◄ Carnets Geol. 14 (10) ►
![]()
Contents
[Introduction] [Geological setting and fossil association] [Stratigraphy and palaeoecology]
[Description] [Discussion] [Conclusions]
and ... [Bibliographic references]
Charles University, Institute of Geology and Palaeontology,
Albertov 6, CZ128 43, Prague 2 (Czech Republic)
K Moravině 11/1689, CZ190 00, Prague Vysočany (Czech Republic)
Published online in final form (pdf) on July 3, 2014
[Editor: Bruno ]
![]()
An enrolled exoskeleton of the holaspid specimen of a tiny agnostid Peronopsis integra (, 1845) entombed inside a conch of the hyolithid ?Buchavalites sp. is described from the middle Cambrian (Drumian) Jince Formation of the Příbram-Jince Basin (Czech Republic). The agnostid is associated with an ichnofossil of the feeding trace classified as Arachnostega-type behaviour. The enrolled attitude of the agnostid exoskeleton suggests that the specimen is a carcass rather than moult. Either the storm disturbance and/or well-protected source of food hypotheses could explain the entombed agnostid. This additional example supports a benthic mode of life in the agnostid P. integra. The studied association of feeding tunnels of an unknown Arachnostega-strategist and Peronopsis preserved inside a hyolithid conch is a case of "frozen" behaviour.
Agnostid palaeoecology; trilobites; Drumian; Cambrian; Jince Formation; Barrandian area; Příbram-Jince Basin; Czech Republic.
O. & V. (2014).- A new type of entombment of Peronopsis (Agnostida) in a hyolithid conch.- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 10, p. 191-198.
Un nouveau type d'enfouissement pour Peronopsis (Agnostida) dans une coquille d'hyolithide.- Un exosquelette recroquevillé d'un spécimen holaspide d'un petit trilobite agnostide, Peronopsis integra (, 1845), a été observé enfoui à l'intérieur de la coquille de l'hyolithide ?Buchavalites sp. provenant de la Formation de Jince attribuée au Cambrien moyen (Drumien) du Bassin de Příbram-Jince (République tchèque). L'agnostide est associé à une trace fossile en lien avec une activité d'alimentation, attribuée à un comportement de type Arachnostega. La disposition recroquevillée de l'exosquelette de l'agnostide suggère que ce spécimen est une carcasse plutôt qu'une mue. L'hypothèse d'une perturbation causée par une tempête et/ou celle d'une source de nourriture bien abritée permettraient d'expliquer l'agnostide enfoui. Ce nouvel exemple confirme un mode de vie benthique chez l'agnostide P. integra. L'association étudiée de terriers alimentaires d'un stratège inconnu de type Arachnostega et d'un Peronopsis préservé à l'intérieur de la coquille d'un hyolithide est un exemple d'action figée.
Paléoécologie des agnostides ; trilobites ; Drumien ; Cambrien ; Formation de Jince ; Aire Barrandienne ; Bassin de Příbram-Jince ; République tchèque.
The conchicolous habit (= "the use by other animals of shells as residences after the original builders have died" following , 1987, p. 240) is very rare in the fossil record. Similarly rare is also inquinilism of invertebrates (e.g., & , 1995; & , 2010; & , 2012). Possible early Palaeozoic examples have been reported from the Cambrian, Ordovician and Silurian of Laurentia (U.S.A.; e.g., , 1993; et al., 2001), Baltica (Sweden; e.g., & , 2012) and West Gondwana (Czech Republic and Portugal; e.g., et al., 2008a; et al., 2009; & , 2014).
The oldest fossils documenting conchicolous habit of an unknown organism were announced by and (2006) from the Early Cambrian of California; somewhat younger examples of entombed agnostids and eodiscid trilobites were described from the middle Cambrian of the Burgess Shale, British Columbia ( et al., 2003) and from the Jince Formation of the Czech Republic ( et al., 2009; & , 2011). Other example of a possible conchicolous habit of trilobites preserved inside of large hyolithids was discussed by et al. (2008a) and et al. (2008) based on fossils from the Buchava Formation (Drumian). Clusters of trilobites, preserved prone or partly enrolled, have been described under disarticulated skeletal elements of large trilobites were described from Middle Ordovician sediments of northern Portugal ( et al., 2009) and the Czech Republic ( & , in press). Diverse trilobites entombed inside cephalopod shells are known from the Middle Ordovician of the Czech Republic (, , unpublished observation), from the Upper Ordovician of the U.S.A. (e.g., et al., 2001), as well as from the Silurian of the U.S.A., the Czech Republic (e.g., et al., 2001; et al., unpublished observation) and also from Sweden ( & , 2012). (1993, Fig. 3.6) figured an enrolled trilobite preserved inside a gastropod body chamber from the Upper Ordovician of the U.S.A.
and (2006) suppose that the hyolith conchs were used as shelters and interpreted these finds as possible evidence of a strategy to avoid predation. et al. (2003), et al. (2008), et al. (2009), et al. (2009) and & (2011) speculated that agnostids and some trilobites might have entered diverse empty tubes and other restricted spaces for a specific purpose, most likely to feed (scavenge on remnants of a dead priapulid worm, hyolithid or large trilobite) or to hide. Other possible explanation of arthropods incude post-mortem transportation or moulting (e.g., et al., 2001; , 2002; & , 2006). et al. (2001) and et al. (2009) documented two rare finds of comparatively small hyolithid conchs containing articulated agnostids in prone attitude from the richly fossiliferous Paradoxides (Paradoxides) paradoxissimus gracilis Trilobite Biozone of the middle Cambrian Jince Formation. The aim of the present contribution is to describe a nearly complete hyolithid conch with an entombed enrolled agnostid; this specimen was recently found by the junior author at the Vystrkov Hill near Jince (see & , 2014, outcrop 6 in Fig. 1). The studied sample is housed in the collections of the Czech Geological Survey, Klárov 3, Praha 1, CZ-118 21, Czech Republic under the designation CW 010. and (2011), and (2012) and and (in press) discussed the feeding, storm disturbance and hiding hypotheses.
The fossil comes from an extensive quarry in green, fine-grained greywacke with thin lenses of shale of the middle Cambrian Jince Formation was exploited at the north-eastern slope of Vystrkov Hill near Jince (49°46'45.922"N, 13°58'2.379"E) from 1970 to the 90's. Lower levels of the quarried succession exposed deposits of the Paradoxides (Paradoxides) paradoxissimus gracilis Trilobite Biozone ( & , 2014). The associated skeletal fauna has been discussed by et al. (2004, 2009), et al. (2008), and and (2014). It consists of abundant trilobites (e.g., Conocoryphe, Paradoxides, Ptychoparia), agnostids (Peronopsis, Hypagnostus and Phalagnostus), echinoderms (e.g., Akadocrinus, Vyscystis, Etoctenocystis, Stromatocysties), bivalved arthropods (Tuzoia, Liangshanella and Konicekion) and hyolithids (e.g., Jincelites, Buchavalites) associated with rare brachiopods, gastropod-like molluscs, fossils distantly resembling Selkirkia and ichnofossils.
Occurrence of the zonal paradoxidid trilobite Paradoxides (P.) paradoxissimus gracilis and the index agnostid Hypagnostus parvifrons makes it possible to correlate a part of the Jince Formation exposed at Vystrkov Hill with middle and higher levels of the Baltic Paradoxides (P.) paradoxissimus Biozone ( & , 2003; & , 2008; & , 2013), as well as with the Caesaraugustian Regional Stage in the West Gondwanan chronostratigraphic sequence (see et al., 2008; et al., 2011). These levels correspond to the Drumian Stage of Cambrian Series 3.
Articulated exoskeletons of the tiny agnostid Peronopsis integra are abundant in the higher levels of the Paradoxides (Eccaparadoxides) pusillus Biozone and in particular in the Paradoxides (Paradoxides) paradoxissimus gracilis Biozone of the Jince Formation (see , 1958; et al., 2004). Similarly, conchs of the small to middle-sized hyolithid genus Buchavalites , 1975, have been found in the Paradoxides (Paradoxides) paradoxissimus gracilis Trilobite Biozone (because of poor preservation, these hyolithids were classified as Orthotheca sp. A by et al., 2004, tab. 5).
Palaeoecology of agnostids
Some authors provided arguments for a planktonic habit (e.g., , 1972, 1975), while others suggested an epifaunal mode of life (, 1977). However, evidence for a benthic mode of life in adult agnostids is accumulating (e.g., , 1974; & , 1987; , 2001; et al., 2003; & , 2011; & , 2014).
Palaeoecology of hyolithids
(1981) favoured a pelagic mode of life for hyolithids, but also pelagic, nektic and benthic habits have been suggested (e.g., , 1962). Many of authors accepted with a vagrant benthic habit (e.g., , 1957; , 1957, 1961; & , 1976, & , 1976; et al., 2011; et al., 2014). Hyolithids have never been considered as infaunal organisms.
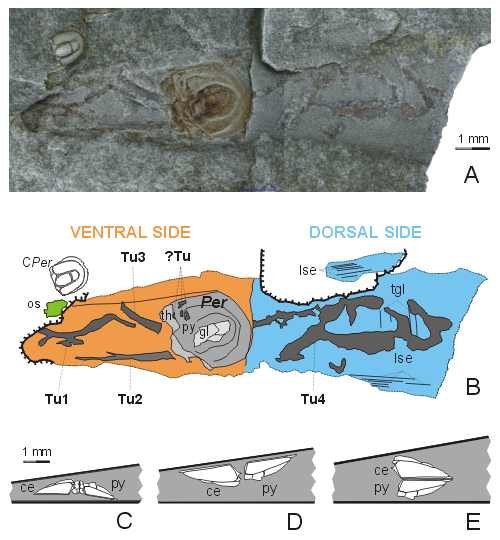
All the specimens are preserved in a small slab of fine shale measuring 22 x 19 mm. The rock surface bears 1) a nearly complete hyolithid conch, 2) an isolated cephalic shield of a small specimen of Peronopsis integra (CPer at
Fig. 1.B ![]() ), and 3) one ossicle of indeterminable echinoderm (os in
Fig. 1.B
), and 3) one ossicle of indeterminable echinoderm (os in
Fig. 1.B ![]() ). The preserved part of the hyolithid conch identified as ? Buchavalites sp. measures 13.35 mm in length and 3.25 mm in its widest visible part; the apical end of the conch is embedded in the sediment, the apertural part is broken off
(Fig. 1.A-B
). The preserved part of the hyolithid conch identified as ? Buchavalites sp. measures 13.35 mm in length and 3.25 mm in its widest visible part; the apical end of the conch is embedded in the sediment, the apertural part is broken off
(Fig. 1.A-B ![]() ).
Estimated length of the complete conch possibly exceeded 23 mm.
).
Estimated length of the complete conch possibly exceeded 23 mm.
Both ventral surface and the visible dorsal surface of the hyolithid conch are slightly compressed but uncrushed. The external surface of the conch is generally smooth; several very fine transverse growth-lines (tgl in
Fig. 1.B ![]() ) and fine narrowly spaced longitudinal
sculptural elements (lse in
Fig. 1.B
) and fine narrowly spaced longitudinal
sculptural elements (lse in
Fig. 1.B ![]() ) are seen in several areas; no trace of muscle scars is visible.
) are seen in several areas; no trace of muscle scars is visible.
Within the conch a specimen of Peronopsis integra is apparent. The external mould of the agnostid pygidium does not show any traces of crushing or biting on its exoskeletal surface. However, antero-laterally the surface of the pygidial shield bears at least four tiny holes of irregular shape (?Tu in
Fig. 1.B ![]() ). The pygidium measures about 2.25 mm in
sagittal length and 2.35 mm in maximum width. The axial part of the pygidial shield includes pressed remains of a glabella (gl in
Fig. 1.B
). The pygidium measures about 2.25 mm in
sagittal length and 2.35 mm in maximum width. The axial part of the pygidial shield includes pressed remains of a glabella (gl in
Fig. 1.B ![]() ). The external surface of the second thoracic segment (th in
Fig. 1.B
). The external surface of the second thoracic segment (th in
Fig. 1.B ![]() ) is preserved at the anterior pygidial margin. The agnostid exoskeleton lies with its pygidial dorsal side directed to the ventrum of the hyolith conch
(Fig. 1.E
) is preserved at the anterior pygidial margin. The agnostid exoskeleton lies with its pygidial dorsal side directed to the ventrum of the hyolith conch
(Fig. 1.E ![]() ). Tiny remains of the glabella are visible inside the pygidial axis and articulated remains of the second thoracic segment indicates that all these remains belong to a complete enrolled specimen with the dorsal side of the cephalon pointing to the dorsal side of the hyolithid conch
(Fig. 1.E
). Tiny remains of the glabella are visible inside the pygidial axis and articulated remains of the second thoracic segment indicates that all these remains belong to a complete enrolled specimen with the dorsal side of the cephalon pointing to the dorsal side of the hyolithid conch
(Fig. 1.E ![]() ). Total length of the articulated prone agnostid
is estimated to be about 5 mm.
). Total length of the articulated prone agnostid
is estimated to be about 5 mm.
Click on thumbnail to enlarge the image.
Figure 1: Agnostid inside a hyolithid conch. A internal mould of conch of ? Buchavalites sp. containing enrolled specimen of Peronopsis integra (, 1845), CW 010. Coated by ammonium chloride. B reconstruction of the conch of ? Buchavalites sp. and the specimen of Peronopsis integra (, 1845). C part of conch of Jincelites vogeli containing articulated specimen of Peronopsis integra, specimen VV 001, modified after et al. (2009, Fig. 3B). D part of the conch of ? Buchavalites sp. containing articulated specimen of Peronopsis integra, specimen VV 002, modified after et al. (2009, Fig. 3D). E part of the conch of ? Buchavalites sp. containing articulated specimen of Peronopsis integra, specimen CW 010. A, B dorsal views, C, D, E lateral views. Tu feeding tunnels of Arachnostega-strategist, ?Tu irregular holes in right antero-lateral surface of pygidial shield of Peronopsis integra, Per Peronopsis, ce cephalic shield, os echinoderm plate, gl glabella, lse longitudinal sculpture elements, py pygidial shield, tgl transverse growth-lines, th thorax.
The generally smooth external surface of the hyolithid conch bears several fine ribbon-like particles of diverse width and length. The presence of three longer ribbon-like elements behind the enrolled agnostid exoskeleton in the very narrow apical portion of the hyolithid conch (Tu1, Tu2 and Tu3 in
Fig. 1.B ![]() ) excludes the assignment of these particles to agnostid faeces. However, the irregular ribbon-like elements could represent either (1) scattered hyolithid soft parts (e.g., remains of the gut filling), or (2) they could belong to tunnels of a tiny burrow system.
) excludes the assignment of these particles to agnostid faeces. However, the irregular ribbon-like elements could represent either (1) scattered hyolithid soft parts (e.g., remains of the gut filling), or (2) they could belong to tunnels of a tiny burrow system.
(1) Remains of hyolithid guts have been rarely observed in the early Cambrian of China ( et al., 1982), western Nevada ( & , 2010) and Canada (, 2003), as well as in the middle Cambrian of the U.S.A. ( & , 1988) and China ( et al., 1992). Recently, et al. (2014) summarized all earlier published data on hyolithid guts proving presence of a simple U-shaped tract with straight branches in all hyolithids showing soft-parts.
Diverticulate character combined with diverse width and diagonal course of these elements in the apertural sector of the conch (Tu1, Tu2 and Tu4 in
Fig. 1.B ![]() ) excludes the interpretation of the ribbon-like particles as hyolithid gut fillings.
) excludes the interpretation of the ribbon-like particles as hyolithid gut fillings.
(2) et al. (2011, p. 375, Fig. 6) reported a rare occurrence of simple to divergent tunnels (= simply looping or ramifying systems) inside diverse invertebrate skeletons from the Barrandian area, including the middle Cambrian Jince Formation. These tunnels presumably represent early "growth stage" of an organism actively searching for food inside skeletons lying on the seafloor or partly embedded in it. Such tunnelling was proposed for the Arachnostega-type behaviour (strategy) and is classified as feeding traces of the ichnogenus Arachnostega (see , 1992; et al., 2011).
We prefer to ascribe the fine ribbon-like and ramifying tunnels to traces of the Arachnostega-type behaviour by an unknown organism (? errant polychaetes, see , 1992, p. 182). The tunnel by burrowing organism fed on decating soft tissue of the hyolithid; the general arrangement of these tunnels obviously copies the disposition of the gut inside a hyolithid conch (see et al., 2014).
Very good, nearly 3D, preservation of the agnostid pygidial shield, associated with parts of the tightly joined second thoracic segment and the remains of the glabella, proves the interpretation that the agnostid is an enrolled carcass rather than a moult. Accordingly, it could be concluded that, when the agnostid
entered the hyolithid conch, it was resting on the sea floor. Unfortunately, the incomplete preservation in a very small rock sample
makes it impossible to decide if the hyolithid was situated in its usual posture with the ligula resting on the sea floor or upside down. The orientation and disposition of the enrolled agnostid exoskeleton differs from entombed agnostids described earlier (cf.
Fig. 1.C-E ![]() ).
).
Similarly, as in other finds of entombed agnostids and eodiscid trilobites, it is not possible to give the exact reason why Peronopsis integra entered the restricted space within the hyolithid conch. However, at least four different hypotheses for an agnostid entering such a specific environment were proposed by et al. (2003) and et al. (2009):
(1) Hiding place. An empty conch provided a hiding place (sheltering) for the small animals during their usual activity;
(2) Disturbance. A hyolithid conch, either with or without the soft tissue, could be an ideal place to survive periods of seafloor disturbance;
(3) Food. A hyolithid conch with decaying soft parts within could provide a well-protected source of food for small benthic scavengers and/or
(4) Moulting in shelter.
(1) Hiding place. Scars attributed to sublethal predation have been only rarely documented in agnostids (, 2003). However, the recently described articulated specimen of Phalagnostus prantli with several small scars on the posterior pygidial border ( et al., 2009) documents the presence of potential predators of small benthic animals in the middle Cambrian of the Příbram-Jince Basin (compare et al., 2012).
Occurrence of potential predators supports the possibility of hiding during usual activity.
(2) Disturbance. Entombed specimens of Peronopsis supports the earlier speculation that some taxa of small agnostids voluntarily entered empty spaces inside various invertebrate skeletons lying on the bottom (e.g., et al., 2003; & , 2011). The enrolled attitude of the exoskeleton favours the seafloor disturbance hypothesis.
(3) Food. Presence of a simply looping to ramifying tunnel system of an Arachnostega-strategist proves that the hyolithid conch contained edible soft tissue. The co-occurrence of feeding tunnels and the enrolled agnostid Peronopsis integra inside the hyolithid conch makes possible the explanation, that the agnostid was attracted to feed on the hyolithid carrion inside the conch. Consequently, the hypothesis of a well-protected source of food for small benthic scavengers is suitable.
Four tiny holes visible at the pygidial surface (?Tu in Fig. 1.B ![]() )
imply that when the producer of the feeding tunnels entered the hyolithid conch, the enrolled agnostid had already been entombed inside this restricted space.
)
imply that when the producer of the feeding tunnels entered the hyolithid conch, the enrolled agnostid had already been entombed inside this restricted space.
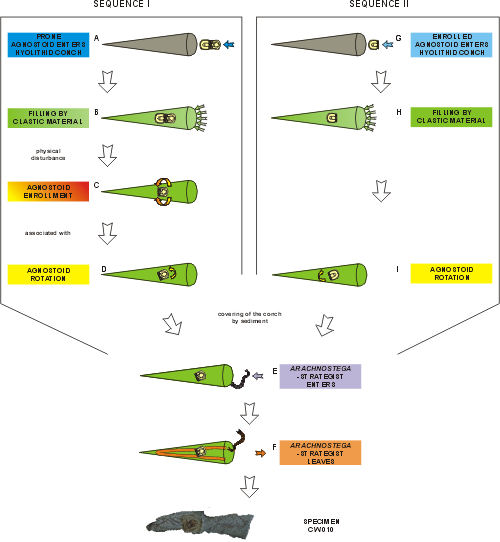
Based on the above discussion, the origin of the described association of the tunnel system of Arachnostega-strategist and Peronopsis inside a hyolithid could be explained by the following two sequences:
Sequence I
Sequence II
The rest of this sequence is the same as in the sequence I.
Click on thumbnail to enlarge the image.
Figure 2: Two simplified sequences leading to possible origin for the association of feeding tunnels produced by Arachnostega- strategist and Peronopsis inside hyolithid conch.
Earlier described entombed specimens of the eodiscid trilobite Pagetia bootes as well as the agnostid Ptychagnostus preacurrens and Peronopsis integra supposedly entered the restricted space inside priapulid worms and hyolithid conchs on its own as part of their normal behaviour ( et al., 2003; et al., 2009). The width of entombed eodiscid trilobites and agnostids ranges from 1.65 to 3.9 mm and reaches 55-89 % of width of the priapulid tube and hyolith conch in the place of entombment (Table 1). Width of the Peronopsis integra described here fits with the earlier observed range, as it reaches 89 % of the hyolith width in the place of entombment.
| Specimen | Entombed species | Entombed inside |
Length of conch |
Max width of conch / tube |
Width of conch / tube in the place with entombed specimen | Width of entombed specimen | Width of entombed specimen /width of conch or tube |
| VV 001A, B |
Peronopsis integra | Hyolithid conch (Jincelites) |
16 mm | 3.9 mm | 2.05-3.00 mm | 1.65 mm | 55-80 % |
| VV 002A, B |
Peronopsis integra | Hyolithid conch (Buchavalites) |
23 mm | 5.5 mm | 1.85-2.05 mm | 1.65 mm | 80-89 % |
| CW 010 | Peronopsis integra | Hyolithid conch (? Buchavalites) |
> 23 mm | > 3.3 mm | 2.6 mm | 2.3 mm | 88 % |
| ROM 543455 | Ptychagnostus preacurrens | Priapulid tube (Selkirkia) | 32.9 mm | 6.0 mm | 5.0-5.3 mm | 3.9 mm | 76 % |
| ROM 54344 | Pagetia bootes | Priapulid tube (Selkirkia) | > 29 mm | 5.7 mm | 5.5 mm | ? | - |
| 4.3 mm | 3.4 mm | 79 % | |||||
| 3.7 mm | 2.8mm | 76 % | |||||
| ROM 54346 | Pagetia bootes | Priapulid tube (Selkirkia) | > 24 mm | 5.7 mm | 4.4-4.8 mm | 2.8 mm | 61 % |
Table 1: Dimensions of entombed agnostids, eodiscid trilobites, hyolithid conchs and tubes of priapulid worm.
An enrolled agnostid specimen preserved inside a hyolithid conch represents a new type of entombment. The enrolled attitude of this agnostid Peronopsis prioritizes the seafloor disturbance hypothesis for explanation of entombed of this specimen. This fits well with the earlier observations that some taxa of small agnostids voluntarily entered empty spaces inside various invertebrate skeletons lying on the bottom (e.g., et al., 2003; & , 2011).
The described association of feeding tunnels produced by an Arachnostega-strategist and Peronopsis inside of a hyolithid conch is designated as a case of "frozen" behaviour sensu (1990) and and (2010).
This research was supported by the project PRVOUK P44 of the Ministry of education, youth and sports of the Czech Republic. We would like to express our thanks to Dr. Joachim (LMU Munich, Germany), Professor Arthur J. (Oregon State University Corvallis, U.S.A.) and Dr. Michal (University of Silesia Sosnowiec, Poland) for helpful comments and suggestions, which improved the clarity of the paper. Dr. A.W.A. (BMNH London, Great Britain) kindly read and helped improve the manuscript. Dr. Petr (Czech Geological Survey, Prague) and Mgr. Martina (Charles University, Prague) kindly commented on an earlier version of this manuscript. Mgr. Luká (Charles University, Prague) is acknowledged for help with photography.
N. & P. (2003).- A core drilling through Cambrian strata at Almbacken, Scania, S. Sweden: trilobites and stratigraphical assessment.- GFF, Stockholm, vol. 125, nº 3, p. 139-156.
L.E. (2003).- Trilobites in Paleozoic predator-prey systems, and their role in reorganization of Early Paleozoic ecosystems. In: P.H., M. & T.A. (eds.), Predator prey interactions in the fossil record.- Kluwer Academic Publisher, p. 55-92.
L.E. & R.A. (1988).- Taxonomy and paleobiology of some Middle Cambrian Scenella (Cnidaria) and hyolithids (Mollusca) from Western North America.- Kansas University Paleontological Contributions, Lawrence, vol. 121, p. 1-22.
M. (1992).- Arachnostega n. ichnog. - burrowing traces in internal moulds of boring bivalves (late Jurassic, Northern Germany).- Paläontologische Zeitschrift, Stuttgart, vol. 66, nº 12, p. 177-185.
E. (1845).- Über einige böhmische Trilobiten.- G. Reimer, Berlin, 47 p.
A.J. (1990).- Evolutionary paleobiology of behavior and coevolution.- Elsevier, London, 725p.
A.J. & G.O. Jr (2010).- Fossil behavior compendium.- CRC Press, Boca Raton, 363 p.
D.S. (1993).- Ecdysis in Flexicalymene meeki (Trilobita).- Journal of Paleontology, Tulsa, vol. 67, nº 6, p. 999-1005.
N.J. (2003).- Exceptional fossil preservation and the Cambrian explosion.- Integrative and Comparative Biology, Oxford, vol. 43, nº 1, p. 166-177.
B.D.E. (2001).- Trilobites. In: D.E.G. & P.R. (eds.), Palaeobiology, II.- Palaeontological Association, Blackwell, p. 387-389.
B.D.E., D.H. & R. (2003).- Cryptic behaviour in trilobites: Cambrian and Silurian examples from Canada, and other related occurrences.- Special Papers in Palaeontology, London, vol. 70, p. 157-173.
R.A., R.H.B. & C.H. (2001).- Trilobites within nautiloid cephalopods.-Lethaia, Oslo, vol. 34, nº 1, p. 37-45.
L., S., J.J., J.S. & D. (2014).- Terreneuvian Orthothecid (Hyolitha) digestive tracts from Northern Montagne Noire, France: Taphonomic, ontogenetic and phylogenetic implications.- PLoS ONE, San Francisco, vol. 9, nº 2, e88583.
H. (1957).- Bryozoans. In: H.S. (ed.), Treatise on marine ecology and paleoecology, vol. 2.- Memoirs of Geological Society of America, Baltimore, Memoir 67, p. 783-800.
J. (1981).- Origin of the Cephalopoda.- Acta Paleontologica Polonica, Warsaw, vol. 26, nº 2, p. 161-191.
A.M. & L.E. (2010).- Census of the Indian Springs Lagerstätte, Poleta Formation (Cambrian), western Nevada, USA.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 295, nº 1-2, p. 236-244.
J. & S. (2014).- Enrolled agnostids from Cambrian of Spain provide new insights about the mode of life in agnostids.- Bulletin of Geosciences, Prague, vol. 87, nº 2, p. 283-290.
O. & P. (in press).- Sheltered gregarious behavior of Middle Ordovician harpetid trilobites.- Palaios, Lawrence.
O., V. & M. (2004).- Stratigraphic distribution of Cambrian fossils in the Příbram-Jince Basin (Barrandian area, Czech Republic).- Senckenbergiana lethaea, Frankfurt am Main, vol. 84, nº 1/2, p. 369-384.
O., R., M., V. & M. (2011).- Arachnostega , 1992 in Cambrian of the Barrandian area (Czech Republic).- Acta Geologica Polonica, Warsaw, vol. 61, nº 4, p. 367-381.
O. & M. (2011).- Agnostid entombed under exoskeletons of paradoxidid trilobites.- Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, Stuttgart, vol. 259, nº 2, p. 207-215.
O. & M. (2014).- Biostratigraphy of Cambrian in the Příbram-Jince Basin (Barrandian area, Czech Republic).- Bulletin of Geosciences, Prague, vol. 87, nº 2, p. 411-427.
O., M. & P. (2008a).- Malformed agnostids from the Middle Cambrian Jince Formation of the Příbram-Jince Basin, Czech Republic.- Bulletin of Geosciences, Prague, vol. 83, nº 1, p. 121-126.
O., M., P. & V. (2008b).- Position of trilobites in Cambrian ecosystem: preliminary remarks from the Barrandian region (Czechia). In: I., R. & D. (eds.), Advances in trilobite research.- Cuadernos del Museo Geominero, vol. 9, Instituto Geológico y Minero de España, Madrid, p. 117-122.
O., V., J., M. & , M. (2009).- Agnostids entombed in hyolith conchs.- Memoirs of the Association of Australasian Palaeontologists, Canberra, vol. 37, p. 481-489.
D.W. (1962).- Small conoidal shells of uncertain affinities. In: R.C. (ed.), Miscellanea, Part W.-Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, p. 98-140.
R.H.B.& H.W.J. (2006).- Crab carapaces preserved in nautiloid shells from the Upper Paleocene of Huesca (Pyrenees, Spain).- Revista Mexicana de Ciencias Geológicas, México, vol. 23, p. 361-363.
R.H.B. & M. (1995).- Decapods in ammonite shells: examples of inquilinism from the Jurassic of England and Germany.- Palaeontology, London, vol. 38 nº 3, p. 63-75.
G., O., O. & A. (2008).- Cambrian. In: T. (ed.), Geology of Central Europe.- Geological Society of London, p. 155-202.
R., J.B., J. & E. (2011).- Correlation between the base of Drumian Stage and the base of middle Caesaraugustan Stage in the Iberian Chains (NE Spain).- Bulletin of Geosciences, Prague, vol. 86, nº 3, p. 545-554.
J.C., A.A., D.C., I. & M. (2009).- Giant trilobites and trilobite clusters from the Ordovician of Portugal.- Geology, Boulder, vol. 37, nº 5, p. 443-446.
J.T., D., A., Y. & C. (2012).- Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian.- Palaeontology, London, vol. 55, nº 2, p. 369-399.
M. & D.L. (2008).- Middle Cambrian trilobites of the suborders Agnostina and Eodiscina from the Oslo Region, Norway.- Palaeontographica A, Stuttgart, vol. 286, nº 1-3, p. 1-87.
J.B. (1974).- Evidence for scavengers from Middle Cambrian sediments.- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, vol. 1974, nº 1, p. 13-17.
A.A. & R.H.B. (2012).- Animal Behavior Frozen in Time: Gregarious Behavior of Early Jurassic Lobsters within an Ammonoid Body Chamber.- PLoS ONE, San Francisco, vol. 7, nº 3, e31893.
H.-L., Z.-W., X., X.-L. & L. (1982).- The Sinian-Cambrian Boundary in Eastern Yunnan, China.- People's Republic of China, 265 p. [In Chinese]
J.-R., Y.-L. & P. (1992).- Some Middle Cambrian hyolithids from Taijiang, Guizhou.- Acta Micropalaeontologica Sinica, vol. 9, nº 3 p. 257-265.
L. (1975).- Objev nové hyolitové fauny ve skryjskotýřovickém kambriu (The discovery of a new hyolithid fauna in the Middle Cambrian of Bohemia).- Bohemia centralis, Praha, vol. 4, 64-71.
L. & A. (1976).- The tabulate coral Hyostragulum, an epizoan with bearing on hyolithid ecology and systematics.- Lethaia, Oslo, vol. 9, nº 1, p. 51-64.
L. & E.L. (1976).- Aspect of the biology of Hyolitha (Mollusca).- Lethaia, Oslo, vol. 9, nº 1, p. 65-82.
M., L. & J.M. (2014).- A complete reconstruction of the Hyolithid skeleton.- Journal of Paleontology, Tulsa, vol. 88, nº 1, p. 160-170.
K.J. & D. (1987).- Morphology, ontogeny, and life habit of Agnostus pissiformis from the Upper Cambrian of Sweden.- Fossils and Strata, Oslo, vol. 19, p. 1-124.
I. (1977).- Agnostid trilobites of the Central Bohemian Ordovician.- Sborník geologických věd, Paleontologie, Praha, vol. 19, p. 7-44.
R.A. (1972).- Mode of life of agnostid trilobites.- 24th International Geological Congress, Montreal, Section 7, p. 33-40.
R.A. (1975).- Species diversity among agnostid trilobites.- Fossils and Strata, Oslo, vol. 4, p. 219-226.
M. (2002).- Krebse aus dem Oberen Muschelkalk von Osthessen und Thüringen. Teil 1, Pseudopemphix alberti (H. v. , 1840).- Veröffentlichungen aus dem Naturkundemuseum Erfurt, vol. 21, p. 15-38.
M. (1958).- Trilobiti českého středního kambria (The trilobites of the Middle Cambrian of Bohemia).- Rozpravy Ústředního Ústavu geologického, Praha, vol. 24, 280 p. (Pls. I-XLVI) [in Czech with English summary]
E. & W.J. (2006).- Earliest record of sheltering strategy for predator avoidance from the Early Cambrian of the Marble Mountains, California.- Geological Society of America Abstracts with Programs, Philadelphia, vol. 38, nº 7, p. 550.
M., O., V. & M. (2008).- Hyoliths with entombed trilobites cryptic behavior of trilobites? In: I., R. & D. (eds.), Advances in trilobite research.- Cuadernos del Museo Geominero, vol. 9, Instituto Geológico y Minero de España, Madrid, p. 411-413.
M., O., M. & V. (2011).- Carinolithidae fam. nov., Carinolithes bohemicus sp. nov. and Slehoferites slehoferi gen. et sp. nov. new hyolithid taxa from the Bohemian middle Cambrian (Skryje-Týřovice Basin Czech Republic).- Palaeobiodiversity and Palaeoenvironments, Berlin, vol. 91, nº 2, p. 101-109.
G.J. (1987).- Evolution and Escalation: An Ecological History of Life.- Princeton University Press, 527 p.
T. & A.T. (2014).- A highly diverse trilobite fauna with Avalonian affinities from the Middle Cambrian Acidusus atavus Zone (Drumian Stage) of Bornholm, Denmark.- Journal of Systematic Palaeontology, London, vol. 12, nº 1, p. 23-92.
E.L. (1957).- "Pteropods" of the Paleozoic. In: H.S. (ed.), Treatise on marine ecology and paleoecology, vol. 2.- Memoirs of Geological Society of America, Baltimore, Memoir 67, p. 827-828.
E.L. (1961).- The operculum and mode life of Hyolithes.- Journal of Paleontology, Tulsa, vol. 35, nº 1, p. 52-61.
M. & S. (2012).- A Silurian Bohemoharpes twice used an empty shell of an orthocone nautiloid as refuge for moulting. In: P. & O. (eds.), The 5th Conference on Trilobites and their relatives.- Czech Geological Survey, Prague, p. 58.