![]()
[Introduction]
[Diorygma] [Bibliographic references]
and ... [Figures]
BrachNet, 20, rue Chaix, F-13007 Marseille (France)
Manuscript online since November 30, 2010
![]()
Various ichnotaxa found in hard substrates are interpreted as "phoronid" trace fossils. Their records are briefly reviewed. An interpretation of Diorygma found in the shells of brachiopods is not compatible with phoronid morphology and anatomy. Criteria for the discrimination of phoronid burrows and borings from those of similar organisms from others are difficult to establish even when the evidence and conclusions made therefrom are sound.
Phoronida; borings; Diorygma; Talpina; Skolithos; ichnotaxa.
(2010).- Fossil Phoronida and related ichnotaxa.- Carnets de Géologie / Notebooks on Geology, Brest, Note brève / Letter 2010/03 (CG2010_L03)
Phoronida fossiles et leurs ichnotaxons induits.- Divers ichnofossiles signalés dans des substrats durs ont été interprétés comme correspondant à des traces de phoronidiens. Leurs descriptions sont brièvement discutées, notamment des interprétations de Diorygma et de Eodiorygma. Découverte perforante dans des coquilles de brachiopodes, Diorygma montre une structure incompatible avec la morphologie et de l'anatomie des phoronidiens. Les critères de discrimination entre les tubes et les perforations des phoronidiens avec ceux d'autres organismes semblables sont très difficiles, voire impossibles, à établir, notamment en l'absence de preuve démontrant qu'il s'agit bien d'une action liée à la présence de phoronidiens.
Phoronida ; perforations ; Diorygma; Talpina; Skolithos; ichnotaxa.
The Phoronida , 1888, or at least the securely monophyletic Proto-lophophorates, emerged as a discrete entity some 800-1,000 Ma ago (, 2008). But fossil phoronids known since the Devonian have preserved the ancestral form and way of life very much like those of their predecessor proto-lophophorates (, 1982, 1984).
Several authors have suggested that fossil tubes or tubicolous burrows and borings are trace fossils of Phoronida, and have ascribed them to various ichnogenera, e.g., Skolithos, Talpina, Diorygma, ... ( & , 1934; , 1955; , 1959; , 1972; , 1961; , 2005).
(1975, 1978) provided good evidence that species of Phoronis , 1856, constructed the burrows of the ichnogenus Talpina von , 1840. In the same vein, (1972) had demonstrated that Conchotrema , 1945, is a junior synonym of Talpina, a finding supported by (1994). Therefore, Talpina ranges back to the Late Devonian (, 1911; & , 1970; & , 2003; , 2004) and is well known from the Jurassic ( et alii, 1994) and the Cretaceous (Voigt, 1972). The Talpina organism bored into diverse calcareous substrates such as calcareous algae, corals, echinoid tests, mollusk shells and the rostra of belemnites (, 1972, 1978). gave criteria for the discrimination of phoronid burrows and borings from similar ones: those made by thallophytes, sponges, bryozoans and "worms". (1970) reported the presence of agglutinated foraminifers dated Late Maastrichtian around the aperture of the tube of a worm-like fossil animal, provisionally determined as Phoronopsis , 1907; he suggested that such a conjunction might have been evidence of a commensal relationship between these organisms. However this relationship has not been confirmed in observations of extant Phoronida.
Tubes of the ichnogenus Talpina ramosa von , 1840, which occur frequently in the guards of Belemnella (, 1979) and Belemnitella d', 1840, have also been described in cavities probably of diagenetic origin in aragonitic corals of the Maastrichtian chalk-tuff (, 1978).
(1978) and (1979) suggested that Tentaculites von , 1820 were related to brachiopods or perhaps more cogently to phoronids. However, noted that the lophophorate feeding system of the phoronids is incompatible with the planktonic mode of life proposed for some tentaculitids. (2005), & (2005), and & (2007) hypothesized that phoronids were perhaps the closest living relatives of Cornulites von , 1820, Tentaculites, and Anticalyptraea , 1867. But to reconstruct cornulitids as 'skeletal phoronids' is completely speculative. There are only two groups to which Cornulites can be compared directly: bryozoans and cnidarians. Both have comparable morphologies and shell structures ( et alii, 2007).
Iotuba chengjiangensis, a form known from three specimens in the Lower Cambrian Chengjiang fauna, has been described and interpreted by & (1997) as a phoronid, for it has a U-shaped gut and is tentaculated.
In studies of Cenozoic borings, holes in mollusk shells have been ascribed to phoronids (, 1994; & d', 1987).
The trace fossil Diorygma atrypophilia1 , 1961,
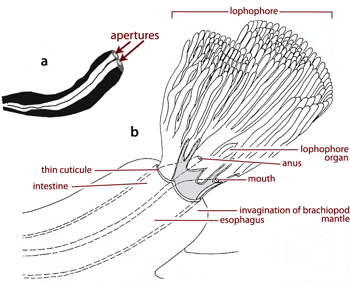
is a tubular protuberance formed by the deposition of a secondary layer of the shell around a vermiform organism housed in the mantle cavity of the atrypide brachiopod Desquamatia subzonata , 1964 from the Givetian,
Middle Devonian (Fig. 1a ![]() ). Although interpreted originally as a result of parasitic boring from the outer surface of the shell (, 1961), the relationships between Desquamatia , 1960,
and Diorygma , 1961,
was interpreted subsequently as being strictly host-specific. Thus, according to
& (1970), Diorygma was considered as having been caused by a phoronid inhabiting the mantle cavity of the brachiopod and sharing its filter-feeding strategy (Fig. 1b
). Although interpreted originally as a result of parasitic boring from the outer surface of the shell (, 1961), the relationships between Desquamatia , 1960,
and Diorygma , 1961,
was interpreted subsequently as being strictly host-specific. Thus, according to
& (1970), Diorygma was considered as having been caused by a phoronid inhabiting the mantle cavity of the brachiopod and sharing its filter-feeding strategy (Fig. 1b ![]() ). During the last few decades the tendency has been to attribute ichno-tubes to fossil phoronids, but the descriptions are rarely subjects of discussion so appear to have been widely accepted. But even forty years later Diorygma merits a short comment regarding several controversial points which were never addressed before.
). During the last few decades the tendency has been to attribute ichno-tubes to fossil phoronids, but the descriptions are rarely subjects of discussion so appear to have been widely accepted. But even forty years later Diorygma merits a short comment regarding several controversial points which were never addressed before.
Each protuberance called Diorygma encloses two contiguous tubes interpreted by
& (1970) as the separated anal and oral tubes of the U-shaped digestive tract of a phoronid. Actually, such a construction has never been described or proved to exist because it is impossible to do so anatomically. During metamorphosis, the phoronid larva named actinotroch (or Actinotrocha , 1846) passes in several minutes from a highly adapted pelagic form to a slender benthic organism organized as a tubicolous adult (see , 1982 and references therein): therefore the adult trunk is formed by eversion of the metasomal sac on the ventral side of the actinotroch (Figs. 2 ![]() - 3
- 3 ![]() ). Consequently, the digestive tract which is attached to the sac wall by the ventral mesentery moves down to occupy its adult U-shaped location. At the same time, mouth and anus are brought into close proximity, and the space between them indicates the location of the dorsal side. In short, the adult structures are emplaced through a 90° rotation of the larval axis. The chitinous tube is secreted during evagination of the metasomal sac which is the cylindrical trunk of the adult (Figs. 2
). Consequently, the digestive tract which is attached to the sac wall by the ventral mesentery moves down to occupy its adult U-shaped location. At the same time, mouth and anus are brought into close proximity, and the space between them indicates the location of the dorsal side. In short, the adult structures are emplaced through a 90° rotation of the larval axis. The chitinous tube is secreted during evagination of the metasomal sac which is the cylindrical trunk of the adult (Figs. 2 ![]() - 3
- 3 ![]() ).
).
The stylized reconstruction of a phoronid (Fig. 1b ![]() ) proposed by
& (1970) depicts a lophophore divided into two parts: the coils are anal and the tentacles at the mouth level are ventral, suggesting a double row of tentacles. In fact, the lophophore of all lophophorates (Phoronida, Bryozoa and Brachiopoda) has a single row of tentacles (, 1976, 1992) (Figs. 3
) proposed by
& (1970) depicts a lophophore divided into two parts: the coils are anal and the tentacles at the mouth level are ventral, suggesting a double row of tentacles. In fact, the lophophore of all lophophorates (Phoronida, Bryozoa and Brachiopoda) has a single row of tentacles (, 1976, 1992) (Figs. 3 ![]() - 4
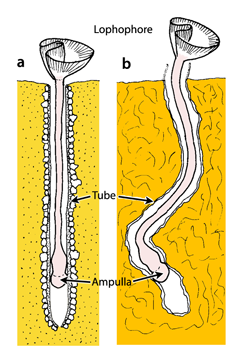
- 4 ![]() ). So these attempted reconstructions are not consonant with the morphology, anatomy and ecology of the Phoronida and they do not appear to be congruent with the longitudinal sections figured by (1961). Furthermore, some phoronid species are boring or encrusting, but their tubes like those of soft substrate dwelling phoronid species are chitinous (Fig. 4
). So these attempted reconstructions are not consonant with the morphology, anatomy and ecology of the Phoronida and they do not appear to be congruent with the longitudinal sections figured by (1961). Furthermore, some phoronid species are boring or encrusting, but their tubes like those of soft substrate dwelling phoronid species are chitinous (Fig. 4 ![]() ) and their outer surface is covered with detritic particles from various sources (see , 1973, 1982; , 1978; , 1979). As a general statement, I reiterate that it is very difficult if not impossible to determine whether or not a phoronid is responsible for a fossil tube or boring. Such a statement may be considered palaeontologically naïve because in palaeontology there are often degrees of certainty about the attribution of both trace and body fossils to any particular biological group. As a biologist, I pointed out that a hypothesis should be supported by a reasonable characterization of the tracemaker. In the present case this attribution to the Phoronidia is based on sound reasoning and evidence from our current knowledge of these organisms. There is a tendency in recent works to attribute ichno-tubes to fossil phoronids (see et alii, 2008) without any reference to the true structure and chemical composition of the tube that may identify them as Phoronida. Moreover, it seems highly speculative to propose a phyletic tree based upon ichno-tubes hypothetically attributed to the Phoronida.
) and their outer surface is covered with detritic particles from various sources (see , 1973, 1982; , 1978; , 1979). As a general statement, I reiterate that it is very difficult if not impossible to determine whether or not a phoronid is responsible for a fossil tube or boring. Such a statement may be considered palaeontologically naïve because in palaeontology there are often degrees of certainty about the attribution of both trace and body fossils to any particular biological group. As a biologist, I pointed out that a hypothesis should be supported by a reasonable characterization of the tracemaker. In the present case this attribution to the Phoronidia is based on sound reasoning and evidence from our current knowledge of these organisms. There is a tendency in recent works to attribute ichno-tubes to fossil phoronids (see et alii, 2008) without any reference to the true structure and chemical composition of the tube that may identify them as Phoronida. Moreover, it seems highly speculative to propose a phyletic tree based upon ichno-tubes hypothetically attributed to the Phoronida.
Finally, the interpretation of & (1970) does not apply to phoronids. Discussing the phoronid relationship to a rather similar structure in Eodiorygma , et , 2004, et alii (2004) wrote: "in some groups are protected distally around the lophophore by a pair of collar folds (, 1979)", but this is a misinterpretation of the collar fold in Phoronopsis as originally described by (1907) – see Phoronida [http://paleopolis.rediris.es/Phoronida/] a WEB site which provides an exhaustive source of data and provides a bibliography updated yearly for each species, both the adult and its actinotroch larva. Many of the species are illustrated by drawings and/or photographs. (2007) indicates: "There is no indisputable fossil evidence of the first appearance of phoronids notwithstanding the phoronid-style of U-shaped borings (Diorygma)". Furthermore, during the last decade, the paper of & (1970) has been quoted as an example of brachiopod bioerosion in at least eight other journals and books, among them & (2002), (2004) who states "this may not be a true trace fossil". & (2005) consider them as possible fossil phoronids.
Nevertheless, among Recent marine groups there are several organisms, e.g., boring bivalve mollusks, capable of creating traces with a double aperture like those of the trace fossil Diorygma. Suspension-feeding organisms do not necessarily have a tentacular apparatus; it is not a requirement for their existence in a brachiopod shell. Suspension feeders or sestonophages require either a sufficiently high density of digestible suspended particles or an adequate supply through water movement. They are grossly classified into two major categories: (i) species which exploit existing water movement, i.e., are unable to generate current on their own; (ii) species which actively produce water movement (, 1982). Thus, mechanisms of suspension feeding are very diverse in marine invertebrates.
1 Diorygma is also a genus of lichenized fungi in the family Graphidaceae.
Grateful thanks to Fernando (Universidad de Oviedo, Spain) and to Karl (Friedrich-Alexander-Universität, Erlangen-Nürnberg, Germany) for their suggestions and to three anonymous referees who reviewed the manuscript. Nestor suggested grammatical and syntactical improvements in the English for which the author is appreciative.
V.V. (1994).- Phoronid borings from the Eocene of the Ukraine [in Russian].- Paleontologicheskii Zhurnal, Moscou, vol. 28, p. 155-160.
M. (1955).- Occurence of fossil Phoronidea-like tubes in several geological formations in Israël.- Bulletin of the Research Council of Israel, Jerusalem, vol. 5, p. 174-177.
M.G., L.E. & L.E. (2004).- The oldest-known metazoan parasite?- Journal of Paleontology, Tulsa, vol. 78, n° 6, p. 1214-1216.
T.K. & F.J. (2002).- Fossil record of parasitism on marine invertebrates with special emphasis on the platyceratid-crinoid interaction.- The Paleontological Society Papers, New Haven, vol. 8, p. 195-209.
G. (1961).- Diorygma atrypophilia n.gen. n.spec. a parasitic organism of Atrypa zonata Schnur.- Acta Palaeontologica Polonica, Warsaw, vol. 6, n° 1, p. 17-28.
R.G. (2004).- A stratigraphy of marine bioerosion.- Geological Society, London, Special Publications, vol. 228, p. 455-479.
R.G. & d' A. (1987).- Bioerosion of the Plio-Pleistocene transgression of southern Italy.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 93, p. 379–442.
J. & G. (1997).- Biology of the Chengjiang fauna.- Bulletin of the National Museum of Natural Science, Taipei, vol. 10, p. 11-105.
B.L. & A. (2005).- Molecular evidence that phoronids are a subtaxon of brachiopods (Brachiopoda: Phoronata) and that genetic divergence of metazoan phyla began long before the early Cambrian.- Organisms Diversity and Evolution, Berlin, vol. 5, n° 4, p. 253-273.
C.C. (1973).- Écologie des Phoronidiens.- Bulletin d'Écologie, Paris, vol. 4, n° 4, p. 339-364.
C.C. (1976).- Le lophophore - structure significative des Lophophorates (Brachiopoda, Bryozoa, Phoronida).- Zoologica Scripta, Oslo, vol. 5, p. 133-137.
C.C. (1979).- British and other Phoronids. In: D.M. & R.S.K. (eds.), Synopses of the British fauna.- Academic Press, London, vol. 13, 57 p.
C.C. (1982).- The biology of Phoronida.- Advances in marine Biology, London, vol. 19, p. 1-89.
C.C. (1984).- On the origin of the Lophophorates. Zeitschrift für zoologische und systematische Evolutionsforschung, Berlin, vol. 22, n° 2, p. 91-94.
C.C. (1990).- Phoronida. In: R.K. & R.G. (eds.), Fertilization, development, and parental care.- Reproductive Biology of Invertebrates, Oxford & IBH, New Dehli, vol. 4, Part B, p. 165-184.
C.C. (1992).- Functional disposition of the lophophore in living Brachiopoda.- Lethaia, Stockholm, vol. 25, p. 291-302.
C.C. (2008).- La naissance du temps et ses conséquences. In: Penser le temps... Collection Orientations et Méthodes, Éditions du Comité des Travaux Historiques et Scientifiques (CTHS), Paris, vol. 11, p. 51-62.
M.A. & C.L. (1934).- Skolithos as a fossil phoronid.- The American Geologist, Minneapolis, vol. 61, p. 341-348.
F.T., T.J. & K.L. (1994).- Growth and disintegration of bivalve-dominated patch reefs in the Upper Jurassic of southern England.- Palaeontology, London, vol. 37, p. 131-171.
J.D. (1907).- New forms of the Hemichordates from South Africa.- Transactions of South Africa Philosophical Society, Cape Town, vol. 17, p. 151-176.
L.G., A.T. & M.P. (2007).- Systematics, shell structure and affinities of the Palaeozoic problematicum Cornulites.- Zoological Journal of the Linnean Society, London, vol. 150, p. 681-699.
K.A. (1959).- Probable cirripede, phoronid and echinoid burrows within a Cretaceous echinoid test.- Palaeontology, London, vol. 1, p. 397-400.
K. (1979).- Silurian tentaculitids from Gotland and Scania.- Fossils and Strata, Oslo, vol. 11, 180 p.
D. & G. (1970).- The probable affinities of the trace fossil Diorygma atrypophilia.- Lethaia, Stockholm, vol. 3, n° 2, p. 163-172.
J.M. (1982).- Ocean management.- In: O. (ed.), Marine Ecology.- Wiley, Chichester, vol. 5, part 1, 642 p.
C.R. (1994).- Jurassic boring phoronids – non-boring insights into fossil record of some soft bodied worms.- Palaeontology Newsletter, London, vol. 24, p. 24.
C. (1979).- Morphology, distribution and role of the epidermal gland in Phoronis psammophila .- Tethys, Marseille, vol. 9, p. 133-136.
J. & R.C. (1970).- Late Devonian-Early Mississippian ichnofossils from westem Montana and northern Utah. In: T.P. & J.C. (eds.), Trace fossils.- Geological Journal Special Issues, Liverpool, vol. 3, p. 407-438.
L. (1954).- Developmental biology of Phoronidea of the Gullmar Fiord area (West coast of Sweden).- Acta Zoologica, Stockholm, vol. 35, 215-257.
C.B., G.A., J.R., L.E. & G.E. (2008).- The scleritome of Eccentrotheca from the Lower Cambrian of South Australia: Lophophorate affinities and implications for tommotiid phylogeny.- Geology, Boulder, vol. 36, n° 2, p. 171-174.
W.P. (2005).- Sedimentology and ichnology of Late Cambrian to Early Ordovician Skolithos sandstone in the Deadwood Formation, northern Black Hills, South Dakota, and southeastern Bear Lodge Mountains, Wyoming.- M.Sc. Degree in Geology, University of Toledo (USA), 220 p.
P.D. & M.A. (2003).- Palaeoecology and evolution of marine hard substrate communities.- Earth-Science Reviews, Amsterdam, vol. 62, n° 1-2, p. 1-103.
A.O. (1911).- A fossil burrowing sponge from the Iowa Devonian.- Bulletin of the Laboratory of Natural History, State University of Iowa, Iowa City, vol. 6, p. 165-166.
K.M. (1978).- Tentaculites: evidence for a brachiopod affinity?- Science, Washington, vol. 201, p. 626-628.
O. (2005).- A new cornulitid genus from the Silurian of Gotland, Sweden.- GFF, Stockholm, vol. 127, p. 205-210
O. & M. (2007).- The tentaculitid affinities of Anticalyptraea from the Silurian of Baltoscandia.- Palaeontology, London, vol. 50, n° 6, p. 1383-1390.
O. & H. (2005).- Observations on the morphology and affinities of cornulitids from the Ordovician of Anticosti Island and the Silurian of Gotland.- Journal of Paleontology, Tulsa, vol. 79, p. 726-737.
E. (1970).- Foraminiferen und (?) Phoronidea als Kommensalen auf den Hartgründen der maastrichter Tuffkreide.- Paläontologische Zeitschrift, Stuttgart, vol. 44, n°1/2, p. 86-92.
E. (1972).- Über Talpina ramosa v. 1840, ein wahrscheinlich zu den Phoroniden gehöriger Bohrorganismus aus der Oberen Kreide, nebst Bemerkungen zu den übrigen bisher beschriebenen kretazischen 'Talpina'-Arten.- Nachrichten der Akademie der Wissenschaften in Göttingen, (II. Mathematisch-physikalische Klasse), vol. 1972, p. 93-126.
E. (1975).- Tunnelbaue rezenter und fossiler Phoronidea.- Paläontologische Zeitschrift, Stuttgart, vol. 49, p. 135-167.
E. (1978).- Phoronidenbaue (Talpina ramosa v. ) aus der maastrichter Tuffkreide.- Natuurhistorisch Genootschap in Limburg, Roermond, vol. 28, p. 3-6.
A. (2007).- Monophyly and intraphylum groupings of the Brachiopoda.- In: P.A. (ed.), Revised Brachiopoda.- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part H, vol. 6, p. 2830-2833.
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Figure 1: a. Longitudinal thin section of Diorygma atrypophilia with the two apertures of the limbs (modified, after , 1961). b. Stylised reconstruction of the phoronid showing its general morphology and inferred life attitude (after & , 1970).
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
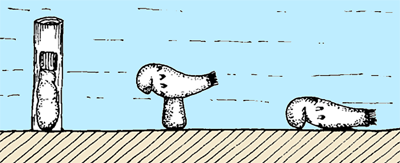
Figure 2: Settlement and metamorphosis of Actinotrocha hippocrepia , 1954 (larva of Phoronis hippocrepia , 1856) on hard calcareous substratum (after , 1954).
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
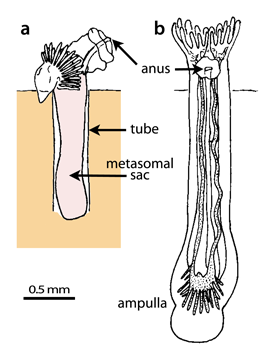
Figure 3: Metamorphosis of Actinotrocha branchiata , 1846 (larva of Phoronis muelleri , 1903) with evagination of the metasomal sac in a soft substratum (a) and a juvenile about one day after metamoprhosis (b) (modified, after , 1990).
Click on thumbnail to enlarge the image.
Cliquer sur la vignette pour agrandir l'image.
Figure 4: Position of the Phoronida in the substrata: (a) embedded vertically in soft sediment; (b) burrowing in hard substrate (modified, after , 1982).