◄ Carnets Geol. 14 (14) ►
![]()
Contents
[Introduction] [Change in species richness ...]
[Oligocene to Upper Miocene palaeobiogeographical reconstructions
...]
[Conclusions] [Bibliographic references] and ... [Appendix]
Muséum national
d'Histoire naturelle, 55, rue Buffon, CP51, 75231 Paris cedex 05 (France)
Published online in final form (pdf) on October 14, 2014
[Editor: Bruno ;
language editor: Stephen ]
![]()
A general overview of the biodiversity of the marine fauna during the Tertiary period is developed in the context of the biogeographical evolution of the European realm. This study combines a reappraisal of the literature with a unique first-hand source of data on the richest marine group (the gastropods) from over a 25 million year period (Early Oligocene to Late Miocene). In total the French deposits have yielded more than 10,000 species from the Eocene to the Upper Miocene. Evidence of significant bias in the fossil record is pointed out for the Palaeocene (Danian) and Upper Eocene (Priabonian). For the period considered (Palaeocene to Late Miocene, ca 50 million years) the second highest diversity is reported in the Late Oligocene. This study reveals also the importance of pre-Miocene extinctions of genera. The Oligocene was when the latitudinal differentiation of the faunas was greatest. The local generic gastropod richness ranges from 59 (Rupelian of Belgium) to 494 (Chattian of Aquitaine). A clear trend towards homogenization appears in the Late Oligocene which leads to the development of a vast biogeographical region named Euro-West Africa. This faunal evolutionary pattern has never been demonstrated before and is a novel feature of the biogeography of the Eastern-Atlantic region.
Biogeography; Tethys; extinction; France; Paleogene; Miocene; molluscs.
P. (2014).- Temporal and latitudinal trends in the biodiversity of European Atlantic Cenozoic gastropod (Mollusca) faunas. A base for the history of biogeographic provinces.- Carnets de Géologie [Notebooks on Geology], Brest, vol. 14, nº 14, p. 273-314.
Évolution temporelle et latitudinale de la biodiversité des faunes de gastéropodes (Mollusca) du Cénozoique atlantique européen. Une base pour l'histoire des provinces biogéographiques.- Une revue globale de la biodiversité des faunes marines du tertiaire est proposée dans le cadre de l'évolution biogéographique du domaine européen. Cette étude est fondée sur une revue critique de la littérature et une source unique de données sur le groupe marin le plus riche (les gastéropodes) examinés durant une période de 25 millions d'années (Oligocène inférieur à Miocène supérieur). Au total, les dépôts français renferment plus de 10 000 espèces dans l'intervalle Éocène à Miocène supérieur. Des biais importants dans les archives fossiles sont soulignés pour le Paléocène (Danien) et l'Éocène supérieur (Priabonien). La richesse générique locale s'étire de 59 genres (Rupélien de Belgique) à 494 genres (Chattien d'Aquitaine). Pour la période considérée (Paleocène à Miocène supérieur, environ 50 Ma), la seconde plus grande diversité (richesse spécifique) est relevée pour l'Oligocène supérieur. L'Oligocène inférieur est la période durant laquelle la différentiation latitudinale des faunes est la plus grande. Une nouvelle tendance apparaît à l'Oligocène supérieur qui conduit à la formation d'une vaste région biogéographique nommée Euro-Ouest Africaine. Cette évolution faunistique n'avait jamais été démontrée avant et apparaît comme un nouveau schéma dans la biogéographie des faunes Est-Atlantiques.
Biogéographie ; Téthys ; extinction ; France ; Paléogene ; Miocène ; mollusques.
Located where the North Sea and tropical Tethyan faunas converge, the French Atlantic Cenozoic domain outranks all others in Europe with its particularly rich molluscan faunas. The preservation of fossil molluscs is also exceptional, being the finest in the World for the Eocene (the famous Lutetian malacofauna of the Paris Basin), the best in the Tethyan realm for the Lower Oligocene (Rupelian = Stampian stage) and the Upper Oligocene (Chattian stage), and one of the best for the Lower and Middle Miocene.
Because molluscs dominate Cenozoic marine fossils ( et al., 2002) and outnumber all other macro-invertebrate taxonomic groups, they provide one of the prime sources of information on the history of the biosphere. Paradoxically we do not know how many species of molluscs occurred in the Cenozoic of the French Atlantic. Therefore, I propose estimates based on two approaches. The first is for Eocene deposits and is founded principally on revised bibliographic data but includes new data and personal research. The second, a more detailed approach, is for the Oligocene and Miocene deposits, with a new generic and species synthesis based on a huge quantity of partly unpublished material collected in the Aquitaine, Loire and Paris basins (, 1997). Finally, a comparison of families' specific richness should provide insight into the global evolution of the malacofauna.
The paleobiogeography (provincialism) of French Atlantic Cenozoic molluscan faunas was based on qualitative observations (, 1979; , 1974, 1988), using restricted sources of data. This new review is the first general overview on the basis of a larger collection of data personally verified (Appendix Table 3) using a multivariate analysis at the generic level. Several questions may then be addressed: How much have the faunas changed after the eastward closure of the Tethys Sea? Was the latitudinal gradient of specific richness reinforced after the closure? Did the faunas affected by long-term global cooling evolve towards more marked provincialism?
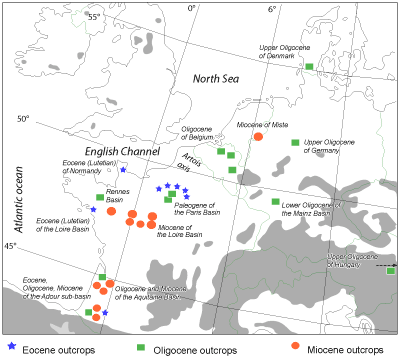
The geographical area taken into account includes the Paris Basin, the Aquitaine Basin and the Loire, Brittany, and Cotentin (Normandy) in France and the southern North Sea Basin (Belgium, Denmark and Germany), the Late Burdigalian/Middle Miocene of the Netherlands, and the Upper Oligocene of Hungary (Paratethys extension)
(Fig. 1 ![]() ).
).
For the comparison of global species richness we used data from the Palaeocene and Paleogene (Eocene and Oligocene) of the Paris Basin, Miocene of the Loire Basin and Oligocene and Miocene of the Aquitaine Basin
(Fig. 1 ![]() ). On the other hand the paleobiogeographic reconstructions for the Oligocene and the Miocene are based on the generic distribution and are limited to malacofaunas from the Lower Oligocene of Belgian and Germany, the Upper Oligocene of Belgian, Germany and Hungary and the Middle Miocene of the Netherlands.
). On the other hand the paleobiogeographic reconstructions for the Oligocene and the Miocene are based on the generic distribution and are limited to malacofaunas from the Lower Oligocene of Belgian and Germany, the Upper Oligocene of Belgian, Germany and Hungary and the Middle Miocene of the Netherlands.
Click on thumbnail to enlarge the image.
Figure 1: Locations of the faunas analyzed in the Cenozoic of the Eastern Atlantic coast basins and its North Sea extension.
During the Palaeogene, the composition of marine molluscan faunas of the Atlantic coasts is determined mainly by the influences of the Nordic realm (warm temperate waters) and the Tethyan realm (tropical and subtropical). The presence or absence of an isthmus between the North Sea Basin and the Paris Basin plays a major role in local biogeographic changes. The Middle Lutetian uplift of the Artois axis interrupted communication between the London-Belgian basins and the Atlantic. The Artois isthmus re-opened only much later, after the Miocene, when exchange was again possible via the English Channel. Meanwhile, we called the sudden invasion of the Paris Basin of molluscs of southern Tethyan origin a "Mesogean bubble" (, 2012). The best example is provided by the upper part of the Paris Basin Stampian (Lower Oligocene, Rupelian Stage) where a significant part of the Pierrefitte Sands fauna, beforehand known only in the latitude of the Aquitaine Basin, appeared suddenly (ca. 30 Ma).
Much later, in the latest Oligocene (Chattian) to Middle Miocene (Langhian and Badenian ages), the progressive closure of the eastern Tethyan seaway and its brief re-opening were the main palaeogeographical events. According to et al. (2007) this intermittent connection was definitively sealed during the middle Badenian (approx. 14-15 Ma).
The severe cooling during middle Miocene time (Serravallian age) is the first reliable evidence of a global change in the climate, suggested to be an incipient glaciation. The "greenhouse period" was definitely over (, 1994). However, before this time, a significant change occurred: the appearance and development of a new Eastern Atlantic faunistic province, during the Early Neogene (, 1983, 1988). (1888, 1909) first pointed out the similarity between European Neogene molluscan faunas and modern faunas of the West African biogeographical province. He considered that the changes were caused by an invasion of West-African species during the Early Miocene. In fact, this suggestion is speculative as (1888, 1909) did not have access to a body of data that was adequate to address the relationships of OligoceneUpper Miocene faunas of the French basins to their North Sea domain European counterparts. Only selected genera or species have been considered.
With respect to terminology we should note that the terms Boreal province (see et al., 2002), Nordic realm, Northern province (e.g., , 1979), Atlantic-Boreal Bioprovince ( & , 1984), Boreal-Celtic province ( & , 2007) and North Sea Basin designate roughly the same geographic and palaeogeographic area. Nevertheless, palaeoclimatological ambiguity exists in that the subtropical Boreal Miocene province is not equivalent to the modern cool-temperate Boreal-Celtic province. In contrast, the Mesogean realm (for example , 1979) is strictly equivalent to Tethyan realm. In fact Mesogean is routinely used in French geology as equivalent to Mediterranean or Tethyan.
The main taxonomic dataset used is the species-level data derived from material collected by the author. For basins for which I have little of my own data, the data result from a critical compilation from the literature (Appendix Table 3). Over the course of 25 years I (alone or with collaborators) have described about 300 new species, but 500-1000 or more, principally of small size, are still undescribed. In the database these species are labelled as "sp.". From a taxonomic point of view these "sp." can be considered as "morphospecies" or "Operational Taxonomic Units" (OTUs). These informal units are here taken to represent specific-level taxa, equivalent to formal "species". During sampling, special attention was given to fragile gastropod specimens and, in particular, to shells with well preserved protoconchs, because this part of the shell contains diagnostic characters. Identification of almost all molluscs was done or checked by the author.
The terms "outcrops" or "sites" are used for sections a few metres in extent. For example, the famous outcrop of Saucats, "Le Peloua" (Burdigalian of Aquitaine; National Geological Reserve of Saucats-La Brède), is the outcrop of a single layer about 40 cm thick. Generally, sampling was conducted in just one homogeneous layer (the richest), which rarely exceeds 2 m in thickness (1 m is the mean value). The lateral extent of outcrops is also generally limited, in the order of a few metres. Conversely, in the Aquitanian of Meilhan, the huge quarry of "Vives" has been extended considerably since 1990 ( et al., 2001b), allowing three separate samples to be taken. The extreme opposite facies consists of very thin detrital layers (4 cm thick) isolated in marl deposits but considered as "outcrops" because they are the only displaced representatives of the shallow-water molluscan fauna at a given time. In summary, almost all outcrops are entities restricted to a limited area (generally less than 20 m wide and 5 m thick), in contrast to the "regional" gathering of most earlier biodiversity studies. A total of 112 outcrops were investigated of which 104 were used to estimate species richness (Table 1). The Oligocene and Lower Miocene outcrops are the best represented.
All the outcrops of the Paris Basin (Paleogene), the Cotentin (Paleogene), the Loire Basin (Miocene) and the northern part of the Aquitaine (Oligocene and Miocene) consist of very shallow-water deposits. Nearshore calcareous facies characterise the Lutetian of the Paris Basin and Cotentin and the Stampian of the Aquitaine Basin. Rupelian ( & , 2012) and other Palaeogene and Eocene deposits of the Paris Basin are represented mostly by sandy facies. Marly facies occur in the southern part of the Aquitaine basin comprising the deeper deposits (circalittoral to upper bathyal for the Upper Oligocene; circalittoral for the Lower and Middle Miocene). True coral reefs are absent from all deposits but large coral colonies (metre-scale) of the zooxanthellate corals have been observed in the Oligocene (particularly Rupelian) and in the Lower Miocene Adour sub-basin.
In an attempt to examine more precisely the results for generic richness, the distances between the main faunas reliably recorded from the Lower Oligocene to the Upper Miocene have been analyzed using multivariate correspondence analysis (CA). The data were transformed into a presence-absence table (Appendix Table 3). Multivariate techniques permit an exploration of the structure of the dataset. In contrast to other multivariate techniques such as principal component analysis (PCA), CA allows simultaneous analysis of variables (columns: malacofaunas) and observations (rows: genera). This method can be employed for datasets that contain numerous zero values in the original matrix. The software package PAST [PAleontological STatistics (, 2013)], developed for structural study of palaeontological data, was used. The rarefaction curve and estimates of species richness were made using the software, EstimatesS 8 (, 2006).
It is a truism to say that early study of the French Cenozoic was central to the development of geology as a science. Among the 18th Century pioneers, Jean-Etienne (1715-1786) was the first to publish a geological monograph and to draw a map, which included the fossiliferous deposits of the Stampian (Rupelian), south of Paris. It is also at this time that the young A.L. (1743-1794), destined to become famous in chemistry, prepared the first geological section in the modern style (, 1989; , 2012). However, it is on the molluscan faunas that the main contributions to early reconstruction of Earth history were founded.
During the 19th Century Jean-Baptiste Pierre Antoine de , chevalier de (1744-1829) and Gérard-Paul (1796-1875) described the faunas collected from the Eocene and Oligocene deposits of the Paris Basin. During the same period Barthélémy de (1800-1887) and Jean-Pierre Sylvestre de (1782-1861) described the molluscs of the Aquitaine Basin and Félix (1801-1860) those of the Loire Basin (Touraine). Jacques Raoul (1822-1882) and Gustave Frédéric (1850-1931) later published monographs on the faunas of the Eocene, Oligocene and Miocene of smaller basins opening on the Atlantic Ocean. Crowning these contributions, Maurice (1850-1924) was the most prolific author in paleomalacology. He described and revised thousands of species and published some memorable monographs (e.g., & , 1904-1913). Albert (1860-1939) continued the work of concerning the Oligocene and Neogene of the Aquitaine Basin and described the Neogene fauna of the Loire Basin. Maxime (1905-1984) later reviewed the faunas of the Miocene basin of the Loire. More recently Louis (1915-1985) and Jacques undertook the description of many small species of the Parisian Eocene disregarded until the mid 20th Century (exactly 924 species, according to , 1999!).
With such a long and rich history, one might expect that these faunas were fully inventoried. However, this is not the case, because complete stratigraphic sequences have not been studied in all cases (e.g., in the Priabonian and the Upper Oligocene of the Aquitaine Basin) and "micro-molluscs" are still relatively neglected. As we shall see, the unexpected richness of the fauna of the Upper Oligocene of the Adour Basin revealed by and (2006) is a good illustration of the likelihood of future discovery of many more species.
Click on thumbnail to enlarge the image.
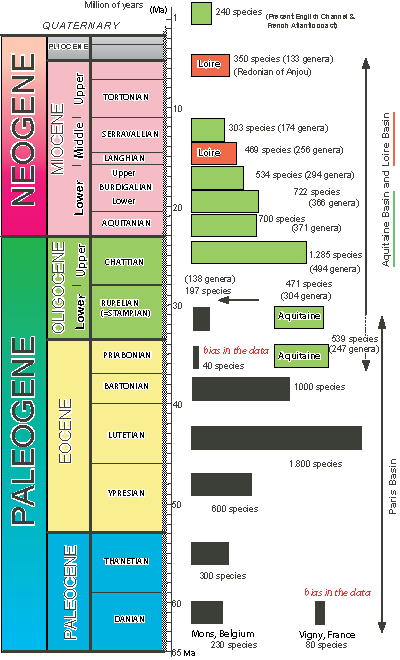
Figure 2: Gastropod richness of discrete faunas from the French Atlantic coast. Time scale is from (1994), with Quaternary boundary modified. Modified from (2012).
The richness of the reliably described faunas of the Eocene, Oligocene and Miocene
(Fig. 2 ![]() ) of the French Atlantic coast ranges from 197 species of gastropods (in the Rupelian of the Paris Basin) to 1800 species in the Lutetian of Paris Basin (,
2008, indicated "only" 1550 referenced species).
) of the French Atlantic coast ranges from 197 species of gastropods (in the Rupelian of the Paris Basin) to 1800 species in the Lutetian of Paris Basin (,
2008, indicated "only" 1550 referenced species).
The most diverse faunas are:
| Lutetian of the Paris Basin | 1800 |
| Upper Oligocene of the Aquitaine Basin | 1285 |
| Bartonian of the Paris Basin | 1000 |
| Lutetian of the Cotentin | 1000 |
| Lutetian of the Loire Basin | 850 |
| Lower Burdigalian of Aquitaine | 722 |
| Aquitanian of Aquitaine | 700 |
| Ypresian of the Paris Basin | 600 |
| Upper Burdigalian of Aquitaine | 534 |
Major sources for the Paleocene and Eocene molluscan record include (1886-1913), fully illustrated in and (1904-1913). The work of & summarized a large volume of literature on 2,500 species of gastropods (updated by and , 1995; , 2008). The most important additional reports are those of (1969, 1970) and and (1980, 1981), with revisions of 924 gastropod species (including numerous new species,, 1999). Other new Eocene data have been provided by J. following intensive collecting and critical reappraisal from the Eocene of the Paris Basin (, 2013) and the Cotentin deposits ( & , 2012). The previous compilation by (1986) is also very useful.
It is especially important to point out the low number of species recorded in the Priabonian (Ludian) and the Danian of the Paris Basin and other Atlantic outcrops, which deserves ecological explanation. The Danian sea includes the Paris area and the adjacent Mons Basin in Belgium (see , 1981, Fig. 21). The Danian fauna of the Paris Basin ( et al., 2000) is mainly known from identifications based on limestone moulds in a subreefal facies from a single outcrop ("Montian" of Vigny, France) and from a core sampling near Mons (type of the abandoned Montian stage), Belgium, 200 kilometres north of Vigny. Only 240 species of gastropods have been recorded (, 1973).
In the case of the late Eocene (Priabonian stage), the "Ludian" facies of the Paris Basin is also poor in molluscs. Marine sand and mud deposits with a well preserved fauna are restricted to a single somewhat brackish environment outcropping only in the Vexin. The index fossil is a potamidid, Potamidopsis vouastensis (, 1905). Similar Potamidopsis communities were also developed in the Middle Eocene (Lutetian, Bartonian) and Oligocene of the Paris Basin. In the Bartonian Potamidopsis crispiacensis (, 1906) community (Mortefontaine Sands, Loisy) as well as in the Priabonian Potamidopsis community (Chavençon outcrop, Oise), I recorded a very similar biodiversity (20 and 22 gastropod species, respectively) compared with the 60 species recorded by (1979) for the whole Priabonian fauna of the Paris Basin. The dramatically reduced species biodiversity in the Paris Basin near the end of the Eocene therefore appears to be an ecological and sampling bias (, 1979) and should not be ascribed to global climatic or other factors. This has been clearly demonstrated by the presence of a rich fauna of molluscs in Priabonian sections of Northern Europe: (1979) estimated that there are at least 732 species of molluscs at Latdorf.
In the Aquitaine Basin, Priabonian outcrops have recently become accessible in the southern Adour sub-basin. Thin detrital layers intercalated in bathyal marls harbour a rich fauna of Priabonian age (NP19/20; et al., 2002). Samples from just one outcrop have yielded at least 480 gastropod species. By comparison, one of the richest outcrops of the Middle Eocene of the Paris Basin (the Bartonian Baron quarry; et al., 1980) has produced 500 species of gastropods following intensive sampling.
The evidently very incomplete malacofaunal record of the Danian and Priabonian suggests that it should not be incorporated into a study of the rate of regional change in the Atlantic European basins. In North America where the two most diverse Eocene molluscan faunas of the Gulf Coast are Early and Middle Bartonian in age (respectively 495 and 580 species, & , 2001), rather than Lutetian, the Priabonian faunas of the Jackson Group are also relatively rich (426 species according to & , 2006). It is therefore not appropriate to accept the Paris Basin Danian and the Priabonian malacofaunas as a reliable basis for a study of the evolution of biodiversity.
The number of confirmed species, 3845, for the Oligocene and Miocene deposits of France is founded principally on my own data. For the Neogene of Aquitaine I identify 1500 species whereas and (1909-1924) and (1925-1935) documented 1337 species (from a total of 1459, erroneously including 122 Chattian species). We conclude that the Neogene faunas of Aquitaine have been relatively well known for a long time. This is not the case for the Oligocene faunas, with the exception of the Stampian (Rupelian) of the Paris Basin, for which we have added only 21 species ( & , 2012) to the 176 species documented by (1892, 1893). In the Stampian of the Aquitaine Basin, (1967a, 1967b, 1968) recorded only 180-190 species, whereas my dataset contains 471 species.
I have identified 1285 species of gastropods in the Upper Oligocene of the southern Aquitaine Basin in stark contrast to the 142 species reported by and (1909-1924) and (1925-1935) for this interval and a dozen species described or reported by and (1971, 1974), and (1970, 1975), (1969) and and (1978). By comparison, the modern Mediterranean Sea, considered a biodiversity hotspot of exceptional planetary value ( et al., 2010), has only 1221 species ( & , 2014). Rich bivalve material has also been collected in the Chattian of the Adour outcrops but has been only very partially studied. A review of Glycymerididae ( & , 1996) revealed the presence of at least seven species, whereas only two were recorded by and (1909-1924). In a preliminary, unpublished inventory, P. recorded 330 species of Bivalvia. Given the number of small species studied only briefly until now, it is possible to estimate the actual richness of the bivalves as between 350 and 450 species. Altogether, the entire Upper Oligocene malacofauna of the Adour Basin probably contains between 1700 and 1800 or more molluscan species. This high number of species contrasts with the much lower number indicated in the Oligocene of the Mediterranean-Iranian Province by (2007, Fig. 5: > 300 species) and deserves explanation.
This discrepancy is the result of a strong bias in the fossil record due to poor preservation of molluscs throughout the entire Mediterranean region in the Oligocene. One of the effects is, for example, the poor representation or lack of micro-molluscs in the dataset. Poor representation due to small size is a classic problem in palaeontology ( et al., 2006). Studies of modern faunas show that molluscs smaller than 10 mm constitute more than 50% of the species. By contrast, macromolluscs larger than 41 mm account only for 8% of the total ( et al., 2002).
This unexpected richness greatly modifies our perception of trends in the evolution of biodiversity in the malacofaunas in the European Cenozoic. The biogeographical consequences are far from being fully identified. However, and (2004) have already noted that many phyletic innovations described as characteristic of the Miocene (within Cypraeoidea in , 1996; Nassariidae in , 1984; Coninae in , 1990) appeared during the Late Oligocene. In the same way, some molecular phylogenic studies reveal an increase in the rate of cladogenesis of some unrelated tropical marine gastropod genera in the Indo-West Pacific (IWP) in Late Oligocene or Early Miocene times ( & , 2008).
Moreover, the recent recognition of the richness of the Upper Oligocene fauna of the Adour Basin is an excellent illustration that serious misinterpretations result from incomplete knowledge of the taxonomic composition of certain deposits. The negative implications are magnified where this gap in reliable data pertains to the stratigraphic record of France a cradle of historical geology, and a country where the fossil record has been regarded as well-known. This is reminiscent of the relative neglect of the Cenozoic molluscs of Westland (New Zealand) as pointed out by (1988).
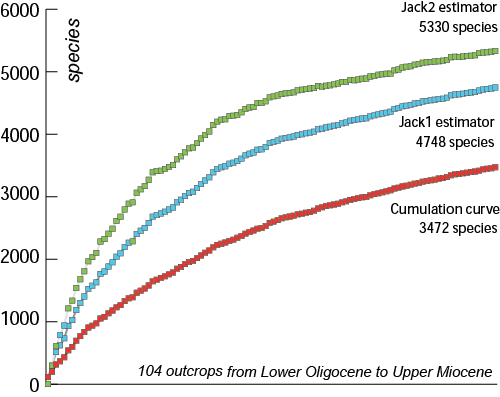
We propose a more precise estimation of the species richness from the data recorded in 104 outcrops that range from the Oligocene to the Upper Miocene of the French Atlantic coast (Table 1). Poorly sampled outcrops (with fewer than 500 specimens) are excluded. We retain 29 Lower Oligocene outcrops, 26 Upper Oligocene, 32 Lower Miocene, 15 Middle Miocene and only 2 Upper Miocene. A total of 275,000 specimens representing 3472 species have been examined.
Table 1:
Stratigraphic distribution and numerical age (from the GeoWhen Database) of 104 outcrops sampled for estimation of species richness (see
Fig. 3 ![]() ):
):
| Stage and numerical age | Paris Basin Number of outcrops |
Aquitaine Basin Number of outcrops |
Loire Basin Number of outcrops |
| Lower Oligocene (Rupelian): 33.9 +/- 0.1 to 28.4 (+/- 0.1 Ma) |
23 | 5 | 1 |
| Upper Oligocene (Chattian): 28.4 +/- 0.1 to 23.03 +/- 0.05 Ma |
26 | ||
| Lowe Miocene (Aquitanian): 23.03 to 20.43 +/- 0.05 Ma |
9 | ||
| Lower Miocene (Burdigalian): 20.43 to 15.97 +/- 0.05 Ma |
23 | ||
| Middle Miocene (Langhian): 15.97 to 13.65 +/- 0.05 Ma |
6 | 6 | |
| Middle Miocene (Serravallian): 13.65 +/- 0.05 to 11.608 +/- 0.005 Ma |
3 | ||
| Upper Miocene (Messinian): 7.246 To 5.332 +/- 0.005 Ma |
2 |
A cumulative species curve was constructed using EstimatesS 8 (,
2006) with 50 randomisations
(Fig. 3 ![]() ). We also used the estimators "jacknife-1" and "jacknife-2", calculated with EstimatesS, in order to evaluate the potential species richness. The cumulative curve is far from saturated and estimator curves indicate a possible numbers of at least 4748 species (Jacknife-1) and up to 5330 species with Jacknife-2. It is interesting to compare these numbers with the 2187 species of gastropods in the modern tropical fauna of New Caledonia collected in a single peri-reefal locality ( et al.,
2002), and with the estimate proposed for a more diverse region of 15,000 hectares at Panglao in the central Philippines (,
2009) of between 4000 and 14,000 species (note the magnitude of uncertainty!).
). We also used the estimators "jacknife-1" and "jacknife-2", calculated with EstimatesS, in order to evaluate the potential species richness. The cumulative curve is far from saturated and estimator curves indicate a possible numbers of at least 4748 species (Jacknife-1) and up to 5330 species with Jacknife-2. It is interesting to compare these numbers with the 2187 species of gastropods in the modern tropical fauna of New Caledonia collected in a single peri-reefal locality ( et al.,
2002), and with the estimate proposed for a more diverse region of 15,000 hectares at Panglao in the central Philippines (,
2009) of between 4000 and 14,000 species (note the magnitude of uncertainty!).
Thus the total of 5330 gastropod species (high-end prediction) estimated for French Atlantic coastal deposits of the Oligocene to Miocene, representing an interval of about 25 million years, seems realistic.
Click on thumbnail to enlarge the image.
Figure 3: Cumulative species curves based on EstimatesS 8 (, 2006) and Jacknife 1 (Jack1) and Jacknife 2 (Jack2) richness estimators. One hundred and four selected sites yielding 3472 species from Lower Oligocene to Upper Miocene have been included. The dataset comprises 3845 species for this interval.
From the data presented here, I suggest that the Palaeogene and Neogene basins of the French Atlantic coast contain at least 10 000 species of gastropods (about 4000-5000 or more for the Eocene; 5000 for the Oligocene and Miocene). In comparison, 4500 species of molluscs (Bivalvia plus Gastropoda) were reported by and (1990) and (1990) in the Cenozoic of New Zealand. However, (2006) considered that "in the best-preserved New-Zealand Cenozoic molluscan faunas, only 43% of the total preservable fauna is retained" (based on et al., 2006), a proportion probably similar for French localities. More recently, (2009) presented an estimate of 6000 species in New Zealand extrapolated from 3396 described species and only 2437 gastropod species. Also for New Zealand, A. reappraisal (pers. comm., 2011) estimates an "eventual" total of 7000 species for the entire Cenozoic (including about 5000 gastropods). (1985) indicated that 1600 species of molluscs had been described from the Tertiary of Australia, but several hundred more are yet to be described. (1984, 1986) and and (2006) reported about 3000 species of molluscs (Gastropoda and Bivalvia) from the Palaeogene of the northern Gulf Coastal Plain (Alabama-Mississippi). Thus, even if we consider that these estimates of regional specific richness are highly underestimated, it may be suggested that no other region in the world has as rich a Palaeogene and Neogene fossil record as the French Atlantic coast.
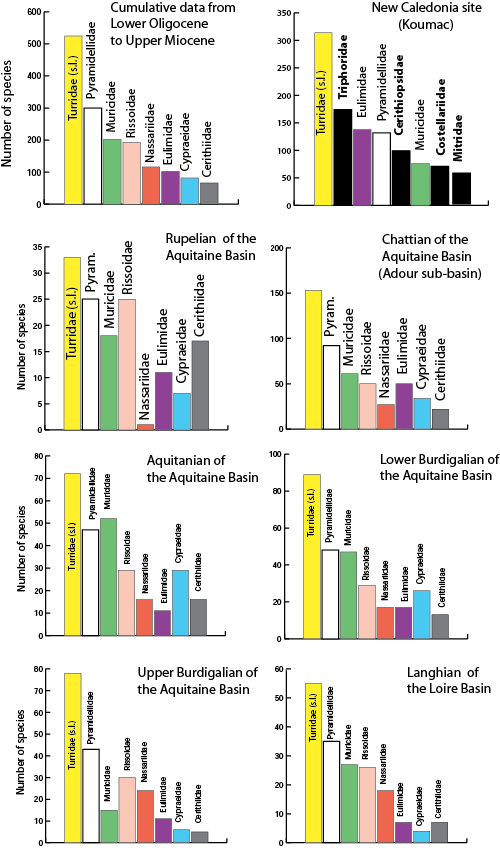
Based on the above reappraisal of West European fossil gastropod richness I selected the five families with the highest number of species from the dataset. Then I did the same for the faunas from the following six stratigraphic units: (1) Lower Oligocene and (2) Upper Oligocene of the Aquitaine Basin; for the Lower Miocene of the Aquitaine Basin: (3) Aquitanian, (4) Lower Burdigalian, (5) Upper Burdigalian; and the Langhian of the Loire Basin represents the Middle Miocene (6). In these faunas, the eight richest families (in decreasing order) are Turridae (sensu lato), Pyramidellidae, Nassariidae, Cerithiidae, Rissoidae, Muricidae, Cypraeidae and
Eulimidae. Figure 4 ![]() shows the change over time of species numbers in these families.
shows the change over time of species numbers in these families.
Click on thumbnail to enlarge the image.
Figure 4: Species richness of the eight most diverse families of Gastropoda in seven selected intervals compared with one modern example. For each fauna, the number of species is given according to the new dataset. Turridae (sensu lato) and Pyramidellidae are generally best represented, followed by Muricidae and Rissoidae. Among the eight most diverse families represented for the modern site of New Caledonia (Koumac), four (in black) are under-represented in the fossil faunas.
In all faunas, the Turridae and Pyramidellidae occupied the first and second places; the Rissoidae and Muricidae generally shared the third and fourth places. Cerithiidae are better represented in the Lower Oligocene, while the Eulimidae are most numerous during the Upper Oligocene and the Cypraeidae in the Lower Miocene. In the Upper Burdigalian and in the Middle Miocene, Nassariidae are especially diverse. The Turridae (sensu lato), including the families (or subfamilies according to different authors) Conidae, Turridae and Drilliidae, are unquestionably the most diverse group of marine molluscs ( et al., 2009). However, the number of species in the parasitic family Pyramidellidae is generally underestimated, probably because hosts have a patchy distribution and the shells of the Pyramidellidae are small and are not identified adequately (, 2009). The high number of species of Muricidae recorded in these Cenozoic deposits is surprising because this family is generally considered to have a relatively low species richness. This may be a bias due to the attractiveness of the Muricidae because of their larger size and varied morphology, permitting easy identification.
When we compare this distribution of fossil malacofaunas with that of the sub-reefal modern fauna of Koumac (New Caledonia), we find that the most species-rich family is also the Turridae (sensu lato), Triphoridae second, Eulimidae third, Pyramidellidae fourth, Cerithiopsidae fifth and Muricidae sixth ( et al., 2002). The main difference between European mid-Cenozoic faunas and modern warm-water faunas is the modern diversification of highly specialized families such as the Triphoridae and Cerithiopsidae, perhaps a peculiarity of coral-reef environments of the Indo-West Pacific.
In the intensively studies Lutetian of the Paris Basin (, 2008), the number of species of Triphoridae and Cerithiopsidae is low, about 20 ( & , 1980, 1981). The most diverse family is the Turridae (sensu lato) with some 200 species. The other highly diverse families are Marginellidae (46 species), Cerithiidae (44 species), Volutidae (41 species), Pyramidellidae (39 species), Epitoniidae (36 species), Turritellidae (35 species) and Buccinidae (34 species). We may therefore question the extreme diversity of the Turridae (sensu lato), the low number of Pyramidellidae and the expanded numbers of the "top eight" families (Marginellidae, Cerithiidae, Volutidae, Turritellidae, Buccinidae). Are they features specific to the Paris Basin during the Lutetian, or is it a global Eocene pattern? The low number of Pyramidellidae is probably an artefact resulting from an underestimate of their species number (, pers. comm., 2011; , 1969, 1970; & , 1995). By comparison, in the quarry at Baron (Bartonian, Paris Basin), et al. (1980) documented more than 500 species of gastropods (half of the recorded gastropods of this sub-stage). The Turridae (sensu lato) comprise only 47 species, while the second group is the Pyramidellidae and the Volutidae, with 21 species each; Cerithiidae and Buccinidae follow, with 16 species. In this locality, there are only three species of Triphoridae and seven species of Cerithiopsidae. Additional data are necessary before we can state with confidence that the hegemony of Neogastropoda (Turridae sensu lato, Volutoidea, Buccinidae), highlighted by the Lutetian and Bartonian record of the Paris Basin, is a general (global) characteristic of the Eocene.
Finally, Oligocene faunas are more similar, in their general composition, to those of the Miocene than those of the Eocene. If we compare the Oligocene and Miocene faunas with a well studied rich European modern malacofauna from the Mediterranean Sea (Spain, Andalusia, et al., 2011), we note that the richest families (in decreasing order) are the same: Pyramidellidae, Rissoidae, Turridae (sensu lato), followed by the Trochidae, Eulimidae, Nassariidae and Muricidae. Such close family biodiversity similarity suggests that the modern Atlantic-Mediterranean biogeographical province has a history reaching back to the Oligocene. However the usefulness of this parameter, intrafamilial species richness, as a proxy for global comparison seems limited, and it reveals only the main trend of the various basins.
The genus is the standard category for this type of biogeographical analysis because fossils are not always well enough preserved for identification. The modern molluscan provinces were initially based on generic distribution (, 1866; , 1880-1887). However since (1974) a criterion of at least 10% endemic species is generally employed in the recognition of modern provinces ( & , 2012). These authors divided the continental shelf into a series of large biogeographic regions that, in turn, contain smaller provinces. As a consequence, some very tiny islands may be recognized as a distinct province (e.g., the Eastern Atlantic Region, consisting only of St Helena Island). Otherwise the identification of the warm-temperate provinces is questionable and they are now included with tropical provinces in larger warm regions ( & , 2012). The endemism rate in the fossil deposits is generally poorly known and particularly difficult to determine at a specific level. In fact, such estimations depend so much on the preservation of the fossils that they must be used cautiously. Consequently, I focus here on significant trends in the evolution of the biogeographical regions (using analysis at generic level), rather than distinguish and describe the various provinces.
The gastropod basinal faunas may be compared in terms of richness. Generic richness varies from 32 (Rupelian of Brittany) to 494 (Chattian of the Adour Basin) with a mean of 220 and a median of 198. Brittany's (Rennes Basin, France) low diversity reflects poor sampling (only three ancient collections are available) but also the uniqueness of this semi-enclosed little basin. New, perfectly preserved material obtained from a drill-hole shows a depositional environment close to anoxic.
However, the low number of genera in the Rupelian of Belgium (59) compared to 139 in the Mainz Basin, 138 in the Paris Basin and 304 in the Aquitaine Basin mainly reflects a latitudinal gradient in species richness, which is particularly clear during the Lower Oligocene.
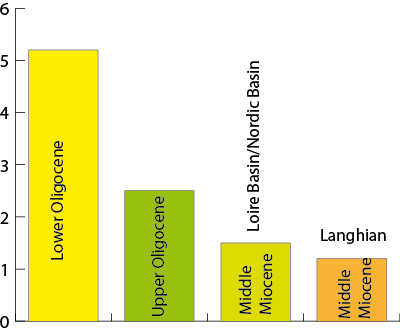
The richest fauna is in the Chattian of the Aquitaine Basin (open to the tropical Atlantic domain which is connected to the Tethys Realm). It includes 494 genera whereas, in the Upper Oligocene of the Nordic Basin, only 198 genera have been recognized giving a ratio of 2.5 (494/198). In the Lower Oligocene the ratio is higher (5.2, with 304 genera in the Aquitaine Basin, and 59 genera in the North Sea Basin). On the other hand, in the Middle Miocene faunas, the ratio between the Loire Basin (256 genera) and the Nordic faunas (173) decreases to 1.5. It is only 1.2 between the more ecologically diverse Langhian of the Aquitaine Basin and the North Sea Basin
(Fig. 5 ![]() ). The Upper Oligocene (Chattian) peak in southern Aquitaine gastropods is consistent with the generic richness of the zooxanthellate corals (z-corals) observed in the Aquitaine Basin ( & ,
1996). However
and (2008) pointed out that this highest generic diversity of Chattian z-corals is exceptional compared to the Paleogene and Neogene coral faunas listed in the Mediterranean Region (including the Aquitaine Basin).
). The Upper Oligocene (Chattian) peak in southern Aquitaine gastropods is consistent with the generic richness of the zooxanthellate corals (z-corals) observed in the Aquitaine Basin ( & ,
1996). However
and (2008) pointed out that this highest generic diversity of Chattian z-corals is exceptional compared to the Paleogene and Neogene coral faunas listed in the Mediterranean Region (including the Aquitaine Basin).
Click on thumbnail to enlarge the image.
Figure 5: The ratios between number of genera in the Aquitaine basin or the Loire Basin and the North Sea Basin express a strong latitudinal gradient of diversity in the Oligocene, and its progressive reduction.
The CA analysis
involved 16 malacofaunas from which 4000 species were distinguished resulting in the identification of 761 genera
(Appendix Table 3).
The data were transformed into a presence-absence table, where each cell is quoted 0 if the genus is absent and 1 if the genus is present
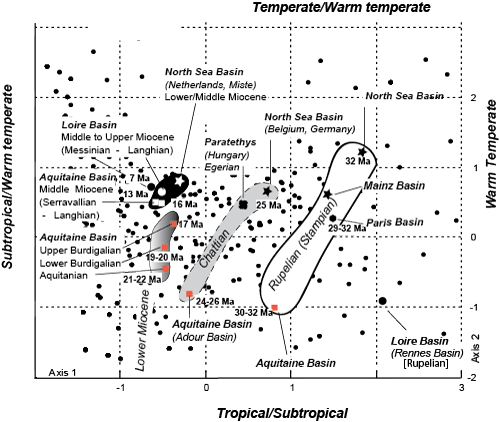
(Appendix Table 3). The two axes (first plane) represent 36.7% of the total variability (information) of the global samples. The first axis distinguishes, on the positive side, the malacofaunas from Oligocene
basins and, on the negative side, malacofaunas from Miocene basins. Axis 2 partly discriminates the North Sea Basin faunas (positive side) from the Aquitaine Basin faunas. The grouping of the 16 main discrete malacofaunas in homogeneous time-period clouds
(Figs. 6 ![]() - 7
- 7 ![]() ) reveals the long-term changes recorded in the malacofaunas of the European Atlantic coasts, from the Oligocene to the Late Miocene.
) reveals the long-term changes recorded in the malacofaunas of the European Atlantic coasts, from the Oligocene to the Late Miocene.
The Rupelian cluster is the widest while the Chattian cloud is reduced. During the Miocene, the size of the clouds was more or less restricted. These results may be expressed alongside the ratios of richness in genera, demonstrating a stronger latitudinal gradient of diversity within the Lower Oligocene (Tethyan Region opposed to the Nordic Region) and its progressive reduction during and after the Upper Oligocene. Although some genera and families, such as the Strombidae, did not reach the Nordic Sea Basin or are poorly represented (Cypraeidae) in Miocene deposits, it appears unequivocal that the Miocene faunas of northern Europe and southern Europe are closer than their Oligocene counterparts.
Click on thumbnail to enlarge the image.
Figure 6: Diagram showing the relationships of Rupelian to Messinian faunas on the French Atlantic coast basins and those of the North Sea basins (Boreal Province) based on multivariate analysis (correspondence analysis) at the generic level.
The first factorial plane (F1 and F2) is used, representing 36.7% of the total variability (information) of the global sample. The dataset contains 761 genera recorded from the Oligocene to the uppermost Miocene. The four clusters show a clear evolution, especially in the relationship between the Nordic Basin and the Aquitaine Basin. Compare the greater extension of the Rupelian cluster (white) on axis 2, in contrast to the focusing of the Middle Miocene (black). Black dots represent the position of molluscan genera.
Click on thumbnail to enlarge the image.
Figure 7:
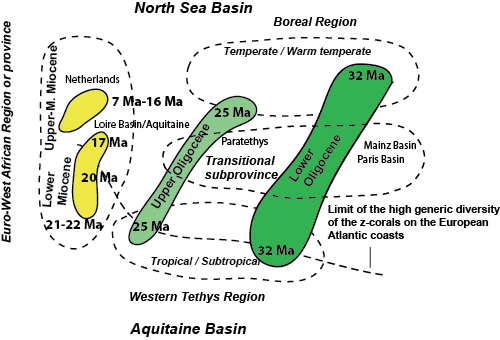
Interpretative diagram based on Figure 6 ![]() , showing the spatio-temporal distribution of Lower Oligocene to Upper Miocene faunas (based on the distribution of 761 genera). For the area considered, three main biogeographical domains can be clearly identified. For the Lower Oligocene we recognized a Boreal Region comprising the North Sea basin, the Western Tethys Region comprising the Aquitaine Basin and a Transitional subunit (Mainz basin, Rennes basin and Paris Basin). Compare the distance between North Sea Basin and Aquitaine Basin during the Lower Oligocene (32 Ma), during the Upper Oligocene (25 Ma) and during the Miocene; the smallest cluster matches the interval 7 to 16 Ma. The North Sea basins and the Aquitaine Basin became a single biogeographical unit, the Euro-West African Region, during the Miocene.
, showing the spatio-temporal distribution of Lower Oligocene to Upper Miocene faunas (based on the distribution of 761 genera). For the area considered, three main biogeographical domains can be clearly identified. For the Lower Oligocene we recognized a Boreal Region comprising the North Sea basin, the Western Tethys Region comprising the Aquitaine Basin and a Transitional subunit (Mainz basin, Rennes basin and Paris Basin). Compare the distance between North Sea Basin and Aquitaine Basin during the Lower Oligocene (32 Ma), during the Upper Oligocene (25 Ma) and during the Miocene; the smallest cluster matches the interval 7 to 16 Ma. The North Sea basins and the Aquitaine Basin became a single biogeographical unit, the Euro-West African Region, during the Miocene.
The genera listed in Table 3 reveal that some classic Tethyan groups of genera, common in the Lower Oligocene of the Paris Basin, such as Ampullinidae (Ampullinopsis, Deshayesia), Potamididae (Potamidopsis), Pseudomelaniidae (Bayania) and Brachytrematidae (Benoistia), are unknown from the Belgian Basin. But most of these genera are common in the Mainz Basin. Conversely, tropical groups of genera comprising the Trochidae (Lesperonia), Turbinidae (several species of Turbo), Angariidae (Angaria), Seraphsidae (Seraphs), Strombidae (Oostrombus, Strombus), Cassidae (Cassis), Pickworthiidae (Mareleptopoma), Cerithiidae (Gourmya), Potamididae (Serratocerithium), Vasidae (Vasum) and Harpidae (Morum, Eocithara), are unknown in the Paris Basin but common in the Aquitaine Basin and most also occur in Italy. The development of coral patches several metres across in the Aquitaine Basin contrasts strongly with the total lack of hermatypic corals in the Stampian of the Paris Basin (, 2012). During the Late Oligocene, some Tethyan groups reached the North Sea Basin, such as the Campanilidae (Campanile), the Harpidae (Morum), and the Olividae (Oliva), while some groups previously common in the Nordic Region also reached the Tethys Region (Nassariidae: Keepingia; Cassidae: Galeodea, etc.). In both domains, some modern genera of Nassariidae (Nassarius) and Turridae (sensu lato) (Perrona) appeared, whereas emblematic genera of the Palaeogene disappeared.
Finally, the pre-Neogene extinction of groups that were particularly common in the Early Oligocene or Late Oligocene deserves to be noted
(Fig. 8 ![]() ). In the Rupelian (Lower Stampian sands of Jeurre and Morigny) of the Paris Basin, 11 samples analysed (size fraction > 4 mm) yielded a total of 6358 specimens ( et al.,
1986). Thirty-three percent of the shells belong to the genera Bayania, Benoistia and Potamidopsis which become extinct at the end of the Oligocene
(Fig. 8
). In the Rupelian (Lower Stampian sands of Jeurre and Morigny) of the Paris Basin, 11 samples analysed (size fraction > 4 mm) yielded a total of 6358 specimens ( et al.,
1986). Thirty-three percent of the shells belong to the genera Bayania, Benoistia and Potamidopsis which become extinct at the end of the Oligocene
(Fig. 8 ![]() ). Thus, this was subject to extinctions that greatly changed the composition of the European mollusc communities. At least 64 genera disappeared at the mid-latitude of Europe and from the Mediterranean realm before the Lower Miocene
(Table 2). This result may be compared with the disappearance of 94
taxa from southern Europe (including the Mediterranean realm) during the Late Oligocene to Early Pliocene
(, 2011).
). Thus, this was subject to extinctions that greatly changed the composition of the European mollusc communities. At least 64 genera disappeared at the mid-latitude of Europe and from the Mediterranean realm before the Lower Miocene
(Table 2). This result may be compared with the disappearance of 94
taxa from southern Europe (including the Mediterranean realm) during the Late Oligocene to Early Pliocene
(, 2011).

Click on thumbnail to enlarge the image.
Figure 8: The Rupelian "losers". All species are from the Paris Basin (except F, I and O, from the Aquitaine Basin, Landes). These species belong to genera coming to extinction during the Oligocene epoch but particularly common in the Paris Basin Stampian (30-40% of the gastropod fauna from the data of et al., 1986) in the Lower Oligocene. The genera Ampullinopsis (I) and Oostrombus (J) contain giant Tethyan species reaching respectively 20 and 25 cm.
A-B, Bayania corrugata (, 1804). (A) from Auvers-Saint-Georges, 27 mm, (B) Pierrefitte, Saint-Hilaire, 20 mm.
C, Potamidopsis trochleare (typical morphotype) from Morigny Sands, "Trou à coquilles", Morigny-Champigny 28 mm.
D, Globulocerithium intratentatum (, 1864). 1-2: Auvers-Saint-Georges, 26.5 mm.
E, Benoistia boblayi (, 1824). Auvers-Saint-Georges, 18 mm.
F, Lesperonia trochoides (, 1870). "Lesperon", Dax, 10 mm.
G, Keepingia gossardii (, 1836). "Trou à coquilles", Morigny-Champigny, 31 mm.
H, Deshayesia cochlearia (, 1823). "Brunehaut", Morigny-Champigny, 29 mm.
I, Oostrombus auricularius (, 1834). "Espibos", Gaas, 140 mm.
J, Raulinia alligata (, 1824). Auvers-Saint-Georges, 8.3 mm.
K, Ampullinopsis crassatina (, 1804). "Brunehaut", Morigny-Champigny, 70 mm.
L, Praerosaria stampinensis (, 1894). "Pierrefitte", Saint-Hilaire, 18 mm.
M, Glibertturricula vervoeneni & , 1985. Core of the "Station de Pompage", Ormoy-La-Rivière, 20 mm.
N, Editharus heberti (, 1864). "Pont Royal", Étréchy, 17.5 mm.
O, Serratocerithium stroppus (, 1823). "Espibos", Gaas, 42 mm.
Table 2: Genera with last European/Mediterranean occurrence in the Oligocene: extinct in the Oligocene; Recent occurrence in the Indo-West Pacific (IP); Recent occurrence in the Western Atlantic (AW):
| Last Oligocene European & Mediterranean appearance | Extinct during the Oligocene | IP occurrence | AW occurrence |
| Aiziella | * | ||
| Ampullinopsis | * | ||
| Aneurychilus | * | ||
| Angaria | * | ||
| Angistoma | * | ||
| Aporrhais (Triaconthium) | * | ||
| Arene | * | ||
| Bartonia | * | ||
| Bayania | * | ||
| Benoistia | * | ||
| Boettgeriola | * | ||
| Boreosiphopsis | * | ||
| Bourdieria | * | ||
| Charitodoron (s.l.) | * | ||
| Cittarium | * | ||
| Conomitra | * | ||
| Crommium | * | ||
| Cryptoconus | * | ||
| Deshayesia | * | ||
| Diastoma | * | ||
| Editharus | * | ||
| Egerea | * | ||
| Ellobium | * | ||
| Eocithara | * | ||
| Fastigiella | * | ||
| Gabrielona | * | * | |
| Glibertturricula | * | ||
| Globulocerithium | * | ||
| Goergesia | * | ||
| Hameconia | * | ||
| Hortia | * | ||
| Iravadia | * | ||
| Keepingia | * | ||
| Keilostoma | * | ||
| Larocheopsis | * | ||
| Lesperonia | * | ||
| Littorinolacuna | * | ||
| Neodesmeria | * | ||
| Nucleolaria | * | ||
| Nystia | * | ||
| Pagodatrochus | * | ||
| Oostrombus | * | ||
| Potamidopsis | * | ||
| Praehyalocylis | * | ||
| Praerosaria | * | ||
| Pseudomalaxis | * | ||
| Raulinia | * | ||
| Sandbergeria | * | ||
| Seraphs | * | ||
| Serratocerithium | * | ||
| Sigatica | * | ||
| Spinobrookula | * | ||
| Sphaerocypraea | * | ||
| Spoelia | * | ||
| Stephopoma | * | * | |
| Subepona | * | ||
| Strebloceras | * | ||
| Streptodictyon (Fredenia) | * | ||
| Taphrostomia | * | ||
| Thereitis | * | ||
| Turbo (Amphiboliturbo) | * | ||
| Turbo (Sabronia) | * | ||
| Vicinocerithium | * | ||
| Volvaria | * |
All the authors consider that the Aquitaine Basin was part of the Western Tethys Region during the Early and Late Oligocene. However et al. (2002, 2007) distinguished an Eastern Atlantic Province for the Oligocene of the Aquitaine and Iberia basins and a Mediterranean/Iranian Province for the Mediterranean to Iran basins, these palaeobiogeographic provinces being subunits of the Western Tethys Region. This opinion is not founded on a critical analysis of the whole fauna but on selected species. In fact, apart from the exceptional preservation of the molluscan faunas in the Oligocene of the Aquitaine Basin, there is no reason to separate them, for example, from their Italian counterparts. All the species cited by et al. (2002) as characteristic gastropods of the Mediterranean/Iranian Province also occur in the Aquitaine Basin. I therefore suggest that this separation is not justified on grounds of endemism.
Although the opinion of (1979) that the same Mesogean/Tethyan tropical region extended from the Aquitaine basin to Iran needs to be reassessed in the light of new faunal discoveries, this remains the best hypothesis. My recent collecting in the classic Italian outcrops (Rupelian of Castelgomberto Hills, Vicentin) confirms the views of the classical authors (e.g., , 1870), who recognized the same faunas in Italian and Aquitanian deposits. In Iran, (2004) identified 42 genera of gastropods from the Abadeh deposits (Lower to Upper Rupelian). For the time being only two (Pseudophasianus and Rimella) are not known from the Lower Oligocene of the Aquitaine basin. However, Pseudophasianus occurs in Italy, while other genera, although shared between the Aquitaine Basin and Iran [Strombus (Tricornis), Peasiella, Astralium], are absent.
The Paris and Mainz basins, being at the same latitudinal position but apparently without direct inter-communication, are situated in a transitional province, intermediate between warmer subtropical and temperate waters, as expressed by their position in the factorial diagrams
(Figs. 6 ![]() - 7
- 7 ![]() ). The absence in these basins of "megathermic" mollusc groups as Strombidae, Vasidae and Cassis and the rarity of z-corals suggest that this subtropical to warm-temperate malacofauna should be included in the large Western Tethys Region as a transitional subunit. In the Paris Basin the endemism (at the specific level) was estimated by
and i (2012) at about 6%. If we follow and
(2012), this rate is too low to consider this area as a separate province. For the Early Oligocene (Rupelian) northern Europe is generally placed in another biogeographical domain (,
1979), generally named the Nordic or Boreal Province or Region
(Fig. 9
). The absence in these basins of "megathermic" mollusc groups as Strombidae, Vasidae and Cassis and the rarity of z-corals suggest that this subtropical to warm-temperate malacofauna should be included in the large Western Tethys Region as a transitional subunit. In the Paris Basin the endemism (at the specific level) was estimated by
and i (2012) at about 6%. If we follow and
(2012), this rate is too low to consider this area as a separate province. For the Early Oligocene (Rupelian) northern Europe is generally placed in another biogeographical domain (,
1979), generally named the Nordic or Boreal Province or Region
(Fig. 9 ![]() ). From my dataset of the Belgian Rupelian the gastropods share only 45% of species with the Paris Basin and 52% with the Mainz Basin, which also opened onto the
palaeo-North Sea.
). From my dataset of the Belgian Rupelian the gastropods share only 45% of species with the Paris Basin and 52% with the Mainz Basin, which also opened onto the
palaeo-North Sea.
We have shown that homogenization in the biogeographical distribution of gastropod genera began on the European Atlantic coasts during the Late Oligocene. This trend was amplified during the Miocene, possibly as a consequence of the tectonic connection between African and Eurasian littoral biotopes at the Oligocene/Miocene transition. This confirms the early intuitive conclusions of (1974, 1988) who claimed that, at that time, a large but somewhat homogeneous tropical to subtropical Eastern Atlantic faunal province succeeded the Palaeogene (pan-tropical) Tethys Province. This Eastern Atlantic Province probably extended from the Netherlands to Angola in the south ( & , 1995). These authors coined the term Euro-West African Province in order to designate this new European/African marine biogeographic area because it has consisted in part of modern West African genera since the Early Miocene.
Click on thumbnail to enlarge the image.
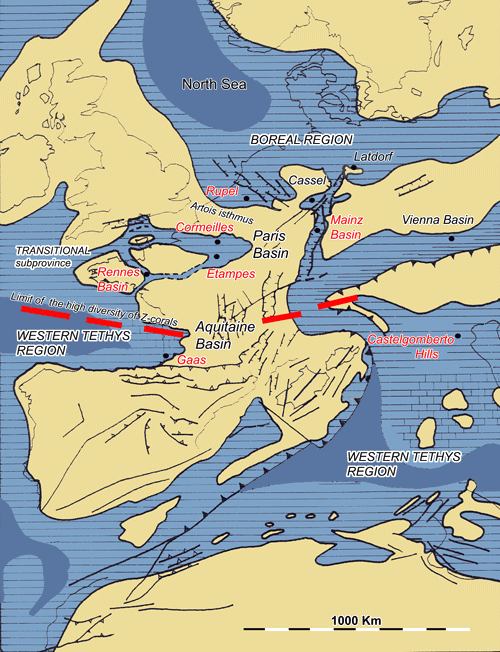
Figure 9: Reconstruction of European seas during the Early Oligocene (Late Rupelian, 32 - 29 Ma) with location of the main biogeographical areas. The main Rupelian localities are in red. Z-corals= zooxanthellate corals (, ed., 2012, modified after et al., 2000).
The close relationship between the modern West African biogeographical province (Tropical Eastern Atlantic province [TEA] of , 1974) and our Neogene Euro-West African paleoprovince is confirmed by generic affinities ( & , 1998). (1974, 1988) preferred the term Mediterranean Province (or Senegal Province, Guinean Province), and et al. (2007) coined Proto-Mediterranean Atlantic Region for the same concept.
In the same way, et al. (2002) used multivariate analyses at the species level in a study of the relationships between Paratethys and Boreal malacofaunas for the Middle Miocene. The Aquitaine Basin was separated by et al. (2002) and placed in a distinct SE North Atlantic Province, but they admitted, following F. (pers. comm. cited by et al., 2002), that a direct connection between Boreal Sea and southeastern North Atlantic is strongly supported by a very close correspondence of the concurrent Miocene faunas from the Aquitaine basin and the North Sea basin. In their demonstration they used a sample from Miste (the Netherlands) as a landmark of species distribution in the Late Burdigalian/Middle Miocene of the North Sea domain. In my analysis, using the same data from Miste (but at the generic level), the multivariate distance between this extremely rich Nordic outcrop (more than 350 species of gastropods, , 1984a) and the Middle Miocene faunas of the Loire and Aquitaine basins is very short, implying that they belonged to the same or very close biogeographical domains. The absence of "megathermic" taxa such as Strombus in the North Sea basin seems to justify the distinction of this region as a subprovince. Strombus disappeared from the French Atlantic Coast during the Middle/Late Miocene and from the Iberian Atlantic Coast during the Pliocene ( et al., 2011). However the recent discovery in the North Sea basin (Germany) of new thermophilous taxa such as Melongena and Cabania ( et al., 2010) suggests that the greatest care must be taken in making a final conclusion.
and
(2007) and et al.
(2011) proposed a reconstruction of the evolution of the Euro-West African region (or Province) from the Pliocene to the present. In the Pliocene the European Atlantic Coast (excluding the Nordic Basin) belonged to a French-Iberian Province, whereas the Mediterranean Sea and the West African Coast were part of a Mediterranean-West African Province. For the modern biogeography et al.
(2011) distinguish the Lusitanian region for the area extending from the English Channel to Mauritania (with two sub-provinces, the Mediterranean and the Mauritanian), with the West African region south of Cap Blanc and the Boreal-Celtic region north of Brittany
(Fig. 10 ![]() ).
).
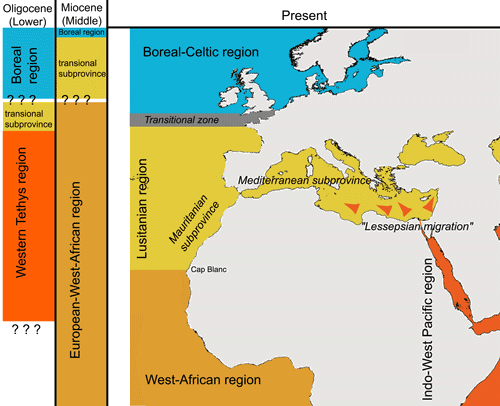
Click on thumbnail to enlarge the image.
Figure 10: Global evolution of molluscan biogeographic regions in the north-eastern Atlantic during the Oligocene and the Miocene. Modern bioprovinces after et al. (2011) and (1953). The "Lessepsian migration" (, 1978; et al., 2003), modern tropicalization of the Mediterranean biota which restores a biogeographical link between the Mediterranean and IWP, is also illustrated. Figure adapted from and (2007).
In order to understand the marine faunal history of a region, it is necessary to discriminate between biodiversity changes that may result simply from variations in the quality of the fossil record and genuine evolution linked to climatic or other changes. The diversity estimates are greatly affected by several factors: the number of faunas that have been studied, how well studied the faunas are, diversity of sampled biotopes (over-represented or absent), taxonomic uncertainty, and the tyranny of a high number of specimens (including demonstration of the species identity of taxa and the difficulty of comparison among such large faunas). Thus, we can say that the present report is an aspirational outline. However, with estimates of the number of gastropod species greatly exceeding 10,000 in the Eocene to Miocene deposits of the French Atlantic region (4000-6000 for the Eocene; more than 5300 estimated species for the Oligocene and Miocene) this first comprehensive synthesis characterizes the exceptional diversity of this region with greater accuracy than before, while highlighting the requirement for further studies of these faunas.
Despite the unquestionably high species richness, some faunas remain only superficially known or unstudied (especially the Upper Eocene and Upper Oligocene molluscs). It is also clear that a continuous record of Oligocene and Miocene marine faunas is
not preserved in all parts of Western Europe. On the other hand, when the local species richness of fossil molluscs is compared with the number of species now living along the modern French Atlantic coast, the drastic change must be underlined. About 240 species of shelled Gastropoda are recorded in the living fauna (S.
, pers. comm., 2010; see
Fig. 2 ![]() ), differing markedly from the 1300 species recorded in the Upper Oligocene deposits of the Adour sub-basin or the 350 species listed in the Upper Miocene (Redonian) of the Loire Basin. However, even if the peak of diversity in the Upper Oligocene is overstated (due to a large bathymetric range), we cannot consider that the Miocene was a period of higher biodiversity than the Oligocene.
Figure 2
), differing markedly from the 1300 species recorded in the Upper Oligocene deposits of the Adour sub-basin or the 350 species listed in the Upper Miocene (Redonian) of the Loire Basin. However, even if the peak of diversity in the Upper Oligocene is overstated (due to a large bathymetric range), we cannot consider that the Miocene was a period of higher biodiversity than the Oligocene.
Figure 2 ![]() indicates, rather, a gradual tendency towards impoverishment of the North Atlantic faunas during Miocene time. In fact, there is no clear peak in biodiversity.
indicates, rather, a gradual tendency towards impoverishment of the North Atlantic faunas during Miocene time. In fact, there is no clear peak in biodiversity.
Changes in the distribution of genera during the Oligocene to Late Miocene reveal a stronger latitudinal gradient in biodiversity within the Early Oligocene Atlantic domain (from the Tethyan region to the North Sea region). During the Early Oligocene (Rupelian) a transitional subtropical area (included to the vast Western Tethys region) is suggested around the latitudes of the Loire, Paris and Mainz basins. At this time, the North Sea Basin clearly belonged to another warm temperate region generally known as the Boreal region. The reduction of the latitudinal gradient during the Late Oligocene and the Miocene led to the development of an enlarged Euro-West African Province
(Fig. 10 ![]() ). The closure of the Tethys seaway at the end of the Badenian (early Middle Miocene) favoured this process with the development of longitudinal exchanges instead of latitudinal exchanges. Indeed the segmented Tethys Ocean parallel to the equator gave birth to a Mediterranean Sea open to the Atlantic Ocean. This Tethys successor was a melting pot and a source of novelty, but cannot be compared with the enormous faunal reservoir preserved in the Eastern Tethys which became the Indo-West Pacific region in the Middle Miocene; its
exceptional richness resulting from very long stability. Some sporadic links have been pointed out between European faunas and the new Indo-West Pacific region during the early Middle Miocene optimum (Badenian stage) through the Paratethys realm. This is indicated by the development of a particularly rich fauna and well expressed by the gastropod family Pickworthiidae (,
2004) and the echinoderm fauna (,
2007). This underlines the long-lasting importance of the Tethys (or its Indo-West Pacific heir) as a reservoir of malacological biodiversity. The depressed Miocene diversity in the Atlantic realm is reinforced during late Neogene times by other climatic events linked
with global step-by-step cooling leading to the late Pleistocene "ice-house" period.
). The closure of the Tethys seaway at the end of the Badenian (early Middle Miocene) favoured this process with the development of longitudinal exchanges instead of latitudinal exchanges. Indeed the segmented Tethys Ocean parallel to the equator gave birth to a Mediterranean Sea open to the Atlantic Ocean. This Tethys successor was a melting pot and a source of novelty, but cannot be compared with the enormous faunal reservoir preserved in the Eastern Tethys which became the Indo-West Pacific region in the Middle Miocene; its
exceptional richness resulting from very long stability. Some sporadic links have been pointed out between European faunas and the new Indo-West Pacific region during the early Middle Miocene optimum (Badenian stage) through the Paratethys realm. This is indicated by the development of a particularly rich fauna and well expressed by the gastropod family Pickworthiidae (,
2004) and the echinoderm fauna (,
2007). This underlines the long-lasting importance of the Tethys (or its Indo-West Pacific heir) as a reservoir of malacological biodiversity. The depressed Miocene diversity in the Atlantic realm is reinforced during late Neogene times by other climatic events linked
with global step-by-step cooling leading to the late Pleistocene "ice-house" period.
Last we note the extinction of groups that were particularly common in the Early Oligocene (Rupelian) or in the Late Oligocene. Thus the Oligocene suffered from extinctions that have greatly changed the composition of molluscan communities.
I thank my colleagues C. , L. , R. , and P. for considerable assistance with field work over a very long time. Thanks to A. , R.H. , D.T. , A. , K.I. and G.J. for helpful comments and review of the text. Jean-Claude provided numerous helpful suggestions and much useful information. The final draft was fully reviewed by S. . This work was supported by the PICS 4723-CNRS "Carbonates production from Greenhouse to Ice-house Earth System" and the project ANR-09-Pext "LOSERS".
T. (1973).- Mollusc fauna of the Hungarian Upper Oligocene (Egerian). Studies in stratigraphy, palaeoecology, palaeogeography and systematics.- Akadémiai Kiadó, Budapest, 393 p. (51 Pls.).
A.G. (1990).- Molluscan generic diversity of New Zealand Neogene stages: Extinction and biostratigraphic events.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 77, p. 279-288.
A.G. (2006).- Marine Mollusca of oxygen isotope stage of the last 2 million years in New Zealand. Part 2. Biostratigraphically useful and new Pliocene to Recent bivalves.- Journal of the Royal Society of New Zealand, Wellington, vol. 36, nº 4, p. 151-338.
A.G. & P.A. (1990).- Cenozoic Mollusca of New Zealand.- New Zealand Geological Survey Paleontological Bulletin, Lower Hutt, vol. 59, 583 p. (57 Pls.).
F. & C. (2008).- Estimating Mediterranean Oligocene-Miocene sea-surface temperatures: An approach based on coral taxonomic richness.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 258, p. 71-88.
P. (2009).- From specimens to data, and from seashells to molluscs: The Panglao Marine Biodiversity Project.- Vita Malacologica, Leiden, nº 8, p. 1-8.
P., P., P. & V. (2002).- Assessing the magnitude of species richness in tropical marine environments: Exceptionally high numbers of molluscs at a New Caledonia site.- Biological Journal of the Linnean Society, London, vol. 75, p. 421-436.
P., P. & A. (2009).- An inordinate fondness for turrids.- Deep-Sea Research Part II, Gloucester Point, vol. 56, p. 1724-1731.
P. (1964, unpublished).- Les gastéropodes du Redonien et leur signification.- Thèse de Doctorat d'État, Paris, 809 p. (6 cartes, 15 Pls.).
P. (1974).- Les gastéropodes du Miocène atlantique.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, t. 78, nº 1, p. 279-285.
P. (1983).- Étude d'une faune de gastéropodes miocènes récoltés par M.M. dans le Sud de l'Angola.- Comunicaçoes dos Serviços Geologicos de Portugal, Lisboa, t. 69, nº 1, p. 161-171.
P. (1988).- Évolution dans le temps et l'espace des gastéropodes marins dans la province Nordique depuis le Miocène.- Bulletin du Muséum national d'Histoire naturelle, Paris, (4e série), vol. 10, section C, nº 2, p. 163-173.
J.C. (1974).- Marine zoogeography.- McGraw-Hill, NewYork, 475 p.
J.C. & B.W. (2012).- A realignment of marine biogeographic provinces with particular reference to fish distributions.- Journal of Biogeography, Oxford, vol. 39, p. 12-30.
B. & C. (1996).- Structural and faunal evolution of Chattian-Miocene reefs and corals in western France and the northeastern Atlantic Ocean. In: E.K., M., W.C. & J.-M. (eds.), Models for carbonate stratigraphy from Miocene reef complexes of the Mediterranean regions.- Concepts in Sedimentology and Paleontology, SEPM, Tulsa, vol. 5, p. 105-127.
C. (1979).- La limite Éocène-Oligocène en Europe Occidentale.- Sciences Géologiques, Strasbourg, Mémoire, nº 54, 280 p.
W.O. (1984).- Systematics of the family Nassariidae.- Bulletins of the Auckland Institute and Museum, Auckland, vol. 14, 356 p. (51 Pls.).
C. (2012).- Le contenu paléontologique : Spongiaires et coraux. In: P. (coord.), Stratotype Stampien.- Biotope, Mèze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 4, p. 228-229.
M., C., J., K., F.B.R., J., E., C.N., J., T., R., M., C., B.S., J.M., R., J., F., K., M.S., A., N., E., de la C.M., H.K., D., D., D., S., J., J.I., C., S., J., X., D., R. & E. (2010).- The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats.- PLoS One, vol. 5, nº 8, p. 1-36 [doi:10.1371/journal.pone.0011842].
R.K. (2006).- EstimateS: Statistical estimation of species richness and shared species from samples, version 8.- Online at http://viceroy.eeb.uconn.edu/estimates/
R.A., P.A., J.S., A.G., C.M. & B.A. (2006).- Completeness of the fossil record: Estimating losses due to small body size.- Geology, Boulder, vol. 34, p. 241-244.
M. (1886-1913).- Catalogue illustré des coquilles fossiles de l'Éocène des environs de Paris.- Annales de la Société royale de Malacologie de Belgique, Bruxelles, 1er fascicule, t. 21 (1886), p. 3-174; 2ème fascicule, t. 22 (1887), p. 1-218; 3ème fascicule, t. 23 (1888), p. 3-328; 4ème fascicule, t. 24 (1889), p. 3-385; 5ème fascicule, t. 26 (1891, published in 1892), p. 3-167; Appendice 1, t. 28 (1893), p. 1-26; Appendice 2, t. 31 (1896), p. 1-94; Appendice 3, t. 36 (1901, published in 1902), p. 9-110; Appendice 4, t. 41 (1906, published in 1907), p. 187-318; Appendice 5: t. 49 (1913), p. 19-238.
M. (1892).- Révision sommaire de la faune du terrain oligocène marin aux environs d'Étampes.- Journal de Conchyliologie, Paris, t. 40, p. 330-375.
M. (1893).- Révision sommaire de la faune du terrain oligocène marin aux environs d'Étampes (suite).- Journal de Conchyliologie, Paris, t. 41, p. 297-363.
M. & A. (1909-1924).- Conchologie Néogénique de l'Aquitaine. Edition in-4 et in-8.- Actes de la Société Linnéenne de Bordeaux, t. I, nº 1, 220 p. (1909); t. I, nº 2, p. 221-428 (1911); t. I, nº 3, p. 429-718 (1912); t. II, nº 1, 204 p. (1913); t. II, nº 2, p. 205-410; t. II, nº 3, p. 411-496 (1914); t. III, nº 1, p. 1-384 (1917); t. III, nº 2, p. 385-695 (1919); t. IV, nº 1, 322 p. (1922); t. IV, nº 2, p. 323-610 (1924).
M. & G. (1904-1913).- Iconographie complète des coquilles fossiles de l'Éocène des environs de Paris.- Hermann, Paris, t. 1 : Pélécypodes, Pls. 1-16 (1904), Pls. 17-38 (1905), Pls. 39-45 (1906) ; t. 2 : Gastéropodes, Pls. 1-9 (1907), Pls. 10-25 (1910), Pls. 26-45 (1911), Pls. 46-65 (1913).
T.A. (1985).- Molluscan biogeography and biostratigraphy of the Tertiary of southeastern Australia.- Alcheringa, Wollongong, vol. 9, p. 3-116.
D.T. III (1984).- Crisis events for Paleogene molluscan faunas in the southeastern United States.- Mississippi Geology, Jackson, vol. 5, nº 2, p. 1-7.
D.T. III (1986).- Punctuated succession of Paleogene mollusks in the northern Gulf Coastal Plain.- Palaios, Lawrence, nº 1, p. 585-589.
D.T. III & P. (2006).- Eocene-Oligocene molluscan extinctions and first appearance data across the North Atlantic in the North American Gulf Coastal Plain and Europe. In: D.R., L.C. & E.A. (eds.), From Greenhouse to Icehouse. The marine Eocene-Oligocene transition.- New York, Columbia University Press, p. 303-340.
C., L. & J. (1980).- Inventaire systématique des mollusques de l'Auversien à "faciès charrié" de Baron (Oise), et remarques paléontologiques.- Bulletin d'Information des Géologues du Bassin de Paris, Paris, t. 17, nº 2, p. 26-48.
L. & P. (2004).- Nouvelles espèces de gastéropodes (Mollusca : Gastropoda) de l'Oligocène et du Miocène inférieur de l'Aquitaine (Sud-Ouest de la France). Partie 3. Cypraeidae et Ovulidae.- Cossmanniana, Leiden, (hors série), t. 4, 164 p. (36 Pls.).
G.F. (1888).- Une coquille remarquable du falun de l'Anjou : Melongena cornuta .- Bulletin de la Société d'Études Scientifiques d'Angers, Angers, p. 1-34 (4 Pls.).
G.F. (1909).- Essai sur l'Étage Aquitanien.- Bulletin pour Servir d'Explication à la Carte géologique de France, Paris, nº 124, t. 19, p. 379-506 (6 Pls.).
S. (1953).- Zoogeography of the sea.- Sidgwick & Jackson, London, 417 p.
F. (1989).- La première coupe historique du stratotype d'Étampes, dressée par en 1767.- Travaux du Comité Français d'Histoire de la Géologie (COFRHIGEO), (Série 3), vol. 3, nº 2, p. 6-17 (1 carte).
P. (1880-1887).- Manuel de conchyliologie et de paléontologie conchyliologique.- F. Savy, Paris, 112 p. (1880), p. 113-304 (1881), p. 305-416 (1882), p. 417-608 (1883), p. 609-688 (1884), p. 689-896 (1885), p. 897-1008 (1886), p. 1009-1367 (1887).
T. (1870).- Beitrag zur Kenntnis der Conchylienfauna des Vicentinischen Tertiargebirges.- Denkschriften der Mathematisch-Naturwissenschaftliche, (Classe Kaiserlichen Akademie der Wissenschaften), Vienna, vol. 30, p. 137-216.
GeoWhen Database (2013).- http://www.stratigraphy.org/bak/geowhen/stages/Chattian.html
J.L., P. & P. (1986).- Biostratigraphie et paléoécologie des gisements types du Stampien de la région d'Étampes (Essonne).- Géologie de la France, Orléans, vol. 1, p. 3-101 (4 Pls.).
M. (1952).- Gastropodes du Miocène moyen du Bassin de la Loire. Deuxième partie.- Mémoire de l'Institut Royal des Sciences Naturelles de Belgiques, Bruxelles, (série 2), vol. 46, p. 241-450 (15 Pls.).
M. (1954).- Pleurotomes du Miocène de la Belgique et de la Loire.- Mémoire de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, vol. 129, 75 p.
M. (1957).- Pélécypodes et Gastéropodes du Rupélien supérieur et du Chattien de la Belgique.- Mémoires de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, (série 2), vol. 137, 98 p. (6 Pls.).
M. (1973).- Révision des Gastropoda du Danien et du Montien de la Belgique.- Mémoires de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, vol. 173, 116 p.
M. & de J. de (1954).- L'Oligocène inférieur belge. In: Volume jubilaire Victor van .- Institut Royal des Sciences Naturelles de Belgique, Bruxelles, vol. 1, p. 281-438 (7 Pls.).
S., D. & C. (coords., 2011).- Moluscos marinos de Andalucía. Vol. 1.- Servicio de Publicaciones e Intercambio Científico, Universidad de Málaga, p. i-xvi & 1-342.
L. (1969).- Clefs de détermination des petites espèces de gastéropodes de l'Éocène du Bassin parisien. II : La famille des Pyramidellidae (partie 1).- Cahiers des Naturalistes, Bulletin des Naturalistes Parisiens, Paris, (n.s.), vol. 23, nº 4, p. 93-109.
L. (1970).- Clefs de détermination des petites espèces de gastéropodes de l'Éocène du Bassin parisien. III : La famille des Pyramidellidae (partie 2).- Cahiers des Naturalistes, Bulletin des Naturalistes Parisiens, Paris, (n.s.), vol. 25, nº 2, 25-36.
L. & J. (1980).- Clefs de détermination des petites espèces de gastéropodes de l'Éocène du Bassin parisien. XII : La famille des Triphoridae.- Cahiers des Naturalistes, Bulletin des Naturalistes Parisiens, Paris, (n.s.), vol. 35, nº 3, p. 41-59.
L. & J. (1981).- Clefs de détermination des petites espèces de gastéropodes de l'Éocène du Bassin parisien. XIV : La famille des Cerithiopsidae.- Cahiers des Naturalistes, Bulletin des Naturalistes Parisiens, Paris, (n.s.), vol. 36, nº 2, p. 17-38.
K. (1995, unpublished).- Revision der marinen Molluskenfauna des Unteren Meeressandes (Oligozän, Rupelium) des Mainzer Beckens.- Dissertation, München, 314 p. (64 Pls.).
K. & H. (2002).- Neues zu den Rissooidea (Mollusca, Gastropoda) aus dem Rupelium (Oligozän) des Mainzer Beckens und des Nordseebeckens.- Courier Forschungsinstitut Senckenberg, Frankfurt, vol. 237, p. 255-273.
Ø. (2013).- PAST PAleontological STatistics Version 3.- http://folk.uio.no/ohammer/past/
M. (2004).- Oligocene gastropod Faunas of the Eastern Mediterranean (Mesohellenic Trough/Greece and Esfahan-Sirjan Basin/Central Iran).- Courier Forschungsinstitut Senckenberg, Frankfurt, vol. 248, p. 93-181.
M., W.E. & F.E. (2002).- Circum-Mediterranean Oligo-Miocene biogeographic evolution - the gastropods' point of view.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 183, p. 103-133.
M., A, O., W.E., U., M. & B. (2007).- Biogeographic responses to geodynamics: A key study all around the Oligo-Miocene Tethyan Seaway.- Zoologischer Anzeiger, Berlin, vol. 246, nº 4, p. 241-256.
J. & A. (1900).- Contribution à l'étude paléontologique des faluns de la Touraine.- Actes de la Société Linnéenne de Bordeaux, t. LV, nº 2, p. 99-130; t. LV, nº 3, p. 131-194; t. LV, nº 4, p. 195-249 (3 Pls.).
A.W. (1984a).- Mollusken uit het Mioceen van Winterswijk-Miste. Een inventarisatie, met beschrijvingen en afbeeldingen van alle aangetroffen soorten.- Koninklijke Nederlandse Natuurhistorische Vereniging, Leiden, 451 p. (82 Pls.).
A.W. (1984b).- Late Oligocene molluscs from a sand-pit near Máriahalom (Hungary): A preliminary study.- Annales Universitatis Scientiarum Budapestinensis de Rolando Eötvös Nominatae Sectio Geologica, Budapest, vol. 24, p. 111-149 (4 Pls.).
R. (1978a).- Die Scaphopoden und Gastropoden des Kasseler Meeressandes von Glimmerode (Niederhessen).- Geologisches Jahrburch, Hannover, (Reihe A), Band A 41, 195 p. (7 Pls.).
R. (1978b).- Die Mollusken des Oberoligozäns (Chattium) im Nordsee-Becken, 1 - Scaphopoda, Archaeogastropoda, Mesogastropoda.- Archiv für Molluskenkunde, Frankfurt, vol. 109, nº 1-3, p. 137-227 (Pls. 9-14).
R. (1979).- Die Mollusken des Oberoligozäns (Chattium) im Nordsee-Becken. 2. Neogastropoda, Euthyneura, Cephalopoda.- Archiv für Molluskenkunde, Frankfurt, vol. 109, nº 4, p. 277-376 (5 Pls.).
R. (2004).- The family Pickworthiidae in the Middle Miocene of the Paratethys (Gastropoda: Caenogastropoda: Littorinoidea?).- Courier Forschungsinstitut und Naturmuseum Senckenberg, Frankfurt, vol. 246, p. 169-186.
E.A. (1996).- Evolutionary radiations in the Cypraeidae. In: J. (ed.), Origin and evolutionary radiation of Mollusca.- Oxford University Press, p. 211-220.
A.J. (1990).- Tempo and mode of evolution in Conidae.- Malacologia, Chicago, vol. 32, nº 1, p. 55-67.
M., K., J.H., W., W.E. & A.P. (2002).- Multivariate hierarchical analyses of Miocene mollusc assemblages of Europe: Paleogeographic, paleoecological, and biostratigraphic implications.- Geological Society of America Bulletin, Boulder, vol. 114, p. 239-256.
A. (2007).- Climate changes in the Early to Middle Miocene of the Central Paratethys and the origin of its echinoderm fauna.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 253, p. 169-207.
E.M. (1973).- Die Gastropoden aus dem Meeressand (Rupelium) des Mainzer Tertiärbeckens.- Abhandlungen des Hessischen Landesamtes für Bodenforschung, Wiesbaden, Heft 67, 170 p. (8 Pls.).
B., C.M. da & E. (2011).- The Lower Pliocene gastropods of the Huelva Sands Formations, Guadalquivir Basin, Southwestern Spain.- Palaeofocus, Mortsel, vol. 4, 90 p. (23 Pls.).
P. & R. von (1998).- Biodiversity patterns of the marine benthic fauna on the Atlantic coast of tropical Africa in relation to hydroclimatic conditions and paleogeographic events.- Acta Oecologica, Fribourg, vol. 19, nº 3, p. 309-321.
J. (1999).- Clefs de détermination des petites espèces de gastéropodes de l'Éocène du Bassin parisien. 42 - Tables des clefs I à XLI (1968-1997).- Cahiers des Naturalistes, Bulletin des Naturalistes Parisiens, Paris, (n.s.), vol. 52, nº 3-4, p. 61-103.
J. (2013).- Fossils. Virtual scientific collection of French Tertiary fossils.- Online at http://www.somali.asso.fr/fossils/index.php
J. & O. (2012).- Les fossiles de l'Éocène moyen du Cotentin. II.- La Biozone des Faluns de Hautteville-Bocage. III.- Historique des espèces originalement du Cotentin.- Cossmanniana, Leiden, (hors série), t. 5, p. 27-197.
J. & J.M. (1995).- Révision des mollusques paléogènes du bassin de Paris. II - Liste des références primaires des espèces.- Cossmanniana, Leiden, t. 3, nº 3, p. 65-132.
P. (1985).- Compléments à la malacofaune oligocène (Stampien) de Gaas (Bassin d'Aquitaine, France), 1. Mollusques saumâtres.- Mededelingen Werkgroep voor Tertiaire en Kwartaire Geologie, Rotterdam, vol. 22, nº 3, p. 125-142 (4 Pls.).
P. (1997, unpublished).- Le domaine atlantique européen au Cénozoïque moyen : Diversité et évolution des gastéropodes.- Thèse de Doctorat du MNHN, 309 p. (+ annexes : 52 p.).
P. (1998).- Nouvelles espèces de gastéropodes (Mollusca : Gastropoda) de l'Oligocène et du Miocène inférieur de l'Aquitaine (Sud-Ouest de la France).- Cossmanniana, Leiden, t. 5, nº 3-4, p. 61-102.
P. (1999).- Nouvelles espèces de gastéropodes (Mollusca : Gastropoda) de l'Oligocène et du Miocène inférieur de l'Aquitaine (Sud-Ouest de la France). Partie 2.- Cossmanniana, Leiden, t. 6, nº 1-2, 68 p. (20 Pls.).
P. (2004).- The European Tertiary Neritiliidae (Mollusca, Gastropoda, Neritopsina): indicators of tropical submarine cave environments and freshwater faunas.- Zoological Journal of the Linnean Society, London, vol. 140, p. 447-467.
P. (2011).- Nouvelles espèces de gastéropodes (Mollusca : Gastropoda) de l'Oligocène et du Miocène inférieur de l'Aquitaine (Sud-Ouest de la France). Partie 4.- Cossmanniana, Leiden, t. 13, p. 49-58 (3 Pls.).
P. (coord., 2012).- Stratotype Stampien.- Biotope, Mèze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 4, 464 p. (1 CD-Rom).
P. & D.T. III (2001).- First fossil record of the genus Pyramidelloides (Mollusca, Gastropoda, Eulimidae) discovered in the Eocene deposits of Alabama.- Comptes Rendus de l'Académie des Sciences, (Sciences de la Terre et des Planètes, Earth and Planetary Sciences), Paris, vol. 333, p. 351-356.
P. & D. (1995).- Senilia (Bivalvia : Arcidae) et Anazola (Gastropoda : Olividae) dans le Miocène d'Angola et de France, témoins d'une paléo-province Ouest-Africaine.- Haliotis, Paris, t. 24, p. 101-108.
P., J.L. & P. (2001a).- Révision des Gastropoda (Mollusca) du stratotype de l'Aquitanien (Miocène inf.) : Site de Saucats "Lariey", Gironde, France.- Cossmanniana, Leiden, (hors série), t. 3, 189 p. (37 Pls.).
P. & P. (1982).- Nouvelles espèces de mollusques de l'Oligocène (Stampien) pour les bassins de Paris et d'Aquitaine.- Archiv für Molluskenkunde, Frankfurt, vol. 112, nº 1-6, p. 165-189.
P. & P. (2012).- Le contenu paléontologique : Mollusques. In: P. (coord.), Stratotype Stampien.- Biotope, Mèze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 4, p. 239-297.
P., P., L. & R. (2001b).- Un site exceptionnel du Miocène inférieur (Aquitanien) : La "Carrière Vives" (Meilhan, Landes, France). Bilan de la campagne de fouilles de juillet - août 1991.- Cossmanniana, Leiden, t. 8, nº 1-4, p. 47-67.
P. & P. (1996).- Les Glycymerididae (Mollusca, Bivalvia) de l'Oligocène supérieur (Chattien) du bassin de l'Adour, France.- Annales de Paléontologie, Paris, (Vert.-Invert.), t. 82, nº 1, p. 3-25.
A. & A.M. (1970).- Présence du genre Melongena dans les faluns de Saint-Étienne-d'Orthe.- Bulletin de la Société de Borda, Dax, 2 p.
A. & A.M. (1975).- Un Cassidaria nouveau du Bassin de l'Adour.- Bulletin de la Société de Borda, Dax, 3 p.
A. & A.M. (1971).- Les Turbo tertiaires du Bassin d'Aquitaine. Bulletin de la Société de Borda, Dax, 13 p.
A. & A.M. (1974).- Le gisement de Quillacq à Saint-Paul-lès-Dax.- Bulletin de la Société de Borda, Dax, 4 p.
R., C., C. & R. (2008).- The Molluscan fauna of the Borgloon Formation in Belgium (Rupelian, Early Oligocene).- Palaeontos, Totnes, vol. 12, 100 p.
P.A. (1988).- Late Miocene deep-water Mollusca from the Stillwater Mudstone at Greymouth, Westland, New Zealand: Paleoecology and systematics.- New Zealand Geological Survey Paleontological Bulletin, Lower Hutt, vol. 55, 120 p. (13 Pls.).
P.A. (2009).- Cenozoic Mollusca. In: D.P. (ed.), The New Zealand inventory of biodiversity: a Species 2000 symposium review. Vol. 1.- Canterbury University Press, Christchurch, p. 180-190 + 231-253.
J.E., W., L., B., J.P., R. & B. (2000).- Late Rupelian (32 - 29 Ma). In: J. et al. (eds.), Atlas Peri-Tethys, palaeogeographical maps.- CCGM/CGMW, Paris, p. 171-178.
H., F. & G. (2012).- Die Molluskenfauna (Hemmoorium, Untermiozän) aus der Kiesgrube Krinke bei Werder (Nordwest-Niedersachen).- Palaeofocus, Mortsel, vol. 3, 155 p. (46 Pls.).
D., E. & B. (2002).- Progrès récents dans la connaissance des gisements cénozoïques en Aquitaine méridionale (Chalosse, Béarn et Bas-Adour; SW France).- Revue de Micropaléontologie, nº spécial "Aquitaine", Paris, t. 45, nº 3, p. 169-194.
G.S. (1994).- Geological time scale (1994).- Comptes Rendus de l'Académie des Sciences, Paris, (série II), t. 238, p. 59-71.
J.M., D. & J.C. (2000).- La faune danienne de Vigny (Val d'Oise, France), importance pour l'étude de la diversification des mollusques au début du Tertiaire.- Comptes Rendus de l'Académie des Sciences, (Sciences de la Terre et des Planètes), Paris, (série II a), t. 330, nº 12, p. 867-873.
J.M. (2008).- Le contenu paléontologique du Lutétien du Bassin de Paris (CD-ROM), Mollusca. In: D. (coord.), Stratotype Stampien.- Biotope, Mèze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 1, 288 p.
A. (1925-1935).- Conchologie néogénique de l'Aquitaine.- Actes de la Société Linnéenne de Bordeaux, t. LXXVII, nº 2, p. 51-194 (1925); t. LXXVIII, p. 199-256 (1927); t. LXXIX, p. 5-264 (1928); t. LXXXII, nº 2, p. 73-126 (1931); t. LXXXIII, p. 5-116 (1931); t. LXXXIV, nº 1, p. 5-128 (1932); t. LXXXIV, nº 2, p. 129-288, (1933); t. LXXXV, nº 1, p. 5-71 (1933); t. LXXXVI, nº 2, p. 257-352 (1935).
A. (1938).- Les mollusques testacés univalves des dépôts helvétiens du Bassin Ligérien. Catalogue critique, descriptif et illustré.- Actes de la Société Linnéenne de Bordeaux, t. LXXXIX, 300 p. (5 Pls.).
W.E., M. & O. (2007).- Miocene Central Paratethys stratigraphy - current status and future directions.- Stratigraphy, vol. 4, nº 2-3, p. 151-168.
J.C. (1981).- Late Cretaceous to Late Eocene palaeogeographic evolution of Southwest Europe.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 36, p. 263-320.
J.C. & P. (2012).- Le contenu géologique : les dépôts du Stampien et leur interprétation du point de vue environnements. In: P. (coord.), Stratotype Stampien.- Biotope, Mèze; Patrimoine géologique, Muséum national d'Histoire naturelle, Paris, vol. 4, p. 75-124.
F.D. (1978).- Lessepsian migration - the influx of Red Sea biota into the Mediterranean Sea by way of the Suez Canal.- Springer, Berlin, 228 p.
D.R. (1994).- The Eocene-Oligocene transition. Paradise lost. In: D.J. & R.K. (eds.), Critical moments in paleobiology and earth history series.- Columbia University Press, 291 p.
F. & F.F. (1984).- Neogene Paratethys, Mediterranean and Indo-Pacific seaways. Implications for the paleobiogeography of marine and terrestrial biotas. In: P.J. (ed.), Fossils and climate.- Wiley, Chichester, p. 171-200.
B. & M. (2014).- The making of the Mediterranean Molluscan biodiversity. In: S. & Z. (eds.), The Mediterranean Sea: Its history and present challenges.- Springer Science+Business Media, Dordrecht p. 285-306.
A.M. (1969).- Découverte d'une nouvelle espèce de gastropode dans les faluns de Saint-Etienne-d'Orthe (Landes).- Bulletin de la Société de Borda, Dax, 3 p.
A.M. & B. (1978).- Le gisement d'Estoti à Saint-Paul-lès-Dax (Landes).- Bulletin de la Société de Borda, Dax, nº 369, p. 103-118; nº 370, p. 237-245.
K.I. (1985).- Two new Upper Oligocene gastropods from the Nord Sea Basin.- Bulletin of the geological Society of Denmark, Copenhagen, 34, p. 199-204.
K.I. & C. (1987).- A Late Oligocene (Chattian B) mollusc fauna from the clay-pit of Galten Brickworks at Norre Vissing, Jylland, Denmark.- Mededelingen Werkgroep voor Tertiaire en Kwartaire Geologie, Rotterdam, vol. 24, nº 3, p. 193-224 (2 Pls.).
K.I. & C. (1990).- A Late Oligocene (Chattian B) Molluscan Fauna from the coastal cliff at Mogenstrup, north of Skive, Jutland, Denmark.- Mededelingen Werkgroep voor Tertiaire en Kwartaire Geologie, Rotterdam, vol. 27, nº 2-3, p. 39-81 (3 Pls.).
C.M. da & B.M. (2007).- On the presence of Spiricella unguiculus & , 1828 (Gastropoda, Notaspidea) in the European Pliocene: Filling the geological gap.- The Veliger, Santa Barbara, vol. 49, nº 1, p. 19-26.
A. & P. (2000).- Opisthobranch Molluscs from the Tertiary of the Aquitaine Basin (South-Western France), with descriptions of seven new species and a new genus.- Palaeontology, London, vol. 43, nº 3, p. 457-479.
A.M. (1967a).- Les gisements de l'Oligocène marin en Aquitaine.- Bulletin de l'Institut Géologique du Bassin d'Aquitaine, Bordeaux, t. 4, p. 196-210.
A.M. (1967b).- Historique des travaux relatifs à la faune malacologique de l'Oligocène aquitain.- Bulletin de l'Institut Géologique du Bassin d'Aquitaine, Bordeaux, t. 4, p. 181-190.
A.M. (1968).- Quelques espèces nouvelles de mollusques de l'Oligocène aquitain.- Bulletin de l'Institut Géologique du Bassin d'Aquitaine, Bordeaux, t. 4, p. 196-209.
G.J. (2011).- The tropical history and future of the Mediterranean biota and the West African enigma.- Journal of Journal of Biogeography, Oxford, vol. 39, nº 1, p. 1-11.
S.T. & F.T. (2008).- Did tectonic activity stimulate Indo-West-pacific Oligo-Miocene speciation in the Indo-West Pacific?.- Evolution, vol. 62, nº 7, p. 1-15 [doi:10.1371/journal.pone.0011842].
S.P. (1868).- A manual of the Mollusca: A treatise of recent and fossil shells. Second edition.- Virtue and Co., London, 543 p. [with "An appendix of recent and fossil conchological discoveries to the Present time" by R. , 86 p. ]. Online at http://www.biodiversityheritagelibrary.org/item/101361#page/7/mode/1up
A., S., G. & J. (2003).- CIESM Atlas of Exotic Species in the Mediterranean. In: F. (ed.), Molluscs. Vol. 3.- CIESM Publishers, Monaco, 376 p.
Source of data and main literature compiled
The major part of the datasets analyzed in this study were assembled through field work by the author and collaborators. The overall sampling effort may be gauged from the 113 person-days allocated to fieldwork in an Aquitaine quarry at Meilhan ( et al., 2001b). The samples were sieved systematically, generally in fresh water. Items larger than 3-4 mm were sorted in the field while the finer fractions (3-0.5 mm) were sorted in the laboratory with the aid of a dissecting microscope. Considerable time was spent in sorting the finer fractions. For basins for which limited data have been collected by the author, the data are augmented from a critical compilation from the literature. "Personal collecting data" denotes exclusive use of personal data, "From literature and personal data" indicates both sources are used, "From literature" denotes exclusive use of bibliographic datasets. "Literature" contains the main publications for the faunas indicated.
Lower Oligocene:
Personal collecting data: Aquitaine Basin (France), Paris Basin (France).
Literature: (1892, 1893), (1967a, 1967b, 1968), et al. (1986), (1985, 2011), & (1982).
From literature and personal data: Belgian, Mainz Basin (Germany) [ & de, 1954; , 1973; , 1995, & , 2002; et al., 2008].
Upper Oligocene:
Personal collecting data: Aquitaine Basin (Adour Basin).
Literature: & (1974), (1969), & (1978), (1998, 1999, 2004).
From literature and personal data: Chattian of Germany and Denmark; Egerian (Hungary)
[, 1957; , 1978a, 1978b, 1979; , 1985; & , 1987, 1990; , 1973; , 1984b]
Lower Miocene:
Personal collecting data: Aquitanian (Aquitaine Basin), Lower Burdigalian (Aquitaine Basin), Upper Burdigalian (Aquitaine Basin).
From literature: & (1909-1924), (1925-1932), et al. (2001a, 2001b), (2011).
Middle Miocene:
Personal collecting data: Langhian (Aquitaine Basin); Langhian (Loire Basin); Serravallian (Aquitaine Basin).
From literature: Langhian (Loire Basin) [, 1938; & , 1900; , 1952, 1954]; North Sea Basin, Hemmoorian (Lower/Middle Miocene: Miste, Netherlands; Werder, Germany) [, 1984a; et al., 2010]
Upper Miocene:
Personal collecting data: Redonian (Loire Basin, France).
From literature: (1964).
Table 3: Datamatrix of genera used for multivariate analysis (correspondence analysis). Data transformed into a presence-absence table, where each cell has a 0 if the genus is absent and a 1 if the genus is present:
Lower Oligocene: [LOBP] Basin of Paris, [LOAQ] Aquitaine Basin, [LONE] North Sea basin, [LOMA] Mainz Basin, [Rennes Basin]. Upper Oligocene: [UOA] Aquitaine (Adour) Basin, [UPN] North Sea basin, [UPPA] Paratethys; Lower Miocene: [LMAA] Aquitanian of Aquitaine Basin, [LMLB] Lower Burdigalian of Aquitaine Basin, Upper Burdigalian of Aquitaine basin [LMUB]; Middle Miocene: [MMLT]: Loire Basin, [MMLA] Langhian of Aquitaine basin, [MMSA] Serravalian of Aquitaine Basin, [MMSA] Middle Miocene of North Sea basin (Miste); Upper Miocene [MLB] (Messinian) of Loire Basin.