![]()
Sommaire
[Introduction] [Méthode de travail]
[Les peuplements définis]
[Conclusions]
[Perspectives de recherches]
[Références bibliographiques] ET ... [Planches photographiques]
Manuscrit en ligne depuis le 22 Novembre 2010
![]()
L'étude de plus de 40 coupes entaillant les "marnes ŕ Creniceras renggeri" du Jura français (Oxfordien inférieur) a permis :
Les principaux résultats obtenus sont :
Oxfordien inférieur ; ammonites ; biochronostratigraphie fine ; stratigraphie séquentielle ; autoécologie ; synécologie.
R. (2010).- L'évolution des peuplements d'ammonites au cours de l'Oxfordien inférieur (Zone ŕ Mariae et Zone ŕ Cordatum) du Jura (Est de la France).- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/07 (CG2010_A07)
The evolution of ammonite associations during the Early Oxfordian (Mariae and Cordatum zones) in the Jura area (eastern France).- The study of more than 40 sections in the "Creniceras renggeri marls" of the French Jura Range (Lower Oxfordian) has found:
The main results of the study are the following:
Lower Oxfordian; ammonites; high-resolution biochronostratigraphy; sequence stratigraphy; autoecoloy; synecology.
Depuis les travaux de P. de et (1900, 1902), les séries du Jura, datées de l'Oxfordien inférieur, ont été revues ŕ l'occasion d'études consacrées ŕ l'étage Oxfordien dans son ensemble (, 1966). Depuis, la biostratigraphie fine de l'Oxfordien inférieur a été affinée par toute une série de travaux, parmi lesquels on peut citer, principalement : (1986), (1989), et (1994), et alii (1997). Ces auteurs ont proposé de subdiviser la Zone ŕ Mariae en six biohorizons, qui ont été acceptés comme standards pour l'Europe occidentale (GFÉJ, 1997). Plus récemment, l'étude de la coupe de Crotenay (département du Jura, France, et alii, 2000) a permis de reconnaître la totalité de ces horizons dans la chaîne du Jura.
À partir de l'an 2000, j'ai entrepris l'étude de la formation des marnes ŕ fossiles pyriteux ou "Marnes ŕ
Renggeri", datées de l'Oxfordien inférieur sur l'ensemble de la chaîne du Jura (quatre départements). J'ai récolté sur une quarantaine de coupes
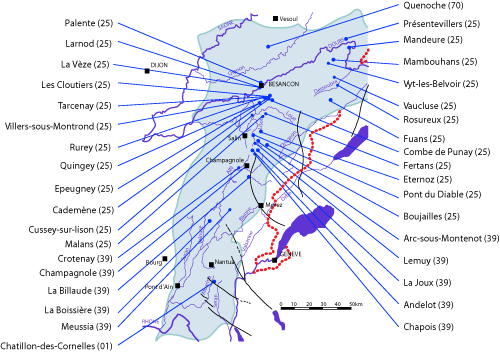
(Fig. 1 ![]() )
plusieurs dizaines de milliers d'ammonites (plus de 90.000 spécimens récoltés dont 78.000 pris en compte dans les statistiques de
population ; Pls. 1
)
plusieurs dizaines de milliers d'ammonites (plus de 90.000 spécimens récoltés dont 78.000 pris en compte dans les statistiques de
population ; Pls. 1 ![]() et 2
et 2 ![]() ) et le découpage biostratigraphique a été élaboré en coordination avec A. , D. et D. . Il en résulte une coupe
synthétique de l'Oxfordien inférieur du Jura valable pour la zone
géographique étudiée oů les marnes ŕ Creniceras renggeri sont
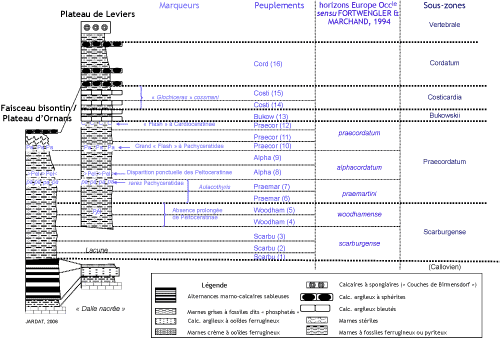
présentes (Fig. 2
) et le découpage biostratigraphique a été élaboré en coordination avec A. , D. et D. . Il en résulte une coupe
synthétique de l'Oxfordien inférieur du Jura valable pour la zone
géographique étudiée oů les marnes ŕ Creniceras renggeri sont
présentes (Fig. 2 ![]() ).
).
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to
enlarge the image.
Figure 1 :
Localisation des gisements.
Figure 1: Location of outcrops.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Figure 2 :
Coupe-type synthétique de l'Oxfordien inférieur du Jura (les épaisseurs
relatives varient selon les gisements et ne sont pas respectées).
Figure 2: Synthetic type section of the Lower Oxfordian of the Jura area
(variations in the thickness of the several sections are not taken into
account).
Le but de cet article est de montrer, non seulement la validité des horizons précédemment définis, mais aussi l'intéręt de l'étude des peuplements ammonitiques pour 1) affiner les découpages biostratigraphiques, 2) proposer si possible une reconstitution des paléoprofondeurs par unité paléogéographique et 3) si possible tenter de corréler les unités biostratigraphiques et celles de la stratigraphie séquentielle. Cette étude trčs détaillée constitue donc un test des hypothčses sur la relation entre peuplements ammonitiques et paléoprofondeurs, proposées par et (1979), (1986, 1992), et les interprétations séquentielles proposées par et alii (1994).
Au sein de chaque gisement, des récoltes ont été effectuées dans des niveaux trčs précis correspondant toujours ŕ un équivalent sédimentaire inférieur ŕ l'extension d'un horizon. Au sein de chaque horizon, cela représente de 4 ŕ 10 prélčvements successifs : lorsque ces prélčvements sont similaires (męme composition en genres et espčces, abondances relatives des divers taxons comparables), ils ont été regroupés en une population unique. Sur l'ensemble du Jura, c'est ainsi prčs de 120 populations différentes qui ont été récoltées et individualisées, chacune d'entre elles renfermant de 150 ŕ 4.000 ammonites. Une quinzaine de populations supplémentaires ne sont représentées que par un faible effectif (de 20 ŕ 150 individus) insuffisant pour une exploitation statistique correcte.
L'analyse statistique des peuplements a été faite en adaptant au Jura les découpages taxonomiques classiquement utilisés dans les "Terres Noires" du bassin du Sud-Est par , et
(1997). Au sein du sous-ordre des Ammonitina, j'ai reconnu une famille
(Pachyceratidae), plusieurs sous-familles (Cardioceratinae, Hecticoceratinae,
Perisphinctinae, Peltoceratinae, Euaspidoceratinae, Taramelliceratinae), parfois des genres (Trimarginites, Creniceras, Scaphitodites, Lissoceratoides) ; au sein du sous-ordre des
Phylloceratina, j'ai distingué le genre Sowerbyceras des autres taxons, regroupés en un seul
ensemble (voir la légende du
Tabl. 3 ![]() ).
).
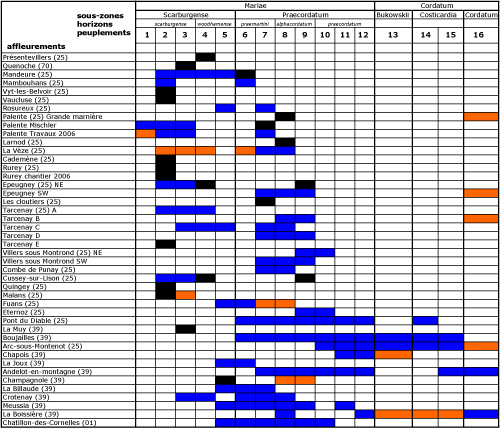
N.B. : il est important de signaler qu'aucune coupe ne couvre l'ensemble de l'intervalle qui sépare l'extręme sommet du Callovien supérieur de l'extręme base de l'Oxfordien moyen. Toutefois, les coupes partielles sont nombreuses et redondantes, ce qui a permis de reconstituer la série complčte par corrélations
successives (voir le
Tabl. 1 ![]() ).
).
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Tableau 1 :
Les populations successives de l'Oxfordien inférieur du Jura - Récapitulatif
des populations étudiées statistiquement et des successions observées. Légende : en orange, niveau reconnu mais population faible ; en
noir, population observée dans le gisement ; en bleu, populations observées en succession sur la męme coupe.
Table 1: Successive populations of the Lower
Oxfordian of the Jura region – A recapitulation of the populations studied
statistically and their successions. Legend: orange: level recognized but
population sparse, black: population in significant numbers in outcrop, blue:
successive populations in one section.
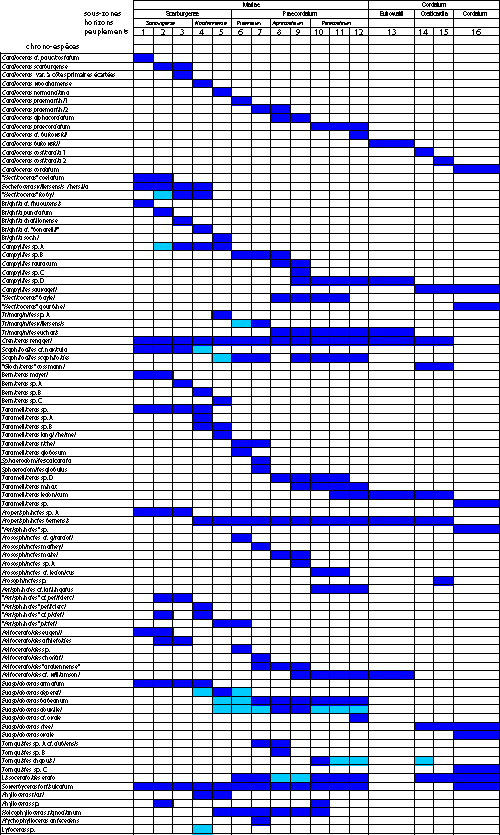
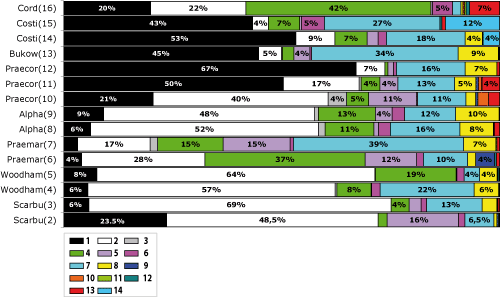
L'examen des taxons d'ammonites récoltés (Tabl. 2 ![]() ), ainsi que l'étude statistique des familles et sous-familles présentes, permet de distinguer 16 peuplements successifs, qui semblent relativement stables pour tout le Jura oů ces niveaux ont pu ętre observés.
), ainsi que l'étude statistique des familles et sous-familles présentes, permet de distinguer 16 peuplements successifs, qui semblent relativement stables pour tout le Jura oů ces niveaux ont pu ętre observés.
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Tableau 2 :
Associations d'ammonites des peuplements Scarbu(1) ŕ Cord(16). Légende : en bleu, formes présentes au sein de chaque récolte
significative et, en bleu clair, formes présentes mais rares.
Table 2: Ammonite associations of populations
Scarbu(1) through Cord(16). Legend: in dark blue
forms present in all significant collections; in light blue forms present only
rarely.
Chaque peuplement est décrit ŕ travers les caractéristiques suivantes :
Marqueurs spécifiques : sont signalées les espčces qui caractérisent avec certitude chaque unité biostratigraphique définie.
Composition : sont signalées les caractéristiques de chaque peuplement en termes d'abondances relatives des différents taxons (familles, sous-familles, genres).
Population-type : correspond ŕ la population la plus abondante et la plus représentative du peuplement ainsi défini.
Peuplement Scarbu(1) : connu sur le seul gisement de Palente-Mischler, il se distingue de la population sus-jacente essentiellement par la présence de Brightia cf. thuouxensis ( et alii), forme proche de l'espčce mais qui possčde des côtes primaires plus resserrées et moins saillantes, associée ŕ des Cardioceras cf. paucicostaum ().
Peuplement Scarbu(2) : ce peuplement a été reconnu dans 14 coupes. Il semble absent au Sud de l'accident de Salins, région marquée par une lacune de sédimentation ŕ la base de l'Oxfordien inférieur (voir , 1966).
Marqueurs spécifiques : Cardioceras scarburgense ( & ), Hecticoceras coelatum (), Berniceras mayeri (de ).
Composition : les peuplements de cet âge sont principalement caractérisés par l'abondance des Cardioceratinae et des Peltoceratinae.
Population-type : la population d'Epeugney N1 (N = 2589 individus,
Tabl. 3 ![]() ).
).
Peuplement Scarbu(3) : cette population a été reconnue dans 10 coupes. Elle n'est que sporadiquement présente au Sud de l'accident de Salins (gisements de Le Muy et Crotenay). La reprise de sédimentation ŕ la base de l'Oxfordien semble trčs irréguličre ŕ cette fine échelle de temps.
Marqueurs spécifiques : Cardioceras scarburgense ( & ) avec certains variants proches de Cardioceras woodhamense () et variants ŕ côtes primaires espacées. Peltoceratinae encore présents ; Hecticoceras coelatum a disparu.
Composition : Les peuplements de cet âge se marquent par une réduction des Cardioceratinae et Peltoceratinae, au bénéfice des Hecticoceratinae.
Population-type : La population de Tarcenay A2b (N = 2196 individus,
Tabl. 3 ![]() ).
).
Peuplement Woodham(4) : ce peuplement a été bien reconnu dans 7 coupes. Il est absent dans certaines localités situées au sud de l'accident de Salins, oů le niveau Woodham(5) surmonte directement la Zone ŕ Lamberti.
Marqueurs spécifiques : Cardioceras woodhamense () abondant, Peltoceratinae absents. Eochetoceras villersensis (d') et "Hecticoceras" kobyi (de ) sont ŕ leur acmé.
Composition : Dominance des Hecticoceratinae, qu'accompagnent des Taramelliceratinae (Taramelliceras) abondants. Au sein des Perisphinctinae, dominance de la lignée des "Perisphinctes" petitclerci-picteti.
Population-type : La population de Cussey-sur-Lison (N = 4287 individus,
Tabl. 3 ![]() ).
).
Peuplement Woodham(5) : ce peuplement a été bien reconnu dans 5 coupes.
Marqueurs spécifiques : Cardioceras woodhamensee cf. var. normandiana (), Taramelliceras langi (de ), microconque de Taramelliceras heimei (de ) ( et alii, 2002), Berniceras sp. nov. (forme dénommée ŕ tort Oppelia mayeri in de , 1900). Apparition d'Euaspidoceras depereti ().
Composition : Se distingue du peuplement Woodham(4) par une inversion des proportions entre Perisphinctinae et Taramelliceratinae.
Population-type : La population de La Joux D7 (N = 2169 individus,
Tabl. 3 ![]() ).
).
Peuplement Praemar(6) : Bien représenté dans les affleurements de l'ensemble du Jura, ce peuplement a été bien reconnu dans 9 coupes.
Marqueurs spécifiques : Cardioceras praemartini () formes primitives, Taramelliceras globosum (de ), apparition de Taramelliceras richei (de ), Scaphitodites scaphitoides (), réapparition des Peltoceratinae. Apparition du genre Prososphinctes au sein des Perisphinctinae. Apparition du brachiopode Aulacothyris.
Composition : Raréfaction des Cardioceratinae, au sein desquels domine le morphe épais ; forte domination des Perisphinctinae qui atteignent leur acmé et abondance des Peltoceratinae ; nette réduction des Hecticoceratinae.
Population-type : La population de La Joux D6 (N = 1426 individus,
Tabl. 3 ![]() ).
).
Peuplement Praemar(7) : Ce peuplement a été reconnu avec certitude dans 16 coupes, d'un bout ŕ l'autre du Jura.
Marqueurs spécifiques : Cardioceras praemartini () formes évoluées, Sphaerodomites, Peltoceratoides choffati (de ), Trimarginites villersensis ( non ), Tornquistes dubiensis ().
Composition : Acmé des Taramelliceratinae, équilibre entre Hecticoceratinae, Perisphinctinae et Peltoceratinae. Cardioceratinae peu abondants, avec morphes minces plus abondants qu'au sein du peuplement précédent. Apparition fugace de Pachyceratidae (Tornquistes dubiensis ()) peu avant le peuplement Alpha(8).
Population-type : La population de Tarcenay C2 (N= 1416
individus,
Tabl. 3 ![]() ).
).
Peuplement Alpha(8) : Cette population a été reconnue avec certitude dans 14 coupes ; elle semble absente au Nord de Besançon.
Marqueurs spécifiques: Cardioceras alphacordatum (), Campylites rauracum ( in de ), Trimarginites eucharis (d'), disparition d'Euaspidoceras depereti () et de Taramelliceras richei (de ), apparition de Taramelliceras sp. nov. et d'Hecticoceras baylei (). Disparition du brachiopode Aulacothyris.
Composition : Permutation majeure entre les Taramelliceratinae + Peltoceratinae et les Hecticoceratinae. Inversion de tendance (augmentation) pour les Cardioceratinae.
Population-type : La population d'Andelot BN2 (N = 2602 individus,
Tabl. 3 ![]() ).
).
Peuplement Alpha(9) : Ce peuplement marque le dernier niveau ŕ fossiles pyriteux de l'Oxfordien sur la quasi-totalité des plateaux situés au Nord de l'accident de Salins. Au-dessus, les niveaux sont stériles sur quelques mčtres, avant de passer aux couches ŕ sphérites. Il a été reconnu avec certitude sur 12 coupes.
Marqueurs spécifiques: Cardioceras alphacordatum () var. ŕ côtes primaires trčs espacées et saillantes, Campylites sp. ŕ côtes élargies, Taramelliceras minax (), réapparition de Scaphitodites scaphitoides ().
Composition : trčs similaire ŕ Alpha(8), mais les Cardioceratinae et les Peltoceratinae augmentent légčrement et annoncent le niveau suivant.
Population-type : La population de Villers NE0 (N= 863 ammonites,
Tabl. 3 ![]() )
)
Peuplement Praecor(10) : reconnu avec certitude sur 7 coupes, situées au Sud de l'accident de Salins, ŕ l'exception d'une mince lentille détectée ŕ Villers-sous-Montrond.
Marqueurs spécifiques : Présence ponctuelle (en "flash") de Pachyceratidae = Tornquistes chapuisi (de ), Cardioceras praecordatum (), Campylites ŕ section épaisse associée ŕ des côtes fines et nombreuses (sp. nov. ?), "Perisphinctes" cf. latilingatus () ŕ tours internes épais, apparition de Peltoceratoides williamsoni () et de Taramelliceras cf. ledonicum (de ).
Composition : La présence ponctuelle de Tornquistes représente 3 ŕ 6% de la population, les Cardioceratinae se développent au détriment des Hecticoceratinae et les Peltoceratinae tendent ŕ remplacer les Perisphinctinae. Les Euaspidoceratinae sont trčs rares.
Population-type : La population d'Andelot CBC (N = 399 ammonites,
Tabl. 3 ![]() )
)
Peuplement Praecor(11) : reconnu avec certitude sur 5 coupes, ce peuplement ne se distingue de Praecor(10) que par la disparition quasi-complčte des Pachyceratidae.
Marqueurs spécifiques: cf. Praecor(10),
Composition : Les Cardioceratinae deviennent le groupe dominant (40% ŕ 50% de l'effectif), au seul détriment des Hecticoceratinae.
Population-type : la population d'Andelot T (N = 677 ammonites,
Tabl. 3 ![]() ).
).
Peuplement Praecor(12) : reconnu sur 5 coupes, ce peuplement précčde de quelques décimčtres le premier banc carbonaté de la série marneuse.
Marqueurs spécifiques: cf. Praecor(10), mais avec des Cardioceratinae qui annoncent Cardioceras bukowskii ().
Composition : Les Cardioceratinae atteignent leur acmé (2/3 de l'effectif), et les Hecticoceratinae leur minimum d'abondance.
Population-type : La population d'Arc-sous-Montenot récoltée dans le dernier mčtre sous le 1er banc carbonaté (N = 152 ammonites,
Tabl. 3 ![]() ).
).
Peuplement Bukow(13) : Reconnu dans 3 coupes, ce peuplement n'est significatif que dans la marničre d'Arc-sous-Montenot. Il est situé entre le premier banc carbonaté signalé ci-dessus et le second banc, appelé "banc bleu" par (1966).
Marqueurs spécifiques : Cardioceras bukowskii () : disparition des morphologies "Scarburgiceras" dans les tours internes, ce qu'avait bien sűr déjŕ noté en 1966 ; Campylites ŕ côtes trčs fines.
Composition : Domination des Cardioceratinae atténuée au profit des Taramelliceratinae ; Peltoceratinae présents et Euaspidoceratinae absents.
Population-type : La population d'Arc-sous-Montenot B1B2 (N = 765 ammonites,
Tabl. 3 ![]() ).
).
Peuplement Costi(14) : n'a été récolté qu'ŕ Arc-sous–Montenot, entre le banc bleu et le banc n° 3 (, 1966).
Marqueurs spécifiques : Cardioceras aff. costicardia (), apparition de Campylites sauvageti (de ), "Glochiceras" cossmanni (de ), apparition d'Euaspidoceras freei ().
Composition : cf. Bukow(13), mais présence des Euaspidoceratinae et apparition des "Glochiceras".
Population-type : La population d'Arc-sous-Montenot B2-B3 (N = 1326 ammonites,
Tabl. 3 ![]() ).
).
Peuplement Costi(15) : reconnu avec certitude dans trois coupes situées sur le plateau de Leviers.
Marqueurs spécifiques : Cardioceras costicardia (), apparition de Creniceras crenatum ( in de , 1922). Apparition de Perisphinctinae dont la morphologie est semblable ŕ celle des formes de l'Oxfordien moyen.
Composition : Acmé de "Glochiceras" cossmanni (de ), disparition des Peltoceratinae et légčre régression des Cardioceratinae.
Population-type : La population récoltée ŕ Andelot-en-Montagne dans les 1,50 m situés sous les couches ŕ sphérites (N = 475 individus,
Tabl. 3 ![]() ).
).
Peuplement Cord(16) : récoltée avec certitude dans la coupe d'Andelot-en-Montagne, au sein des marnes qui s'intercalent entre le 1er et le 3e banc ŕ sphérites, ainsi qu'ŕ La Boissičre (39)
Marqueurs spécifiques : Cardioceras cordatum () (identiques ŕ ceux de la population d'Herznach, voir & , 1986), divers Perisphinctinae, Euaspidoceras ovale (), "Hecticoceras" gourbinei (de ), Tornquistes sp. semblable aux formes figurées par A. de , 1922 (pl. XV, fig. 7), sous le nom de genre Sphaeroceras.
Composition : elle annonce ŕ l'évidence les peuplements de l'Oxfordien moyen ; enrichissement massif en Perisphinctinae et abondance des Lissoceratoides ; les Campylites concurrencent ŕ nouveau les Cardioceratinae.
Population-type : La population récoltée ŕ Andelot-en-Montagne (N = 1148 individus,
Tabl. 3 ![]() ).
).
À Andelot, le dernier banc ŕ sphérites est surmonté de marnes renfermant Cardioceras vertebrale () ŕ morphologie primitive. Peu épais (1m), ce niveau marneux est lui-męme surmonté directement par les facičs ŕ spongiaires dits "Couches de Birmenstorf".
Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Tableau 3 :
Populations types de l'Oxfordien inférieur du Jura. 1 : Cardioceratinae, 2 : Hecticoceratinae /
Ochetoceratinae, 3 : Trimarginites, 4 : Perisphinctinae, 5 :
Peltoceratinae, 6 : Euaspidoceratinae, 7 : Taramelliceras, 8 :
Creniceras, 9 : Scaphitodites, 10 : Pachyceratidae, 11 :
Sowerbyceras, 12 : autres Phylloceratidae, 13 : divers 14 :
"Glochiceras".
Table 3: Typical populations in the Lower Oxfordian of the Jura region. 1 : Cardioceratinae, 2 : Hecticoceratinae /
Ochetoceratinae, 3 : Trimarginites, 4 : Perisphinctinae,
5 : Peltoceratinae, 6 : Euaspidoceratinae, 7 : Taramelliceras,
8 : Creniceras, 9 : Scaphitodites, 10 :
Pachyceratidae, 11 : Sowerbyceras, 12 : other Phylloceratidae,
13 : other diverse forms, 14 : "Glochiceras".
La subdivision proposée pour la Zone ŕ Mariae et la Zone ŕ Cordatum par & (1994) a été retrouvée et ŕ nouveau validée par les peuplements d'ammonites, de la męme façon que dans le Boulonnais par et alii (1993), et dans le Weymouth (Sud de l'Angleterre) par (2000 et données inédites).
Dans le Jura, la précision des récoltes, associée ŕ la grande abondance des ammonites, permet d'envisager de subdiviser l'horizon ŕ woodhamense en deux unités : l'unité ŕ woodhamense s.s. (peuplement Woodham(4)) et l'unité oů abonde la var. normandiana de (peuplement Woodham(5)). Cette subdivision a été retrouvée dans les sondages ANDRA du bassin de Paris, par comparaison avec les faunes du Jura ( et alii, 2006, détermination des ammonites oxfordiennes par et ).
Habituellement difficile ŕ individualiser, la Sous-zone ŕ Costicardia est désormais bien définie grâce ŕ la mise en évidence des deux peuplements successifs Costi(14) et Costi(15), qui s'intercalent entre les niveaux Bukow(13) et Cord(16) connus dans toute l'Europe Occidentale (, 1986).
On peut découper la succession des peuplements ammonitiques en cinq ensembles :
En se servant des hypothčses autécologique et synécologique des ammonites proposées par (1992), on peut interpréter :
On pourra ici remarquer, comme l'avaient déjŕ suggéré & (1997) dans leur étude consacrée au cycle de 2e ordre Bathonien inférieur – Oxfordien inférieur d'Europe occidentale, que l'évolution des peuplements ammonitiques est dans l'ensemble compatible avec les séquences de troisičme ordre de la "charte" séquentielle proposée par EXXON (de et alii, 1993). Une version valable pour toute l'Europe (de et alii, 1998) a été publiée peu aprčs), avec toutefois des décalages ponctuels entre variations fauniques et variations eustatiques. Plusieurs facteurs peuvent ętre invoqués ici pour expliquer de tels décalages. Premičrement, on dispose avec les faunes d'ammonites du Jura d'un enregistrement biostratigraphique plus complet et détaillé que dans les plates-formes et bassins étudiés par ailleurs en Europe occidentale (Bourgogne, Normandie, Alpes du Sud). Deuxičmement, par sa situation de bordure de bassin, le Jura est en terme de paléo-profondeur trčs sensible aux variations eustatiques mais aussi tectoniques. Troisičmement, cette situation de bordure induit pour le Jura des conditions paléo-écologiques spécifiques qui se traduisent par des compositions fauniques d'ammonites trčs particuličres en comparaison d'autres secteurs géographiques. Par conséquent, il devient difficile de paralléliser avec certitude les paléoprofondeurs dont témoignent les peuplements d'ammonites du Jura avec celles des plates-formes et bassins d'Europe occidentale.
Une telle étude, oů la finesse biostratigraphique est associée ŕ une forte biodiversité des faunes, ouvre la possibilité d'analyser paléontologiquement et phylogénétiquement des groupes rares (Perisphinctinae de la lignée des "espčces" petitclerci - picteti) encore mal définis en terme de bio-espčces (Hecticoceratinae) et de compléter, pour la Zone ŕ Mariae, une biozonation parallčle de l'Oxfordien d'Europe occidentale fondée sur les Cardiocertinae et les Peltoceratinae.
La poursuite de récoltes de plus en plus complčtes au sein de chaque compartiment paléo-tectonique permet d'envisager, ŕ terme, 1) l'étude du contrôle tectonique de la sédimentation, avec ses répercussions sur les faunes et 2) de mesurer encore plus finement la sensibilité de peuplements isochrones aux contraintes de la paléogéographie.
Ce travail a été réalisé en collaboration avec les chercheurs de l'UMR CNRS 5561 "Biogéosciences" de l'Université de Dijon. Il n'aurait pas été possible sans les repérages effectués depuis de nombreuses années par Christian , naturaliste d'Ardon prčs Champagnole (Jura), ŕ qui l'on doit la découverte et la redécouverte de plusieurs gisements majeurs du Callovo-Oxfordien jurassien. Philippe a également contribué ŕ l'enrichissement du matériel récolté. L'auteur remercie chaleureusement Didier et Jacques pour leur investissement dans l'amélioration minutieuse du manuscrit.
A. (1939).- The ammonite succession at the Woodham Brick Company's Pit, Akenan Street Station, Buckinghamshire, and its bearing on the classification of the Oxford Clay.- The Quarterly Journal of the Geological Society, London, vol. XCV, part 2, p. 135-222, pls. VIII-XI.
A. (1995, inédit).- Les Aspidoceratidae (Ammonitina) en Europe Occidentale au Callovien supérieur et ŕ l'Oxfordien inférieur.- Thčse de Doctorat de l'Université de Bourgogne, Dijon, 454 p., 15 pls.
A. & P. (1999).- Données nouvelles sur Euaspidoceras depereti (, 1917), Ammonitina (Aspidoceratidae) de la sous-zone ŕ Scarburgense (Oxfordien inférieur).- Revue de Paléobiologie, Genčve, vol. 18, n° 1, p. 259-271.
N.D. (2000).- Ammonite assemblages of the Upper Oxford Clay (Mariae Zone) near Weymouth, Dorset.- Proceedings of the Dorset Natural History and Archeological Society, Dorchester, n° 121, p. 77-100.
P., A., P.-Y., R. & D. (2000).- Une série oxfordienne de référence, ŕ mi-chemin des régions boréales et téthysiennes (Jura central, France).- Revue de Paléobiologie, Genčve, vol. 19, n° 2, p. 381-397, 3 pls.
G., D. & D. (1994).- Tectonique synsédimentaire et variations du niveau marin pendant le dépôt de la formation des Terres noires (Callovien supérieur - Oxfordien moyen ; Bassin du Sud-Est, France).- Comptes Rendus de l'Académie des Sciences, Paris, (Série II), t. 319, p. 559-565.
S. & D. (1979).- Réflexions sur la répartition des Ammonoďdés dans l'Est et le Sud du Bassin parisien au Callovien supérieur (zone ŕ Lamberti) et ŕ l'Oxfordien inférieur (zone ŕ Mariae).- Bulletin du Bureau de Recherches Géologiques et Miničres, Orléans, (2čme série), section IV, Géologie générale, n° 2, p. 103-112.
R. (1914).- Études sur les Oppeliidées de Dives et Villers-sur-Mer.- Mémoire de Paléontologie, Mémoires de la Société Géologique de France, Paris, n° 48, 26 p., Pls. I-II.
R. (1966).- L'Oxfordien dans la moitié Sud du Jura français. Étude stratigraphique.- Nouvelles Archives du Museum d'Histoire Naturelle de Lyon, n° 8 (t. 1-2), 624 p.
D. (1989).- Les "Terres noires" d'âge Callovien supérieur ŕ Oxfordien moyen des chaînes subalpines du Sud (Diois, Baronnies, Dévoluy) : nouvelles données biostratigraphiques.- Comptes Rendus de l'Académie des Sciences, Paris, (Série II), t. 308, p. 531-536.
D. & D. (1994).- Nouvelles unités biochronologiques de la zone ŕ Mariae (Oxfordien inférieur).- Géobios, Villeurbanne, mémoire spécial n° 17, p. 203-209.
D., D. & A. (1997).- Les coupes de Thoux et de Savournon (SE de la France) et la limite Callovien-Oxfordien.- Géobios, Villeurbanne, n° 30, fasc. 4, p. 519-540.
P.-C. de, G., T., T., D., R. & P.R. (1993).- Depositional sequence cycles, transgressive-regressive facies cycles, and extensional tectonics; example from the southern subalpine Jurassic basin, France.- Bulletin de la Société géologique de France, Paris, vol. 164, p. 709-718.
P.-C. de, J., T. & P.R. (1998).- Mesozoic and Cenozoic sequence stratigraphy of European basins.- Society of Economic Paleontologists and Mineralogists, Special Publication, Tulsa, n° 60, p. 3-13 ; Appendix : p. 763-781 ; Mesozoic and Cenozoic chronostratigraphic charts n° 1 ŕ 8.
A. de (1922).- L'Oxfordien moyen des environs de Niort.- Bulletin de la Société géologique de France, Paris, (Série 4), t. XXI, fasc. 7-9, p. 297-316, pl. XV.
R.A. (1992).- The Oxfordian ammonites succession near Liesberg BE et Péry BE, northern Switzerland.- Eclogae geologica Helvetiae, Basel, vol. 83 n° 1, p. 177-199.
R.A. & D. (1986).- Les faunes de Cardioceratinae (Ammonoidea) du Callovien terminal et de l'Oxfordien inférieur et moyen (Jurassique) de la Suisse septentrionale : stratigraphie, paléoécologie, taxonomie préliminaire.- Géobios, Villeurbanne, n° 15, fasc. 4, p. 239-254.
P. de & A. (1900).- Étude sur les mollusques et brachiopodes de l'Oxfordien inférieur du Jura lédonien.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 27, 196 p.
P. de & A. (1902).- Étude sur les mollusques et brachiopodes de l'Oxfordien supérieur et moyen du Jura lédonien.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 19-21, 303 p.
P. de & E. (1896-1897).- Étude sur les mollusques et brachiopodes de l'Oxfordien supérieur et moyen du Jura bernois.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 23-24, 158 p.
P. de & E. (1898-1899).- Étude sur les mollusques et brachiopodes de l'Oxfordien inférieur du Jura bernois.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 25-26, 220 p.
V. (1928).- Contribution ŕ la connaissance de la faune des marnes ŕ Creniceras renggeri, dans la Franche-Comté septentrionale. Étude sur les Oppeliidés.- Travaux du Laboratoire de Géologie de la Faculté des Sciences de Lyon, fasc. XII, mém. 10, 60 p., 3 pls.
D. (1986, inédit).- L'évolution des Cardioceratinae d'Europe Occidentale dans leur contexte paléobiogéographique.- Thčse Doctorat es Sciences de l'Université de Bourgogne, Dijon, 601 p., 22 pls.
D. (1992).- Ammonites et paléoprofondeur : les faits, les interprétations.- Paleovox, Lyon, n° 1, p. 49-68.
D. & R. (1992).- Les ammonites du niveau vert de Zalas (Oxfordien inférieur, Pologne du Sud) : condensation ou concentration de faunes.- Bulletin of the Polish Academy of Sciences, Earth Sciences, Warszawa, vol. 40, n° 1, p. 55-65.
D., P., A., J. & Q. (2002).- Very small ammonites (micromorphs) from Lower Oxfordian marls (Mariae zone). In : H., K. & A. (eds.), Cephalopods – Present and Past.- Abhandlungen der Geologischen Bundesanstalt, Wien, Band 57, p. 467-478, 1 pl.
D. & J. (1997).- Enregistrement des variations morphologiques et de la composition des peuplements d'ammonites durant le cycle régressif/transgressif de 2e ordre Bathonien inférieur-Oxfordien inférieur en Europe occidentale.- Bulletin de la Société géologique de France, Paris, vol. 168, n° 2, p. 121-132.
L.F. (1939).- The Ammonite zones of the Upper Oxford clay of Warboys Huntingtonshire.- Bulletin of the Geological Survey of Great-Britain, London, n° 1, p. 82-96.
J., D., D., A. & R. (2006).- Les ammonites du Callovien-Oxfordien des sondages ANDRA dans l'Est du bassin de Paris : synthčse biochronostratigraphique, intéręts paléoécologique et paléobiogéographique.- Comptes Rendus Geoscience, Paris, n° 338, p. 834-853.
J.-P., D., A. & D. (1993).- The Callovian and Oxfordian of the Boulonnais area in Northern France: new biostratigraphic data.- Acta Geologica Polonica, Warszawa, vol. 43, n° 3-4, p. 169-182.

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 1 :
1a-c Cardioceras cf. paucicostatum (), peuplement
Scarbu(1), Palente-Mischler ; 2a-c Cardioceras scarburgense ( & ), peuplement
Scarbu(2), Rurey C ; 3a-c Cardioceras scarbugense var. ŕ côtes primaires espacées, peuplement
Scarbu(3), Palente T3 ; 4a-c Cardioceras woodhamense (), peuplement
Woodham(4), Crotenay A1b ; 5a-c Cardioceras woodhamense var. normandiana (), peuplement
Woodham(5), La Joux B4 ; 6a-c Cardioceras praemartini () variété primitive, peuplement
Woodham(6), Boujailles ; 7a-c Cardioceras alphacordatum (), peuplement Alpha(8), Andelot BN2 ; 8a-c Cardioceras alphacordatum () variété ŕ côtes primaires trčs espacées et saillantes, peuplement Alpha(9), Villers-sous-Montrond NE0 ; 9a-c Cardioceras praecordatum (), forme typique, peuplement
Praecor(11), Andelot T ; 10a-b Cardioceras praecordatum (), spécimen annonçant Cardioceras bukowskii (), peuplement
Praecor(12), Andelot T ; 11a-b Cardioceras bukowskii (), peuplement
Bukow(13), Arc-sous-Montenot B1B2 ; 12a-c Cardioceras costicardia (), peuplement
Costi(14), Arc-sous-Montenot B3B4 ; 13a-c Cardioceras costicardia (), peuplement
Costi(15), Boujailles ; 14a-c Cardioceras costicardia (), peuplement
Costi(15), Arc-sous-Montenot B4-Sph ; 15a-c Cardioceras costicardia (), peuplement
Costi(14) ŕ Costi(15), Arc-sous-Montenot B ; 16a-c Cardioceras cordatum (), peuplement
Cord(16), Andelot ; 17a-c idem, variant proche des formes de la Sous-zone ŕ Vertebrale ; 18a-c idem
(toutes les échelles graphiques représentent 1 cm).
Plate 1: 1a-c Cardioceras cf. paucicostatum (),
Scarbu(1) population at Palente-Mischler; 2a-c Cardioceras scarburgense ( & ),
Scarbu(2) population at Rurey C; 3a-c Cardioceras scarbugense var. with widely spaced primary ribs,
Scarbu(3) population at Palente T3; 4a-c Cardioceras woodhamense (),
Woodham(4) population at Crotenay A1b; 5a-c Cardioceras woodhamense var. normandiana (),
Woodham(5) population at La Joux B4; 6a-c Cardioceras praemartini () primitive
variety,
Woodham(6) population at Boujailles; 7a-c Cardioceras alphacordatum (),
Alpha(8) population at Andelot BN2; 8a-c Cardioceras alphacordatum ()
variety with ribs very widely spaced and raised,
Alpha(9) population at Villers-sous-Montrond NE0; 9a-c Cardioceras praecordatum (),
typical form,
Praecor(11) population at Andelot T; 10a-b Cardioceras praecordatum (),
specimen grading toward Cardioceras bukowskii (),
Praecor(12) population at Andelot T; 11a-b Cardioceras bukowskii (),
Bukow(13) population at Arc-sous-Montenot B1B2; 12a-c Cardioceras costicardia (),
Costi(14) population at Arc-sous-Montenot B3B4; 13a-c Cardioceras costicardia (),
Costi(15) population at Boujailles; 14a-c Cardioceras costicardia (),
Costi(15) population at Arc-sous-Montenot B4-Sph; 15a-c Cardioceras costicardia (),
population from Costi(14) to Costi(15) at Arc-sous-Montenot B; 16a-c Cardioceras cordatum (),
Cord(16)
population at
Andelot; 17a-c idem, a
variant resembling the forms of the Vertebrale
Subzone; 18a-c idem
(all the scales represent a length of 1 cm).

Cliquer sur la miniature pour agrandir l'image.
Click on thumbnail to enlarge the image.
Planche 2 :
1a-c Brightia cf. thuouxensis ( et alii), peuplement
Scarbu(1), Palente-Mischler ; 2a-c "Hecticoceras" coelatum (), peuplement
Scarbu(2), Epeugney 1 ; 3a-c Berniceras mayeri (de ), peuplement
Scarbu(2), Rurey C ; 4a-c "Perisphinctes" petitclerci (de ), peuplement
Scarbu(3) ŕ Woodham(4), Crotenay AR ; 5a-c Eochetoceras villersensis (d')
morphe microconque hersilia (d'), peuplement
Woodham(4), Cussey-sur-Lison 2 ; 6a-c "Hecticoceras" kobyi (de ), peuplement
Woodham(4), Crotenay A2 ; 7a-f Taramelliceras langi (de ), peuplement
Woodham(5), Rosureux B0 ; 8a-c Berniceras sp. nov., peuplement
Woodham(5), La Joux D14 ; 9a-d Trimarginites villersensis ( non ), peuplement
Praemar(7), Combe de Punay ; 10a-b Peltoceratoides choffati (de ), peuplement
Praemar(7), Combe de Punay ; 11a-c Trimarginites eucharis (d'), peuplement Alpha(8), Tarcenay D1 ; 12a-c
"Hecticoceras" baylei (), peuplement Alpha(9), Villers-sous-Montrond NE ; 13a-c Campylites sp. nov. ? forme ŕ section épaisse ornée de côtes fines et nombreuses, peuplement
Praecor(11), Arc-sous-Montenot ; 14a-b Tornquistes chapuisi (de non ),
peuplement
Praecor(10), Arc-sous-Montenot ; 15a-e "Glochiceras" cossmanni (de ), peuplement
Costi(15), Pont du Diable ; 16a-b Tornquistes sp., peuplement Cord(16), Andelot ; 17a-c Euaspidoceras ovale (), peuplement
Cord(16), Andelot (toutes les échelles graphiques représentent 1 cm).
Plate 2: 1a-c Brightia cf. thuouxensis ( et alii),
Scarbu(1) population at Palente-Mischler; 2a-c "Hecticoceras" coelatum (),
Scarbu(2) population at Epeugney 1; 3a-c Berniceras mayeri (de ),
Scarbu(2) population at Rurey C; 4a-c "Perisphinctes" petitclerci (de ),
population from
Scarbu(3) to Woodham(4) at Crotenay AR; 5a-c Eochetoceras villersensis (d')
microconch form of hersilia (d'),
Woodham(4) population at Cussey-sur-Lison 2; 6a-c "Hecticoceras" kobyi (de ),
Woodham(4) population at Crotenay A2; 7a-f Taramelliceras langi (de ),
Woodham(5) population at Rosureux B0; 8a-c Berniceras sp. nov.,
Woodham(5) population at La Joux D14; 9a-d Trimarginites villersensis ( non ),
Praemar(7) population at Combe de Punay; 10a-b Peltoceratoides choffati (de ),
Praemar(7) population at Combe de Punay; 11a-c Trimarginites eucharis (d'),
Alpha(8) population at Tarcenay D1; 12a-c
"Hecticoceras" baylei (),
Alpha(9) population at Villers-sous-Montrond NE; 13a-c Campylites sp. nov. ?
a form with a thick section ornamented with numerous narrow fine
ribs, Praecor(11) population at Arc-sous-Montenot; 14a-b Tornquistes chapuisi (de non ),
Praecor(10) population at Arc-sous-Montenot; 15a-e "Glochiceras" cossmanni (de ),
Costi(15) population at Pont du Diable; 16a-b Tornquistes sp., Cord(16) population at Andelot; 17a-c Euaspidoceras ovale (),
Cord(16) population at Andelot (all the scales represent a length of 1
cm).