![]()
[Introduction]
[Material and geological setting] [Results]
[Discussion] [Bibliographic references]
and ... [Figures]
Department of Geology, University of Tartu, Ravila 14A, 50411 Tartu (Estonia)
Department of Geology, The College of Wooster, Wooster, OH 44691 (USA)
Manuscript online since December 14, 2010
![]()
Large plug- or slightly amphora-shaped borings have been found in the hardground marking the boundary between Early and Middle Ordovician rocks in northeastern Estonia. These borings cut large bioclasts of the trilobite Megistaspis and cannot be assigned with certainty to any known ichnotaxon. They indicate that the diversity of early borings may have been greater than was recognized previously.
Bioerosion; Gastrochaenolites; hardground; Early Ordovician; Baltica.
O. & M.A. (2010).- Early large borings from a hardground of Floian-Dapingian age (Early and Middle Ordovician) in northeastern Estonia (Baltica).- Carnets de Géologie / Notebooks on Geology, Brest, Note brève / Letter 2010/04 (CG2010_L04)
Premières grandes traces de perforation découvertes dans un fond durci d'âge Floien-Dapingien (à la transition de l'Ordovicien inférieur-moyen) dans le NE de l'Estonie (Baltique).- De grandes perforations cylindriques ou légèrement renflées, en forme d'amphore, ont été découvertes dans le fond durci qui sépare l'Ordovicien inférieur de l'Ordovicien moyen. Ces perforations recoupent de gros bioclastes dont des débris de trilobites (du genre Megistaspis). Elles ne peuvent être rattachées avec certitude à aucun ichnotaxon connu. Par conséquent la diversité des perforations primitives est certainement plus importante que ce qui était jusqu'à présent admis.
Bioérosion ; Gastrochaenolites ; fond durci ; Ordovicien inférieur ; Baltique.
Bioerosional trace fossils produced by macroborers are known since the Cambrian where they are represented by two genera Trypanites ( et alii, 1978) and Oichnus (, 2007). Respectively, these borings are relatively small and simple tubes and small circular holes in shells. The earliest large borings (Gastrochaenolites oelandicus & , 2001) are from the Lower Ordovician of Baltica ( & , 2001). Another Lower Ordovician Gastrochaenolites (identified as Gastrochaenolites isp.) is found in Utah ( et alii, 2004). In addition to Trypanites, other possible large borings in hardgrounds are common in the Lower Ordovician of Baltica ( et alii, 1996, 2000; & , 2001). The Ordovician was a time of great biodiversification (GOBE: & , 1983; , 1995; , 2003; et alii, 2010). Diversification of boring organisms, termed the Ordovician Bioerosion Revolution, was a function of this event. The number of ichnogenera increased from two (Cambrian) to at least eight (Ordovician, & , 2006).
Possible borings from the Lower to Middle Ordovician hardgrounds of Estonia have been known for a long time (, 1960, 1961; et alii, 2000). However, the boring nature and taxonomic affinities of the organisms that produced these structures has been controversial. There are difficulties involved in discriminating true borings (formed through bioerosion) from lithified burrows, particularly if the latter have been scoured or otherwise eroded. In Estonia, probable large early borings (Billingen to Volkhov Stages) have been affiliated with Amphorichnus , 1966, and Conichnus , 1966 ( et alii, 2000), but these ichnotaxa are definitely burrows and not borings ( pers. obs.). It has long been accepted that burrows and borings be given discrete ichnogenus and ichnospecies names, even if they are identical morphologically, because they represent dissimilar behaviors (, 1975). Large borings from the hardgrounds of the Volkhov sequence of neighboring northwestern Russia have been identified recently as Gastrochaenolites oelandicus. However, these traces have considerable morphological variability: drop-like forms are definitely G. oelandicus), but there are also spherical varieties (probable G. oelandicus), and pencil-like or conical borings/burrows ( & , 2005).
This ichnotaxonomic confusion indicates a possibility that not all earliest Ordovician East Baltic large conical trace fossils are true borings in hardgrounds, but
that some may be burrows excavated into non-lithified sea floor. This paper
attempts to:
1) evaluate these structures in northeastern Estonia as regards their origin as
borings,
2) discuss the ichnosystematics of these trace fossils, and
3) review the evolution of bioerosion in the Lower Ordovician of Baltica.
The Saka section (northeastern Estonia) (Fig. 1 ![]() ) is part of the Baltic Klint which is one of the most extensive outcrops of Lower Palaeozoic rocks in the world (,
2008). The section is 2 km west of the village of Saka. A trench about 20-30 m
in width and depth was cut into the Klint to accomodate a sewage pipe from nearby Kohtla-Järve to the sea.
Consequently, a section spanning
the Lower Cambrian Tiskre Formation up to the Middle Ordovician Aseri Formation is exposed. The
older part of the section is masked, but strata
of the Volkhov Stage are well exposed.
) is part of the Baltic Klint which is one of the most extensive outcrops of Lower Palaeozoic rocks in the world (,
2008). The section is 2 km west of the village of Saka. A trench about 20-30 m
in width and depth was cut into the Klint to accomodate a sewage pipe from nearby Kohtla-Järve to the sea.
Consequently, a section spanning
the Lower Cambrian Tiskre Formation up to the Middle Ordovician Aseri Formation is exposed. The
older part of the section is masked, but strata
of the Volkhov Stage are well exposed.
Seven borings were studied from the Saka section. These occur in a supposed hardground at the boundary between the Hunneberg and Volkhov stages. The large plug-shaped borings were bored into the upper surface of the dolomitized glauconitic packstone of the Hunneberg Stage. The trace fossils were interpreted to be borings if they cut biomineralized trilobite bioclasts (Megistaspis) in the glauconitic packstone.
In 1961 described a variety of possible borings from the northern Estonian sections of the Baltic Klint. Possible borings range widely in morphology: from cone-shaped (northeastern Estonia) to vase and drop-shaped (northwestern Estonia).
The hardground with large conical trace fossils is the boundary between the Lower and Middle Ordovician, and its age is either latest
Early Ordovician (Floian) or earliest Middle Ordovician (Dapingian). Its surface and the interiors of the trace fossils are impregnated with
iron-bearing minerals and are a rusty brown color. The trace fossils are filled
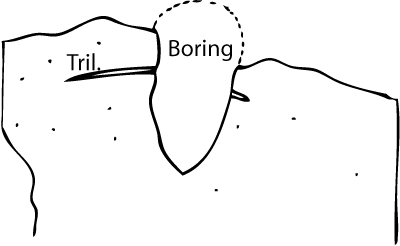
with sediments richer in glauconite than the rock forming the hardground. One large trace completely cuts a Megistaspis pygidium in the upper third of its length
(Fig. 2 ![]() ).
The plug- or slightly amphora-shaped straight trace fossils have a circular
cross-section. They are 10-18 mm in diameter and up to 30 mm deep. Their lower ends are sharp and slightly pointed
(Fig. 2
).
The plug- or slightly amphora-shaped straight trace fossils have a circular
cross-section. They are 10-18 mm in diameter and up to 30 mm deep. Their lower ends are sharp and slightly pointed
(Fig. 2 ![]() ).
).
That the large conical, slightly amphora-shaped traces in the Lower Ordovician glauconitic packstone are borings is supported by the observation that they sometimes cut large bioclasts (Megistaspis trilobites). The large Megistaspis skeletons are relatively massive. The traces were bored into a lithified sea-floor - hardground. If the rock had not been lithified (a lime mud) prior to their excavation, the Megistaspis bioclasts would have been pushed aside by the digging organism and not cut through. We agree with et alii (2004) that the cutting of a trilobite bioclast by Gastrochaenolites (Lower Ordovician) is direct evidence that this trace fossil is a true boring.
Conical slightly amphora-shaped borings are difficult to assign to any known ichnogenus
(Fig. 2 ![]() ). They are similar to Gastrochaenolites oelandicus in size and
in the absence of parallel walls in the shaft. However, they lack a
narrow aperture so are not club-shaped ( & ,
1984).Therefore they cannot be assigned to Gastrochaenolites. Gastrochaenolites isp. from the
Lower Ordovician of Utah ( et alii,
2004) differs from the eastern Estonian borings in being more
pouch-like
rather than conical or amphora-shaped and does not have a slightly pointed lower
end. The other vertical borings in Lower Ordovician
strata are those of Trypanites, but this fossil has parallel shaft walls
and is usually much smaller in diameter than the Utah trace fossil. Thus, the large borings described here could
be a previously undescribed ichnotaxon, but further studies on the morphology and variability of the borings are necessary before formal
description is feasible. Possible borings from a nearby section at Künnapõhja of the same age (,
1961) may be conspecific with those found in Saka
(Fig. 3
). They are similar to Gastrochaenolites oelandicus in size and
in the absence of parallel walls in the shaft. However, they lack a
narrow aperture so are not club-shaped ( & ,
1984).Therefore they cannot be assigned to Gastrochaenolites. Gastrochaenolites isp. from the
Lower Ordovician of Utah ( et alii,
2004) differs from the eastern Estonian borings in being more
pouch-like
rather than conical or amphora-shaped and does not have a slightly pointed lower
end. The other vertical borings in Lower Ordovician
strata are those of Trypanites, but this fossil has parallel shaft walls
and is usually much smaller in diameter than the Utah trace fossil. Thus, the large borings described here could
be a previously undescribed ichnotaxon, but further studies on the morphology and variability of the borings are necessary before formal
description is feasible. Possible borings from a nearby section at Künnapõhja of the same age (,
1961) may be conspecific with those found in Saka
(Fig. 3 ![]() ).
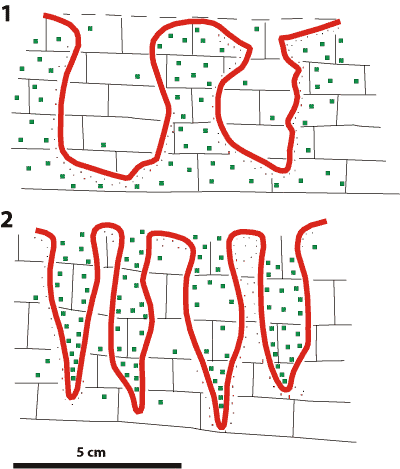
On the other hand, large possible borings from the Väike-Pakri section (northwestern Estonia)
and of the same age
are probably Gastrochaenolites oelandicus
(Fig. 3
).
On the other hand, large possible borings from the Väike-Pakri section (northwestern Estonia)
and of the same age
are probably Gastrochaenolites oelandicus
(Fig. 3 ![]() ).
).
Most examples of Gastrochaenolites are interpreted as domichnia of boring bivalves. Based on the absence of preserved body parts of the tracemaker in some Ordovician examples, it has been suggested that the borer might have been a shell-less, soft-bodied invertebrate of unknown affinity ( & , 2001). It is possible that the maker of the large borings in the Saka hardground was a soft-bodied animal, but different in morphology or behavior from the entity which made G. oelandicus. The biological affinities of the Saka large hardground borers are unclear, but similarity to plug-shaped burrows ( et alii, 1988) such as Conichnus (incl. Amphorichnus) may point to an affinity with actinarian sea anemones. Plug-shaped dominichnia are associated with anemones from the earliest Early Cambrian (, 2003). Thus, some of these burrowers may have been adapted to life in carbonate hardgrounds using chemical boring. Hardgrounds became very common in Ordovician calcite seas ( & , 2003) and this development may correlate with the appearance of first large borers as a part of GOBE (Great Ordovician Biodiversification Event). The Saka borings suggest that the diversity of initial large borers and/or boring behaviors may have been greater than previously thought. Whether this was only a regional phenomenon in the Baltica or was part of a global change will be determined from future studies.
O.V. is grateful to the target financed project (from the Estonian Ministry of Education and Science) SF0180051s08 (Ordovician and Silurian climate changes, as documented from the biotic changes and depositional environments in the Baltoscandian Palaeobasin) and to a Grant (from the Paleontological Society) for financial support.
We are grateful to two reviewers for their constructive suggestions.
J.S., A.A. & J.M. de (2004).- Macroborings (Gastrochaenolites) in Lower Ordovician hardgrounds of Utah: Sedimentologic, paleoecologic, and evolutionary implications.- Palaios, Lawrence, vol. 19, p. 543-550.
A., T., L. & O. (2000).- The Billingen and Volkhov Stages in the northern east Baltic: detailed stratigraphy and lithofacies zonation.- Proceedings of the Estonian Academy of Sciences, Geology, Tallinn, vol. 49, p. 3-16.
A.V., J.V., P.V. & E.A. (1996).- Detailed lithostratigraphy of the Ordovician lower Volkhovian limestone along the eastern part of the BalticLadoga Glint, northwestern Russia.- GFF, Stockholm, vol. 118, p. 1924.
A.A. & R.G. (2001).- Bioerosional innovation for living in carbonate hardgrounds in the Early Ordovician of Sweden.- Lethaia, Oslo, vol. 34, p. 1-12.
W. (1975).- Trace fossils and problematica. In: C. (ed.), Miscellanea.- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part W (Second Edition), 269 p.
S. (2003).- The Proterozoic and Earliest Cambrian trace fossil record: patterns, problems and perspectives.- Integrative and Comparative Biology, McLean, vol. 43, p. 219-228.
S.R.A. & R.G. (1984).- Ichnological nomenclature of clavate borings.- Palaeontology, Oxford, vol. 27, p. 793807.
D.R., N.P. & S.G. (1978).- Initial diversification of macroboring ichnofossils and exploitation of the macroboring niche in the Lower Paleozoic.- Paleobiology, Washington, vol. 4, p. 163170.
R. (1966).- O vertikalnikh norkach zaryvania v ordovikskikh izvestniakakh Pribaltiki [On vertical burrows in the Ordovician limestones of the Peribaltic].- Akademiya Nauk SSSR, Paleontologicheskiy Institut, St. Petersburg, p. 200206 (in Russian).
R. & A.V. (2005).- Trace fossils. In: A., T., & M. (eds.), Cambrian and Ordovician of St. Petersburg Region.- 6th Baltic Stratigraphical Conference, St. Petersburg, p. 33-38.
A.I. (2003).- Ordovician radiation. In: D.E.G. & P.R. (eds.), Palaeobiology II.- Blackwell Publishing, Oxford, p. 49-52.
K. (1960).- O litostratigraphii volkhovskogo i kundaskogo gorizontov v Estonii [On the lithostratigraphy of the Volkhov and Kunda stages in Estonia].- Akademiya Nauk Estonskoi SSR, Geologicheskii Institut, Trudy, Tallinn, vol. 5, p. 45-87.
K. (1961).- Diskontinuiteedipinnad volhovi ja kunda lademes.- Geoloogiline kogumik, Tartu, p. 16-25.
S.G., R.W., & R.G. (1988).- The ichnotaxonomy of Conostichus and other plug-shaped ichnofossils.- Canadian Journal of Earth Sciences, Ottawa, vol. 25, p. 866892.
J.J. Jr & P.M. (1983).- Diversification, faunal change, and community replacement during the Ordovician radiations. In: M.J.S. & P.L. (eds.), Biotic interactions in recent and fossil benthic communities.- Plenum Press, New York, p. 673-717.
J.J. Jr (1995).- The Ordovician radiations: diversification and extinction shown by global genus-level taxonomic data. In: J.D., M.L. & S.C. (eds.), Ordovician odyssey.- Short Papers for the Seventh International Symposium on the Ordovician System, SEPM, California, p. 393-396.
P.D. & M.A. (2003).- Palaeoecology and evolution of marine hard substrate communities.- Earth-Science Reviews, Amsterdam, vol. 62, p. 1-103.
O. (2008).- Stop B3: Baltic Klint at Saka and Valaste. In: O., L., P. & T. (eds.), The Seventh Baltic Stratigraphical Conference.- Abstracts and Field Guide, Geological Society of Estonia, Tallinn, p. 105-106.
M.A. (2007).- Macroborings and the evolution of bioerosion. In: W. III (ed.), Trace fossils: concepts, problems, prospects.- Elsevier, Amsterdam, p. 356-367
M.A. & T.J. (2006).- Patterns and processes in the Ordovician bioerosion revolution.- Ichnos, Philadelphia, vol. 13, p. 109-112.
Y.D., R.B., J.X., J.F. & X. (2010).- Principal aspects of the Ordovician biotic radiation.- Science China Earth Sciences, Beijing, vol. 53, n° 3, p. 382394.
Click on thumbnail to enlarge the image.
Figure 1: Location of the Saka outcrop in northeastern Estonia (red square).

Click on thumbnail to enlarge the image.
Figure 2: A plug-shaped vertical boring (filling sediment removed) cutting a trilobite pygidium (Megistaspis) in a hardground from Saka, at the upper surface of the Päite Member, marking the boundary between the Floian and Dapingian stages. The figured specimen is deposited in the Geological Museum, Museum of Natural History, University of Tartu (TUG): TUG 1405-1.
Click on thumbnail to enlarge the image.
Figure 3: 1) Section of a possible hardground on the Väike-Pakri cliff with borings tentatively assigned to Gastrochaenolites oelandicus, cut into the upper surface of the Päite Member, the boundary between the Floian and Dapingian stages (after , 1961). 2) Section of a possible hardground with large plug-shaped probable borings, like those found in Saka: at the upper surface of the Päite Member, the boundary between the Floian and Dapingian stages (after , 1961). Hardground surface: brown; impregnation by glauconite: green.