![]()
Contents
[Introduction] [Materials and methods]
[Historical overview ...]
[Systematic paleontology]
[Geographical and stratigraphical distribution ...]
[Mode of life ...]
[Phylogeny]
[Homoeomorphy]
[Conclusion]
[Appendix] [Bibliographic references] and ... [Table]
Department of Geology, University of Pécs, 6 Ifjúság Street, H-7624 Pécs (Hungary);
Corresponding address: 63 Fenyofa Street, H-2030 Érd (Hungary)
Manuscript online since December 24, 2010
![]()
The Engonoceratidae may well have originated on the shallow marine carbonate platforms of the southern margin of the Mediterranean Province of the Tethyan Realm during the
earliest Albian (?latest Aptian). In the entire Tethyan Realm and beyond adaptive radiation of the group was rapid and successful during the early Albian as indicated by endemic centres in the Peruvian Basin and the Western Interior Sea (USA). Later the group successfully enlarged its distribution, and invaded some provinces
(Mowry Sea, Canada) of the Boreal Realm. The mode of life of engonoceratids seems to have been
nektoplanktonic, epipelagic, and stenohaline, restricted to shallow water and platform or in some cases extremely shallow (littoral and
lagoonal) facies which may have helped their radiation. The group is distinctive and consists of nine genera and up to a hundred species, although its origin is still obscure. Their appearance may have been triggered by the oceanic anoxic event
(OAE 1b) and their rapid rise may have been helped by their shallow water mode of life and the global mid-Cretaceous warming and rise in sea level. The fall of the engonoceratids coincides with the
end-Cenomanian rapid transgression, which may have changed their shallow water habitats. Today the group is considered to have been a successful colonizer, a reliable stratigraphical indicator for shallow marine environments and a pioneer taxon for recognition of transgressive phases.
Based on new material and sutural analysis, Engonoceras duboisi is revised and assigned to Parengonoceras. A set of features consisting of a simplified ceratitid suture, lanceolate and compressed oxycone shell together comprising a shallow marine ecotype is pointed out as a successful and repeated morphotype among Mesozoic
Ammonoidea, for it was repeated three times during ammonoid evolution.
Ammonites; Albian; Cenomanian; phylogeny; systematics; homeomorphy.
L. (2010).- Systematics, phylogeny and homeomorphy of the Engonoceratidae , 1900 (Ammonoidea, Cretaceous) and revision of Engonoceras duboisi , 1989.- Carnets de Géologie / Notebooks on Geology, Brest, Article 2010/08 (CG2010_A08)
Systématique, phylogénie et homéomorphie
des Engonoceratidae , 1900
(Ammonoidea, Crétacé) et révision de Engonoceras duboisi
, 1989.- Les Enganoceratidae apparaissent à l'Albien basal (ou peut-ętre dčs l'Aptien terminal) dans les séries de
plates-formes carbonatées de la marge sud de la province méditerranéenne du domaine téthysien. Leur efficacité évolutive leur permet de coloniser rapidement l'ensemble du domaine téthysien et męme au delà comme le montre la présence de groupes endémiques dans le Bassin Péruvien et dans la mer intérieure occidentale des États-Unis. Plus tard, le groupe élargira encore plus son aire de distribution en envahissant certaines régions du domaine boréal (mer de Mowry, Canada).
Les Enganoceratidae semblent avoir eu un mode de vie nectoplanctonique, épipélagique et sténohalin au sein de mers chaudes peu à trčs peu profondes (rivages, lagons), dont la fréquence a favorisé leur évolution. Ce groupe, dont l'origine est obscure, est bien individualisé. Il comprend neuf genres et une centaine d'espčces. Leur apparition semble avoir été provoquée par la crise anoxique OAE 1b et leur rapide expansion favorisée par l'extension progressive des mers chaudes et épicontinentales qui accompagne la montée des eaux du milieu du Crétacé. Le déclin des Enganoceratidae semble lié à l'accélération de la montée des mers qui caractérise la fin du Cénomanien, sans doute par modification substantielle de leurs habitats peu profonds. Ce groupe est considéré comme un colonisateur efficace et un bon indicateur de milieux de faible profondeur en ambiance transgressive.
À partir de nouvelles études stratigraphiques et structurales, il est proposé une révision de l'espčce
Engonoceras duboisi qui doit ętre rattachée au genre Parengonoceras. Par ailleurs, il est mis en évidence que le fait de présenter des sutures simplifiées de
type Cératite et une coquille lancéolée à oxycône comprimé est symptomatique
d'ammonites adaptées à des milieux marins peu profonds. Ce modčle est, en effet, répété trois fois au long de l'évolution des Ammonoďdes
mésozoďques.
Ammonites ; Albien ; Cénomanien ; phylogénie ; systématique ; homéomorphie.
Engonoceratid ammonites form a distinct but perplexing group among Cretaceous Ammonoidea. Their systematics is still confusing and their origin is obscure. After the period of great monographs on engonoceratid ammonites (, 1903; , 1907; , 1910; , 1940; in et alii, 1947; , 1953; , 1955; , 1956; , 1970, 1982; , 1979), twelve new species have been introduced and forty species invalidated. This prompts a review of the group, and a discussion on the ecology and facies dependency of certain of its taxa. Recent debates on the systematics of Engonoceratidae also invite a review of the systematic position, ecology, distribution, phylogeny and homoeomorphy of this group of fossils and of the species assigned it.
The following abbreviations indicate the repositories of specimens mentioned in the text: HNHM, Hungarian Natural History Museum, Budapest, Hungary; LC, Lumachella Collection (private collection of Mr. Zoltán , 29 Szatmár Street, Budapest, H-1194 Hungary); MHNG-GEPI, Collection Muséum d'Histoire naturelle de Genčve, Switzerland. Suture terminology is after & (1970): E, external lobe; L, lateral lobe; U, umbilical lobe; I, internal lobe. Note that et alii (2003) revised the sutural terminology of the Paleozoic prolecanitid ammonites, which implies its modification for Mesozoic ammonoids too, but this modification has not been published yet. All dimensions are in millimetres: D, diameter; Wb, whorl breadth; Wh, whorl height; U, umbilicus; myr: million year. Species names in brackets indicate invalidation.
The name Ceratites was introduced by de (1825) for a group of Triassic ammonites with simplified suture lines. As early research developed, Ceratites-like fossils of Cretaceous age were discovered, so early palaeontologists included these fossils into the ceratitid group, naming them as Cretaceous Ceratites (, 1848, p. 30) or "pseudo-ceratites based on an apparent resemblance between the sutures of Triassic ceratitids and those of Cretaceous engonoceratids" (, 1903; G. , 1940a, 1940b; & , 1976; , 1981; et alii, 1998a). The first engonoceratid ammonite was published by d' (1841) as Ammonites Vibrayeanus [= Neolobites vibrayeanus] from France, and by (1848) as Ammonites Syriacus [= Knemiceras syriacum] from Lebanon, and Ammonites pierdenalis [= Engonoceras pierdenalis] from Texas. (1848) also called attention to the importance of the group referring to them as Cretaceous ceratitids ('Kreide-Ceratit'). Although (1848) reported other "ceratitids" (whose suture lines superficially resemble those of the Triassic Ceratites) such as Ammonites Ewaldi and Ammonites Robini [= Metatissotia ewaldi and M. robini], these species belong to discrete taxa (Acanthoceratoidea) with no direct phyletic connection to the engonoceratids. The family name Engonoceratidae was introduced by (1900, p. 585) to include Protengonoceras, Engonoceras, Metengonoceras, and Neolobites. Later (1903, p. 144) established the family Knemiceratidae for Knemiceras. Based on the suture line (1912, 1928, 1931) placed the group in the Pulchelliidae, an assignment accepted by G. (1940b, p. 1066). (1922, 1924) introduced Hypengonoceras, and Parengonoceras. The last described member of the family is Platiknemiceras (, 1954).
In the mid 1900's (1941) and (1942) referred certain placenticeratid ammonites (Placenticeras mintoi, and P. planum) to the Engonoceratidae and (1956, p. 486) regarded Neolobites as incertae sedis again an indication of taxonomic uncertainty. Hypengonoceras was regarded as a placenticeratid ammonite (, 1960) at that time the only known Lower Cretaceous placenticeratid. Later (1970) summarized the systematic debates and content of the family, and & (1989, p. 365) revised the systematic position of the most discussed member, Hypengonoceras, which they regarded as having been derived probably from Parengonoceras. et alii (1996, p. 126) did not accept this rationale proposing a different origin for the genus: "Hypengonoceras, despite some resemblances to Engonoceratidae, seems to belong here" [= Placenticeratidae , 1900] and confirmed that the family had an uncertain origin, "but some genera closely resemble Pulchelliidae but are probably not closely related" ( et alii, 1996, p. 130) and placed the family in the Hoplitoidea with these eight genera included: (Engonoceras, Hypengonoceras, Knemiceras, Metengonoceras, Neolobites, Parengonoceras, Platiknemiceras, and Protengonoceras). At present, the systematic position of Hypengonoceras is again disputed ( & , 2004). (2002) resurrected the genus Glottoceras 1875, a view supported by & (2004, p. 20). As yet there is no unequivocal approach regarding the systematic position of the group. (1968, p. 747) had already emphasized the impossibility of placing the group properly until its sutural development is clearly known. He refers this family doubtfully to the Pulchelliidae or the Hoplitidae. (1970) gave a detailed analysis of the engonoceratid sutural history. In the past forty years many species were introduced but no significant work was done on the sutural ontogeny of the Engonoceratidae. Although & (2004) discussed the systematic position of the group placing it in the Pulchelliatoidea, this attribution is not yet widely accepted. et alii (2004), (2004), & (2005), et alii (2007), and et alii (2008) place the group in the Hoplitoidea, while (2008) refers it to the Pulchelliatoidea. Most recently, (2010, p. 169) rejects the pulchelliatoid origin of Engonoceratidae, suggesting that the genus Subpulchellia/Mogharaeceras is most likely an endemic offshoot of Barremites so cannot be the ancestor of the Engonoceratidae. Further uncertainty is indicated by (in , 2008b, p. 160) regarding the systematic position of European Engonoceras sensu stricto. & (2004, p. 11) stressed a pulchelliatoid origin based on a probable relation with the Barremian/Aptian Subpulchellia, but did not include a thorough discussion. (2010) proposes a new taxonomic rank for engonoceratid ammonites, the Superfamily Engonoceratoidea , 1900. This taxonomic approach has certain advantages: on one hand it emphasizes the integrity of the group, thus highlighting its morphological independence. On the other hand this approach leaves open the problem of the origin of the group. As mentioned, (2010) rejected the pulchelliatoid origin of engonoceratid ammonites. If so, the most plausible explanation for its origin is the hoplitoids. This view may be strengthened by new data on the ecology of Aptian Parahoplites ( et alii, 2009). Consequently, the systematics and origin of Engonoceratidae are still open issues. Recently Engonoceratidae was considered to consist of nine genera (Engonoceras, Glottoceras, Hypengonoceras, Knemiceras, Metengonoceras, Neolobites, Parengonoceras, Platiknemiceras, and Protengonoceras), and 143 species (Table 1) of which 38 are invalid. They are listed in the section discussing the described genera and their species. The genus Pseudengonoceras referred to by (1981, p. 517) is a typing error for Parengonoceras (P. , pers. comm., 2009).
Order Ammonitida , 1884
Suborder Ammonitina , 1889
Superfamily Hoplitoidea , 1890
Family Engonoceratidae , 1900
= Knemiceratidae , 1903, p. 144; Neolobitinae & 1958, p. 125
Genus Glottoceras , 1875, p. 372
(Fig. 1 ![]() )
)
Type species. Glottoceras attenuatum , 1875, from the Lower Albian of Peru.

Click on thumbnail to enlarge the image.
Figure 1: Glottoceras attenuatum (, 1903). Reproduction of the original specimen of (1903, pl. 17, figs. 13-14).
Diagnosis. Typical engonoceratid coiling with narrow umbilicus. Cross section is rather compressed and pentagonal, or oval in some species. Venter is flat to slightly concave or rounded in adult stage. Ornamentation consists of rather strong, sometimes coarse prorsiradiate ribs, which may originate at the umbilical shoulder from bullae. Ventrolaterally bullae may also appear. Suture consists of many adventive and auxiliary elements. Relative breadth of saddles and lobes are similar. Saddles are bifid, lobes are finely frilled.
Occurrence. Glottoceras (sensu , 2002) is restricted to the Albian of South America.
Included species:
Remarks. (1956, p. 449) had already pointed out that South American 'Knemiceras' are different from the Knemiceras attenuatum described by (1940). On the basis of Art. 23.9 of (1999), (2002) resurrected Glottoceras to group the South American species of Knemiceras. (2002) and & (2004, p. 20) consider the genus Glottoceras to be restricted to these Andean species. In the assignment of species to this group, Knemiceras andinum must be included in Glottoceras if the rationale of & (2004, p. 20) is accepted.
Genus Engonoceras & ,
1881, p. 140
= Engonhoplitoides , 1940, p. 441; Epigonoceras ,
1956, p. 400
(Fig. 2 ![]() )
)
Type species. Ammonites pierdenalis , 1848, from the Cretaceous of Fredericksburg, Texas.

Click on thumbnail to enlarge the image.
Figure 2: Engonoceras pierdenalis (, 1848). Reproduction of the original figure of (1848, p. 31, pl. 6, figs. 8-10).
Diagnosis. Diagnosis is after et alii (1996, p. 130): "Involute, inner whorls very compressed with narrow, flat or sulcate venter; later flexuous striae normally repeated by weak, flat, flexuous or straight ribs ending in small ventrolateral clavi placed alternately and, in some shells, joined across venter by zigzagging ribs; umbilical and lateral tubercles may be present, and venter of last whorl may be rounded. Suture with more elements than Knemiceras and with saddles normally all entire except that outermost are bifid; external lobe normally with strongly divergent branches."
Occurrence. Engonoceras is widespread in its distribution. It is abundant with many species in North America (Colorado, Kansas, New Mexico, Oregon, Texas) and in Mexico (Sonora) and South America (Colombia, Peru) but also in the Tethys: North Africa (Algeria, Morocco, Tunisia), the Near East (Lebanon, Syria) and Asia (Borneo). It is also reported from Europe (England). Stratigraphically it ranges from the Early Albian to the Middle Cenomanian.
Included species:
Remarks. Although (1848, pl. 6, fig. 10) in his description of the type species indicates that all the saddles of the suture are bifid: "the saddle is rounded, however with one small secondary lobe in the middle", subsequent designations refer to entire or nearly entire saddles with only one bifid element being typical for Engonoceras ( et alii, 1996; et alii, 1998a, p. 5). According to (1956, p. 444) most species of Engonoceras have sutures with entire saddles and are ornamented with three or two rows of tubercles. In some species (e.g. E. complicatum) there are more (up to five) bifid or sometimes trifid saddles (E. elegans). Saddles are generally rounded, lobal necks are narrow. Saddles are always broader than lobes. (1940, p. 441) established Engonhoplitoides on the basis of the subdivision of the lateral lobe of the suture line, remarking that the shell morphology of Engonhoplitoides is closer to that of Engonoceras, while its suture line is more nearly like that of Knemiceras. This separation was discredited by et alii (1996).
Genus Neolobites , 1882, p. 389
(Fig. 3 ![]() )
)
Type species. Ammonites Vibrayeanus d', 1841, p. 322, from the Cretaceous of Sarthe (France).

Click on thumbnail to enlarge the image.
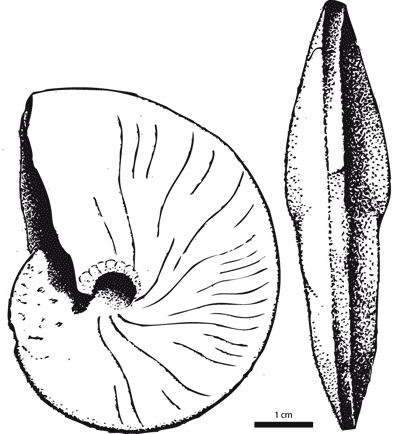
Figure 3: Neolobites vibrayeanus (d', 1841). Reproduction of the original figure of d' (1841, p. 322, pl. 96, figs. 1-3).
Diagnosis. Highly variable, oxycone and lanceolate engonoceratid, with small, shallow umbilicus. Suture is extremely simplified consisting of rounded, narrow lobes and wide rounded saddles. Cross section is variable from compressed to slightly inflated. Venter is variable from wide to narrow trapezoidal or simply rounded and in some species ornamented by fine crenulations. Sculpture is variable, too, ranging from smooth, unornamented forms to flexuous and ventrally ornamented forms.
Occurrence. Neolobites is restricted to the Middle (N. fourtaui) and Late (N. vibrayeanus) Cenomanian being a successful shallow marine taxon among Engonoceratidae. The genus is characterized by distinct morphotypes and populations in the Tethyan Realm and seamingly preferred lower latitudes and subtropical/tropical settings ( & , 2005, p. 942). It is reported from South America (Bolivia, Colombia, Peru, and Venezuela), Africa (Algeria, Egypt, Libya, Morocco, Niger, and Tunisia), Europe (France, Portugal, and Spain), and the Near and Middle East (Israel, Jordan, Lebanon, Oman, and Syria).
Included species:
Remarks. & (2005) stated that only N. vibrayeanus, N. fourtaui and N. peroni deserve discrete specific status. et alii (1981, p. 28) convincingly ruled out N. bedoti from Neolobites. It was placed in Metengonoceras ( et alii, 1992). Reported new species are based on fragmented and worn specimens. Therefore the sculpture and suture appear to be simplified (e.g. N. bassleri, N. kummeli). & (1981, p. 24) argued that N. peroni is also no more than an intraspecific variety of N. vibrayeanus. and (2005) did not discuss the earlier authors' opinion, but maintain N. peroni as a distinct species based on slight morphological differences between N. vibrayeanus and N. peroni. Taking into consideration the extreme morphological variety of the populations of Neolobites, the present author accepts the opinion of & (1981) on N. peroni, and considers this species an intraspecific variety of N. vibrayeanus. It is possible that N. fourtaui will also be found to fall into the intraspecific range of N. vibrayeanus, especially taking into consideration the eco-morphological analysis of & (2005). The other possibility, that future research will elucidate, is that N. fourtaui and N. vibrayeanus form two discrete chronospecies, namely N. fourtaui is older and N. vibrayeanus is younger. The Early Cenomanian N. bedoti of (1912, p. 31) has been assigned to Metengonoceras by & (1984, p. 105) because of the very similar morphologies of Neolobites and Metengonoceras. Based on these similarities, the most probable ancestor of Neolobites is Metengonoceras ( & , 2001).
Genus Knemiceras ,
1898, p. 200
= Cnemidoceras , 1900, p. 24; Cnemioceras ,
1900, p. 85; Knemoceras ,
1902, p. 7;
Omaimaiceras , 1955; Iranoknemiceras ,
1981, p.
258
Type species. Ammonites Syriacus , 1848, from the Neocomian of Lebanon

Click on thumbnail to enlarge the image.
Figure 4: Knemiceras syriacum (, 1848). Reproduction of the original figure of (1848, p. 21, pl. 6, figs. 1-3).
Diagnosis. Diagnosis is after et alii (1996, p. 130): "compressed to moderately inflated; sides flat and parallel or converging; venter flat or slightly concave; ribs moderately to very strong, sparse, rounded or flat, arising singly or in pairs from stout umbilical tubercles; ending in ventrolateral clavi or crossing venter. Suture with frilled lobes and slightly frilled, rarely entire saddles; commonly irregular."
Click on thumbnail to enlarge the image.
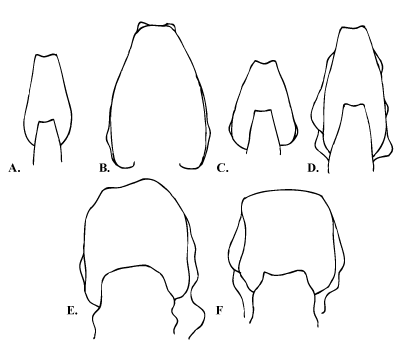
Figure 5: Highly variable cross sections of Knemiceras species. A.) K. compressum (, 1903, pl. 16, fig. 16); B.) K. compressum (, 1970, text-fig. 9c); C.) K. subcompressum ( et alii, 1997, fig. 2f); D.) K. kazhdumiense (, 1981, pl. 3, fig. 1c); E.) K. douvillei (, 1981, pl. 4, fig. 1b); F.) K. persicum (, 1981, pl. 5, fig. 1b).
Occurrence. Separation of South American forms of Knemiceras as Glottoceras by (2002) does not restrict the geographic distribution of the remaining Knemiceras species, which are reported from Europe (France, Portugal, Spain), North Africa (Algeria, Egypt, Tunisia), Near East (Israel, Lebanon, Syria), Middle East (Iraq, Iran, Oman), and also from South America (Colombia, Peru). Stratigraphically, Knemiceras appears in the earliest Albian (?latest Aptian) and disappears in the Early Cenomanian.
Included species:
Remarks. Resurrection of Glottoceras by (2002) did not solve the systematic uncertainties around Knemiceras. He considered the genus to be a geographical variant and leaves open the problem of the systematic position of the remaining Knemiceras species (e.g. K. attenuatum spinosum from Australia: , 1963). Other occurrences of 'Knemiceras' attenuatum from Tunisia (, 1956, pl. 2, fig. 3a-b) and Iran (, 1981, pl. 6, fig. 2) should be convincingly placed somewhere in the Engonoceratidae. & (1989, p. 364) proposed that Hypengonoceras ibericum be placed in Knemiceras on the basis that quadrituberculate ornament is unknown in Hypengonoceras. Bifid (e.g. K. compressum, K. subcompressum) or irregular (K. douvillei, K. iraniense) saddles are present in the suture line at any growth stage, distinguishing all Knemiceras species from Engonoceras and Platiknemiceras. The extreme intraspecific variation seen in Knemiceras ( & , 1989, p. 383) especially in its inflation and ornamentation ( & , 1991) is easily observable in the cross- sections of the body chambers of various species (Fig. 5). Cross-sections of Knemiceras range from the slender lanceolate forms (K. compressum) through wider (K. kazhdumiense, K. subcompressum) and subcircular (K. douvillei) and rectangular (K. persicum) forms. (1961) decreased this great variation within the genus by distinguishing the thin, unsculptured and less ornamented forms as Platiknemiceras. The extreme variability of shell morphology of Knemiceras may have been linked to the mode of life.
Genus Protengonoceras , 1903, p. 153
(Fig.
6 ![]() )
)
Type species. Engonoceras Gabbi , 1898, p. 197, from the Cretaceous of Texas (USA).
Click on thumbnail to enlarge the image.
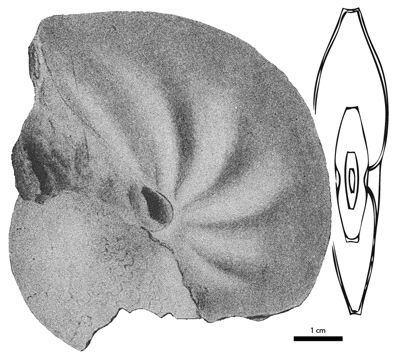
Figure 6: Protengonoceras gabbi (, 1898). Reproduction of the original specimen of (1898, p. 197).
Diagnosis. Diagnosis is after et alii (1996) and et alii (1998a). Conch is like Engonoceras but lacks tubercles at any stage and has feeble, smooth ribs only on adult stage. Cross section is compressed to very compressed and tabulate or may be rounded at adult stage. Where ventrolateral shoulder is present, it is always sharp. The suture line is almost straight and consists of many low adventive saddles, which are entire. Adventicious lobes are narrow and incised.
Occurrence. Protengonoceras is reported from the Early and Middle Albian of Peru, Texas, Mexico and Egypt. (1981) reported two species from the latest Albian and earliest Cenomanian of Iran.
Included species:
Remarks. (1903, p. 157) referred 's Sphenodiscus emarginatus to both Protengonoceras and Engonoceras. Without detailed description it can only be considered a nomen nudum so cannot be maintained. (1903, p. 157) referred to this ambiguity: "not having seen any specimens of the species, I can not say positively that it is a member of this genus", therefore this reference is omitted. (1981) reported two new species from the Late Albian-Early Cenomanian of Iran. However, all other species are reported from the Early and Middle Albian, and no occurrences are known from the Late Albian or younger. (1981) introduced P. prestati possessing well preserved dorsolateral and lateral bullae and a suture line with bifid saddles typical of Knemiceras. Protengonoceras lacks tubercles and never presents bifid and/or incised saddles. On that basis, P. prestati belongs rather to Knemiceras; however the single figured specimen (, 1981, pl. 6, fig. 3a-b) does not permit a reliable revision.
Genus Metengonoceras ,
1903, p. 179
= Epengonoceras , 1924, p. 508
(Fig. 7
![]() )
)
Type species. Metengonoceras inscriptum , 1903, from the Albian of Decatur, Texas.

Click on thumbnail to enlarge the image.
Figure 7: Metengonoceras inscriptum , 1903. Reproduction of the original figure of (1903: p. 180, pl. 25, figs. 5-9).
Diagnosis. Diagnosis is after (1987, p. C2). Metengonoceras includes very compressed, almost smooth ammonites having very narrow umbilicus and narrow, flattened venters, which may be rounded on the adult body chamber. Sculpture consists of falcoid growth lines. On some species, weak arcuate ribs appear a little above the midflank. Rarely faint umbilical bullae may appear. Suture is simplified, consisting of closely shaped numerous auxiliary and adventive elements. Most of the saddles are wide, being undivided and rounded. Larger lobes may be frilled moderately.
Occurrence. Metengonoceras occurred in the Gulf of Mexico during the Middle Albian ( & , 1989). Many species flourished in the Western Interior of the USA. Repeated speciation took place there (R.W. , 2007) from which certain Metengonoceras species invaded northward (Mowry Sea, Canada) and eastward (Normandy, France). During the Cenomanian Metengonoceras enlarged its territory outside North America. It is reported from Europe (France, Germany), Africa (Algeria, Egypt, Niger, Nigeria, Tunisia), and the Middle East (Jordan) as well as from South America (Brazil, Ecuador) reached through the Saharan seaway. Metengonoceras tended to become more acute and sharply keeled during the Cenomanian.
Included species:
Remarks: (1952) extensively reviewed the type species of Epengonoceras (E. dumbli) and concluded that the genus is not separable from Metengonoceras. This view was accepted by & (1984, p. 100) and is also followed here. R.W. (2007) reports stratigraphical ranges of engonoceratid ammonites from North America showing that certain Metengonoceras species occur in the Late Albian (M. teigenense, M. aspenanum) and Cenomanian (M. dumbli, M. acutum) suggesting that in North America Metengonoceras species range from earliest Late Albian to the latest Cenomanian.
Genus ? Hypengonoceras , 1922, p. 112
(Fig. 8 ![]() )
)
Type species. Placenticeras Warthi , 1895, from the Albian Utatur Group of southern India.
 |
 |
Click on thumbnail to enlarge the image.
Figure 8: Hypengonoceras decaryi (, 1963). Reproduction of the figures of & (1989: figs. 112-113).
Diagnosis. Diagnosis is after & (1989, p. 362). "Umbilicus is narrow. The shell compressed, the flanks generally little rounded. Sculpture consists of low falcoid ribs, often very weak and only visible under oblique lighting. Umbilical tubercles may be present, but are never very prominent. Venter is flat to concave with alternating ventral clavi at least at some stage. Dimorphism pronounced, but mainly restricted to differences in size. Suture variable; in typical forms some saddles are little divided and 'pincer-like' with bifid folioles; in others as incised as in a normal placenticeratid pattern."
Occurrence. Hypengonoceras is reported from the Late Albian of Europe (France), Africa (Madagascar, Morocco, Mozambique, Zululand), Near East (Israel), India, and probably Sakhalin (Japan). & (1984, Table 1) noted a doubtful occurrence in the Late Cenomanian of the Utatur Group, South India but they did not figure or describe it, so this occurrence is omitted.
Included species:
Remarks. Although & (1989) indicated the placenticeratid-like pattern of the suture of Hypengonoceras, they assigned the genus to the Engonoceratidae. This clearly relates to the debate on the systematic position of Hypengonoceras for it is the most disputed among the Engonoceratidae and is again under reconsideration ( & , 2004). This continuing dispute is based on the general differences between the Engonoceratidae and the Placenticeratidae. The Engonoceratidae are characterized by simplified ceratitic saddles (best seen on all species of Protengonoceras, Engonoceras, Metengonoceras, and Neolobites) in contrast to the frilled and finely incised saddles and lobes of Placenticeratidae. In this scheme, Parengonoceras and Hypengonoceras are transitional between the Engonoceratidae and the Placenticeratidae ( & , 1989, p. 364); however the first-named genus was placed in the Engonoceratidae without dispute - partly because of its known stratigraphical distribution (Early and Middle Albian) and because Parengonoceras possesses two true adventive lobes (, 1970). Only H. ibericum is questioned as a member of the genus by & (1989, p. 364) because quadrituberculate ornament is unknown in Hypengonoceras. They proposed that H. ibericum be placed in Knemiceras. Since the thorough discussion of Hypengonoceras by & (1989) no significant progress toward clarifying the position of the genus has been published; therefore the opinion of & (1989) is maintained here.
Genus Parengonoceras , 1924, p. 508
(Fig. 9 ![]() )
)
Type species. Amaltheus Ebrayi , 1882, from the Albian of Cosne, Ničvre, France.

Click on thumbnail to enlarge the image.
Figure 9: Parengonoceras ebrayi (, 1882). Reproduction of the original figure of (1882, p. 7, pl. 1).
Diagnosis. Coiling is typically engonoceratid. Conch is evolute. At early stages the cross section is lanceolate with tabulate venter. The lanceolate cross-section varies between wide triangular to acute and compressed. At adult stage the cross-section remains lanceolate or becomes oval. Venter is smooth or slightly grooved. Umbilical shoulder is low and always rounded and unornamented. Flanks are smooth in most cases or ornamented by ventrolateral, lateral or dorsolateral bullae. Falcoid ribs may appear ventrolaterally. Venter is usually narrow or in some species wide at adult stage and ornamented by weak tubercles. In some species these tubercles develop strongly. Suture line consists of two adventive and many auxiliary elements. Lobes are always irregularly incised. Saddles are never entire, always weakly or strongly incised.
Occurrence. Although Parengonoceras is widely considered to be restricted to the Early and Middle Albian ( & , 1989, p. 365; et alii, 1996, p. 130), many species (mainly South American) are reported from the Late Albian (P. barbacoense, P. caneroti, P. discoides, P. duplicatum, P. elegans). Furthermore, in Europe and Asia it is also known from the Late Albian (P. zagrosiense). Based on our recent knowledge, the stratigraphical distribution of Parengonoceras spans the whole of the Albian.
Included species:
Remarks. In his accurate analysis, (1970, p. 1030-1032) examined the sutural development of Parengonoceras during ontogeny, proving that it has two distinct adventive lobes. Parengonoceras has the most complex suture lines among the Early and Middle Albian Engonoceratidae with irregularly and finely frilled deep lobes and saddles. This suture line is distinctive among other Early and Middle Albian engonoceratid ammonites and is considered to be the forerunner of the suture seen in Hypengonoceras and therefore, a phylogenetic connection exists between these genera. All species of Parengonoceras have rather compressed and smoothly ornamented shells (although fine tubercles may appear on flanks) with frilled suture, which clearly distinguish them from all other genera of Albian and Cenomanian engonoceratids. The proposed Parengonoceras - Hypengonoceras lineage and the suture remarkably different from those of other engonoceratid species suggest a polyphyletic origin for the Engonoceratidae.
Genus Platiknemiceras ,
1954, p. 174
= Platyknemiceras , 1959, p. 77
(Fig. 10 ![]() )
)
Type species. Platiknemiceras bassei , 1954, from the Albian of Spain.
Click on thumbnail to enlarge the image.
Figure 10: Platiknemiceras bassei , 1954. Reproduction of the figure of (1954, p. 175, refigured by , 1961).
Diagnosis. Diagnosis is after (1954, p. 175). Discoidal shape, flanks convex in the middle, being widest next to umbilicus, which is narrow. Umbilical wall is vertical, the last whorl almost entirely embracing. Sculpture is reduced consisting of attenuated ribs that arise at the umbilicus. Ribs may be flexuous in course but not tuberculate. Venter is smooth and forms a shallow groove. Suture is ceratitic.
Occurrence. Platiknemiceras appears in the Early Albian. Most species occur in Early-Middle Albian and some range into the Late Albian (P. deserti, P. jullieni) and higher, into the earliest Cenomanian (P. valencianum). Geographically the genus is reported from Europe (Spain, France), Africa (Algeria, Egypt), South America (Colombia, Peru), North America (Texas, USA), the Middle East (Iran) and Japan.
Included species:
Remarks. (1961) discussed the genus thoroughly and grouped some species of Knemiceras (K. deserti, K. gracile, K. hachourii, K. subcomplicatum, and K. sp. nov. [later described as P. flexuosum et alii, 1998a]), and Engonoceras (E. jullieni) under Platiknemiceras because of common morphological features: thinly discoidal shape with extreme reduction of ornamentation; this grouping restricts the scope of the genus Knemiceras. Platiknemiceras is close to Parengonoceras (, 1961, p. 354) but it includes only compressed and discoidal forms having extremely reduced sculpture. P. caseyi sp. nov. was listed previously (, 1954; & , 1964) as Engonoceras aff. stolleyi (), an attribution corrected by et alii (1980, p. 334). Since (1961) revised the genus there have been no further contributions regarding the content and systematics of the taxon; however four new species have been described. Most recently (2010, p. 171) discussed the species assigned of Platiknemiceras. He considers the genus to be restricted to P. bassei, P. hachourii, and P. flexuosum. (2010) transfers back to Knemiceras some of the species that (1961) referred to Platiknemiceras (P. gracile, P. subcomplicatum, and P. deserti). Unfortunately, (2010) did not consider the other five species (P. caseyi, P. colombiana, P. julieni, P. sequanense, and P. valencianum) of the genus so his revision is not complete. For the moment this genus is considered to be polyphyletic for it is based on morphological features common to all.
[Engonoceratidae gen. et sp. nov. ,
1973
= Borissiakoceras sp. ( & ,
2009, p. 881)]
This tiny specimen is reported from the Albian of New Zealand (, 1973, p. 106). The specimen is not strongly evolute (two whorls are visible) atypical for Engonoceratidae. Nor is the suture is typical of Engonoceratidae: therefore the generic assignment is not supported. & (1979, p. 116) had already referred it to Borissiakoceras, but without further analysis. This was provided by & (2009) who also assigned it to Borissiakoceras. However poor preservation and a limited number of specimens do not permit a more precise distinction.
The mean generic range of Jurassic/Cretaceous ammonites is 7.3 myr per ammonite genus ( & , 1983). Engonoceratid genera have longer ranges. The longevity of most of the genera is remarkable: seven genera existed for more than 10 myr (Engonoceras, Glottoceras, Knemiceras, Metengonoceras, Parengonoceras, Platiknemiceras, and Protengonoceras). Only two genera (Hypengonoceras and Neolobites) had shorter ranges (4 myr) than the average mean generic range. Originally the Engonoceratidae were said to have appeared during the Barremian (, 1938, p. 486), but later works did not support this view. Now it is widely accepted that engonoceratids appeared in the latest Aptian-earliest Albian (, 1956; , 1981; , 2010) in shallow marine platform environments of the southern Tethys (Algeria) and migrated to the peri-Tethyan territories (, 1957; , 1965). It is noteworthy that G. (1940b, p. 1066) had already speculated that some engonoceratids possibly were present in the Aptian of Texas, an observation supported by et alii (2004) when revising the original collection of Ferdinand , that included an Engonoceras sp. from the ?Upper Aptian of Texas. Obviously, the adaptive radiation of the group was rapid during the Early Albian, for its genera played an important role in the faunas of the Peruvian Basin (South America) and the Western Interior Sea (USA). During Middle and Late Albian times the group occupied the Far East and Australian provinces (Australia, Borneo, India, Japan, Madagascar and South Africa).
In the Cenomanian it successfully invaded some portions of the Boreal Realm (Mowry Sea, Canada) for it became ubiquitous in temperate watery environments everywhere in palaeolatitudes up to the 60 degrees where it co-occurs with Boreal ammonites (, 2009). In the Mowry Sea the adaptive radiation of engonoceratid ammonites was so successful that the invader Metengonoceras displaced the endemic Neogastroplites from its original morphoplace (, 2004) as recorded by changes in its morphotypes (, 2000). The taxon became extinct during the Cenomanian. The last surviving species of the engonoceratids are reported from the latest Cenomanian of Texas ( et alii, 1981), Niger and Nigeria ( et alii, 1992, 1994; et alii, 1998) and the southern Mediterranean ( & , 2005; , 2007; et alii, 2008). Table 1 summarizes the stratigraphical ranges of engonoceratid genera. Although (1935) reported a Metengonoceras dumbli from the Early Turonian of Niger, this dating is not supported by the detailed biostratigraphical study of et alii (1992), which showed that M. dumbli ranges only to the latest Cenomanian in Niger. The general climate warming of the mid-Cretaceous combined with an increase in subtropical upper ocean temperature ( et alii, 2003), may have helped the radiation of engonoceratids along ancient continental shelf seas. Changes in sea level may be an additional favourable factor. The highest relative sea level documented occurred during the Mesozoic ( et alii, 2003a). These fluctuations may have been accompanied by changes in the oceanic circulation that aided in increasing productivity and opened new ecological niches, which together resulted in a decline of provincialism in epicontinental seas. Some of these factors may have contributed to a general decline of ammonoid diversity ( et alii, 2003a, p. 395). However, the well documented decrease in the richness of ammonoid species significantly pre-dates the spread of anoxic deposits ( et alii, 2003b) and surely contributed to the dramatic changes of shallow water habitats that were preferred by engonoceratids, that in the end may have led to their extinction. "Suddenly" raised sea levels would have reduced the number of barriers between those epicontinental and shallow water habitats and endemic centres in which engonoceratids lived.
Based on our recent knowledge, all engonoceratid genera preferred shallow marine environments, and some (Knemiceras and Neolobites) occupied extremely shallow (tidal, lagoonal) milieux. However, it took a long time to arrive at this conclusion, which began seventy years ago.
G. (1940a) was among the first to discuss the growing importance of palaeoecology, pointing out that only oxyconic ammonites (as Engonoceras and Oxytropidoceras) are numerous in the epineritic area of the Cretaceous of Texas (Western Interior Sea, USA) where echinoids, oysters, and thick-shelled gastropods abundant. G. (1940a) estimated the epineritic zone to occupy water depths of 40 to 180 metres, which corresponds well with the lower limit of the euphotic zone. G. (1940b, p. 1066) considered that the group preferred shallow water and platform facies: "Knemiceras, Engonoceras and Oxytropidoceras appear to have had their greatest development in the broad, shallow, clear seas of Texas". The habitat-dependency of ammonites was also analyzed by (1991) for the Cenomanian-Turonian ammonites of the Western Interior. It became obvious that most ammonites are not restricted to a particular depth or facies, although compressed and discoidal forms (e.g. the Engonoceratidae) tended to occur in environments where the depth was less than 50 to 100 metres (, 2003). (1984) also analyzed thoroughly the Western Interior basin, its endemism and faunal provinces concluding that during the Cretaceous the maximum depth of thebasin was never greater than 200-500 metres with a significant area covered by less than a 50 metres. This shallow, warm, trophic environment offered excellent conditions for speciation among many ammonites (including engonoceratids) thus creating an endemic centre (, 1972). Based on his extensive research on bivalves, (1973) defined a Western Interior Endemic Centre (WIEC). Later et alii (1993) enlarged the scope of the WIEC to include other molluscs emphasizing that the WIEC was a very important pool of genetic variability and speciation for many molluscs. This direct link between the water depth and the blossoming of the engonoceratids is remarkable. Its validity supported by (1990, 1996) who from a theoretical standpoint pointed out the special ecological needs of engonoceratids and that they preferred shallow (30-50 metres) water. Ecology of the South American Engonoceratidae was first discussed by (1970, p. 1022) concluding that the South American Engonoceratidae lived on a subsiding carbonate platform. (1995a) also recognized the importance of engonoceratids in shallow marine facies: "Knemiceratinae (Knemiceras, Platiknemiceras, Parengonoceras) represent important ammonoid "pioneer" genera for mid-Cretaceous transgressive facies: they occur frequently near the base of marine, often glauconitic sediments overlying a continental clastic succession in their Tethyan (s.l.) realm." Engonoceratids frequently occur in glauconitic sediments. Parengonoceras duboisi and Knemiceras saadense are present in the glauconitic sand/sandstone of Salazac (Gard, France: , 1940; , 1989) and in the latest Albian glauconitic limestone of Pénzesgyor (Bakony Mts. Hungary; , 1990a, 1990b), furthermore Parengonoceras attenuatum spinosum of (1963) from the Early Albian of Bathurst Island (Australia) is found in a glauconite sand. et alii (1981) reported an engonoceratid (Metengonoceras dumbli) from the Late Cenomanian of stable Europe (Angers, NW France), also in a glauconitic sand. (1997) noted a Knemiceras from a glauconite bed in the Albian of Jordan. & (1981) described Parengonoceras caneroti from a lignitic deltaic series of latest Albian age at Castellon (Spain). This occurrence also supports the shallow water dependency of engonoceratids. Although & (1991) considered engonoceratid ammonites to be indicative of a neritic environment they did not discuss their shallow water dependency.
It became obvious very early on, that the presence of some engonoceratids may be indicative of certain environments. (1903, p. 148) called attention to the ostreid and bryozoid encrustations of Knemiceras syriacum shells. More recently (2008) too discussed shell encrustations on Knemiceras found in fluvial, deltaic, littoral or sublittoral facies. The encrustation may have occurred during post-mortem drifting, rather than during life in natural habitats. However, & (1976) and (1996) found that the post-mortem dispersal of ammonoids is rather the exception than the rule. This finding is very probably applicable to Knemiceras. Accepted as a generalization, nekroplanktonic dispersal on a large scale must be abandoned; a Knemiceras from the Albian of Jordan with oyster encrustation (, 2008) makes very likely the probability of a shallow water habitat. (1955) pointed out that the Knemoceras [sic!] beds are linked to basal conglomerate in Borneo. et alii (1986) remarked that various species of Knemiceras occur in a littoral facies of the southern Atlas Mountains, Africa. et alii (1997) report Knemiceras from a delta-influenced carbonate ramp in the Albian of Sinai (Egypt), again supporting the conclusion that many occurrences of Knemiceras are coincident with the early stages of transgressive systems tracts. & (2008) report Knemiceras from intercalated non-marine-shallow marine beds of Middle Albian age in Israel. Knemiceras uhligi from the Albian of Oman occurs in a shallow marine (average depth 35 metres) epicontinental sea, which might not have been more than 80 metres deep ( et alii, 2000, p. 223). But ( & , 1994; et alii, 2009) report that Late Albian 'Knemiceras beds' in Tunisia involve a unique range of ecological needs. (1956) began the attempts to explain the ecology of the highly variable Knemiceras. She postulated that as Knemiceras lived in a stable environment in the neritic zone at the margin of the old Saharan mainland where there was no deposition of detritus or perturbation, so conditions there were optimum for the rapid expansion of its population, and the stress that would have hindered adaptation did not exist. Consequently, the genetic potential and variability of Knemiceras was wasted in useless polymorphism. (1956) said that the mode of life of Knemiceras species was more benthic than neritic. She outlined a completely opposing view of the mode of life and habitats of Knemiceras and other engonoceratids. & (1991) proposed that the great variability of shell form and ornamentation can be explained by their inhabintance of a shallow and labile (= epicontinental) environment. Recently (2008) reported Knemiceras from the euphotic, shallow marine deposits of Late Albian age in the Zagros Mountain (SW Iran). Again it is the only ammonite found in that shallow marine environment. The shallow water dependency of Knemiceras is supported by the frequent presence of Knemiceras on shallow marine platforms ( & , 1994) and in fluvial-lagoonal deposits with dinosaur remnants ( et alii, 2000). Generally, Knemiceras is the only ammonite in shallow water environments of the Arabian Plate (, 2010). Dinosaur experts also remark that during Cenomanian times the fragmented dinosaur remains found in continental and deltaic facies followed by lagoonal and coastal plain deposits topped by marine limestones containing the first truly marine animals such as Neolobites ( et alii, 2005, p. 168). & (2005) noted that where Neolobites occurs in great abundance, it is on shallow-marine shelves. Neolobites is always linked to transgression and/or flooding periods in South-America ( & , 1993, p. 600). Distribution of Cenomanian carbonate platforms and Neolobites occurrences are correlated to a fine degree in northern Africa and the Americas ( & , 1993). The possible shallow marine dependency of Engonoceratidae is strongly supprted by (2007) who considers Neolobites as the endemic and intra-shelf taxon of the group. That Neolobites is a Cenomanian intra-shelf (e.g. shallow marine) taxon is strong support for the same mode of life for its predecessor, the Albian engonoceratids; however the Neolobites in Bolivia (, 1968; & , 1992) does not support its endemism at the genus level.
With regard to the dispersal of Ammonoidea, (1993) analyzed the post-hatching early life of 71 selected Cretaceous species of Ammonoidea. He concluded (based on density calculations of the shells), that at the hatching stage most of them were floaters with a positive buoyancy, strongly suggestive of a planktonic mode of life, thus permitting the rapid migration of all Cretaceous Ammonoidea. But positive buoyancy cannot explain the shallow water dependency of all engonoceratids, a relationship strengthened by 's (1988) report of well preserved and abundant engonoceratid fauna (Parengonoceras, Protengonoceras, Engonoceras pierdenalis, E. stolleyi, and E. uddeni) from the Middle Albian open shelf of Lampazos, Sonora (Mexico). (1981) offered an interesting working hypothesis to explain the connection between the distribution of engonoceratids and the depth of seas in which they occur. He noted that thin forms [of engonoceratids] are found in the western regions and the thickest in eastern areas. This difference would suggest some regional control in the depth of the seas. But recent knowledge of the geographical and facies distribution of engonoceratids, negates this idea. Thin forms are found in the Far East (Platiknemiceras caseyi in the Early Albian of Japan ( et alii, 1980).
Some Upper Albian ammonoid habitats in southeastern France were reviewed by et alii (2005). But this work did not discussed the habitats of the engonoceratids, although engonoceratids occur in that region (cf. , 1940; , 1989). Based on the available data engonoceratids certainly lived in the neritic and epipelagic domain of et alii (2005) occupying extremely shallow marine habitats. But we have more evidence for the connection between transgression and the rapid distribution of engonoceratids. The "pioneerism" of engonoceratids sensu (1995a) is linked to their shallow water habitancy, for the colonizing populations of engonoceratids were able to occupy rapidly the new areas of shallow water formed by transgressive seas, and thus to inhabit pristine ecological niches. The invasion of engonoceratids into the Peruvian province ( & , 2004) is closely linked to a transgression. In Early Albian times, the stem-engonoceratids which originated at the southern margin of the Tethys invaded the Peruvian Basin using shallow water seaways. A similar event occurred during the Cenomanian. (2007) following up on earlier works noted that the Cenomanian shallow water seaway between the southern margin of the Tethys and South America (Saharan seaway), Metengonoceras migrated southward during latest Cenomanian times (, 2007, fig. 1). (1935) and et alii (1992) had already reported this southward advance on the Late Cenomanian deposits of Niger. The appearance of M. oliveirai in Sergipe, Brazil (, 1967, 1986; , 1994) substantiates this southward migration of the seaway.
(1938, p. 510) attempted to erect a phylogeny of Engonoceratidae, proposing that the first engonoceratid was the Aptian Protengonoceras with no direct link to other engonoceratids. The first Albian engonoceratid genus was Engonoceras and its offsprings were Metengonoceras, Parengonoceras, Hoplitoides, Knemiceras, Neolobites, and Placenticeras. (1952) suggested that in all probability, the Engonoceratidae were derived directly from the Parahoplitidae. (1957, p. 39) proposed that if the pseudoceratitic development of the Albian species is indeed the expression of a development from the deshayesitid lineage, then the superfamily name Placenticeratoidea of (1900, p. 584) would be applicable. Summarizing the earlier results, (1960, p. 208) concluded: "origin of the Engonoceratidae has never been settled." Since that statement, there has been no significant development concerning the origin of the engonoceratids. Almost forty years later, the situation is unaltered from the state that et alii (1998a, p. 1) remarked: "evolutionary origin of the group is uncertain". Recently & (2001) outlined a phylogenetic chart for Engonoceratidae proposing that the most possible ancestor was Knemiceras ( & , 2001, fig. 2) with a questioned relation between Parengonoceras/Hypengonoceras and the rest of the group. So uncertainty remains an open field for investigation.
Speculation on the origin of Engonoceratidae and its possible parahoplitid origin now is strengthened by new results on the ecology of Parahoplites. et alii (2009, p. 904) report Parahoplites from a lagoonal environment in north-central Tunisia. In addition to the shallow marine habitat of Parahoplites, et alii (2005) proposed that the relationship of the mode of life of the Acanthohoplitinae to the water surface (incl. Parahoplites) supports the probability of a relationship. The similarities of the environments of Parahoplites and Engonoceratidae furnish a plausible explanation for the origin of the Engonoceratidae.
Recently, & (2004) and (2008) placed the Engonoceratidae in the superfamily Pulchelliatoidea based on morphological analogies. However, the ecological data on Pulchelliidae sensu stricto do not fully support this lineage. Recently et alii (2004) proposed a nektobenthic mode of life for the Pulchelliidae. If the Engonoceratidae is in fact derived from the Pulchelliidae, then their differences in modes of life and palaeoenvironments require an explanation. Other data ( et alii, 2008) also refer to neritic platform environment with an abundant pulchelliid fauna. Very recently (2010) rejected the pulchelliatoid origin for the Engonoceratidae.
A search for the evolutionary driving force behind the genesis of the Engonoceratidae suggests that the global oceanic anoxic event offers a plausible explanation. Dramatic changes in the pelagic ecosystem may have been triggered by the long anoxic event (OAE 1b) and progressively increased ecological pressure on those ammonite populations inhabiting shallow marine and platform environments. They were forced to occupy pristine extremely shallow ecological niches, previously unused. The evolution of the supposed parahoplitid ancestors continued apace and the first Engonoceratidae (Knemiceras, Platiknemiceras, Parengonoceras, and Protengonoceras) appeared in the latest Aptian-earliest Albian times in the shallow water environments of one or all three large shelves in North Africa, the proto-Gulf of Mexico, and the Caribbean Province.
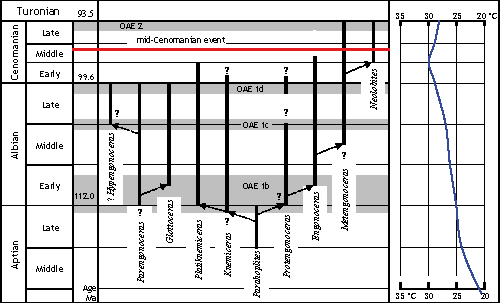
Click on thumbnail to enlarge the image.
Figure 11: Stratigraphical distribution of engonoceratid genera and supposed phylogeny of Engonoceratidae. Data from (1938), (1960, 1961), & (1989), et alii (1996), & (2004). Cretaceous time-scale after et alii (2004). OAE 1b-d dating from et alii (2006), OAE 2 dating from et alii (1999). Seawater temperature curve from et alii (2003).
The origin of Engonoceratidae is almost certainly polyphyletic as indicated by the remarkable differences between the sutures of Parengonoceras
- Hypengonoceras and those of the remainder of the group, along with the apparently contemporaneous occurrences of these species in shallow marine environments in both Americas and North Africa. However, the enigma of the systematic position of Hypengonoceras, the most problematic genus of the Engonoceratidae may also be the result of a polyphyletic origin. &
(1983, p. 868) suggested that Hypengonoceras may be an off-shoot of an early Late Albian placenticeratid, but as seems more probable,
(1960, p. 208) proposed that Hypengonoceras was derived from Parengonoceras or some allied members of the Engonoceratidae. This uncertainty of origin is shown in
Fig. 11 ![]() by the discrete tracing of possible lineages. The most probable course of evolution was from ?Parahoplites to Protengonoceras. Evolution from Protengonoceras toward Engonoceras (or from Engonoceras toward Protengonoceras proposed by & ,
2001) and Metengonoceras and ultimately Neolobites is reasonable based on shell morphology and the simplification of suture over time. Lately,
(2010, p. 169) supports this concept assembles these genera into a phylogenetic lineage. The other lineage is more speculative; however &
(2004) suggested that the South American Engonoceratidae had a platiknemiceratid origin. But this is not a solution applicable to the other species, for a direct lineage from Parengonoceras to Hypengonoceras is feasible. Many have speculated on the existence of a direct phylogenetic connection between the Engonoceratidae and the Placenticeratidae (,
1930, p. 390; et alii,
1957, p. L109), but
(1960, p. 208) and &
(1989, p. 365) negated the possibility that the Engonoceratidae was the direct ancestor of the Placenticeratidae. Therefore the proposed phylogenetic connection with Hypengonoceras as an off-shoot of an early placenticeratid ( & ,
1983) must be abandoned (Fig. 11
by the discrete tracing of possible lineages. The most probable course of evolution was from ?Parahoplites to Protengonoceras. Evolution from Protengonoceras toward Engonoceras (or from Engonoceras toward Protengonoceras proposed by & ,
2001) and Metengonoceras and ultimately Neolobites is reasonable based on shell morphology and the simplification of suture over time. Lately,
(2010, p. 169) supports this concept assembles these genera into a phylogenetic lineage. The other lineage is more speculative; however &
(2004) suggested that the South American Engonoceratidae had a platiknemiceratid origin. But this is not a solution applicable to the other species, for a direct lineage from Parengonoceras to Hypengonoceras is feasible. Many have speculated on the existence of a direct phylogenetic connection between the Engonoceratidae and the Placenticeratidae (,
1930, p. 390; et alii,
1957, p. L109), but
(1960, p. 208) and &
(1989, p. 365) negated the possibility that the Engonoceratidae was the direct ancestor of the Placenticeratidae. Therefore the proposed phylogenetic connection with Hypengonoceras as an off-shoot of an early placenticeratid ( & ,
1983) must be abandoned (Fig. 11 ![]() ). Today's opinion: the Engonoceratidae became extinct without descendants in latest Cenomanian times in the Western Interior Province (USA). However the dating of the end of the Cenomanian is debated by
R.W. et alii
(2009), as is the closing of the Saharan seaway (Niger and Nigeria).
). Today's opinion: the Engonoceratidae became extinct without descendants in latest Cenomanian times in the Western Interior Province (USA). However the dating of the end of the Cenomanian is debated by
R.W. et alii
(2009), as is the closing of the Saharan seaway (Niger and Nigeria).
The apparent similarities between the engonoceratid and ceratitid suture lines tempted palaeontologists to speculate on the cause of the repetition of similar structures (cf. , 1914, p. 30 on the Neolobites suture line). Recently et alii (2005) analyzed the ecology and immigration patterns of approximately 500 specimens of ceratitid Ammonoidea found in strata of the Late Anisian to Early Ladinian age in Germany. Their study was based on the morphological variability of ammonoid shells. The Germanic Basin was the site of a shallow sea during most of the Anisian and Ladinian ( et alii, 2005, p. 19). It was occupied repeatedly by incursions of ammonoids, crinoids and brachiopods coincident with intervals of high sea level. After each of these immigrations new species of Ceratites evolved ( et alii, 2005, p. 25) just as we had seen among Cretaceous engonoceratids in the Peruvian Basin and in the Western Interior Sea (USA). Cretaceous species of the engonoceratids exhibit remarkable morphological convergence very likely that of the Triassic species used in the ecological analysis of et alii (2005). The repetition of suture lines, external shell morphologies, together with records of a shallow marine habitat, successive migrations, and successful adaptation in both Middle Triassic and mid-Cretaceous forms suggest that this pattern is a very successful ammonoid eco-morphotype in shallow marine habitats. This conclusion is supported by the morphological convergences of the Triassic Ceratites nodosus nodosus ( et alii, 2005, fig. 4) and the Cretaceous Knemiceras syriacum (, 1848, figs. 2-3).
Other morphological convergences occur (Fig. 12 ![]() ) among the Triassic Discoceratites semipartitus ( et alii,
2005, fig. 4), the Albian-Cenomanian Metengonoceras teigenense ( & ,
1989, pl. 2, figs. 1-2) and the Maastrichtian Sphenodiscus pleurisepta ( et alii,
2005, fig. 7a-b). The Triassic scheme of colonization followed by speciation follows the same pattern as that of engonoceratids during the Albian. The endemic centres in the Americas were rapidly colonized by engonoceratids, followed by speciation ( & ,
2004). This suggests that the repetition of the Triassic eco-morphotype in the engonoceratids was caused by their occupation of nearly identical ecological niches and habitats.
) among the Triassic Discoceratites semipartitus ( et alii,
2005, fig. 4), the Albian-Cenomanian Metengonoceras teigenense ( & ,
1989, pl. 2, figs. 1-2) and the Maastrichtian Sphenodiscus pleurisepta ( et alii,
2005, fig. 7a-b). The Triassic scheme of colonization followed by speciation follows the same pattern as that of engonoceratids during the Albian. The endemic centres in the Americas were rapidly colonized by engonoceratids, followed by speciation ( & ,
2004). This suggests that the repetition of the Triassic eco-morphotype in the engonoceratids was caused by their occupation of nearly identical ecological niches and habitats.
Another interesting convergence is that of the engonoceratids and another group of Ammonoidea, the Maastrichtian sphenodiscids.
et alii (2005) reported several species of Sphenodiscus (S. lobatus and S. pleurisepta) from the Maastrichtian of La Popa Basin, Mexico, a shallow water coastal deposit. The oxyconic Sphenodiscus was well adapted to continuous swimming with excellent acceleration skills ( & ,
1996, p. 211). Shell morphology, suture lines and palaeoenvironments of these sphenodiscid ammonites surprisingly repeat that of the Albian-Cenomanian engonoceratids. et alii
(2005, p. 62) concluded: "sphenodiscid-dominated faunas […] are thus clearly related to bathymetry, i.e. the shallow-water assemblages are very low in diversity and represent specialization to an environment in which ammonoids could not easily survive". It seems obvious that during ammonoid evolution, the combination of shallow water habitats, the oxy- and/or platyconic, brevidome shell morphology and simplified, Ceratites-like sutures occurred at least three times, one of them the Albian-Cenomanian engonoceratids. In all cases, this pattern was associated with successful migration, colonization and speciation. This set of characters is summarized in
Fig. 12 ![]() .
.

Click on thumbnail to enlarge the image.
Figure 12: Strong similarities in shell shapes among certain ammonoid taxa. Repeated eco-morphotype pattern among Mesozoic Ammonoidea. The proposed eco-morphotype comprises a set of features consists of nektonic, active swimmer mode of life in shallow marine settings, simplified and ceratitic suture and involute, oxycone-platycone shell form. Mesozoic time-scale after et alii, 2004.
Engonoceratid ammonites are important time indicators of chronostratigraphic time in strata of shallow marine origin ranging in age from the ?latest Aptian times to the youngest of Cenomanian formations. Where they occur in the Tethyan Faunal Realm, they are excellent indicators of a shallow water environment. Today engonoceratids play an important role in correlation, biostratigraphy and ecological studies, but trhey are limited in distribution to temperate water. Engonoceratids are a versatile, and successful group of Cretaceous Ammonoidea, ecological pioneers that were successful in occupying previously inaccessible pristine ecological niches. The range of suture lines, shell form, all is response to a shallow water habitat created a morphological pattern that proved successful not only for the engonoceratids, but also for other Mesozoic Ammonoidea.
Engoncoeras duboisi , 1989, is known only from 2 localities and represented by a handful specimen. New specimen from Hungary led the present author for thorough morphological analysis, which resulted the below revision of the species.
Parengonoceras duboisi (, 1989)
(Figs. 13A-B ![]() - 14
- 14 ![]() - 15
- 15 ![]() )
)
v 1989 Engonoceras duboisi sp. nov. : p. 56, pl. 2, figs. 1-3.
1989 Engonoceras sp. cf. E. duboisi sp. nov. : p. 57, pl. 2, fig. 4.
v 1990a Engonoceras duboisi ; : p. 11, pl. 1, figs. 1-4.
v 2007 Engonoceras duboisi ; et alii: p. 101, pl. 15, figs. 1, 4.
A |
 |
B |
 |
Click on thumbnail to enlarge the image.
Figure 13: Parengonoceras duboisi (, 1989) from the latest Albian (Stoliczkaia dispar Zone) of Tilos forest (Bakony Mts. Pénzesgyor, Veszprém County, Hungary). Specimen NHNM M2010.760.1. A.) lateral view, B.) dorsal view.
Click on thumbnail to enlarge the image.
Figure 14: Suture line of Parengonoceras duboisi (, 1989) at whorl height of 51 mm. Specimen HNHM M2010.760.1.
Material. Three internal moulds from the Late Albian (Stoliczkaia dispar Zone) of Tilos forest (Pénzesgyor, Bakony Mts, Hungary) and three worn internal moulds from the Late Albian (Stoliczkaia dispar Zone) of Salazac, Gard (France).
Occurrence. (1989) reported it from the latest Albian of Salazac, Gard (France). (1990a) described it from Hungary (Tilos Forest, Bakony Mts.). et alii (2007) copied the figures of (1990a). New collection by the present author from the Tilos Forest (Bakony Mts., Hungary) yielded a single specimen described here for the first time supporting the distribution of the species for the latest Albian of France and Hungary. The new record of the Late Albian P. duboisi from the Bakony Mts. is important as a connector between the rich and diverse western and less well known eastern Tethyan Parengonoceras species during the Albian. It suggests a southern Tethyan migration route because the fact that the Alcapa Tectonic Unit (which also includes the Bakony Mts.) was part of the Apulia microplate bordered by the southern edge of the Penninic Ocean ( & , 2004). The revealed third specimen of P. duboisi from Hungary refers to the rarity of this species in the Bakony Mts, Hungary: 1:10,000 (Z. Evanics pers. comm.).
Description. Medium sized incomplete but shelly phragmocones and internal moulds. Coiling is typical of the Engonoceratidae. The conch is discoidal and strongly involute and oxycone
(Fig. 13 ![]() ). Last whorl embraces the previous ones. The umbilicus is small and shallow. Umbilical wall is low and the umbilical shoulder is gently rounded. Flanks converge and are almost unsculptured. The cross section is lanceolate. Venter is smooth and narrow. Finer or stronger prorsiradiate falcoid ribs arise at midflank, which end at the ventro-lateral shoulders. Six fine and falcoid ribs are on the last quarter of whorl; ribs end at fine tubercles on ventral shoulder. Body chamber is not preserved. Suture line
(Fig. 14
). Last whorl embraces the previous ones. The umbilicus is small and shallow. Umbilical wall is low and the umbilical shoulder is gently rounded. Flanks converge and are almost unsculptured. The cross section is lanceolate. Venter is smooth and narrow. Finer or stronger prorsiradiate falcoid ribs arise at midflank, which end at the ventro-lateral shoulders. Six fine and falcoid ribs are on the last quarter of whorl; ribs end at fine tubercles on ventral shoulder. Body chamber is not preserved. Suture line
(Fig. 14 ![]() ) is typical for Parengonoceras showing two adventive lobes. Suture is characterized by pincer-like and finely incised lobes and saddles. Pincer-like saddles are irregular polyphillic. Lobes are also incised irregularly.
) is typical for Parengonoceras showing two adventive lobes. Suture is characterized by pincer-like and finely incised lobes and saddles. Pincer-like saddles are irregular polyphillic. Lobes are also incised irregularly.
Click on thumbnail to enlarge the image.
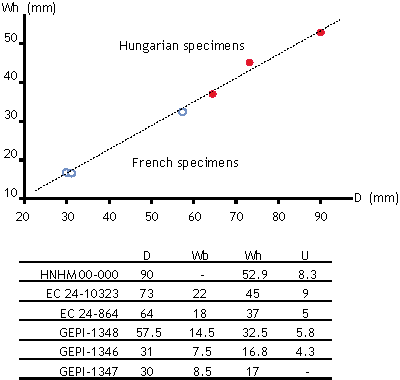
Figure 15: Size distribution of Hungarian (solid) and French (open) specimens of Parengonoceras duboisi (, 1989).
Remarks. P. duboisi is most similar to P. elegans and Platiknemiceras hachourii (). The Early Albian Platiknemiceras hachourii has a slightly more slender cross-section, and the striae on the lower flanks are finer in P. duboisi and the suture is less frilled than on P. hachourii. The extreme reduction of ornamentation in P. duboisi is also not present in P. hachourii. The ornamentation is typical of Parengonoceras as is the diameter and whorl height ratios
(Fig. 15 ![]() ). P. duboisi is most similar to P. elegans in its shell ornamentation. The small, ventrolateral falcoid ribs of
P. elegans (,
1970, pl. 3, fig. 1a-c) are comparable with P. duboisi. However P. duboisi shows fewer ribs per whorl than P. elegans. The superficially similar Engonoceras and Parengonoceras species are clearly distinguished from each other based on suture lines
- if present. Normally the saddles of Engonoceras are all entire except that the outermost, which is bifid ( et alii,
1996, p. 130). The present species show only frilled saddles in all stages in the specimens. All saddles of the holotype are incised, mostly bifid (,
1989, pl. 2, figs. 2-3) being worn specimens, therefore the frilled suture is not well preserved, however there are no entire saddles at any stage. The Hungarian specimens are well preserved, partly shelly specimens with well observable suture revealing more incised saddles. Biometry of the French and Hungarian specimens refers the same species
(Fig. 15
). P. duboisi is most similar to P. elegans in its shell ornamentation. The small, ventrolateral falcoid ribs of
P. elegans (,
1970, pl. 3, fig. 1a-c) are comparable with P. duboisi. However P. duboisi shows fewer ribs per whorl than P. elegans. The superficially similar Engonoceras and Parengonoceras species are clearly distinguished from each other based on suture lines
- if present. Normally the saddles of Engonoceras are all entire except that the outermost, which is bifid ( et alii,
1996, p. 130). The present species show only frilled saddles in all stages in the specimens. All saddles of the holotype are incised, mostly bifid (,
1989, pl. 2, figs. 2-3) being worn specimens, therefore the frilled suture is not well preserved, however there are no entire saddles at any stage. The Hungarian specimens are well preserved, partly shelly specimens with well observable suture revealing more incised saddles. Biometry of the French and Hungarian specimens refers the same species
(Fig. 15 ![]() ), too. Based on the well developed and frilled saddles and lacking entire saddles at any stage, this species is convincingly placed into Parengonoceras. Recently the uncertainty of the systematic position of P. duboisi is also referred by
(2010, p. 169) excluding it from the genus Engonoceras. This conclusion is supported here.
), too. Based on the well developed and frilled saddles and lacking entire saddles at any stage, this species is convincingly placed into Parengonoceras. Recently the uncertainty of the systematic position of P. duboisi is also referred by
(2010, p. 169) excluding it from the genus Engonoceras. This conclusion is supported here.
I acknowledge the valuable comments on the initial draft by Herbert , Jean-Louis and Christian . My special thanks to Robert W. for providing vital papers and essential criticism of draft as well as to Peter F. to clarify 'Pseudengonoceras'. I acknowledge the detailed remarks of Jens , which greatly improved the text. Special thanks to Margaret , Jeffrey and Mikel for sending important papers. I am deeply indebted for the great help and encouragement of Hugh G. on Albian ammonites found in the 1990's and especially for his careful comments on this draft, which greatly improved the quality of the paper. I am also indebted to the late William who in the early 1990's introduced me to the engonoceratids. I acknowledge the technical support of Attila (Hungarian Natural History Museum, Budapest) for housing the specimen treated here and granting permission for use the facilities of the Museum. I am grateful to Mr. Zoltán (private collector) who provided his Hungarian engonoceratid specimens for this research. Finally, I acknowledge the comments of the referees of this paper, especially the detailed remarks of Christian . Last but not least I acknowledge the continuous help of the Editor, Bruno , and the careful linguistic improvement provided by Nestor .
H. & L. (1994).- Sur l'âge des couches à Knemiceras de Tunisie méridionale. Caractérisation de l'Albien supérieur (Zone à Inflata et Zone à Substuderi).- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Plančtes), t. 319, p. 337-340.
H., S. & C. (2008).- Palaeogeography at the Cenomanian-Turonian boundary in southern Central Tunisia: The basins of Gafsa area.- Abstracts, Thirty third International Geological Congress, 6-14 August, 2008, Oslo, Norway.
R. (2008).- Lithostratigraphy and biostratigraphy of some Lower Cretaceous outcrops from Northern Sinai, Egypt.- Cretaceous Research, London, vol. 29, p. 603-624.
H. & M. (1991).- Découverte d'ammonites albiennes dans le massif apennin de la Maiella (plate-forme Lazio-abruzzaise Italie centrale) : précision sur la durée du hiatus bauxitique médio-Crétacé.- Cretaceous Research, London, vol. 12, p. 81-90.
M.F. & G.I. (2001).- Upper Cenomanian-Lower Turonian ammonites from north and central Sinai, Egypt.- El Minia Bulletin, vol. 13-14, p. 17-60.
M.F., A. & A. (2008).- Late Cenomanian-Early Turonian ammonites of Jordan.- Revue de Paléobiologie, Genčve, vol. 27, p. 43-71.
B., M. & J. (1981).- Étude stratigraphique et paléontologique du Crétacé supérieur et Paléocčne du Tinrhert-W et Tademait-E (Sahara algérien).- Documents des Laboratoires de Géologie de Lyon, Hors Série, vol. 6, p. 15-173.
F. & P. (1984).- Présence du genre Knemiceras (Ammonoidea, Engonoceratidae) dans l'Albien moyen de l'Aube (France).- Bulletin d'Information des Géologues du Bassin de Paris, vol. 21, p. 21-25.
F., G. & A. (1996).- Révision des ammonites du Cénomanien supérieur et du Turonien inférieur du Tinrhert (Sahara algérien) : implications biostratigraphiques.- Bulletin du Muséum national d'Histoire naturelle, Paris, (4čme Série), Section C, vol. 18, p. 179-232.
F., W.A., G. & P. (2002).- Metengonoceras teigenense et , 1989 : une ammonite exotique d'origine nord-américaine dans le Cénomanien inférieur de Basse Normandie (France).- Bulletin trimestriel de la Société géologique de Normandie et des Amis de Muséum du Havre, t. 87, p. 5-28.
F. (2008).- Support for a Vraconnian Stage between the Albian sensu stricto and the Cenomanian (Cretaceous System).- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2008/02 (CG2008_M02), 83 p.
B.S. (1997).- Sedimentology and paleogeography of the regressive-transgressive Kurnub Group (Early Cretaceous) of Jordan.- Sedimentary Geology, Amsterdam, vol. 112, p. 69-88.
C. & J. (1977).- Ammoniten und der Utrillas-Schichten (Mittelkreide) in der östlichen Provinz Albacete, SE Spanien.- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, vol. 1977, p. 1-14.
W.J., B. & C.W. (1957).- Mesozoic Ammonoidea. In: R.C. (Ed.), Mollusca 4.- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, p. L80-L437.
S. (1956).- Contribution à l'étude des Engonoceratidae (les couches à Knemiceras du Sud Tunisien).- Annales des Mines et de la Géologie, Tunis, vol. 20, p. 1-47.
K. & R.K. (1984).- Cenomanian-Turonian transition in the Cretaceous of Southern India.- Bulletin of the Geological Society of Denmark, Copenhagen, vol. 33, p. 21-30.
R., C. & A.B. (2004).- Palaeoecological significance of Barremian ammonite assemblages and facies variations from Southwest Mexico.- Lethaia, Oslo, vol. 37, p. 223-234.
R., R., C. & J. (2008).- Lower Barremian (Lower Cretaceous) ammonites from the Veloz Formation of Central Cuba: biostratigraphic and paleobiogeographic implications. In: J.G. (ed.), Abstracts and Program Eighteenth Caribbean Geological Conference.- Sociedad Dominicana de Geología, Santo Domingo, p. 14-15.
É. (1937-1940).- Les Céphalopodes crétacés des massifs côtiers syriens. Part 1-2.- Notes et Mémoires sur le Moyen-Orient, Paris, vol. 2, p. 165-200; vol. 3, p. 411-473.
É. (1954).- Description d'une nouvelle espčce d'ammonite tunisienne appurtenant au genre Neolobites , 1882 : Neolobites medeninensis nov. sp. et considérations sur çe genre.- Bulletin de la Société des Sciences Naturelles de Tunisie, Tunis, vol. 6, p. 197-204.
J.R. (1954).- Los Engonoceratidos en Espańa. In: Homenaje póstumo al Dr. D. Francisco .- Facultad de Ciencias de la Universidad de Barcelona, p. 173-178.
J.R. (1959).- Primer suplemento a la "Sinopsis de las especies nuevas del Cretáceo de Espańa".- Boletin del Instituto Geológico y Mineralógico de Espańa, Madrid, vol. 70, p. 1-77.
R.J. (1991).- Sutural amplitude of ammonite shells as a paleoenvironmental indicator.- Lethaia, Oslo, vol. 24, p. 219-225.
V.A. (1956).- Cretaceous System in northern Peru.- Bulletin of the American Museum of Natural History, New York, vol. 108, p. 357-493.
M., S., E., D., M., M. & C. (2000).- Dinosaurs and other fossil vertebrates from fluvial deposits in the Lower Cretaceous of southern Tunisia.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 157, p. 227-246.
M., A., Y. & M. (1985).- L'Albien moyen supérieur à Knemiceras frome la base de la grande transgression crétacée au Tebaga de Medenine (Tunisie meridionale).- Comptes Rendus de l'Académie des Sciences, Paris, (Série II, Sciences de la Terre et des Plančtes), t. 300, p. 965-968.
P.-Y. & J. (1978).- Les bassin occidental portugais de l'Albien au Campanien. In: R.A. & G. (eds.), Événements de la partie moyenne du Crétacé.- Annales du Muséum d'Histoire Naturelle de Nice, vol. 4 (1976), p. I.1.-I.14.
P.-Y. & J. (1979).- Essai de synthčse paléogéographique et paléobiostratigraphique du bassin occidental portugais au cours du Crétacé supérieur.- Cięncias da Terra, Lisboa, vol. 5, p. 121-144.
P.-Y., A. & J. (1979).- Mid Cretaceous events Iberian Field Conference 77, Guide I, Partie Portugal.- Cuadernos de Geología Ibérica, Madrid, vol. 5, p. 31-124.
B. (1926).- Dos Neolobites.- Boletín de la Sociedad Geológica del Perú, Lima, vol. 2, p. 39-41.
J. (1898).- Ueber Ammonites Pedernalis v. .- Zeitschrift der Deutschen Geologischen Gesellschaft, Hannover, Band 50, p. 183-201.
E. (1910).- Monografía
geológica y paleontológica del Cerro de Muleros cerca de Ciudad Juárez, Estado de Chihuahua
y descripción de la fauna Cretácea de La Encantada, Placer de Guadalupe, Estado
de Chihuahua.- Boletín del Instituto Geológico de México, vol. 25, 193
p.
online at http://ia700303.us.archive.org/6/items/boletndelinsti251910inst/boletndelinsti251910inst.pdf
E. (1928).- Cretaceous ammonites from Texas and northern Mexico.- University of Texas, Bulletin, Austin, vol. 2748, p. 143-312.
J.C., M., M., A., P. & P. (1982).- El Albense superior en el sector de Huescar (Provincia de Granada, Cordilleras Beticas).- Cuadernos de Geología Ibérica, Madrid, vol. 8, p. 703-720.
T.J., E., B., W.V., C.L. & J. (1999).- The record of global change in mid-Cretaceous (Barremian-Albian) sections from the Sierra Madre, northeastern Mexico.- Journal of Foraminiferal Research, Lawrence, vol. 29, p. 418-437.
L. (1968).- Hallazgo del ammonite Neolobites en la Caliza Miraflores y de huellas de dinosaurios en la Formación El Molino y su significado para la determinación de la edad del "Grupo Puca".- Boletín del Instituto Boliviano del Petróleo, La Paz, vol. 8, p. 16-28.
M. (1940).- Sur la découverte de Knemiceras aff. saadense et sp. dans le Vraconien de Salazac (Gard).- Compte Rendu sommaire des Séances de la Société géologique de France, Paris, fasc. 8-9, p. 87-88.
M. (1952).- Sur la découverte de Knemiceratinae (ammonites albiennes) en Équateur, en Colombie et au Vénézuela.- Comptes rendus hebdomadaires des séances de l'Académie des Sciences, Paris, t. 234, p. 2633-2635.
I.M. (1967).- Sôbre a ocorręncia de um cefalópodo no Cretáceo da área de Estância, Estado de Sergipe, Brasil.- Instituto de Geocięncias, Boletim de Geologia, Universidade Federal do Rio de Janeiro, vol. 1, p. 71-77.
I.M. (1986).- O Cretáceo inferior do nordeste do Brasil.- Anuário do Instituto de Geocięncias, Universidade Federal do Rio de Janeiro, vol. 1986, p. 103-125.
L. (1848).- Über Ceratiten.- Physikalische Mathematische Abhandlungen der Koeniglichen Akademie der Wissenschaften zu Berlin, p. 1-31.
L. (1990a).- First record of Engonoceratidae , 1900 (Ammonoidea, Cretaceous) from Hungary.- Revue de Paléobiologie, Genčve, vol. 9, p. 9-14.
L. (1990b).- Albai és cenomán ammonoideák palaeobiogeográfiai értékelése az alp-kárpáti és a mediterrán Tethys régióban.- általános Földtani Szemle, Budapest, vol. 25, p. 305-325.
L.G., B. & L. (2005).- Modification de la structure des populations d'ammonites dans l'Aptien inférieur sommital et l'Aptien supérieur du domaine Vocontien (SE France). In: D. (ed.), Impact environnemental sur la dynamique des populations au Crétacé.- Volume des résumés, École des Mines de Paris, p. 15-17.
L.G. (2007).- Endémisme et cosmopolitisme des faunes d'ammonites de la plaque arabique au Crétacé moyen (Aptien-Turonien). In: L.G., S. & D. (eds.), Relations entre les marges septentrionale et méridionale de la Téthys au Crétacé.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/02 (CG2007_M02), p. 49-55.
L.G., E., E. & W.J. (2007).- The Albian ammonite successions of the subandean zone of Ecuador. In: E. & I. (eds.), Fourth European Meeting on the Palaeontology and Stratigraphy of Latin America.- Cuadernos del Museo Geominero, Instituto Geológico y Minero de Espańa, Madrid, vol. 8, p. 57-61.
L.G. (2010).- Systematic palaeontology of Aptian and Albian ammonites from southwest Iran. In: F.S.P. van, M.I., F. & H.J. (eds.), Aptian Stratigraphy and Petroleum Habitat of the Eastern Arabian Plate.- GeoArabia Special Publications 4, vol. 1, p. 167-195.
A. & G. (1936).- Reconocimiento geológico de los yacimientos petrolíferos del departamento de Puno.- Boletín del Cuerpo de Ingenieros de Minas del Perú, Lima, vol. 115, 100 p.
J. & M. (1981).- La faune albienne de Traiguera (province de Castellon-Espagne).- Documents des Laboratoires de Géologie de Lyon, Hors Série, vol. 6, p. 227-249.
J., P., B., I., J. & J-P. (1986).- Comparative study of the lower and mid-Cretaceous sequences on different Maghrebian shelves and basins - their place in the evolution of the North African Atlantic and Neotethysian margins.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 55, p. 213-232.
R. (1957).- The Cretaceous ammonite genus Leymeriella, with a systematic account of its British occurrences.- Palaeontology, London, vol. 1, p. 28-59.
R. (1960).- Hengestites, a new genus of Gault ammonites.- Palaeontology, London, vol. 2, part 2, p. 200-209.
R. (1961).- The Cretaceous (Albian) ammonite genus Platiknemiceras Bataller.- The Annals and Magazine of Natural History, London, (Thirteenth Series), vol. 4, p. 353-357.
R. (1978).- A monograph of the Ammonoidea of the Lower Greensand. Part VIII.- Monograph of the Palaeontographical Society, London, Issue 554, part of vol. 132, p. 583-632.
G.W. (1941).- Ammonites from the Bagh Beds.- Proceedings of the Indian Academy of Sciences, Bangalore, (Section B), vol. 14, p. 271-276.
P. (1886).- Recueil d'études paléontologiques sur la faune crétacique du Portugal. Volume I. Espčces nouvelles ou peu connues.- Commisăo do Serviço Geologico, Lisbon, vol. 1, p. 1-40.
W.A. (1987).- The Upper Cretaceous (Cenomanian) ammonites Metengonoceras dumbli () and M. acutum .- U.S. Geological Survey, Bulletin, Washington, vol. 1690, p. C1-C7.
W.A. & W.J. (1989).- The Ammonite Metengonoceras , 1903 from the Mowry Shale (Cretaceous) of Montana and Wyoming.- U.S. Geological Survey, Bulletin, Washington, vol. 1787, p. L1-L11.
W.A. & W.J. (1990).- Upper Cenomanian ammonites from the Woodbridge Clay member of the Raritan Formation in New Jersey.- Journal of Paleontology, Tulsa, vol. 64, p. 845-846.
W.A. & G.R. (1972).- Stratigraphy and ammonite fauna of the Graneros Shale and Greenhorn Limestone near Pueblo, Colorado.- U.S. Geological Survey, Professional Paper, Washington, vol. 645, p. 1-108.
M. (1963).- Atlas des fossiles caractéristiques de Madagascar (Ammonites). Fascicule X (Albien).- Service géologique, Tananarive, 184 p.
M. (1965).- Nouvelles ammonites néocrétacées sahariennes.- Annales de Paléontologie (Invertébrés), Paris, vol. 51, p. 165-220.
M. (1966).- Les Céphalopodes crétacés du bassin côtier de Tarfaya. Relations stratigraphiques et paléontologiques.- Notes et Mémoires Service géologique du Maroc, Rabat, vol. 175, p. 9-148.
M. & A. (1968).- Ammonites crétacées des Rides prérifaines (région du Dahar-en-Nsour).- Notes du Service géologique du Maroc, Rabat, vol. 28, p. 19-27.
M. (1981).- Faune albo-cénomanienne de la formation des marnes de Kazhdumi, région du Fars - Khuzestan (Iran).- Documents des Laboratoires de Géologie de Lyon, Hors Série, vol. 6, p. 251-291.
M.H. (1862).- Géologie et
paléontologie de la région sud de la province de Constantine.- Mémoires de la Société
d'Émulation de la Provence, Marseille, t. II, 366 p.
online at http://www.archive.org/download/geologieetpaleon00coquiala/geologieetpaleon00coquiala.pdf
G.H. (1853).- Études sur les Mollusques fossiles du département de l'Yonne.- Bulletin de la Société des Sciences historiques et naturelles de l'Yonne, Auxerre, fasc. 7, 319 p.
P., J. & J. (1998).- Ammonites faunal exchanges between South Tethysian platforms and South Atlantic, during the uppermost Cenomanian - lowermost Turonian, in the Benue Trough (Nigeria).- Géobios, Villeurbanne, vol. 31, p. 187-214.
P. (2007).- Échanges et colonisations fauniques (Ammonitina) entre Tethys et Atlantique sud au Crétacé superieur : voies atlantiques ou sahariennes? In: L.G., S. & D. (eds.), Relations entre les marges septentrionale et méridionale de la Téthys au Crétacé.- Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/02 (CG2007_M02), p. 16-19.
F.W. (1893).- A contribution to the invertebrate paleontology of the Texas Cretaceous.- Texas Geological Survey Fourth Annual Report (for 1892), Austin, p. 139-294.
F.W. (1894).- New and little known vertebrates from the Neocomian of Kansas.- The American Geologist, Minneapolis, vol. 14, p. 1-12.
F.W. (1900).- Buchiceras (Sphenodiscus) belviderensis and its varieties.- The Colorado College Studies, Colorado Springs, vol. VIII, p. 27-31.
L. & A. (2004).- Mesozoic plate tectonic reconstruction of the Carpathian region.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 210, p. 1-56.
P. (1965).- Découverte de Knemiceratinae : Parengonoceras et Platiknemiceras (Ammonites) dans l'Albien inférieur du Bassin parisien.- Comptes Rendus sommaire de la Société géologique de France, p. 332-333.
P. (1979).- Les ammonites de l'Albien inférieur et moyen dans le stratotype de l'Albien : gisements, paléontologie, biozonation. In: P., F., J.-J., R., P., D., P., H., D. & G. (eds.), L'Albien de l'Aube.- Les Stratotypes Français, Comité Français de Stratigraphie, Ed. CNRS, Paris, vol. 5, p. 51-193.
H. (1890).- Sur la classification des Cératites de la Craie.- Bulletin de la Société géologique de France, Paris, (3čme Série), t. XVIII, p. 275-292.
H. (1912).- Évolution et classification des Pulchelliidés.- Bulletin de la Société géologique de France, Paris, (4čme Série), t. XI, p. 285-320.
H. (1916).- Les Terrains Secondaires dans le Massif du Moghara à l'Est de l'Isthme de Suez.- Mémoires de l'Académie des Sciences, Paris, (2čme Série), vol. 54, 184 p.
H. (1928).- Les Ammonites de la Craie supérieure en Égypte et au Sinaď.- Académie des Sciences de l'Institut de France, Mémoires, Paris, vol. 60, p. 1-10.
H. (1931).- Contribution à la Géologie de l'Angola. Les Ammonites de Salinas.- Boletim do Museu e Laboratorio Mineralogico e Geologico da Universidade de Lisboa, vol. 1, p. 17-46.
G. (1953).- Ammonites nouvelles des Monts du Mellégue.- Bulletin du Service de la Carte géologique d'Algérie, Alger, (1čre Série), Paléontologie, vol. 16, 76 p.
O. (1910).- Bearbeitung der Cephalopoden der Schweinfurtschen Sammlung und über die Entwicklung des Turons in Ägypten, (Obere Kreide Ägyptens).- Zeitschrift der Deutschen Geologischen Gesellschaft, Hannover, Band 62, p. 379-387.
O. (1914).- Die Cephalopoden der Schweinfurthschen Sammlung aus der Oberen Kreide Ägyptens.- Zeitschrift der Deutschen Geologischen Gesellschaft, Hannover, Band 66, p. 179-216.
F. (1979).- Zonation of the Cretaceous of central Colombia by ammonites.- Publicaciones Geológicas Especiales del Ingeominas, Bogotá, vol. 2, 186 p.
F. (1981).- On the supposed Aptian occurrence of the ammonite genus Neodeshayesites in Colombia and Venezuela; with an appendix on Neodeshayesites karsteni (Marcou).- Geología Norandina, Bogotá, vol. 3, p. 45-51.
P. (1880-1887).- Manuel de conchyliologie et de paléontologie conchyliologique.-
Librairie F. Savy, Paris, 11 fascicules, 1369 p.
online at http://www.archive.org/download/manueldeconchyli00fisc/manueldeconchyli00fisc.pdf
R. (1975).- Die geologische Entwicklung von Süd-Mozambique seit der Unterkreide und die Ammoniten-Fauna von Unterkreide und Cenoman.- Geologisches Jahrbuch, Hannover, (Reihe B), Heft 12, 324 p.
R. (1935).- Le Crétacé et le Tertiaire du Sahara soudanais (Soudan, Niger, Tchad).- Archives du Muséum national d'Histoire naturelle, Paris, (6čme Série), vol. 13, p. 3-97.
W.M. (1877).- Description of a collection of fossils, made by Doctor Antonio in Peru.- Journal of the Academy of Natural Sciences of Philadelphia, (series 2), vol. VIII, p. 263-336.
T.K. & S. (2007).- Ornamental polymorphism in Placenticeras kaffrarium (Ammonoidea; Upper Cretaceous of India): evolutionary implications. In: N.H., R.H. & R.H. (eds.), Cephalopods present and past: new insights and fresh perspectives.- Springer Verlag, Dordrecht, p. 97-120.
A., R., M., B., J.F., J., A., M., B., M.I., A. & R. (2004).- Segunda fase de post-rifting: Cretácico Superior. In: J.A. (ed.), Geología de Espańa.- Sociedad Geológica de Espańa y Instituto Geológica y Mineralógica de Espańa, Madrid, p. 509-522.
O.F. (1995a).- Knemiceras uhligi () dans le Chaîne Ibérique méridionale (Albien supérieur, Espagne).- Batallería, Barcelona, vol. 5, p. 5-17.
O.F. (1995b).- Zur Alb-Cenoman-Transgression im südlichen Keltiberikum (Spanien).- IGPS Abstracts Profil, Stuttgart, vol. 8, p. 473-482.
O.F., J. & M. (1997).- On some Albian ammonites from northeastern Sinai (Egypt).- Paläontologische Zeitschrift, Stuttgart, vol. 71, p. 221-229.
P.A. (1942).- Sur la stratigraphie des calcaires crétaciques de la foręt de Touvois.- Compte Rendu sommaire des Séances de la Société géologique de France, vol. 9, p. 78-79.
R., J. & L. (1993).- Late Albian-Middle Cenomanian carbonate platform of Betic and Iberian basins (Spain). In: J.A.T., R.W. & J.-P. (eds.), Cretaceous carbonate platforms.- AAPG Memoirs, vol. 56, p. 271-281.
C. (1988).- Estratigrafía y geología estructural de las rocas sedimentarias cretácicas del área del Lampazos, Sonora.- Universidad Nacional Autónoma de México, Instituto de Geología, Revista, vol. 7, p. 148-162.
F., J.G. & A.G. (2004).- A geologic time scale 2004.- Cambridge University Press, 589 p.
B. (1915).- Fauna cretacea dell'Egitto raccolta dal Figari Bey.- Palaeontographia Italica, Pisa, vol. 21, p. 189-232.
D., F., S., F., M., F. & W. (2008).- Le Cénomanien supérieur-Turonien de l'Atlas saharien (Algérie).- Bulletin de la Société Géologique de France, vol. 179, n° 6, p. 593-603.
A. (1912).- Le Crétacé de la Loire-Inférieure et de la Vendée.- Bulletin de la Société Scientifique naturelle de l'Ouest de la France, Nantes, (3čme Série), t. II, n° 1-2, p. 1-38.
J. (2002).- A proposal on the classification of systems tracts: application to the allostratigraphy and sequence stratigraphy of the Cretaceous Colombian Basin. Part 2: Barremian to Maastrichtian.- Geología Colombiana, Bogotá, vol. 27, p. 27-49.
G. de (1825).- Monographiae Ammoniteorum et Goniatiteorum Specimen.- H.W. Hazenberg, Lugduni Batavorum, 168 p.
N.S. (1955).- Geological accounts of West-Borneo.- Geological Survey Department, British Territories in Borneo, Bulletin, Kuching, Sarawak, vol. 2, 285 p.
E. (1900).- Céphalopodes.- Revue critique de Paléozoologie,
Paris, 4čme année, n°1, p. 19-24 & 78-87.
online at http://ia700104.us.archive.org/11/items/revuecritiquedep41900coss/revuecritiquedep41900coss.pdf
R.A. (1973).- Clarence and Raukumara Series (Albian - ?Santonian) Ammonoidea from New Zealand.- Journal of the Royal Society of New Zealand, Wellington, vol. 3, p. 71-123.
M. (1999).- La sédimentation de haut niveau marin du Crétacé supérieur de l'Atlas saharien oriental et des Aurés. Stratigraphie séquentielle. Analyse quantitative des biocénoses, évolution paléogéographique et contexte géodynamique.- Thčse Doctorat čs Sciences, Université des Sciences et de la Technologie Houari Boumediene (USTHB), Alger, 802 p.
A. (2006).- Northern Iraq study.- Cambridge Carbonates Ltd, UK, 386 p.
A. (1875).- The Jurassic and Cretaceous ammonites collected in South America by Prof. James , with an appendix upon the Cretaceous ammonites of Prof. 's collection.- Proceedings of the Boston Society of Natural History, vol. 17, p. 365-378.
A. (1889).- Genesis of the Arietidae.- Smithsonian Contributions to Knowledge, Washington, vol. 673, p. 1-238.
A. (1900).- Cephalopoda. In: K.A. (ed.), Text-book of Paleontology.- Macmillan, London - New York, p. 502-604.
A. (1903).- Pseudoceratites of the Cretaceous.- U.S. Geological Survey, Monographs, Washington, vol. 44, 1-351.
Y. (2009).- An Early Albian Arctic-type ammonite Arcthoplites from Hokkaido, northern Japan, and its paleobiogeographic and paleoclimatological implications.- Journal of Asian Earth Sciences, Oxford, vol. 34, p. 46-50.
I.C.Z.N. (1999).- International Code of Zoological Nomenclature, Fourth Edition.- International Trust for Zoological Nomenclature, London, xxix + 306 p.
online version at
http://www.nhm.ac.uk/hosted-sites/iczn/code/
C., W. & A. (2005).- Maastrichtian shallow-water ammonites of northeastern Mexico.- Revista Mexicana de Ciencias Geológicas, Mexico, vol. 22, p. 48-64.
A., W., S.J., R.W., T., J., S. van der & L.J.A. (2000).- Origin and correlation of disconformity surfaces and marker beds, Nahr Umr Formation, northern Oman.- Society for Sedimentary Geology, Special Publication, Tulsa, vol. 69, p. 209-225.
D.K. & J.A. (1996).- Buoyancy and hydrodynamics in Ammonoids. In: N.H., K. & R.A. (eds.), Ammonoid paleobiology.- Topics in Geobiology, Plenum Press, New York, vol. 13, p. 170-224.
É. & T. (1992).- Las secuencias sedimentarias de la formacion miraflores y su significado cronoestratigrafico.- Revista Tecnica de YPFB, Santa Cruz, vol. 12, p. 257-264.
É. & A. (1993).- The Cenomanian - Turonian transition on the Peruvian margin.- Cretaceous Research, London, vol. 14, p. 585-605.
É. (1995).- La sedimentación albiana - turoniana en el sur del Perú (Arequipa - Puno - Putina).- Sociedad Geológica del Perú, Lima, Volumen jubilar Alberto , p. 135-157.
E.G. (1973).- Cretaceous Bivalvia. In: A. (ed.), Atlas of paleobiogeography.- Elsevier, Amsterdam, p. 353-384.
E.G. (1984).- Paleobiogeography and evolutionary response dynamic in the Cretaceous Western Interior seaway of North America. In: G.E.G. (ed.), Jurassic-Cretaceous biochronology and paleogeography of North America.- Geological Association of Canada, Special Paper, St. John's, n° 27, p. 273-306.
E.G., B.B., J.I., W.P., P.J. & T. (1993).- Molluscan biostratigraphy of the Cretaceous Western Interior Basin, North America. In: W.G.E. & E.G. (eds.), Evolution of the Western Interior Basin.- Geological Association of Canada, Special Paper, St. John's, n° 39, p. 397-434.
W.J. & W.A. (1976).- Aspects of ammonite biology, biogeography, and biostratigraphy.- Special Papers in Palaeontology, London, vol. 17, 94 p.
W.J. & H.C. (1979).- Cretaceous faunas from Zululand and Natal, South Africa. The ammonite superfamily Haplocerataceae , 1884.- Annals of the South African Museum, Cape Town, vol. 77, p. 85-121.
W.J. & P. (1981).- Upper Cenomanian ammonites from the environs of Saumur, and provenance of the type of Ammonites vibrayeanus and Ammonites geslinianus.- Cretaceous Research, London, vol. 2, p. 19-49.
W.J., P. & J.M. (1981).- Upper Cenomanian ammonites from Anjou and the Vendée, Western France.- Palaeontology, London, vol. 24, p. 25-84.
W.J. & C.W. (1983).- Ammonites polyopsis , 1837, and the Cretaceous ammonite family Placenticeratidae , 1900.- Palaeontology, London, vol. 26, p. 855-873.
W.J. & P. (1984).- A revision of the ammonite faunas of the Cenomanian. 2 The families Binneyitidae, Desmoceratidae, Engonoceratidae, Placenticeratidae, Hoplitidae, Schloenbachiidae, Lyelliceratidae and Forbesiceratidae.- Cretaceous Research, London, vol. 5, p. 93-161.
W.J. (1988).- Late Cenomanian and Turonian ammonite faunas from northeast and central Texas.- Special Papers in Palaeontology, London, vol. 39, 131 p.
W.J. & W.A. (1990).- Cenomanian ammonite faunas from the Woodbine Formation and lower part of the Eagle Ford Group, Texas.- Palaeontology, London, vol. 33, p. 75-154.
W.J. & M.D. (1991).- Mid-Cretaceous ammonites and associated microfossils from the Central Oman Mountains.- Newsletters on Stratigraphy, Stuttgart, vol. 25, p. 127-154.
W.J., N.H. & W.A. (1998a).- Engonoceratid ammonites from the Glen Rose limestone, Walnut clay, Goodland limestone, and Comanche Peak limestone (Albian) in Texas.- American Museum Novitates, New York, n° 3221, 40 p.
W.J., W.A., A.S., J.M. & N.H. (1998b).- Ammonites from the Weno Limestone (Albian) in Northeast Texas.- American Museum Novitates, New York, n° 3236, 46 p.
W.J., J.M., W.A. & N.H. (2004).- A revision of the ammonite types described in F. 's Die Kreidebildungen von Texas und ihre organishen Einschlüsse.- Acta Geologia Polonica, Warszawa, vol. 54, p. 433-445.
W.J. (2004).- Ammonites from the Pawpaw Shale (Upper Albian) in northeast Texas.- Cretaceous Research, London, vol. 25, p. 865-905.
W.J., W.A., J.M. & A.S. (2005).- Upper Albian and Lower Cenomanian ammonites from the Main Street Limestone, Grayson Marl and Del Rio Clay in northeast Texas.- Cretaceous Research, London, vol. 26, p. 349-428.
H.C. & W.J. (1989).- Cretaceous faunas from Zululand and Natal, South Africa: The ammonite family Placenticeratidae , 1900; with comments on the systematic position of the genus Hypengonoceras , 1924.- Annals of the South African Museum, Cape Town, vol. 98, p. 241-408.
C., W., D. & A.G. (2005).- Morphological fluctuations of ammonoid assemblages from the Muschelkalk (Middle Triassic) of the Germanic Basin - indicators of their ecology, extinctions, and immigrations.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 221, p. 7-34.
M.M., E.F. & M.J. (1947).- Mesozoic fossils of the Peruvian Andes.- Studies in Geology, Johns Hopkins University, Baltimore, vol. 15, 150 p.
C.A. & K. (1986).- Micropaleontology and biostratigraphy of the coastal basins of West Africa.- Journal of African Earth Sciences, Oxford, (Special Series), vol. 4, p. 1-102.
D., V., J. & C. (2003).- The A-mode sutural ontogeny in prolecanitid ammonoids.- Palaeontology, London, vol. 46, p. 1123-1132.
F. (1895).- Untersuchungen über die Südindische Kreideformation.- Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients, Wien, Band 9, p. 97-203.
P.G.L.W. (1902).- Die Fauna der Kreide von Temojoh in West-Borneo.- Sammlungen des Geologisches Reichs-Museums in Leiden, (series I), vol. 7, p. 1-28.
V. & S. (2008).- Arthropod trace diversity on fossil leaves from the mid-Cretaceous of Negev, Israel.- Alavesia, Vitoria-Gasteiz, vol. 2, p. 239-245.
J. & J. (1970).- Significance of sutures in phylogeny of Ammonoidea.- Paleontological Contributions of the University of Kansas, Lawrence, vol. 47, p. 1-32.
J.-L. (1989).- Les genres Engonoceras et , 1881 et Hypengonoceras , 1922 dans l'Albien supérieur (z. à dispar) de Salazac, Gard, France.- Revue de Paléobiologie, Genčve, vol. 8, p. 51-63.
J.-L. (2008).- A revision of Amaltheus Ebrayi de , 1882, type species of the genus Parengonoceras , 1924.- Revue de Paléobiologie, Genčve, vol. 27, p. 249-267.
J. & M.A. (2001).- A new placenticeratid ammonite from the Albian (Lower Cretaceous) of California and the inferred history of some placenticeratid and engonoceratid ammonites.- Neues Jahrbuch für Geologie und Paläontologie, Monatshefte, Stuttgart, vol. 2001, n° 6, p. 343-358.
J. & H-G. (2009).- Late Cretaceous ammonites from the Bou Angueur Syncline (Middle Atlas, Morocco) - stratigraphic and paleobiogeographic implications.- Palaeontographica A, Stuttgart, vol. 289, p. 45-87.
J., M., M. & M.E.H. (2009).- Aptian (Lower Cretaceous) biostratigraphy and cephalopods from north central Tunisia.- Cretaceous Research, London, vol. 30, p. 895-910.
Z. (1981).- A Late Albian Hypengonoceras (Ammonoidea) from the 'Bentonite-Bed' at Makhtesh Ramon, Southern Israel.- Israel Journal of Earth Sciences, Tel Aviv, vol. 30, p. 35-38.
C.I. (1908).- Contribución al conocimiento sobre algunos ammonites del Perú.- Tipografía el Perú, Lima, 22 p.
C.I. (1925).- Algunos fósiles del Perú.- Boletin de la Sociedad Geológica del Perú, Lima, vol. 1, p. 23-30.
M., H.G., A., P.A. & J. (2009).- Early Late Albian (Cretaceous) of the central region of the Basque-Cantabrian Basin, northern Spain: biostratigraphy based on ammonites and orbitolinids.- Cretaceous Research, London, vol. 30, p. 385-400.
P. de (1882).- Études sur les faunes des couches du Gault de Cosne (Ničvre).- Mémoires de la Société Paleontologique de Suisse, Genčve, vol. 9, p. 1-118.
N.P. & N.P. (1958).- Superfamily Engonocerataceae. In: N.P. & V.V. (eds.), Fundamentals of palaeontology: Molluscs - Cephalopods II.- Gosudarstvenniye Nauchno-Tekhnicheskoe Izdatel'stvo, p. 125-128 [in Russian].
G.I. (1955).- Études paléontologiques sur la faune crétacique du massif du Moghara (Sinaď, Égypte).- Publications de l'Institut du Désert d'Égypte, Héliopolis, Le Caire, vol. 8, 192 p.
E.A. (1979).- Late Albian and Early Cenomanian Grayson ammonite biostratigraphy in North-Central Texas.- Journal of Paleontology, Tulsa, vol. 53, p. 1013-1022.
E.A. (1982).- Early Cenomanian cephalopods from the Grayson Formation of north-central Texas.- Cretaceous Research, London, vol. 3, p. 241-259.
R., A. & R. (1994).- Distribución de los ammonites del Cretácico inferior de la Cordillera Costera Catalana e Ibérica Oriental.- Cuadernos de Geología Ibérica, Madrid, vol. 18, p. 337-354.
L. & M. (2008).- Relative sea-level changes of the Lower Cretaceous deposits in the Chotts area of southern Tunisia.- Turkish Journal of Earth Sciences, Ankara, vol. 17, p. 835-845.
J.R. & J. (1980).- Ammoniten und Alter der Mittelkreide-Transgression in der westlichen Provinz Valencia, Spanien.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 159, p. 256-272.
T. (1954).- The Cretaceous System in the Japanese Islands.- Japan Society for the Promotion of Science, Tokyo, 324 p.
T. & T. (1964).- Hinagu.- Geological Map of Japan, scale 1:50,000, Tokyo, Explanatory Text, 147 p.
T., T. & Y. (1980).- Cephalopod faunule from the Cretaceous Yatsushiro Formation (Kyushu) and its implications.- Transactions and Proceedings of the Palaeontological Society of Japan, Tokyo, (new series), vol. 118, p. 325-338.
C. & H. (1996).- Les ammonites du Cénomanien supérieur et du Turonien inférieur de la région de Gafsa-Chotts, Tunisie du Centre-Sud.- Géobios, Villeurbanne, vol. 29, p. 3-49.
C. & H. (2005).- Précision sur les successions d'ammonites du Cénomanien-Turonien dans la région de Gafsa, Tunisie du Centre-Sud.- Revue de Paléobiologie, Genčve, vol. 24, p. 111-199.
C. & M. (2002).- Quelques ammonites du Cénomanien-Turonien de la région d'Errachidia-Boudnid-Erfoud (partie méridionale du Haut Atlas Central, Maroc).- Revue de Paléobiologie, Genčve, vol. 21, p. 759-779.
C., K., J. & B. (1992).- Les ammonites du Niger (Afrique occidentale) et la transgression transsaharienne au cours du Cénomanien-Turonien.- Géobios, Villeurbanne, vol. 25, p. 55-100.
C., K., J., B. & A. (1994).- Nouvelles données sur les ammonites du Niger oriental (Ténéré, Afrique occidentale) dans le cadre de la transgression du Cénomanien-Turonien.- Géobios, Villeurbanne, vol. 27, p. 189-219.
C., H., G. & J. (2003a).- Cenomanain (early Late Cretaceous) ammonoid faunas of Western Europe. Part II: diversity patterns and the end-Cenomanain anoxic event.- Eclogae Geologicae Helvetiae, Basel, vol. 96, p. 381-398.
C., H., J. & G. (2003b).- Cenomanian ammonoid diversity patterns and the end-Cenomanian anoxic event. In: Mesozoic paleoceanography.- Séance spécialisée de la Société géologique de France, (Paris, 10-11 juillet 2003), Abstract Volume, p. 22.
P., I.H. & W.J. (1983).- Cenomanian ammonites from northern Aquitaine.- Cretaceous Research, London, vol. 4, p. 317-339.
W.L. (1942).- Paleontology of the Eagle Ford Group of North and Central Texas.- Journal of Paleontology, Tulsa, vol. 16, p. 192-220.
J.A., L., J.-L., R., O., T. & R. (2008a).- Precisiones sobre la edad de la base de la Fm. Escucha, mediante ammonoideos, en la subcuenca de la Salzedella, Cuenca del Maestrat (E Cordillera Ibérica).- Geo-Temas, Madrid, vol. 10, p. 1269-1272.
J.A., R. & H. (2008b).- Consideraciones bioestratigráficas sobre los ammonites albienses de la sección del Jebel Naimia, Cadena norte de los Chotts (Túnez). In: J.I., L. & J.C. (eds.), XXIV Jornadas de la Sociedad Espańola de Paleontología.- Museo del Jurásico de Asturias, Colunga, Spain, p. 160-161.
L. & G.I. (1955).- Nouvelles observations stratigraphiques et paléontologiques sur l'Albien du massif du Moghara (Sinai-Égypte).- Travaux du Laboratoire de Géologie de Grenoble, vol. 31, p. 269-274.
M. & H. (1999).- Mid-Cretaceous ammonites from the coastal chain of Syria. In: K. (ed.), Cephalopods - Present and Past.- Fifth International Symposium, (Vienna, 6-9 September 1999), Abstract Volume, Geologische Bundesanstalt, Vienna, p. 88.
M., A.K. & H. (2003).- Le Crétacé moyen de la chaîne des Palmyrides (Syrie centrale).- Geodiversitas, Paris, vol. 25, p. 429-443.
M. & V. (1881).- Ueber Ammonitiden aus den Hilsbildungen Norddeutschlands.- Palaeontographica, Stuttgart, Band 27, p. 129-203.
F.E., F. & D.F. (2005).- Theropod pedal unguals from the Late Cretaceous (Cenomanian) of Morocco, Africa.- Revista del Museo Argentino de Ciencias Naturales, Buenos Aires, (new series), vol. 7, p. 167-175.
A.C.V.D. d' (1840-1842).- Paléontologie française, terrains crétacés, I - Céphalopodes.- Masson édit., Paris, 662 p. + 148 Pls.
H.G. (1999).- Correlation of Albian European and Tethyan ammonite zonations and the boundaries of the Albian Stage and substages: some comments.- Scripta Geologica, Amsterdam, Special Issue, vol. 3, p. 129-149.
H.G. & J. (2006).- Late Albian ammonites from offshore Suriname: implications for biostratigraphy and palaeobiogeography.- Cretaceous Research, London, vol. 27, p. 717-727.
H.G. (2007).- Ammonite biostratigraphy of the Albian in the Kirchrode II borehole, Hannover, Germany.- Cretaceous Research, London, vol. 28, p. 921-938.
E.L. (1956).- An Engonoceras from Central Oregon.- Journal of Paleontology, Tulsa, vol. 30, p. 398-402.
K.N. (1996).- Mesozoic Ammonoids in space and time. In: N.H., K. & R.A. (eds.), Ammonoid paleobiology.- Topics in Geobiology, Plenum Press, New York, vol. 13, p. 755-794.
A. & P. (1890).- Description des invertébrés fossiles des terrains crétacés de la région sud des Hauts-Plateaux de la Tunisie recueillis en 1885 et 1886 par P. .- Exploration scientifique de la Tunisie, Imprimerie nationale, Paris, 103 p.
L. (1907).- Études de paléontologie tunisienne. I. Céphalopodes des terrains secondaires; systčme crétacique.- Carte géologique de la Tunisie, Rudeval, Paris, 438 p.
L. (1910).- Sur quelques Ammonites du Crétacé algérien.- Mémoires de la Société Géologique de France, Paris, n° 42, (Paléontologie), vol. XVII, fasc. 2-3, 86 p.
A.H. (1905).- Fossil Cephalopoda described by and , in the Museum of the University of Texas.- The Biological Bulletin, Woods Hole, vol. 8, p. 365-366.
E., C., S.M.F., G., S. & P. (2003).- Thermal evolution of Cretaceous marine waters inferred from oxygen isotope composition of fish tooth enamels.- Paleoceanography, Washington, vol. 18, p. 7.1-7.12.
P.F. (1981).- Early Cretaceous ammonite biostratigraphy and biogeography. In: M.R. & J.R. (eds.), The Ammonoidea: the evolution, classification, mode of life and geological usefulness of a major fossil group.- The Systematics Association, Academic Press, London, Special Volume, vol. 18, p. 499-529.
S., F. & O. (2005).- Ammonoid abundance variations related to changes in trophic conditions across the Oceanic Anoxic Event 1d (Latest Albian, SE France).- Palaios, Lawrence, vol. 20, p. 121-141.
J.B. & A.A. (1931).- Mollusks from the Aspen Shale (Cretaceous) of southwestern Wyoming.- Proceedings of the United States National Museum, Washington, vol. 78, article 17, p. 1-24.
O. (1970).- Über die Gattungen Parengonoceras , Knemiceras und Neophlycticeras (Ammonoidea) aus den Anden Venezuelas.- Eclogae Geologicae Helvetiae, Basel, vol. 63, p. 1021-1057.
O. (1982).- The Cretaceous ammonites of Venezuela.- Maraven S.A., Caracas, 132 p.
R. & W.J. (1991).- Phenotypic plasticity in a Cretaceous ammonite analyzed by multivariante statistical methods: A methodological study. In: M.K., B. & J. (eds.), Evolutionary biology.- Plenum Press, New York, p. 411-426.
R. (2008).- A review of the post-mortem dispersal of cephalopod shells.- Palaeontologia Electronica, London, vol. 11, n° 3, 12A, 13 p.
E., L.G., A., É., R., M. & M. (1998).- La transgresión de cretáceo inferior en el margen andino (Perú y Ecuador): datos preliminares.- Boletín de la Sociedad Geológica del Perú, Lima, vol. 88, p. 73-86.
E. (2002).- La transgression albienne dans le Bassin Andin (Pérou) : Biostratigraphie, paléontologie (ammonites) et stratigraphie séquentielle.- Strata, Toulouse, vol. 38, p. 1-380.
E., L.G., É. & B. (2003).- Revision bioestratigrafica del Albiano inferior a superior basal en La Cuenca Andina Peruana (Peru central y norte).- Boletín Sociedad Geológica del Perú, Lima, vol. 95, p. 59-74.
E. & L.G. (2004).- Origin, phylogeny, faunal composition, and stratigraphical significance of the Albian Engonoceratidae (Pulchelliaceae, Ammonitina) of Peru.- Journal of South American Earth Sciences, Oxford, vol. 17, p. 11-23.
F. (1938).- Les ammonites jurassiques et crétacées. Essai de genera.- Masson et Cie, Paris, 554 p.
S.S. (1966).- On Placenticeras mintoi , 1908, from the Bagh Beds (Cretaceous) of India.- Paläontologische Zeitschrift, Stuttgart, vol. 40, p. 144-146.
O.H. (1968).- Studien zur Stammesgeschichte der Ammoniten. Lieferung 7.- Abhandlungen der Mathematisch-Naturwissenschaftlichen Klasse, Akademie der Wissenschaften und der Literatur in Mainz, Frankfurt am Main, vol. 3, p. 733-901.
D. (2000).- Ecological character displacement in adaptive radiation.- The American Naturalist, Chicago, vol. 156, p. S4-S16.
G. (1940a).- Paleoecological factors controlling distribution and mode of life of Cretaceous ammonoids in the Texas area.- Journal of Paleontology, Tulsa, vol. 14, p. 299-323.
G. (1940b).- Cephalopods from the Cretaceous Trinity Group of the South-Central United States.- The University of Texas, Publication, Austin, vol. 3945, p. 969-1107.
R.W. (1970).- Paleoecology and paleontology of the Lower Cretaceous Kiowa Formation, Kansas.- Paleontological Contributions of the University of Kansas, Lawrence, vol. 52, p. 1-94.
R.W. & C. (1991).- Paleontology and biostratigraphy of Cretaceous rocks, Lampazos area, Sonora, Mexico. In: E. & C. (eds.), Studies of Sonoran Geology.- Geological Society of America, Special Papers, Boulder, vol. 254, p. 51-68.
R.W. (2007).- Calibration of the Albian/Cenomanian boundary by ammonite biostratigraphy: U.S. Western Interior.- Acta Geologica Sinica, Beijing, vol. 81, p. 940-948.
R.W., D.G., R.W., B.L. & F.E. (2003).- Integrated Albian-Lower Cenomanian Chronostratigraphy Standard, Trinity River Section, Texas. In: R.W. (ed.), Cretaceous Stratigraphy and Paleoecology, Texas and Mexico.- Perkins Memorial Volume, GCSSEPM Foundation Special Publications in Geology, Houston, CD, GCS211, n° 1, p. 277-334.
R.W., F.E., D.G. & J.M. (2009).- Numerical age calibration of the Albian/Cenomanian boundary.- Stratigraphy, New York, vol. 6, p. 17-32.
Y. (1993).- Post-hatching early life history of Cretaceous Ammonoidea.- Lethaia, Oslo, vol. 26, p. 133-145.
M.P.N. (2008).- Calcareous algae from the Cretaceous of Zagros Mountain (SW Iran).- World Applied Sciences Journal, Cairo, vol. 4, p. 803-807.
L.R.L. de (1994).- Fossil molluscs of Brazil.- Boletim do Instituto Geológico, Săo Paulo, vol. 11, p. 1-202.
J.P. (1914).- Acceleration of development in fossil Cephalopoda.- Leland Stanford Junior University Publications, University Series, Stanford University, California, 30 p.
A. (1910).- Die Fauna des Aptien und Albien im nördlichen Perú. Partie I. Cephalopoden.- Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Stuttgart, Beilage-band 30, p. 313-382.
L.F. (1922).- On Cretaceous Ammonoidea from Angola collected by Professor J.W. , D.Sc., F.R.S.- Transactions of the Royal Society of Edinburgh, vol. 53, p. 91-160.
L.F. (1924).- On a new ammonite (Engonoceras iris, sp. nov.) from the Gault of Folkestone.- The Annals and Magazine of Natural History, London, (Ninth Series), vol. 14, p. 504-508.
L.F. (1930).- The fossil fauna of the Sanama Range and some neighbouring areas. Part 5. The Lower Cretaceous Ammonoidea with notes on Albian Cephalopoda from Hazara.- Palaeontologia Indica, (New Series), Geological Survey of India, Memoirs, Calcutta, vol. XV, p. 51-66.
L.F. (1931).- A monograph of the Ammonoidea of the Gault. Part VIII.- Monograph of the Palaeontographical Society, London, vol. LXXXIII, n° 379, p. 313-378.
H. von & O. (1908).- Über die Notwendigkeit einer Revision des Genus Neolobites .- Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin, Jahre 1908, p. 253-286.
L.W. (1952).- Larger invertebrate fossils of the Woodbine Formation (Cenomanian) of Texas.- U.S. Geological Survey, Professional Paper, Washington, vol. 242, 226 p.
J.D. & S.J. (2009).- Biostratigraphy and macroinvertebrate palaeontology of the petroleum-rich Belfast Mudstone (Sherbrook Group, uppermost Turonian to mid-Santonian).- Cretaceous Research, London, vol. 30, p. 873-884.
O., L., L. & I. (2007).- Aptian-Campanian ammonites of Hungary.- Geologica Hungarica, Budapest, (Series palaeontologica), vol. 57, p. 1-187.
R., P. & H. (2006).- Recent advances in research on terrestrial and marine sequences from the mid-Cretaceous Oceanic Anoxic Events (OAEs).- Scientific Drilling, Washington, n° 2, p. 50-51.
E.E. (2001).- Geology of Egypt and Lybia.- A.A. Balkema Publishers, Rotterdam, 461 p.
E.W. (1907).- The Ammonites of the Bagh Beds.- Records of the Geological Survey of India, Calcutta, vol. 36, p. 109-125.
P.D. & P.W. (1983).- Evolutionary tempo in Jurassic and Cretaceous ammonites.- Paleobiology, Lawrence, vol. 9, p. 183-198.
T. (2002).- Stratigraphy and correlation of the Lower Cretaceous exposures across the Dead Sea Transform with emphasis on tracing the Amir Formation in Jordan.- Israel Journal of Earth Sciences, Jerusalem, vol. 51, p. 55-78.
G.E.G. (1990).- New developments in ecology of Jurassic-Cretaceous ammonoids. In: G., F., S. & M. (eds.), Fossili, evoluzione, ambiente.- Atti II° Convegno Internazionale (Pergola, 25-30 Ottobre 1987), Technostampa, Ostra Vetere, p. 459-478.
G.E.G. (1996).- Ammonoid life and habitat. In: N.H., K. & R.A. (eds.), Ammonoid paleobiology.- Topics in Geobiology, Plenum Press, New York, vol. 13, p. 607-707.
J. (1979).- Mid Cretaceous events. Iberian field conference 77. Guide II Partie. Itinéraire géologique à travers le Crétacé moyen des Chaînes Vascogotiques et Celtibériques (Espagne du Nord).- Cuadernos de Geología Ibérica, Madrid, vol. 5, p. 127-214.
F. & F. (2005).- The upper Cenomanian (Cretaceous) ammonite Neolobites vibrayeanus (d', 1841) in the Middle East: taxonomic and palaeoecologic remarks.- Cretaceous Research, London, vol. 26, p. 930-946.
M. (1996).- The Cenomanian of northern Cantabria (N-Spain): facies development and sequential subdivision. In: W. (ed.), Jost Symposium.- Cretaceous stratigraphy, paleobiology and paleobiogeography (Tübingen, 7-10 March 1996), Abstracts, University of Kiel, p. 181-187.
G.D. & A. (1998).- Palynology, palynofacies, geochemistry and invertebrate paleontology of the Upper Cretaceous Napo Formation, Oriente Basin, Ecuador.- Thirty-first Annual AASP Meeting (October 27-31, 1998, Ensenado, Baja California, Mexico), Abstracts, p. 52.
C.W. (1952).- A classification of Cretaceous ammonites.- Journal of Paleontology, Tulsa, vol. 26, p. 213-222.
C.W. (1963).- Cretaceous ammonites from Bathurst Island, northern Australia.- Palaeontology, London, vol. 6, p. 597-614.
C.W., J.H. & M.K. (eds.) (1996).- Cretaceous Ammonoidea. In: R.L. (ed.), Mollusca 4 (revised).- Treatise on Invertebrate Paleontology, Geological Society of America, New York; University of Kansas, Lawrence, Part L, 362 p.
M.M. (2003).- Controls on shell shape in Acanthoceratid ammonites from the Cenomanian-Turonian Western Interior seaway. In: P.J. (ed.), High-resolution approaches in stratigraphic paleontology.- Topics in Geobiology, Plenum Press, New York, vol. 21, p. 195-226.
M.M. (2004).- Neogastroplites meets Metengonoceras: morphological response of an endemic hoplitid ammonite to a new invader in the mid-Cretaceous Mowry Sea of North America.- Cretaceous Research, London, vol. 25, p. 927-944.
K. (1972).- Cretaceous paleogeography: implications of endemic ammonite faunas.- Bureau of Economic Geology, University of Texas Geological Circular, Austin, vol. 72, p. 1-13.
K.A. (1884).- Cephalopoda. In: K.A. (ed.), Handbuch der Paläontologie, Band 1, Abteilung 2, Lieferung 3.- Druck und Verlag von R. Oldenbourg, München und Leipzig, p. 329-522.
T., M., H. & M.H. (2009).- Seismic sequence stratigraphy, basin structuring, and hydrocarbon implications of Cretaceous deposits (Albian-Maastrichtian) in central Tunisia.- Cretaceous Research, London, vol. 30, p. 1-21.
| Species | Stratigraphical distribution | Geographical distribution | References | |
| Glottoceras , 1875 | Albian | South America | ||
| 1 | andinum (, 1970) | Upper Albian | Venezuela | , 1970 |
| 2 | attenuatum , 1875 | ?Cenomanian | Celendín, Peru | , 1903; , 1938 |
| Albian-Cenomanian | Oriente Basin, Ecuador | & , 1998 | ||
| Albian | Une, Une Formation, Colombia | , 2002 | ||
| Middle Albian | northern Peru | et alii, 1947; , 1956 | ||
| Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| 3 | bassleri ( in et alii, 1947) | Aptian | En Huanco, Celendin, Peru | et alii, 1947 |
| 4 | crassicostatum (, 1910) | Middle Albian | Peru | et alii, 1947 |
| Lower-Middle Albian | Pongo de Rentemo, Peru | et alii, 1998 | ||
| 5 | crassinodosum (, 1910) | Middle Albian | Condebamba, northern Peru | , 1910; et alii, 1947 |
| Lower-Middle Albian | Peru | et alii, 2003; & , 2004 | ||
| 6 | gabbi (, 1903) | ?Cenomanian | Quebrada de Huari, Peru | , 1903; , 1938 |
| Middle Albian | Celendín, Sunchubamba, northern Peru | , 1956 | ||
| upper Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| 7 | gracilecostatum (, 1910) | Lower Albian | northern Peru | et alii, 2003 |
| 8 | laraense (, 1970) | Middle Albian-Upper Albian | Misahualli, Ecuador; Venezuela | et alii, 2007 |
| 9 | largum , 2002 | Lower Albian | La Cuenca, Peru | et alii, 2003 |
| 10 | libertadense (, 1952) | Albian | Colombia | , 1952 |
| 11 | moorei ( in et alii, 1947) | upper Middle Albian | Peru | et alii, 2003; & , 2004 |
| Middle Albian | Peru | et alii, 1947 | ||
| Misahualli, Ecuador | et alii, 2007 | |||
| 12 | ollonense (, 1877) | Lower Albian | La Cuenca, Peru | et alii, 2003 |
| Sunchubamba, northern Peru | , 1956; et alii, 1998 | |||
| 13 | ovale (, 1956) | Middle Albian | Uchupata, northern Peru | , 1956 |
| 14 | pacificum (, 1956) | Lower Albian | Celendín, Peru | , 1956 |
| 15 | pegnai (, 1979) | Cretaceous | Colombia | , 1979 |
| 16 | raimondii (, 1908) | Middle Albian | Cajamarca, Sihuas, Pomachaca, northern Peru | , 1910; et alii, 1947; , 1956; et alii, 1998 |
| upper Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| 17 | semicostatum (, 1910) | Middle Albian | Cochamarca, northern Peru | , 1910; et alii, 1947 |
| upper Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| 18 | seminodosum (, 1910) | Middle Albian | Peru | & , 2004 |
| 19 | sommermeieri ( in et alii, 1947) | Middle Albian | Peru | et alii, 1947 |
| Misahualli, Ecuador | et alii, 2007 | |||
| 20 | spinosum (, 1910) | Middle Albian | northern Peru | et alii, 1947 |
| Lower Albian (mammillatum Zone) | Bathurst Island, North Australia | , 1963 | ||
| 21 | tardum (, 1956) | Middle Albian | Sihuas, northern Peru | , 1956 |
| Lower Albian | La Cuenca, Peru | et alii, 2003 | ||
| 22 | triangulare (, 1956) | Middle Albian | Sihuas, northern Peru | , 1956 |
| 23 | typicum (, 1910) | Middle Albian | northern Peru | & , 2004 |
| Engonoceras & , 1881 | Lower Albian-Middle Cenomanian | Tethyan | ||
| 1 | belviderense (, 1894) | Lower Cretaceous | Belvidere, Kansas | , 1903; , 1987 |
| Upper Albian | Texas, USA | et alii, 2003 | ||
| Neocomian | Kansas, USA | , 1970; et alii, 1998a | ||
| Middle-Upper Albian | Sonora, Mexico | , 1938; & , 1991 | ||
| 2 | commune , 1903 | Cretaceous | Texas | , 1903 |
| 3 | complicatum , 1903 | Albian | Texas | , 1903; et alii, 1998a |
| Upper Albian | El Mizab, Meknes, Atlas, Morocco | & , 1968 | ||
| 4 | elegans et alii, 1998a | Albian | Texas | et alii, 1998a |
| 5 | emarginatum (, 1893) | Albian | Pleasant Point, Texas, USA | , 1903 |
| 6 | gibbosum , 1903 | Albian | Texas, USA | , 1903; et alii, 1998a |
| Middle Albian | et alii, 2003 | |||
| 7 | grimsdalei , 1931 | Upper Albian | Weymouth, Dorset, England | , 1931; , 1956 |
| 8 | hilli , 1898 | Albian | Texas, USA | et alii, 1998a |
| North Peruvian Andes | et alii, 2003 | |||
| Middle-Upper Albian | Texas, USA | et alii, 2003 | ||
| 9 | iris , 1924 | Middle Albian | Folkestone, England | , 1938; , 1924 |
| 10 | jezzinense , 1940 | Albian | Djebel Tebega, Tunisia | , 1956 |
| Jezzine, Lebanon | , 1940 | |||
| 11 | khenchelaensis (, 1940) | Albian-Cenomanian | Syria | et alii, 1998a |
| Albian | Bhamdoum, Lebanon | , 1961 | ||
| 12 | olgaluciae , 1979 | Middle Albian | Colombia | , 1979 |
| 13 | pierdenalis (, 1848) | Upper Cretaceous | Fredericksburg, Texas | , 1848 |
| Albian | Texas | , 1903; , 1938 | ||
| Upper Albian | Chihuahua, Mexico | , 1910 | ||
| Middle Albian | Sonora, Mexico | , 1988; & , 1991 | ||
| 14 | pinax (, 1902) | Cenomanian | Temojoh, West Borneo | , 1902; , 1940; , 1955 |
| 15 | retardum , 1903 | lowest Cenomanian | Pottsboro, Texas, USA | , 1903; & , 1989; et alii, 2005 |
| 16 | serpentinum (, 1900) | Lower Cenomanian | Texas | , 1982 |
| Upper Albian | , 1938; et alii, 1996 | |||
| uppermost Albian (dispar Zone) | , 1903; et alii, 1998b; , 2004; et alii, 2005 | |||
| 17 | subjectum , 1903 | uppermost Albian (dispar Zone) | Texas | , 1903; et alii, 1998b |
| 18 | stolleyi , 1898 | Albian | Texas | et alii, 1998a |
| Upper Albian | Chihuahua, Mexico | , 1910 | ||
| Middle-Upper Albian | Sonora, Mexico | & , 1991 | ||
| Middle Albian | , 1988 | |||
| La Cuenca, Peru | et alii, 2003 | |||
| 19 | thomasi , 1907 | Cenomanian | Rekkame, Middle Atlas, Morocco | & , 2009 |
| U. Albian-L. Cenomanian | Guelta, Tunisia | , 1938; , 1989 | ||
| 20 | toussainti , 1907 | U. Albian-L. Cenomanian | Tebaga, Tunisia | , 1938; , 1989 |
| 21 | uddeni (, 1900) | Comanche series, Kiowa shale | Kansas, USA | , 1903 |
| Middle Albian | Sonora, Mexico | , 1988 | ||
| 22 | vicorpense (, 1940) | Albian-Cenomanian | Syria | et alii, 1998a |
| Upper Albian | Djebel Naimia, Chotts, Tunisia | et alii, 2008b | ||
| 23 | wilkinsoni , 1956 | ?Upper Albian | Oregon, USA | , 1956 |
| 24 | zumoffeni (, 1940) | Albian-Cenomanian | Syria | et alii, 1998a |
| Neolobites , 1882 | Cenomanian | Tethyan | ||
| 1 | bassleri , 1926 | Cenomanian | Caminaca, Peru | , 1926; & , 1936 |
| Middle-Upper Cenomanian | Altiplano, Peru | , 1995; et alii, 1998 | ||
| 2 | brancai & , 1908 | U. Cenomanian- L. Turonian | Egypt | & , 1908; , 1914; , 1928 |
| Upper Cenomanian | Central High Atlas, Morocco | & , 2002 | ||
| Upper Cenomanian | Gafsa, Central Tunisia | & , 2005 | ||
| 3 | bussoni , 1965 | Cenomanian | Tinrhert, Sahara, Algeria | , 1965; et alii, 1981 |
| Upper Cenomanian | Ourém, Portugal | & , 1978, 1979 | ||
| 4 | choffati , 1903 | Cenomanian | Portugal | , 1903 |
| 5 | fourtaui , 1907 | Cenomanian | Egypt, Tunisia | , 1907; , 1915 |
| Upper Cenomanian | Tarfa, Arabic Desert | , 1928 | ||
| Tinrhert, Sahara, Algeria | , 1965; et alii, 1981 | |||
| Middle-Upper Cenomanian | Palmyrids, Central Syria | et alii, 2003 | ||
| 6 | isidis , 1915 | Cretaceous | Egypt | , 1915 |
| Upper Cenomanian | Gafsa, Central Tunisia | & , 1996 | ||
| 7 | kummeli , 1956 | base Upper Cenomanian | Celendín, Northern Peru | , 1956 |
| Bolivia | , 1968; & , 1992 | |||
| 8 | maresi (, 1862) | Cenomanian | Egypt | , 1862; , 1915; & , 1984 |
| 9 | medeninensis , 1954 | Upper Cenomanian | Tunisia | , 1954; & , 2005 |
| 10 | peroni , 1903 | Upper Cenomanian | Tinrhert, Sahara, Algeria | , 1910; , 1965; et alii, 1981 |
| Cenomanian | North Africa | , 1903 | ||
| Egypt | , 1914; , 1928 | |||
| Provence (France), Tunisia | , 1938 | |||
| 11 | pervinquieri & , 1908 | Upper Cenomanian | Egypt | & , 1908; & , 2005 |
| schweinfurthi & , 1908 | U. Cenomanian-L. Turonian | Egypt | & , 1908; , 1914; , 1928 | |
| Turonian | Egypt | , 1910 | ||
| 12 | vibrayeanus (d', 1841) | Cretaceous | Vibraye, Sarthe, France | d', 1841; , 1848 |
| Cenomanian | Celendín, Northern Peru | , 1926 | ||
| El Kisba, High Atlas, Morocco | & , 2009 | |||
| France | , 1942; et alii, 1996 | |||
| France, Portugal | , 1903 | |||
| Upper Cenomanian | Jordan | et alii, 2008 | ||
| Central Oman | & , 1991 | |||
| Estremadure, Lisbon, Portugal | et alii, 1979 | |||
| Vouillac, France | et alii, 1983 | |||
| Tinrhert, Sahara, Algeria | , 1965; et alii, 1981; et alii, 1996 | |||
| Egypt, Jordan | & , 2005 | |||
| Gafsa, Central Tunisia | & , 2005 | |||
| Tanout, Niger | et alii, 1992 | |||
| Burgos, Spain | , 1979 | |||
| Knemiceras , 1898 | ?Upper Aptian-Albian | Tethyan | ||
| as Knemiceras sp. | Upper Albian | Río Aguamulac, Huescar, Spain | et alii, 1982 | |
| Middle Albian | Aube, France | & , 1984 | ||
| 1 | aegyptiacum , 1955 | Upper Albian-Lower Cenomanian | Kazhdumi, southern Iran | , 1981 |
| Upper Albian | Gafsa, Central Tunisia | et alii, 2008 | ||
| Oued Zitoun, North Tunisia | & , 1986 | |||
| 2 | arambourgi , 1940 | Cretaceous | Syria, Lebanon | , 1940; & , 1991 |
| 3 | collignoni , 1955 | Cretaceous | Sinai, Egypt | , 1955 |
| Upper Albian-Lower Cenomanian | Kazhdumi, southern Iran | , 1981 | ||
| 4 | compressum , 1903 | ?Cenomanian | Beirut, Lebanon | , 1903; , 1938 |
| Albian | Nar, Kartang, Jahrum, southern Iran | , 1981 | ||
| Upper Albian | Venezuela | , 1970 | ||
| Oued Zitoun, North Tunisia | & , 1986 | |||
| Djebel Naimia, Chotts, Tunisia | et alii, 2008b; & , 2008 | |||
| Middle Albian | Bilbao, northern Spain | et alii, 2009 | ||
| 5 | douvillei , 1940 | Cretaceous | Djebel Dourra Syria | , 1940 |
| Upper Albian-Lower Cenomanian | Kazhdumi, southern Iran | , 1981 | ||
| Upper Albian | Lurestan, Iran | , 2007 | ||
| 6 | dubertreti , 1940 | Cretaceous | Djebel Dourra, Syria | , 1940 |
| Albian | Mderedj, Lebanon | , 1961 | ||
| Upper Albian | Lurestan, Iran | , 2007 | ||
| Lower-Middle Albian | Central Oman | & , 1991 | ||
| 7 | flexiloculosum , 1940 | Cretaceous | Syria | , 1940 |
| Middle Albian | Dead Sea, Israel | , 2002 | ||
| Lower Albian | Guajira, NE Colombia | , 2002 | ||
| 8 | iraniense , 1981 | U. Albian-L. Cenomanian | Kazhdumi, southern Iran | , 1981 |
| Middle-Upper Albian | Bangestan, SW Iran | , 2010 | ||
| 9 | kazhdumiense , 1981 | U. Albian-L. Cenomanian | Kazhdumi, southern Iran | , 1981 |
| Middle-Upper Albian | Bangestan, SW Iran | , 2010 | ||
| 10 | orientalis , 1955 | Middle Albian | Moghara, Egypt | , 1955 |
| 11 | persicum , 1981 | U. Albian-L. Cenomanian | Kazhdumi, southern Iran | , 1981 |
| Lower Albian | Bangestan, Lurestan, Iran | , 2007, 2010 | ||
| 12 | rittmanni , 1955 | Middle Albian | Moghara, Egypt | , 1955; & , 1955; , 2001 |
| 13 | saadense ( & , 1890) | Cretaceous | Tunisia | & , 1890 |
| Upper Albian | Salazac, Gard, France | , 1940; , 1989; , 2008 | ||
| Algeria | , 1910; , 1938; , 1941 | |||
| 14 | saharae , 1965 | Albian | Tebaga de Kebili, South Tunisia | , 1965 |
| Sinai, Egypt | et alii, 1997 | |||
| 15 | ? sinaiticum | Upper Albian | Gafsa, Central Tunisia | et alii, 2008 |
| 16 | spathi , 1955 | U. Albian-L. Cenomanian | Nar, Gavbasht, southern Iran | , 1981 |
| Upper Albian | Oued Zitoun, North Tunisia | & , 1986 | ||
| Lower Albian | northern Sinai, Egypt | et alii, 1997; , 2001; , 2008 | ||
| Bangestan, SW Iran | , 2010 | |||
| 17 | subcompressum , 1903 | ? Cenomanian | Gilead Mts. Jordan; Mt. Lebanon, Syria | , 1903; et alii, 1997 |
| 18 | syriacum (, 1848) | Lower Cretaceous | Lebanon | , 1848 |
| Angola | , 1931; , 1938 | |||
| Cenomanian | Mt. Lebanon, Syria | , 1903 | ||
| U. Albian-L. Cenomanian | Kazhdumi, southern Iran | , 1981 | ||
| Upper Albian | Portugal | , 1938 | ||
| Lurestan, Iran | , 2007 | |||
| Syria | et alii, 1996; et alii, 2003 | |||
| Middle-Upper Albian | northern Sinai, Egypt | et alii, 1997 | ||
| Jawan area, NW Iraq | , 2006 | |||
| Middle Albian | Tebaga, central Tunisia | et alii, 1985 | ||
| Moghara, Egypt | , 1916, 1928; & , 1955 | |||
| Dead Sea, Israel | , 2002 | |||
| Cajamarca, Uchupata, Peru | , 1956; et alii, 1998 | |||
| Lower Albian | northern Sinai, Egypt | , 2008 | ||
| 19 | uhligi (, 1886) | ? Cenomanian | Portugal | , 1903; , 1938 |
| Upper Albian | Santander, Cantabria, Spain | , 1996 | ||
| Teruel, Valencia, Spain | , 1995a, 1995b | |||
| Estremadure, Lisbon, Portugal | , 1941; et alii, 1979, , 1981 | |||
| Salinas (Angola); Moghara (Egypt) | , 1928, 1931 | |||
| Lurestan, Iran | , 2007 | |||
| Middle-Upper Albian | northern Sinai, Egypt | et alii, 1997 | ||
| Tebaga, central Tunisia | , 1965 | |||
| Muscat, Oman | et alii, 2000 | |||
| Lower Albian | Lurestan, Iran | , 2007 | ||
| northern Sinai, Egypt | , 2008 | |||
| Protengonoceras , 1903 | Lower/Middle Albian-?Lower Cenomanian | Tethyan | ||
| 1 | dufaurei , 1981 | Lower Cenomanian | Kazhdumi, southern Iran | , 1981 |
| 2 | gabbi (, 1898) | Albian | Sonora (Mexico), Texas | , 1903; et alii, 1998a |
| Upper Albian | northern Sinai, Egypt | , 2008 | ||
| Middle Albian | Texas | et alii, 1996 | ||
| Peru | et alii, 1998 | |||
| 3 | planum , 1903 | Upper Cretaceous | Dallas County, Texas | , 1903; , 1942 |
| 4 | ? prestati , 1981 | U. Albian-L. Cenomanian | Kazhdumi, southern Iran | , 1981 |
| Metengonoceras , 1903 | Middle Albian-Cenomanian | cosmopolitan | ||
| as Epengonoceras sp. | Upper Albian | Lower Saxony Basin, Germany | , 2007 | |
| as Metengonoceras sp. | uppermost Cenomanian | Sahara, Africa | , 2007 | |
| lower Upper Cenomanian | Sayreville, NJ, USA | & , 1990 | ||
| 1 | acutum , 1903 | Upper Cenomanian | North Central Texas | , 1903; , 1942; , 1988; & , 1990; et alii, 2003 |
| 2 | ambiguum , 1903 | Lower Cretaceous | Texas, USA | , 1903 |
| Albian | et alii, 1998a | |||
| Aptian-Albian | , 1938 | |||
| 3 | arnaudi , 1912 | Cenomanian | Touvois, Vendée, France | , 1942 |
| Charentes, France | et alii, 1981 | |||
| 4 | aspenanum ( & , 1931) | Cenomanian | Montana, Wyoming, USA | & , 1989; , 2004 |
| uppermost Albian | Montana, USA | et alii, 2003 | ||
| 5 | bedoti (, 1912) | Lower Cenomanian | Vendée, France | , 1942; et alii, 1981; & , 1984; et alii, 1992 |
| 6 | bravoense (, 1928) | Cenomanian [M. (P.) rhacioformis Zone] | Texas | , 1979 |
| lowermost Cenomanian | Texas, USA | et alii, 2005 | ||
| Albian-Cenomanian | Oriente Basin, Ecuador | & , 1998 | ||
| 7 | ? dibbleyi | Upper Cenomanian | Ouled nail, Saharan Atlas, Algeria | , 1999; et alii, 2008 |
| 8 | douvillei , 1912 | Cenomanian | Vendée, France | et alii, 1981 |
| 9 | dumbli (, 1893) | Cenomanian | Texas, USA | , 1942; et alii, 1996 |
| uppermost Cenomanian | Jordan | et alii, 2008 | ||
| Benue trough, Nigeria | et alii, 1998 | |||
| Upper Cenomanian | Mts. Iguellala, Niger | et alii, 1992 | ||
| Sinai, Egypt | & , 2001 | |||
| Le Mans, France | & , 1984 | |||
| Angers, France | et alii, 1981 | |||
| Middle-Upper Cenomanian | Gafsa, Central Tunisia | & , 2005 | ||
| Colorado, USA | & , 1972 | |||
| North Central Texas | & , 1990; et alii, 2003 | |||
| 10 | inscriptum , 1903 | Lower Cretaceous | Texas, USA | , 1903 |
| Upper Albian | et alii, 1996 | |||
| Middle Albian | & , 1989 | |||
| Albian | et alii, 1998a | |||
| Aptian-Albian | , 1938 | |||
| 11 | nigeriensis , 1935 | Upper Cenomanian | Niger | et alii, 1981 |
| 12 | oliveirai , 1967 | Albian-Cenomanian | Rio Fundo, Sergipe, Brazil | , 1986; , 1994 |
| 13 | teigenense & , 1989 | Cenomanian | Montana, USA | & , 1989; , 2004 |
| Lower Cenomanian | Normandie, France | et alii, 2002 | ||
| U. Albian-lowermost Cenomanian | Montana, USA | et alii, 2003 | ||
| 14 | tolveiense , 1912 | Cenomanian | Vendée, France | et alii, 1981 |
| Hypengonoceras , 1922 | Upper Albian | Tethyan | ||
| as Hypengonoceras sp. | Upper Albian | Makhtesh Ramon, Israel | , 1981 | |
| as Hypengonoceras sp. ind. | Upper Albian | Salazac, Gard (France) | , 1989 | |
| 1 | chouberti , 1966 | Upper Albian | Tarfaya, Morocco | , 1966; & , 1989 |
| 2 | decaryi , 1963 | Upper Albian | Madagascar | , 1963 |
| Zululand, Mozambique | , 1975; & , 1979 | |||
| Zufia, Navarra, Spain | , 1979 | |||
| 3 | fauremuretae , 1966 | Upper Albian | Tarfaya, Morocco | , 1966; & , 1989 |
| 4 | ? ibericum & , 1977 | Albian | Albacete, SE Spain | & , 1977; & , 1989 |
| 5 | tarfayense , 1966 | Upper Albian | Tarfaya, Morocco | , 1966; & , 1989 |
| 6 | warthi (, 1895) | Albian | Albacete, SE Spain | & , 1977 |
| Upper Albian | South India | , 1960; et alii, 1996 | ||
| Diego Suarez, Madagascar | , 1960 | |||
| Parengonoceras , 1924 | Albian | Tethyan | ||
| as Parengonoceras sp. juv. | Upper Albian | Suriname | & , 2006 | |
| as Parengonoceras sp. | Lower Albian | SW Iran | , 2010 | |
| as Parengonoceras sp. | Lower Albian | Kent, England | , 1957, p. 42 | |
| 1 | algerianum (, 1953) | lowermost Albian | Mellégue, Algeria | & , 2004 |
| 2 | azlense (, 1940b) | Upper Albian | Fort Worth, Texas | , 1940b |
| 3 | barbacoense , 1970 | Upper Albian | Barbacoas, Venezuela | , 1970 |
| 4 | bussoni , 1965 | Upper Albian | Tebaga de Kebili, South Tunisia | , 1965 |
| 5 | caneroti & , 1981 | uppermost Albian | Castellon, Spain | & , 1981 |
| Lower Albian | Maestrat Basin, Spain | et alii, 2008a | ||
| 6 | champaraense , 1956 | Middle Albian | Sihuas, northern Peru | , 1956 |
| Lower Albian | La Cuenca, Peru | et alii, 2003 | ||
| 7 | discoides , 1970 | Upper Albian | Barbacoas, Venezuela | , 1970 |
| 8 | duartei (, 1979) | Lower Albian | Corral Quemado, Peru | et alii, 2003; & , 2004 |
| 9 | duboisi (, 1989) | Upper Albian | Salazac, Gard, France | , 1989; , 2008 |
| Bakony Mts., Hungary | , 1990a, 1990b | |||
| 10 | duplicatum , 1970 | Upper Albian | Barbacoas, Venezuela | , 1970 |
| Basque-Cantabrian Basin, N Spain | , 1970; et alii, 2009 | |||
| 11 | ebrayi (, 1882) | Albian | Cosne, France | , 1928, , 1938 |
| Upper Albian | Santander, Cantabria, Spain | , 1996 | ||
| Middle Albian | France | et alii, 1996 | ||
| Perchois, Aube, France | , 1979; , 2008 | |||
| Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| Maestrat Basin, Spain | et alii, 2008a | |||
| 12 | elegans , 1970 | Upper Albian | Barbacoas, Venezuela | , 1970 |
| 13 | guadaloupaeforme (, 1910) | Middle Albian | northern Peru | , 1910; et alii, 1998 |
| Cajamarca, Condebamba, northern Peru | , 1956; et alii, 1998 | |||
| Lower Albian | La Cuenca, Peru | et alii, 2003 | ||
| 14 | haasi , 1956 | Lower Albian | Celendín, northern Peru | , 1956 |
| upper Lower Albian | Peru | et alii, 2003; & , 2004 | ||
| 15 | icaunensis (, 1853) | Albian | Ravennes, Yonne, France | , 1853; , 1978; , 2008 |
| 16 | mahmoudi (, 1953) | lowermost Albian | Mellégue, Algeria | , 1953; & , 2004 |
| England | , 1978 | |||
| 17 | nodosum (, 1940b) | Upper Albian | Glen Rose, Texas | , 1940b |
| 18 | pernodosum (, 1910) | Lower Albian | Cajabamba, northern Peru | , 1910; et alii, 2003; & , 2004 |
| Middle Albian | northern Peru | , 1956; et alii, 1998 | ||
| La Cuenca, Peru | et alii, 2003 | |||
| 19 | roemeri (, 1893) | Cenomanian | Texas | , 1905 |
| Upper Albian | Glen Rose, Texas | , 1940b | ||
| Albian | Texas | , 1903; et alii, 1998a | ||
| 20 | tetranodosum (, 1925) | Middle Albian | Celendín, Cajamarca, northern Peru | , 1956 |
| uppermost Lower Albian | Cajamarca, northern Peru | et alii, 1998 | ||
| 21 | trinitense (, 1940b) | Upper Albian | Fort Worth, Texas | , 1940b |
| 22 | zagrosiense , 1981 | ? Lower Cenomanian | Kazhdumi, southern Iran | , 1981 |
| 23 | ziczag (, 1952) | Middle Albian | Uchupata, northern Peru | , 1956; et alii, 1998; & , 2004 |
| Platiknemiceras , 1954 | uppermost Aptian-Lower Cenomanian | Tethyan | ||
| 1 | bassei , 1954 | Lower-Middle Albian | Spain | , 1961 |
| Lower Albian | et alii, 1996 | |||
| Maestrat Basin, Cataluna, Spain | et alii, 1994; et alii, 2008a | |||
| uppermost Aptian | SW Iran | , 2010 | ||
| 2 | caseyi et alii, 1980 | Lower Albian | Kyushu, Japan | et alii, 1980 |
| 3 | colombiana , 1979 | upper L. Albian-lower M. Albian | Colombia | , 1979, 1981; , 1999 |
| 4 | deserti (, 1955) | Upper Albian | Utrillas Formation, SE Spain | et alii, 1993 |
| Lower Albian | Moghara, Egypt | , 1955; & , 1955; , 1961 | ||
| Lurestan, Iran | , 2007 | |||
| 5 | flexuosum et alii, 1998a | Lower-Middle Albian | Texas, USA | et alii, 1998a |
| upper Lower Albian | Peru | & , 2004 | ||
| 6 | gracile (, 1916) | Albian | Mderedj, Lebanon | , 1940; , 1961 |
| Upper Albian | Utrillas Formation, Spain | et alii, 1993 | ||
| Oued Zitoun, North Tunisia | & , 1986 | |||
| Gafsa, Central Tunisia | & , 2008 | |||
| Middle Albian | Tebaga, central Tunisia | , 1965; et alii, 1985 | ||
| Dead Sea, Israel | , 2002 | |||
| Lower Albian | Moghara, northern Sinai, Egypt | , 1928; & , 1955; & , 1999; , 2001; , 2008 | ||
| 7 | hachourii (, 1953) | lowermost Albian | Mellégue, Algeria | , 1953; & , 2004 |
| basal Albian | Guajira, NE Colombia | , 2002 | ||
| 8 | jullieni (, 1940) | Upper Albian | El Mizab, Meknes, Atlas, Morocco | & , 1968 |
| Albian | Khenchela, Algeria | , 1961 | ||
| 9 | sequanense , 1979 | Lower-Middle Albian | Aube, France | , 1979 |
| 10 | subcomplicatum (, 1940) | Albian | Lebanon | , 1940; , 1961 |
| 11 | valencianum & , 1980 | uppermost Albian-lowermost Cenomanian | Valencia, Spain | & , 1980; et alii, 2004 |
Table 1: Geographical and stratigraphical distribution of engonoceratid species. Type species are indicated in bold.