◄ Carnets Geol. 15 (8) ►
![]()
Contents
[1. Introduction]
[2. Geological setting and studied sections]
[3. Material and methods]
[4. Ammonite succession and
zonation]
[5. Conclusions] and ...
[Bibliographic references]
Departamento de Estratigrafía y Paleontología, Facultad de
Ciencias, Universidad de Granada, Campus de Fuentenueva s/n. E-18002 Granada
(Spain)
Departamento de Estratigrafía y Paleontología, Facultad de
Ciencias, Universidad de Granada, Campus de Fuentenueva s/n. E-18002 Granada
(Spain)
Published online in final form (pdf) on June 11, 2015
[Editor: Bruno ;
language editor: Stephen ]
![]()
A new zonation for the lower Valanginian in the Betic Cordillera is presented. It is based on the study of 16 sections located near Caravaca and Cehegín (Region of Murcia). From bottom to top, the following interval zones, defined by the first appearance of the index-species, are distinguished:
The assemblages characterizing each of these biozones can be recognized throughout the Mediterranean region.
Lower Valanginian; Cretaceous; ammonite; biostratigraphy; Spain; Mediterranean region.
M. & J.M. (2015).- Lower Valanginian ammonite biostratigraphy in the Subbetic Domain (Betic Cordillera, southeastern Spain).- Carnets Géol., Madrid, vol. 15, nº 8, p. 71-88.
Biostratigraphie par ammonites du Valanginien inférieur du Domaine Subbétique (Cordillères Bétiques, SE de l'Espagne).- Une nouvelle zonation pour le Valanginien inférieur de la Cordillère Bétique est présentée. Elle est fondée sur l'étude de 16 coupes localisées près de Caravaca et de Cehegín (région de Murcie). De bas en haut, définies par la première apparition de l'espèce-index, les zones d'intervalle suivantes ont été reconnues :
Les associations qui caractérisent chacune de ces unités biostratigraphiques peuvent être reconnues sur l'ensemble de la région méditerranéenne.
Valanginien inférieur ; Crétacé ; ammonites ; biostratigraphie ; Espagne ; région méditerranéenne.
Knowledge of the lower Valanginian ammonite biostratigraphy in the Mediterranean region has evolved little during the last two decades. Indeed, the current standard zonation for this interval until very recently (et al., 2003; et al., 2011) was based largely on the proposals of & (1995a, 1995b), (1996), and & (1999) in SE France. These proposals, however, differed significantly from the zonal scheme developed, around the same time, by (1995) and & (1995). These differences involved not only the bioevents chosen to define the biostratigraphic units, but also the stratigraphic range and even the conception of some key species ( & , 1999).
In order to resolve these discrepancies, we undertook a detailed analysis of the ammonite stratigraphic distribution throughout the lower Valanginian. To this end, we carried out a systematic sampling of 16 sections in the eastern Subbetic domain (Betic Cordillera, southeastern Spain), some of them previously not studied. This enabled us to accurately identify the sequence of ammonite bioevents and propose a more precise zonation for this interval in the Betic Cordillera. An outline of these results was previously presented ( & , 2013a, 2013b), and some of our proposals were accepted by the attendants to the 5th Meeting of the " Group" (Ankara, 31 August 2013) to be included in the last version of the standard Mediterranean zonation ( et al., 2014). In this paper, we develop this zonal scheme giving a detailed account of the bioevents that define and characterize the different units that compose it and discussing its correlation with the schemes previously proposed by other authors.
All the studied sections belong to the Subbetic Zone, a tectonic domain which corresponds roughly to the pelagic realm of the southern paleomargin of Iberia during the Alpine cycle (Mesozoic and Paleogene). The lithologic successions are made of marl-limestone rhythmites of the Miravetes Formation (, 1969), in which ammonites are, by far, the main components of the macrofauna. Some slumped intervals are present in the sections of the Sierra de Quípar-Peña Rubia-Mai Valera area, which behaved as a relative topographic high during the Jurassic and the Early Cretaceous (, 1993; , 2004).
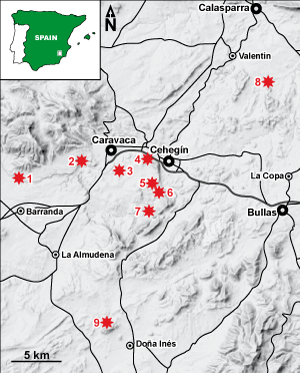
The sections sampled, all of them located near Caravaca and Cehegín (region of Murcia, SE
Spain), are the following
(Fig. 1 ![]() ):
):
The sections Y.CL2,
Y.Qp2, Y.B, Y.T, and Y.P have been taken as reference sections and
are represented, respectively, in Figs. 2 ![]() - 3
- 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() - 6
- 6 ![]() .
.
|
|
|
Figure 1: Geographic location of the sections studied. (1) Las Oícas (section Y.O); (2) Miravetes (section Y.Mv); (3) Prado Borda (sections Y.B, Y.B1 and Y.T); (4) Mai Valera (section Y.P); (5) Puente de la Virgen (section Y.N); (6) Cañada Luenga (sections M.CL, Y.CL2 and Y.Qp2); (7) Barranco del Garranchal (sections Y.G and Y.Q2); (8) Cerro Cambrones (section Y.V); (9) Tornajo (sections Y.J1, Y.J2 and Y.J3). |
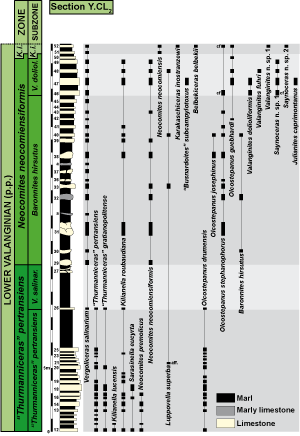
Figure 2: Distribution of the most significant ammonite species in section Y.CL2. |
|
|
|
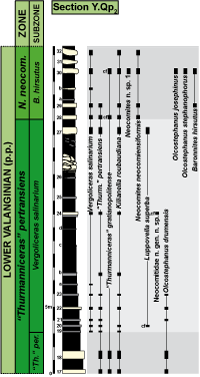
Figure 3: Distribution of the most significant ammonite species in section Y.Qp2. |
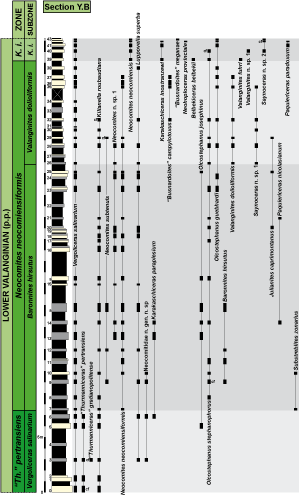
Figure 4: Distribution of the most significant ammonite species in section Y.B. |
|
|
|
|
Figure 5: Distribution of the most significant ammonite species in section Y.T. |
Figure 6: Distribution of the most significant ammonite species in section Y.P. |
For the present study more than 10,000 ammonites were collected by bed-by-bed sampling from the interval studied in these sections. The fieldwork was carried out by the authors between 1980 and 2012. Most of the specimens are preserved as calcareous internal casts and come from the limestone beds, but some limonitized forms were found in the marly interbeds. Although many of the ammonites are flattened by compaction, their preservation is good enough to allow most of them to be identified to the species level. The assemblages are almost exclusively composed of Mediterranean taxa (Haploceratidae, Neocomitidae, Olcostephanidae, Oosterellidae, Bochianitidae, Phylloceratidae, and Lytoceratidae) and only very few specimens of the typically Boreal Platylenticeratinae were recorded.
The ammonite material is housed at the paleontological collections of the University of Granada. The specimens studied have been numbered following the usual method in these collections. Thus, each specimen is identified with a reference code composed of four elements separated by periods (e.g., Y.T.19.32). The first letter refers to the collector(s) (in this case, Y = and ), the second and third elements indicate respectively the section (T) and the bed (19) where the specimen was collected. The last element is a series number (32 in the example) for each specimen coming from the same bed.
With a few exceptions, the systematic arrangement proposed by (2005, 2006) and et al. (2009) has been followed in this paper. Nevertheless, some generic names will be cited in quotation marks (namely "Thurmanniceras" and "Busnardoites") indicating that the assignment of the species to that genus is provisional and requires a thorough review. Furthermore, the interpretation of some problematic species (such as Neocomites neocomiensiformis or Karakaschiceras biassalense) will also be discussed below.
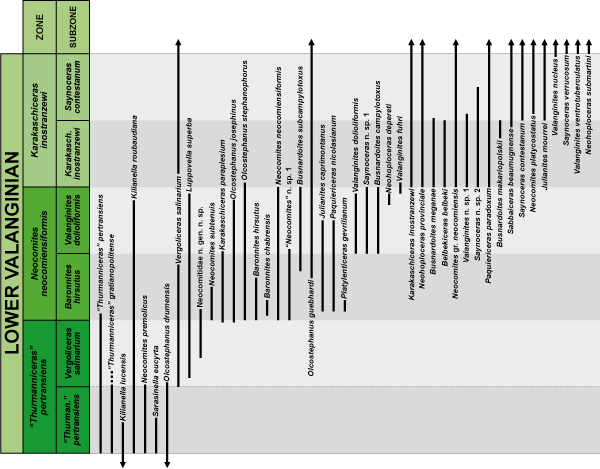
The analysis of
the stratigraphic distribution of the species identified in the studied sections
has enabled us to distinguish three interval biozones for the lower Valanginian
in the Betic Cordillera (Figs. 7 ![]() - 8
- 8 ![]() ).
These biozones are, from bottom to top, the "Thurmanniceras" pertransiens Zone, the Neocomites neocomiensiformis Zone, and the Karakaschiceras inostranzewi Zone, all of them defined at their base
by the FAD of the index species. Each of these zones has been, in turn,
subdivided into two subzones: a lower one whose base obviously coincides with
that of the zone, and an upper one defined at their base by the FAD of its
index-species.
).
These biozones are, from bottom to top, the "Thurmanniceras" pertransiens Zone, the Neocomites neocomiensiformis Zone, and the Karakaschiceras inostranzewi Zone, all of them defined at their base
by the FAD of the index species. Each of these zones has been, in turn,
subdivided into two subzones: a lower one whose base obviously coincides with
that of the zone, and an upper one defined at their base by the FAD of its
index-species.
Click on thumbnail to enlarge the image.
Figure 7: Stratigraphic range of the most significant ammonite species in the framework of the zonation proposed for the lower Valanginian in the Betic Cordillera.
Click on thumbnail to enlarge the image.
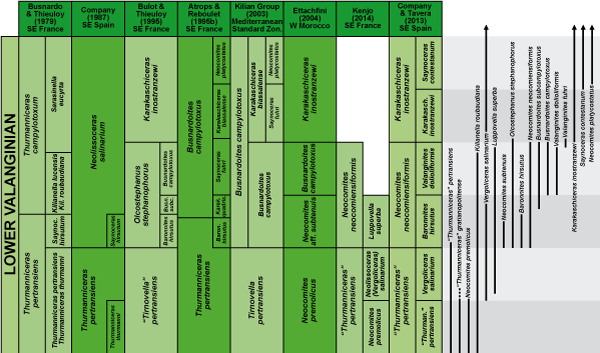
Figure 8: Correlation of the zonation proposed for the lower Valanginian in the Betic Cordillera with other zonal schemes.
Definition and characterization
The lower boundary of this zone is defined by the FAD of "Thurmanniceras" pertransiens, which, as generally admitted, closely correlates with the FAD of Calpionellites darderi ( & , 1979; et al., 1994; et al., 1996; et al., 2000; et al., 2011; but see , 2014, for a different interpretation). It is well known that the FAD of Calpionellites darderi currently marks the base of the Valanginian stage ( et al., 1996; & , 2012).
The base of this zone coincides with a conspicuous faunal renewal. Species that characterized the uppermost Berriasian beds, like Berriasella calisto, Fauriella boissieri, Tirnovella alpillensis, "Thurmanniceras" otopeta, and Erdenella paquieri, disappear and are replaced by new forms such as "Thurmanniceras" pertransiens, "Thurmanniceras" gratianopolitense (), Neocomites premolicus , Kilianella roubaudiana (), and Sarasinella eucyrta (). They are accompanied by several taxa inherited from lower levels such as Kilianella lucensis (), Olcostephanus drumensis , the last representatives of the genera Kilianiceras and Protancyloceras and some species of broad stratigraphic distribution. The first specimens of Luppovella superba () and Vergoliceras salinarium () are recorded in the upper part of this zone.
Subdivision
The significant differences between the assemblages from the lower and upper parts of this zone allow it to be subdivided into two subzones: a lower "Thurmanniceras" pertransiens Subzone and an upper Vergoliceras salinarium Subzone. The "Th." pertransiens Subzone is characterized by an abundant and highly diverse fauna dominated by the index species. "Thurmanniceras" gratianopolitense, Kilianella roubaudiana, Neocomites premolicus and Olcostephanus drumensis are also frequent, while Kilianella lucensis and Sarasinella eucyrta are less common and seem to disappear in this interval.
The FAD of Vergoliceras salinarium, here considered a senior synonym of Vergoliceras extracornutum (), marks the base of the upper subzone. In contrast to the underlying subzone, the assemblage of this unit is little diversified and overwhelmingly dominated by the index species. "Thurmanniceras" pertransiens occurs, although scarcely, throughout this subzone, whereas "Thurmanniceras" gratianopolitense, Neocomites premolicus, and Olcostephanus drumensis become extinct in its lower part. The first, rare specimens of Luppovella superba have been recorded in this interval accompanied by other not yet described neocomitid taxa.
Discussion
This unit was used for the first time by & (1968) as a subzone of the classic Kilianella roubaudiana Zone, and elevated to the rank of zone by & (1973a, 1973b). In its original sense, it included the 'horizon à Kilianella aff. pexiptycha et Thurmannites aff. pertransiens' of (1939), equivalent to the current uppermost Berriasian "Thurmanniceras" otopeta Subzone. Some years later & (1979a) modified the initial conception by separating the "Th." otopeta beds as an independent zone. Nevertheless, the first formal definition of the "Thurmanniceras" pertransiens Zone was given by (1987), who drew its base at the FAD of the index species. It is in this sense that this unit has been employed by most subsequent authors.
"Thurmanniceras" pertransiens is a well-known species that has been reported throughout the Mediterranean region. It is frequent in basinal facies, especially in the lower part of the zone, but becomes rare or even absent in outer shelf paleoenvironments, where Neocomites premolicus and/or "Thurmanniceras" gratianopolitense largely dominate the assemblages. This led (2004) to choose N. premolicus as index of the basal zone of the Valanginian in the western High Atlas. This species has also been employed by (2014) to characterize the lower subzone of the "Thurmanniceras" pertransiens Zone in southeastern France.
The FAD of Vergoliceras salinarium marks the base of the upper subzone of this zone, coinciding with the start of a marl-dominated interval in the lithologic succession. According to the data of (1994) the FO of the calcareous nannofossil species Calcicalathina oblongata, takes place at the same level. (2014) has also recognized this subzone in southeastern France. The index species, present all over the Mediterranean area, has a broad stratigraphic range, reaching the upper Valanginian. It has also been occasionally reported from the Berriasian ( & , 1976; & , 1995; , 1999; & , 2004), but in the specimens figured by these authors the keel is only present on the adapertural part of the body chamber, suggesting that they belong to a different species.
Definition and characterization
The FAD of Neocomites neocomiensiformis () marks the base of this zone. A new sharp faunal change takes place around this level, with the extinction of "Thurmanniceras" pertransiens and the appearance, besides the index species, of Baronnites hirsutus ( & ), Baronnites chabrensis , & , Olcostephanus stephanophorus (), Olcostephanus josephinus (), Karakaschiceras paraplesium (), and Neocomites subtenuis . Other species, such as "Busnardoites" subcampylotoxus , "Busnardoites" campylotoxus (), Neohoploceras depereti , Olcostephanus guebhardi , Valanginites dolioliformis (), Valanginites fuhri (, & ), Paquiericeras nicolasianum (), and Julianites caprimontanus () have their first records in higher levels within this stratigraphic interval. In addition, some undescribed neocomitid and olcostephanid forms have also been recorded, and the holdover species Kilianella roubaudiana, Luppovella superba and Vergoliceras salinarium are still common in the Neocomites neocomiensiformis Zone.
Subdivision
The Neocomites neocomiensiformis Zone can be subdivided into two subzones: the Baronnites hirsutus Subzone below, and the Valanginites dolioliformis Subzone above. The species Baronnites hirsutus, Baronnites chabrensis and Neocomites subtenuis, as well as a few specimens of Platylenticeras gevrilianum (), have been found to be restricted to the lower part of the B. hirsutus Subzone. Paquiericeras nicolasianum, Julianites caprimontanus, "Busnardoites" subcampylotoxus, and Olcostephanus guebhardi have been first recorded in higher levels of this subzone.
The lower boundary of the Valanginites dolioliformis Subzone is identified by the FAD of the index species, accompanied by "Busnardoites" campylotoxus and Saynoceras n. sp. 1. Some specimens of Neohoploceras depereti have been recorded from the upper part of this stratigraphic interval, whereas Valanginites fuhri characterizes a distinctive interval at its top and the base of the overlying Karakaschiceras inostranzewi Zone. Kilianella roubaudiana, Luppovella superba, and most of the species that had appeared in the Neocomites neocomiensiformis Zone become extinct throughout this subzone.
Discussion
& (2013a, b) suggested the use of Neocomites neocomiensiformis as index species for the middle zone of the lower Valanginian. Their proposal was accepted by the Kilian Group and incorporated into the current version of the standard Mediterranean zonation ( et al., 2014).
The species Neocomites neocomiensiformis has usually been misinterpreted (see synonymy list in , 2005). Indeed, most authors have given this name to several late Valanginian and early Hauterivian forms. However, as pointed out by (1996) and et al. (2003), the ammonite assemblage described by (1902) and revised by (1975) from the Upper Teschen Shales clearly corresponds to the early Valanginian. Despite it, this species has rarely been properly reported from this age (, 1995, Pl. 1, fig. 14; et al., 2010, Fig. 5A, 5B; , 2010, Pl. 4, fig. 4; , 2014, Pl. 1, fig. 5, Pl. 2, figs. 1-3), and its representatives have been erroneously attributed to "Busnardoites" campylotoxus (, 1987, Pl. 11, figs. 12-13; , 1996, Pl. 3, figs. 1-2; , 2004, Pl. 16, fig. 8), to "Busnardoites" roberti (, 1995, Pl. 11, figs. 1-2), to Neocomites neocomiensis (, 1901, Pl. 3, fig., 6; et al., 1981, Pl. 2, fig. 2), or to "Thurmanniceras" pertransiens (, 1993, Pl. 8, figs. D-E; et al., 1997, Pl. 3, fig. 11).
This misinterpretation of N. neocomiensiformis underlies the widespread use of "Busnardoites" campylotoxus to characterize the entire middle and upper parts of the lower Valanginian (, 1971; , 1973; & , 1979a, 1979b; , 1982; & , 1990; & , 1995a; et al., 2003). By contrast, several authors (, 1995; & , 1995; , 2001, 2003; , 2004) have emphasized that true "B." campylotoxus has a much shorter stratigraphic range, limited to the interval corresponding to our Valanginites dolioliformis Subzone. Our observations (see above) confirm this last point of view and, hence, we consider N. neocomiensiformis to be a more appropriate index for the middle part of the lower Valanginian. Its total range entirely covers this stratigraphic interval, and its known paleogeographic distribution spreads over a large part of the Mediterranean region, from Morocco to Romania (see references above).
The Neocomites neocomiensiformis Zone would be equivalent to the Olcostephanus stephanophorus Zone proposed by (1995) for the middle part of the lower Valanginian in SE France, and originally conceived as the interval between the FAD of O. stephanophorus, which coincides with the FAD of N. neocomiensiformis, and the base of the Karakaschiceras inostranzewi Zone. There are, however, two main reasons that have led us to choose N. neocomiensiformis, instead of maintaining O. stephanophorus, as index for this zone. First, O. stephanophorus has a more restricted geographic distribution, being known only from Spain, France, and Hungary (, 2005; et al., 2010), since the specimen from Pakistan figured by & (1993, Fig. 3n, 3o) does not appear to belong to this species. Second, the markedly cadicone morphology of this species favors a strong dorsoventral crushing by diagenetic compaction in marly sediments (dominant in our sections), which considerably complicates its identification.
The Baronnites hirsutus Subzone was introduced by & (1979a, 1979b), and subsequently used as a more or less formal biohorizon by (1987), (1995), & (1995), and (1996). For all these authors, the extension of this unit nearly coincided with the total range of the index species. As defined in the present paper, however, it occupies a broader stratigraphic extension corresponding to both the Baronnites hirsutus and the Busnardoites subcampylotoxus Horizons of the zonal scheme proposed by (1995). In our sections, the first specimens of "Busnardoites" subcampylotoxus have, in fact, been recorded shortly above the extinction of B. hirsutus. However, "B." subcampylotoxus is a rare species, especially in the lower part of its stratigraphic range, as most of the specimens collected by us come from the Valanginites dolioliformis Subzone. For his part, (1995) mentioned the presence of this species in a few particular beds in two platform margin sections (Carajuan and Preynes), but reported only some uncertain juvenile specimens from the Vocontian Basin sections. As can be seen, the records of "B." subcampylotoxus are scarce and scattered, hindering the accurate estimation of its real stratigraphic range, and therefore it does not seem, for the moment at least, advisable to use this species to characterize any stratigraphic unit.
& (1995a) defined a Karakaschiceras quadristrangulatum Horizon at the base of their Busnardoites campylotoxus Zone, and overlying the Baronnites hirsutus Horizon. In our opinion, the specimens attributed to K. quadristrangulatum by (1996, Pl. 1, figs. 11-12) are typical morphotypes of "Busnardoites" subcampylotoxus. Consequently, the K. quadristrangulatum Horizon would be equivalent to the Busnardoites subcampylotoxus Horizon of (1995) and to the upper part of our Baronnites hirsutus Subzone.
The species Baronnites hirsutus has hitherto been found only in France and Spain (see references in , 2005). Nevertheless, the presence of Neocomites subtenuis in the Baronnites hirsutus Subzone allows the direct correlation of this unit with the Neocomites aff. subtenuis Zone of the zonal scheme established by (2004) in Morocco. The Baronnites hirsutus Subzone would also equate with the Luppovella superba Subzone proposed by (2014) for the lower part of the Neocomites neocomiensiformis Zone. Although L. superba has a broader geographic distribution than B. hirsutus, we consider that the choice of L. superba to characterize this stratigraphic interval is not appropriate, since this species is rare in basin environments ( himself only reported four fragmentary specimens from the Vergol section) and, moreover, it is already present in the upper part of the "Thurmanniceras" pertransiens Zone.
The Valanginites dolioliformis Subzone was erected by & (2013a, b). We consider Valanginites paludensis to be a junior synonym of V. dolioliformis. This species has been reported from Morocco, Spain and France. Our V. dolioliformis Subzone nearly equates with the Busnardoites campylotoxus Horizon as defined by (1995) (and later used by & , 1995; , 2001, 2003, and , 2004). In fact, the FAD of true "B." campylotoxus, the event employed by (1995) to draw the base of his B. campylotoxus Horizon, practically coincides with that of V. dolioliformis in our sections (see also , 1995; , 2001, and , 2004). However, we prefer to use V. dolioliformis rather than "B." campylotoxus as index for this unit because V. dolioliformis is much more frequent in our region, and because "B." campylotoxus has been previously used as index species for several biostratigraphic units with a quite different sense, which is explicitly against the recommendations of the International Stratigraphic Guide.
&
(1995a) distinguished, in the middle part of their Busnardoites campylotoxus Zone, a Saynoceras fuhri Horizon, the extent of which would coincide with
the total range of the index species. Nevertheless, as pointed out by
(1996), they included in S. fuhri not
only the adult calcareous specimens that matched the original description of the
species and that came from two beds situated just below the 'Plaquettes
Rousses' in the La Charce section, but also a handful of strongly
bituberculate, pyritized nuclei collected in the Vergol section. Thanks to the
kindness of our colleague Stéphane , we have had the opportunity
to examine resin casts of some of these bituberculate nuclei. In fact, they are quite different from the inner stages of true Valanginites
fuhri, which only have umbilical tubercles and fasciculate ribs (hence
justifying the transfer of this last species to the genus Valanginites).
On the contrary, they are identical to the forms we have here called Saynoceras
n. sp. 1, an undescribed species, similar to Saynoceras
verrucosum (), that occurs throughout the V. dolioliformis Subzone (see
above and Fig. 7 ![]() ). Therefore, the S. fuhri Horizon of &
(1995a)
would correlate almost exactly with our V.
dolioliformis Subzone.
). Therefore, the S. fuhri Horizon of &
(1995a)
would correlate almost exactly with our V.
dolioliformis Subzone.
Definition and characterization
This zone occupies the stratigraphic interval between the FAD of Karakaschiceras inostranzewi () and the FAD of Saynoceras verrucosum (). A distinct turnover in the ammonite fauna also occurs around the base of this zone. As mentioned above, most of the species present in the underlying Neocomites neocomiensiformis Zone disappear in its upper part, and only a few survive in the K. inostranzewi Zone. Among them, Valanginites fuhri becomes extinct at the very base of the zone, and Olcostephanus stephanophorus, somewhat higher. By contrast, Olcostephanus guebhardi and Vergoliceras salinarium are still frequent throughout this zone and pass into the upper Valanginian.
Together with the zonal index, other species that appear at the base of the Karakaschiceras inostranzewi Zone are Neocomites gr. neocomiensis ()-teschenensis (), Neohoploceras provinciale (), "Busnardoites" meganae , Belbekiceras belbekii , Paquiericeras paradoxum , Valanginites n. sp. 1, and Saynoceras n. sp. 2. Furthermore, Neocomites platycostatus , Sabbaiceras beaumugnense (), "Busnardoites" makariopolskii , Julianites mourrei (), and Saynoceras contestanum have been recorded in higher levels within this zone.
(1995) and et al. (1995) mentioned the co-occurrence of "Busnardoites" campylotoxus and Valanginites paludensis (= V. dolioliformis) with Karakaschiceras inostranzewi at the top of formation 7 in some North-Provençal Platform sections. However, we have never found "B." campylotoxus or V. dolioliformis in the K. inostranzewi Zone. In Morocco, (2001, 2003) and (2004) did not report that association either, except in some strongly condensed sections (, 2004, Figs. 20-21). It can be assumed, therefore, that a certain degree of condensation must affect the so called 'bancs à Karakaschiceras' in the Arc of Castellane.
Subdivision
Two subzones can be distinguished within this zone. The lower Karakaschiceras inostranzewi Subzone is characterized by the association of the index species with "Busnardoites" meganae and Belbekiceras belbekii. A few specimens attributable to "Busnardoites" makariopolskii have also been recorded in the upper part of this unit.
No species has been found to be restricted to the upper Saynoceras contestanum Subzone. The index species, together with Neocomites platycostatus, Sabbaiceras beaumugnense and Julianites mourrei appear at the base of this subzone and range into the upper Valanginian. Valanginites n. sp. 1 and Saynoceras n. sp. 2, which are already present in the previous subzone, have their last records in this interval.
Discussion
The Karakaschiceras inostranzewi Zone was introduced by (1995) to replace some previous informally used biostratigraphic units, such as the Sarasinella eucyrta Subzone of & (1979b) and the Karakaschiceras biassalense Horizon of et al. (1993) and & (1993). Although (1995) convincingly justified the choice of K. inostranzewi as index of this unit, & (1995a) had already retained K. biassalense to characterize the uppermost unit of their lower Valanginian biostratigraphic scheme. This option was subsequently followed by (2001, 2003) and (2004), and integrated in the standard zonation proposed by et al. (2003). We agree with (1995) in considering K. biassalense unsuitable to characterize this stratigraphic interval. Actually, we have collected no specimen strictly attributable to K. biassalense from these beds. Most of our specimens clearly belong to K. inostranzewi, and even the more finely ribbed forms, which superficially resemble K. biassalense, do not show the very high whorl expansion rate that is characteristic of this species. The same can be said of most of the specimens coming from this level and assigned in the literature to K. biassalense (e.g., & , 2004, Pl. 1, fig. 4; , 2001, Pl. 27, figs. 5-7, Pl. 28, fig. 1; , 2004, Pl. 32, figs. 1-5). On the contrary, we have collected specimens very similar to the holotype of K. biassalense in the upper part of the Saynoceras verrucosum Zone (see also the specimens coming from the same interval figured as K. pronecostatum by , 1995, Pl. 13, figs. 1-5). In addition, it should be noted that, whereas the holotype of K. inostranzewi was found in Mangush (currently Prokhladnoe), that of K. biassalense comes from Biassala (currently Verkhorechie), the same locality as the holotype of Neohoploceras karakaschi (), a species that we have recorded precisely in the upper part of the S. verrucosum Zone. We can conclude that the finely ribbed specimens of Karakaschiceras found in the uppermost part of the lower Valanginian do not belong to K. biassalense, but they would rather be extreme morphotypes of K. inostranzewi. Therefore, K. biassalense cannot be kept as index species for this interval, and has to be replaced by K. inostranzewi. This proposal was accepted by the Group and integrated in the last version of the standard Mediterranean zonation ( et al., 2014).
We consider our Karakaschiceras inostranzewi Zone to be equivalent to the Karakaschiceras biassalense Subzone of et al. (2003). et al. (2014) pointed out that some confusion exists about the conception of this last unit, which could be understood to be defined at its base by the appearance of K. biassalense (actually K. inostranzewi) or, alternatively, by the appearance of Valanginites fuhri (species that was selected by et al., 2003, as index of the lower horizon of this subzone). In any case, the difference would have little significance since, as stated above, the FO of V. fuhri only very slightly predates that of K. inostranzewi. Nevertheless, it has to be stressed that the Saynoceras fuhri Horizon of et al. (2003) is not equivalent at all to the Saynoceras fuhri Horizon of & (1995a) since, whereas the latter nearly correlates with our Valanginites dolioliformis Subzone (see discussion above), the former would approximately equate our K. inostranzewi Subzone.
The Saynoceras contestanum Subzone proposed in this paper would correlate in turn with the Eristavites platycostatus Horizon of & (1995a) and et al. (2003). Two reasons lead us to use S. contestanum rather than Neocomites platycostatus as index for this stratigraphic interval. Firstly, we have serious doubts on the taxonomic status of N. platycostatus. In our opinion, most of the specimens assigned to this taxon in the literature could be no more than extreme morphotypes of Neocomites gr. neocomiensis-teschenensis (see , 1987; , 2004). Secondly, these morphotypes, which become relatively common in the Saynoceras verrucosum Zone, are still extremely rare in the uppermost lower Valanginian beds in our sections. Something similar seems to happen elsewhere, as some authors have found these forms only in the S. verrucosum Zone (, 1995; , 2001, 2003; , 2004).

Click on thumbnail to enlarge the image.
Figure 9: A: "Thurmanniceras" pertransiens (Y.CL2.26.1, section Y.CL2, "Th." pertransiens Zone, V. salinarium Subzone). B: Vergoliceras salinarium (Y.T.19.32, section Y.T, N. neocomiensiformis Zone, B. hirsutus Subzone). C: Neocomites neocomiensiformis (Y.V.R'.1, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). D: Baronnites hirsutus (Y.N.5.19, section Y.N, N. neocomiensiformis Zone, B. hirsutus Subzone). E: Neocomites subtenuis (Y.T.8.2, section Y.T, N. neocomiensiformis Zone, B. hirsutus Subzone). F: "Busnardoites" subcampylotoxus (Y.V.5.38, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). G: "Busnardoites" campylotoxus (Y.V.6.14, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). H: Olcostephanus guebhardi (Y.V.5.6, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). I: Valanginites dolioliformis (Y.V.6.47, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). J: Valanginites fuhri (Y.V.R'.17, section Y.V, N. neocomiensiformis Zone, V. dolioliformis Subzone). K: Karakaschiceras inostranzewi (Y.V.12.75, section Y.V, K. inostranzewi Zone, K. inostranzewi Subzone). L: Saynoceras contestanum (Y.P.6.45, section Y.P, K. inostranzewi Zone, S. contestanum Subzone).
The analysis of the vertical distribution of more than 10000 specimens from 16 sections in the Subbetic Domain has provided an accurate record of the succession of ammonite bioevents occurring during the early Valanginian in the Mediterranean region. We have identified in this interval three significant faunal turnovers that allow us to distinguish the three biozones, each of them with two subzones, composing the new zonal scheme proposed here.
The "Thurmanniceras" pertransiens Zone is maintained as defined by (1987). At the lower boundary of this zone, typical upper Berriasian species are replaced by new forms such us "Thurmanniceras" pertransiens, Kilianella roubaudiana, Neocomites premolicus and Sarasinella eucyrta. The FAD of Vergoliceras salinarium defines the boundary between the two subzones of this zone.
The N. neocomiensiformis Zone is featured by a strong diversification of the olcostephanids (with the appearance of the genera Baronnites, Saynoceras and Valanginites, together with several new species of Olcostephanus), the emergence of the first oosterellids (genera Paquiericeras and Julianites) and a nearly complete renewal of the neocomitid stock.
Further changes, mainly at the species level, take place at the base of the Karakaschiceras inostranzewi Zone, with the disappearance of most of the species present in lower stratigraphic levels and their replacement by new taxa, such as Karakaschiceras inostranzewi, Neohoploceras provinciale, Neocomites gr. neocomiensis-teschenensis, Neocomites platycostatus, Sabbaiceras beaumugnense, Paquiericeras paradoxum and others that already herald the late Valanginian assemblages.
Although the zonation proposed is intended only for the Betic Cordillera, the assemblages characterizing each of the units can be recognized throughout the Mediterranean Region.
We want to thank our colleague Antonio P. for his photographic assistance, as well as David for his review of the first English version of this paper. We are also grateful to the reviewers (María B. , Stéphane and Zdeněk ) for their very constructive comments. Our work has been co-financed by Project CGL2011-23759 (Spanish Ministry of Economy and Competitiveness) and Research Group RNM-178 (Junta de Andalucía).
R. (1994).- Nannofósiles del Cretácico de la Cordillera Bética (sur de España). Bioestratigrafía.- Universidad de Granada, 413 p.
R., M. & J.M. (2000).- The Berriasian-Valanginian boundary in the Mediterranean region: new data from the Caravaca and Cehegín sections, SE Spain.- Cretaceous Research, vol. 21, p. 1-21.
F., W. & , J. (1975).- The Berriasian of Caravaca (Prov. of Murcia) in the subbetic zone of Spain and its importance for defining this stage and the Jurassic-Cretaceous boundary. In: Colloque sur la limite Jurassique-Crétacé, Lyon-Neuchâtel, septembre 1973.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, nº 86, p. 14-22.
F. & J. (1979).- Les faunes de calpionelles du Berriasien supérieur/Valanginien. In: R., J.-P. & M., eds., Hypostratotipe mesogéen de l'étage Valanginien (Sud-Est de la France).- Éditions du C.N.R.S, Paris, p. 99-109.
H., M. & J.-P. (1981).- Les Calcaires du Fontanil des environs de Grenoble: leur place dans la stratigraphie du Néocomien entre le Jura et le domaine vocontien.- Eclogae Geologicae Helvetiae, Bâle, vol. 74, p. 109-137.
F. & S. (1995a).- Le Valanginien-Hauterivien basal du bassin vocontien et de la bordure provençale : zonation et corrélations.- Comptes Rendus de l'Académie des Sciences, Paris, (série IIa), vol. 320, p. 985-992.
F. & S. (1995b).- Neolissoceras (Carinites) nouveau sous-genre d'ammonites du Valanginien du bassin vocontien (SE de la France).- Comptes Rendus de l'Académie des Sciences, Paris, (série IIa), vol. 321, p. 1203-1210.
E. (1995).- Lower Cretaceous (Valanginian-Early Aptian) ammonite succession in the Sviniţa region (SW Rumania).- Géologie Alpine, Grenoble, Mémoire H.S., vol. 20 (1994), p. 113-167.
E.J. & I.A. (2000).- New and poorly known ammonites from South-West Crimea.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Bruxelles, (Sciences de la Terre), vol. 70, p. 89-120.
É., L.G. & J.-C. (1994).- La coupe de référence de Montbrun-les-Bains (Drôme, SE France): un stratotype potentiel pour la limite Berriasien-Valanginien.- Comptes Rendus de l'Académie des Sciences, Paris, (série IIa), vol. 318, p. 101-108.
L. (1993).- Valanginian ammonite fauna from the Kisújbánya Basin (Mecsek Mts., South Hungary) and its palaeobiogeographical significance.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 188, p.103-131.
L.G. (1995, unpublished).- Les formations à ammonites du Crétacé inférieur dans le Sud-Est de la France (Berriasien à Hauterivien) : biostratigraphie, paléontologie et cycles sédimentaires.- PhD Thesis, Muséum National d'Histoire Naturelle, Paris, 375 p.
L.G., É., M., S., S., P.J., H., F., J., G. & P.F. (1996).- The Valanginian stage.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, Bruxelles, vol. 66-supp., p. 11-18.
L.G. & J.-P. (1993).- Implications chronostratigraphiques de la révision de l'échelle biostratigraphique du Valanginien supérieur et de l'Hauterivien du Sud-Est de la France.- Comptes Rendus de l'Académie des Sciences, Paris, (série II), vol. 317, p. 387-394.
L.G. & J.-P. (1995).- Les biohorizons du Valanginien du Sud-Est de la France : un outil fondamental pour les corrélations au sein de la Téthys occidentale.- Géologie Alpine, Grenoble, Mémoire H.S., vol. 20 (1994), p. 15-41.
L.G., J.-P., H. & G. (1995).- The Lower Cretaceous of the South Vocontian basin and margins.- Géologie Alpine, Grenoble, Mémoire H.S., vol. 20 (1994), p. 383-399.
L.G., J.-P., E. & J. (1993).- Le cadre stratigraphique du Valanginien supérieur et de l'Hauterivien du Sud-Est de la France: définition des biochronozones et caractérisation de nouveaux horizons.- Géologie Alpine, Grenoble, vol. 68 (1992), p. 13-56.
R., , J., M & B. (2003).- Le Crétacé inférieur de la Veveyse de Châtel (Ultrahelvétique des Préalpes externes; canton de Fribourg, Suisse).- Revue de Paléobiologie, Genève, vol. 22, p. 1-174.
R. & J.-P. (1979a).- Les zones d'ammonites du Valanginien. In: R., J.-P. & M., eds., Hypostratotype mesogéen de l'étage Valanginien (Sud-Est de la France).- Éditions du C.N.R.S, Paris, p. 58-68.
R. & J.-P. (1979b).- Conclusions. In: R., J.-P. & M., eds., Hypostratotype mesogéen de l'étage Valanginien (Sud-Est de la France.- Éditions du C.N.R.S, Paris, p. 127-134.
M. (1987).- Los ammonites del Valanginiense del sector oriental de las Cordilleras Béticas (SE de España).- Universidad de Granada, 294 p.
M. & J.M. (1982).- Los ammonites del tránsito Berriasense-Valanginiense en la región de Cehegín (prov. de Murcia, SE de España).- Cuadernos de Geología Ibérica, Madrid, vol. 8, p. 651-664.
M. & J.M. (2013a).- Lower Valanginian ammonite biostratigraphy in the Betic Cordillera (southeastern Spain): new data. In: 9th International Symposium on the Cretaceous System, Ankara (Turkey).- Abstract Book, Middle East Technical University, Ankara, p. 118-119 (abstract).
M. & J.M. (2013b).- Ammonites del Valanginiense inferior (Cretácico inferior) de la Cordillera Bética. Nuevos datos bioestratigráficos. In: XXIX Jornadas de la Sociedad Española de Paleontología, Córdoba (Spain).- Libro de Resúmenes, Real Jardín Botánico de Córdoba, Córdoba, p. 143-144 (abstract).
P. (1971).- Le Crétacé inférieur de l'arc subalpin de Castellane entre l'Asse et le Var : stratigraphie et sédimentologie.- Mémoires du Bureau de Recherches Géologiques et Minières, Paris, nº 68, 313 p.
M. (2004).- Les ammonites néocomiennes dans l'Atlas atlantique (Maroc). Biostratigraphie, paléontologie, paléobiogéographie et paléoécologie.- Strata, Toulouse, (série 2), vol. 43, 223 p.
P., D., A., G. & , N. (1997).- Valanginian and early Hauterivian ammonite successions in the Mt Catria group (Central Apennines) and in the Lessini Mts (Southern Alps), Italy.- Paleopelagos, Roma, vol. 7, p. 59-100.
A.N. & P.F. (1993).- The first Early Cretaceous ammonite faunas from Baluchistan.- Cretaceous Research, vol. 14, p. 91-100.
I., N.M.M., G.D., J. & J. (2010).- Integrated isotope and biostratigraphy of a Lower Cretaceous section from the Bakony Mountains (Transdanubian Range, Hungary): A new Tethyan record of the event.- Cretaceous Research, vol. 31, p. 525-545.
G.A. de (2004).- Bioestratigrafía y eventos del Cretácico Inferior en las Zonas Externas de la Cordillera Bética.- Universidad de Jaén, 658 p.
P.J. (1982).- Ammonite biostratigraphy of the uppermost Tithonian, Berriasian, and lower Valanginian along the Río Argos (Caravaca, SE Spain).- Scripta Geologica, Leiden, vol. 65, p. 1-81.
P.J. & L. (1990).- Preliminary ammonite zonation for the Lower Cretaceous of the Mediterranean region.- Géologie Alpine, Grenoble, vol. 66, p. 123-127.
P.J. & H. (1995).- Biostratigraphy and sequence stratigraphy of the Berriasian-lowest Aptian (Lower Cretaceous) of the Río Argos succession, Caravaca, SE Spain.- Cretaceous Research, vol. 16, p. 195-230.
P.J., S., M.B., P., M., F., R., M., C., J., A., I., N., P.F., P., Z., J. & M.G.E. (2003).- Report on the 1st International Workshop of the IUGS Lower Cretaceous Ammonite Working Group, the ' Group' (Lyon, 11 July 2002).- Cretaceous Research, vol. 24, p. 89-94.
N.M.M. (2003).- Mediterranean Neocomian belemnites, part 2: the Berriasian-Valanginian boundary in southeast Spain (Río Argos, Cañada Lengua and Tornajo).- Scripta Geologica, Leiden, vol. 126, p. 121-183.
S. (2014, unpublished).- Biostratigraphie intégrée à nannofossiles calcaires et ammonoïdes : développement et implications pour la définition et la valorisation des stratotypes d'unité et de limite. L'exemple des étages Berriasien et Valanginien et de leur limite (~140 Millions d'années).- PhD Thesis, Université Claude Bernard, Lyon, 226 p.
J. (2005).- Lower Cretaceous Ammonites I. Perisphinctaceae 1: Himalayitidae, Olcostephanidae, Holcodiscidae, Neocomitidae, Oosterellidae. In: W., ed., Fossilium Catalogus I: Animalia, pars 139.- Backhuys, Leiden, 484 p
J. (2006).- Lower Cretaceous Ammonites II. Perisphinctaceae 2: Polyptychitidae. In: W., ed., Fossilium Catalogus I: Animalia, pars 141.- Backhuys, Leiden, p. 1-186.
J. & P.J. (1999).- Ammonite stratigraphy of the Valanginian to Barremian for the Mediterranean region.- Scripta Geologica, Leiden, Spec. Issue 3, p. 97-127.
J., R., B., Y. & Z. (2009).- Lower Cretaceous Ammonites IV. Boreophylloceratoidea, Phylloceratoidea, Lytoceratoidea, Tetragonitoidea, Haploceratoidea including the Upper Cretaceous representatives. In: W., ed., Fossilium Catalogus I: Animalia, pars 146.- Backhuys, Leiden, 416 p.
I.V. (1999).- Berriasian cephalopods of the Crimea and the Caucasus.- Proceedings of the Georgian Academy of Sciences Geological Institute, Tbilisi, (new series), vol. 112, 188 p. [in Russian]
G. & J. (1968).- Tithonique supérieur et Berriasien de l'Ardèche et de l'Hérault. Corrélation des ammonites et des calpionelles.- Geobios, Lyon, vol. 1, p. 7-70.
G. (1939).- Les Palehoplitidae tithoniques et berriasiens du Sud-Est de la France.- Mémoires de la Société Géologique de France, Paris, (nouvelle série), nº 41, 303 p.
J.G. & L.A. (2012).- Cretaceous. In: F.M., J.G., M. & G., eds., The geologic time scale 2012.- Elsevier, Amsterdam, p. 793-853.
J. (1969).- Étude géologique de l'ouest de la province de Murcie (Espagne).- Mémoires de la Société Géologique de France, Paris, (nouvelle série), nº 111, 270 p.
D. & E. (1976).- Les céphalopodes des couches de Carhaga (Tithonique supérieur-Barrémien inférieur).- Mémoires de l'Institut de Géologie et de Géophysique, Bucharest, vol. 24, p. 153-201.
S., I. & D. (2011).- Berriasian-Valanginian boundary in Bulgaria.- Review of the Bulgarian Geological Society, Sofia, vol. 72, p. 91-97.
A. & R. (2004).- Ammonite-supported microfossil and nannofossil stratigraphy of the Tithonian-Hauterivian limestones in selected sections of the Branisko succession, Pieniny Klippen Belt (Poland).- Studia Geologica Polonica, Kraków, vol. 123, p. 133-197.
S. (1996).- L'évolution des ammonites du Valanginien-Hauterivien inférieur du bassin vocontien et de la plate-forme provençale (Sud-Est de la France): relations avec la stratigraphie séquentielle et implications biostratigraphiques.- Documents des Laboratoires de Géologie de Lyon, Villeurbanne, vol. 137 (1995), 371 p.
S. & F. (1999).- Comments and proposals about the Valanginian-Lower Hauterivian ammonite zonation of south-eastern France.- Eclogae Geologicae Helvetiae, Basel, vol. 92, p. 183-197.
S., P.F., J.A., M.B., R., Y., M., C., V., A., B., V., H., Z., E.J., D., S., T.N., L.G., J.L., I.A., P. & O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Dijon, France, 30th August 2010).- Cretaceous Research, vol. 32, p. 786-793.
S., O., M.B., R., M., V., M., M., J.A., J., E.J., M.K., I., C., S., A., S.N., P.F. & J.M. (2014).- Report on the 5th International Meeting of the IUGS Lower Cretaceous Working Group, the " Group" (Ankara, Turkey, 31st August 2013).- Cretaceous Research, vol. 50, p. 126-137
J. & J.-P. (1973a).- Coupe A. III-2: Barret-le-Bas, Les Sausses. In: Colloque sur la limite Jurassique-Crétacé, Lyon-Neuchâtel, septembre 1973. Livret guide des excursions.- Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon, Villeurbanne, Hors série 1, p. 90-95.
J. & J.-P. (1973b).- Coupe A. IV-1: La Faurie-Pusteau. In: Colloque sur la limite Jurassique-Crétacé, Lyon-Neuchâtel, septembre 1973. Livret guide des excursions.- Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon, Villeurbanne, Hors série 1, p. 101-105.
J. (1993).- Análisis de la cuenca Subbética durante el Jurásico y el Cretácico en la transversal Caravaca-Vélez Rubio.- Universidad de Granada, 460 p.
G. (1901).- Les ammonites pyriteuses des marnes valangiennes du Sud-Est de la France.- Mémoires de la Société Géologique de France, Paris, (Paléontologie), nº 23, 66 p.
J.-P. (1973).- The occurrence and distribution of boreal ammonites from the Neocomian of southeast France (Tethyan Province). In: R. & P.F., eds., The Boreal Lower Cretaceous.- Geological Journal, Liverpool, special issue 5, p. 289-302.
V. (1902).- Über die Cephalopodenfauna der Teschener und Grodischter Schichten.- Denkschriften der Kaiserlichen Akademie der Wissenschaften, Wien, (Matematisch-Naturwissenschaftliche Classe), vol. 72, p. 1-87.
Z. (1975).- Zur Revision der Ammoniten von den Oberen Tĕín-Schichten (Valendis-Stufe).- Sborník Geologických Vĕd, Praha, (Paleontologie), vol. 17, p. 71-103.
Z. (2010).- Early Cretaceous ammonites from the Butkov Quarry (Manín Unit, Central Western Carpathians, Slovakia).- Acta Geologica Polonica, Warsaw, vol. 66, p. 393-415.
G.W. van (1969, unpublished).- Geological investigations in the region west of Caravaca, south-eastern Spain.- PhD Thesis, University of Amsterdam, 143 p.
M.G.E. (2001).- Die tiefe Unter-Kreide (Berrias bis Unter-Hauterive im Sudwestmarokkanischen Becken: Ammonitenfauna, Bio- und Sequenzstratigraphie.- PhD Thesis, Ruhr-Universität, Bochum, 142 p. [available online at http://www-brs.ub.ruhr-uni-bochum.de/netahtml/HSS/Diss/WippichMaxGE/diss.pdf]
M.G.E. (2003).- Valanginian (Early Cretaceous) ammonite faunas from the western High Atlas, Morocco, and the recognition of western Mediterranean 'standard' zones.- Cretaceous Research, vol. 24, p. 357-374.