◄ Carnets Geol. 14 (8) ►
![]()
Sommaire
[Introduction] [Discussion] [Conclusion]
et ... [Références]
BrachNet, 20, Rue Chaix, F-13007 Marseille (France)
Manuscrit en ligne depuis le 19 juin 2014
[Éditeur scientifique : Bruno ;
éditeur technique : Christian C. ;
chargé de l'édition en langue anglaise : Phil ]
![]()
Anomia turbinata, ou Anomie conique, (= Novocrania turbinata) a été décrite par (1795) dans le Bathyal des côtes du Royaume des Deux-Siciles, avec comme synonyme Patella anomala , 1776 (= N. anomala). Longtemps considérée comme la forme méditerranéenne de N. anomala, son histoire est brièvement décrite. Récemment, N. turbinata a été considérée par plusieurs auteurs, comme une espèce valide, mais sur quelques caractères ne correspondant pas à ceux de la description originelle ; ils apparaissent comme des variations des caractères de N. anomala. La présence conjointe des deux "espèces" dans plusieurs localités surtout sur le plateau continental tend à accréditer leur synonymie. Celle-ci a été récemment corroborée par des analyses moléculaires et est discutée en prenant aussi en compte les caractéristiques des bassins méditerranéens et leur histoire depuis le Miocène.
Novocrania ; taxinomie ; anomala ; turbinata ; synonymie ; mer Méditerranée.
C.C. (2014).- Novocrania turbinata synonyme de N. anomala.- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 8, p. 159-171.
Novocrania turbinata synonym of N. anomala.- Anomia turbinata, or conical Anomia (= Novocrania turbinata), was described by (1795) in the bathyal environment off the coast of the Kingdom of the Two Sicilies, Patella anomala , 1776 (= N. anomala) being considered a synonym. The history of this species, commonly considered as the Mediterranean form of N. anomala, will be described. Recently, several authors have described N. turbinata as a valid species on the basis of shell variations, as compared to N. anomala. After analysis of the taxonomic validity of these characters, both species are considered as synonymous. That is supported by their occurrence in various localities, mainly in the continental shelf. Their synonymy has been corroborated by molecular analyses and is discussed with reference to the characteristics of the Mediterranean basins and their history since the Miocene.
Novocrania; taxonomy; anomala; turbinata; synonymy; Mediterranean Sea.
Anomia turbinata, ou Anomie conique, a été décrite par
(1795) (Fig. 1 ![]() ; Tableau 1
; Tableau 1 ![]() ) sur une branche du scléractiniaire Dendrophyllia ramea (en fait, il s'agit de Dendrophyllia cornigera avec laquelle elle est parfois confondue - H. , 2014, communication personnelle). Elle a été récoltée dans un fond qui se rencontre en mer Méditerranée (ici en Sicile) dans la partie supérieure de la pente continentale (étage Bathyal supérieur) entre 150 et 600 m de profondeur. Le peuplement est caractérisé par des branches mortes et vivantes de Dendrophyllia sur lesquelles se fixent notamment diverses espèces de brachiopodes : Novocrania anomala, N. turbinata, Mergerlia truncata (et ses formes synonymes monstruosa et echinata), Terebratulina retusa et Megathiris detruncata (Fig. 2
) sur une branche du scléractiniaire Dendrophyllia ramea (en fait, il s'agit de Dendrophyllia cornigera avec laquelle elle est parfois confondue - H. , 2014, communication personnelle). Elle a été récoltée dans un fond qui se rencontre en mer Méditerranée (ici en Sicile) dans la partie supérieure de la pente continentale (étage Bathyal supérieur) entre 150 et 600 m de profondeur. Le peuplement est caractérisé par des branches mortes et vivantes de Dendrophyllia sur lesquelles se fixent notamment diverses espèces de brachiopodes : Novocrania anomala, N. turbinata, Mergerlia truncata (et ses formes synonymes monstruosa et echinata), Terebratulina retusa et Megathiris detruncata (Fig. 2 ![]() ; Pl. 1
; Pl. 1 ![]() ). Les deux premières espèces sont mentionnées par
(1795) dans la légende de sa planche XXX (Fig. 1
). Les deux premières espèces sont mentionnées par
(1795) dans la légende de sa planche XXX (Fig. 1 ![]() ), respectivement sous Anomia truncata et Anomia Caput Serpentis (sic).
), respectivement sous Anomia truncata et Anomia Caput Serpentis (sic).

Click on thumbnail to enlarge the image.
Figure 1 : Extrait en fac-similé des figures sur Anomia turbinata (Anomie Conique) de la
Planche XXX de (1795), avec les légendes originales :
Figure 1: Extract in facsimile of the figures on Anomia turbinata (Conical Anomie) of the
Plate XXX of (1795), with the original captions:
EXPLICATION DE LA PLANCHE XXX.
On a dessiné dans cette Planche toutes les especes d'Anomies, aussi-bien que
l'anatomie de leurs Mollusques. (...)
Fig. 15. Branche de Madrepore Rameuse (Madrepora Ramea Linn.). Le Zoo-
phite logé dans les bouts étoilés des branches de cette Madrepore, étend ses
prolongemens de maniere, que tous ce bouts-là paroissent peints d'une jolie
couleur jaune, et quelquefois de rouge. Lorsqu'on la tire de la mer, elle le
dispute en beauté aux plus jolies productioms marines. Outre la Millepore Cel-
lulaire f, et la Réticulaire h, on trouve attaché à la dite branche l'Anomie en-
forme d'écaille, a (Anomia Squamula Linn.), qu'on a déssiné un peu grossie
dans la Fig. 18: les deux Anomies Tronquées, b (Anomia Truncata Linn.): la Tête
de Serpent, y (Anomia Caput Serpentis Linn.); et l'Anomie Conique c (Anomia
Turbinata *), dont les valves sont déssinées à part en d et en e: l'inférieure d
est tellement fixée à la Madrepore, qu'il est très-difficile de l’en détacher. (...)
Fig. 21. Le battant inferieur de l'Anomie Conique, grossi.
Fig. 24. Le lobe supérieur du manteau du Mollusque de l'Anomie Conique, contenant
l'ovaire ramifié dd.
Fig. 22. Le Mollusque de cette Anomie adhérent à son battant inférieur. a, a,
c, c sont les tendons des muscles adducteurs. L'abdomen est placé dans l'espa-
ce ee, la bouche en s: ir; ir sont les ouïes contournées naturellement en for-
me de cornes de belier, et garnies de cils tout à l'entour.
Enfin la Fig. 23 montre ce même Animal avec ses ouïes bb renversées en dessus,
pour mettre à nud le lobe inférieur du manteau zz, et l'ovaire n, n, qui l'orne
agréablement.
Comme la provenance exacte de cette branche n'est pas donnée par (1795), il faut donc nous reporter au titre de son livre : Testacea utriusque Siciliae (...) = Coquilles de l'une et l'autre Sicile (...) : en 1795, le Royaume des Deux-Siciles était formé par le Royaume de Naples et par la Sicile, la capitale étant Naples. Dans sa préface (, 1795, vol. 1) indique que les récoltes venaient de la mer Adriatique, la mer de Sicile, la mer Tyrrhénienne. Giuseppe Saverio (1746-1825) était un physicien, médecin et naturaliste italien. Il s'installe à Naples en 1790. Il fut le tuteur du prince-héritier François de . Ses collections sont au Museo Zoologico, Centro Musei delle Scienze Natural, Napoli.

Click on thumbnail to enlarge the image.
Figure 2 : Fond rocheux dans le Bathyal supérieur (255 m de profondeur) au large de l'Ile de Porquerolles (Provence, France) avec des Novocrania anomala (N), des Megathiris detruncata (M) et de nombreuses Megerlia truncata, dont certaines présentent la forme monstruosa avec déformation de la coquille en fonction du substrat. Ces espèces se trouvent aussi sur des branches du scléractiniaire Dendrophyllia cornigera (Pl. 1 ![]() ).
).
Figure
2: Hard substrate in the upper Bathyal zone (255 m depth) off Porquerolles Island (Provence, France) with Novocrania anomala (N), Megathiris detruncata (M) and numerous Megerlia truncata, some of which have the form called monstruosa with shell deformation induced by the substrate. Those species are also found on branches of the stony coral Dendrophyllia cornigera (Pl. 1 ![]() ).
).
La diagnose de Anomia turbinata donnée de
(1795, p. 189) est la suivante :
"Testae characteres. Testa conica, transversim striata, integerrima; valva altera subrotunda, parasitica, crassa, imperforata" : elle est suivie dans la même page par un paragraphe intitulé
"Testae descriptio" (= description de la coquille). Cet auteur mentionne la synonymie avec Crania anomala de
(1788) décrite et représentée dans Zoologia Danica (,
1788 ; Fig. 3 ![]() ). La diagnose originale de avait été publiée en
1776 sous le nom de Patella anomala, comme suit :
"testa rudi, sulca, orbiculari, vertice submarginali" ; elle est suivie d'une croix signifiant
"a me, detectae indicantur" (espèce découverte par moi : ,
1776, p. VII). L'espèce N. turbinata est connue en Méditerranée depuis le Pliocène, notamment en Italie (,
1865) et en Espagne (,
1977 ; & ,
2007), tout comme N. anomala en Italie (,
1979 ; ,
1987 ; ,
1983, 1988,
1994a, 1996a ; ,
2001 ; et al.,
2005 ; & , 2006; et al.,
2009), en Grèce ( et al.,
2006 ; ,
2007), ainsi qu'à l'Oligocène en France ( et al.,
2013).
). La diagnose originale de avait été publiée en
1776 sous le nom de Patella anomala, comme suit :
"testa rudi, sulca, orbiculari, vertice submarginali" ; elle est suivie d'une croix signifiant
"a me, detectae indicantur" (espèce découverte par moi : ,
1776, p. VII). L'espèce N. turbinata est connue en Méditerranée depuis le Pliocène, notamment en Italie (,
1865) et en Espagne (,
1977 ; & ,
2007), tout comme N. anomala en Italie (,
1979 ; ,
1987 ; ,
1983, 1988,
1994a, 1996a ; ,
2001 ; et al.,
2005 ; & , 2006; et al.,
2009), en Grèce ( et al.,
2006 ; ,
2007), ainsi qu'à l'Oligocène en France ( et al.,
2013).
Les deux espèces actuelles nommées Novocrania anomala et N. turbinata (Tableau 1) ont toujours fait débat quant à leur état comme espèces séparées ou comme synonymes. Donc, sans faire ici un historique exhaustif, quelques faits marquants méritent d'être relatés avant de conclure sur la place de chacune au sein du genre.

Click on thumbnail to enlarge the image.
Figure 3 : Fac-similé des figures sur Patella - Crania anomala de la
planche de (1788), avec les légendes originales (en p. 4 comme indiqué sur la figure) ci-dessous.
Figure 3:
Extract in facsimile of the figures on Patella - Crania anomala of 's
(1788) plate, with original captions (on p. 4 as stated on the figure)
below.
Fig. 1. Saxum tribus patellis passim obsitum ac vnius
ouaria ramosa derelicta, testamque Serpulae ac
frustum Milleporae polymorphae exhibet.
Fig. 2. Faciam exteriorem
Fig. 6. interiorem patellae
Fig. 3. vero ipsum inhabitatorem naturali magnitudine offert.
Fig. 4. Animalculum cincinnatum ligamentis et muscu-
lis testae adhaerens aduersa parte aucta magni-
tudine monstrat.
Fig. 5. lobos spirales cirratos e testa euulsos auersa parte exhibet.
Fig. 7. ouaria ramosa musculorumque vestigia aucta magnitudine.
Fig. 8. ouula maxime aucta.
En l'absence d'une diagnose vraie (voir Commission Internationale de Nomenclature Zoologique, 1999, et Conclusion) permettant d'identifier Novocrania anomala et N. turbinata, l'usage du nom de l'une ou de l'autre a toujours été laissé à l'appréciation de l'auteur qui identifiait des cranies de Méditerranée. En fait, les auteurs méditerranéens souvent rapportaient leurs cranies à turbinata, car elle représentait une forme méditerranéenne, tandis que les autres auteurs plaçaient turbinata comme synonyme de N. anomala - citons : (1817), (1819), (1834), (1862), (1878), (1888), & (1891), (1899), ... Les exemplaires méditerranéens étaient donc identifiées sous anomala ou turbinata au cours du XIXe siècle, puis turbinata fut considéré comme synonyme d'anomala jusqu'à la publication de (1988) qui relança le débat.
(1888) note : " (1871) seems disposed to regard Crania turbinata as a variety of the northern C. anomala."
Dans sa thèse, faite au Laboratoire Arago à Banyuls (France), (1885) écrit : "L'espèce de Banyuls ne peut être que la Crania rostrata ; elle répond en effet à sa description. Mais il y a si peu de différences avec la Crania anomala, qui ne diffère guère, elle non plus de la Crania Pourtalesi, qu'il est possible que ces trois Cranies ne soient que des variétés de la même espèce."
(1921) regroupe des espèces de Crania méditerranéennes (dont Crania turbinata) sous le nom de Crania kermès da et (in da , 1770), sauf Crania rostrata , 1828, tout en notant : "The specific relations between the various Mediterranean Cranias require more thourough investigation."
| Novocrania anomala (, 1776) | Novocrania turbinata (1795 |
| [Patella anomala , p. 237, 1776] | [Anomia turbinata , p. 189, 1795] |
| Hår-Krøllen (Danemark) - Baltique | Côtes du royaume des Deux-Siciles (Italie) – mer Méditerranée |
|
Synonymes |
|
| Orbicula anomala , p. 435, 1799 | Patella kermes da et in da , 1770 |
| Patella distorta , 1808 | Crania personata , 1818 (non ) |
| Orbicula norvegica , 1819 | Crania ringens , 1828 |
| Anomia turbinata , 1817 (non ) | Crania rostrata , 1828 |
| Orbicula norvegica , 1817 | Orbicula turbinata & , 1836 |
| Crania personata (part.) , 1818 | |
| Discina ostreoides , 1819 (not ) | * Patella anomala , 1776, est synonyme selon (1795) |
| Criopus anomalus , 1822 | |
| Crania anomala , 1846 | |
| Criopus orcadensis | |
| Crania anomala var. alba , 1869 | |
Tableau 1 : Localité-type et synonymes pour les deux espèces de Novocrania anomala et N. turbinata. Les espèces de craniidés ont souvent été décrites sous les noms de genres Patella, Anomia ou Orbicula, puis elles ont été mises dans le genre Crania , 1781 (voir ,
2009). Ensuite, &
(1986) créent le genre Neocrania pour les espèces actuelles, devenu Novocrania & ,
2001.
Table 1:
Type locality and synonyms for both species Novocrania anomala and N. turbinata. Craniids species have often been described under the genus names Patella, Anomia or Orbicula. Later they were placed in the genus Crania , 1781 (see ,
2009). Then, &
(1986) created the genus Neocrania for the living species, changed to Novocrania & ,
2001.
(1979) avait confirmé la présence d'une seule espèce (N. anomala) en mer Méditerranée, en rappelant des variations dans la coquille liée principalement au substrat. Ceci avait déjà été souligné auparavant, notamment par (1888). Ces observations ont été corroborées au cours de mes nombreuses récoltes de N. anomala en mer Méditerranée (, 1988a, 1988b).
C'est la description de (1988) qui a relancé le débat sans faire une étude exhaustive de N. turbinata et le doute s'est à nouveau installé. Néanmoins, dans l'attente d'une étude morpho-anatomique significative permettant de trancher entre ces deux "espèces", je maintenais la possibilité que N. turbinata puisse quand même être une espèce distincte ( & , 2005 ; , 2010, 2012). Suite à des travaux signalant la présence de N. turbinata dans l'océan Pacifique (voir ci-dessous), il était nécessaire de reprendre l'historique de cette espèce, ce qui est l'objet du présent travail.
Rappelons que la localité-type de N. turbinata est située dans l'étage
Bathyal sur les côtes du royaume des Deux-Siciles (Italie), peut-être dans la baie de Naples où habitait G.S. (Fig. 6 ![]() ). Or,
(1988) indique que, dans cette baie,
"there are specimens lacking the prominently raised group of anterior muscle
scars", ce qui correspond bien aux figures de
(1795) (Fig. 1
). Or,
(1988) indique que, dans cette baie,
"there are specimens lacking the prominently raised group of anterior muscle
scars", ce qui correspond bien aux figures de
(1795) (Fig. 1 ![]() ). Dès lors, l'identification par
(1988) devient sujet à caution, et cet auteur poursuit :
"whereas recognized Crania anomala (), I believe the eastern Mediterranean specimens discussed herein should be attributed to the species turbinata."
). Dès lors, l'identification par
(1988) devient sujet à caution, et cet auteur poursuit :
"whereas recognized Crania anomala (), I believe the eastern Mediterranean specimens discussed herein should be attributed to the species turbinata."
Seules quatre valves dorsales, récoltées entre 44 et 49 m, ont été décrites par (1988) ; il mentionne que, selon (1862), les valves ventrales de anomala et turbinata (exemplaires au British Museum, Londres) sont "similarly calcified and closely comparable in morphologies." En revanche, il note aussi qu'il y aurait une différence en calcification des valves ventrales entre turbinata et anomala d'après (1979), ce qu'il n'a pu vérifié, et il écrit "I believe to be true." Néanmoins, il n'exclut pas un possible cline de N. anomala depuis l'océan Atlantique jusqu'au bassin oriental méditerranéen, en suggérant qu'il n'y aurait qu'une seule espèce atlantico-méditerranéenne N. anomala avec des variations géographiques de certains caractères.
&
(2001) reprennent les critères utilisés par
(1988) pour séparer des individus qu'ils attribuent soit à anomala, soit à turbinata, les uns et les autres pouvant être présents dans une même localité en mer Méditerranée et dans l'océan Atlantique. Concernant la bathymétrie des localités méditerranéennes, étudiées par ces auteurs, elles sont toutes situées sur le plateau continental, entre quelques mètres et 44 m, et on y trouve principalement N. turbinata, mais dans deux localités profondes, au-delà de 100 m dans le Bathyal, seule N. anomala a été identifiée. Ceci confirme les résultats des campagnes le long des côtes provençales et corses par la RCP-CNRS 728 entre 1983 et 1988 (,
1988a, 1988b), montrant que la densité maximale de N. anomala en mer Méditerranée se situe au-delà de 100m de profondeur. À rappeler que N. turbinata a été décrite par
(1795) du Bathyal (Fig. 1 ![]() ). Ce type fond a été dragué à de nombreuses reprises dans l'étage Bathyal supérieur au large des côtes de Provence et de Corse et aussi observé lors de plongées en submersible (,
1987) (Figs. 2
). Ce type fond a été dragué à de nombreuses reprises dans l'étage Bathyal supérieur au large des côtes de Provence et de Corse et aussi observé lors de plongées en submersible (,
1987) (Figs. 2 ![]() , 6
, 6 ![]() ; Pl. 1
; Pl. 1 ![]() ). et al.
(2002) donne la même distribution bathymétrique pour ces deux espèces dans le bassin oriental méditerranéen. Enfin, et al.
(2004) mentionnent Novocrania anomala comme "Cemented to cave walls and roofs; boulders; coralligène; mainly shallow, infralittoral-bathyal - Recent: NE Atlantic, Mediterranean",
tandis que N. turbinata est "Cemented to cave walls and roofs; boulders; coralligène; mainly shallow, infralittoral-circalittoral - Recent: NE Atlantic, Mediterranean; - Miocene: Italy" donc avec une distribution limitée au plateau continental.
). et al.
(2002) donne la même distribution bathymétrique pour ces deux espèces dans le bassin oriental méditerranéen. Enfin, et al.
(2004) mentionnent Novocrania anomala comme "Cemented to cave walls and roofs; boulders; coralligène; mainly shallow, infralittoral-bathyal - Recent: NE Atlantic, Mediterranean",
tandis que N. turbinata est "Cemented to cave walls and roofs; boulders; coralligène; mainly shallow, infralittoral-circalittoral - Recent: NE Atlantic, Mediterranean; - Miocene: Italy" donc avec une distribution limitée au plateau continental.
Si cette répartition bathymétrique devait se confirmer, il faudra alors créer une nouvelle espèce et restreindre N. anomala à l'étage Bathyal en considérant que l'autre espèce bathyale N. turbinata est synonyme de N. anomala, ou bien nommer l'espèce profonde N. turbinata en mer Méditerranée avec N. anomala comme espèce vicariante dans l'océan Atlantique. Il faut remarquer que les espèces méditerranéennes profondes, généralement eurybathes, ont souvent des espèces vicariantes vivant à faible profondeur dans l'océan Atlantique, ou une large répartition géographique extra-méditerranéenne (voir & ,
2004). Enfin, en profondeur, c'est sur la pente continentale méditerranéenne que Novocrania anomala montre les plus fortes densités (Fig. 4 ![]() ; Pl. 1
; Pl. 1 ![]() ).
).
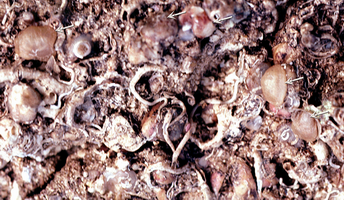
Click on thumbnail to enlarge the image.
Figure 4 : Fond rocheux avec des Novocrania anomala (flèches) dans l'étage Bathyal supérieur près de Calvi (Corse) à 210 m de profondeur. On distingue aussi quelques anciennes valves ventrales.
Figure 4:
Bedrock with Novocrania anomala (arrows) in the upper Bathyal
zone off Calvi (Corsica) at 210 m depth. Some older ventral valves can be
observed.
Le critère pris en compte par &
(2001) pour N. turbinata est une forte calcification de la valve ventrale, alors que N. anomala n'en montrerait qu'une faible. Or, chez les Novocrania méditerranéennes profondes, cette valve est toujours plus ou moins calcifiée (Fig. 4 ![]() ) et le même constat a été fait sur les N. anomala, récoltés dans le golfe de Gascogne au large d'Avilés (Espagne) à 120 m de profondeur (Fig. 5
) et le même constat a été fait sur les N. anomala, récoltés dans le golfe de Gascogne au large d'Avilés (Espagne) à 120 m de profondeur (Fig. 5 ![]() ). Concernant la calcification des valves de brachiopodes, son importance dépend des caractéristiques physico-chimiques des couches d'eaux atlantiques et méditerranéennes, qui sont à prendre en compte dès que la calcification des coquilles est évoquée (,
1997) : celle-ci a tendance à être plus forte en profondeur jusqu'à la CCD (Carbonate Compensation Depth). En outre, ce critère n'a aucune incidence dans la taxinomie ; pourtant, il est parfois utilisé selon son degré d'importance dans les formes fossiles pour créer des (fausses) nouvelles espèces (voir ,
2002).
). Concernant la calcification des valves de brachiopodes, son importance dépend des caractéristiques physico-chimiques des couches d'eaux atlantiques et méditerranéennes, qui sont à prendre en compte dès que la calcification des coquilles est évoquée (,
1997) : celle-ci a tendance à être plus forte en profondeur jusqu'à la CCD (Carbonate Compensation Depth). En outre, ce critère n'a aucune incidence dans la taxinomie ; pourtant, il est parfois utilisé selon son degré d'importance dans les formes fossiles pour créer des (fausses) nouvelles espèces (voir ,
2002).

Click on thumbnail to enlarge the image.
Figure 5 : Valve ventrale de Novocrania anomala sur un fond rocheux vers 120 m de profondeur au large d'Avilés (Golfe de Gascogne, Espagne : Campagne Fauna II, juin 1991, programme Fauna Ibérica, N/O García del Cid).
Figure 5:
Ventral valve of Novocrania anomala on a rocky bottom at 120 m depth off Avilés (Bay of Biscay, Spain: Fauna II Expedition, June 1991, Fauna Ibérica program, R/V García del Cid).
Quant à la valve dorsale, & (2001) utilisent aussi la calcification qui modifie les impressions musculaires laissées sur la valve. Malheureusement, aucune étude des variations de ces critères dans l'une et l'autre valve sur un échantillonnage représentatif n'a été faite, avec un schéma comparatif permettant de bien appréhender les différences fondamentales entre anomala et turbinata, notamment dans la disposition des muscles dans chaque valve, et non sur leur seules impressions dans les valves dorsales. Mais, sans corroborer le possible cline Ouest-Est de (1988), ces deux auteurs créent deux morphotypes qu'ils considèrent chacun comme des espèces distinctes, en précisant "while we provisionally recognise the two forms here as separate species, this will need to be confirmed by molecular biology studies."
Les récoltes de Novocrania tant en plongée dans les grottes sous-marines des environs de Marseille, qu'au large des côtes de Provence et de Corse (,
1988a, 1988b), au-delà de 100 m de profondeur, n'ont pas permis de distinguer au-delà des variations des caractères à l'existence de deux espèces distinctes. Il faut aussi souligner que la présence de deux espèces d'un même genre dans une même biocœnose est un fait exceptionnel qui implique que ces espèces aient au moins un caractère phylogénétique qui les distingue nettement. Des cas sont bien connus dans divers groupes zoologiques, ce sont fréquemment les caractères taxinomiques liés à la reproduction et à l'éthologie qui distinguent deux espèces voisines. En conséquence, les variations indiquées tant par
(1988) que par &
(2001) ne peuvent suffire pour séparer deux espèces. Il est indispensable d'avoir une diagnose complète basée sur tous les caractères phylogénétiques disponibles et établis selon la méthode cladistique, ceux-ci sont aussi les caractères taxinomiques à utiliser dans la diagnose. Ces auteurs n'expliquent pas sur quelle diagnose ils ont identifié leurs exemplaires comme étant N. turbinata. À comparer avec N. anomala, description et figures de N. turbinata par
(1795) (Fig. 1 ![]() ) sont bien plus précises que celles de
(1788), et, la première a une localité-type littorale et l'autre
bathyale. Néanmoins, la distribution bathymétrique de N. anomala s'étend depuis quelques mètres jusqu'à 1478 m.
) sont bien plus précises que celles de
(1788), et, la première a une localité-type littorale et l'autre
bathyale. Néanmoins, la distribution bathymétrique de N. anomala s'étend depuis quelques mètres jusqu'à 1478 m.
et al. (2008) signalent N. turbinata dans le Miocène supérieur/Pliocène inférieur aux Açores, d'après de seules valves dorsales, de même pour la signalisation d'individus vivants aux îles Chafarines par & de (2013). Dans la liste des espèces présentes en Nouvelle-Zélande, et al. (2009) puis (2011) mentionnent Novocrania turbinata, espèce dite cosmopolite, depuis l'Éocène inférieur jusqu'à nos jours. Déjà & (2001) avaient proposé une certaine similitude d'exemplaires du Pacifique avec N. turbinata. Si une aussi grande longivité, quelque 50 MA, pour une espèce n'est pas exceptionnelle, elle doit être assise sur une étude approfondie des caractères taxinomiques et de leurs variations à la fois géographiques et temporelles, tout en démontrant le caractère cosmopolite dans toutes ses acceptations.
(2014) a décrit les muscles chez six espèces de craniidés dont Novocrania anomala and N. turbinata : aucune différence n'a été notée, sauf "two pedestals and a median spike that on three sides hang above the ventral mound" chez N. turbinata et N. japonica qui est la seule à être illustrée. Il s'agit probablement du résultat d'une calcification de la valve.
En se basant sur des analyses moléculaires, et al. (2014) arrivent à la conclusion que des exemplaires identifiés par & (2001) comme appartenant à N. turbinata font partie du clade N. anomala qui s'étend dans le NE Atlantique et dans le bassin méditerranéen occidental jusqu'en mer Adriatique : il y a donc extension de ce clade dans le bassin oriental dont cette mer est partie intégrante. Rappelons que la localité-type de turbinata est située dans le bassin occidental, voire en mer Adriatique. Ceci répond à l'interrogation des deux derniers auteurs indiquant "this will need to be confirmed by molecular biology studies." Le fait de ne pas corroborer les assertions de (1988) et de & (2001) peut permettre à et al. (2014) de rejeter la "traditional morphology", mais sans la définir : nous l'assimilerons à la systématique phénétique, mais sûrement pas aux caractères phylogénétiques résultant d'une analyse cladistique, qui ne peuvent être restreints à la seule coquille. D'ailleurs, les caractères phylogénétiques morpho-anatomiques sont le résultat d'expressions génétiques que les molécularistes ne savent pas encore attribuer à des gènes identifiés. Et il convient de ne pas oublier que les caractères écologiques ont toute leur place dans une telle analyse. Le rejet de la systématique classique par les molécularistes a été récemment fustigé par l'académicienne et embryologiste Nicole (2012).
On peut donc conclure qu'une seule espèce N. anomala est présente dans le bassin occidental et au moins partiellement dans le bassin oriental de la mer Méditerranée et qu'en conséquence les exemplaires ayant servi à la description originale de (1795) sous turbinata sont à mettre en synonymie avec N. anomala. En cela, et al. (2014) ne font que corroborer ce que des auteurs avaient déjà avant eux publié au cours de plus d'un siècle.
La proposition d'un clade téthysien ( et al.,
2014) selon le schéma Méditerranée orientale > Cap Vert > Caraïbes est une curiosité qui soulève plusieurs points intéressants. Concernant le bassin oriental de la Méditerranée, les explications données sur son histoire ne correspondent plus aux connaissances actuelles (voir & ,
2004) : en effet, avec la fermeture du passage Indo-Pacifique-Atlantique au Burdigalien (Miocène), le bassin oriental ne communique plus qu'avec le bassin occidental de la mer Méditerranée ; or, depuis cette époque, ses caractéristiques physiques et biologiques, soumises aux variations des époques glaciaires, rendent toute communication difficile à travers le détroit siculo-tunisien (Fig. 6 ![]() ). Alors, que représente ces Novocrania dont la distribution est relativement isolée dans la mer du Levant ? Ce clade téthysien met aussi en exergue l'importance, pourtant décriée, de la morphologie, quand et al.
(2014) mentionnent "members of this molecular clade show considerable diversity of form" : cette hétérogénéité morphologique et géographique est-elle compatible avec l'expression génétique de ce clade ? Le débat est ouvert, mais les analyses moléculaires ne sauraient résoudre le problème sans une excellente étude morpho-anatomique complémentaire.
). Alors, que représente ces Novocrania dont la distribution est relativement isolée dans la mer du Levant ? Ce clade téthysien met aussi en exergue l'importance, pourtant décriée, de la morphologie, quand et al.
(2014) mentionnent "members of this molecular clade show considerable diversity of form" : cette hétérogénéité morphologique et géographique est-elle compatible avec l'expression génétique de ce clade ? Le débat est ouvert, mais les analyses moléculaires ne sauraient résoudre le problème sans une excellente étude morpho-anatomique complémentaire.
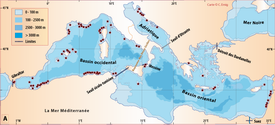
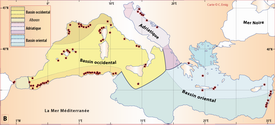
Click on thumbnail to enlarge the image.
Figure 6 : Répartition de Novocrania en mer Méditerranée.- A. En fonction de la géographie et de la bathymétrie, avec les principales limites dans les échanges des masses d'eaux, qui ont évolué depuis le Miocène. B. en fonction des caractéristiques faunistiques (modifiée, d'après ,
1970). La circulation des masses d'eau et les variations du niveau de la mer au cours des glaciations sont des facteurs primordiaux dans l'évolution des faunes méditerranéennes ( & ,
2004). Actuellement, ce genre ne comprend probablement qu'une seule espèce dans cette mer : N. anomala qui est citée
par la majorité des auteurs, quelques-uns citant N. turbinata, la plupart avec N. anomala. En outre, une référence à des animaux vivants ou à des coquille vides n'est pas toujours clairement indiquée ; dans le deuxième cas, l'âge des valves vides peut atteindre plusieurs centaines de milliers d'années.
Pour établir cette carte,
de nombreux travaux ont été compulsés
(suit une liste non exhaustive : ,
1836 ; ,
1886 ;
, 1896 ;
, 1914 ;
, 1915 ; & ,
1941 ; ,
1962 ; ,
1964 ;
, 1967 ; ,
1971 ;
& ,
1986 ; & ,
1987 ; , 1988 ; ,
1989 ; et
al., 1991 ; ,
1994b ; & ,
1995 ; ,
1996b ; et al.,
1999 ;
et al., 2000 ; ,
2003, sous presse ; ,
2003 ; & ,
2003 ;
et al., 2006 ; et al.,
2006 ;
et al., 2009 ;
et al., 2011 ;
et al., 2014 ;
...).
Figure 6: Distribution of Novocrania in the Mediterranean Sea.- A.
Controled by geography, bathymetry, and the main boundaries of water
mass exchange, which have evolved since the Miocene. B. According to faunal characteristics (modified from , 1970). The circulation of water masses and changes in sea level during glaciations are key factors in the evolution of the Mediterranean fauna ( & , 2004).
At present, probably only one species of this genus occurs in the Mediterranean Sea.
This is N. anomala, which has been identifed by almost all authors.
Some cite N. turbinata occurring with N. anomala.
Information on the shells, living or empty, is rarely indicated; the age of empty valves may reach several hundreds of thousands of
years.
Many
studies were looked through to build this map (a non-exhaustive list
follows: , 1836; ,
1886;
, 1896;
, 1914;
, 1915; & ,
1941; ,
1962; ,
1964;
, 1967; ,
1971;
& ,
1986; & ,
1987; , 1988; ,
1989; et
al., 1991; ,
1994b; & ,
1995; ,
1996b; et al.,
1999;
et al., 2000; ,
2003, in press; ,
2003; & ,
2003;
et al., 2006; et al.,
2006;
et al., 2009;
et al., 2011;
et al., 2014; ...).
En l'état actuel de nos connaissances et des données apportées par les divers auteurs et discutées ci-dessus, nous considérons Novocrania turbinata comme synonyme de N. anomala. La distribution de cette espèce en mer Méditerranée (Fig. 6 ![]() ) ne peut s'expliquer qu'au travers des bouleversements que cette mer a subi au cours de sa récente formation depuis le Burdigalien (Miocène). Selon des données dans d'autres groupes zoologiques, une colonisation depuis la Téthys vers le futur océan Atlantique, voire vers ceux Indien et Pacifique, avant la fermeture définitive du bassin oriental n'est pas à exclure. En mer Méditerranée, la distribution bathymétrique de N. anomala doit aussi prendre en compte les fortes variations du niveau de la mer au cours des glaciations successives, notamment la dernière, celle du Würm, il y a environ 40 kA, ainsi que la disparition de la faune chaude au Sicilien (Pléistocène), puis les remplacements répétés de faunes tempérées et froides jusqu'à nos jours (voir & ,
2004). La circulation anti-cyclonique des masses d'eau dans les bassins méditerranéens donne le sens d'une possible colonisation.
) ne peut s'expliquer qu'au travers des bouleversements que cette mer a subi au cours de sa récente formation depuis le Burdigalien (Miocène). Selon des données dans d'autres groupes zoologiques, une colonisation depuis la Téthys vers le futur océan Atlantique, voire vers ceux Indien et Pacifique, avant la fermeture définitive du bassin oriental n'est pas à exclure. En mer Méditerranée, la distribution bathymétrique de N. anomala doit aussi prendre en compte les fortes variations du niveau de la mer au cours des glaciations successives, notamment la dernière, celle du Würm, il y a environ 40 kA, ainsi que la disparition de la faune chaude au Sicilien (Pléistocène), puis les remplacements répétés de faunes tempérées et froides jusqu'à nos jours (voir & ,
2004). La circulation anti-cyclonique des masses d'eau dans les bassins méditerranéens donne le sens d'une possible colonisation.
Au niveau de Gibraltar, les caractéristiques physico-chimiques des masses d'eau, d'une part, et les inversions du courant entrant lors des périodes glaciaires entre l'océan Atlantique et la mer Méditerranée, d'autre part, n'ont guère favorisé de larges échanges ; la mer d'Alboran a été la seule région colonisée depuis l'Atlantique, d'où son particularisme faunistique (Fig. 6B ![]() ).
).
La lecture approfondie des travaux antérieurs, dont les originaux, nous est apparue nécessaire pour pouvoir progresser dans la connaissance de la biodiversité des brachiopodes. Le respect du Code International de Nomenclature Zoologique (1999) permet de donner à une espèce une diagnose "c'est-à-dire un résumé des caractères* qui différencient ce taxon nominal des taxons comparables" (* il faudrait préciser de tous les caractères) ; il y a parfois confusion entre les termes diagnose et description, qui sont pourtant bien distincts. Concernant les brachiopodes actuels, les caractères taxinomiques ne peuvent évidemment pas être restreints à la seule coquille, alors que les parties dites molles du corps et l'écologie peuvent et doivent apporter leur contribution pour établir la diagnose et ainsi faciliter une identification. Ce n'est pas parce que cela est difficile, voire impossible, sur les formes fossiles qu'il faut s'en priver aujourd'hui dans les formes actuelles.

Click on each thumbnail to enlarge the corresponding image.
Planche 1 :
Récoltes de branches du scléractiniaire Dendrophyllia cornigera, avec de nombreux brachiopodes, Novocrania anomala, Megerlia truncata (dont la forme appelée antérieurement Pantellaria/Megerlia monstruosa), de rares Platidia anomioides. Sur ces branches on trouve aussi quelques Terebratulina retusa et Megathiris detruncata (espèce cryptique), non photographiées ici. En C, une Megerlia fixée sur une Novocrania. Provenance : st. BM 30 : 139-147 m, 42°41.67'N - 8°48.11'E (N/O Catherine-Laurence - Nord de la baie de Calvi, W Corse) ; st. A36 : 175-180 m, 42°58'36"N - 5°36'24"E ; st. A 91 : 195-200 m, 42°59.10'N - 5°33.40'E (N/O Antedon - Sud de La Ciotat, Banc des Blauquières, Provence).
Plate 1:
Set of branches of the stony coral Dendrophyllia cornigera with many brachiopods,
i.e., Novocrania anomala, Megerlia truncata (and occurrence of the form formerly called Pantellaria/Megerlia monstruosa), rare Platidia anomioides. Terebratulina retusa and Megathiris detruncata
(a cryptic species) are not pictured here. On fig. C, Megerlia fixed on Novocrania. Sources: st. BM 30: 139-147 m, 42°41.67'N - 8°48.11'E (R/V Catherine-Laurence - N off Bay of Calvi, W Corsica); st. A36: 175-180 m, 42°58'36"N - 5°36'24"E; st. A 91: 195-200 m, 42°59.1'N - 5°33.40'E (R/V Antedon - South of La Ciotat, Blauquières
Bank, Provence).
Je voudrais souligner le travail réalisé par les divers organismes publics et privés en rendant accessible en ligne la grande majorité des publications consultées pour rédiger cette note [notamment Gallica-Bibliothèque Nationale de France ; Google Books ; BHL-Biodiversity Heritage Library, USA ; Animal base-Universitätsbibliothek Göttingen]. Mes chaleureux remerciements aux deux lecteurs-arbitres et néanmoins amis, Fernando (Université d'Oviedo, Espagne) et Aleksandra (Institut de Paléobiologie, Varsovie, Pologne), pour leurs critiques, corrections et commentaires.
F. & C.C. (2005).- Filo Brachiopoda. In: Fauna Ibérica.- Museo de Ciencias Naturales, CSIC Madrid, vol. 27, p. 57-177, 188-207, 215-239 & 255-276.
A. & C.C. (1995).- Lophophorata. In : Check list delle specie della fauna italiana.- Calderini, Bologne, vol. 108, p. 10-23.
P. (1987).- Stratigraphie des dépôts pliocènes et quaternaires du Détroit de Messine.- Documents et Travaux de l'I.G.A.L., Cergy-Pontoise, vol. 11, p. 59-81.
L., E. & D. (2009).- Brachiopodi neogenici e pleistocenici dell'Emilia (Parte seconda).- Parva Naturalia, Plaisance, vol. 8, p. 3-42.
M.A., P. & B. (2013).- Upper Oligocene (Chattian) brachiopod fauna from the Aquitaine Basin, southwestern France and its paleoenvironmental implications.- Geodiversitas, Paris, vol. 35, p. 579-606.
H.-M. de (1818).- Cranie, Crania. In : Dictionnaire des Sciences naturelles.- Levrault, Strasbourg, vol. 11, p. 312.
E. (2001).- Osservazioni sui brachiopodi neogenici e pleistocenici dell'Emilia-Romagna.- Parva Naturalia, Plaisance, 2000-2001, p. 45-81.
C.H.C. (1988).- Some brachiopods from the Eastern Mediterranean Sea.- Israel Journal of Zoology, Londres, vol. 35, p. 151-169.
C. (1970).- Écologie de l'étage bathyal dans la Méditerranée occidentale.- Mémoires de l'Institut océanographique de Monaco, vol. 2, 146 p.
J.P. (1967).- Les sédiments meubles à brachiopodes de la marge continentale algérienne.- Bulletin du Muséum National d'Histoire naturelle de Paris, (2e Série), vol. 39, nº 4, p. 779-792.
S., C.N., C. & A. (2000).- First survey of sessile communities on subtidal rocks in an area with hydrothermal vents: Milos Island, Aegean Sea.- Hydrobiologia, Gehnt, vol. 426, nº 1, p. 113–121.
B.L., A. & C. (2014).- Craniid brachiopods: aspects of clade structure and distribution reflect continental drift (Brachiopoda: Craniiformea). Zoological Journal of the Linnean Society, Londres, vol. 171, nº 1, p. 133-150.
Commission Internationale de Nomenclature Zoologique (1999).- Code International de Nomenclature Zoologique.- International Trust for Zoological Nomenclature, Londres, 4e édition, 306 p.
J., C.C. & M. (1991).- Els lofoforats i afins. In : Invertebrats no artròpodes.- Història Natural dels Països Catalans, Enciclopèdia Catalana, Barcelone, vol. 8, p. 335-374 [en catalan].
E.M. da (1770).- Conchology, or, natural history of shells.- T. Jones, Londres, 26 p. (12 Pls.).
G. (1798).- Tableau élémentaire de l'histoire naturelle des animaux.- Baudouin, Paris, 710 p. (Pls. I-XIV).
G. (1834).- The animal kingdom.- Whittaker, Londres, vol. 12, 601 p. [Orbicula p. 133].
W.H. (1871).- Report on the Brachiopoda obtained by the United States Coast Survey Expedition in charge of L.F. Pourtales, with a revision of the Craniidæ and Discinidæ.- Bulletin of the Museum of comparative Zoology, Cambridge (MA), vol. 3, nº 1, p. 1-45 (2 Pls.).
W.H. (1921).- Annotated list of the Recent Brachiopoda in the Collection of the United States National Museum, with descriptions of thirty-three new forms.- Proceedings of the U.S. National Museum, Washington, vol. 57, nº 2314, p. 261-377.
T. (1888).- A monograph of the Recent Brachiopoda. Part III.- Transactions of the Linnean Society of London, (serie 2), vol. 4 (Zool.), p. 183-248 (Pls. 26-30).
E. (1818).- Cranie (). In : Dictionnaire des Sciences Naturelles.- Levrault, Strasbourg, vol. 11, p. 312-314.
G.P. & H. (1836).- Obicules (Orbicula). In : Histoire naturelle des animaux sans vertèbres : présentant les caractères généraux et particuliers de ces animaux, leur distribution, leurs classes, leurs familles, leurs genres, et la citation des principales espèces qui s'y rapportent.- Baillière, Paris, 2e édition, tome 7, p. 313-319.
I. (1979).- Il Pleistocene in facies batiale di Valle Palione (Grammichele, Catania).- Bollettino Malacologico, Prato, vol. 15, nº 5-6, p. 85-156.
I. & G. (1987).- Les fonds à Errina aspera et Pachylasma giganteum.- Documents et Travaux de l'IGAL, Cergy-Pontoise, vol. 11, p. 243-247.
L.W. (1817).- A descriptive catalogue of recent shells, arranged according to the Linnæan method; with particular attention to the synonymy.- J. and A. Arch, Londres, vol. I, 580 p. [Anomia turbinata p. 286].
C.C. (1987).- Offshore brachiopods investigated by submersible.- Journal of experimental marine Biology and Ecology, vol. 108, Millport, p. 261-273.
C.C. (1988a).- Document technique nº 1 - A + B - Campagnes en mer "BRACORS 1 - 6" (1983-1987).- CNRS-RCP 728, 13 p. En ligne sur : http://paleopolis.rediris.es/Phoronida/EMIG/RCP-728/BRACors-RCP728.pdf
C.C. (1988b).- Document technique nº 2 - A + B - Campagnes en mer "BRAPROV 0 - 9" (1983-1987).- CNRS-RCP 728, 13 p. En ligne sur : http://paleopolis.rediris.es/Phoronida/EMIG/RCP-728/BRAProv-RCP728.pdf
C.C. (1997).- Bathyal zones on the Mediterranean continental slope: An attempt.- Publicaciones especiales del Instituto Español de Oceanografía, Madrid, vol. 23, p. 23-33.
C.C. (2002).- Tools for linguloid taxonomy: the genus Obolus (Brachiopoda) as an example.- Carnets de Géologie [Notebooks on Geology], Maintenon, Article 2002/01 (CG2002_A01), 9 p.
C.C. (2003).- Brachiopoda, Phoronida. In : Checklist of the species of the Italian fauna. En ligne sur : http://www.faunaitalia.it/checklist/introduction.html (dernière consultation le 06/05/2014).
C.C. (2009).- Nummulus brattenburgensis and Crania craniolaris (Brachiopoda, Craniidae).- Carnets de Géologie [Notebooks on Geology], Brest, Article 2009/08 (CG2009_A08), 11 p.
C.C. (2010).- Brachiopoda.- Biologie marina Mediterranea, Gênes, vol. 17 (suppl. 1), p. 586-588.
C.C. (2012).- Révision des espèces de brachiopodes décrits par A. .- Carnets de Géologie [Notebooks on Geology], Brest, Article 2012/02 (CG2012_A02), p. 15-30.
C.C. (sous presse).- Les Brachiopodes des côtes françaises métropolitaines.- Bulletin de la Société des Sciences naturelles de l'Ouest de la France, Nantes, hors-série.
C.C., F. & M.A. (2014).- World Brachiopoda database. En ligne sur : http://www.marinespecies.org/brachiopoda (dernière consultation le 06/05/2014).
C.C. & P. (2004).- Faune profonde en Mer Méditerranée : les échanges historiques, géographiques et bathymétriques.- The Mediterranean deep-sea fauna: historical evolution, bathymetric variations and geographical changes.- Carnets de Géologie [Notebooks on Geology], Maintenon, Article 2004/01 (CG2004_A01), 10 p.
F., P.G. & B. (2011).- Recent Brachiopoda of the marine protected area "Secche di Tor Paterno", Central Tyrrhenian Sea.- Cahiers de Biologie Marine, Roscoff, vol. 5, nº 2, p. 193-202.
P. & D.-P. (1891).- Sur la répartition stratigraphique des Brachiopodes de mer profonde, recueillis durant les expéditions du Travailleur et du Talisman.- Compte-Rendu de l'Académie des Sciences de Paris, vol. 111, p. 247-249.
J. (1822).- The philosophy of zoology, or, A general view of the structure, functions, and classification of animals.- A. Constable & Co., Edimbourg, vol. 2, 618 p.
D. & G. (2007).- Perspectivas de protección y conservación de los yacimientos paleontológicos de braquiópodos pliocenos de Murcia.- XXIIIa Jornadas de Paleontología (Caravaca de la Cruz, Murcia, 2007), Resumen, p. 80-81.
L. (1971).- Braquiópodos de Baleares.- Boletin de la Sociedad de Historia natural de Baleares, Palma de Majorque, vol. 16, p. 95-104.
F.W. (1828).- Beitrag zur Monographie der Gattung Crania.- Schüller, Krefeld, 12 p. (1 Pl.).
J.G. (1915).- Datos para la fauna española (Moluscos y Braquiópodos).- Boletin de la Real Sociedad española de Historia natural, Madrid, vol. 16, p. 235-246.
N. (2011).- Micromorphic brachiopods from the Early Miocene (Otaian) of Northland, New Zealand.- New Zealand Journal of Geology and Geophysics, Wellington, vol. 54, p. 75-87.
International Commission on Zoological Nomenclature (1999).- Code International de Nomenclature Zoologique.- The International Trust for Zoological Nomenclature, Londres, 4e édition, p. 127-306.
J.G. (1878).- On the Mollusca procured during the "Lightning" and "Porcupine" expeditions 1868-1870. Part 1.- Proceedings of the zoological Society of London, vol. 1878, p. 393-416 (Pls. XXII-XXIII).
L. (1885).- Recherches sur l'anatomie des brachiopodes inarticulés.- Thèse Dr. es-Sciences, Faculté des Sciences de Paris, 143 p.
L. (1886).- Recherches sur l'anatomie des brachiopodes inarticulés.- Archives de Zoologie expérimentale et générale, Paris, (série 2), vol. 4, p. 161-303.
E. (2007).- Pliocene brachiopods from Rhodes Island (Cape Vagia section).- Bulletin of the Geological Society of Greece, Byronas, vol. 40, nº 1, 13 p.
A., M.A. & S.P. (2008).- Novocrania turbinata (Brachiopoda) from the Early Pliocene of the Azores (Portugal).- Acta Geologica Polonica, Varsovie, vol. 58, nº 4, p. 473-478.
M., A., J. & M. (2005).- Distribution and habitats of Acesta excavata (Bivalvia: Limidae) with new data on its shell ultrastructure. In : A. & J.M. (eds.), Cold-water corals and ecosystems.- Springer-Verlag, Berlin, p. 173-205.
J.P. [P.A. de de] (1819).- Histoire naturelle des animaux sans vertèbres, présentant les caractères généraux et parti- culiers de ces animaux, leur distribution, leurs classes, leurs familles, leurs genres, et la citation des principales espèces qui s'y rapportent; précédée d'une introduction offrant la détermination des caractères essentiels de l'animal, sa distinction du végétal et des autres corps naturels, enfin, l'exposition des principes fondamentaux de la zoologie.- Déterville, Paris, vol. 6 (1e partie), 343 p. [Cranie, p. 237-240].
E. & M. (1941).- Notes sur les espèces lamarckiennes de brachiopodes vivants.- Journal de Conchyologie, Paris, vol. 84, nº 2, p. 183-196 ; nº 3, p. 279-299.
N. (2012).- Dans le secret des êtres vivants - Itinéraire d'une biologiste.- Laffont, Paris, 487 p.
W.E. (1852).- Genus 1. Criopus. In : Molluscorum Britanniæ synopsis: A synopsis of the Mollusca of Great Britain.- Van Voorst, Londres, p. 358.
D.E. & C.H.C. (1986).- Neocrania n. gen., and a revision of Cretaceous-Recent brachiopod genera in the Family Craniidae.- Bulletin of the British Museum of natural History (Geology), Londres, vol. 40, nº 4, p. 141-160.
D.E. & C.H.C. (2001).- Novocrania, new name for the genus Neocrania and , 1986 (Brachiopoda, Craniida); preoccupied by Neocrania , 1978 (Insecta, Lepidoptera).- Bulletin of the British Museum of natural History (Geology), Londres, vol. 57, nº 1, p. 5.
C. (1988).- Braquiópodos del Banco de Chella (Mar de Alborán, Mediterráneo Occidental).- Acta geológica hispánica, Barcelone, vol. 23, p. 311-319.
A. (1899).- Les coquilles marines au large des côtes de France : faune pélagique et faune abyssale : description des familles, genres et espèces.- Baillière, Paris, 198 p. [Brachiopoda p. 181-186].
A. (1979).- The Recent Brachiopoda of the Mediterranean Sea.- Bulletin de l'Institut océanographique de Monaco, vol. 72, nº 1434, p. 1-112.
A. (2003).- Marine fauna of the Mljet National Park (Adriatic Sea, Croatia). 3. Brachiopoda.- Natura Croatica, Zagreb, vol. 12, nº 4, p. 233-243.
A., C. N., C., H. & G. (2002).- New records of Recent brachiopods from the eastern Mediterranean Sea.- Annali del Museo civico di Storia naturale "G. Doria", Gênes, vol. 94, p. 407-418.
A., C.N., i C. & H. (2004).- The present-day Mediterranean brachiopod fauna: diversity, life habits, biogeography and paleobiogeography.- Scientia marina, Barcelone, vol. 68 (suppl. 1), p. 163-170.
A. & S.L. (2001).- Shell morphology and geographical distribution of Neocrania (Brachiopoda, Recent) in the eastern North Atlantic and Mediterranean Sea.- Systematics Association Special Volume Series, Londres, vol. 63, p. 71-79.
A., P. & F. (2009).- New record of Novocrania (Brachiopoda, Craniida) from Madeira, with notes on Recent brachiopod occurrences in the Macaronesian archipelagos.- Arquipelago - Life and Marine Sciences, Ponte Delgada, vol. 24, p. 17-22.
S. (1845).- Index molluscorum litora Scandinaviae occidentalia habitantium.- Konglida Vetenskaps-Akademiens Förhandlinger, Stockholm, vol. 1, p. 135-204.
D.A.B., M.A., H.J., R.A., D.E., D.I., J.B., A.J. & J.H. (2009).- Brachiopoda. In: D. (ed.), New Zealand inventory of biodiversity, Kingdom Animalia, Radiata, Lophotrochozoa, and Deuterostomia.- New Zealand Inventory of Biodiversity, Christchurch, vol. 1, p. 255-267.
C. (1962).- Études des fonds chalutables de la Méditerranée occidentale (Écologie et pêche). Résultats des campagnes des navires océanographiques "Président-Théodore-Tissier" 1957 à 1960 et "Thalassa" 1960 et 1961.- Revue des Travaux de l'Institut des Pêches marititimes, Nantes, vol. 26, nº 2, p. 163-218.
G. (1815).- XIII. An account of some new and rare marine British shells and animals.- Transactions of the Linnean Society of London, vol. XI, p. 179-204 [Patella distorta, p. 195-196, Pl. XIII, fig. 5].
C., C.N., S., A., A.M., S., M., M., F., M. & P. (1999).- Biodiversity of marine sessile epifauna at an Aegean island subject to hydrothermal activity: Milos, eastern Mediterranean Sea.- Marine Biology, Kiel, vol. 135, p. 729-739.
O.F. (1776).- Zoologiae Danicae Prodromus, seu Animalium Daniae et Norvegiae indigenarum chracteres, nomina, et synonyma imprimis popularium.- Typis Hallagerii, Havniae (Copenhague), 282 p. [Patella anomala nº 2870, p. 237 ; Anomia aculeata nº 3005, p. 249]. En ligne sur :
http://books.google.fr/books?id=LXSp6LuBwu0C&hl=fr&pg=PP1#v=onepage&q&f=false (dernière consultation le 06/05/2014).
O.F. (1788).- Zoologia Danica, seu animalium Daniae et Norvegiae rariorum ac minus notorum descriptiones et historia.- Weygand, Copenhague & Leipzig, 1e édition, vol. 1, 606 p. [Patella anomala, p. 4, pl. V]. (*)
D. (1977).- Les brachiopodes du Pliocène I de la région d'Ahuilas (Sud d'Almeria, Espagne).- Annales de Paléontologie (Invertébrés), Paris, vol. 63, nº 1, p. 59-75.
P. (1914).- Liste des mollusques du Golfe de Tunis.- Bulletin de la Société d'Histoire naturelle de l'Afrique du Nord, Alger, tome 5, nº 1, p. 12-27.
G.S. (1795a).- Praefatio. In : Testacea utriusque Siciliae eorumque historia et anatome. Tabulis aeneis illustrata.- Parme, tome 1, p. i-x.
G.S. (1795b).- Testacea utriusque Siciliae eorumque historia et anatome. Tabulis aeneis illustrata.- Parme, tome 2, 356 p. ; tome 4, Pl. XXX. En ligne sur : https://archive.org/details/TestaceautriusqIIPoli (dernière consultation le 06/05/2014).
L. (1862).- Monography of the genus Orbicula. Monography of the genus Crania.- Conchologia Iconica, Londres, vol. 13, p. 67-71 (Pl. 1).
D. (1964).- Contribution à l'étude du Rech Lacaze-Duthiers, vallée sous-marine des côtes du Roussillon.- Vie et Milieu, Banyuls, vol. 15, nº 1, p. 1-46.
J. (2014).- The muscles, body wall and valve-opening mechanism of extant craniid (inarticulated) brachiopods.- Journal of natural History, Londres, p. 1-22.
L. & de J.M. (2013).- First record of Novocrania turbinata (, 1795) (Brachiopoda, Craniata, Craniidae) from the Chafarinas Islands.- Zoologica Baetica, Grenade, vol. 24, p. 189-192.
J.I. (1989).- Verzeichnis der rezenten Brachiopoden (Brachiopoda) von den iberischen Küsten und den angrenzenden Meeren.- Bonner zoologische Beiträge, vol . 40, nº 2, p. 141-154.
A. (1836).- Catalogus Conchyliorum regni neapolitani quae usque adhuc reperit.- Sereti, Naples, 19 p. (1 Pl.).
C.F. (1817).- Orbicule, Orbicula. In : Essai d'un nouveau système des habitations des vers testacés.- Schultz, Copenhague, p. 176-177 (Pl. 21).
G. (1865).- Paleontologia malacologia delle rocce terziarie del distretto di Messina: Classe Brachiopodi.- Memoria della Società Italiana di Scienze naturali, Milan, vol. 1, p. 5-88.
M., S. & E. (2006).- Reconstructing palaeoenvironments in a Pliocene–Pleistocene Mediterranean microbasin.- Facies, Erlangen, vol. 52, p. 361-380.
R. (1896).- Zoologische Ergebnisse. VIII. Brachiopoden, gesammelt auf den Expeditionen S.M. Schiffes "Pola" 1890-1894.- Denkschriften der Kaiserliche Akademie der Wissenschaften in Wien, (Mathematisch-Naturwissenschaftliche Klasse), vol. 63, p. 37-38.
E. (1983).- Paleoecologia e biostratigrafia delle calcareniti a brachiopodi di Castro (Lecce).- Bolletin della Società dei Naturalisti in Napoli, vol. 92, p. 347-413.
E. (1988).- Analisi paleoecologica di un affioramiento di Calcareniti di Gravina.- Atti del 4º Simposio di Ecologia e Palaeoecologia delle Comunità Bentoniche, Sorrente, p. 443-454.
E. (1994a).- Neogeno Salento brachiopod palaeocommunities.- Bolletino della Società paleontologica italiana, Modène, vol. 33, nº 2, p. 197-213.
E. (1994b).- Brachiopods from bio- and thanatocoenoses of the Isca submarine cave (Sorrento Penisula).- In: R. et al. (eds), Studies on ecology and paleoecology of benthic communities.- Bolletino della Società paleontologica italiana, Modène, special vol. 2, p. 313-323.
E. (1996a).- Biostratigrafia e paleoecologie delle Calcarentiti di Gravina nei dintorni di Cerignola (brachiopodi e foraminiferi).- Memorie della Società Geologica Italiana, Rome, vol. 51, p. 197-207.
E. (1996b).- Notes on living brachiopod ecology in a submarine cave off the Campania coast, Italy.- Proceedings of the 3rd international brachiopod congress (Sudbury, 1995), Balkema, Rotterdam p. 227-231.
E., G. & P. (2006).- Bioerosion on brachiopod shells of a thanatocoenosis of Alboran Sea (Spain).- Ichnos, Philadelphia, vol. 13, nº 3, p. 175-184.
E. & R. (2006).- I brachiopodi delle calcareniti di Castro Marina (Lecce).- Thalassia Salentina, Porto Cesareo, vol. 29, suppl., p. 301-310.
J., M., D., A., F., R., J., C.M. & M. (2006).- Flora y fauna de la reserva marina y reserva de pesca de la Isla de Alborán.- Secretaría General de Pesca Marítima, Ministerio de Agricultura, Pesca y Alimentación, Madrid, 269 p.
J. & A.A. (1986).- Braquiopodos de los fondos de Corallium rubrum () proximos a la Isla de Alboran (S.E. de España).- Boletín del Instituto Español de Oceanografía, Madrid, vol. 3, nº 4, p. 111-114.
W. (1819).- A conchological dictionary of the British Islands.- Booth, Londres, 272 p.
P. & H. (2003).- Mediterranean and Atlantic Invertebrate Guide.- Conchbooks, Harxheim, 305 p.
(*) C'est cette édition que cite (1795). Cependant, il y a eu une édition en 1779 dans laquelle Patella anomala est en p. 9 et dont je n'ai pu trouver les planches.