◄ Carnets Geol. 15 (17) ►
![]()
Contents
[1. Introduction]
[2. Geological background and localities] [3. Material and methods]
[4. Results] [5. Discussion] [6. Conclusions]
and ... [Bibliographic references]
Department of Geology, University of Tartu, Ravila 14A, EE-50411 Tartu (Estonia)
Department of Geology, College of Wooster, Wooster, Ohio 44691 (USA)
School of Earth Sciences, 155 South Oval Mall, Ohio State University, Columbus 43210 (USA)
Institute of Geology, Tallinn University of Technology, Ehitajate tee 5, EE-19086 Tallinn (Estonia)
Published online in final form (pdf) on November 18, 2015
[Editor: Brian ; technical editor: Christian ]
![]()
Rare pits attributed to Tremichnus have been found in crinoids from the Silurian of Estonia. The Rhuddanian Tremichnus is the earliest symbiont in crinoid columnals of Baltica. These pits presumably were domiciles of unknown organisms. Tremichnus had a negative effect on the host crinoid as demonstrated by swollen columnals. Tremichnus in the Silurian of Estonia is less common than similar traces in the Silurian of nearby Gotland. The most important aspect of this study is the rarity of this interaction in these samples in contrast to most other samples of comparable age elsewhere. These structures have a very patchy distribution.
Trace fossils; Tremichnus; symbiosis; crinoids; Silurian; Estonia.
O., M.A., W.I. & U. (2015).- Tremichnus in crinoid pluricolumnals from the Silurian of western Estonia (Baltica).- Carnets Geol., Madrid, vol. 15, nº 17, p. 239-243.
Tremichnus dans des segments de tiges de crinoïdes du Silurien d'Estonie occidentale (bouclier balte).- Quelques petites cavités attribuées à Tremichnus ont été trouvées dans des crinoïdes provenant du Silurien d'Estonie. Ce Tremichnus rhuddanien est le symbiote le plus ancien observé dans des segments de tiges de crinoïdes du bouclier balte. Ces cavités correspondent vraisemblablement à des habitats d'organismes inconnus. Tremichnus a eu un impact négatif sur le crinoïde hôte comme en attestent les renflements des tiges. Tremichnus est moins fréquent dans le Silurien d'Estonie que des traces semblables dans le Silurien du Gotland voisin. L'aspect le plus significatif de cette étude est la rareté de cette interaction dans ces échantillons à la différence de la plupart des échantillons du même âge provenant d'autres localités. Ces structures ont une répartition très irrégulière.
Traces fossiles ; Tremichnus ; symbiose ; crinoïdes ; Silurien ; Estonie.
The Phanerozoic record of symbiosis often relies on trace fossils (, 2005). These trace fossils commonly include borings associated with shell repair in various invertebrates. The earliest symbiotic borings are known from Ordovician brachiopods, bryozoans and echinoderms ( & , 1988; , 2004; et al., 2014). Paleozoic symbiotic borings are especially common in echinoderms (, 1985; et al., 2014) and in the shells of brachiopods and mollusks (, 2004). The Ordovician was a time of major diversification of invertebrates, and many new symbiotic interactions appeared by the Late Ordovician involving mostly larger colonial animal hosts, but also echinoderms (, 2005). During the Silurian the diversity of symbiotic interactions increased further (, 2005). The most common endobiotic symbiont in crinoids is Tremichnus 1978, 1985). Tremichnus is an embedment structure formed by an organism that inhibited growth of the crinoid host stereom , 1978, 1985; & 2014; et al., 2015).
Trace fossils in the Silurian of Baltica are poorly known, and most are not related to symbiosis. Traces caused by symbionts have previously been described from Silurian corals and stromatoporoids of Estonia ( et al., 2014; & , 2014a, 2014b). Silurian crinoids of Estonia are relatively well studied ( et al., 2012, 2015), but their ichnofossils have mostly remained undescribed.
The aims of this paper are: 1) to describe Tremichnus from crinoids of the Silurian of Estonia; 2) to assess the frequency of Tremichnus in crinoids; 3) to discuss the ethology of Tremichnus in the crinoids.
During the Silurian the Baltica palaeocontinent was located in equatorial latitudes, slowly moving northward ( et al., 2004; & , 2005). The western part of modern Estonia was covered by the shallow epicontinental Baltic Basin (Fig. 1 ![]() ). The tropical Baltic Basin was characterized by diverse biota and environments. Organic buildups were common, especially coral-stromatoporoid reefs ( & 1997).
). The tropical Baltic Basin was characterized by diverse biota and environments. Organic buildups were common, especially coral-stromatoporoid reefs ( & 1997).
Click on thumbnail to enlarge the image.
Figure 1: Locality map of crinoids with symbiotic borings.
& (1977) described the paleoenvironments of the Baltic Basin in detail. They reported the following facies belts: tidal flat/lagoonal, shoal, open shelf, transitional (i.e., basin slope), and a basin depression. The first three facies belts formed a carbonate platform. A deep pericratonic basin with fine-grained siliciclastic deposition formed the latter two facies belts ( & , 1997).
Hilliste quarry (GPS: 58.873889; 22.971944) (Fig. 1 ![]() ) is located 1km east from Pühalepa Church. The fossiliferous limestones of the Hilliste Formation (Rhuddanian) have abundant favositids, stromatoporoids, and encrusting algae; they belong to the shoal facies belt. Sepise alvar (GPS: 58.455128; 22.007986) (Fig. 1
) is located 1km east from Pühalepa Church. The fossiliferous limestones of the Hilliste Formation (Rhuddanian) have abundant favositids, stromatoporoids, and encrusting algae; they belong to the shoal facies belt. Sepise alvar (GPS: 58.455128; 22.007986) (Fig. 1 ![]() ) is a low escarpment limestone exposure located in Sepise Village. Fossiliferous limestones of the Jaagarahu Formation (Sheinwoodian) exposed here are rich in tabulate corals and stromatoporoids.
) is a low escarpment limestone exposure located in Sepise Village. Fossiliferous limestones of the Jaagarahu Formation (Sheinwoodian) exposed here are rich in tabulate corals and stromatoporoids.
An echinoderm collection from the Silurian of Estonia containing more than 1000 crinoid specimens, mostly columnals, was searched for signs of syn vivo borings. The studied collection, acquired by numerous collectors during the past 50 years, is housed in the Institute of Geology, Tallinn University of Technology. In addition, crinoid specimens collected during fieldwork in the Hilliste quarry were examined for borings. Hilliste quarry specimens are housed in the Natural History Museum (Museum of Geology), University of Tartu.
Tremichnus is very rare in the Silurian crinoids of Estonia. It occurs only in a single specimen from the Hilliste Formation and in a single specimen from the Jaagarahu Formation.
One crinoid pluricolumnal with a single Tremichnus was found in the Rhuddanian (Hilliste Formation) of Hiiumaa Island (Fig. 2 ![]() ). The boring is oval with a somewhat irregular outline, and is moderately deep in the column wall. It has a rounded edge and shows partial repair. The columnal is strongly swollen around the pit. The pit is 4.7 mm long and 2.0 mm wide.
). The boring is oval with a somewhat irregular outline, and is moderately deep in the column wall. It has a rounded edge and shows partial repair. The columnal is strongly swollen around the pit. The pit is 4.7 mm long and 2.0 mm wide.

Click on thumbnail |
Figure 2: A crinoid pluricolumnal with single Tremichnus from the Rhuddanian (Hilliste Formation) of Hiiumaa Island (Hilliste quarry) (TUG 1692-1). |
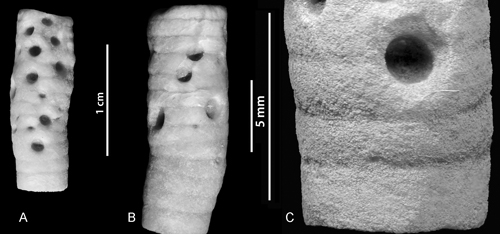
One crinoid pluricolumnal (Fig. 3A-C ![]() ) with multiple (n=14) Tremichnus was found in the Jaagarahu Formation (Sheinwoodian) of Saaremaa Island. The borings are smaller and larger conical pits that penetrate the column wall around the circumference of the stem. These pits have circular outlines, rounded edges and straight-sided walls. The crinoid pluricolumnal appears to be slightly swollen at the pits. The diameter of the outer edge is 0.2 to 1.9 mm (n=14, mean 0.9 mm, sd=0.47).
) with multiple (n=14) Tremichnus was found in the Jaagarahu Formation (Sheinwoodian) of Saaremaa Island. The borings are smaller and larger conical pits that penetrate the column wall around the circumference of the stem. These pits have circular outlines, rounded edges and straight-sided walls. The crinoid pluricolumnal appears to be slightly swollen at the pits. The diameter of the outer edge is 0.2 to 1.9 mm (n=14, mean 0.9 mm, sd=0.47).
Click on thumbnail to enlarge the image.
Figure 3: A-C. A crinoid pluricolumnal with multiple Tremichnus from the Sheinwoodian (Jaagarahu Formation) of Saaremaa Island (Sepise) (GIT 405-165).
The syn vivo nature of the pit is indicated by the strongly swollen column. In addition, the edges of the pit show partial repair. This is the earliest record of a symbiont in the columnals of crinoids from Baltica. The shape of the pit defies simple characterization. It differs from typical Tremichnus by its slightly irregular outline, but this may be due to partial repair. Overall, the structure is subelliptical. The top is broadly rounded, but the lowest portion tapers to a point until the final portion which is expanded into small bulb. The Rhuddanian specimen is a typical embedment structure formed by an organism that inhibited growth of the crinoid host stereom. There is evidence of the exterior columnal surface in the floor of the pit. This suggests that an epizoan settled on the exterior of the column but did not bore in but occupied space as the crinoid secreted extra stereom around it (, personal comm.). The most likely motive for settling on a crinoid columnal was to create a domicile. The exact nature of this crinoid-symbiont association is uncertain, but occupying a portion of the columnal wall may have had a negative effect on the host. The swollen pluricolumnal also indicates a negative effect of the symbiont on its host. It is possible that the symbiont may have consumed nutrients from the host crinoid tissues. Alternatively, it may have simply benefited from a higher tier for feeding if it was a suspension feeder. This trace resembles somewhat the barrel-shaped inflations of crinoid stems with one entrance hole described by (1974). They also have edges around the pit often bent slightly inward, indicating that crinoid tried to close the opening. (1974) suggested that metabolic waste products from the parasite might be responsible for the malformation of the surrounding columnals. She also argued that in some cases the contact with alien tissue might have been the stimulus for a histological response by the crinoid.
The pits in the Sheinwoodian pluricolumnal are slightly different from typical Tremichnus because they are more conical than bowl-shaped. The Sheinwoodian pluricolumnal is somewhat swollen at the pits. If this is not an artefact of preservation, it may indicate reaction of the crinoid. In this case the pits are similar to the parasitic pits known from crinoids since the Ordovician (, 1985; et al., 2014). The syn vivo origin of the pits is also supported by their distribution around the column. It is difficult to estimate the exact nature of this crinoid-symbiont relationship. Numerous deep pits in the column may have had negative effects on the host. The symbionts may have benefitted from a higher tier if they were suspension feeders. Alternatively, or in addition, they may have consumed nutrients from the crinoid. The symbionts also likely benefitted from a domicile and additional protection by the crinoid skeleton against the predators. Similar symbiotic pits (Type 1), possibly created by parasites, have been described from crinoid columns of the Sheinwoodian of Gotland (, 1974). These pits (type 1 in the Gotland crinoids) are also located all around the stem and contain both smaller and larger pits. The crinoid collection studied here was comprised of numerous specimens from the Sheinwoodian of Saaremaa, but no other symbiotic pits were discovered. This kind of symbiotic pit may have not been common everywhere in the Silurian of Baltica. The producers of Tremichnus were potentially host-specific, as demonstrated by their decidedly non-random distribution among crinoid taxa in the Silurian of New York (, 1978). The absence of pits in most stems may simply reflect the fact that these were not the required hosts, which were rare in this setting.
Symbiosis was common in Paleozoic crinoids. There is a rich record of various invertebrates living symbiotically with crinoids, mostly various symbiotic boring organisms that lived within the crinoid columns. Symbionts in crinoid columnals are often common (, 1974; , 1974; , 1976; , 1978, 1985; & , 1983; , 1990; , 1992; et al., 2014). In addition to symbiotic boring organisms, there were also cornulitid worms, microconchids, tabulate corals, bryozoans and rhynchonellid brachiopods (, 1974; & , 1998) that grew attached to the stems of living crinoids. Crinoids could also attach to an erect column of other crinoids (, 1992). The Silurian of Estonia is well studied and rich in shelly fossils ( & , 1997). However, crinoid symbionts have not been reported previously. This probably represents a study bias as the earlier literature is mostly devoted to the stratigraphic distribution of fossils and less to paleoecology.
Crinoid symbionts have been previously described from Baltica only by (1974), and it is clear that they did not form an important part of symbiotic interactions in this region at this time. Most records of symbiosis involve colonial organisms with massive skeletons, such as stromatoporoids and tabulate corals (, 1987; & , 2014a, 2014b). Skeletal volume probably played an important role in symbiont host selection, although why this was is unclear. Crinoids, with their relatively low-volume skeletons, were less attractive hosts than the large colonial animals.
Tremichnus is an embedment structure formed by an organism that inhibited growth of the crinoid host stereom. This is particularly likely in the case of the Rhuddanian specimen in which there is evidence for exterior surfaces of columnals in the floor of the pit. This suggests that an epizoan settled on the exterior of the column, did not bore into it but occupied space while the crinoid secreted extra stereom around it.
Tremichnus is rare in the crinoid columns from the Silurian of Estonia. This is not a study bias and can presumably be explained by the small number of suitable host species among the crinoid fauna. The rarity of crinoid-Tremichnus interactions here contrasts with other examples of comparable age elsewhere.
Financial support to O.V. was provided by the Palaeontological Association Research Grant, Paleontological Society Grant, Estonian Research Council projects ETF9064 and IUT20-34. This paper is a contribution to IGCP 591 "The Early to Middle Palaeozoic Revolution". W.A. and M.W. were supported by the National Geographic Society (NGS 60031112) and the Fund at The College of Wooster. We are grateful to G. , Institute of Geology, Tallinn University of Technology, for photographing the specimens. We are also grateful to C.E. and B. for the constructive reviews.
W.I., M.A. & O. (2012).- Crinoids from the Silurian of western Estonia.- Acta Palaeontologica Polonica, Warsaw, vol. 57, p. 613-631.
W.I., M.A. & O. (2015).- Wenlock and Pridoli (Silurian) crinoids from Saaremaa, western Estonia (Phylum Echinodermata).- Journal of Paleontology, Lawrence, vol. 89, p. 72-81.
T.K. (1990).- Non-predatory drilling of Mississippian crinoids by platyceratid gastropods.- Palaeontology, Oxford, vol. 33, p. 743-748.
C.E. (1978).- Host-specific pit-forming epizoans on Silurian crinoids.- Lethaia, Hoboken, vol. 11, p. 217-232.
C.E. (1985).- Tremichnus: a new ichnogenus of circular-parabolic pits in fossil echinoderms.- Journal of Paleontology, Lawrence, vol. 59, p. 625-635.
L.R.M. & T.H. (2005).- Baltica from the late Precambrian to mid Palaeozoic: the gain and loss of a terranes's identity.- Earth Science Reviews, vol. 72, p. 39-66.
C. (1974).- Epizoans on Silurian-Devonian crinoids.- Lethaia, Hoboken, vol. 7, p. 287-301.
T.E. (1992).- Paleoecology of hardground encrusting and commensal crinoids, Middle Ordovician, Tennessee.- Journal of Paleontology, Lawrence, vol. 66, p. 129-147.
S. (1987).- Stromatoporoid-coral intergrowths in a Silurian biostrome.- Lethaia, Hoboken, vol. 20, p. 371-380.
M.J., R.A. & P.M. (2004).- The Silurian Period. In: F.M., J.G. & A.G. (eds.), A Geologic Time Scale 2004.- Cambridge University Press, p. 188-201.
D.L. & W.I. (1983).- Biotic interactions among Recent and among fossil crinoids. In: M.J.S. & P.L. (eds.), Biotic interactions in Recent and fossil benthic communities.- Plenum, New York, p. 377-427.
H. & R. (1977).- Model of facies and sedimentology for Paleobaltic epicontinental basin. In: D.L. (ed.), Facies and fauna Silurian of Baltica.- Institute of Geology AN ESSR, Tallinn, 89-121 [In Russian].
T.J. & M.A. (1988).- Parasitism of Ordovician bryozoans and the origin of pseudoborings.- Palaeontology, Oxford, vol. 31, p. 939-949.
S.E. & K.B. (1998).- Secondary tiering on crinoids from the Waldron Shale (Silurian: Wenlockian) of Indiana.- Journal of Paleontology, Lawrence, vol. 72, p. 887-894.
A. & A. (1997).- Geology and Mineral Resources of Estonia.- Estonian Academy Publishers, Tallinn, 436 pp.
L. (2005).- Palaeoecology and diversity of endosymbionts in Palaeozoic marine invertebrates: trace fossil evidence.- Lethaia, Hoboken, vol. 38, p. 89-99.
J.R., T.J. & C.E. (2014).- A swollen crinoid pluricolumnal from the Upper Ordovician of northern Kentucky, USA: the oldest record of an amorphous paleopathologic response in Crinoidea?- Estonian Journal of Earth Sciences, Tallinn, vol. 63, p. 317-322.
O. (2004).- The earliest known Trypanites borings in the shells of articulate brachiopods from the Arenig (Ordovician) of Baltica.- Proceedings of the Estonian Academy of Sciences, Geology, Tallinn, vol. 53, p. 257-266.
O. & M.-A. (2014a).- Endobiotic rugosan symbionts in stromatoporoids from the Sheinwoodian (Silurian) of Baltica.- PLoS ONE, San Francisco, vol. 9, nº 2, e90197, 7 p.
O. & M.-A. (2014b).- Symbiotic worms in biostromal stromatoporoids from the Ludfordian (Late Silurian) of Saaremaa, Estonia.- GFF, Stockholm, vol. 136, p. 503-506.
O., M.A. & M.-A. (2014).- Symbiotic endobiont biofacies in the Silurian of Baltica.- Palæogeography, Palæoclimatology, Palæoecology, vol. 404, p. 24-29.
J.M. (1974).- Presumed myzostomid infestation of an Ordovician crinoid.- Journal of Paleontology, Lawrence, vol. 48, p. 506-513.
J.R. (1976).- Phosphannulus on Paleozoic crinoid stems.- Journal of Paleontology, Lawrence, vol. 50, p. 218-225.
M., A., M., D., J.K., J.W.M., C. & K.S.S. (2015).- In defence of an iconic ichnogenus - Oichnus , 1981.- Annales Societatis Geologorum Poloniae, Warsaw, vol. 85, nº 3, p. 445-451.
M.A., E.A. & W.I. (2014).- Parasitism of a new apiocrinitid crinoid species from the Middle Jurassic (Callovian) of southern Israel.- Journal of Paleontology, Lawrence, vol. 88, p. 1212-1221.
J.-P. & M.K. (2014).- Sedilichnus, Oichnus, and Tremichnus: 'small round holes in shells' revisited.- Journal of Paleontology, Lawrence, vol. 88, p. 895-905.