◄ Carnets Geol. 14 (4) ►
![]()
Contents
[1. Introduction] [2. Material and methods]
3. Systematic micropaleontology]
[4. Age and distribution]
[5. Palaeoenvironmental interpretation] [6.
conclusions] [Bibliographic references] and ... [Figure cross-comparison]
Museu de Ciències Naturals de Barcelona (Paleontologia), Passeig Picasso s/n, 08003
Barcelona (Spain)
PEMEX, Av. Sitio Grande Nu 2000, Fracc. Carrizal,
Villahermosa, 86038 Tabasco (Mexico)
Departament d'Estratigrafia, Paleontologia i Geociències
Marines, Facultat de Geologia, Universitat de Barcelona, Martí i Franquès s/n, 08028
Barcelona (Spain)
Manuscript online since March 17, 2014
[Editor: Bruno ;
language editor: John ]
![]()
The larger foraminifera found in the material from the Chichen Itzá Formation of Campeche (SE Mexico) were studied in detail. The architectural analyses allow the description of four new species and a new genus, namely, two agglutinated conical foraminifera, Coskinolina yucatanensis n.sp. and Pseudofallotella drobneae n.gen. n.sp., one new alveolinid, Borelis hottingeri n.sp., and one new rotaliid, Dictyoconoides boneti n.sp. The larger foraminifera of the Chichen Itzá Formation are distributed across two different associations: one includes Pseudofallotella drobneae, Neotaberina sp., Rhabdorites sp., Borelis hottingeri, Dictyoconoides boneti, Kathina sp., Miscellaneidae, Ranikothalia soldadensis and Hexagonocyclina cristensis, while the other assemblage is composed of Coskinolina yucatensis, Fallotella causae, Rhabdorites sp., Borelis floridanus and Cincoriola cf. ovoidea. Some of these genera, including Neotaberina and Dictyoconoides, are cited in the Caribbean paleobioprovince for the first time in this paper. The first association is dated as Thanetian (SBZ 3? to SBZ 4) by the presence of Ranikothalia soldadensis. Though the age of the second association remains uncertain, a similar age has been inferred.
Paleocene; Yucatán; Mexico; larger foraminifera; biostratigraphy
V., J.A. & J. (2014).- Paleocene larger foraminifera from the Yucatán Peninsula (SE Mexico).- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 4, p. 41-68.
Les grands foraminifères paléocènes de la péninsule du Yucatán (SE du Mexique).- Les grands foraminifères observés dans du matériel provenant de la Formation Chichen Itzá de l'État de Campeche (SE Mexique) ont été étudiés en détail. Des analyses de leur architecture ont permis de décrire quatre nouvelles espèces et un nouveau genre, deux foraminifères coniques à test agglutinant, Coskinolina yucatanensis n.sp. et Pseudofallotella drobneae n.gen. n.sp., un nouvel alvéolinidé, Borelis hottingeri n.sp., et un nouveau rotaliidé, Dictyoconoides boneti n.sp. Les grands foraminifères de la Formation Chichen Itzá sont groupés dans deux associations distinctes : une première avec Pseudofallotella drobneae, Neotaberina sp., Rhabdorites sp., Borelis hottingeri, Dictyoconoides boneti, Kathina sp., Miscellaneidae, Ranikothalia soldadensis et Hexagonocyclina cristensis, une seconde avec Coskinolina yucatensis, Fallotella causae, Rhabdorites sp., Borelis floridanus et Cincoriola cf. ovoidea. Quelques-uns de ces genres, notamment Neotaberina et Dictyoconoides, sont signalés ici pour la première fois dans la province paléobiogéographique caraïbéenne. La première association est datée du Thanétien (SBZ 3? à SBZ 4) du fait de la présence de Ranikothalia soldadensis. Bien qu'il y ait encore une incertitude quant à l'âge de la seconde association, nous avons retenu la même position stratigraphique.
Paléocène ; Yucatán ; Mexique ; grands foraminifères ; biostratigraphie
Los macroforaminíferos del Paleoceno de la península de Yucatán (SE México).- Los macroforaminíferos encontrados en el material procedente de la Formación Chichen Itzá en Campeche (SE México) han sido estudiados en detalle. Los análisis arquitecturales han permitido la descripción de cuatro nuevas especies y un nuevo género, a saber, dos aglutinados cónicos, Coskinolina yucatanensis n.sp. y Pseudofallotella drobneae n.gen. n.sp.; un nuevo alveolínido, Borelis hottingeri n.sp., y un nuevo rotálido, Dictyoconoides boneti n.sp. Los macroforaminíferos de la Formación Chichen Itzá están distribuidos en dos asociaciones diferentes: una de ellas incluye Pseudofallotella drobneae, Neotaberina sp., Rhabdorites sp., Borelis hottingeri, Dictyoconoides boneti, Kathina sp., Miscellaneidae, Ranikothalia soldadensis y Hexagonocyclina cristensis, mientras que la otra asociación está formada por Coskinolina yucatensis, Fallotella causae, Rhabdorites sp., Borelis floridanus y Cincoriola cf. ovoidea. Algunos de estos géneros, como Neotaberina y Dictyoconoides, se citan en la paleobioprovincia del Caribe por primera vez. La primera asociación ha sido datada como Tanetiense (SBZ 3? a SBZ 4) por la presencia de Ranikothalia soldadensis. A la segunda asociación le ha sido inferida una edad similar.
Paleoceno; Yucatán; México; macroforaminíferos; biostratigrafía
Els macroforaminífers del Paleocè de la península de Yucatán (SE Mèxic).- Els macroforaminífers trobats al material procedent de la Formació Chichen Itzá en Campeche (SE Mèxic) han estat estudiats en detall. Les anàlisis arquitecturals han permès la descripció de quatre noves espècies i un nou gènere, a saber, dos aglutinats cònics, Coskinolina yucatanensis n.sp. i Pseudofallotella drobneae n.gen. n.sp.; un nou alveolínid, Borelis hottingeri n.sp., i un nou rotàlid, Dictyoconoides boneti n.sp. Els macroforaminífers de la Formació Chichen Itzá es troben distribuïts en dues associacions diferents: una inclou Pseudofallotella drobneae, Neotaberina sp., Rhabdorites sp., Borelis hottingeri, Dictyoconoides boneti, Kathina sp., Miscellaneidae, Ranikothalia soldadensis i Hexagonocyclina cristensis, mentre que l'altra està formada per Coskinolina yucatensis, Fallotella causae, Rhabdorites sp., Borelis floridanus i Cincoriola cf. ovoidea. Alguns d'aquests gèneres, com Neotaberina i Dictyoconoides, es citen a la paleobioprovíncia del Carib per primera vegada. La primera associació ha estat datada com Tanetià (SBZ 3? a SBZ 4) per la presència de Ranikothalia soldadensis. A la segona associació li ha estat inferida una edat similar.
Paleocè; Yucatán; Mèxic; macroforaminífers; biostratigrafia
Larger foraminifera represent one of the most successful groups of fossils through geological times. They can be found repeatedly in the geological record from Paleozoic to recent times, but it was during the Permian, Jurassic, Cretaceous and Paleogene when larger foraminifera were particularly important in terms of abundance and taxonomic diversity (, 1982), becoming one of the most important fossil groups in biostratigraphy. The use of larger foraminifera in biostratigraphy is conditional on the correct identification of the taxa, and this identification requires detailed architectural analysis. External and internal characters must be described in detail in order to make correct identifications, which avoid contradictions and inconsistences in the biostratigraphical and paleogeographical distribution of larger foraminiferal taxa and enhance their value as a tool to date shallow-water deposits. A relatively recent example of the usefulness of larger foraminifera in biostratigraphy was the definition of the shallow benthic zones (SBZs) for the Paleogene deposits of the Tethyan paleobioprovince ( et al., 1998). Tethyan Paleogene larger foraminifera were studied and described in detail prior to using them as biostratigraphical markers. This biozonation cannot be extrapolated to the Caribbean paleobioprovince because the Paleogene larger foraminifera of that area are still poorly known and taxonomic revisions are needed in order to clarify their taxonomic and biostratigraphic position. During last century, authors such as (1929, 1945, among others), (1941, 1944, 1957, among others), (1944, 1975, 1996), and (1968), and (1974), and (1975), (1984, 1995), and (1993), (1997), et al. (2007), and (2009) and et al. (2013) have revised the taxa of the Paleogene of the Caribbean bioprovince, but much more research is needed to build a reliable future biozonation equivalent to the SBZ of the Tethyan paleobioprovince.
This paper focuses on the enhancement of the biostratigraphic value of the larger foraminifera taxa found in the Paleocene rocks of the Yucatán Peninsula (SE Mexico). In particular, the main goals of this paper are: 1) to present a detailed taxonomic study of the larger foraminifera, 2) to characterise the larger foraminifera assemblages, assigning ages and paleoenvironmental interpretations, and 3) to make a tentative trans-Atlantic correlation comparing the evolutionary trends in the taxa involved with those of their consorts from the Tethyan paleobioprovince.
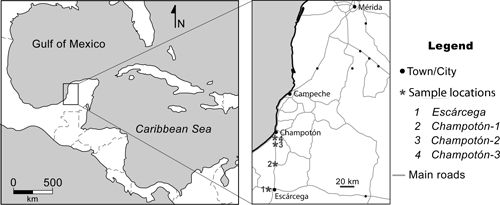
The material was collected during the field trips carried out in the 1990s by the micropalaeontologist Prof. Esmeralda , from the Universitat Autònoma de Barcelona, and PEMEX geologists. The samples studied in this work were collected from the Chichen Itzá Formation in several outcrops along the road between Escárcega and Campeche (Campeche State, SE Mexico) (Fig. 1 ![]() ). A long, continuous stratigraphic section could not be obtained because of the scarcity of large outcrops in this flat area of Mexico. The Chichen Itzá Formation was mentioned for the first time by and
(1960) to describe a group of carbonate rocks outcropping in the surroundings of the archaeological locality of Chichen Itzá. The formation was formally described two years later by the same authors ( & ,
1963), after studying in detail all the Cenozoic formations of the Yucatán Peninsula. In the original description, the Chichen Itzá Fm. was defined as being mainly composed of fossiliferous limestones exhibiting some variations in terms of lithology and age, which led the authors to describe three different superposed members: Xbacal (lower Eocene), Pisté (middle Eocene) and Chumbec (middle-upper Eocene). The fossil assemblages of these members are all very rich in larger
foraminifera, but the material studied in this paper comes exclusively from the lowest unit, the Xbacal Mb. This member consists of thin-bedded to massive limestones and marls (100-200 m thick) that outcrop in the southern sector of the state of Campeche.
). A long, continuous stratigraphic section could not be obtained because of the scarcity of large outcrops in this flat area of Mexico. The Chichen Itzá Formation was mentioned for the first time by and
(1960) to describe a group of carbonate rocks outcropping in the surroundings of the archaeological locality of Chichen Itzá. The formation was formally described two years later by the same authors ( & ,
1963), after studying in detail all the Cenozoic formations of the Yucatán Peninsula. In the original description, the Chichen Itzá Fm. was defined as being mainly composed of fossiliferous limestones exhibiting some variations in terms of lithology and age, which led the authors to describe three different superposed members: Xbacal (lower Eocene), Pisté (middle Eocene) and Chumbec (middle-upper Eocene). The fossil assemblages of these members are all very rich in larger
foraminifera, but the material studied in this paper comes exclusively from the lowest unit, the Xbacal Mb. This member consists of thin-bedded to massive limestones and marls (100-200 m thick) that outcrop in the southern sector of the state of Campeche.
Click on thumbnail to enlarge the image.
Figure 1. Geographical location of the studied samples. Escárcega, X: -90,80566803180; Y: 18,61150448100. Champotón-1, X: -90,71832692450; Y: 18,97268548020. Champotón-2, X: -90,73470254970; Y: 19,19421930200. Champotón-3, X: -90,73379612940; Y:19,26029960940.
The material analysed in this paper was previously studied by (1997), but it was necessary to undertake a comprehensive review of the specimens described there to correct some identifications. Detailed analysis of around 400 random sections of larger foraminifera were used to identify the morphotypes. The figured material from Yucatán Peninsula is housed in the micropaleontological collection of the Universitat Autònoma de Barcelona, Spain, under the catalogue code 95-Me.
Phylum Foraminifera
Order Textulariida & , 1896
Superfamily Ataxophragmioidea , 1877
Family Coskinolinidae , 1965
Genus Coskinolina , 1875
Type species: Coskinolina liburnica , 1875
Remarks: Taking into consideration the description of (1875) and the posterior revisions made by (1974) and and (1980), the genus Coskinolina (type species C. liburnica , 1875) can be defined as follows: agglutinated tests with pseudo-keriothecal texture, early trochospiral, pfenderinid coiling, and later becoming uniserial with numerous low chambers rapidly enlarging in diameter. Chambers exhibit marginal apertures and endoskeleton consisting of interseptal pillars. No exoskeleton is present in the periphery of the chamber lumen.
The pseudo-keriothecal texture in Coskinolina, called "closely spaced, transverse pores" in
(1960), "keriothecal substructure" in and
(1980) and "pseudo-keriothecal wall texture" in and
(2007), has also been observed in specimens of other Paleocene agglutinated conical taxa such as Pfendericonus & ,
1980, and Coleiconus & ,
1980, but it has been difficult to identify among the population of Coskinolina studied in this work. Specifically, the pseudo-keriothecal texture is only visible in some specimens (see
Fig. 3.D ![]() ); in others, it is not observed and this is attributed to it being masked by diagenetic processes. Hence, it is worth underlining the difficulty of using this characteristic to identify taxa.
); in others, it is not observed and this is attributed to it being masked by diagenetic processes. Hence, it is worth underlining the difficulty of using this characteristic to identify taxa.
Coskinolina yucatanensis n.sp.
1997 Coskinon? sp. , p. 78, Pl. 3, figs. 1-6
Holotype. Fig. 2.B ![]() (95-ME-38-01)
(95-ME-38-01)
Paratypes. Figs. 2.A ![]() (95-ME-38-02), 2.H
(95-ME-38-02), 2.H ![]() (95-ME-38-03), 2.I
(95-ME-38-03), 2.I ![]() (95-ME-38-04), 3.L
(95-ME-38-04), 3.L ![]() (95-ME-38-05).
(95-ME-38-05).
Type locality. Champotón-1; X: -90,71832692450; Y: 18,97268548020 (Fig. 1 ![]() ).
).
Type level. Thanetian (SBZ 3? and SBZ 4), Chichen Itzá Formation.
Derivation of name. Named after the Yucatán Peninsula, where the type material was collected.
Material. Sample Champotón-1 (Fig. 1 ![]() ).
).
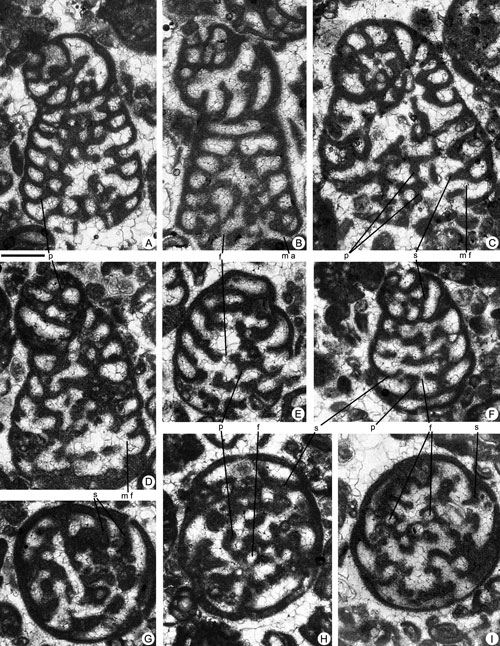
Description. The test is agglutinated with irregular conical morphology. The size of the shell in megalospheric generation reaches about 1.6 mm in length and 1.2 mm in diameter. Microspheric forms have not been found. Megalospheric specimens begin with a spherical proloculus in subapical position of about 0.14 mm in diameter (see
Fig. 3.L ![]() ) and this is followed by 1 ½ to 2 trochospiral whorls. In this early trochospiral stage of growth, 6-7 chambers compose a whorl. The final stage of the ontogeny consists of 7-8 chambers with slightly increasing diameter in a uniserial arrangement (see
Fig. 2.A-B
) and this is followed by 1 ½ to 2 trochospiral whorls. In this early trochospiral stage of growth, 6-7 chambers compose a whorl. The final stage of the ontogeny consists of 7-8 chambers with slightly increasing diameter in a uniserial arrangement (see
Fig. 2.A-B ![]() ). The marginal chamber walls are thick and chamber sutures are almost flush. Adult chambers show an endoskeleton that consists of interseptal pillars (see
Fig. 2.C, F
). The marginal chamber walls are thick and chamber sutures are almost flush. Adult chambers show an endoskeleton that consists of interseptal pillars (see
Fig. 2.C, F ![]() ), continuous from one chamber to the next. The pillared endoskeleton appears later during the trochospiral stage of growth. No exoskeleton is present in the marginal chamber lumen. Numerous foramina are distributed in the centre of the septum (see
Fig. 2.H
), continuous from one chamber to the next. The pillared endoskeleton appears later during the trochospiral stage of growth. No exoskeleton is present in the marginal chamber lumen. Numerous foramina are distributed in the centre of the septum (see
Fig. 2.H ![]() ), this being strongly concave in some cases (see
Fig. 3.E
), this being strongly concave in some cases (see
Fig. 3.E ![]() ). Marginal foramina are smaller in size and arranged in more or less circular pattern. Foramina and pillars are continuously arranged from one chamber to the next.
). Marginal foramina are smaller in size and arranged in more or less circular pattern. Foramina and pillars are continuously arranged from one chamber to the next.
Differences and similarities. The new species C. yucatanensis differs from other species of the genus by having a smaller overall size of the shell and by its low conical -almost cylindrical in some cases- uniserial stage. The megalosphere in C. yucatanensis n.sp. is about 0.14 mm in diameter followed by 12-14 chambers trochospirally arranged in 1 ½ to 2 whorls, whereas in the type species C. liburnica the megalosphere has a diameter of up to 0.4 mm followed by a deuteroconch and 5-11 chambers trochospirally arranged in ½ to 1 ½ whorls. The maximum cone diameter observed in C. yucatanensis n.sp. is around 1.2 mm, and in C. liburnica is 1.75-2 mm. The megalospheric generation of the new species also shows strong differences with respect to the other species attributed to the genera Coskinolina. Coskinolina roberti (, 1905) has a megalosphere of 0.35-0.4 mm in diameter, a nepionic stage composed of about 8-10 chambers arranged in ½ to 1 ½ whorls and a maximum cone diameter of around 3 mm. Coskinolina douvillei (, 1930) shows a megalosphere of around 0.35 mm in diameter followed by a deuteroconch and 6-10 chambers arranged in a trochospiral (almost planispiral in some cases) of about half a whorl. Coskinolina perpera & , 1980, exhibit a megalosphere of 0.34-0.45 mm followed by a large deuteroconch and 5-11 nepionic chambers trochospirally arranged in 1/3 to 2/3 whorls.
Three other species have been reported as belonging to the genus Coskinolina: C. depressa (, 1952) and C. dispansa (, 1952) described from the Middle-Upper Eocene of Somalia by (1952), and C. cuvillieri (, 1960) found in the Upper Eocene of France (, 1960). Though the differences between these three species are still under debate due to a lack of material and of detailed studies, the Caribbean species C. yucatanensis n.sp. can be clearly differentiated from these three Tethyan relatives by its smaller size and its nepionic trochospiral architecture.
Finally, the Eocene Indo-Pacific species Coskinolina rotaliformis (, 1957) is not a truly Coskinolina because it shows a simple exoskeleton based on spaced beams (see Pl. 233, figs. 11-26 in , 1957).
Associated larger foraminifera. This form occurs associated with Fallotella causae , 1997, Cincoriola cf. ovoidea (, 1958), Rhabdorites sp. and Borelis floridanus , 1941.
Click on thumbnail to enlarge the image.
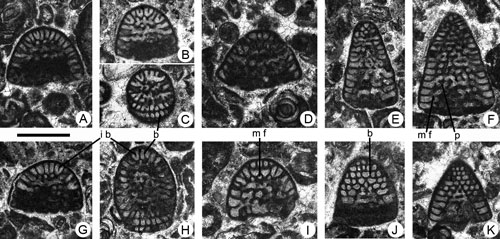
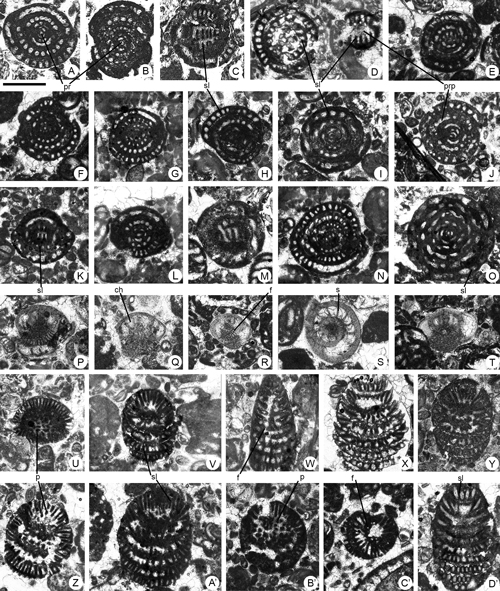
Figure 2. Coskinolina yucatanensis n.sp. from the Paleocene of Campeche (SE Mexico). Scale bar = 0.3 mm. A-F, longitudinal, slightly oblique sections. G-I, transversal sections. A-B, C-I, megalospheric specimens. C, microspheric? specimen. A, 95-Me-38-02 (Paratype). B, 95-Me-38-01 (Holotype). C, 95-Me-38-06. D, 95-Me-38-07. E, 95-Me-38-08. F, 95-Me-38-09. G, 95-Me-38-10. H, 95-Me-38-03 (Paratype). I, 95-Me-38-04 (Paratype).
Abbreviations: f: foramen; m a: marginal aperture; m f: marginal foramen; p: pillar; s: septum. See , 2006, for definition of terms.
Click on thumbnail to enlarge the image.
Figure 3. Coskinolina yucatanensis n.sp. from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm for pictures A-C and E-O; 0.075 mm for picture D. A-C, G-O, oblique sections. Note the embryo in apical position in picture L. D, detail of the chamber wall of the specimen 3 showing the pseudo-keriothecal structure. E-F, longitudinal sections. A, 95-Me-38-11-12. B, 95-Me-38-13. C, 95-Me-38-14. D, 95-Me-38-15. E, 95-Me-38-16. F, 95-Me-38-17. G, 95-Me-38-18. H, 95-Me-38-19. I, 95-Me-38-20. J, 95-Me-38-21. K, 95-Me-38-05. L, 95-Me-38-22 (Paratype). M, 95-Me-38-23. N, 95-Me-38-24. O, 95-Me-38-56.
Abbreviations: ch: chamber; f: foramen; p: pillar; s: septum. See , 2006, for definition of terms.
Superfamily Orbitolinoidea , 1890
Family Orbitolinidae , 1980
Subfamily Dictyoconinae , 1965
Genus Fallotella , 1954
Type species Fallotella alavensis , 1954
Description. The shell is agglutinated with conical shape and a reduced trochospiral early stage of growth. The chamber wall is thin without any ultrastructure. The megalosphere is located in an apical position and it is followed by a kidney-shaped deuteroconch. The external wall is relatively thick (no epidermis). The exoskeleton consists of first and second order radial beams. No rafters are present, but a slight ridge or thickening parallel to the septa is located at the central part of the chamber, inside the surface of the external wall. The endoskeleton consists of pillars with a rounded transverse section that changes to bean-shaped or irregular in the distal part of the chamber. The apertural face is slightly convex and shows apertures distributed in the central part and in a single row around the periphery (marginal apertures).
Remarks. According to one of the most important papers dealing with Tertiary conical larger foraminifera ( & , 1980), the genus Fallotella has a simple to moderately complex exoskeleton "consisting either of beams only or of beams and rafters". Among the "Fallotella morphotypes" figured in the present paper and those of and (1980), the nature of the external wall is similar: somewhat thinner than in Ataxophragmiids and thicker than an orbitolinid epiderm, but two different types of exoskeleton can be distinguished, with and without rafters. The exoskeleton of the adult chambers of the type species F. alavensis consists of two orders of beams (primary and secondary), no rafters are present but only a slight central thickening in later stages of growth that cannot be considered a true rafter. Therefore, the forms having a more complex exoskeleton consisting of one generation of rafters and several generations of beams should be removed from this genus.
Fallotella causae , 1997 emend.
1997 Fallotella causi n.sp. , p. 71-73; Pl. 1, figs. 1-5, Pl. 2, fig. 5
2007 Fallotella sp. et al., p. 365; Pl. 1, fig. 11
Paralectotypes. Fig. 4.B ![]() (95-ME-36-02), 4.C
(95-ME-36-02), 4.C ![]() (95-ME-36-03), 4.I
(95-ME-36-03), 4.I ![]() (95-ME-36-04), 4.J
(95-ME-36-04), 4.J ![]() (95-ME-36-05), 4.K
(95-ME-36-05), 4.K ![]() (95-ME-36-06).
(95-ME-36-06).
Type locality. Escárcega; X: -90,80566803180; Y: 18,61150448100 (Fig. 1 ![]() ).
).
Type level. Thanetian (SBZ 3? and SBZ 4), Chichen Itzá Formation.
Derivation of name. Given in honour of Prof. Esmeralda (Universitat Autònoma de Barcelona), distinguished specialist of larger foraminifera.
Material. Samples Escárcega and Champotón-1 (Fig. 1 ![]() ).
).
Remarks on the emendation. The original name given by (1997) to this species was F. causi, named after Prof. Esmeralda . The International Code of Zoological Nomenclature (1999) states "31.1.2. A species-group name, if a noun in the genitive case (see Article 11.9.1.3) formed directly from a modern personal name, is to be formed by adding to the stem of that name -i if the personal name is that of a man, -orum if of men or of man (men) and woman (women) together, -ae if of a woman, and -arum if of women; [...]". Following these indications, the name of the species F. causi has been emended to F. causae. Moreover, it was necessary to designate paralectotypes, which show the architectural characteristics not visible in the holotype of (1997).
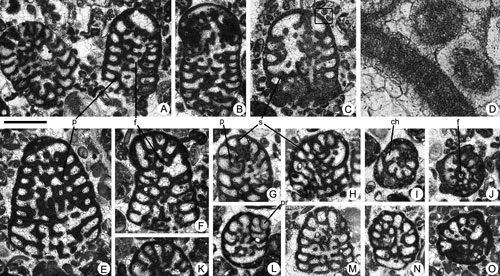
Description. The test is agglutinated and high conical in shape. The dimorphism between microspheric and megalospheric forms is restricted to the early spiral stages. The cone surface is smooth and the wall is thin. The nepionic stage of the megalospheric generation consists of a protoconch with a diameter of 0.12-0.14 mm in apical position followed by a small, kidney-shaped deuteroconch (see
Fig. 4.F ![]() ). The next 3-5 chambers are spirally arranged in a very low trochospiral. The sections of the specimens interpreted as microspheric forms by their early, tight stages of growth are not perfectly centred
(Fig. 4.B
). The next 3-5 chambers are spirally arranged in a very low trochospiral. The sections of the specimens interpreted as microspheric forms by their early, tight stages of growth are not perfectly centred
(Fig. 4.B ![]() ), thus their embryos cannot be accurately ascertained. The adult conical shell shows a maximum axial length of 1.5 mm with a maximum diameter observed of around 1 mm. There are 13-14 chambers per 1 mm of axial cone length. The endoskeleton and exoskeleton appear early in the course of ontogeny, later in the spiral, nepionic stage or in the first chamber of the uniserial, adult stage (see
Fig. 4.I
), thus their embryos cannot be accurately ascertained. The adult conical shell shows a maximum axial length of 1.5 mm with a maximum diameter observed of around 1 mm. There are 13-14 chambers per 1 mm of axial cone length. The endoskeleton and exoskeleton appear early in the course of ontogeny, later in the spiral, nepionic stage or in the first chamber of the uniserial, adult stage (see
Fig. 4.I ![]() ). The exoskeleton consists of beams of first and second order intercalated (see
Fig. 4.J, M
). The exoskeleton consists of beams of first and second order intercalated (see
Fig. 4.J, M ![]() ), larger and shorter respectively. There are no rafters, but a slight thickening in the peripheral part of the chamber has been observed in the latest stages of growth (see
Fig. 4.B
), larger and shorter respectively. There are no rafters, but a slight thickening in the peripheral part of the chamber has been observed in the latest stages of growth (see
Fig. 4.B ![]() ). In young specimens with a cone diameter of 0.5 mm, there are only 4 beams per quadrant (see
Fig. 4.L
). In young specimens with a cone diameter of 0.5 mm, there are only 4 beams per quadrant (see
Fig. 4.L ![]() ), while in the adult forms, with a cone diameter about 1 mm, there are 5-6 beams and 4-6 intercalated beams per quadrant. The endoskeleton is composed of thin pillars with a subcircular transverse section in the proximal part of the chamber, and bean-shaped or irregular outline towards the distal part (see
Fig. 4.M
), while in the adult forms, with a cone diameter about 1 mm, there are 5-6 beams and 4-6 intercalated beams per quadrant. The endoskeleton is composed of thin pillars with a subcircular transverse section in the proximal part of the chamber, and bean-shaped or irregular outline towards the distal part (see
Fig. 4.M ![]() ). The apertures are relatively large and distributed in the central part of the apertural face.
). The apertures are relatively large and distributed in the central part of the apertural face.
Differences and similarities. Fallotella causae differs from the type species F. alavensis mainly by its smaller size. The former shows a protoconch with a diameter of 0.12-0.14 mm, 1.5 mm of axial cone length and 1 mm of maximum cone diameter. The latter exhibits a protoconch with a diameter of 0.15-0.18 mm, 2.1 mm of axial cone length and 1.5 mm of maximum cone diameter.
Remarks. The other species ascribed to the genus Fallotella, as F. cookei (, 1928), show well-developed rafters. Therefore, it should be removed from the genus Fallotella.
Associated larger foraminifera. This form occurs associated with Coskinolina yucatensis n.sp., Cincoriola cf. ovoidea, Rhabdorites sp. and Borelis floridanus.
Click on thumbnail to enlarge the image.
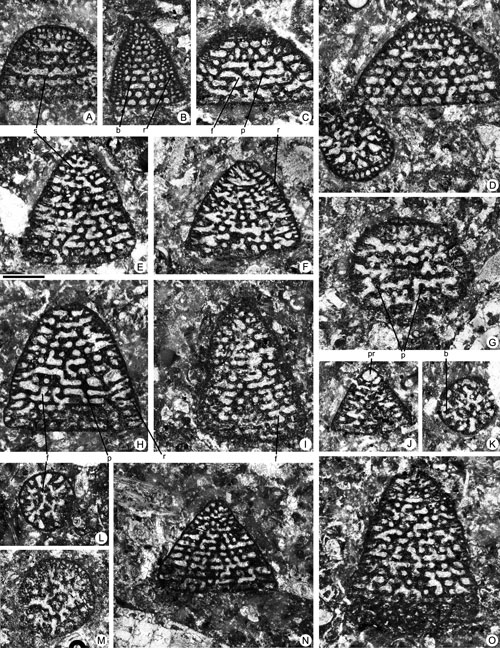
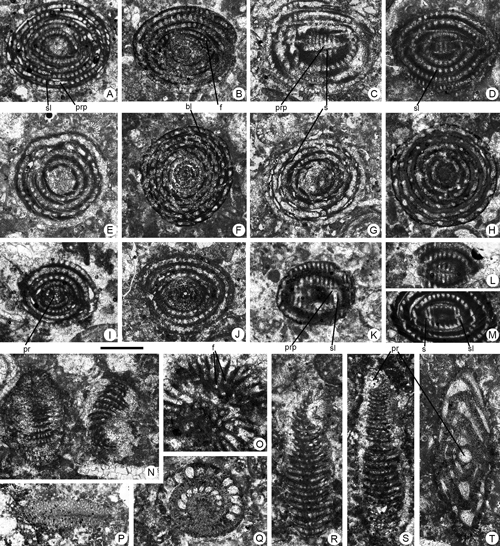
Figure 4. Fallotella causae , 1997, from the Paleocene of Campeche (SE Mexico). Scale bar = 0.3 mm. A, D-G, I, axial sections. B-C, H, longitudinal sections close to the axial plane. J-M, transversal sections. A, 95-Me-36-01. B, 95-Me-36-02 (Paralectotype). C, 95-Me-36-03 (Paralectotype). D, 95-Me-36-07. E, 95-Me-36-08. F, 95-Me-36-09. G, 95-Me-36-10. H, 95-Me-36-11. I, 95-Me-36-04 (Paralectotype). J, 95-Me-36-05 (Paralectotype). K, 95-Me-36-06 (Paralectotype). L, 95-Me-36-12. M, 95-Me-36-13.
Abbreviations: b: beam; i b: intercalary beam; f: foramen; m t: marginal thickening; p: pillar; pr: proloculus. See , 2006, for definition of terms.
Click on thumbnail to enlarge the image.
Figure 5. Fallotella causae , 1997, from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm. A-D, G-K, oblique sections. E-F, longitudinal sections. A, 95-Me-36-14. B, 95-Me-36-15. C, 95-Me-36-16. D, 95-Me-36-17. E, 95-Me-36-18. F, 95-Me-36-19. G, 95-Me-36-20. H, 95-Me-36-21. I, 95-Me-36-22. J, 95-Me-36-23. K, 95-Me-36-24.
Abbreviations: b: beam; i b: intercalary beam; m f: marginal foramen (aperture); p: pillar. See , 2006, for definition of terms.
Genus Pseudofallotella n.gen.
Derivation of name: Pseudo- (latin word of "false") has been given due to the resemblance to Fallotella specimens.
Type species. Pseudofallotella drobneae n.sp.
Diagnosis. Agglutinated, high to low-conical shells. The chamber wall is thin with no ultrastructure. In megalospheric generation, embryo placed in apical position and composed of protoconch followed by deuteroconch. Embryo of microspheric generation unknown. Adult discoidal chambers increasing in diameter through course of ontogeny; uniserially arranged. Exoskeleton of one generation of rafters and primary, secondary and sporadically, tertiary beams. Endoskeleton of pillars alternating in position from one chamber to the next. Pillars with a rounded periphery in transversal section, changing to bean-shaped towards distal part of chamber. Apertural face flat to slightly convex with multiple apertures. Apertures distributed in centre of apertural face and in single row at periphery (marginal apertures).
Remarks. As pointed out above, and (1980) grouped together different morphotypes with "simple exoskeletons with or without one rafter series but combined with a thick, structure less marginal chamber wall". In fact, the genus Fallotella grouped forms with both types of exoskeleton, consisting of rafters or equatorial slight thickening ("rudimentary rafters"). These authors also affirmed that the characteristics of the marginal wall and the type of the exoskeleton reflect basic cellular activities. This assertion would support using both types of exoskeleton as criteria to separate taxa. However, because of the complexity of some larger foraminifera groups, in particular the Tertiary conical group, a precise hierarchy of the morphological characters used in taxonomic classification has not yet been agreed on, despite the efforts of many authors (see & , 1987, introduction to , 2006, and & , 2007). Such is the case of the families Coskinolinidae and Orbitolinidae. Some taxa ascribed to them have been transferred from subgeneric to generic status in subsequent research (e.g., Coleiconus, Coskinon, Daviesiconus, etc.). In any case, it is generally accepted that larger foraminifera species are defined in terms of dimensions, and the presence or absence of architectural elements must be considered as criteria of supraspecific rank.
Taking all these issues into consideration and after studying many morphotypes of conical larger foraminifera, the two, Fallotella and Pseudofallotella exoskeleton types can easily be distinguished in random thin sections. Leaving aside the biological or ecological meaning of the presence of a rafter in the marginal chamber wall, the usefulness of using this feature of the exoskeleton to identify taxa in almost all types of random sections from centred to tangential sections and considering the generic criterion of presence/absence of architectural elements, have lead us to consider these two types of exoskeleton as diagnostic criteria of generic rank. Therefore, a new genus Pseudofallotella has been created with the aim of bringing together all the forms previously ascribed to the genus Fallotella but differing with its type species F. alavensis by having a dissimilar exoskeleton exhibiting rafters from early stages of growth. The species F. cookei, F. kochanskae and F. kochanskae persica, which also show well developed rafters, should be revised as they should likely be assigned to the new genus Pseudofallotella.
Pseudofallotella drobneae n.sp.
1997 Dictyoconus sp. , p. 76; Pl. 2, figs. 1-2; Pl. 4, figs. 1-5
Holotype. Fig. 7.E ![]() (95-ME-47-01).
(95-ME-47-01).
Paratypes. Figs. 6.O ![]() (95-ME-47-02), 7.A
(95-ME-47-02), 7.A ![]() (95-ME-47-03), 7.C
(95-ME-47-03), 7.C ![]() (95-ME-47-04).
(95-ME-47-04).
Type locality. Champotón-3, X: -90,73379612940; Y: 19,26029960940 (Fig. 1 ![]() ).
).
Type level. Thanetian (SBZ 3? and SBZ 4), Chichen Itzá Formation.
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Derivation of name. Given in honour of Dr Katica , distinguished specialist of larger foraminifera.
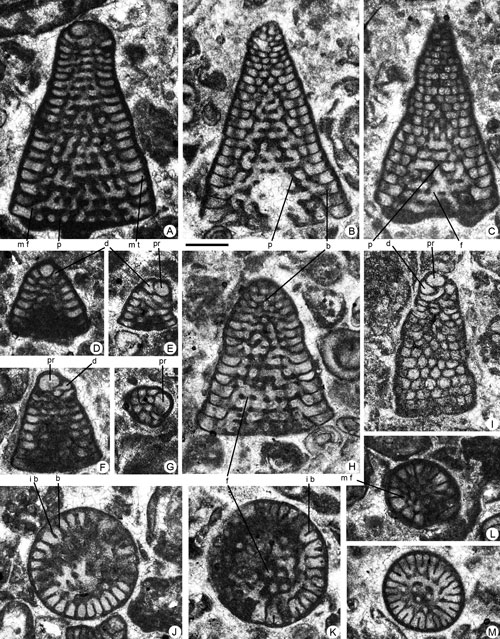
Description. Specimens show medium to high conical shells. The wall is thin and the cone surface is smooth. The megalospheric and microspheric generations show a slight dimorphism. The nepionic stage of megalospheric generation begins with a sub-spherical protoconch of around 0.22 mm in apical, slightly eccentric position and a bean-shaped deuteroconch
(Fig. 6.J ![]() ). The embryo is followed by uniserial chambers that rapidly acquire a discoidal shape exhibiting endo- and exoskeleton. The nepionic stage of the microspheric generation is unknown. Megalospheric forms have a maximum axial length of around 1.4 mm and a maximum cone diameter of 1.8 mm. The exoskeleton is constituted by two to three intercalary beams (secondary and tertiary) between adjacent primary beams (see
Fig. 7.A, D
). The embryo is followed by uniserial chambers that rapidly acquire a discoidal shape exhibiting endo- and exoskeleton. The nepionic stage of the microspheric generation is unknown. Megalospheric forms have a maximum axial length of around 1.4 mm and a maximum cone diameter of 1.8 mm. The exoskeleton is constituted by two to three intercalary beams (secondary and tertiary) between adjacent primary beams (see
Fig. 7.A, D ![]() ), and one generation of rafters (see
Figs. 7.K
), and one generation of rafters (see
Figs. 7.K ![]() ,
8.J
,
8.J ![]() ). Large specimens with a maximum axial length of 2.5 mm and the maximum cone diameter of 2.4 mm have been interpreted as belonging to the microspheric generation
(Figs. 6.O
). Large specimens with a maximum axial length of 2.5 mm and the maximum cone diameter of 2.4 mm have been interpreted as belonging to the microspheric generation
(Figs. 6.O ![]() ,
7.F
,
7.F ![]() ). The rafters are more developed in large forms (possibly microspheric forms). At a cone diameter of between 0.7-1 mm there are 5-7 primary beams and 10-11 intercalated beams. The apertural face is slightly convex and shows a cribate distribution with marginal apertures in a circular arrangement (see
Fig. 7.A
). The rafters are more developed in large forms (possibly microspheric forms). At a cone diameter of between 0.7-1 mm there are 5-7 primary beams and 10-11 intercalated beams. The apertural face is slightly convex and shows a cribate distribution with marginal apertures in a circular arrangement (see
Fig. 7.A ![]() ).
).
Differences and similarities. The specimens of F. cookei differ from P. drobneae n.sp. by their more flattened shell of low conical shape and larger megalosphere. The new species P. drobneae exhibits a protoconch of 0.20-0.22 mm and maximum cone diameters of 1.8 mm for A forms and 2.4 mm for B forms. Fallotella cookei shows a protoconch of 0.28-0.32 mm and the maximum cone diameters observed are 2.5 mm for A forms and 3.9 mm for B forms. P. drobneae n.sp. differs from F. kochanskae and F. kochanskae persica by its larger shell. The maximum axial length in F. kochanskae and F. kochanskae persica does not reach more than 1.4 mm, whereas there are specimens of P. drobneae n.sp. as long as 2.5 mm.
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis ( & , 1941), Hexagonocyclina cristensis (, 1924), Dictyoconoides boneti n.sp., Kathina sp., Miscellaneidae indet., Neotaberina sp., Rhabdorites sp. and Borelis hottingeri n.sp.
Click on thumbnail to enlarge the image.
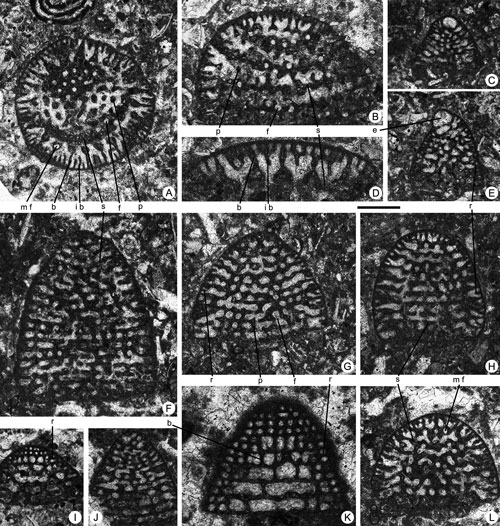
Figure 6. Pseudofallotella drobneae n.gen. n.sp. from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm. A, C, D, G, oblique sections. B, E-F, H-I, N-O, longitudinal sections. J, axial section. K-M, transverse sections. A, 95-Me-47-09. B, 95-Me-47-10. C, 95-Me-47-11. D, 95-Me-47-12-13. E, 95-Me-47-14. F, 95-Me-47-15. G, 95-Me-47-16. H, 95-Me-47-17. I, 95-Me-47-18. J, 95-Me-47-19. K, 95-Me-47-20. L, 95-Me-47-21. M, 95-Me-47-22. N, 95-Me-47-23. O, 95-Me-47-02 (Paratype).
Abbreviations: b: beam; f: foramen; p: pillar; r: rafter; s: septum. See , 2006, for definition of terms.
Click on thumbnail to enlarge the image.
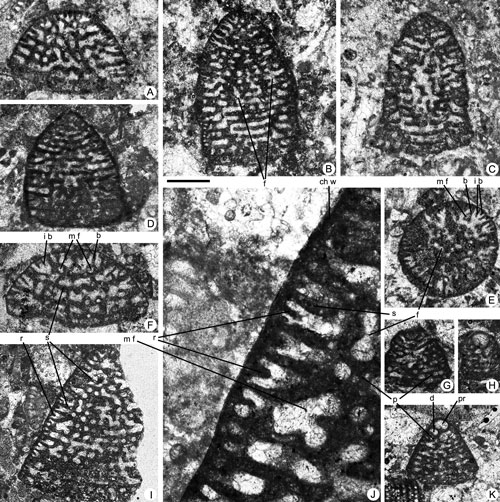
Figure 7. Pseudofallotella drobneae n.gen. n.sp. from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm for pictures A-C and E-L; 0.3 mm for picture D. A, transverse, oblique section. B, D, G-H, L, oblique sections. C, E, axial sections. F, J, longitudinal sections. I, K, tangential sections. A, 95-Me-47-03 (Paratype). B, 95-Me-47-24. C, 95-Me-47-04 (Paratype). D, 95-Me-47-25. E, 95-Me-47-01 (Holotype). F, 95-Me-47-26. G, 95-Me-47-27. H, 95-Me-47-28. I, 95-Me-47-29. J, 95-Me-47-30. K, 95-Me-47-31. L, 95-Me-47-32.
Abbreviations: f: foramen; p: pillar; r: rafter; s: septum; b: beam; i b: intercalary beam; m f: marginal foramen (aperture); pr: proloculus. See , 2006, for definition of terms.
Click on thumbnail to enlarge the image.
Figure 8. Pseudofallotella drobneae n.gen. n.sp. from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm for pictures A-I and K; 0.15 mm for picture J. A, F, oblique sections. B-D, G, I, longitudinal sections. E, transversal section. H, K, axial section. J, detail of the marginal chamber wall of specimen I. A, 95-Me-46-06. B, 95-Me-46-07. C, 95-Me-46-08. D, 95-Me-46-09. E, 95-Me-46-10. F, 95-Me-46-11. G, 95-Me-46-12. H, 95-Me-46-13. I, 95-Me-46-14. J, 95-Me-46-15. K, 95-Me-46-30.
Abbreviations: d: deuteroconch; f: foramen; b: beam; i b: intercalary beam; m f: marginal foramen (aperture); chw: chamber wall; p: pillar; pr: proloculus; r: rafter; s: septum. See v for definition of terms.
Order Miliolida & , 1896
Superfamily Alveolinacea , 1839
Family Alveolinidae , 1839
Genus Borelis , 1808
Type species: Borelis melonoides , 1808 = Nautilus melo & , 1798
Remarks. This genus is characterized by a glomerulus with streptospiral arrangement, septula aligned from one chamber to the next, preseptal passage and apertures in a single row. The absence of floors in the chamberlets of our specimens implies that they do not belong to Quasiborelis , 1967.
In the Tethyan domain, the genus Borelis appears in the geological record for the first time in Neogene deposits. The presence of this genus in the Paleocene rocks of Yucatán would indicate that it occupied the ecological niche of Glomalveolina , 1962, which is very abundant in the Tethys but absent in the Caribbean paleobioprovince.
Borelis floridanus , 1941
1941 Borelis gunteri var. floridana. , p. 35; Pl. 18, figs. 3-4
1944 Borelis floridana . , p. 121; Pl. 9, figs. 1-5
1957 Borelis floridanus . , p. 143; Pl. 2, figs. 3-4
1960 Borelis floridanus . & , p. 11; Pl. 1, fig. 7
non 1960 Borelis floridanus . & , p. 11; Pl. 1, figs. 4-6
1981 Quasiborelis floridanus (). , p. 24; Pl. 4, fig. 4
1997 Quasiborelis? floridanus (). , p. 82-83; Pl. 5, fig.s 1-2, 4-8; Pl. 6, figs. 5-6
2013 "Quasiborelis" floridanus . et al., p. 176; figs. 5.12-5.14
Material. Samples Champotón-1 (Fig. 1 ![]() ).
).
Description. Specimens have porcelaneous, spherical shells. Dimorphism has not been observed. The nepionic stage (glomerulus) consists of a small proloculus of around 4 to 6 µm followed by one to two whorls of non-subdivided chambers streptospirally arranged
(Fig. 9.A-B ![]() ). The adult chambers are arranged in a planispiral growth pattern and they show septula subdividing the chamber lumen into chamberlets. Septula are aligned from chamber to chamber
(Fig. 9.A-C
). The adult chambers are arranged in a planispiral growth pattern and they show septula subdividing the chamber lumen into chamberlets. Septula are aligned from chamber to chamber
(Fig. 9.A-C ![]() ) and the preseptal passage occupies 1/3rd of the chamber. No postseptal passage has been identified. The outline of the chamberlets in axial section is subcircular in the first whorls and subrectangular, higher than wide, in the latest stages of growth. The apertures are aligned in a single row. The maximum equatorial diameter observed is about 1 mm.
) and the preseptal passage occupies 1/3rd of the chamber. No postseptal passage has been identified. The outline of the chamberlets in axial section is subcircular in the first whorls and subrectangular, higher than wide, in the latest stages of growth. The apertures are aligned in a single row. The maximum equatorial diameter observed is about 1 mm.
Associated larger foraminifera. This form occurs associated with Fallotella causae, Coskinolina yucatensis, Rhabdorites sp. and Cincoriola cf. ovoidea.
Click on thumbnail to enlarge the image.
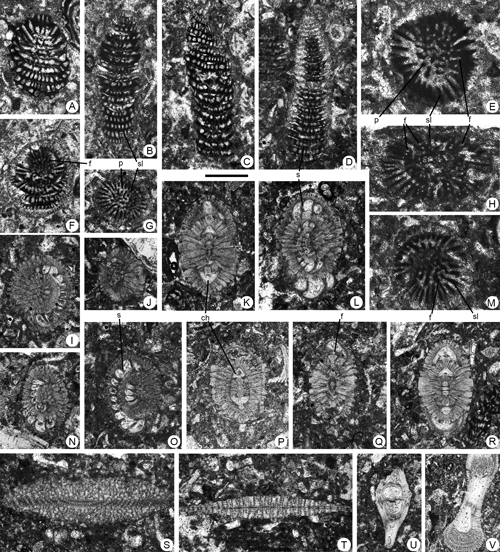
Figure 9. Borelis floridanus , 1941 (A-O), Cincoriola cf. ovoidea (, 1958) (P-T), Neotaberina sp. (U-V, X-B') and Rhabdorites sp. (W, C'-D') from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm. A-B, F-H, J, L, axial sections. C-D, K, M, tangential sections. E, I, U-D', oblique sections. O, equatorial, slightly oblique section. P-R, T, axial, slightly oblique sections. S, transverse section. Note the aligned position of the foramina between two adjacent chambers in Cincoriola cf. ovoidea (, 1958) (Q-R). A, 95-Me-38-25. B, 95-Me-38-26. C, 95-Me-38-27. D, 95-Me-38-28-29. E, 95-Me-38-30. F, 95-Me-38-31. G, 95-Me-38-32. H, 95-Me-38-33. I, 95-Me-38-34. J, 95-Me-38-35. K, 95-Me-38-36. L, 95-Me-38-37. M, 95-Me-38-38. N, 95-Me-38-39. O, 95-Me-38-40. P, 95-Me-38-41. Q, 95-Me-38-42. R, 95-Me-38-43. S, 95-Me-38-44. T, 95-Me-38-45. U, 95-Me-38-46. V, 95-Me-38-47. W, 95-Me-38-48. X, 95-Me-38-49. Y, 95-Me-38-50. Z, 95-Me-38-51. A', 95-Me-38-52. B', 95-Me-38-53. C', 95-Me-38-54. D', 95-Me-38-55.
Abbreviations: ch: chamber; f: foramen; p: pile; pr: proloculus; prp: preseptal space; s: septum; sl: septulum. See , 2006, for definition of terms.
Borelis hottingeri n.sp.
1960 Borelis floridanus . & , p. 11; Pl. 1, figs. 4, 6
1997 "Borelis" sp. , p. 84; Pl. 6, figs. 7-8; Pl. 7, figs. 1-8
Holotype. Fig. 10.I ![]() (95-ME-46-01)
(95-ME-46-01)
Paratypes. Fig. 10.A ![]() (95-ME-46-02), 10.F
(95-ME-46-02), 10.F ![]() (95-ME-46-03), 10.H
(95-ME-46-03), 10.H ![]() (95-ME-46-04), 10.K
(95-ME-46-04), 10.K ![]() (95-ME-46-05).
(95-ME-46-05).
Type locality. Champotón-2, X: -90,73470254970; Y: 19,19421930200 (Fig. 1 ![]() ).
).
Type level. Thanetian (SBZ 3? and SBZ 4), Chichen Itzá Formation.
Derivation of name. Given in honour of Prof. Lukas , distinguished specialist of larger foraminifera.
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Description. The shell has a porcelaneous, spherical to ovoidal morphology. Dimorphism has not been observed. The early stages of growth of the megalospheric generation (glomerulus) consist of a small proloculus of about 5 µm in diameter followed by streptospiral whorls
(Fig. 10.I ![]() ). The adult chambers are arranged planispirally and are subdivided into chamberlets by septula that are aligned from one chamber to the next (see
Fig. 10.C, L
). The adult chambers are arranged planispirally and are subdivided into chamberlets by septula that are aligned from one chamber to the next (see
Fig. 10.C, L ![]() ). The preseptal passage extends from pole to pole and occupies about 1/3rd of the chamber. No postseptal passage has been identified. The apertures are aligned in a single row
(Fig. 10.B
). The preseptal passage extends from pole to pole and occupies about 1/3rd of the chamber. No postseptal passage has been identified. The apertures are aligned in a single row
(Fig. 10.B ![]() ). The maximum equatorial diameter observed is about 1.2 mm. The specimens studied are characterized by having a slight flosculinisation in their chambers (thickening of the basal layer in the equatorial zone). The outline of the chamberlets in axial section is circular to subrectangular. For 7-8 planispiral whorls the axial diameter is around 1.5 mm, equatorial diameter around 1.2 mm and the elongation index is 1.25.
). The maximum equatorial diameter observed is about 1.2 mm. The specimens studied are characterized by having a slight flosculinisation in their chambers (thickening of the basal layer in the equatorial zone). The outline of the chamberlets in axial section is circular to subrectangular. For 7-8 planispiral whorls the axial diameter is around 1.5 mm, equatorial diameter around 1.2 mm and the elongation index is 1.25.
Click on thumbnail to enlarge the image.
Figure 10. Borelis hottingeri n.sp. (A-M), Rhabdorites sp. (N-O, R-S), Hexagonocyclina cristensis (, 1924) (P), and Ranikothalia soldadensis ( & , 1941) (Q, T) from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm for pictures A-N, P-S; 0.3 mm for picture O. A, oblique-centred section. B, D, oblique sections close to the axial plane. C, K-M, tangential sections. E, G, Q, oblique sections close to the equatorial plane. F, equatorial section. H, subequatorial section. I, J, R-S, axial sections. P, oblique section. N, tangential (left) and equatorial section (right). Note that the shells are partially dissolved. O, transverse, slightly oblique section. Note the absence of pillars in the central part of the chamber, typical from the genus Rhabdorites. A, 95-Me-46-02 (Paratype). B, 95-Me-46-16. C, 95-Me-46-17. D, 95-Me-46-18. E, 95-Me-46-19. F, 95-Me-46-03 (Paratype). G, 95-Me-46-30. H, 95-Me-46-04 (Paratype). I, 95-Me-46-01 (Holotype). J, 95-Me-46-20. K, 95-Me-46-05 (Paratype). L, 95-Me-46-21. M, 95-Me-47-33. N, 95-Me-46-22-23. O, 95-Me-46-24. P, 95-Me-46-25. Q, 95-Me-46-26. R, 95-Me-46-27. S, 95-Me-46-28. T, 95-Me-46-29.
Abbreviations: bl: basal layer; f: foramen; pr: proloculus; prp: preseptal space; s: septum; sl: septulum. See , 2006, for definition of terms.
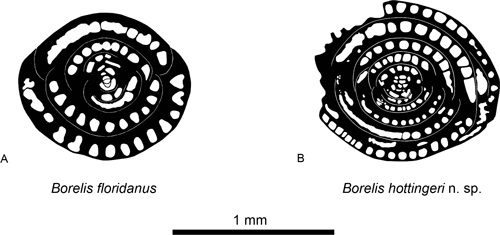
Differences and similarities. Borelis hottingeri n.sp. differs from B. floridanus by its larger size, its more elongated morphology and its tighter spiral
(Fig. 11 ![]() ).
).
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti, Kathina sp., Miscellaneidae indet., Neotaberina sp., Rhabdorites sp. and Pseudofallotella drobneae.
Click on thumbnail to enlarge the image.
Figure 11. 1, Drawing of Borelis floridanus , 1941 (specimen represented in Fig. 9.A). 2, Drawing of Borelis hottingeri n.sp. (specimen represented in Fig. 10.I). Note the differences between the two species in the outline of the chamberlets in axial section and the size of the glomerulus. Scale bar = 1 mm.
Superfamily Soritoidea , 1839
Family Soritidae? , 1839
Remarks. The Soritidae Family, as conceived by and (1987), includes several morphotypes with very dissimilar test features. Soritidae includes specimens with simple endoskeleton, cylindrical/conical tests and simple aperture (e.g., genus Praerhapydionina , 1943), but also includes morphotypes with complex endoskeleton, discoidal tests and multiple apertures (e.g., archaiasines). This complex group (soritids sensu lato) requires revision prior to correctly assigning morphotypes to different families or subfamilies. We tentatively include Neotaberina , 2007, and Rhabdorites , 1996, within this family due to their architectural similarities to some soritids.
Genus Neotaberina , 2007
Type species: Neotaberina neaniconica , 2007
Neotaberina sp.
Figs. 9.U-V, X-B' ![]() , 12.C, E-H, M
, 12.C, E-H, M ![]()
1997 Pseudorhapydionina?, , Pl. 9, fig. 9
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Description. Specimens show porcelaneous, conical to sub-cylindrical shells. The embryonic and nepionic stages are not adequately known, but the specimens of this species seems to start with a small proloculus followed by chambers arranged planispirally. In the ephebic growth stage chambers acquire a uniserial arrangement resulting in a conical to sub-cylindrical shell-shape
(Figs. 9.A' ![]() ,
12.C
,
12.C ![]() ). The apertural face is convex. The adult chambers show an endoskeleton consisting of septula and pillars
(Fig. 12.E, G
). The apertural face is convex. The adult chambers show an endoskeleton consisting of septula and pillars
(Fig. 12.E, G ![]() ). Radial septula alternate regularly with radial rows of foramina
(Fig. 12.E, H
). Radial septula alternate regularly with radial rows of foramina
(Fig. 12.E, H ![]() ). The septula are interrupted in the adaxial area of the chamber, where pillars take place. The endoskeletal elements and foramina are aligned from one chamber to the next. The morphotype studied reach a maximum axial length of 1.6 mm and a diameter of 0.7 mm.
). The septula are interrupted in the adaxial area of the chamber, where pillars take place. The endoskeletal elements and foramina are aligned from one chamber to the next. The morphotype studied reach a maximum axial length of 1.6 mm and a diameter of 0.7 mm.
Differences and similarities. The specimens of Neotaberina from the Paleocene of SE Mexico described in this work can be clearly differentiated from the type species N. neaniconica , 2007, from the late Middle Eocene of Iran by its size and complexity. Neotaberina neaniconica is characterized by reaching an axial length of at least 2.5 mm and a diameter of the cone of 1.8 mm, whereas Neotaberina sp. shows an axial length of 1.6 mm and a diameter of 0.7 mm. Moreover, the latter is simpler exhibiting fewer septula per mm.
Remarks. The genus Neotaberina is first mentioned for the Caribbean area in this work. Taking into consideration the associated larger foraminifers and the nature of the test (size and complexity) of Neotaberina sp., this morphotype seems to be older than the Tethyan Eocene form N. neaniconica. Unfortunately, the absence in our material of sections of the nepionic stage means that the Caribbean morphotype cannot be described as a new species.
Associated larger foraminifera. This form occurs associated with Pseudofallotella drobneae, Borelis hottingeri, Rhabdorites sp., Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti, Kathina sp., and Miscellaneidae indet.
Genus Rhabdorites , 1996
Type species: Rhapydionina malatyaensis , 1976
Rhabdorites sp.
Figs. 9.W, C'-D' ![]() ,
10.N-O, R-S
,
10.N-O, R-S ![]() ,
12.A-B, D
,
12.A-B, D ![]()
1997 Rhabdorites cf. malatyaensis, , Pl. 10, figs. 1-8.
Material. Samples Champotón-1, Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Remarks. Some specimens with porcelaneous shells and uniserial, conical shapes have been identified among the material studied and have been attributed to the genus Rhabdorites , 1996. The small number of specimens identified means that it is not possible to describe the morphotype to species level, but its basic architecture has been identified. It consists of radial partitions placed in the peripheral part of the saucer-shaped chambers, interpreted as septula, and multiple apertures
(Fig. 10.O ![]() ). The radial rows of foramina alternate with septula. No pillars are present in the adaxial part of the chamber. The megalosphere seems to be relatively large and followed by spirally arranged chambers building half a whorl
(Fig. 10.S
). The radial rows of foramina alternate with septula. No pillars are present in the adaxial part of the chamber. The megalosphere seems to be relatively large and followed by spirally arranged chambers building half a whorl
(Fig. 10.S ![]() ).
).
The presence of Rhabdorites in the Caribbean paleobioprovince during Paleocene times was indicated by (1997). The present paper supports that statement in the light of new material from the Yucatán Peninsula.
Associated larger foraminifera. This form occurs associated with Fallotella causae, Coskinolina yucatensis, Cincoriola cf. ovoidea, Neotaberina sp., Borelis floridanus, Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti, Kathina sp., Miscellaneidae indet., Borelis hottingeri and Pseudofallotella drobneae n.sp.
Suborder Rotaliina & , 1896
Family Miscellaneidae in , 1952
Miscellaneidae indet.
1997 Miscellanea sp., , Pl. 18, figs. 1-8.
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Remarks. The specimens attributed to the Family Miscellaneidae show lamellar-perforate shells, planispiral-involute chamber arrangement and external ornamentation based on pustules produced by the addition of successive outer lamellas (see
Fig. 12.Q-R ![]() ). The aperture seems to be single, small and in an interiomarginal position
(Fig. 12.Q
). The aperture seems to be single, small and in an interiomarginal position
(Fig. 12.Q ![]() ), but more material is needed in order to confirm this feature as well as the presence of an umbilical plate. Having single or multiple apertures is the taxonomic criterion to separate the two subfamilies of miscellaneids: Miscellaneinae in & , 1959, includes forms with simple aperture; and Miscellanitinae ,
2009, groups forms with multiple apertures. The abundance of miscellaneiforms in the Paleocene of the Caribbean paleobioprovince combined with the lack of detailed structural analysis call for further research on this group in the near future.
), but more material is needed in order to confirm this feature as well as the presence of an umbilical plate. Having single or multiple apertures is the taxonomic criterion to separate the two subfamilies of miscellaneids: Miscellaneinae in & , 1959, includes forms with simple aperture; and Miscellanitinae ,
2009, groups forms with multiple apertures. The abundance of miscellaneiforms in the Paleocene of the Caribbean paleobioprovince combined with the lack of detailed structural analysis call for further research on this group in the near future.
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti n.sp., Kathina sp., Neotaberina sp., Rhabdorites sp., Borelis hottingeri and Pseudofallotella drobneae.
Click on thumbnail to enlarge the image.
Figure 12. Rhabdorites sp. (A-B, D), Neotaberina sp. (C, E-H, M), Miscellaneidae indet. (I-L, N-R), Hexagonocyclina cristensis (, 1924) (S-T), and Ranikothalia soldadensis ( & , 1941) (U-V), from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm for pictures A-D, F-G, I-L, N-V; 0.3 mm for pictures E, H, M. A-D, longitudinal sections. E, G-H, M, transverse sections. F, I, L, N-O, S, oblique sections. K, U, axial sections. J, P, tangential sections. Q, R, T, subaxial sections. In pictures E, H and M, note the presence of pillars in the central part of the chamber, and foramina that alternate with septula, both characteristics are typical from the genus Neotaberina. A, 95-Me-47-34. B, 95-Me-47-35. C, 95-Me-47-36. D, 95-Me-47-37. E, 95-Me-47-38. F, 95-Me-47-39. G, 95-Me-47-40. H, 95-Me-47-41. I, 95-Me-47-42. J, 95-Me-47-43. K, 95-Me-47-44. L, 95-Me-47-45. M, 95-Me-47-46. N, 95-Me-47-47. O, 95-Me-47-48. P, 95-Me-47-49. Q, 95-Me-47-50. R, 95-Me-47-51. 19. S, 95-Me-47-52. T, 95-Me-47-53. U, 95-Me-47-54. V, 95-Me-47-55.
Abbreviations: ch: chamber; f: foramen; p: pillar; sl: septulum. See , 2006, for definition of terms.
Family Discocyclinidae , 1928
Genus Hexagonocyclina , 1944
Type species: Orbitoclypeus? cristensis , 1924
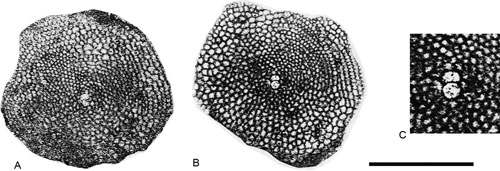
Hexagonocyclina cristensis (, 1924)
Figs. 10.P ![]() , 12.S-T
, 12.S-T ![]() ,
13
,
13 ![]()
1929 Discocyclina cristensis (). , p. 8-9; Pl. 2, figs. 1-2
1944 Hexagonocyclina cristensis (). , p. 362-363
1960 Discocyclina (Discocyclina) cristensis (). & , p. 14-15; Pl. 3, figs. 1-4
1981 Discocyclina cristensis (). , p. 48-49; Pl. 26, figs. 3-4
1984 Discocyclina cristensis (). , p. 128-129; Pl. 8, figs. 16-17
1997 Hexagonocyclina cf. cristensis (). , p. 113-116; Pl. 24, figs. 1-6
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Description. Lamellar perforate specimens with discoidal morphology. The embryonic apparatus consists of protoconch, deuteroconch and two auxiliary chambers at the junction between protoconch and deuteroconch (see
Fig. 13.C ![]() ). The equatorial diameter is around 1.6 mm and the diameter of the protoconch is 80 µm. Adult chamberlets are hexagonal.
). The equatorial diameter is around 1.6 mm and the diameter of the protoconch is 80 µm. Adult chamberlets are hexagonal.
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis, Dictyoconoides boneti, Kathina sp., Miscellaneidae indet., Neotaberina sp., Rhabdorites sp., Borelis hottingeri, and Pseudofallotella drobneae.
Click on thumbnail to enlarge the image.
Figure 13. Hexagonocyclina cristensis (, 1924) from the Paleocene of Campeche (SE Mexico). Scale bar = 1 mm. A-B, equatorial section. C, detail of the embryonic apparatus. Note the presence of the auxiliary chambers. A, 95-Me-47-56. B, 95-Me-47-57.
Family Nummulitidae , 1827
Genus Ranikothalia , 1944
Type species: Nummulites nuttalli , 1927
Ranikothalia soldadensis ( & , 1941)
Figs. 10.Q, T ![]() , 12.U-V
, 12.U-V ![]() , 14
, 14 ![]() , 15.D-F
, 15.D-F ![]() , 16.A-D
, 16.A-D ![]()
1941 Miscellanea soldadensis n.sp. & , p. 36; Pl. 4, figs. 8-9
1944 Ranikothalia soldadensis ( & ). , p. 23; Pl. 4, fig. 19; Pl. 5, figs. 24, 26?
1945 Miscellanea soldadensis ( & ). , p. 30; Pl. 5, figs. 2-5
1960 Operculina catenula ( & ). & , p. 11-14; Pl. 2, figs. 1-5
1975 Ranikothalia soldadensis ( & ). , p. 539; Pl. 1, figs. 1, 4; Pl. 2, figs 1, 6, 8; Pl. 6, figs. 1, 3; Pl. 7, figs. 1-5, Pl. 8, figs. 1-3
1996 Ranikothalia soldadensis ( & ). , p. 1185; Pl. 5.4; Pl. 9.1
1997 Ranikothalia soldadensis ( & ). , p. 145-151; Pl. 30, figs. 1-4; Pl. 31, figs. 1-3; Pl. 32, figs. 1-6; Pl. 33, fig. 1
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Description. The microspheric and megalospheric forms show flattened lenticular shell-shape with rounded periphery. The ornamentation is formed by radial filaments slightly curved at the periphery and piles concentrated in the umbonal area. The microspheric generation is characterized by having a marginal cord strongly thickened up to 1/3rd of the chambers height
(Fig. 14.A ![]() ). The maximum equatorial diameter observed in this form is around 8.25 mm and the thickness in axial section 0.7-1 mm with 3 ½ to 5 whorls and 72 to 100 chambers in total, around 35 chambers being present in the last whorl. The maximum diameter in megalospheric forms varies from 2.5-3 mm and the thickness from 0.6-0.9 mm for 3 to 3 1/2 whorls and 45 to 52 chambers in total, there being about 25 chambers in the last whorl. The embryonic apparatus of the megalospheric generation is isolepidine, and consists of a subspherical proloculus with a maximum diameter of 0.180-0.300 mm and a deuteroconch of about 0.200 mm in diameter (see
Fig. 14.C-D
). The maximum equatorial diameter observed in this form is around 8.25 mm and the thickness in axial section 0.7-1 mm with 3 ½ to 5 whorls and 72 to 100 chambers in total, around 35 chambers being present in the last whorl. The maximum diameter in megalospheric forms varies from 2.5-3 mm and the thickness from 0.6-0.9 mm for 3 to 3 1/2 whorls and 45 to 52 chambers in total, there being about 25 chambers in the last whorl. The embryonic apparatus of the megalospheric generation is isolepidine, and consists of a subspherical proloculus with a maximum diameter of 0.180-0.300 mm and a deuteroconch of about 0.200 mm in diameter (see
Fig. 14.C-D ![]() ). The following chambers are planispiral with regular spiral growth and involute arrangement, except for microspheric forms that are evolute in the last whorls. The chambers are higher than long with a subrectangular outline in equatorial section. The septa are straight or slightly inclined and curved backward to the top of the chambers. The ceiling of the chambers is arched or sometimes flattened.
). The following chambers are planispiral with regular spiral growth and involute arrangement, except for microspheric forms that are evolute in the last whorls. The chambers are higher than long with a subrectangular outline in equatorial section. The septa are straight or slightly inclined and curved backward to the top of the chambers. The ceiling of the chambers is arched or sometimes flattened.
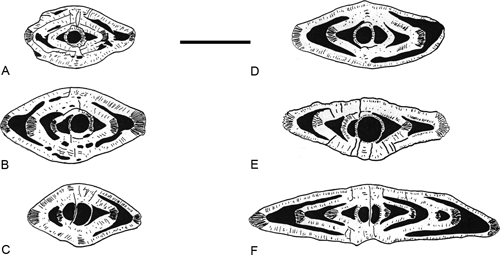
Differences and similarities. Ranikothalia soldadensis differs from the Caribbean species Ranikothalia bermudezi ( & ,
1941) by its flatter morphology, tighter spiral and smaller test size
(Fig. 15 ![]() ).
).

Click on thumbnail to enlarge the image.
Figure 14. Ranikothalia soldadensis ( & , 1941) from the Paleocene of Campeche (SE Mexico). Scale bar = 1 mm. A, axial section, microspheric form. B, equatorial section, microspheric form. C-F, equatorial sections, megalospheric forms. G-H, axial sections, megalospheric forms. A, 95-Me-47-58. B, 95-Me-47-59. C, 95-Me-47-60. D, 95-Me-47-61. E, 95-Me-47-62. F, 95-Me-47-63. G, 95-Me-47-64. H, 95-Me-47-65.
Click on thumbnail to enlarge the image.
Figure 15. Drawings of the Ranikothalia bermudezi (, 1934) (A-C), and Ranikothalia soldadensis ( & , 1941) (D-F) from the Paleocene of Campeche (SE Mexico). Scale bar = 1 mm. Note the differences between the two species in the axial outline. Specimens of R. soldadensis from the Paleocene of Campeche (SE Mexico); specimens of R. bermudezi from the Paleocene of Chiapas (SE Mexico).
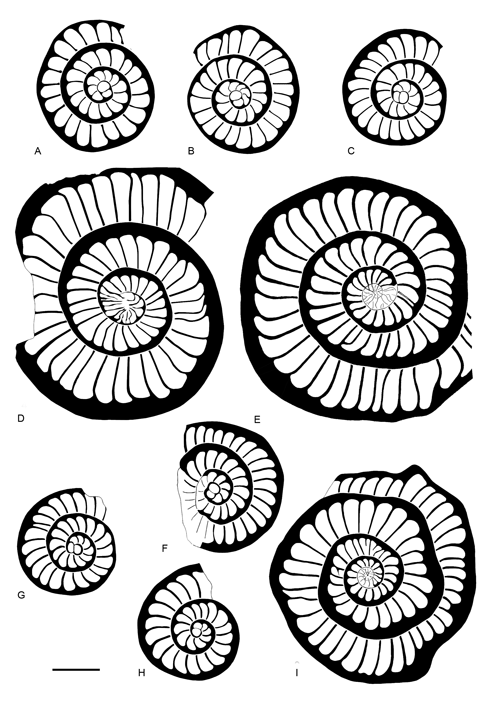
Remarks. Considering the external ornamentation, the planispiral growth from involute to evolute arrangement of the chambers, its outline and distribution, we consider that the forms attributed in this work to Ranikothalia soldadensis belong to the same phylogenetic branch as Ranikothalia sindensis (, 1927) from the Tethyan realm. There is, however, sufficient controversy regarding the Tethyan specimens attributed to R. sindensis, that it is worth mentioning here. This species was described by (1927) as Operculina sindensis in Paleocene rocks (Khairabad limestones Formation) from Pakistan. Some years later, and (1937) constrained the biostratigraphic range of R. sindensis to the upper Thanetian-lower Eocene? age, equivalent to SBZ 4-SBZ 5? according to et al. (1998). The controversy appeared when (1977) described and illustrated R. sindensis from Ruisseau de la Mède (France) and Dor M'Sid (Libya). Though (1927) and and (1937) only illustrated the external surface of the test with no equatorial sections of R. sindensis (op. cit.: see Pl. 19, fig. 10-13 and Pl. 4, fig. 21), the morphotype described by seems to be very different showing smaller microspheric specimens (5.5 mm versus 7-12 mm). Moreover, 's forms were dated as lower Thanetian or SBZ 3. Thus, R. sindensis sensu (1977) must be separated as a different species from the original species of .
The specimens of R. soldadensis studied differ from R. sindensis sensu by having larger microspheric forms (8.2 mm versus 5.5 mm), a larger diameter proloculus in megalospheric forms (0.240-0.260 mm versus 0.180-0.225 mm) and a laxer spiral in both generations (see
Fig. 16 ![]() ). Thus, the morphotype described in this paper as R. soldadensis can be considered a more evolved form than R. sindensis described by
(1977), and both would be in the same phylogenetic branch.
). Thus, the morphotype described in this paper as R. soldadensis can be considered a more evolved form than R. sindensis described by
(1977), and both would be in the same phylogenetic branch.
In contrast, R. soldadensis is architecturally closer to the R. sindensis from Pakistan. Unfortunately, as noted above, (1927) and and (1937) did not illustrate any equatorial sections, but only the external surface of the test (op. cit.: see Pl. 19, fig. 10-13 and Pl. 4, fig. 21). Therefore, verifying the possible synonymy of R. soldadensis with R. sindensis would require the re-characterization of the holotype or the topotypes, and this is far beyond the scope of this paper. Nonetheless, the external similarities between the two species suggest that the range of R. soldadensis is above SBZ 3, likely SBZ 4.
The recently described Ranikothalia polatliensis (2013) from the Tethyan paleobioprovince seems to be also closely related to R. soldadensis, but the former differs from the latter by both its larger proloculus (0.350-0.450 mm versus 0.180-0.300 mm) and loosely coiled early whorls.
Associated larger foraminifera. This form occurs associated with Hexagonocyclina cristensis, Dictyoconoides boneti, Kathina sp., Miscellaneidae indet., Neotaberina sp., Rhabdorites sp., Borelis hottingeri, and Pseudofallotella drobneae.
Click on thumbnail to enlarge the image.
Figure 16. Drawings of equatorial sections of Ranikothalia soldadensis ( & , 1941) (A-D) from the Paleocene of Campeche (SE Mexico), Ranikothalia sindensis (, 1927) (E-F) from Ruisseau de la Mède (France), and Ranikothalia sindensis (, 1927) (G-I) from Dor M'Sid (Libya). Scale bar = 1 mm.
Family Rotalidae , 1839
Subfamily Rotaliinae , 1839
Subfamily Lockhartinae , 2014
Genus Dictyoconoides , 1925
Type species Conulites cooki , 1861
Dictyoconoides boneti n.sp.
1997 Lockhartia? sp. , p. 99; Pl. 17, figs. 1-9.
Holotype. Fig. 17.C ![]() (95-ME-47-05).
(95-ME-47-05).
Paratypes. Fig. 17.O ![]() (95-ME-47-06), 17.P
(95-ME-47-06), 17.P ![]() (95-ME-47-07), 17.R
(95-ME-47-07), 17.R ![]() (95-ME-47-08).
(95-ME-47-08).
Type locality. Champotón-3, X: -90,73379612940; Y: 19,26029960940. (Fig. 1 ![]() ).
).
Type level. Thanetian (SBZ 3? and SBZ 4), Chichen Itzá Formation.
Material. Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Derivation of name. Given in honour of Federico , who worked on the stratigraphy of the Yucatán Peninsula.
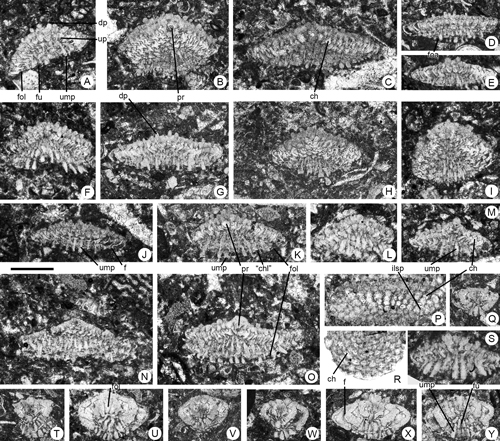
Description: Specimens show lamellar-perforate, low-conical shells. No B-Forms have been found. The A-Forms show the low-trochospiral morphology with evolute dorsal side and flattened or slightly convex ventral side. The oblique longitudinal section showed the chamber arrangement in two simultaneous spirals
(Fig. 17.B-C ![]() ). The longitudinal length reaches 0.650 mm with a diameter of 2 mm. The wall is bilamellar-perforate and thick with coarse pores on the dorsal side; the periphery is rounded, without keels. The dorsal side shows the piles located at the junction of chamber and whorl sutures, these sutures being slightly depressed
(Fig. 17.A
). The longitudinal length reaches 0.650 mm with a diameter of 2 mm. The wall is bilamellar-perforate and thick with coarse pores on the dorsal side; the periphery is rounded, without keels. The dorsal side shows the piles located at the junction of chamber and whorl sutures, these sutures being slightly depressed
(Fig. 17.A ![]() ). The umbilical or ventral side is filled with numerous vertical piles and shows funnels subdivided by numerous folia
(Fig. 17.K, O
). The umbilical or ventral side is filled with numerous vertical piles and shows funnels subdivided by numerous folia
(Fig. 17.K, O ![]() ); between two adjacent piles there are spaces similar to "chamberlets"
(Fig. 17.K
); between two adjacent piles there are spaces similar to "chamberlets"
(Fig. 17.K ![]() ). The apertures are interiomarginal and located near the periphery on the umbilical side. Proloculus are located in apical position with a maximum diameter of 0.120 mm.
). The apertures are interiomarginal and located near the periphery on the umbilical side. Proloculus are located in apical position with a maximum diameter of 0.120 mm.
Remarks. This new species differs from Dictyoconoides flemingi described by and (1937) from Paleocene of Pakistan by being smaller in size and by the presence of abundant piles on the dorsal side.
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis, Hexagonocyclina cristensis, Kathina sp., Miscellaneidae indet., Neotaberina sp., Rhabdorites sp., Borelis hottingeri, and Pseudofallotella drobneae.
Click on thumbnail to enlarge the image.
Figure 17. Dictyoconoides boneti n.sp. (A-P, R-S), and Kathina sp. (Q, T-Y) from the Paleocene of Campeche (SE Mexico). Scale bar = 0.6 mm. A-B, D, H-J, Q, T-W, oblique sections close to the axial plane. C, E, F-G, K-O, Q, S, X-Y, axial sections. P, R, transverse, slightly oblique sections. A-M, megalospheric specimens. N-P, microspheric specimens. A, 95-Me-47-65. B, 95-Me-47-66. C, 95-Me-47-05 (Holotype). D, 95-Me-47-67. E, 95-Me-47-68. F, 95-Me-47-69. G, 95-Me-47-70. H, 95-Me-47-71. I, 95-Me-47-72. J, 95-Me-47-73. K, 95-Me-47-74. L, 95-Me-47-75. M, 95-Me-47-76. N, 95-Me-47-77. O, 95-Me-47-06 (Paratype). P, 95-ME-47-07 (Paratype). Q, 95-Me-47-78. R, 95-ME-47-08 (Paratype). S, 95-Me-47-79. T, 95-Me-47-80. U, 95-Me-47-81. V, 95-Me-47-82. W, 95-Me-47-83. X, 95-Me-47-84. Y, 95-Me-47-85.
Abbreviations: ch: chamber; "ch": "chamberlet"; dp: dorsal pile; f: foramen; foa: foliar aperture; fol: folia; fu: funnel; ilsp: interseptal interlocular space; pr: proloculus; ump: umbilical pile; up: umbilical plate. See , 2006, for definition of terms.
Subfamily Kathininae , 2014
Genus Kathina , 1954
Type species: Kathina delseota , 1954
Kathina sp.
1997 Kathina? sp. , Pl. 16, figs. 4-5.
Material: Samples Champotón-2 and Champotón-3 (Fig. 1 ![]() ).
).
Description. Specimens have lamellar-perforate, low conical shells. No B-Forms have been found. The A-Forms show low-trochospiral morphology with rounded, unkeeled periphery. The dorsal side is flattened or slightly convex and the umbilical side markedly convex. The wall of the chambers is thick with coarse pores. The longitudinal length reaches 0.6 mm with a diameter of 0.95 mm. The folia are fused in umbilical zone forming piles; between these piles there are funnels connected with spiral canal (see
Fig. 17.Y ![]() ). The apertures are arched in interiomarginal position and located below the dorsal side
(Fig. 17.X
). The apertures are arched in interiomarginal position and located below the dorsal side
(Fig. 17.X ![]() ). The dorsal side is smooth and show piles only at the junction of the first chamber sutures.
). The dorsal side is smooth and show piles only at the junction of the first chamber sutures.
Remarks. This form differs from Kathina major, K. delseota and K. selveri characterized by (1954) in Paleocene rocks from Qatar in its smaller size and differs from Kathina pernavuti characterized by (1972) in Paleocene rocks from Anatolia (Turkey) in its less flattened morphology. However, the scarcity of this form in our samples does not allow greater taxonomic precision.
Associated larger foraminifera. This form occurs associated with Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti, Miscellaneidae indet., Neotaberina sp., Rhabdorites sp., Borelis hottingeri, and Pseudofallotella drobneae.
Genus Cincoriola , 1958
Type species: Punjabia ovoidea , 1956
Cincoriola cf. ovoidea (, 1958)
1997 Cincoriola? sp. , p. 98; Pl. 15, figs. 1-3
1998 Cincoriola?. , p. 77, figs. 15-17
1998 ?Kathina melona . et al. p. 174, 196; Pl. 3, fig. c; Pl. 14, figs. 1-3
Material: Sample Champotón-1 (Fig. 1 ![]() ).
).
Description: The shell is lamellar perforate with thick walls and trochospiral chamber arrangement. The ventral side is highly convex. The septa cover all the umbilical face and show numerous foramina. The foramina between two continuous chambers are aligned in parallel spirals (see
Fig. 9.R ![]() ).
).
Remarks. Due to the scarcity of specimens of this form in our material we prefer to use the term confer for the specific assignment.
Associated larger foraminifera. This form occurs associated with Fallotella causae, Coskinolina yucatensis, Rhabdorites sp., and Borelis floridanus.
Two different associations of larger foraminifera have been distinguished in the material studied. The first one is represented by Fallotella causae, Coskinolina yucatensis n.sp., Rhabdorites sp., Borelis floridanus and Cincoriola cf. ovoidea and the second by Pseudofallotella drobneae n.sp., Neotaberina sp., Rhabdorites sp., Borelis hottingeri n.sp., Ranikothalia soldadensis, Hexagonocyclina cristensis, Dictyoconoides boneti n.sp., Kathina sp. and Miscellaneidae indet.
The presence of Ranikothalia soldadensis allows the second association to be dated as Thanetian, SBZ 3? or SBZ 4 (Paleocene as pointed out by & , 1975). However, no index fossil has been found in the stratigraphic interval containing the first association. The similarities of some of the larger foraminifera found in the two assemblages, such as Rhabdorites and Neotaberina, suggest a similar age to the second association, though a slightly older age cannot be ruled out. Inferring an older age to the first association with respect to the second would be justified by the absence of the relatively complex conical form Pseudofallotella in the former, and its presence in the latter. Moreover, in the Tethyan paleobioprovince the presence of Fallotella alavensis is restricted to the SBZ 3; therefore, a similar age for the Caribbean consort F. causae is plausible.
The larger foraminifera associations defined in et al. (2013) for the Paleocene of Chiapas State (SE Mexico), one of them with Taberina cubana , 1946, and the other one with Ranikothalia bermudezi (, 1934), are younger than, at least, the association of R. soldadensis. More regional stratigraphic studies are needed, however, to constrain the ages and the paleobiogeographic distribution of all the assemblages.
The evolutionary trends shown by Paleogene agglutinated conical foraminifera on both sides of the Atlantic were thoroughly discussed in & (1980). However, the lack of representatives of conical forms in the Paleocene of the Caribbean paleobioprovince prevented the authors from providing a complete comparison of the morphotypes of equivalent time levels during this interval. Thus, one of the major arguments on which & (1980) based their conclusions about the evolution of conical forms was that common taxa in the two paleobioprovinces seemed to appear in different stratigraphic levels (specimens the genus Fallotella in the Caribbean paleobioprovince were known only from the middle Eocene deposits). This assumption allowed the authors to conclude that species found in the two areas developed in parallel lineages without any phylogenetic relationships and that the evolution of Paleogene conical foraminifera was independent on the two sides of the Atlantic. The descriptions presented in this paper of the Paleocene conical forms found in the Yucatán peninsula (Mexico) shed new light on this issue. The presence of representatives of the genera Coskinolina and Fallotella in the Paleocene of the Caribbean paleobioprovince means that the distribution patterns in the two areas are similar and confirms the similar evolution of the architectural features of the taxa. The similar evolutionary trends are also confirmed by the presence, on both sides of the Atlantic, of Eocene complex forms with subepidermal network such as Dictyoconus (in the Tethyan) and Cushmania (in the Caribbean).
Concerning the other taxa of larger foraminifera studied in this work, representatives of Ranikothalia, Cincoriola, Kathina and ortophragminids were present on both sides of the Atlantic during Paleocene times. On the other hand, the genera Borelis, Neotaberina, Rhabdorites and Dictyoconoides were restricted to the Caribbean paleobioprovince at that time (representatives are stratigraphically higher in Tethyan paleobioprovince). The differences between the representatives, at least at the specific level, of the taxa found in the Paleocene deposits on both sides of the Atlantic seems to confirm that Caribbean and Tethyan paleobioprovinces can be clearly differentiated by their fossil assemblages. However, further research is needed to improve our knowledge of the taxa and their paleobiogeographic and stratigraphic distribution (endemisms, processes of evolutionary convergence, migratory fluxes, ).
Previous work concerning the palaeoenvironmental distribution of larger Paleogene foraminifera has been reported by (1988), & (1988), et al. (1996), et al. (2002), & (2004), et al. (2004), et al. (2006) and et al. (2013). In our work, the data obtained from different isolated outcrops in Campeche, and those from Chiapas ( et al., 2013), make it possible to comment on the ecological requirements of the morphotypes studied from the Paleocene times in the Caribbean area. The paleoenvironmental interpretation of the previous study of & (1999) dealing with the microfacies of the Xbcal Mb. has been considered in this review.
In recent times, the larger foraminifera are among the major sediment producers in the shallow-marine domain of tropical-subtropical climatic areas ( et al., 1981) characterized by stable nutrient-deficient environments (mesotrophic to oligotrophic) (, 1988). These features involve an anti-estuarine circulation, the absence of rejuvenation currents (up-welling) and the absence of nutrient input from emerged continents (runoff) (, 1990). Thus, considering the architectural similarities of the taxa studied in this paper and those of the current foraminifera (e.g., nummulitids and alveolinids), we infer similar ecological requirements for the fossil assemblages.
According to their morpho-functional characteristics, the genera Taberina, Neomurciella and Raadshoovenia (discoidal-flabelliform and planar morphology of test, and peripheral distribution of apertures) have similarities with modern soritid Amphisorus. Hence, we assume that they have similar environmental requirements. In the Red Sea, at depths of up to 35 meters, Amphisorus hemprichii lives under normal marine conditions and epiphytically on Halophila leaves ( & , 1984). The predominance of these genera and the presence of microspheric forms of the genera with long life-times (annual, not seasonal) imply the presence of perennial vegetation (vascular plants) and the development of seagrass cover in an inner-ramp under oligotrophic conditions in the Paleocene outcrops from Chiapas.
The current forms of the genus Borelis live at depths of 10 to 60 meters mostly between 25 and 35 meters, in normal-salinity, low-energy nutrient-poor marine environments with very low terrigenous input, and on either soft or hard substrates. Borelis lives on Halophila leaves or on hard substrate ( & , 1984).
We consider that Rhabdorites and Neotaberina with planispiral to uniserial arrangement of the chambers and the kind of apertures (cribate) have the same palaeoenvironmental requirements as current forms of peneroplids. These forms live as phytal dwellers or on the soft substrate (, 1991) and at a maximum depth of around 70 meters, though they are more likely to be found within the top 40 meters ( & , 1984).
Regarding the genus Ranikothalia, as a nummulitid, it is able to colonize the environment within deeper photic zones under oligotrophic conditions. For instance, in the Red Sea the genus Operculina can live around a depth of 60 m and Heterostegina around 100 m as free-forms under oligotrophic conditions ( & , 1984).
The ecological requirements of other larger foraminifera without current equivalent forms can also be inferred from their morphostructure. For example, the genera Fallotella, Coskinolina (orbitolinids) and Dictyoconoides (rotalid) have no equivalents since their extinction at the Eocene-Oligocene boundary ( & , 2007). The similar aperture distribution (at the base of the conical test) of these genera indicates that these forms could not live on the ecological soft substrate with a basal trophic level. As a hypothesis, we consider that these forms were able to live as free-forms on the vegetal substrate or on bioclastic debris.
Finally, the orthophragminids such as the Hexagonocyclina genus are considered to be the larger foraminifers that lived the deepest, up to the lower limit of the photic zone ( & , 1992).
Considering the data obtained in this work and by et al.
(2013), the four assemblages described from the Paleocene rocks of Campeche and Chiapas can be interpreted as deposits of an inner to middle ramp setting
(Fig. 18 ![]() ). However, some particularities can be observed: whereas Taberina and its associates of Chiapas are considered to live in seagrass, the association with Ranikothalia and Hexagonocyclina indicate the deeper part of the inner ramp or even the middle ramp.
). However, some particularities can be observed: whereas Taberina and its associates of Chiapas are considered to live in seagrass, the association with Ranikothalia and Hexagonocyclina indicate the deeper part of the inner ramp or even the middle ramp.

Click on thumbnail to enlarge the image.
Figure 18. Paleoenvironmental distribution of the Paleocene larger foraminifera from Chiapas and Yucatán Peninsula (SE Mexico).
The detailed study of the larger foraminifera found in the Chichen Itzá Formation in Campeche (SE Mexico) has allowed us to describe three new species and one new genus of larger conical foraminifera, Fallotella causae, Coskinolina yucatanensis and Pseudofallotella drobneae. In addition, one new species of alveolinid, Borelis hottingeri, and one new species of rotaliid, Dictyoconoides boneti, have been identified among the material studied.
Two larger foraminiferal associations interpreted as deposited in the deeper part of the inner ramp, or even the middle ramp, have been defined. One is characterized by the presence of Coskinolina yucatensis, Fallotella causae, Rhabdorites sp., Borelis floridanus and Cincoriola cf. ovoidea, and the other by Pseudofallotella drobneae, Neotaberina sp., Rhabdorites sp., Borelis hottingeri, Dictyoconoides boneti, Kathina sp., Miscellaneidae, Ranikothalia soldadensis, and Hexagonocyclina cristensis. The genera Cincoriola, Neotaberina, Rhabdorites and Dictyoconoides have been cited in the Caribbean paleobioprovince for the first time in this paper.
The two foraminiferal associations in Campeche described in this paper, interpreted as deposits of an inner to middle ramp setting, together with those defined in Chiapas by et al. (2013) characterize the Paleocene of southern Mexico. Combining data from these two studies, the estimated age for the two assemblages of Campeche is Thanetian (SBZ 3? and SBZ 4).
The financial support for the project by the Spanish Ministry of Science and Innovation (CGL 2011-27869) is gratefully acknowledged. Many thanks are also due to the Documentation Centre of the Museu de Ciències Naturals de Barcelona for its support in supplying references. The very useful suggestions and corrections of the reviewers of the present paper, K. , V. and E. are also gratefully acknowledged. We also thank John , who ultimately revised the English text of the manuscript.
G., F. & J. (1998).- Depositional history of a Paleogene carbonate ramp (Western Cephalonia, Ionian Islands, Greece).- Geologica Romana, vol. 34, 205 p.
A. (1952).- I macroforaminiferi della serie del Carcàr (Eocene medio e superiore) in Somalia e la loro distribuzione stratigrafica.- Palaeontographia Italica, Pisa, vol. XLVII (Nuova Serie, vol. XVII, 1950-1951), p. 99-131.
J. & A. (2004).- Ecology of extant nummulitids and other larger benthic foraminifera: applications in palaeoenvironmental analysis.- Earth Science Reviews, Amsterdam, vol. 67, p. 219-265.
J., V.P. & A. (2006).- The Middle Eocene Seeb Formation of Oman: An investigation of acyclicity, stratigraphic completeness, and accumulation rates in shallow marine carbonate settings.- Journal of Sedimentary Research, Tulsa, vol. 76, p. 1137-1161.
J.A. (1997).- Macroforaminíferos del Terciario inferior del sureste de México.- Ph. Doctoral Thesis, Universitat Autònoma de Barcelona, 207 p. (45 Pls.).
J.A. & E. (1999).- Las microfacies con macroforaminíferos del Terciario inferior de la península de Yucatán (SE de México).- Revista Asociación de Geólogos y Geofísicos Españoles del Petróleo, Madrid, Libro Homenaje a José , p. 19-32.
J. (1981).- Claves para la determinación de macroforaminíferos de México y del Caribe, del Crétacico Superior al Mioceno Medio.- Instituto Mexicano del Petróleo, Subdirección de Tecnología de Exploración, México, 219 p.
J. & F. (1960).- Microfauna del Eoceno Inferior de la Península de Yucatán.- Paleontología Mexicana, Instituto de Geología, México, vol. 7, p. 1-18.
J. & F. (1963).- Las Formaciones Cenozoicas de la parte mexicana de la península de Yucatán.- Ingeniería Hidráulica en México, Mexico, vol. 17, p. 63-71.
J. & M. (1968).- Les Orbitolinidae de l'Éocène de la région des Caraïbes.- Archives des Sciences, Genève, vol. 21, fasc. 1, p. 5-20 ( 3 Pls.).
C.M.B. (1944).- The larger foraminifera from San Juan de los Morros, State of Guarico, Venezuela.- Bulletin of American Paleontology, Ithaca, vol. 28(114), p. 355-404.
C.M.R. (1975).- Geology and paleontology of Soldado Rock, Trinidad (West Indies) Part 2: The larger foraminifera.- Eclogae Geologicae Helvetiae, Basel, vol. 68, nº 3, p. 533-589 (30 Pls.)
C.M.B. (1996).- The larger foraminifera of Trinidad (West Indies).- Eclogae Geologicae Helvetiae, Basel, vol. 89, nº 3, p. 1137-1309.
W.S. (1941).- Stratigraphic and paleontologic studies of wells in Florida. Nº 1.- Geological Bulletin, Tallahassee, vol. 16, 73 p.
W.S. (1944).- Stratigraphic and paleontologic studies of wells in Florida. Nº 3.- Geological Bulletin, Tallahassee, vol. 26, 168 p.
W.S. (1957).- Larger foraminifera from Eniwetok Atoll drill holes.- United States Geological Survey, Professional Paper, Washington, vol. 260/5, p. 739-784 (19 Pls.).
V., K. & A. (2004).- Paleoenviromental model for Eocene foraminiferal limestones of the Adriatic carbonate platform (Istrian Peninsula).- Facies, Erlangen, vol. 50, p. 61-75.
L. M. (1927).- The Ranikot beds at Thal (North-West frontier provinces of India).- Quarterly Journal Geological Society of London, vol. 83, p. 260-290, Pl. 5
L.M. & E.S. (1937).- The Eocene beds of the Punjab Salt Range.- Palaeontologia Indica, Calcutta, vol. 24, nº 1, 79 p. (7 Pls.).
R.C. (1960).- Revision of the Family Orbitolinidae.- Micropaleontology, vol. 6, New York, p. 249-270 (6 Pls.).
K. & V. (2009).- Palaeobiogeography of the Late Cretaceous to Paleogene larger Miliolids from tropical to subtropical sea belts (Neotethys to Caribbean).- Bulletin de la Société géologique de France, Paris, vol. 180, nº 4, p. 317-331.
W.W. (1991).- Distribution and substrate preference of Peneroplis planatus and P. arietinus from the Halophila meadow near Wadi Taba, Eilat, Israel.- Journal of Foraminiferal Research, Lawrence, vol. 21, p. 218-221.
C. & J. (1992).- Morphostructure and paleobiology of Discocyclina , 1870.- Journal of Foraminiferal Research, Lawrence, vol. 22, nº 2, p. 147-165.
S.H. & R. Jr (1974).- Cenozoic reef biofacies: Tertiary larger foraminifera and scleractinian corals from Chiapas, Mexico.- Northern Illinois University Press, DeKalb, 387 p.
L. (1988).- Diversification in algal symbiont-bearing foraminifera: A response to oligotrophy? In: Benthos'86.- Revue de Paléobiologie, Genève, vol. spec. 2, p. 789-797.
L. (1977).- Les foraminifères operculiniformes.- Mémoires du Muséum d'Histoire naturelle, Paris, vol. 40, 159 p.
L. (1982).- Larger foraminifera, giant cells with a historical background.- Naturwissenschaften, Berlin, vol. 69, p. 361-371.
L. (1990).- Significance of diversity in shallow benthic foraminifera.- Atti Quarto Simposio di Ecologia e Paleoecologia delle Comunità Bentoniche, Museo Regionale di Scienze Naturali, Torino, p. 35-51.
L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets de Géologie [Notebooks on Geology], Brest, Memoir 2006/02 (CG2006_M02), 126 p. DOI: 10.4267/2042/5832
L. (2007).- Revision of the foraminiferal genus Globoreticulina , 1978, and of its associated fauna of larger foraminifera from the late Middle Eocene of Iran.- Carnets de Géologie [Notebooks on Geology], Brest, Article 2007/06 (CG2007_A06), 51 p. DOI: 10.4267/2042/9213
L. (2009).- The Paleocene and earliest Eocene foraminiferal Family Miscellaneidae: neither nummulitids nor rotaliids.- Carnets de Géologie [Notebooks on Geology], Brest, Article 2009/06 (CG2009_A06), 41 p. DOI: 10.4267/2042/28794
L. (2014).- Paleogene larger rotaliid foraminifera from the western and central Neotethys.- Springer Verlag, Heidelberg, 200 p.
L. & K. (1980).- Early Tertiary conical imperforate foraminifera.- Razprave, Dissertationes (Slovenska Akademia Znanosti in Umetnostni), Lljubljana, (classis IV), vol. 22, nº 3, p. 187-276 (22 Pls.).
International Commission on Zoological Nomenclature (1999).- International Code of Zoological Nomenclature (4th edition).- The International Trust for Zoological Nomenclature, The Natural History Museum, London. URL: http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp
H.G. & C.M.B. (1975).- Geology and paleontology of Soldado Rock, Trinidad (West Indies). Part I: Geology and biostratigraphy.- Eclogae Geologicae Helvetiae, Basel, vol. 68, nº 2, p. 365-430 (2 Pls.).
H.L. (1957).- Micropaleontology of the Oldsmar limestone (Eocene) of Florida.- Micropaleontology, New York, USA, vol. 3, nº 2, p. 137-154.
A.R. & H. (1987).- A.R. & H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold Co, New York, 2 vols. 970 + 212 p. (847 Pls.).
J.-F. (1984).- Le Sénonien supérieur-Tertiaire du Chiapas (S.E. Mexique) et ses macroforaminifères.- Ph. Doctoral Thesis, Université de Nice; Travaux du Centre de Recherches micropaléontologiques "Jean ", vol. 4, 322 p.
J.-F. (1995).- Pseudorhapydionina moulladei, nouveau macroforaminifère du Paléocène du Chiapas, SE Mexique.- Revue de Micropaléontologie, Paris, vol. 38, nº 2, p. 169-176.
A. (1960).- Aperçu sur la microfaune de l'Éocène du Médoc.- Revue de Micropaléontologie, Paris, vol. 3, p. 31-36 (2 Pls.).
Z. & L. (1984).- The Gulf of Aqaba ecological micropaleontology.- Ecological Studies, Heidelberg, vol. 50, 354 p.
J., P., J., A., S., J. & C. (2013).- The Paleogene pre-rift to syn-rift succession in the Dhofar margin (northern Gulf of Aden): Stratigraphy and depositional environments.- Tectonophysics, Utrecht, vol. 607, p. 1-16.
E. & R.M. (1993).- Jamaican Paleogene larger foraminifera. In: R.M. & E. (eds.), Biostratigraphy of Jamaica.- Geological Society of America, Boulder, Memoir 182, p. 283-345.
J., E. & J. (2002).- A model for the palaeoenvironmental distribution of larger foraminifera based on late Middle Eocene deposits on the margin of the South Pyrenean basin (NE Spain).- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 179, p. 43-56.
R. (1974).- Der Typus der Foraminiferen-gattung Coskinolina 1875.- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 11, p. 702-706.
J., L., E., K., C., A.K., G., R., J., J. M., H., E., A., Y., J. & E. (1998).- Larger Foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene.- Bulletin de la Société géologique de France, Paris, vol. 169, nº 2, p. 281-299.
J., C., J. & P.P. (2007).- Cainozoic larger foraminifera from Dominican Republic.- Boletín Geológico y Minero, Madrid, vol. 118, nº 2, p. 359-384.
E. (1972).- Systematic study of new species of the genera Fabularia and Kathina from Paleocene.- Türkiye Jeoloji Kurumu Bülteni, Ankara, vol. 15, p. 277-249 (8 Pls.).
E. (1998).- Foraminiferal description and biostratigraphy of the Paleocene-Lower Eocene shallow-water limestones and discussion on the Cretaceous-Tertiary boundary in Turkey.- Mineral Research and Exploration (MTA), Monography Series, Ankara, nº 2, 117 p. (68 Pls.).
E. (2013).-
Description of two new families, three new species and re-description of four known genera and one
subfamily from the larger benthic foraminifera of Paleocene in Turkey.- Bulletin
of the Mineral Research and Exploration, Ankara, vol. 15, p. 27-53 (7 Pls.).
G. (1875).- Neue Beobachtungen in den Schichten der liburnischen Stufe.- Verhandlungen der kaiserlich-königlichen geologischen Reichsanstalt, Wien, Jahrgang 1875, p. 334-338.
A.H. (1954).- Lower Tertiary foraminifera of the Qatar Peninsula.- British Museum (Natural History), London, 96 p. (15 Pls.).
A. & J. (1988).- Sedimentología y distribución de foraminíferos en medios litorales y de plataforma mixta (Eoceno Medio y Superior, Cuenca del Ebro Oriental).- Boletín Geológico y Minero, Madrid, vol. 94, p. 871-885.
A., J. & I. (1996).- Paleoecological Interpretation of transitional environments in Eocene carbonates (NE Spain).- Palaios, Lawrence, vol. 11, p. 141-160.
T.W. (1929).- Descriptions of new species of foraminifera of the genus Discocyclina from the Eocene of Mexico.- Proceedings of the United States National Museum, Washington, vol. 76, art. 3 (2800), p. 1-18.
T.W. (1945).- American old and middle Tertiary larger foraminifera and corals. Part 1. American Paleocene and Eocene larger foraminifera.- Geological Society of America Memoirs, Boulder, vol. 9, p. 1-167.
T.W. & W.S. (1941).- Preliminary report on the Cretaceous and Tertiary larger foraminifera of Trinidad British West Indies.- Special Papers of the Geological Society of America, Boulder, vol. 30, 135 p.
V., A., J. & E. (2013).- Architecture and age of the foraminiferal genus Taberina , 1945.- Journal of Foraminiferal Research, Lawrence, vol. 43, nº 2, p. 170-181.
E. & L. (2007).- Agglutinated conical foraminifera from the Lower-Middle Eocene of the Trentinara Formation (southern Italy).- Facies, Erlangen, vol. 53, p. 509-533.
C., J.S. & G.Z. (1981).- Stable isotopes in Recent larger foraminifera.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 33, p. 253-270.