◄ Carnets Geol. 15 (7) ►
![]()
Contents
[1. Introduction]
[2. Geological background and locality]
[3. Material and methods] [4. Results]
[5. Discussion]
[Bibliographic references] and ... [Table]
Department of Geology, University of Tartu, Ravila 14A, 50411 Tartu (Estonia)
Institute
of Geology, Tallinn University of Technology, Ehitajate tee 5, Tallinn, 19086 (Estonia)
Published online in final form (pdf) on June 11, 2015
[Editor: Bruno ;
language editor: Phil ]
![]()
The Ordovician hardground faunas of Estonia are not diverse. They include echinoderm holdfasts (i.e., eocrinoids and crinoids), edrioasteroids, bryozoans (both hemispherical trepostomes and stalked ptilodictyids) and cornulitids. The earliest hardground faunas appeared in the Dapingian (i.e., bryozoans and echinoderms). The Estonian hardground faunas are less diverse than the North American ones. North American hardgrounds seem to be more heavily encrusted than the Estonian ones. These differences may be due to the paleogeographic distances, different climates and different sedimentation environments of the paleocontinents.
Hardground; Cornulites; Bryozoa; echinoderms; Ordovician; Baltica.
O. & U. (2015).- Some encrusted hardgrounds from the Ordovician of Estonia (Baltica).- Carnets Géol., Madrid, vol. 15, nº 7, p. 63-70.
Quelques fonds durcis et encroûtés de l'Ordovicien d'Estonie, bouclier balte (Baltica).- La faune des fonds durcis ordoviciens est peu diversifiée. On y observe des structures d'ancrage d'échinodermes (d'éocrinoïdes et de crinoïdes), des édrioasteroïdes, bryozoaires (à la fois des trépostomes hémisphériques et des ptilodictyidés pourvus de tiges ) et des cornulitidés. Les premières peuplement de fonds durcis apparaissent au Dapingien : il s'agit alors de bryozoaires et d'échinodermes. La faune des fonds durcis estoniens est moins diversifiée que celle d'Amérique du Nord. Les fonds durcis d'Amérique du Nord semblent être plus fortement encroûtés que ceux d'Estonie. Ces différences pourraient être attribuables aux distances entre ces deux paléocontinents, à des différences climatiques ou d'environnements de dépôt.
Fond durci ; Cornulites ; bryozoaires ; échinodermes ; Ordovicien ; bouclier balte ; Baltica.
Carbonate hardgrounds are synsedimentarily cemented carbonate layers that have been exposed on an ancient seafloor ( & , 1992). Hardgrounds form excellent attachment surfaces for encrusting and bioeroding (i.e., boring) organisms (, 1982). These organisms otherwise dwell on carbonate cobbles or exoskeletons of various invertebrates. Carbonate hardgrounds are more common in calcite sea conditions, such as in the Ordovician, than in aragonite seas. Calcite seas favored early cementation of carbonate sediments in the seafloor ( & , 1992). Ordovician hardground faunas are relatively well documented. They are among the best studied hardground faunas in general, especially the North American examples. Much less is known of Ordovician hardground faunas in Baltica and the eastern Baltic. The earliest hardground ichnofaunas of Baltica include Trypanites and Gastrochaenolites? from the latest Early Ordovician ( & , 2010). Ripple marks associated with the hardground in the Vasalemma quarry were described in detail by & (2008). They also mentioned the occurrence of Trypanites borings and the edrioasteroid Cyathocystis. Another hardground is located above the reef bodies in the Vasalemma quarry. This hardground is remarkable for its numerous cornulitid encrusters.
This paper attempts to: 1) identify encrusted hardgrounds in the Ordovician of Estonia, 2) identify hardground encrusters in the Ordovician of Estonia, 3) find trends in the record of hardground encrustation in the Ordovician of Estonia, 4) find possible controls on the hardground encrustation in the Ordovician of Estonia, 5) compare hardground encrustation in Baltica with that of North America.
The area of Estonia
(Fig. 1 ![]() )
was covered by a shallow epicontinental
sea in the Ordovician. The Ordovician sequence of northern Estonia is relatively
complete
(Fig. 2
)
was covered by a shallow epicontinental
sea in the Ordovician. The Ordovician sequence of northern Estonia is relatively
complete
(Fig. 2 ![]() )
and represented mostly by carbonate rocks, except for the
terrigenous Lower Ordovician part. Ordovician carbonate rocks are exposed in
northern Estonia as a wide belt from the Narva River in the east to Hiiumaa
Island in the west ( & ,
2007). The drift of
Baltica from the southern high latitudes (i.e., temperate climate zone)
to the tropical realm caused a drastic climatic change ( & ,
1990; et al., 1992).
Sedimentation rate of carbonates increased during the warming of the climate.
Due to climatic change deposits that are characteristic of an arid and tropical
climate appeared in the Estonian Ordovician sequence ( & ,
2007). In the Early and Middle Ordovician, when the Baltic sedimentation basin
was situated in a temperate climate zone, these types of tropical deposits were
completely lacking (, 1973). The first signs of a warm climate
in the early Katian include the appearance of tabulate corals, stromatoporoids
and reefs, but it was not until the Hirnantian that they became prevalent (
& ,
2007).
)
and represented mostly by carbonate rocks, except for the
terrigenous Lower Ordovician part. Ordovician carbonate rocks are exposed in
northern Estonia as a wide belt from the Narva River in the east to Hiiumaa
Island in the west ( & ,
2007). The drift of
Baltica from the southern high latitudes (i.e., temperate climate zone)
to the tropical realm caused a drastic climatic change ( & ,
1990; et al., 1992).
Sedimentation rate of carbonates increased during the warming of the climate.
Due to climatic change deposits that are characteristic of an arid and tropical
climate appeared in the Estonian Ordovician sequence ( & ,
2007). In the Early and Middle Ordovician, when the Baltic sedimentation basin
was situated in a temperate climate zone, these types of tropical deposits were
completely lacking (, 1973). The first signs of a warm climate
in the early Katian include the appearance of tabulate corals, stromatoporoids
and reefs, but it was not until the Hirnantian that they became prevalent (
& ,
2007).
Click on thumbnail to enlarge the image.
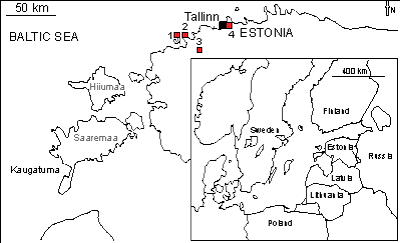
Figure 1: Map with locations of encrusted Ordovician hardgrounds in Estonia (modified after , 2006). 1 - Väike-Pakri Island, 2 Pakri Cliff at Cape Pakri, 3 Vasalemma quarry, and 4 - Suhkrumägi outcrop.
The large Vasalemma quarry is situated in Vasalemma settlement in northwestern
Estonia
(Fig. 1 ![]() ), west of the Keila-Riisipere railway. Limestones of the quarry
are excavated by Nordkalk AS down to the rippled upper surface of the Pääsküla
Member of the Kahula Formation ( & ,
2008). The
Vasalemma Formation consists of a succession of biodetrital grainstones up to 15
m thick with numerous intercalated reef bodies, composed of bryozoan
framestone-bindstone, echinoderm bindstone, receptaculitid-bryozoan-microbial
framestone, and tabulate bafflestone. The reef bodies reach diameters of more
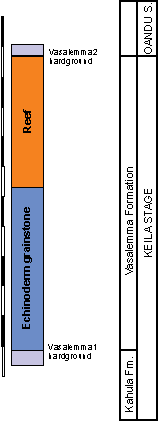
than 50 m ( et al., 2014). The studied hardground surfaces are located above the reef bodies and at
the base of Vasalemma Formation (Fig. 3
), west of the Keila-Riisipere railway. Limestones of the quarry
are excavated by Nordkalk AS down to the rippled upper surface of the Pääsküla
Member of the Kahula Formation ( & ,
2008). The
Vasalemma Formation consists of a succession of biodetrital grainstones up to 15
m thick with numerous intercalated reef bodies, composed of bryozoan
framestone-bindstone, echinoderm bindstone, receptaculitid-bryozoan-microbial
framestone, and tabulate bafflestone. The reef bodies reach diameters of more
than 50 m ( et al., 2014). The studied hardground surfaces are located above the reef bodies and at
the base of Vasalemma Formation (Fig. 3 ![]() ).
).
|
|
|
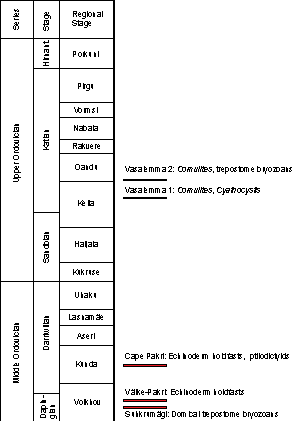
Figure 2: Stratigraphy of the Middle and Upper Ordovician of Estonia (modified after et al., 2008). Location of encrusted hardgrounds marked. |
Figure 3: Stratigraphic locations of hardgrounds in the Vasalemma quarry. This section is modified after et al. (2014). |
Hardground cobbles from Väike-Pakri Island Two large cobbles from Toila Formation (Volkhov Regional Stage, Dapingian) of Väike-Pakri Island have a total area of 40.8 cm2. Studied material came from the collections of Institute of Geology, Tallinn University of Technology.
Hardground pebbels from Suhkrumägi Two large pebbles from Suhkrumägi, Toila Formation (Volkhov Regional Stage, Dapingian) with total area 21.5 cm2. Studied material came from the collections of Institute of Geology, Tallinn University of Technology.
Vasalemma 1 hardground Nine hardground samples with total area 818 cm2 from Vasalemma quarry. This hardground forms a boundary between Kahula and Vasalemma Formation of the Keila Regional Stage (early Katian). Studied material came from the collections of Institute of Geology, Tallinn University of Technology.
Vasalemma 2 hardground - Three hardground slabs collected from the Vasalemma quarry were studied for encrustation and bioerosion using the collections of Institute of Geology, Tallinn University of Technology. The studied hardground area was 562 cm2. Only the hardground upper surface was exposed for the study. This hardground forms the upper surface of Vasalemma Formation (early Katian).
Sample orientation was marked during collecting. Samples were manually cleaned with water and soft brushes. Maximal number of Trypanites borings was counted in 4cm2 using a cm grid drawn on a transparent film ( et al., 2004). A centimeter grid was also used to measure the area of the studied hardground surface. A millimeter grid drawn on a transparent film was used to measure the area covered by cornulitid and bryozoan encrusters. An Olympus B 061 binocular microscope equipped with caliper (accuracy 0.1 mm) was used to measure cornulitid specimens. The hardground samples were photographed using Nikon 7000 digital camera.
Previously studied hardgrounds
Cape Pakri hardground (, pers. obs.) The encrusters compose of
discoidal echinoderm holdfasts and a ptilodictyid bryozoan holdfasts (Figs. 2 ![]() & 4
& 4 ![]() ;
Table) cemented to the hardground surface within the Pakri Formation
(Darriwilian). The encrusters cover only 4.65 cm2 (0.37%) of the
total hardground sample studied (1260 cm2). Sample
represents sandy limestone of relatively shallow water and nearshore origin (
& , 1997)
;
Table) cemented to the hardground surface within the Pakri Formation
(Darriwilian). The encrusters cover only 4.65 cm2 (0.37%) of the
total hardground sample studied (1260 cm2). Sample
represents sandy limestone of relatively shallow water and nearshore origin (
& , 1997)

Click on thumbnail to enlarge the image.
Figure 4: Ptilodictyid bryozoan (Br) (TUG 1681-2) cemented to the hardground from Pakri Formation (Darriwilian) of Pakri Cliff, Cape Pakri, NW Estonia. Small disc below the bryozoan belongs possibly to echinoderms (TUG 1681-3).
The hardground cobbles from Väike-Pakri Island
The Dapingian hardground cobble
studied (Toila Formation of Volkhov Regional Stage)
is formed of pure grey limestone and has a slightly lens-shaped profile. Its
surface is strongly mineralized by pyrite and has a much darker coluor than the
matrix. The surface of cobble is relatively smooth and has numerous Trypanites borings. A discoidal echinoderm holdfast is cemented to
upper surface of the cobble (Figs. 2 ![]() & 5
& 5 ![]() ;
Table). The total encrustation
area is 0.07 cm2 of 40.8 cm2 (0.2%). Sample represents
limestone of relatively shallow water and near shore origin ( &
, 1997).
;
Table). The total encrustation
area is 0.07 cm2 of 40.8 cm2 (0.2%). Sample represents
limestone of relatively shallow water and near shore origin ( &
, 1997).

Click on thumbnail to enlarge the image.
Figure 5: Discoid echinoderm holdfast cemented to the hardground from Volkhov Regional Stage (Dapingian) of Suhkrumägi outcrop, northern Estonia (GIT 156-356).
Hardground
pebbles from Suhkrumägi - Studied Dapingian
(Toila Formation of Volkhov Regional Stage) limestone pebbles have an
irregular shape and slightly lens-shaped profile. Their surface is slightly
bumpy and is strongly mineralized by pyrite and has a dark colour that contrast
to the grey colour of the matrix. There are numerous Trypanites
borings in the pebbles. Three small bryozoans (possibly trepostomes) are
cemented to the pebble (Figs. 2 ![]() & 6
& 6 ![]() ;
Table). There is an indication (i.e.,
good preservation) that the eroded cobbles were secondary colonized by
bryozoans. The total encrustation area is 0.16cm2 of 21.5 cm2
(0.7%). Sample represents limestone of relatively shallow water and near shore
origin ( & ,
1997).
;
Table). There is an indication (i.e.,
good preservation) that the eroded cobbles were secondary colonized by
bryozoans. The total encrustation area is 0.16cm2 of 21.5 cm2
(0.7%). Sample represents limestone of relatively shallow water and near shore
origin ( & ,
1997).

Click on thumbnail to enlarge the image.
Figure 6: Two domical trepostome bryozoans and a possible ptilodictyid cemented to the hardground from Volkhov Regional Stage (Dapingian) of Väike-Pakri Island, NW Estonia (GIT 381-595).
Vasalemma 1 hardground
The surface of hardground has a dark brownish-grey colour due to mineralization by pyrite, contrasting to the grey limestone matrix. The surface is eroded and cuts the bioclasts in the hardground. We found just a single eroded cornulitid cemented to the hardground upper surface. The encrustation area of the studied hardground samples is 0.2 cm2 of 818 cm2 (0.02%). and (2008) reported also encrustation by edrioasteroid echinoderm Cyathocystis, which we did not find. The sample represents relatively shallow water and near shore sediments ( & , 1997) (Table).
Vasalemma 2 hardground (above reefs)
The hardground surface has a darker grey colour than the rest of rock
matrix, which is light grey in colour (Figs. 2 ![]() - 3
- 3 ![]() & 7
& 7 ![]() ;
Table). The
hardground occurs on the upper surface of a reef limestone layer. Its surface is
relatively smooth, with possible marks of microbioerosion. Bioclasts in the
hardground are abraded on the hardground surface. Its ledges have rounded edges.
Crypts are formed under the hardground ledges. The hardground ledges are 2 to 9
mm thick. The top surface of hardground is encrusted by cornulitids and
branching bryozoans (Fig. 8
;
Table). The
hardground occurs on the upper surface of a reef limestone layer. Its surface is
relatively smooth, with possible marks of microbioerosion. Bioclasts in the
hardground are abraded on the hardground surface. Its ledges have rounded edges.
Crypts are formed under the hardground ledges. The hardground ledges are 2 to 9
mm thick. The top surface of hardground is encrusted by cornulitids and
branching bryozoans (Fig. 8 ![]() ). The total encrustation area is 7.4 cm2
(1.3% of the total hardground surface studied).
). The total encrustation area is 7.4 cm2
(1.3% of the total hardground surface studied).

Click on thumbnail to enlarge the image.
Figure 7: Upper surface of the hardground, Vasalemma Formation, early Katian, Vasalemma quarry, NW Estonia (GIT 362-96).
Numerous
Cornulites sp. tubes occur on the
upper surface of the hardground (area 562 cm2, N=18) (Fig.
5 ![]() ). These
tubes lack preferred orientation. Their maximal diameters vary from 1.1 mm to
3.5 mm (mean 2.59 mm, sd=0.64, N=18). Most tubes are of similar large size, but
juveniles also occur. Some tubes were broken off from the hardground and their
bases were subsequently overgrown by other cornulitids. Overgrowth occurs
between four specimens of cornulitids. Weathered and broken off tubes (tube
bases) occur on the hardground (N=4). Distribution of cornulitids on the
hardground is patchy. Cornulitids cover 1.3% of the studied hardground surface
(562 cm2), but in some places they may encrust up to 4% of the
hardground surface.
). These
tubes lack preferred orientation. Their maximal diameters vary from 1.1 mm to
3.5 mm (mean 2.59 mm, sd=0.64, N=18). Most tubes are of similar large size, but
juveniles also occur. Some tubes were broken off from the hardground and their
bases were subsequently overgrown by other cornulitids. Overgrowth occurs
between four specimens of cornulitids. Weathered and broken off tubes (tube
bases) occur on the hardground (N=4). Distribution of cornulitids on the
hardground is patchy. Cornulitids cover 1.3% of the studied hardground surface
(562 cm2), but in some places they may encrust up to 4% of the
hardground surface.
Cornulites sp. tubes are straight to moderately curved, up to 1.7 mm long, and with diameters ranging from 1.1 mm to 3.5 mm (mean 2.59). The divergence angle of the tubes is 10.5o. The tubes are externally covered with very faint and dense longitudinal striation and moderately developed perpendicular annulation. Their bases are not widened to slightly widened, especially at annular crests. The lumen of the tubes is covered with prominent and regular annulation. The distance between annular crests at the tube diameter 3 mm is 0.7 to 1.0 mm. Cornulites sp. resembles most closely Cornulites? sp. D (, 2013) from the Vasalemma Formation by its moderately conical shell and prominent annulation in the tube lumen. It differs from C.? sp. D by presence of external faint longitudinal striation and less developed perpendicular ornamentation. The tube divergence angle is also slightly smaller in C. sp. than in C.? sp. D (12o).
One
branching trepostome bryozoan encrusts a cornulitid tube (Fig. 8 ![]() ). The bryozoan
encrustation area is 2.5 cm2 (0.45% of the total hardground area).
). The bryozoan
encrustation area is 2.5 cm2 (0.45% of the total hardground area).
In addition to cornulitids and bryozoans, a few Trypanites borings occur in the upper surface of the hardground. Trypanites borings range from 0.2 to 2.1 mm in diameter. There is a maximum five Trypanites borings per 4 cm2 of the hardground upper surface, but some areas (up to 10 cm2) have no borings.

Click on thumbnail to enlarge the image.
Figure 8: Cornulites sp. encrusted by a branching bryozoan, with Trypanites borings in the hardground, Vasalemma Formation, early Katian, Vasalemma quarry, NW Estonia (GIT 362-96).
Vasalemma 2 hardground
We interpret the palaeoenvironment of the Vasalemma hardground community as a shallow sea floor of normal salinity below the tidal zone on the basis of fauna and sediments ( et al., 2014). The relatively dark color of the hardground surface as compared to the rest of rock matrix is probably due to slight pyritization during diagenesis. The hardground was most likely cemented under sediment cover and exposed later to winnowing, which also exposed the cryptic surfaces. The occurrence of abraded bioclasts of reef origin in the hardground indicate that the hardground was abraded before the cornulitid and bryozoan encrustation. The sediment layer below the hardground may have still been soft or less cemented than the hardground during the abrasion; this conclusion is supported by the occurrence of crypts.
The relatively low bioerosion density on the studied Vasalemma hardground for the Late Ordovician ('s pers. obs.) could indicate a relatively short exposure time for the hardground, before it was buried by sediments, or a relatively low number of bioeroding organisms in the Vasalemma hardground community. However, considering the small size of the studied hardground sample and the usual patchy distribution of Trypanites borings, it is likely that the samples do not represent the most bioeroded part of the hardground.
Cornulitid tubeworms form an important and primitive group of the encrusting tentaculitoids. Cornulitids are presumably phylogenetically closely related to free-living tentaculitids (, 2010). The biological affinities of cornulitids have long been debated. They most likely represent Lophothrochozoa ( & , 2012) and could belong to stemgroup phoronids ( et al., 2010). Their palaeoecology is relatively well documented and understood as they generally retain their original position on the substrate after fossilisation ( & , 2003). In contrast to their close relatives, the microconchids that lived in waters of various salinities, cornulitid tubeworms are found only in normal marine sediments ( et al., 2012). Shallow marine sediments, especially those associated with carbonate platforms, are usually rich in cornulitid fossils. Cornulitids were most abundant in the Late Ordovician to Devonian, but their fossils are known also from the Carboniferous (, 2010). Six adaptive strategies have thus far been described in cornulitids, indicating a diverse ecology (, 2010).
Cornulitids show that hardground was exposed to encrusters long enough to allow at least two generations of cornulitids to colonize it, and then bryozoans to encrust the cornulitids. The numerical dominance of large sized Cornulites sp. specimens resulted presumably from colonization event/s where most of specimens reached adult size after the settling on the hardground. High juvenile mortality is common in many modern invertebrates. The situation with the Vasalemma hardground cornulitids can either be explained by low juvenile mortality or weaker preservation potential of juvenile specimens. The empty juvenile tubes could have been destroyed by the hydrodynamic activity of the waters above the hardground. The random orientation of the tubes indicates the lack of unidirectional water current or random settlement of larvae or lack of larvae reorientation after settlement. If there was unidirectional water current, the cornulitids would have oriented their apertures up current to achieve better position for suspension feeding. Oriented cornulitids have been reported from Ordovician to Carboniferous brachiopods (, 1967; , 2010) where they presumably took advantage of the feeding currents of brachiopods.
Middle Ordovician hardgrund encrusters
The Middle Ordovician hardground faunas of the Estonia are not diverse. They include only bryozoans and echinoderms. Among the bryozoans both domical colonies and stalked colonies (possible ptilodictyids) occurred. Echinoderms were represented at least by three different stemmed forms (i.e., eocrinoids or crinoids).
Trends in the hardground encrustation in the Ordovician of Estonia
All samples derive from relatively shallow water environments and no obvious bathymetric or sedimentological trends can be established for the Ordovician of Estonia ( & , 1997). However, there is a stratigraphical trend in the taxonomic composition of hardground encrusters in the Ordovician of Estonia.
In the Dapingian the first hardground encrusters appeared in the Ordovician of Estonia. They included bryozoans (possibly trepostomes), and echinoderm holdfasts (i.e., eocrinoids or crinoids). During the Darriwilian, ptilodictyid bryozoans appeared, but in general the early Middle Ordovician and late Middle Ordovician hardground faunas of Estonia were similar. In the Late Orodovician (early Katian) edrioasteroids (i.e., Cyathocystis) and cornulitids (i.e., Cornulites) appeared. Most notable is the late appearance of edrioasterioids in the Ordovician hardground communities of Estonia. Similarly, cornulitids are known from the late Darriwilian of Estonia (, 2013), but they colonized hardgrounds later in the early Katian.
There seems to be no remarkable stratigraphic trend in encrustation intensities in the Ordovician of Estonia, which is surprising considering the great climatic change in Baltica ( & , 1990; & , 1997) while moving from the temperate zone to tropics and the diversification of faunas during the Great Ordovician Biodiversification Event (GOBE) ( et al., 2004).
The hardground encrusters of the Ordovician of Estonia compared to the other regions
The Middle Ordovician hardgrounds of Estonia are very sparsely encrusted (<1% of the hardground area) as compared to known North American analogues. 10.5% of the Kanosh Shale hardground area is covered by encrusters (et al., 1992) which is a magnitude more than any known Middle Ordovician example from Estonia. Similar to Estonian (Baltica) faunas, North American Middle Ordovician hardground faunas contain bryozoans and echinoderms ( et al., 1992).
The Late Ordovician Vasalemma 2 hardground has a relatively small number (N=1) of branching bryozoans in the association, which usually cover the largest area on the other Ordovician hardgrounds, especially North American ones ( & , 1978; & , 2003). The Vasalemma 2 hardground fauna is unique for its numerical dominance by Cornulites sp. What is the reason for the high abundance of Cornulites sp. in this association? They may have been opportunistic suspension feeding encrusters, successful in conditions of low competition with other suspension feeders. Similar opportunistic behavior has been described in microconchids ( & , 2011; et al., 2012, 2013), a group of spirorbiform tubeworms descended from the same ancestor as the cornulitids ( & , 2012).
The relatively low skeletal coverage (1.3%) of the Vasalemma 2 hardground and an extremely low skeletal coverage (0.02%) of the Vasalemma 1 hardground contrast with the higher coverage of Late Ordovician analogues from North America ( & , 1978; & , 1984). This could represent a paleobiogeographic difference caused by a slightly colder climate in Baltica and possibly differences in faunal composition and sedimentation ( & , 1997). It can also be explained by low nutrient levels in the local sea water of Baltica (low productivity, e.g., et al., 2002; et al., 2015; et al., 2015) or by a large area of hardground being occupied by soft-bodied organisms in the community, which did not preserve as fossils. Alternatively, low skeletal coverage may indicate a short exposure time of the hardground ( et al., 2002).
The North American Middle Ordovician and Late Ordovician hardground faunas ( & , 1978; & , 1984; et al., 1992) seem to be more diverse than the Estonian ones (Baltica). This may be due to a different faunal composition caused by paleogeographic distance. During the Ordovician North America was located in a tropical zone while Baltica was located in a temperate zone until the earliest Katian. Thus, most likely the lower diversity of Estonian Ordovician hardground faunas was mostly caused by the different climates of the paleocontinents. It is a general trend that tropical faunas are more diverse than temperate ones ( & , 1999).
Financial support to O.V. was provided by the Palaeontological Association Research Grant, Estonian Research Council projects ETF9064 and IUT20-34. This paper is a contribution to IGCP 591 "The Early to Middle Palaeozoic Revolution". We are grateful to G. , Institute of Geology, Tallinn University of Technology for photographing the specimens. We are also grateful to M.A. , College of Wooster, for comments on the manuscript.
R.S.K. & R.N. (1999).- An introduction to marine ecology.- Blackwell Science, Oxford, 3rd Edition, 286 p.
C.E. & M.E. (1984).- Morphology, faunas and genesis of Ordovician hardgrounds from southern Ontario, Canada.- Palæogeography, Palæoclimatology, Palæoecology, vol. 46, p. 233290.
C.E. & W.D. (1978).- Preservation and paleoecology of a Middle Ordovician hardground community.- Paleobiology, Washington, vol. 4, p. 329348.
D.W. (1962).- Small conoidal shells of uncertain affinities. In: C.D., ed., Treatise on Invertebrate Paleontology.- Geological Society of America, New York; University of Kansas, Lawrence, Part W, p. 130143.
L., L., P. & T., eds. (2008).- The Seventh Baltic Stratigraphical Conference: Abstracts and Field Guide.- Geological Society of Estonia, Tallinn, 46 p.
L. & A. (2008).- Ripple marks as indicators of Late Ordovician sedimentary environments in Northwest Estonia.- Estonian Journal of Earth Sciences, Tallinn, vol. 57, p. 1122.
V. (1973).- Aspects of carbonate sedimentation in the Ordovician of Baltoscandia.- Lethaia, Oslo, vol. 6, p. 1134.
B., L. & O. (2014).- Age, facies and geometry of the Sandbian/Katian (Upper Ordovician) pelmatozoan-bryozoan-receptaculitid reefs of the Vasalemma Formation, northern Estonia.- Facies, Erlangen, vol. 60, p. 963986.
H.L., E. & M.J. (2002).- Mollusc shell encrustation and bioerosion rates in a modern epeiric sea: taphonomy experiments in the Java Sea, Indonesia.- Palaios, Lawrence, vol. 17, p. 171191.
M.-A. & O. (2007).- Excursion B2: Lower Paleozoic geology and corals of Estonia. In: Excursion Guidebook. 10th International Symposium on Fossil Cnidaria and Porifera (August 18-22, 2007).- Institute of Geology at Tallinn University of Technology, p. 5455.
T. (1982).- Cambrian to Cretaceous changes in hardground communities.- Lethaia, Oslo, vol. 15, p. 309323.
A. & A. (1997).- Geology and Mineral Resources of Estonia.- Estonian Academy Publishers, Tallinn, 436 p.
D. (1967).- Die Lebensweise von Mucrospirifer , 1931 (Brachiopoda).- Palæogeography, Palæoclimatology, Palæoecology, vol. 3, p. 381392.
E.F. von (1820).- Die Petrefaktenkunde auf ihrem jetzigen Standpunkte durch die Beschreibung seiner Sammlung versteinerter und fossiler Überreste des Thier- und Planzenreichs der Vorwelt erläutert.- Becker'sche Buchhandlung, Gotha, 436 p.
C.R. & W.S. (1990).- Revised world maps and introduction. In: W.S. & C.R., eds., Palaeozoic palaeogeography and biogeography.- The Geological Society of London, Memoirs, London, vol. 12, p. 121.
L., P. & E. (2004).- Environmental and substrate control on Paleozoic bioerosion in corals and stromatoporoids, Anticosti Island, Eastern Canada.- Palaios, Lawrence, vol. 19, p. 292306.
P.D., O. & M.A. (2010).- Evolution of biomineralization in "lophophorates".- Special Papers in Palaeontology, Oxford, vol. 84, p. 317333.
P.D. & M.A. (2003).- Palaeoecology and evolution of marine hard substrate communities.- Earth Science Reviews, vol. 62, p. 1-103.
T.H., M.A., R. van der, A., N. & E. (1992).- Baltica. A synopsis of VendianPermian palaeomagnetic data and their palaeotectonic implications.- Earth-Science Reviews, vol. 33, p. 133152.
O. (2010).- Adaptive strategies in the evolution of encrusting tentaculitoid tubeworms.- Palæogeography, Palæoclimatology, Palæoecology, vol. 292, p. 211221.
O. (2013).- Cornulitid tubeworms from the Ordovician of eastern Baltic.- Carnets Géol., Madrid, Letter 2013/03 (CG2013_L03), p. 131-138
O. & M.A. (2010).- Early large borings from a hardground of Floian-Dapingian age (Early and Middle Ordovician) in northeastern Estonia (Baltica).- Carnets Géol., Madrid, Letter 2010/04 (CG2010_L04), 4 p.
O. & M. (2012).- Phenetic phylogenetics of tentaculitoids - extinct problematic calcareous tube-forming organisms.- GFF, Stockholm, vol. 134, p. 145156.
B.D., M.L., F. & I., eds. (2004).- The Great Ordovician Biodiversification Event.- Columbia University Press, New York, 484 p.
M.A. & T.J. (1992).- Hardgrounds and hardground faunas.- University of Wales, Aberystwyth, Institute of Earth Studies Publications, Aberystwyth, vol. 9, p. 1131.
M.A., T.J., T.E., C.D. & L.E. (1992).- The development of an Early Ordovician hardground community in response to rapid sea-floor calcite precipitation.- Lethaia, Oslo, vol. 25, p. 1934.
M.A., S. & P.D. (2015).- Middle Jurassic (Callovian) cyclostome bryozoans from the Tethyan tropics (Matmor Formation, southern Israel).- Bulletin of Geosciences, Prague, vol. 90, p. 5163.
M., T., B., M., M.K. & A.V. (2015).- Paleoecology and sedimentary environment of the Late Devonian coral biostrome from the Central Devonian Field, Russia.- Palæogeography, Palæoclimatology, Palæoecology, vol. 424, p. 6175.
M. & W. (2011).- Microconchid tubeworms across the upper Frasnian lower Famennian interval in the Central Devonian Field, Russia.- Palaeontology, Oxford, vol. 54, p. 14551473.
M., P.D. & O. (2013).- Early Triassic (Spathian) post-extinction microconchids from western Pangaea.- Journal of Paleontology, Lawrence, vol. 87, p. 159165.
M., O. & M. (2012).- Invasion of freshwater and variable marginal marine habitats by microconchid tubeworms an evolutionary perspective.- Geobios, Villeurbanne, vol. 45, p. 603610.
Table: Encrusted hardgrounds in the Ordovician of Estonia.
| Hardground | Taxa | Area encrusted |
Total hardground area encrusted |
Total hardground area studied |
| Hardground cobbles from Väike-Pakri Island |
Discoidal
echinoderm holdfast |
0.07 cm2 (0.2%) | 0.07 cm2 (0.2%) | 40.8 cm2 |
| Hardground pebbles from Suhkrumägi | Trepostome bryozoans | 0.16 cm2 (0.7%) | 0.16cm2 (0.7%) | 21.5 cm2 |
| Cape Pakri hardground |
Discoidal
echinoderm holdfasts and bryozoans |
4.65 cm2 (0.37%) | 4.65 cm2 (0.37%) | 1260 cm2 |
| Vasalemma 1 hardground | Cornulites sp. | 0.2 cm2 (0.02%) | 0.2 cm2 (0.02%) | 818 cm2 |
| Vasalemma 2 hardground | Cornulites sp. | 7.4 cm2 (1.3%) | 7.4 cm2 (1.3%) | 562 cm2 |
| Branching bryozoans | 2.5 cm2 (0.45%) |