mesohibolites uhligi (, 1913), ANG174.2-175, I. giraudi Subzone, RGM 361478
(VR)
figs. 13-14. Mesohibolites uhligi (, 1913), ANG187.1-2, Pc. waagenoides Subzone, RGM 583652
figs. 15-16. Mesohibolites uhligi (, 1913),
COM082-083 (= ANG187.2-188), Pc. waagenoides Subzone, RGM 583797
(VR)
figs. 17-18. Mesohibolites uhligi? (,
1913) or M. (?) aff. inguriensis (,
1938), ANG187.2 top, Pc. waagenoides Subzone, RGM 583656

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 15. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites ekimbontchevi ,
1965, ANG197 base, Pc. waagenoides Subzone, RGM 583681
figs. 3-4. Mesohibolites ekimbontchevi ,
1965, ANG201-202, Pc. waagenoides Subzone, RGM 583699
figs. 5-6. Mesohibolites nalchikensis ? ,
1939, ANG210, D. forbesi Zone, RGM 583745
figs. 7-8. Mesohibolites uhligi ? (, 1913) or M. (?) aff. inguriensis (,
1938), Méouilles, ?M. sarasini Subzone, RGM 583839 (VR)
figs. 9-10. Mesohibolites elegans (, 1913), ANG230-231, D. forbesi Zone, RGM 583775
figs. 11-12. Mesohibolites uhligi (, 1913), Méouilles, ?M. sarasini Subzone, RGM 583837 (VR)
figs. 13-14. Mesohibolites ekimbontchevi ,
1965, ANG200-202, Pc. waagenoides Subzone, RGM 583697 (VR)
figs. 15-16. Mesohibolites elegans (,
1913), ANG215 top, D. forbesi Zone, RGM 583749
figs. 17-18. Mesohibolites elegans (,
1913), ANG225-226, D. forbesi Zone, RGM 543022
figs. 19-20. M. uhligi ? (, 1913) or M. (?) aff. inguriensis (,
1938), Méouilles, ?M. sarasini Subzone, RGM 583838 (VR)

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 16. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Shvetsovia elegantoidea (,
1965), ANG161.2-3, G. sartousiana Zone, RGM 583135a (VR)
figs. 3-4. Shvetsovia elegantoidea (,
1965) (imm.), ANG161.2-3, G. sartousiana Zone, RGM 583135b (VR)
figs. 5-6. Shvetsovia elegantoidea (,
1965) (imm.), ANG161.2-3, G. sartousiana Zone, RGM 583135c (VR)
figs. 7-8. Mesohibolites (?) inguriensis (,
1938), ANG231-232, D. deshayesi
? Zone, RGM 543003 (VR)
figs. 9-10. Mesohibolites (?) inguriensis (,
1938), ANG223-224, D. forbesi Zone, RGM 583752
figs. 11-12. Mesohibolites fallauxi (, 1883), ANG (231)-238, D. deshayesi Zone, RGM 583789 (ex situ)
figs. 13-14. Mesohibolites (?) horeshaensis (,
1938),
COM121-122 (= ANG230-231), D. forbesi Zone, RGM 583829 (VR)
figs. 15-16. Mesohibolites fallauxi (, 1883) (juvenile),
COM121-122 (= ANG230-231), D. forbesi Zone, RGM 583833 (VR)
figs. 17-18. Mesohibolites georgicus ,
1973, ANG225-226, D. forbesi Zone, RGM 543021 (VR)
fig. 18a. ibid. internal view.
figs. 19-20. Mesohibolites georgicus ,
1973, ANG231-232, D. deshayesi? Zone, RGM 583780 (VR)
figs. 21-22. Mesohibolites (?) inguriensis (,
1938) (imm.),
COM121-122 (= ANG230-231), D. forbesi Zone, RGM 583832
figs. 23-24. Mesohibolites georgicus ,
1973 (imm.), ANG225-226, D. forbesi Zone, RGM 583754
figs. 25-26. Mesohibolites fallauxi (, 1883), ANG229-230, D. forbesi Zone, RGM 583767

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 17. In all specimens ventral side is to the left, unless otherwise indicated.
figs. 1-2. Mesohibolites georgicus ,
1973, ANG228-229, D. forbesi Zone, RGM 583760
(VR)
figs. 3-4. Mesohibolites georgicus ,
1973,
COM120-121 (= ANG229-230), D. forbesi Zone, RGM 583826
figs. 5-6. Mesohibolites georgicus ,
1973, ANG231- (232), D. deshayesi? Zone, RGM 543004
figs. 7-8. Mesohibolites georgicus ,
1973 (imm.), ANG228-229, D. forbesi Zone, RGM 583759
figs. 9-10. Mesohibolites georgicus ,
1973 (juv.), ANG231- (232), D. deshayesi? Zone, RGM 543005
figs. 11-12. Mesohibolites georgicus ,
1973 (imm.), ANG228-229, D. forbesi Zone, RGM 583761 (VR)

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 18:
fig. A. The first beds of the section (ANG001 to ANG 015), Pl. ligatus and B. balearis
zones. Bed-numbers are indicated in red.
fig. B. The first beds of the Barremian are exposed in the lower left part of the picture. The rather thick beds of the Pt. colombiana (ANG090 to ANG093) and the K. nicklesi (from ANG094 on) are as well. View approximately to the west, along the road of the hamlet of Angles. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 19:
The transition from the marly-calcareous to the calcareous dominated beds in the latest Early Barremian. The boundary between the Early and Late Barremian is indicated by the blue arrow in bed ANG147.2. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 20:
fig. A. Small-scaled fault system in the B. barremense - G. sartousiana
subzones. The view is approximately to the west, along the road of the hamlet of Angles. Bed-numbers are indicated in red.
fig. B. View of the marly beds with two thin marly-calcareous beds (ANG161.3 and ANG161.4) on top of ANG161.2 (G. sartousiana Subzone). Typical development of marly intervals in the calcareous dominated earliest Barremian sediments. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 21:
fig. A. Typical development of the calcareous earliest Barremian B. barremense Subzone. Bed-numbers are indicated in red.
fig. B. Base of the B. barremense Subzone with relative thick marly bed (ANG151.3-152) at the base of the section. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 22:
Typical development compressional fault pattern in the B. barremense Subzone. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 23:
The B. barremense Subzone with regular alternations of thin and thick beds of calcareous rock, intercalated with either very thin marly layers and some more pronounced marly beds. Stratigraphical
column to the right. Bed-numbers are indicated in red.

Click on thumbnail to enlarge the image
[Cliquer sur la miniature pour agrandir l'image]
Plate 24:
fig. A. The upper part of the section with a small fault, note bed ANG216 = ANG220. Bed-numbers are indicated in red.
fig. B. The boundary beds between the Barremian and Bedoulian. The boundary is placed in the middle part of ANG202 (based on the occurrence of the ammonite Pa. oglanlensis in a nearby section). Bed-numbers are indicated in red.