◄ Carnets Geol. 14 (7) ►
![]()
Contents
[Introduction] [Geological and stratigraphic settings] [Materials and methods]
[Results and discussion] [Concluding remarks] and ... [References]
Department of Botany, LaTrobe University, Bundoora, Victoria 3086 (Australia)
Department of Ecology and Evolutionary Biology, The University of Kansas, 1200 Sunnyside Avenue, Lawrence, Kansas 66045 (USA)
UNESP - Universidade Estadual Paulista, Center for Geosciences Applied to Petroleum (UNESPetro) and Departamento de Geologia Aplicada, Caixa Postal 178, Av. 24 A, nş 1515, Bela Vista, CEP13506-900 - Rio Claro - SP (Brazil)
Published online in final form (pdf) on May 8, 2014
[Editor: Michel ; copy editor: Bruno ]
![]()
Fossil specimens of Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), representing the first confirmation of the genus in the fossil record, were discovered in thin sections of Albian limestones from the Riachuelo Formation, Sergipe Basin, and in thin sections of Albian - Cenomanian limestones from the Ponta do Mel Formation, Potiguar Basin in north-eastern Brazil. A detailed morphological-anatomical account of the species is provided, and its placement in Heydrichia is discussed in relation to current classification proposals. Comparisons with the four other known species of the genus, all non-fossil, show that H. poignantii is the only known species of Heydrichia in which thalli are encrusting to sparsely warty to horizontally layered with overlapping lamellate branches that commonly appear variously curved or arched, and in which thalli have sporangial complexes that become buried in the thallus. The evolutionary history of Heydrichia remains uncertain, but available data suggest that the genus may have diverged from the sporolithacean genus Sporolithon, known as early as Hauterivian times (c. 129.4-132.9 ± 1 Ma) from Spain (and newly reported here from Switzerland), or it may have arisen from a graticulacean alga such as Graticula, dating from mid-Silurian times (c. 427-435 Ma). Current data also suggest that Heydrichia is more likely to have arrived in Brazil from Central Atlantic waters than from higher latitude South Atlantic waters. This implies that currently living species in southern Africa probably arose later from ancestors further equatorward in the South Atlantic, although confirming studies are needed. All non-fossil species of Heydrichia are known only from the southern hemisphere.
Heydrichia; Sporolithaceae; Sporolithon; fossil coralline red algae; Brazil; Switzerland; Hauterivian; Albian; Cenomanian.
W.J., B. & D. (2014).- Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), a 100 million year old fossil coralline red alga from north-eastern Brazil, and a new Hauterivian record of Sporolithon from Switzerland.- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 7, p. 139-158.
Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), une algue rouge coralline fossile du Nord-Est du Brésil, vieille de quelques 100 millions d'années, et nouvelle découverte en Suisse d'un Sporolithon d'âge hauterivien.- Des spécimens fossiles de Heydrichia (?) poignantii, sp. nov. (Sporolithaceae, Sporolithales, Rhodophyta), représentant le enregistrement fossilifčre confirmé du genre, ont été découverts en lames minces dans des calcaires albiens de la Formation de Riachuelo, Bassin de Sergipe, et dans des calcaires d'âge Albien - Cénomanien de la Formation de Ponta do Mel, Bassin de Potiguar, au NE du Brésil. Les caractéristiques morphologiques et anatomiques de cette espčce sont détaillées et son attribution au genre Heydrichia est débattue en tenant compte des propositions de la classification actuelle. Des comparaisons avec les quatre autres espčces recencées du genre, toutes inconnues ŕ l'état fossile, montre que H. poignantii est la seule espčce connue du genre Heydrichia dans laquelle les thalles sont encroűtants ŕ modérément verruqueux, ŕ horizontalement stratifiés avec recouvrements de leurs excroissances lamellaires qui, fréquemment, paraissent diversement arquées ou cintrées et dans laquelle les compartiments sporangiques qui s'enfouissent dans ce thalle. L'histoire évolutive de Heydrichia demeure incertaine, mais les données disponibles suggčrent que le genre pourrait s'ętre différencié ŕ partir du genre Sporolithon de la Famille des Sporolithacées, connu déjŕ dans l'Hauterivien (c. 129.4-132.9 ± 1 Ma) d'Espagne (et nouvellement découvert en Suisse, comme nous le rapportons dans cette publication), mais il pourrait tout aussi bien ętre issu d'une algue de la Famille des Graticulacées, telle que Graticula, trouvée dans des séries d'âge Silurien moyen (aux environs de -427 ŕ -435 Ma). Les éléments en notre possession suggčrent également que Heydrichia a plus probablement été apportée sur les rivages du NE brésilien par les eaux de l'Océan Atlantique Central plutôt que par celles de l'Océan Atlantique Sud méridional. Ceci implique que les espčces actuelles observées dans la partie méridionale du continent africain sont probablement arrivées plus tardivement, issues d'ancętres plus septentrionaux dans l'Océan Atlantique Sud : des recherches complémentaires seront nécessaires pour confirmer cette hypothčse. Toutes les représentants modernes du genre Heydrichia n'ont été observés que dans l'hémisphčre sud.
Heydrichia ; Sporolithaceae ; Sporolithon ; algues rouges corallines fossiles ; Brésil ; Suisse ; Hauterivien ; Albien ; Cénomanien.
The coralline red algal Family Sporolithaceae (Sporolithales, Corallinophycidae, Florideophyceae, Rhodophyta) Note 1 includes two genera with living species: Sporolithon (1897) and Heydrichia et al. (1994). The Order Sporolithales differs from the related Order Corallinales in producing tetrasporangia in calcified compartments rather than in conceptacles, and in having cruciate rather than zonately arranged tetraspores ( et al., 2010). Both Orders include fossil and non-fossil genera and species.
Unlike tetraspores and tetrasporangia, which seldom are encountered in fossil material, sporangial compartments in taxa of Sporolithales are more commonly preserved. Sori are composed of aggregations of sporangial compartments, each originally containing a tetrasporangium. Compartments may also be solitary. In Heydrichia, sporangial compartments are surrounded by an involucre of smaller-diameter filaments whose cells differ in appearance from ordinary vegetative cells. A sporangial compartment together with the surrounding involucre is known as a sporangial complex ( et al., 1995, p. 98). By contrast, sporangial complexes do not occur in Sporolithon because sporangial compartments are not surrounded by involucres. However, involucres are known in carposporangial conceptacles of both genera (see et al., 1995, p. 98-99 and et al., 2002, p. 210 for further comments on involucres).
In the palaeontological record, Sporolithon is known as far back as the Hauterivian (c. 129.4-132.9 ± 1 Ma) of Spain ( et al., 1995, as Archaeolithothamnium; et al., 2007, as Sporolithon) and is newly reported here from Switzerland. These Hauterivian records represent several of the earliest known unequivocal occurrences of the genus and of fossil coralline red algae in general ( et al., 2010, p. 527).
By contrast, specimens explicitly identified as Heydrichia have not been reported previously in the fossil record. et al.
(2008, Pl. 2, fig. C), however, depicted a specimen identified as Sporolithon sp. in strata of Late Albian to Early Cenomanian age (c. 100.5 ± 2 Ma) found in cores taken in the uppermost part of the Ponta do Mel Formation in the Potiguar Basin north of Macau in north-eastern Brazil that we recognize here as the first effectively published record of fossil material referable to Heydrichia
Note 2. We also have discovered additional material referable to Heydrichia in
thin sections of Albian age material (c. 100.5-113 ± 1 Ma) at two localities in the Riachuelo Formation, Sergipe Basin, north-eastern Brazil. Collections from these three localities appear to be conspecific and are regarded here to represent a species new to science, described here as Heydrichia (?) poignantii, sp. nov.
Note 3. An account of the species based on thin sections
(Fig. 1 ![]() ) from the three known localities is provided, and the palaeontological and biological implications are discussed, including the matter of whether apparent sporangial complexes in which involucres have not been preserved could instead be trace fossils produced by micro-borers.
) from the three known localities is provided, and the palaeontological and biological implications are discussed, including the matter of whether apparent sporangial complexes in which involucres have not been preserved could instead be trace fossils produced by micro-borers.
The development of Albian-Cenomanian shallow-water carbonate deposits in Cretaceous basins along the north-eastern coast of Brazil ( et al., 2000, p. 352, Fig. 1), including the Sergipe and Potiguar basins, coincided with the break-up of continental Gondwana, the separation of the South American and African plates, and the opening of the Central and South Atlantic oceans. The depositional, diagenetic and stratigraphic aspects of microfacies from the Riachuelo Formation (Sergipe Basin) have been analyzed by et al. (2013), while the carbonate composition and the reconstruction of the depositional and diagenetic history of the Ponta do Mel Formation (Potiguar Basin) have been described by (1990), with further data provided by et al. (2008). A first paleophycological report on the Riachuelo Formation (Sergipe Basin) was provided by et al. (1991).
et al. (2008) also published a cursory inventory of the fossilized remains of calcareous red and green algae from Petrobras exploration well RNS-1, discussed earlier algal records from the Potiguar and Sergipe basins, and noted that fossilized calcareous red and green algae are poorly documented in southern hemisphere basins.
Rock samples containing coralline algae were cut into rectangular pieces c. 40 mm x 45 mm x 10 mm thick using a diamond blade circular saw running under a continuous water stream. The coralline-bearing algal face of each piece was polished with a lapidary cast plane and then placed on a hotplate for 1-2 h for drying and degassing before bonding to a clean glass slide with an epoxy adhesive, which was allowed to polymerize (c. 1 h for warmed samples; 12-24 h for cold samples). Each rock sample was then reduced in thickness with manual or mechanical lapping until the section was 20-30 µm thick. Any loose particles remaining after lapping were removed with a soft brush or by gentle sonication before covering with an ethanol-cleaned coverslip using Canada balsam and allowed to dry.
Nomenclatural terminology follows the International Code of Nomenclature for Algae, Fungi and Plants ( et al., 2012), hereafter abbreviated ICN. For purposes of typification, the term 'specimen' is used as defined in ICN Art. 8.2. Thus a specimen may consist of a single organism, parts of one or several organisms, or of multiple small organisms belonging to a single species made at one time. A 'holotype' (ICN Art. 9.1) consists of a single specimen as defined in ICN Art. 8.2 (also see Art. 8.3). The type (epitypes excepted) of the name of a fossil species is always a specimen (ICN Art. 8.5).
Morphological and anatomical terminology follows
(1988) and et al.
(2002), and growth-form terminology follows et al.
(1993) and et al.
(2012, p. 173). In dorsiventrally organized branches or thalli, vertical sections are cut more or less perpendicular to the thallus surface (e.g.,
Figs. 2-3 ![]() ,
4-6
,
4-6 ![]() ,
7
,
7 ![]() ,
11-12
,
11-12 ![]() ). In vertical sections, cells of filaments may be cut longitudinally or transversely or obliquely, depending on their position in the thallus and their orientation to the thallus surface. Paradermal sections of dorsiventrally organized branches or thalli are cut parallel to the thallus surface. In paradermal sections taken close to the dorsal thallus surface (e.g.,
Figs. 8-9
). In vertical sections, cells of filaments may be cut longitudinally or transversely or obliquely, depending on their position in the thallus and their orientation to the thallus surface. Paradermal sections of dorsiventrally organized branches or thalli are cut parallel to the thallus surface. In paradermal sections taken close to the dorsal thallus surface (e.g.,
Figs. 8-9 ![]() ), all cells are cut transversely. Cell sizes are measured in cells cut longitudinally. Cell measurements are taken from mid-cell wall to mid-cell wall; 'sporangial complex' cavity measurements do not include the involucre.
), all cells are cut transversely. Cell sizes are measured in cells cut longitudinally. Cell measurements are taken from mid-cell wall to mid-cell wall; 'sporangial complex' cavity measurements do not include the involucre.
Herbarium abbreviations follow (2014, continuously updated). Thin sections used for the account of Heydrichia (?) poignantii have been deposited in PC (Herbier Cryptogamique, Département Systématique et Évolution, Muséum National d'Histoire Naturelle, Paris, France). The thin section of the Swiss Hauterivian Sporolithon is housed at the Muséum d'Histoire naturelle de la Ville de Genčve, Switzerland.
Names and dates of chronostratigraphic units are those of the International Commission on Stratigraphy ( et al., 2013). UTM data were calculated using a geographic-UTM converter available at http://www.geoplaner.com
Taxonomic outcomes are based on the morphological-anatomical species concept (, 2012; et al., 2013, p. 82). The almost total lack (except for et al., 2008) of molecular sequence data for fossil coralline red algae precludes any cross comparisons of outcomes based on such data sets with those based exclusively on morphological-anatomical data.
Map 2 ![]() was generated using the ODSN (Ocean Drilling Stratigraphic Network) Plate Tectonic Reconstruction Service established by GEOMAR, Research Center for Marine Geosciences/Kiel and the Geological Institute of the University Bremen (URL:
http://www.odsn.de/odsn/services/paleomap/paleomap.html). Parameters used to generate base map were as follows:
cartographic projection: Lambert Azimuthal; move plates relative to: Magnetic Ref.
Frame; guideline interval 15°; annotation interval 15°; reconstruction
age: 100 Ma; map boundaries: 90° North, -120° West, 90° East, 90° South;
frame type: thin lines; plate fragments as at 100Ma: grey; present-day
shorelines: black. Oceanic shading, place names, locality dots, and position of Rio Grande Rise - Walvis Ridge were added subsequently, and the colour of plate fragments was changed to
light grey.
was generated using the ODSN (Ocean Drilling Stratigraphic Network) Plate Tectonic Reconstruction Service established by GEOMAR, Research Center for Marine Geosciences/Kiel and the Geological Institute of the University Bremen (URL:
http://www.odsn.de/odsn/services/paleomap/paleomap.html). Parameters used to generate base map were as follows:
cartographic projection: Lambert Azimuthal; move plates relative to: Magnetic Ref.
Frame; guideline interval 15°; annotation interval 15°; reconstruction
age: 100 Ma; map boundaries: 90° North, -120° West, 90° East, 90° South;
frame type: thin lines; plate fragments as at 100Ma: grey; present-day
shorelines: black. Oceanic shading, place names, locality dots, and position of Rio Grande Rise - Walvis Ridge were added subsequently, and the colour of plate fragments was changed to
light grey.
Uniform resource locators (URL, i.e., web addresses) cited for online resources were operational at the time of final manuscript submission.
Scale bar lengths listed in figure legends do not include the white highlight borders.
Heydrichia (?) poignantii , & , sp. nov.
Previous references in publications and theses:
1990 Archaeolithothamnion.- , 1990, p. 29 (as Archaeolithothamnium); p. 59, "foto 9", "algal vermelhas corallináceas" (coralline red alga); Formaçăo Ponta do Mel, (?) Upper Albian - Cenomanian, Potiguar basin, NE Brazil.
2008 Sporolithon sp.- et al., 2008, p. 312, Pl. 2, fig. C; Ponta do Mel Formation, (?) Upper Albian - Cenomanian, Potiguar basin, NE Brazil.
Diagnosis: Heydrichia (?) poignantii, with the characteristics of Heydrichia (sensu et al., 1995, p. 98-99); differing from other species in having a thallus that is encrusting to sparsely warty to horizontally layered with overlapping lamellate branches that commonly appear variously curved or arched, and in producing sporangial complexes that become buried in the thallus.
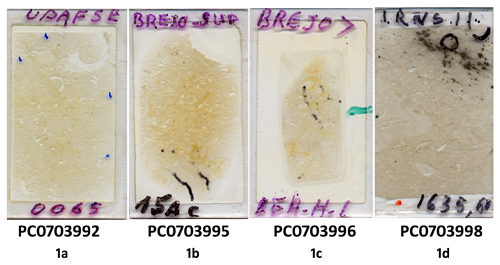
Holotype preparation: PC0703992 (thin section UPAFSE 0065), prepared from a rock sample collected by Dimas on 5 February 2009).
Fig. 1a ![]() .
.
Holotype specimen: The holotype specimen, designated here, and depicted in
Figs. 2-3 ![]() ,
4, 6
,
4, 6 ![]() ,
7
,
7 ![]() , consists of five pieces of thallus with empty sporangial complexes occurring on
thin section PC0703992 (UPAFSE 0065)
(Fig. 1a
, consists of five pieces of thallus with empty sporangial complexes occurring on
thin section PC0703992 (UPAFSE 0065)
(Fig. 1a ![]() ). With one exception, these pieces all extend to the same place along the edge of the
thin section and thus may belong to a single thallus. Thallus material beyond the edge was lost during
thin section preparation.
). With one exception, these pieces all extend to the same place along the edge of the
thin section and thus may belong to a single thallus. Thallus material beyond the edge was lost during
thin section preparation.
Click on thumbnail to enlarge the image.
Fig. 1: Heydrichia poignantii [thin sections examined; all deposited in PC]. Fig. 1a: PC0703992 (UPAFSE 0065). Thin section containing holotype material from Fazenda Paty, Riachuelo Formation, Sergipe Basin, Brazil. Fig. 1b: PC0703995 (Brejo Sup., 15A - C). Thin section containing material from Pedreira (Quarry) Brejo, Riachuelo Formation, Sergipe Basin, Brazil. Fig. 1c: PC0703996 (Brejo>, 15A - H-L). Thin section containing additional material from Pedreira (Quarry) Brejo, Riachuelo Formation, Sergipe Basin, Brazil. Fig. 1d: PC0703998 (RNS 11). Thin section containing material from Petrobras Well 1, north of Macau, Ponta do Mel Formation, Potiguar Basin, Brazil.
Click on thumbnail to enlarge the image.
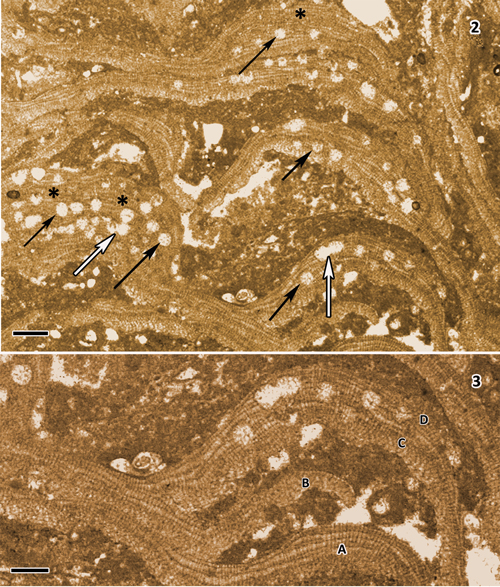
Figs. 2-3: Heydrichia poignantii [holotype material, PC0703992 (thin section UPAFSE 0065)]. Fig. 2: Vertical section of holotype material with numerous, solitary, scattered empty sporangial complexes (black arrows) and several conjoined sporangial complexes (white arrows). Note several small, thickened warty protuberances (black *) at lower left and upper right. Scale bar = 200 µm. Fig. 3: Enlarged view of part of lower left part of holotype material in Fig. 2 showing four layers of horizontal branching (A, B, C, D), all derived from a single parent branch (at left). Scale bar = 100 µm.

Click on thumbnail to enlarge the image.
Figs. 4-6: Heydrichia poignantii [Figs. 4, 6, holotype material, PC0703992 (thin section UPAFSE 0065); Fig. 5, PC0703995 (Brejo thin section Brejo Sup., 15A-C)]. Fig. 4: Vertical section of young poorly preserved thallus branch apex. Note monomerous construction with a thin ventral core region (v) in which filaments run parallel to thallus surface, and a peripheral region (p) in which filaments curve upwards to thallus surface. Arrows denote poorly preserved epithallial cells. Scale bar = 40 µm. Fig. 5: Vertical section of older part of poorly preserved thallus branch showing monomerous construction with a thin ventral core (v) cut obliquely and a more extensive peripheral region (p) and several empty buried sporangial complexes (c). Sporangial involucres are not preserved. Scale bar = 50 µm. Fig. 6: Vertical section of older portion of a thallus branch with poorly preserved, flattened epithallial cells (arrows). Scale bar = 25 µm.

Click on thumbnail to enlarge the image.
Figs. 7-10: Heydrichia poignantii [Fig. 7, holotype material, PC0703992 (thin section UPAFSE 0065); Figs. 8-10, PC0703996 (Brejo thin section Brejo>, 15A - H-L)]. Fig. 7: Vertical section of thallus showing conceptacle compartments (c) encased by remains of involucres (white arrows), and possible cell fusions (black arrows). Scale bar = 50 µm. Fig. 8: Paradermal section of thallus cut through three sporangial complexes. Note darkened remnants of involucres bordering each empty sporangial complex and polygonal shape of ordinary vegetative cells. Scale bar = 75 µm. Fig. 9: Enlarged view of part of paradermal section depicted in Fig. 8. Note more or less polygonal shape of vegetative cells, central dark spots denoting primary pit connections to subtending cells, and possible cell fusions (arrows). Scale bar = 25 µm. Fig. 10: Low magnification largely paradermal section of thallus fragment showing sporangial complexes (white arrows), two trace fossil annelid borings (black arrows) and a large trace fossil bivalvia boring (BB). Green arrowheads denote edge of thallus fragment. Scale bar = 200 µm.
Thin section PC0703992 (UPAFSE 0065) also includes six other fragments (two with cavities) that resemble holotype material vegetatively, but these fragments have been excluded from consideration as type material because they lack clear sporangial complexes and thus cannot definitely be identified as H. poignantii.
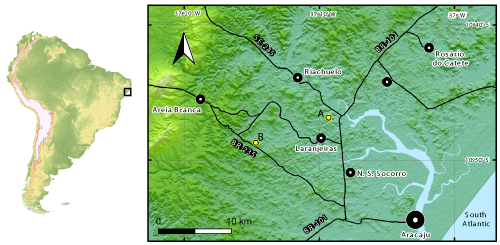
Type locality (locus typicus): Fazenda Paty, north west of Aracaju, State of Sergipe, Brazil
(Map 1 ![]() , locality B). UTM 24S 0691860E 8804268N (latitude: 10°48'27.256"S; longitude: 37°14'42.47" W, see Google Maps: -10.807572 -37.245131).
, locality B). UTM 24S 0691860E 8804268N (latitude: 10°48'27.256"S; longitude: 37°14'42.47" W, see Google Maps: -10.807572 -37.245131).
Geological age: undifferentiated Albian. Geological stratum (stratum typicum): Riachuelo Formation, Sergipe Basin, north-eastern Brazil.
Images:
Figs. 1a ![]() ,
2-3
,
2-3 ![]() ,
4, 6
,
4, 6 ![]() ,
7
,
7 ![]() .
.
Click on thumbnail to enlarge the image.
Map 1: Heydrichia poignanti collection localities in the Sergipe Basin, Brazil. A = Brejo, north west of Aracaju, State of Sergipe; B = Fazenda Paty, north west of Aracaju, State of Sergipe.
Etymology: The specific epithet honours Prof. Alain-François (1931-2011, see http://deptsec.ku.edu/~ifaaku/Poignant.html) for his numerous contributions to our understanding of fossil coralline red algae. Prof. was one of the first persons to study Cretaceous red algae from Brazil, and also encouraged the second author (B. ) to study fossil and modern "calcareous" algae during his student days.
Additional thin sections examined:
PC0703995 (thin section Brejo Sup., 15A-C). Pedreira (Quarry) Brejo, north west of Aracaju, State of Sergipe, Brazil (see
Map 1 ![]() , locality A). UTM 24S 0701684E 8807626N (latitude: 10°46'36.141"S; longitude: 37°9'19.767" W, see Google Maps: -10.776706, -37.155492). Rock sample collected by Dimas on 4 February 2009.
, locality A). UTM 24S 0701684E 8807626N (latitude: 10°46'36.141"S; longitude: 37°9'19.767" W, see Google Maps: -10.776706, -37.155492). Rock sample collected by Dimas on 4 February 2009.
Geological age: Late Albian (Upper Albian strata). Geological stratum: Riachuelo Formation, Sergipe Basin, north-eastern Brazil.
Images:
Figs. 1b ![]() ,
5
,
5 ![]() .
.
PC0703996 (thin section Brejo>, 15A - H-L). Pedreira (Quarry) Brejo, north west of Aracaju, State of Sergipe, Brazil (see
Map 1 ![]() , locality A). UTM 24S 0701684E 8807626N (latitude: 10°46'36.141"S; longitude: 37°9'19.767" W, see Google Maps: -10.776706, -37.155492). Rock sample collected by Dimas on 4 February 2009.
, locality A). UTM 24S 0701684E 8807626N (latitude: 10°46'36.141"S; longitude: 37°9'19.767" W, see Google Maps: -10.776706, -37.155492). Rock sample collected by Dimas on 4 February 2009.
Geological age: Late Albian (Upper Albian strata). Geological stratum: Riachuelo Formation, Sergipe Basin, north-eastern Brazil.
Images:
Figs. 1c ![]() ,
8-10
,
8-10 ![]() .
.
PC0703998 (thin section RNS-11). Petrobras Well 1, 30 km north of Macau, State of Rio Grande do Norte, SAD 69 (Brazil): UTM 24S 776981.92E 9461584.57N as indicated by ANP-BDEP (latitude 4°51'59.198"S, longitude 36°30'9.655"W; see Google Maps: -4.866444, -36.502682). Rock sample from a depth of 1635.60 m, collected by Petrobras Oil Company team members in 1976.
Geological age: Late Albian-Early Cenomanian. Geological stratum: Ponta do Mel Formation, Potiguar Basin, north of Macau, Brazil.
Images:
Fig. 1d ![]() ; ,
1990, p. 59, 'foto 9'; et al.,
2008, Pl. 2, fig. C.
; ,
1990, p. 59, 'foto 9'; et al.,
2008, Pl. 2, fig. C.
Other organisms present on the thin sections examined:
Fazenda Paty, Riacheulo Formation, Sergipe Basin (PC0703992):
bivalvia; gastropoda; echinids; foraminifera (including Mississippiana sp.); algae (including Elianella elegans, Parachaetetes sp., Pycnoporidium sinuosum)
Pedreira (Quarry) Brejo, Riacheulo Formation, Sergipe Basin (PC0703995, PC0703996):
bivalvia (including pycnodontids, ?Lopha sp., boring bivalvia); gastropoda; echinids; serpulids; foraminifera (including Trocholina silvai); bryozoans; algae (including Marinella lugeoni, Girvanella sp.); trace fossils/ichnotaxa (bivalvia: Gastrochaenolites ichnosp.; annelida: Trypanites ichnosp.)
Petrobras Well 1, 30 km north of Macau, Ponta do Mel Formation, Potiguar Basin (PC0703998):
bivalvia (including boring bivalvia); gastropoda; echinids; foraminifera (including Trocholina silvai); algae (including Elianella elegans, Parachaetetes sp., Pycnoporidium sinuosum, Sporolithon ? amphiroaeformis); trace fossils/ichnotaxa (bivalvia: Gastrochaenolites ichnosp.)
Morphology and anatomy: The holotype of Heydrichia (?) poignantii is encrusting to warty to layered in growth-form. Layered portions have flattened, overlapping lamellate branches that commonly appear variously curved or arched
(Figs. 2-3 ![]() ). Up to four derivative branches arising from a single parent branch
(Fig. 3
). Up to four derivative branches arising from a single parent branch
(Fig. 3 ![]() ) have been observed. Several low, local mound-like or warty protuberant outgrowths up to 440 µm tall also were evident
(Fig. 2
) have been observed. Several low, local mound-like or warty protuberant outgrowths up to 440 µm tall also were evident
(Fig. 2 ![]() ) and may have made local portions of a mainly layered thallus look sparsely warty or lumpy in surface view. Unbranched portions of thalli were up to 1200 µm thick, and individual branches were mostly 80-165 µm thick. Substrate relations are not evident in the holotype collection, and thus we are uncertain whether individuals grew attached to rocks or to other substrata. There is no evidence to suggest that the holotype was living as a rhodolith.
) and may have made local portions of a mainly layered thallus look sparsely warty or lumpy in surface view. Unbranched portions of thalli were up to 1200 µm thick, and individual branches were mostly 80-165 µm thick. Substrate relations are not evident in the holotype collection, and thus we are uncertain whether individuals grew attached to rocks or to other substrata. There is no evidence to suggest that the holotype was living as a rhodolith.
The holotype is pseudoparenchymatous and dorsiventrally organized. Thallus construction is monomerous, consisting of a single system of branched filaments that form a ventral core running more or less parallel to the thallus surface and a dorsal peripheral region where portions of filaments or their derivatives curve upwards and terminate at the thallus surface in epithallial cells
(Fig. 4 ![]() ). Within filaments, cells cut longitudinally look more or less rectangular (e.g.,
Figs. 6
). Within filaments, cells cut longitudinally look more or less rectangular (e.g.,
Figs. 6 ![]() ,
7
,
7 ![]() ) while cells cut transversely look more or less angular
(Figs. 8-9
) while cells cut transversely look more or less angular
(Figs. 8-9 ![]() ). Core and peripheral filament cells are mostly 19-50 µm long and 9-14 (-17) µm in diameter. Evident epithallial cells
(Figs. 4, 6
). Core and peripheral filament cells are mostly 19-50 µm long and 9-14 (-17) µm in diameter. Evident epithallial cells
(Figs. 4, 6 ![]() ) are poorly preserved but appear flattened and are mostly 6-10 µm high and 14-25 µm in diameter. Cells below the epithallial cells
(Figs. 4, 6
) are poorly preserved but appear flattened and are mostly 6-10 µm high and 14-25 µm in diameter. Cells below the epithallial cells
(Figs. 4, 6 ![]() ), generally regarded as subapical meristematic cells, are mostly about the same size or slightly shorter than cells beneath them. Some cells of adjacent filaments appear to have possible cell fusions
(Figs. 7, 9
), generally regarded as subapical meristematic cells, are mostly about the same size or slightly shorter than cells beneath them. Some cells of adjacent filaments appear to have possible cell fusions
(Figs. 7, 9 ![]() ), but these may be artefacts of oblique sectioning through cells of adjacent filaments. Secondary pit connections were not evident, and haustoria and trichocytes were not seen.
), but these may be artefacts of oblique sectioning through cells of adjacent filaments. Secondary pit connections were not evident, and haustoria and trichocytes were not seen.
Sporangial complexes with preserved remnants of involucres (Figs.
7-8 ![]() ; et al.,
2008, Pl. 2, fig. C) occurred in some thalli from all three localities, but more commonly, involucres were not obviously preserved
(Figs. 2-3
; et al.,
2008, Pl. 2, fig. C) occurred in some thalli from all three localities, but more commonly, involucres were not obviously preserved
(Figs. 2-3 ![]() ,
5
,
5 ![]() ). Sporangial complexes had become buried in the thallus after presumed release of spores, and those near the surface no longer had evident pore canals
Note 4. The apparent absence of pore canals might be a consequence of non-preservation or an artefact of sectioning. Vertical sections that are cut slightly obliquely to sporangial complexes may not, by chance, pass through the pore canal. Empty complexes were mostly 35-90 µm in diameter and 65-100 µm high.
). Sporangial complexes had become buried in the thallus after presumed release of spores, and those near the surface no longer had evident pore canals
Note 4. The apparent absence of pore canals might be a consequence of non-preservation or an artefact of sectioning. Vertical sections that are cut slightly obliquely to sporangial complexes may not, by chance, pass through the pore canal. Empty complexes were mostly 35-90 µm in diameter and 65-100 µm high.
Most complexes were solitary and scattered, but some were paired or in small clusters, and some appeared to be partially conjoined
(Fig. 2 ![]() ). Paired or
clustered sporangial complexes also occur in several non-fossil species of Heydrichia (H. cerasina: & van der ,
2012, p. 20,
Fig. 26; H. groeneri: & ,
1995, p. 53,
Fig. 7, p. 54, Fig. 20; H. woelkerlingii: et al.,
1995, p. 121,
Fig. 19). et al. (1995, p. 98-99) also noted that when complexes become crowded and abut, the adjoining filamentous regions (including involucres) can become crushed and are no longer evident. This might also happen in fossil material, either when thalli were still alive or as a result of non-preservation during the fossilization process.
). Paired or
clustered sporangial complexes also occur in several non-fossil species of Heydrichia (H. cerasina: & van der ,
2012, p. 20,
Fig. 26; H. groeneri: & ,
1995, p. 53,
Fig. 7, p. 54, Fig. 20; H. woelkerlingii: et al.,
1995, p. 121,
Fig. 19). et al. (1995, p. 98-99) also noted that when complexes become crowded and abut, the adjoining filamentous regions (including involucres) can become crushed and are no longer evident. This might also happen in fossil material, either when thalli were still alive or as a result of non-preservation during the fossilization process.
Closely situated complexes also can retain their integrity. In Figure
8 ![]() , for example, the two empty sporangial complexes at lower right are closely paired but have remained separated by vegetative filaments.
, for example, the two empty sporangial complexes at lower right are closely paired but have remained separated by vegetative filaments.
In vertical sections of thalli (Figs. 2-3 ![]() ,
5
,
5 ![]() ,
7
,
7 ![]() ), empty sporangial complexes look more or less ovoid to nearly circular, while in paradermal sections
(Figs. 8, 10
), empty sporangial complexes look more or less ovoid to nearly circular, while in paradermal sections
(Figs. 8, 10 ![]() ), empty sporangial complexes look more or less circular.
), empty sporangial complexes look more or less circular.
Gametangial thalli were not found.
Empty sporangial complexes vs trace fossils (micro-borings/tunnels): Because the Brazilian material is imperfectly preserved, involucres were not always evident in the chambers, and unequivocal pore canals were not observed. In addition, sporangia presumably had been discharged prior to complexes becoming buried in the thallus. This raises the question as to whether the empty structures lacking preserved involucres that we interpret here as sporangial complexes could, instead, be trace fossils (micro-borings/tunnels) produced by boring organisms.
According to & (2009), trace fossil assemblages in hard substrates dominated by coralline red algae have scarcely been investigated, and in their study of Middle Miocene rhodoliths from Italy, two categories of trace-fossil borings were distinguished. Trace borings with chamber diameters of 300 µm or more were categorized as macro-borings, while trace borings with chamber diameters of 10-40 µm were termed micro-borings. In our specimens, chamber diameters were 35-90 µm, similar in size to micro-borings. More generally, micro-borings include trace fossils with diameters of less than 100 µm ( & , 2007).
Micro-borings can be produced by algae, fungi, bacteria, sponges or polychaete worms ( & , 2007; & , 2009) and also have been termed tunnels or micro-tunnels. The borings (tunnels) may be wedge-shaped or tubular, can be branched or unbranched, can vary in diameter, and can run perpendicular to or more or less parallel to the surface of the host, and can form networks termed micro-galleries. Examples in Middle Miocene rhodoliths are illustrated in & (2009) and in et al. (2010), who distinguished several kinds based on differences in the above features. When cut transversely, individual tunnels look more or less rounded in section; when cut longitudinally, tunnels look cylindrical or tubular in section. At branch or convergence points in branched tunnels and in networks of micro-galleries, tunnels form a "Y" pattern in longitudinal view and look conjoined in transverse view.
Several lines of evidence have led us to conclude that even though unequivocal pore canals were not observed and involucres were not always evident, the hole-like structures in the Brazilian material are far more likely to be empty sporangial complexes than trace fossil micro-borings. Firstly, involucres were detected bordering some complexes
(Figs. 7-8 ![]() ; et al.,
2008, Pl. 2, fig. C) in specimens from all three localities, strongly supporting the conclusion that these are reproductive structures and not micro-borings. We are unaware of any reports of micro-borer tunnels that have
involucre-like linings. We also note that et al.
(2009, p. 419, Pl. 4,
figs. 4-5) found structures indicative of possible involucres associated with the reproductive structures in the lectotype of Sporolithon taiwanensis () et al., and they suggested that further studies are needed to determine the affinities of this species to Heydrichia. et al.
(2009, p. 419), however, make no mention of micro-borings.
; et al.,
2008, Pl. 2, fig. C) in specimens from all three localities, strongly supporting the conclusion that these are reproductive structures and not micro-borings. We are unaware of any reports of micro-borer tunnels that have
involucre-like linings. We also note that et al.
(2009, p. 419, Pl. 4,
figs. 4-5) found structures indicative of possible involucres associated with the reproductive structures in the lectotype of Sporolithon taiwanensis () et al., and they suggested that further studies are needed to determine the affinities of this species to Heydrichia. et al.
(2009, p. 419), however, make no mention of micro-borings.
Secondly, no branched tube-like tunnels (i.e., longitudinally cut micro-tunnels) or networks of micro-galleries are evident in our material. If such structures were present and oriented more or less perpendicularly to the coralline thallus surface, then vertical sections through the coralline thallus should show branched, tubular tunnels cut longitudinally, but none were found
(Figs. 2-3 ![]() ,
5
,
5 ![]() ,
7
,
7 ![]() ). Similarly if branched tunnels or micro-galleries were present and oriented more or less parallel to the thallus surface, then paradermal sections cut parallel to the coralline thallus surface should show branched, tubular tunnels or networks of micro-galleries cut longitudinally, but branched micro-borings were not evident in the paradermal thallus section available to us
(Figs. 8, 10
). Similarly if branched tunnels or micro-galleries were present and oriented more or less parallel to the thallus surface, then paradermal sections cut parallel to the coralline thallus surface should show branched, tubular tunnels or networks of micro-galleries cut longitudinally, but branched micro-borings were not evident in the paradermal thallus section available to us
(Figs. 8, 10 ![]() ). The absence of evidence for branched, longitudinally cut tubular micro-tunnels or networks of micro-galleries strongly suggests that the rounded structures in our material are far more likely to be reproductive structures (sporangial complexes) produced by the alga than micro-tunnels produced by micro-borers.
). The absence of evidence for branched, longitudinally cut tubular micro-tunnels or networks of micro-galleries strongly suggests that the rounded structures in our material are far more likely to be reproductive structures (sporangial complexes) produced by the alga than micro-tunnels produced by micro-borers.
Thirdly, the conjoined empty sporangial complexes in our material could be interpreted as Y-shaped junctures of branched micro-tunnels, but the absence of evidence that branched tunnels actually occur precludes this possibility. As already noted, conjoined sporangial complexes also are known in non-fossil species of Heydrichia, so their occurrence in the fossil H. poignantii is not without precedent.
Finally, the only two micro-borings found in our material (Fig. 10 ![]() , black arrows) were unbranched, varied in diameter from 75 to 140 µm when seen in longitudinal view, lacked any evidence of
involucre-like linings, and showed no other features suggestive of sporangial complexes. These micro-borings may belong to the ichnogenus Trypanites; at least one ichnospecies placed in Trypanites by &
(2007) produces borings of the dimensional range found in our material. The only other kind of boring found
(Fig. 10
, black arrows) were unbranched, varied in diameter from 75 to 140 µm when seen in longitudinal view, lacked any evidence of
involucre-like linings, and showed no other features suggestive of sporangial complexes. These micro-borings may belong to the ichnogenus Trypanites; at least one ichnospecies placed in Trypanites by &
(2007) produces borings of the dimensional range found in our material. The only other kind of boring found
(Fig. 10 ![]() , BB) was a macro-boring produced by a bivalve belonging to the ichnogenus Gastrochaenolites.
, BB) was a macro-boring produced by a bivalve belonging to the ichnogenus Gastrochaenolites.
Based on the above, the possibility that the structures we interpret as empty sporangial complexes are trace fossils produced by micro-borers seems highly unlikely. It is much more likely that when further material with better preservation of reproductive structures is found, the occurrence of sporangial complexes will be unequivocally confirmed.
Generic placement: The appearance and vegetative structure of Heydrichia (?) poignantii is concordant with that of various other coralline red algae, all of which are referred to the subclass Corallinophycidae ( & , 2007) within the Rhodophyta. Heydrichia is unique amongst known genera of Corallinophycidae in producing sporangial complexes, i.e., sporangia borne in compartments surrounded by involucres ( et al., 2000, p. 192-193; et al., 2002, p. 210). The occurrence of sporangial complexes, even though empty, provides a strong basis for placement of our Brazilian material in that genus.
In addition to the occurrence of involucres, we can confirm that thalli are non-geniculate and lack haustoria, and we have found possible cell fusions but not secondary pit-connections, all of which are characteristic of Heydrichia ( et al., 2002; & van der , 2012). We cannot, however, be absolutely certain that the Brazilian material belongs to Heydrichia because other features considered characteristic of the genus (see & van der , 2012, p. 15) have not been preserved in our fossil material. Thus we do not know whether the outer walls of epithallial cells are flared at the distal corners or whether tetrasporangia are borne on multiple-celled stalks, produce apical plugs and bear cruciately arranged spores. Consequently, while we definitely accept the name and the species, and all features evident in the holotype support inclusion in the genus Heydrichia, we have added a question mark after the generic name to indicate slight uncertainty on generic placement. Adding a question mark does not affect valid publication of the name, as explained in Footnote 3 above.
Comparisons with other species of Heydrichia: Four other species of Heydrichia. have been described: H. cerasina & van der (2012), H. groeneri & (1995), H. homalopasta & (2001), and the type species, H. woelkerlingii et al. (1994). All four are known only from non-fossil material. Species comparisons were most recently made by et al. (2002, p. 224, Table 2) and & van der (2012, p. 19, Table 1), but neither set of authors explicitly indicated which characters they considered diagnostic of species. Moreover, in their key to southern African species, & van der (2012, p. 17) also stated that the characters used were not necessarily diagnostic of the species to which they pertain but rather were used for specimen identification only.
We follow (1988, p. 226) and (2010, p. 63) (also see et al., 2005, p. 132; et al., 2009, p. 225) who define as diagnostic those characters that unambiguously separate one taxon from another (of equal taxonomic rank) or permit an organism to be assigned to a particular taxon. Based on current knowledge and the above definition, we have identified five characters (Table 1) that in combination separate each of the five species from the other four. Growth-form and the occurrence of overlapping horizontal layers of lamellate branches, while listed separately for easier recognition, are linked: a thallus or portions of a thallus with a layered growth-form by definition has horizontally layers of overlapping lamellate branches, whereas thalli or portions of thalli lacking horizontal layers of overlapping lamellate branches have other growth-forms.
| Species | Thallus thickness | Growth form | Occurrence of horizontal layers of overlapping lamellate branches | Sporangial compartment aggregation in thallus | Occurrence of buried sporangial complexes in thallus |
| H. poignantii | up to 1200 µm | encrusting to layered to warty | present | mostly solitary with some paired or in small clusters | present |
| H. cerasina | up to 1400 µm | warty | absent | solitary or in clusters of 2-5 | absent |
| H. groeneri | up to 800 µm | encrusting | absent | aggregated into large clusters | absent |
| H. homalopasta | up to 1000 µm | encrusting | absent | solitary | absent |
| H. woelkerlingii | up to 15 mm | encrusting to locally lumpy | absent | solitary or aggregated into large clusters | absent |
Table 1: Characters and character states currently considered diagnostic of species of Heydrichia based on data in et al. (1995), et al. (2002), & van der (2012) and the present study. Characters unique to H. poignantii are highlighted. Growth-form terminology follows et al. (1993).
As noted by et al. (1993), growth-forms involve an intergrading network. A given species may have a single known growth-form or more than one growth-form, even within the same thallus. Encrusting to locally lumpy thalli have been recorded for H. woelkerlingii, and several small warty protuberances occurred in H. poignantii, while only a single growth-form is known in each of the other non-fossil species of Heydrichia (Table 1). All characters and character states in Table 1 are readily observable in fossil and non-fossil specimens.
H. poignantii is the only known species of Heydrichia in which thalli are encrusting to warty to horizontally layered with overlapping lamellate branches, and have sporangial complexes that become buried in the thallus (Table 1). By contrast, thallus thickness in H. poignantii is similar to that in H. cerasina, H. groeneri, and H. homalopasta, and additionally, sporangial complexes also can be solitary or in clusters of 2-5 or more in H. cerasina and H. woelkerlingii.
The absence of sporangia in our material of H. poignantii leaves open the slight possibility that what we interpret as tetrasporangial compartments/complexes could in fact be small uniporate carposporangial conceptacles. Carposporangial conceptacles in all four non-fossil species also have involucres. & (1995, p. 52) reported that carposporangial conceptacle chambers in H. groeneri were 49-112 µm in diameter and 44-99 µm high, thus markedly overlapping the dimensions of the chambers (35-90 µm in diameter and 65-100 µm high) in H. poignantii. The diagnostic differences between H. poignantii and H. groeneri listed in Table 1, however, are not affected, however, because they pertain to the vegetative thallus, the comparative aggregation of compartments into large groups, and the burying of sporangial complexes rather than the dimensions of conceptacle/sporangial complex chambers. Carposporangial conceptacles in the remaining three species are larger in diameter: 220-280 µm in H. cerasina ( & van der , 2012, p. 15), 110-150 µm in H. homalopasta ( et al., 2002, p. 216) and 210-250 µm in H. woelkerlingii ( et al., 1994, p. 178).
The diagnostic value of characters relating to male conceptacles and to female/carposporangial conceptacles could not be evaluated due to the absence of gametangial specimens in our material of H. poignantii. Comparisons of these features amongst non-fossil species are provided by et al. (2002, p. 224, Table 2) and by & van der (2012, p. 19, Table 1). & van der (2012, p. 15, 17), however, concluded from their data that the occurrence of branched vs unbranched spermatangial filaments and the distribution of these within male conceptacles could not be considered diagnostic of Heydrichia as a genus. The absence of data on male conceptacles in H. groeneri as well as H. poignantii precludes a full assessment of the diagnostic value of the above spermatangial characters at species level.
Distribution of Heydrichia in time and space: The discovery of Heydrichia poignantii in Albian age deposits in north-eastern Brazil confirms that Heydrichia not only is represented in the fossil record but has been present in marine environments for at least 100 million years.
The evolutionary history of Heydrichia remains uncertain. Based on an analysis involving data from SSU (18S rDNA) gene sequences of 39 non-fossil species of corallines, molecular clocks, and the fossil record, et al. (2010, p. 526, Fig. 2) estimated that Heydrichia diverged from Sporolithon approximately 98.5 Ma (Cenomanian times). Our data suggest that any such evolutionary divergence occurred somewhat earlier (Albian times).
However, it also is possible that Heydrichia has had a much earlier and different origin. &
(1998, 2000) concluded that mid-Silurian (c. 427-435 Ma) specimens identified as Graticula gotlandica () & from Wales (UK) and Sweden were coralline red algae that possessed calcified sporangial compartments and otherwise resembled taxa of the
Sporolithaceae. The presumed calcified compartments ( & ,
1998, p. 187,
Fig. 2) occur in irregular groups and look like those of Heydrichia poignantii
(Figs. 2 ![]() ,
7
,
7 ![]() ). and
(1998, p. 189-190) referred Graticula to a distinct family, the Graticulaceae
Note 5, in part because of the nearly 300 million year gap (mid-Silurian/Early Cretaceous) between documented records of Graticula and the earliest confirmed records of Sporolithaceae
Note 6. They also suggested that morphological-anatomical differences (presence or absence of trichocytes; arrangement of compartments) between the families were unlikely to be sufficient to distinguish them if they were documented to be contemporaneous. Unfortunately, the absence of gene sequence data for Graticula, which is unknown outside of the mid Silurian, has precluded molecular assessments of any phylogenetic relationships between graticulaceans and
sporolithaceans.
). and
(1998, p. 189-190) referred Graticula to a distinct family, the Graticulaceae
Note 5, in part because of the nearly 300 million year gap (mid-Silurian/Early Cretaceous) between documented records of Graticula and the earliest confirmed records of Sporolithaceae
Note 6. They also suggested that morphological-anatomical differences (presence or absence of trichocytes; arrangement of compartments) between the families were unlikely to be sufficient to distinguish them if they were documented to be contemporaneous. Unfortunately, the absence of gene sequence data for Graticula, which is unknown outside of the mid Silurian, has precluded molecular assessments of any phylogenetic relationships between graticulaceans and
sporolithaceans.
and (1998, p. 193) also concluded that recognition of Graticula as a coralline implied that the Graticulaceae constituted the ancestral group of coralline red algae. This conclusion infers that Heydrichia may have a graticulacean ancestry rather than diverging directly from Sporolithon, as suggested in the molecular clock model of et al. (2010), which did not include Graticula due to the absence of suitable molecular data.
H. poignanti was growing during the period when the Atlantic Ocean was developing and South America had recently become separated from Africa
(Map 2 ![]() ) (also see ,
2002; et al., 2008, p. 245,
Fig. 2). In the context of continental drift, therefore, it would not be surprising if fossil material of Heydrichia occurred in western Africa, particularly in the coastal region from Ghana through Angola, to which north-eastern Brazil was juxtaposed/joined prior to and just after separation.
) (also see ,
2002; et al., 2008, p. 245,
Fig. 2). In the context of continental drift, therefore, it would not be surprising if fossil material of Heydrichia occurred in western Africa, particularly in the coastal region from Ghana through Angola, to which north-eastern Brazil was juxtaposed/joined prior to and just after separation.
Click on thumbnail to enlarge the image.
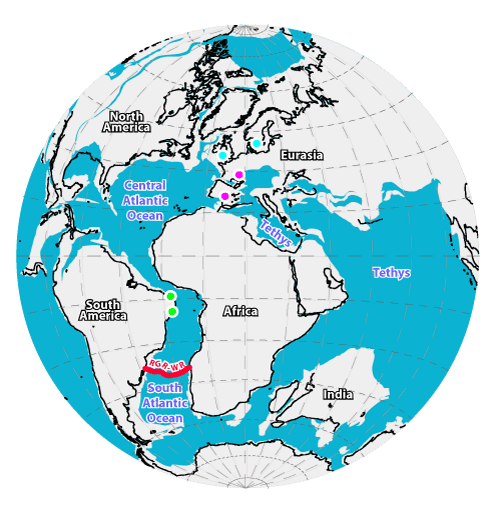
Map 2: Lambert Azimuthal map projection with reconstruction of continents associated with developing Atlantic Ocean 100Ma, when Heydrichia poignanti was living. Late (Upper) Albian land masses shaded in
light grey. Oceanic regions shaded in turquoise blue. Black lines denote present-day shorelines. Approximate position of the Rio Grande
Rise-Walvis Ridge barrier (RGR-WR) indicated in Red. Green dots denote collection sites of fossil Heydrichia (the more southerly dot includes both localities shown on
Map 1 ![]() ); pink dots denote Hauterivian records of Sporolithon in Spain and Switzerland; blue dots indicate mid-Silurian records of graticulaceans in Wales and Sweden. Gridline interval is 15°. Base map generated using the ODSN Plate Tectonic Service; details given in
Materials and Methods.
); pink dots denote Hauterivian records of Sporolithon in Spain and Switzerland; blue dots indicate mid-Silurian records of graticulaceans in Wales and Sweden. Gridline interval is 15°. Base map generated using the ODSN Plate Tectonic Service; details given in
Materials and Methods.
Of the four known non-fossil species of Heydrichia, three (H. cerasina, H. groeneri, H. woelkerlingii) are based on type material from Namibia or South Africa ( et al., 1994; & , 1995; & van der , 2012), and the first two are unknown outside that region. This also supports the possibility of finding fossil material of the genus in western (and southern) Africa. Similarly, the occurrence of fossil material of Heydrichia in eastern South America suggests the possibility that non-fossil species occur along that coast, particularly in more temperate waters. To date, however, there are no records of non-fossil material from eastern South America.
The fourth non-fossil species, H. homalopasta is based on a type from temperate eastern Australia ( & , 2001; et al., 2002), and it and H. woelkerlingii also have been recorded from temperate New Zealand ( et al., 2005; et al., 2009; , 2012). H. homalopasta also is documented from the Chatham Islands (44°00'S, 176°30'W) ( et al., 2005; et al., 2009) to the east of mainland New Zealand, raising the possibility that it may also occur further towards the west coast of South America. Annual seawater temperatures in the Chatham Islands range from 10° to 18°C.
At present there are no confirmed fossil or non-fossil records of Heydrichia from the northern hemisphere Note 7. et al. (2009, p. 419), however, suggest that compartments in the type of the fossil species Sporolithon taiwanensis () et al. might be surrounded by involucres, a diagnostic character of Heydrichia but not Sporolithon ( et al., 1995, p. 98-99), but et al. (2009, p. 419) noted that any possible affinity of S. taiwanensis to Heydrichia cannot be determined with certainty until further material from the type locality is collected and examined. The type is Pliocene - Pleistocene in age (1.8-5.3 Ma) and was collected from Taitung County, Taiwan.
Palaeogeographic and palaeoecologic reconstructions: Palaeogeographic and palaeoecologic reconstructions must be approached with great caution due to the sparseness of adequate sampling and the consequent scarcity of data ( et al., 2008, p. 318). For data on palaeoclimatic conditions, see (2002).
Based on present knowledge, habitats that contained Heydrichia poignantii also harboured other calcareous green algae (Brasiliporella, Clypeina, Linoporella (?), Neomeris) and red algae (Elianella, Lithothamnion, Marinella, Parachaetetes, Pycnoporidium, Sporolithon) ( et al., 2008, 2012a, 2012b). These algae are generally considered indicative of tropical environments. et al. (2008) emphasized that their inventory was preliminary, but they indicated that their results and well as those from earlier studies of other authors suggest that during the mid-Cretaceous, calcareous algal biodiversity may have been low on both sides of the South Atlantic Ocean.
After considering possible options concerning the geographic origin of these algae in north-eastern Brazil, et al.
(2008, p. 318-319) concluded that they probably arrived from the Central Atlantic Ocean and reached the early South Atlantic Ocean after the South American and African continents began to separate and the two oceans merged
(Map 2 ![]() ). Other groups of organisms are also thought to have followed a similar route (e.g., see ,
2000; & de ,
2000; , 2000; ,
2004; , 2009; & ,
2012; also see references in et al.,
2008, p. 319).
). Other groups of organisms are also thought to have followed a similar route (e.g., see ,
2000; & de ,
2000; , 2000; ,
2004; , 2009; & ,
2012; also see references in et al.,
2008, p. 319).
One can only speculate in a very preliminary manner about the palaeogeographic origins of Heydrichia, as the only known fossil specimens are from north-eastern Brazil. As noted above, however, there are at least two possible evolutionary origins of the genus: divergence from the sporolithaceaen genus Sporolithon, or divergence from a graticulacean alga such as Graticula. The oldest known fossils of Sporolithon come from early and late Hauterivian (c. 129.4-132.9 ± 1 Ma) deposits in eastern and south-eastern Spain and in Switzerland
(Figs. 11-12 ![]() ;
Map 2
;
Map 2 ![]() ), while Graticula is known only from mid-Silurian (c. 427-435 Ma) deposits in Wales and Sweden
(Map 2
), while Graticula is known only from mid-Silurian (c. 427-435 Ma) deposits in Wales and Sweden
(Map 2 ![]() ). We know of no records of either group from southern South Atlantic latitudes that predate Albian times. This suggests that Brazilian material of Heydrichia is more likely to have arrived from the Central Atlantic rather than from the southern South Atlantic. Moreover, during Aptian-Albian times, the Rio Grande Rise
- Walvis Ridge
Barrier (Map 2
). We know of no records of either group from southern South Atlantic latitudes that predate Albian times. This suggests that Brazilian material of Heydrichia is more likely to have arrived from the Central Atlantic rather than from the southern South Atlantic. Moreover, during Aptian-Albian times, the Rio Grande Rise
- Walvis Ridge
Barrier (Map 2 ![]() ) prevented full oceanic circulation in the South Atlantic and thus restricted incursions of marine organisms from the south (,
1999; & ,
2007, p. 1208-1209, Fig. 7; & ,
2012, p. 794).
) prevented full oceanic circulation in the South Atlantic and thus restricted incursions of marine organisms from the south (,
1999; & ,
2007, p. 1208-1209, Fig. 7; & ,
2012, p. 794).
Arrival of Heydrichia from the Central Atlantic also implies that one or more of the non-fossil species of Heydrichia found in Namibia and South Africa ( & van der , 2012) may ultimately have been derived from as yet unrecorded fossil or non-fossil taxa further north in the South Atlantic Ocean. Confirming studies of yet to be discovered non-fossil and fossil material from the west coast of Africa, particularly from Namibia to Ghana, are needed.
New Hauterivian record of Sporolithon: The occurrence of the related genus Sporolithon in Hauterivian strata (c. 129.4-132.9 ± 1 Ma) from Switzerland is recorded here for the first time
(Figs. 11-12 ![]() ). These deposits originated in the western part of the Tethys
(Map 2
). These deposits originated in the western part of the Tethys
(Map 2 ![]() ), which was open to the Central Atlantic Ocean during Hauterivian times but not during Albian times.
), which was open to the Central Atlantic Ocean during Hauterivian times but not during Albian times.
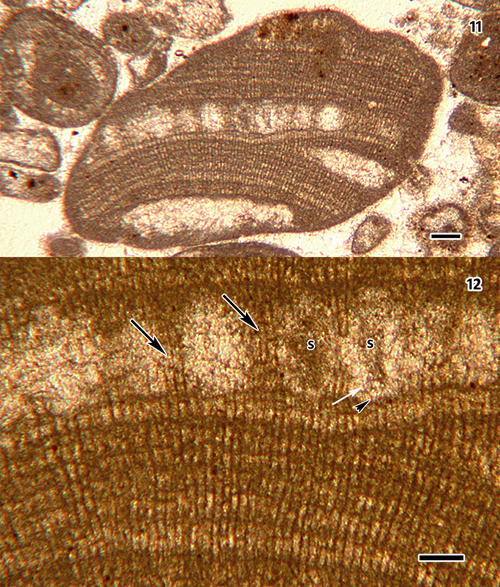
Click on thumbnail to enlarge the image.
Figs. 11-12: Sporolithon sp. [Specimen on thin section 7148 (15), housed at Muséum d'Histoire naturelle de la Ville de Genčve, Switzerland]. Fig. 11: Vertical section of thallus fragment containing a single buried sorus with evident sporangial compartments, and two cavities that may constitute remnants of additional sori. Scale bar = 100 µm. Fig. 12: Higher magnification of part of sorus showing sterile filaments (black arrows) between sporangial compartments, presumed remnants of two sporangia (s), and subtending vegetative filaments. Note partial remnant of sporangial compartment wall (white arrow); black arrowhead denotes base of compartment where stalk cell originally was. Scale bar = 50 µm.
Material examined: Thin section 7148 (15) [thin section 15 of sample 7148]. Vaulion, district of Jura-Nord Vaudois, canton of Vaud, Switzerland. UTM 32N 297415 E5173446 (latitude: 46°41'2"N; longitude: 6°21'3"E; see Google Maps: 46.68388928, 6.35083391). Rock sample collected by Jean & Bernard on 29 October 1993; thin section housed at the Muséum d'Histoire naturelle de la ville de Genčve, Switzerland. See et al. (1994) for a preliminary description of the Vaulion section, from which the Sporolithon sample comes.
Geological age: Late Hauterivian (Upper Hauterivian strata, probably Ligatus Zone, Ha5).
Geological stratum: "Urgonien blanc" ("niveaux ŕ
coraux"). Images: Figs. 11-12 ![]() .
.
The single, more or less transversely ovoid thallus fragment found
(Fig. 11 ![]() ) is c. 1.5 mm in greatest diameter, has a single buried sorus with evident sporangial compartments, and includes two cavities that may constitute remnants of additional sori. The occurrence of sori of sporangial compartments places the fragment in the Sporolithaceae and in Sporolithon.
) is c. 1.5 mm in greatest diameter, has a single buried sorus with evident sporangial compartments, and includes two cavities that may constitute remnants of additional sori. The occurrence of sori of sporangial compartments places the fragment in the Sporolithaceae and in Sporolithon.
Thallus growth-form cannot be determined with certainty, although the fragment may have been part of a rhodolith. While not evident, thallus construction is inferred to be monomerous; no dimerous species of Sporolithon are known. Epithallial cells were not preserved; vegetative cells are mostly (11-) 16-30 µm long and (6-) 8-13 µm in diameter. Unequivocal cell fusions and secondary pit-connections were not seen.
Sporangial compartments are c. 80-105 µm high and 70-80 µm in diameter, and most are separated by several interspersed sterile filaments (termed 'tetrasporangial paraphyses' by ,
1993) that are 3-4 cells long
(Fig. 12 ![]() ). Involucres were not evident. Presumed remnants of tetrasporangia occur in two of the compartments
(Figs. 11-12
). Involucres were not evident. Presumed remnants of tetrasporangia occur in two of the compartments
(Figs. 11-12 ![]() ), but subtending stalk cells and individual spores (presumably cruciately formed) were not observed. A layer of somewhat more conspicuous cells subtends the sorus
(Fig. 11
), but subtending stalk cells and individual spores (presumably cruciately formed) were not observed. A layer of somewhat more conspicuous cells subtends the sorus
(Fig. 11 ![]() ), but these are not distinctly more elongate than cells underneath them
(Fig. 12
), but these are not distinctly more elongate than cells underneath them
(Fig. 12 ![]() ).
).
The species to which the Swiss Hauterivian fragment of Sporolithon belongs is somewhat uncertain. Morphological-anatomical species concepts have been clarified for some non-fossil species (e.g., , 1993; et al., 2002), but as noted by et al. (2009, p. 417), concepts for fossil species (which constitute the vast majority of described species of Sporolithon) scarcely exist.
The only recent detailed account of Hauterivian material of Sporolithon is that of et al. (2007), who after studying relevant types and other specimens, recognized and compared two species in Late Hauterivian deposits of the Llŕcova Formation (Maestrat Basin, Iberian Chain, E. Spain): S. phylloideum ( & ) et al. and S. rude () & .
An analysis of characters used by (1993, p. 192, Table 2) and by et al. (2007, p. 87, Table 1) with those found in the Swiss fragment, however, suggests that the Swiss material may be specifically distinct from both S. phylloideum and S. rude. (1993) concluded that the presence or absence of an elongated cell layer beneath developing sporangial compartments/sori was of diagnostic significance at species level. According to et al. (2007), such an elongated layer occurs in S. phylloideum but not S. rude. The absence of such a layer in the Swiss fragment suggests that it is not conspecific with S. phylloideum.
In all species studied by (1993), sterile filaments ("tetrasporangial paraphyses") were interspersed between sporangial compartments. et al. (2007) reported such filaments in S. phylloideum. In S. rude, by contrast, filaments sometimes were present between compartments but in other cases were lost, giving the sorus the appearance of a multiporate conceptacle. The only intact sorus in the Swiss fragment had filaments between sporangial compartments, leaving uncertainty as to whether the fragment was conspecific with S. rude. Adding to this uncertainty are some difference in sporangial compartment height (130-170 µm in S. rude; 80-105 µm in the Swiss fragment), and while monomerous growth was clearly evident in S. rude, none was not evident in the Swiss fragment, thus precluding detailed comparisons of various vegetative features.
Of the five currently living species of Sporolithon studied by (1993), the Swiss Hauterivian fragment most closely resembles S. molle () , whose type is from the Red Sea. Fossil material of S. molle is recorded from Messinian age (7.246 - 5.333 Ma) reefs in the western and central Mediterranean Sea ( et al., 2009), but, according to those authors, the species apparently disappeared from the Mediterranean during the Messinian Salinity Crisis.
Sori resembling multiporate conceptacles are unknown both in S. molle and the Swiss Hauterivian fragment, but conceptacle compartment diameter differs somewhat (25-45 µm in S. molle; 70-80 µm in the Swiss fragment), and comparisons of various vegetative features are limited by the absence of obvious monomerous growth in the Swiss Hauterivian sample. Consequently, additional Swiss material of Hauterivian age from the same locality is needed before unequivocal identification is possible.
Living material of S. molle is reported from the Red Sea, the Persian Gulf, Indonesia, Japan, and tropical eastern Australia (, 1993; & , 1996; et al., 2000; et al., 2004; & , 2011; et al., 2013).
The main outcomes of this study are as follows:
We are indebted to Paulo , UNESPetro Associate Reseacher, for his invaluable assistance with the field work in Sergipe Basin. Sincere thanks are due to Masasuke , Davide , Daniela , and Yasufumi for providing excellent reviews that led to important improvements in the manuscript. Thanks also are due to Adele for assistance with several of the photographic images, to Jean and Bernard for sharing information on their Swiss material, and to the Department of Botany, La Trobe University, for the ongoing provision of facilities for WJW. This research is associated with the "Carbonatos do Brasil Project" linked to the Brazilian Sedimentology/Stratigraphy Net sponsored by Petrobras.
E., J.M., J.C., G.E., G., A.L., G., P.J. & D. (2013).- Deglacial mesophotic reef demise on the Great Barrier Reef.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 392, p. 473-494.
J., F. & J.C. (2010).- Integrating phylogeny, molecular clocks, and the fossil record in the evolution of coralline algae (Corallinales and Sporolithales, Rhodophyta).- Paleobiology, Gainesville, vol. 36, nº 4, p. 519-533.
M. (2009).- Paleogeografia do Atlântico Sul no Aptiano: um novo modelo a partir de dados micropaleontológicos recentes.- Boletim de Geocięncias da Petrobras, Rio de Janeiro, vol. 17, nº 2, p. 331-351.
M., J., C. & E. (2000).- Cretaceous dinoflagellate provincialism in Brazilian marginal basins.- Cretaceous Research, London, vol. 21, nº 2-3, p. 351-366.
C., J.-P. & L. (1995).- Hauterivian shallow marine calcareous biogenic mounds: S.E. Spain.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 119, nº 1-2, p. 3-17.
R.L.M de (2004).- Paleoceanografia e a evoluçăo do Atlântico Sul no Albiano.- Boletim de Geocięncias da Petrobras, Rio de Janeiro, vol. 12, nº 2, p. 1-19.
D.J. & R.K. (2007).- Systematic ichnology of microborings from the Cenozoic White Limestone Group, Jamaica, West Indies.- Scripta Geologica, Leiden, vol. 134, p. 77-108.
J.C., A., F.R. & J. (2009). Coralline algae (Corallinales, Rhodophyta) in western and central Mediterranean Messinian reefs. Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 275, p. 113-128.
C. & R. (1998).- Ordovician and Silurian coralline red algae.- Lethaia, Oslo, vol. 31, nº 3, p. 185-195.
C. & R. (2000).- Graticula and its derivatives, replacement name for the alga Craticula.- Lethaia, Oslo, vol. 33, nº 2, p. 82.
A., D., G. & P. (2010).- Re-deposited rhodoliths in the Middle Miocene hemipelagic deposits of Vitulano (Southern Apennines, Italy): Coralline assemblage characterization and related trace fossils.- Sedimentary Geology, Amsterdam, vol. 225, p. 50-66.
A. & P. (2009).- Trace fossil assemblages in rhodoliths from the Middle Miocene of Mt. Camposauro (Longano Formation, Southern Apennines, Italy).- Studi Trentini di Scienze Naturali, Acta Geologica, Trento, vol. 83 (2008), p. 165-176. Note: Vol. 83 is dated 2008 but appeared in 2009.
B., J., R., H. & R. (1994).- Révision de "l'Aptien" de Boveresse et nouvelle attribution chronostratigraphique de l'Urgonien du Jura neuchâtelois et vaudois (Suisse).- Publications du Département de Géologie et Paléontologie, Université de Genčve, nº 14, p. 25-55.
K.M.. S.C., P.LO. & X.J. (2013).- The ICS international chronostratigraphic chart.- Episodes, Bangalore, vol. 36, nº 3, p.199-204.
P.J., J.C., M. & J.M. (2004).- Holocene deep water algal buildups on the Eastern Australian Shelf.- Palaios, Lawrence, vol. 19, p. 598-609.
O. & E. (1996).- Marine algae of the Jubail marine wildlife sanctuary, Saudi Arabia. In: F., A.H. & I.A. (eds.), A marine wildlife sanctuary for the Arabian Gulf. Environmental research and conservation following the 1991 Gulf War Oil Spill.- NCWCD, Riyard and Senckenberg Research Institute, Frankfurt am Main, p. 199-289.
D. (2000).- Global stratigraphy, palaeobiogeography and palaeoecology of Albian-Maastrichtian pithonellid calcispheres: impact on Tethys configuration.- Cretaceous Research, London, vol. 21, nº 2-3, p. 315-349.
R.V. (1999).- Walvis Ridge barrier: its influence on palaeoenvironments and source rock generation deduced from ostracod distributions in the early South Atlantic Ocean. In: N.R., R.H. & V.S. (eds.), The oil and gas habitats of the South Atlantic.- Geological Society Special Publication, London, vol. 153, p. 293-302.
T., J.E.S., D.R., K. & W. (2009).- Common Coralline Algae of Northern New Zealand. An Identification Guide.- NIWA Information Series, Auckland, nº 70, 249 p.
B., P.-Y. & A.-F. (1991).- Constructions bio-sédimentaires laminées, Lithothamnium et Parachaetetes de la Formation Riachuelo (Albien) du bassin de Sergipe (Nord-Est du Brésil).- Geocięncias, Sâo Paulo, 10, p. 169-181.
B., D. & I.I. (2008).- Calcareous algae from Upper Albian - Cenomanian strata of the Potiguar basin (NE Brazil).- Geologia Croatica, Zagreb, vol. 61, nº 2-3, p. 311-320 (Pls. 1-2).
B., D. & I.I. (2012b, online first).- A new mid-Cretaceous Neomeris (dasycladacean alga) from the Potiguar Basin, Brazil.- Facies, Erlangen, vol. 59 (2013), nº 1, p. 221-230.
B., D., I.I. & P. (2012a, online first).- Brasiliporella, a new mid-Cretaceous dasycladacean genus: the earliest record of the Tribe Batophoreae.- Facies, Erlangen, vol. 59 (2013), nº 1, p. 207-220.
M.D. (2012).- How many species of algae are there?.- Journal of Phycology, Somerset, vol. 48, nº 5, p. 1057-1063.
A.S., W.J., T., K. & W. (2005).- Coralline Algae of Central New Zealand. An Identification Guide to Common 'Crustose' Species.- NIWA Information Series, Auckland, nº 57, 145 p.
A.S., W.J., J.M. & C.F.D. (2013).- A monographic account of Australian species of Amphiroa (Corallinaceae, Rhodophyta).- Australian Systematic Botany, Melbourne, vol. 26, nº 2, p. 81-144.
A.S., W.J. & A.J.K. (2002).- The Sporolithaceae (Corallinales, Rhodophyta) in south-eastern Australia: taxonomy and 18S rRNA phylogeny.- Phycologia, Lawrence, vol. 41, nº 3, p. 207-227.
A.S., W.J. & A.J.K. (2009).- The genus Amphiroa (Lithophylloideae, Corallinaceae, Rhodophyta) from the temperate coasts of the Australian continent, including the newly described A. klochkovana.- Phycologia, Lawrence, vol. 48, nº 4, p. 258-290.
D.L. (2010).- Terms used in bionomenclature.- Global Biodiversity Information Facility, Copenhagen, 216 p. Online at http://links.gbif.org/gbif_terms_nomenclature_guide_en_v1.pdf (last visited 06 November 2013).
F. (1897).- Corallinaceae, inbesondere Melobesieae.- Berichte der Deutschen Botanischen Gesellschaft, Berlin, Band XV, p. 34-71 [Effective publication date: 25.ii.1897 (see & , 1998, p. 680)]. Online at http://biodiversitylibrary.org/page/5001350 (last visited 08 December-2013).
J.R., J.C., J., W.J. & J.M. (2008).- Analysis of ancient DNA from fossil corallines (Corallinales, Rhodophyta).- Journal of Phycology, Moss Landing, vol. 44, nº 2, p. 374-383.
Y., D. & W.J. (2009).- Re-assessment of the type collections of fourteen corallinalean species (Corallinales, Rhodophyta) described by W. (1942-1960).- Palaeontology, London, vol. 52, nº 2, p. 401-427.
Y., D. & W. J. (2012).- Typification and reassessment of seventeen species of coralline red algae (Corallinales and Sporolithales, Rhodophyta) described by W. during 1954-1978.- Journal of Systematic Palaeontology, London, vol. 10, nº 1, p. 171-209.
D.W. & Y.M. (1995).- Heydrichia groeneri sp. nov.: a new species of crustose coralline alga (Rhodophyta, Sporolithaceae) from South Africa and Namibia.- Phycologia, Lawrence, vol. 34, nº 1, p. 51-57.
E.A.M. & I. de (2000).- Late Cretaceous foraminiferal biogeography (Families Bolivinidae, Buliminellidae, Gavelinellidae, Siphogenerinoididae, Turrilinidae) in northeastern Brazilian shelf and central West African basins.- Cretaceous Research, London, vol. 21, nº 2-3, p. 381-405.
L., C.E., L. & G.W. (2010).- Multigene phylogenetic analyses support recognition of the Sporolithales ord. nov.- Molecular Phylogenetics and Evolution, San Diego, vol. 54, n º 1, p. 302-305.
L. & G.W. (2007).- A nuclear phylogeny of the Florideophyceae (Rhodophyta) inferred from combined EF2, small subunit and large subunit ribosomal DNA: establishing the new red algal subclass Corallinophycidae.- Molecular Phylogenetics and Evolution, San Diego, vol. 43, nº 3, p. 1118-1130.
M., P.J. & J.C. (2000).- Coralline algal nodules off Fraser Island, eastern Australia. Facies, Erlangen, vol. 42, p. 25-34.
J.G. (2000).- Continental break up and the distribution of fishes of Western Gondwana during the Early Cretaceous.- Cretaceous Research, London, vol. 21, nº 2-3, p. 281-314.
G. & E. van der (2012).- Heydrichia cerasina sp. nov. (Sporolithales, Corallinophycidae, Rhodophyta) from the southernmost tip of Africa.- Phycologia, Lawrence, vol. 51, nº 1, p. 11-21.
S. & Y. (2011).- Rhodoliths from deep fore-reef to shelf areas around Okinawa-jima, Ryukyu Islands, Japan. Marine Geology, Amsterdam, vol. 282, p. 215-230
J. (Chairman), F.R., W.R., V., W., D.L., P.S., S., K., J., van W.F., G.F., J.H. (Members) & N.J. (Secretary of the Editorial Committee) (2012).- International Code of Nomenclature for algae, fungi and plants (Melbourne Code) adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011.- Regnum Vegetabile, vol. 154, Königstein, xxx + 240 p. Online at http://www.iapt-taxon.org/nomen/main.php (last visited 15 January 2014).
W.A. (2012).- Phylum Rhodophyta: red algae. In: D.P. (ed.), New Zealand inventory of biodiversity. Volume Three. Kingdoms Bacteria, Protozoa, Chromista, Plantae, Fungi.- Canterbury University Press, Christchurch, p. 327-346.
J.A. & N.B. (2007).- Subsurface seepage of seawater across a barrier: a source of water and saltto peripheral salt basins.- Geological Society of America Bulletin, Boulder, vol. 119, nº 9-10, p. 1201-1217.
S.F. & J.-P. (2012).- Early Cretaceous ostracod biostratigraphy of eastern Brazil and western Africa: An overview.- Gondwana Research, London, vol. 22, nº 3-4, p. 772-798.
C.R. (2002, continuously updated).- PALEOMAP website.- Online at http://www.scotese.com (last visited 5 January 2014).
G.J.S. (1990, unpublished).- Fácies, modelo deposicional e diagenese da seqüencia carbonática albo-cenomaniana (Formaçao Ponta do Mel) da Bacia Potiguar.- Dissertaçao de Mestrado, Instituto de Geociencias, Universidade Federal do Rio de Janeiro, 153 p.; Anuário do Instituto de Geocięncias (1993), Rio de Janeiro, vol. 16, p. 108-109 (Resumos das teses de mestrado).
B. (2014, continuously updated).- Index Herbariorum: A global directory of public herbaria and associated staff.- New York Botanical Garden's Virtual Herbarium. Online at http://sweetgum.nybg.org/ih/ (last visited 14 January 2014).
S., J., J.C. & C. (2007).- Late Hauterivian coralline algae (Rhodophyta, Corallinales) from the Iberian Chain (E. Spain). Taxonomy and the evolution of multisporangial reproductive structures.- Facies, Erlangen, vol. 53, p. 79-95.
R.A. & M.A. (2001).- Heydrichia homalopasta sp. nov. (Sporolithaceae, Rhodophyta) from Australia.- Botanica Marina, Berlin, vol. 44, nº 3, p. 237-244.
R.A., Y.M. & D.W. (1994).- Heydrichia woelkerlingii gen. et sp. nov., a newly discovered non-geniculate red alga (Corallinales, Rhodophyta) from Cape Province, South Africa.- Phycologia, Lawrence, vol. 33, nº 3, p. 177-186.
R.A., W.J., A.S. & M.S. (1995).- An account of the red algal genus Sporolithon (Sporolithaceae, Corallinales) in southern Australia.- Australian Systematic Botany, Melbourne, vol. 8, p. 85-121.
C.V.G, P. de, A.J.V. & R.C. (2013).- Depositional, diagenetic and stratigraphic aspects of microfacies from Riachuelo Formation, Albian, Sergipe Basin, Brazil.- Geologia USP, (série cientifica), Saő Paulo, vol. 13, nº 4, p. 29-48.
G., M., P. & D. (2000).- Revision and re-documentation of M. 's species of Archaeolithothamnion from the Tertiary Piedmont Basin (NW Italy).- Revista Italiana di Paleontologia e Stratigrafia, Milano, vol. 106, no. 2, p. 191-202.
E. (1993).- The genus Sporolithon (Sporolithaceae fam. nov., Corallinales, Rhodophyta) from the Spermonde Archipelago, Indonesia.- Phycologia, Lawrence, vol. 32, nº 3, p. 184-196.
W.J. (1988).- The Coralline red algae: An analysis of the genera and subfamilies of non-geniculate Corallinaceae.- British Museum (Natural History) and Oxford University Press, London and Oxford, xi + 268 pp.
W.J., L.M. & A. (1993).- Growth-forms in non-geniculate coralline red algae (Corallinales, Rhodophyta).- Australian Systematic Botany, Melbourne, vol. 6, nº 4, p. 277-293.
W.J. & D. (1998).- Appendix 3. Effective Publication Dates. In: W.J. & D. (eds.), Non-geniculate Coralline red algae and the Paris Museum: Systematics and scientific history.- Publications scientifiques du Muséum/A.D.A.C., Paris, p. 671-682 [Effective publication date: 31.xii.1998 (stated on p. 1)].
The use of a question mark after the genus name indicates slight taxonomic uncertainty about generic placement of the new species, as fully explained in Results and Discussion, but does not affect valid publication of the name [see Art 36.1, last sentence, and Art. 52, Note 1 of the International Code of Nomenclature ( et al., 2012)].
The matter of whether presumed sporangial complexes in which involucres, sporangia and pore canals have not been preserved are reproductive structures or, instead, trace fossils (tunnels) produced by micro-borers is fully considered below.
As already noted, however, the possibly ancestral Graticula occurs in northern hemisphere mid-Silurian deposits.