◄ Carnets Geol. 15 (18) ►
![]()
Contents
[1. Introduction]
[2. Geographical and Geological setting]
[3. Methodology]
[4. Stratigraphy and microfacies] [5. Systematic micropaleontology]
[6. Comments on the associated porcellaneous and lamellar-perforate foraminifera]
[7. Conclusions]
and ... [Bibliographic references]
Departament de Geologia (Paleontologia), Universitat Autònoma de Barcelona, E-08193 Cerdanyola del Vallès (Bellaterra) (Spain)
Repsol Campus, Méndez Álvaro 44, 28045 Madrid (Spain)
Departament de Geologia (Paleontologia), Universitat Autònoma de Barcelona, E-08193 Cerdanyola del Vallès (Bellaterra) (Spain)
Published online in final form (pdf) on November 18, 2015
[Editor: Bruno ; technical editor: Christian ;
language editor: John A. ]
![]()
The Font de les Bagasses marls and marly limestones (Montsec Mountains, NE Spain) host a rich assemblage of larger foraminifera, that was described for the first time in the early 20th Century from the currently submerged locality of Tragó de Noguera (Marginal Mountains). In this paper, a detailed architectural study has been carried out on selected, agglutinated larger foraminifera, which include Choffatella aff. rugoretis , Hemicyclammina chalmasi (), Cuneolina conica , Cuneolina cylindrica , and Dictyopsella kiliani . Moreover, it presents their biostratigraphic range and the range of the associated porcellaneous and lamellar-perforate foraminifera in the study area. The assemblage is Early Campanian in age.
Larger foraminifera; systematics; biostratigraphy; Upper Cretaceous; Pyrenees; NE Spain.
S., C. & E. (2015).- Selected agglutinated larger foraminifera from the Font de les Bagasses unit (Lower Campanian, southern Pyrenees).- Carnets Geol., Madrid, vol. 15, nº 18, p. 245-267.
Quelques grands foraminifères agglutinés de l'unité de la Font de les Bagasses (Campanien inférieur, Sud Pyrénées).- Les marnes et marno-calcaires de la Font de les Bagasses (Serra del Montsec, Nord-Est de l'Espagne) présentent une riche association de grands foraminifères décrite pour la première fois au début du 20e siècle dans la localité, aujourd'hui submergée, de Tragó de Noguera (Serres Marginals). Nous présentons ici une étude détaillée de l'architecture des grands foraminifères agglutinés : Choffatella aff. rugoretis , Hemicyclammina chalmasi (), Cuneolina conica , Cuneolina cylindrica et Dictyopsella kiliani . Ce travail est complété par les répartitions stratigraphiques de ces espèces et par celles d'autres grands foramininifères, porcelanés et lamellaires-perforés, qui leur sont associés dans le secteur étudié. L'association est attribuée au Campanien inférieur.
Grands foraminifères ; systématique ; biostratigraphie ; Crétacé supérieur ; Pyrénées ; NE Espagne.
The larger foraminifera of the southern Pyrenees and particularly those attributed to the Late Cretaceous have been studied since the end of the 19th Century. The pioneer was the Spanish engineer Luis Mariano who, during the works for the first geological map of Lleida province (NE of Spain), collected abundant material containing larger foraminifera. This material was sent to the French naturalist M.C. who, in 1899,
described various genera and species (e.g., Dictyopsella
chalmasi, Dictyopsella kiliani, Vidalina hispanica, Nummofallotia cretacea,
and Lacazina elongata) from the locality of Tragó de Noguera (Marginal Mountains, Lleida, Fig. 1 ![]() ). After 's work, the so-called "Tragó de Noguera fauna" became a reference for the study of the "Upper Santonian" larger foraminifera until the 1960s, when the locality was submerged under the water of the Santa Anna reservoir. However, "Tragó de Noguera fauna" is well represented in the Font de les Bagasses Unit in the nearby Montsec Mountains and since that time studies have continued there (,
1966; , 1966; , 1983; et al., 2009; & , 2009; et al., 2014, among others). However, a detailed inventory on the larger foraminifer fauna has not been carried out yet. Some groups such as lacazinids ( et al., 1989), meandropsinids ( & , 2009) or orbitoidids ( et al., 2014) have received the attention of researchers, while others lack detailed studies and an accurate biostratigraphic distribution.
). After 's work, the so-called "Tragó de Noguera fauna" became a reference for the study of the "Upper Santonian" larger foraminifera until the 1960s, when the locality was submerged under the water of the Santa Anna reservoir. However, "Tragó de Noguera fauna" is well represented in the Font de les Bagasses Unit in the nearby Montsec Mountains and since that time studies have continued there (,
1966; , 1966; , 1983; et al., 2009; & , 2009; et al., 2014, among others). However, a detailed inventory on the larger foraminifer fauna has not been carried out yet. Some groups such as lacazinids ( et al., 1989), meandropsinids ( & , 2009) or orbitoidids ( et al., 2014) have received the attention of researchers, while others lack detailed studies and an accurate biostratigraphic distribution.
Click on thumbnail to enlarge the image.
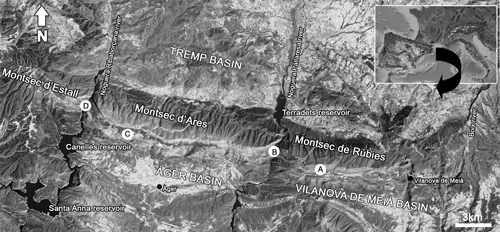
Figure 1: Location of the study area (about 75 km N of Lleida, Catalonia). Note the position of the Santa Anna reservoir with the submerged old locality of Tragó de Noguera. The capital letters refer to the stratigraphic sections studied in this paper: A. Font de la Plata section; B. Terradets section; C. Clot d'Olsí section, D. Montrebei section.
Therefore, the aim of this paper is to contribute to the knowledge of foraminifera from the Font de les Bagasses Unit by presenting a detailed architectural analysis of the main agglutinated larger foraminifera and their stratigraphic distribution. A tentative range chart including agglutinated forms, together with the main larger porcellaneous and lamellar-perforate foraminifera, is also given as an useful tool for future geological studies.
The Montsec Mountains extend continuously E-W for more than 50 kilometers in the Southern Pyrenees, between the Tremp and Àger-Vilanova de Meià basins (Lleida, NE of Spain). The Boix, Noguera Pallaresa and Noguera Ribagorçana rivers, which run N-S,
individualize three adjacent geographic units, from East to West: Montsec de Rúbies, Montsec d'Ares, and Montsec d'Estall (Fig. 1 ![]() ).
).
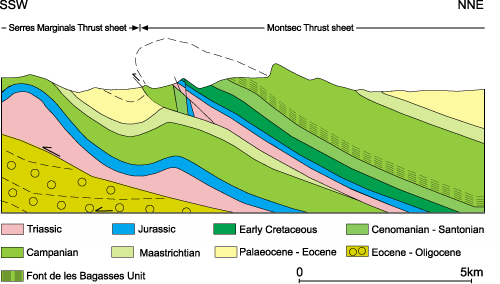
From a geological point of view, the Montsec Mountains belong to the Montsec Thrust Sheet, a Southern Pyrenean
upper thrust sheet detached from its basement over Upper Triassic evaporites. At the South, it shows a simple anticline structure representing the front of the Montsec thrust, while northwards it develops a broad syncline supporting a piggyback basin filled with Palaeogene rocks, the Tremp-Graus Basin (, 1992; & , 2000). The Montsec Mountains include a complete Mesozoic sequence, mainly
Upper Cretaceous, of about 1500 m in thickness (Fig. 2 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 2: Simplified geological cross-section of the Montsec Thrust Sheet along the Montrebei pass (Noguera Ribagorçana River). Modified from & (2000).
The deposits attributed to the Font de les Bagasses Unit ("Font de les Bagasses marls and clays" in , 1977) uniformly extend E-W along the southern slope of the mountain, dipping 35º-40º to the North. They rest unconformably on the deposits of the La Cova Unit (uppermost Lower Coniacian to Middle Santonian according to et al., 2011), passing gradually upwards to the deposits of the Terradets Unit (, 1977; , 1986), with a base dated latest Early to Middle Campanian (, 2009).
Palaeogeographically, the Upper Cretaceous deposits of the Montsec Mountains were deposited in the southern margin of the Pyrenean Basin that extended approximately E-W and opened to the Atlantic Ocean (Bay of Biscay). The basin was locaced at about 30ºN latitude ( et al., 1985; et al., 1999). The climate during the Santonian-Campanian was warm ( et al., 2003) and the global sea-level curve indicates a level 50 m higher than the present day for the Early Campanian ( et al., 2005).
The age "Late Santonian" traditionally attributed to the Font de les Bagasses deposits (, 1966; , 1966; , 1977; & , 1983; & , 1989a, 1989b; et al., 1999; among others) has been recently contested ( et al., 2014). New data published in this last paper, obtained by means of isotope strontium stratigraphy, indicate a younger age for the whole unit. Thus, the age attributed to the larger foraminifera from the Font de les Bagasses Unit is Early Campanian, corresponding to the ammonite zones of Placenticeras tridorsatum and Menabites delawarensis, and to the planktonic foraminiferal zone of Globotruncanita elevata (see the correspondence in the global time scale of et al., 2012).
The work is based on field observations and 137 samples collected from four stratigraphic sections on the well-exposed slopes of the Montsec de Rúbies and Montsec d'Ares. From East to West, the sections are (Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() ):
):
A) Font de la Plata section measured in the Font de la Plata gully about 2 km east of the Rúbies abandoned village; coordinates: UTM ETRS89 31T X: 328852.3 and Y: 4654008.6. Samples from this section are labelled SG-7 to SG-43.
B) Terradets section measured along the Noguera-Pallaresa River in the well-exposed trench of the C-13 road (a field picture of this section is shown in et al., 2014, Fig. 4A); coordinates: UTM ETRS89 31T X: 324903.0 and Y: 4655864.0. Samples from this section are named MARG-122 to MARG-177.
C) Clot d'Olsí section measured along the Pardina gully near the Ager-Alçamora road; coordinates: UTM ETRS89 31T X: 311246.8 and Y: 4657201.0. Samples from this section are labelled CO-28 to CO-68.
D) Montrebei section measured along the left margin of the Noguera-Ribagorçana River in the Montrebei pass; coordinates: UTM ETRS89 31T X: 307891.0 and Y: 4660320.0. Samples from this section are named ME-23 to ME-40.
Ten samples are from washed sediments while 127 are from consolidated rocks. The samples from unconsolidated sediments were washed over two sieves with holes of 0.35 and 0.21 mm respectively, and larger foraminifera were picked, photographed and sectioned. The external features of the foraminifera shells were photographed using optical and scanning electron microscopes. Oriented sections were prepared following the classical techniques for studying internal characteristics in agglutinated foraminifera (see , 1967). Larger foraminifera pictures are at standard magnifications of x25 and x50. More than 350 thin-sections were obtained from hard samples to study the larger foraminifera structures in random sections and the microfacies analysis. All the larger foraminifera presented in this paper were identified at genus and species levels. The genera were defined by the presence or absence of structural elements combined with chamber arrangement, while the species are defined by quantitative morphological characteristics.
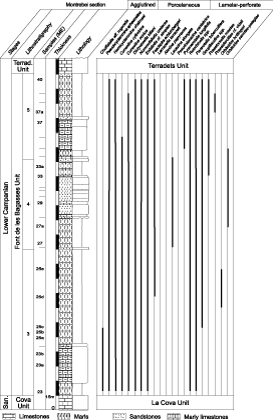
Figure 3: La Font de la Plata stratigraphic section with the larger foraminifera distribution. |
Figure 4: Terradets stratigraphic section with the larger foraminifera distribution. |
|
Figure 5: Clot d'Olsí stratigraphic section with the larger foraminifera distribution. |
Figure 6: Montrebei stratigraphic section with the larger foraminifera distribution. |
All the material studied in this work was deposited in the collection of Paleontology of the Department of Geology (Universitat Autònoma de Barcelona). The figured specimens are labeled as PUAB 82410, 82412, and 84458-84473.
Based on field work, palaeontological and sedimentological studies, the Font de les Bagasses Unit is divided into five stratigraphic subunits that are, from bottom to top (Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() ):
):
4. a. Subunit 1
This basal subunit, which overlies La Cova Unit by means of an erosive contact (et al., 1999), is composed of yellow sands, clays and coarsely grained sandstones with some intercalated microconglomerate
beds; it is interpreted to be deposited in a fluvial setting. This subunit is well represented in the Montsec de Rúbies, with a maximum thickness of 20m in the Font de la Plata section, but disappears to the West of the Terradets section (Figs. 3 ![]() -
4
-
4 ![]() ). In the uppermost part of the subunit, some sandy limestones with fragmented marine bivalves indicate the transition from fluvial to littoral marine deposits (, 1997). This subunit lacks larger foraminifera.
). In the uppermost part of the subunit, some sandy limestones with fragmented marine bivalves indicate the transition from fluvial to littoral marine deposits (, 1997). This subunit lacks larger foraminifera.
4.b. Subunit 2
The subunit 2 (called "Boix River platform" in et al., 1999) overlies the previous subunit in the sections of Font de la Plata and Terradets, disappearing to the West. It is composed of well-bedded nodular limestones with
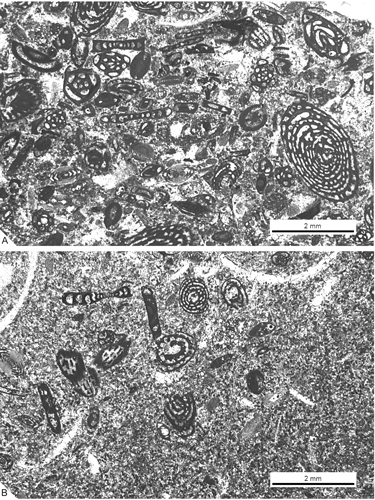
pelletoidal-packstone texture containing a diverse fauna of gastropods, bivalves (including rudists), echinoids, larger foraminifera and some sponge spicules (Fig. 7 A-B ![]() ). Dasycladacean algae occurring in this subunit are frequently recrystallized. The unit reaches a maximum thickness of 10 m in the Font de la Plata section and disappears west of the Terradets section (Pla d'Escumó area). Among the foraminifera, the
porcellaneous Pseudolacazina cantabrica et al. and Alexina papyracea & are the main components of the facies associated with the agglutinated foraminifera (Figs. 3
). Dasycladacean algae occurring in this subunit are frequently recrystallized. The unit reaches a maximum thickness of 10 m in the Font de la Plata section and disappears west of the Terradets section (Pla d'Escumó area). Among the foraminifera, the
porcellaneous Pseudolacazina cantabrica et al. and Alexina papyracea & are the main components of the facies associated with the agglutinated foraminifera (Figs. 3 ![]() -
4
-
4 ![]() ).
).
Click on thumbnail to enlarge the image.
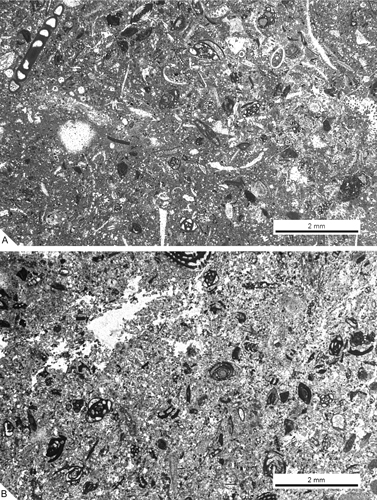
Figure 7: Microfacies of the subunit 2. A. Pelletoidal packstone with porcellaneous larger foraminifera as major components; from SG-8. B. Peletoidal packstone-wackestone with agglutinated and porcellaneous foraminifera; from SG-10.
4.c. Subunit 3
In the Font de Plata and Terradets sections, the subunit 3 overlies the subunit 2 and in the Clot d'Olsí and Montrebei sections the subunit 3 overlies directly the deposits of the La Cova Unit (Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() ). It is composed of gray and yellow argillaceous marls intercalated with nodular marly limestone with pelletoidal-packstone texture, and more rarely wackestones (Fig. 8 A-B
). It is composed of gray and yellow argillaceous marls intercalated with nodular marly limestone with pelletoidal-packstone texture, and more rarely wackestones (Fig. 8 A-B ![]() ). Fine quartz grains are rare. In the Clot d'Olsí and Montrebei sections the subunit shows intraformational slumps. The thickness varies from 135 to 210m. The faunal content is abundant and diverse: foraminifera, gastropods, bivalves (oysters, rudists and indeterminate bivalves), corals, brachiopods, and echinoid fragments are the main components (, 2000). Gastropods, rudists and other indeterminate bivalves are frequently recrystallized. Sporadically, green algae are also present in some levels, with fragments of crustose rhodophyte algae occurring in the uppermost part of the subunit. Larger foraminifera are abundant and highly diverse (see their distribution in Figs. 3
). Fine quartz grains are rare. In the Clot d'Olsí and Montrebei sections the subunit shows intraformational slumps. The thickness varies from 135 to 210m. The faunal content is abundant and diverse: foraminifera, gastropods, bivalves (oysters, rudists and indeterminate bivalves), corals, brachiopods, and echinoid fragments are the main components (, 2000). Gastropods, rudists and other indeterminate bivalves are frequently recrystallized. Sporadically, green algae are also present in some levels, with fragments of crustose rhodophyte algae occurring in the uppermost part of the subunit. Larger foraminifera are abundant and highly diverse (see their distribution in Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() ).
).
Click on thumbnail to enlarge the image.
Figure 8: Microfacies of the subunit 3. A. Wackestone-packstone with P. sphaeroidea, porcellaneous foraminifera, spicule sponges and fragmented dasycladacean algae (close to the scale bar: an oblique transverse section of Trinocladus tripolitanus ); from samples M-125. B. Wackestone with agglutinated foraminifera, small benthic and dasycladacean algae; from M-145.
4.d. Subunit 4
The boundary between this subunit and the underlying subunit 3 is marked by the presence of coarse-grained quartz in the Font de les Bagasses deposits. However, the coarse terrigenous deposits are not uniformly distributed in the unit: in the Font de la Plata and Montrebei sections, discontinuous lenticular-shaped calcareous sandstones are interbedded within argillaceous marls and marly limestones in intervals of 50-to-100m (Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() ), while in the Terradets and Clot d'Olsí sections the terrigenous intervals do not exceed a few meters (Figs. 4
), while in the Terradets and Clot d'Olsí sections the terrigenous intervals do not exceed a few meters (Figs. 4 ![]() - 5
- 5 ![]() ). The fauna of invertebrates and larger foraminifera appear frequently fragmented (Fig. 9 A-B
). The fauna of invertebrates and larger foraminifera appear frequently fragmented (Fig. 9 A-B ![]() ).
).

Click on thumbnail to enlarge the image.
Figure 9: Microfacies of the subunit 4. A. Sandstone with isometric irregular quartz grains, elongate bivalve fragments, some rotaliids and agglutinated foraminifera; from SG-40. B. Packstone with irregular quartz grains, fragmented bryozoans, echinoids, bivalves, worms and lamellar-perforate foraminifera; from Me-29.
4.e. Subunit 5
This uppermost subunit is composed of grey or yellow marls and marly limestones alternating with nodular and, more rarely, bioclastic limestones. The microfacies study shows wackestone, packstone and grainstone textures (Fig. 10 A-B ![]() ). Rarely, coarse-grained idiomorphic quartz grains are present. Bryozoans and sponge spicules are the dominant components of the marly limestones associated mainly with large specimens of Larrazetia larrazetti . The nodular limestones contain brachiopods, rudists and other bivalves, and echinoderms; the larger foraminiferal fauna is dominated by Lacazina elongata . In constrast, the bioclastic limestones include mainly Praestorsella roestae (), Praesiderolites praevidali (), and Pararotalia tuberculifera (). The limestones of the Terradets Unit conformably overlie this subunit.
). Rarely, coarse-grained idiomorphic quartz grains are present. Bryozoans and sponge spicules are the dominant components of the marly limestones associated mainly with large specimens of Larrazetia larrazetti . The nodular limestones contain brachiopods, rudists and other bivalves, and echinoderms; the larger foraminiferal fauna is dominated by Lacazina elongata . In constrast, the bioclastic limestones include mainly Praestorsella roestae (), Praesiderolites praevidali (), and Pararotalia tuberculifera (). The limestones of the Terradets Unit conformably overlie this subunit.

Click on thumbnail to enlarge the image.
Figure 10: Microfacies of the subunit 5. A. peletoidal wackestone with sponge spicules, fragmented bryozoans and lamellar-perforate foraminifera; from M-170. B. Packstone-grainstone with crustose rhodophyte algae, bryozoans and echinoid fragments, and small foraminifera; from M-167.
In the taxonomic classification of larger foraminifera, it is commonly accepted that genera are identified by the presence or absence of shell structures like exoskeleton, endoskeleton and supplemental skeleton combined with the chamber arrangement. Species are related mainly to the adult size and the size of chamber elements (, 1978; , 1981), although a great number of oriented and random sections have been required for correctly interpreting the shell structures and comparing the biometric data of the shells at homologous growth stages. The ascription of genera to taxonomically higher-levels is controversial due to the lack of a comprehensive systematic classification based on phylogenetic characteristics. As a result, the successive classifications of & (1964; 1988; 1992; 1994) have been mainly used in foraminiferal works, frequently without discussion.
In recent years, the integration of molecular data in foraminiferal studies has allowed the highest-levels foraminiferal classification (see for instance, et al., 2013, and the papers cited therein). For categories below the class, multiple classifications are proposed, but only a few of these are clearly presented, as the hierarchy of taxonomic criteria used for such classifications remains obscure and is often inconsistent. For the agglutinated foraminifera, (2014) updated a new classification partially based on the and (1988; 1992; 1994) and (2004) classifications. However, some criteria proposed by (2014) for grouping larger, complex foraminifera are only partially defined. Thus, in this paper the phylum and class categories proposed by et al. (2013) are accepted, but 's (2014) classification is only partially followed, and some remarks or new proposals are added to this classification (see below). Finally, the hierarchy of taxonomic criteria sketched by (1978, Table 1) has been taken into account in the elaboration of our proposals. In order to facilitate the comparison between the architectural characteristics of the genera of agglutinated foraminifera described in this paper, the architectural and structural terms are those referred to by (2006).
Phylum Foraminifera (, 1826) et al., 2013
Class Globothalamea et al., 2013
Order Loftusiida & in , 2004
Remarks: Following the authors of the Order Loftusiida, it includes all agglutinated foraminifera with coiled arrangement of chambers in early stage, that later may be uncoiled, and with complex exoskeleton. The simple forms with the same chamber arrangement are grouped in the Order Lituolida , 1885. Therefore, in the future, further studies are needed to explore the relationship between taxa included in both orders.
Suborder Loftusiina & in , 2004
Remarks: (2014) refers to the Suborder Loftusiina as "test free or attached, multilocular, coiling or uncoiling; wall agglutinated, microgranular, with imperforate epidermis (outer layer) and an inner alveolar layer; aperture terminal in the initial stage, later multiple, rows of rounded areal openings". Therefore, with reference to the order description, the type and distribution of the apertures are added.
Observations: In (2014) the term "epidermis" is used in a wider meaning than in its original description (, 1948), which restricted the term epidermis for walls with polygonal subepidermal network (, 1967). The inner alveolar layer of (2014) refers to the "alveolar network" of (1906) comprising the alveolar and subepidermal exoskeleton types of (1967). The former consists of a tapestry of one or several generations of rounded outline alveoli, with no differentiation between beams and rafters. The latter has differentiated beams and rafters forming a regular polygonal subepidermal network similar to the pigeonholes. The thin epidermis is restricted to this last group of exoskeleton type.
Superfamily Loftusoidea , 1884
Remarks: Foraminifera with planispiral (or streptospiral) arrangement of chambers that may become uncoiled, peneropliform or cyclical in the adult stage, with exoskeleton structures and one or several apertures.
(2014) included the following four families in this superfamily: the Jurassic Mesoendothyridae with a pseudokeriothecal exoskeleton (see , 1988, for details); the Late Jurassic to recent Cyclamminidae with an alveolar exoskeleton (sensu , 1967); and the Jurassic-Cretaceous Spirocyclinidae with polygonal subepidermal exoskeleton. The fourth Family Loftusiidae has the same structural characteristics as Spirocyclinidae, but the axis of coiling is elongated. The Family Haurinidae , 1988, with similar disposition of chambers and exoskeleton structures, has been ascribed to the Suborder Orbitolina by (2004, 2014) (see the discussion below).
Family Spirocyclinidae (, 1887) , 1950
Remarks: The Family Spirocyclinidae was constructed by (1887) to include four agglutinated genera of larger foraminifera (Spirocyclina, Dicyclina, Cuneolina and Orbitolina), having in common exoskeleton elements forming a polygonal subepidermal network, but with a different chamber arrangement.
(1950) emended the Spirocyclinidae, resulting in the following description: planispiral-to-peneropliform or uncoiled agglutinated shells with one or multiple apertures. Therefore, the biserial genera Dicyclina and Cuneolina and the uniserial Orbitolina were removed from Spirocyclinidae.
All representatives of the Spirocyclinidae have an exoskeleton constituted by beams and rafters forming a polygonal subepidermal network covered by a fine epidermis. According to (1950), both taxa, with or without exoskeleton elements, were included in Spirocyclinidae. But & (1988), not in accordance with (1950), restricted the Family Spirocyclinidae to the genera with endoskeleton elements, while most of the genera lacking such elements were placed in the Family Cyclamminidae. The same classification was also followed by (2014). However, Cyclammina (type-genus of the Family Cyclamminidae) possesses alveolar exoskeleton and not a regular polygonal subepidermal network as in Spirocyclinidae. Therefore, the two families should remain separate, and the genera with polygonal subepidermal patterns should be ascribed to the Family Spirocyclinidae instead of the Cyclamminidae. Following (1978), the presence or absence of endoskeleton elements is considered here of lower category than the exoskeleton elements and the communication system of apertures.
Subfamily Choffatellinae , 1958
Remarks: In this work, the Subfamily Choffatellinae, included by (2014) in the Family Cyclamminidae, is removed from this last family and transferred back to Spirocyclinidae. Accordingly to (1958), it is characterized by planispiral to uncoiled or peneropliform-to-cyclical shells with polygonal subepidermal exoskeleton, multiple aligned apertures, and lacking endoskeletal elements.
Choffatella , 1905
(Type-species: C. decipiens , 1905)
Description: Agglutinated, lens-compressed shell with multiple openings placed in one row in the median plane of the apertural face. The chambers are planispiral-evolute arranged. Choffatella has exoskeleton elements (beams and rafters) forming a well-developed polygonal subepidermal network. It lacks endoskeleton, although & (1988) talked about an "endoskeleton formed by thick massive septa". The septa are pierced by multiple foramina aligned from one septum to the next.
Age: After & (1988), the genus Choffatella ranges from the Late Jurassic to the Late Cretaceous (Cenomanian). (1968) and (1980) enlarged its stratigraphic range to the Santonian. Our specimens attributed to this genus are collected in deposits attributed to the Lower Campanian ( et al., 2014).
Choffatella aff. rugoretis , 1968
(Fig. 11 A-F ![]() )
)
Synonymies:
aff. 1968 Choffatella rugoretis , Pl. 4, figs. 6-13.
aff. 1972 Gendrotella rugoretis () ,
Figs. 5-6.
1977 Choffatella rugoretis () , Fig. 3.
1978 Choffatella rugoretis , et al., Fig. 2.
Description: Finely agglutinated, lens-shaped foraminifera with one row of openings aligned in the median plane of the apertural face. The A-generation consists of an embryo formed by a spherical protoconch and a hemispherical deuteroconch followed by planispiral-evolute chambers disposed in two or two and half whorls. The earliest chambers have a quadrangular outline, but they become rapidly low and elongate to build an operculiniform spire. The number of chambers in the last whorl varies from 24 to 27. The septa are arched and pierced by multiple, regularly distributed intercameral foramina. The average size of the adult megalospheric shells is 2.6 mm. The scarce and poorly preserved B-forms can reach 3.0 mm and up to three whorls of planispiral chambers.

Click on thumbnail to enlarge the image.
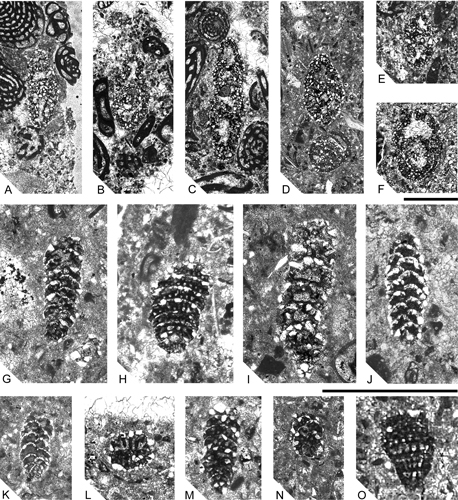
Figure 11: A-C. Choffatella aff. rugoretis: subequatorial sections (near the equatorial plane) showing the planispiral, partially evolute chamber arrangement and multiple aligned intercameral foramina; from CO-32. D-F. Choffatella aff. rugoretis: lateral view showing the elongated chambers and sutures (scanning microphotography); the epidermis is poorly preserved. from SG-36. G-I: Hemicyclammina chalmasi:G-H; Hemicyclammina chalmasi: external views showing the inflate chambers and depressed sutures (scanning microphotography); from SG-36. I. Hemicyclammina chalmasi: equatorial section of an extraordinarily well preserved specimen showing the large intercameral foramen and its particular position in the septum (hemi-septum from , 1953); from SG-36. Scale bar 1 mm.
Remarks: (1972) re-studied the 's types of C. rugoretis and he concludes that the species attributed by (1968) to the genus Choffatella lacked an exoskeleton. Thus, (1972) stated that 's taxon cannot be assigned to the genus Choffatella, a Jurassic-Early Cretaceous genus exhibiting an exoskeleton consisting of beams and rafters organized in a polygonal subepidermal network. Therefore, he erected the new genus Gendrotella for it.
However, material from the type locality (Martigues, Bouches-du-Rhône, South-East of France) has yielded specimens morphologically identical to those described by (1968) and bearing exoskeleton elements; moreover, the specimens of C. rugoretis showed by (1968, Pl. 4, figs. 9-10) exhibit short beams and rafters forming a delicate network in the inner surface of the chamber, interpreted here as a subepidermal type of exoskeleton. Thus, the species is re-installed within the genus Choffatella, and Gendrotella becomes a non-valid genus after the ICNZ.
Similarities and differences: The specimens from the Font de les Bagasses Unit are larger (almost the double size) than the size of Martigues (type-locality of C. rugoretis), and designated as Choffatella aff. rugoretis. Further studies are needed to determine if the size difference is due to evolution (Santonian in the 's specimens and Early Campanian in the Pyrenean specimens) or related to a deeper water environment in the Font de les Bagasses Unit than in the Martigues deposits.
Stratigraphical distribution: In the Font de les Bagasses, C. aff. rugoretis occurs in the four measured sections. It has been identified mainly in the washed sediments, but some random sections also occur in the consolidated rocks. See the distribution in Figs. 3 ![]() -
4
-
4 ![]() -
5
-
5 ![]() -
6
-
6 ![]() .
.
Geographical distribution: It has been only mentioned from the Pyrenean basin (North of Spain and South and South-East of France).
Subfamily Amijiellinae , 1966
Remarks: Agglutinated foraminifera with polygonal subepidermal exoskeleton. The chambers with planispiral-involute arrangement have only one aperture. It lacks endoskeletal elements.
Hemicyclammina , 1953
(Type-species: Hemicyclammina sigali , 1953)
Description: Agglutinated, lens-shaped foraminifer with rounded-to-subacute periphery. The chambers have a planispiral-involute arrangment. The aperture is unique, large, located near the base of the septum and with circular-to-elliptic outline (hemi-septum after , 1953). Hemicyclammina has a well-developed polygonal subepidermal network with two orders of beams. It lacks endoskeleton.
Age: After & (1988), the age of Hemicyclammina is Early Cretaceous and Cenomanian for this genus but, with the ascription of "Dictyopsella" chalmasi to this genus (et al., 1979), the range extends as far as the Early Campanian.
Hemicyclammina chalmasi (, 1899)
(Fig. 11 G-I ![]() )
)
Synonymies:
1899 Dictyopsella chalmasi , Pl. 8, fig. 4.
1977 Hemicyclammina chalmasi () ,
Figs. 4-5.
1978 Hemicyclammina chalmasi () et al.,
Figs. 1- 5.
cf. 1999 Hemicyclammina chalmasi () , Pl. 1, figs. 1-6.
cf. 2004 Hemicyclammina chalmasi () & Pl.1,
figs. 1-9.
cf. 2010 Hemicyclammina chalmasi () et al., Pl. 1, figs. 1-5.
Description: Coarsely agglutinated, lens-shaped shells with a more or less rounded periphery. The diameter varies from 1.3 to 1.5 mm. The slightly inflated chambers
have a planispiral-involute arrangment, with two-to-three spiral whorls. The sutures are depressed,
exhibiting a lobate outline. Two generations have been observed, interpreted as A and B generations ( et al., 1978,
Figs. 1-4), although the adult size does not present any
differences. The A-generation consists of a spherical embryo followed by two whorls of planispiral chambers, which generally number 8 or 9 in the last whorl, but some specimens can reach 11 chambers. In the B-generation, the small microsphere is followed by three whorls of spiral-involute chambers with 6-8 chambers in the last whorl. The septa are oblique. The exoskeleton is well developed with long beams and less pronounced rafters, but can only be observed in the thin-sections of the isolated specimens (Fig. 13 I ![]() ). The Hemicyclammina sections coming from hard rocks (Fig. 13 A-F
). The Hemicyclammina sections coming from hard rocks (Fig. 13 A-F ![]() ) are difficult to identify and are
designated in this work as H. cf. chalmasi. However, they coincide with the sections shown by (1999), & (2004) and et al. (2010).
) are difficult to identify and are
designated in this work as H. cf. chalmasi. However, they coincide with the sections shown by (1999), & (2004) and et al. (2010).
Similarities and differences: The specimens of the Font de les Bagasses Unit are identical to those described by (1899) from the Tragó de Noguera under the name of Dictyopsella chalmasi, that have been transferred to the genus Hemicyclammina by et al., 1978.
Hemicyclammina chalmasi differs from H. sigali (species-type of the genus) due to its larger size, the complexity of the skeleton, and the septa perpendicular to the external wall.
Stratigraphical distribution: The age of H. chalmasi is Early Campanian. In the Font de les Bagasses Unit, it is observed in all sections, although infrequently. The sections from consolidated rocks are badly preserved.
Geographical distribution: It has been mentioned in the Pyrenees and the central Europe ( & , 2004) and Balkans (, 1999; et al., 2010).
Subfamily Amijiellinae , 1988
Remarks: After (1988), the Subfamily Amijiellinae consists of planispiral to uncoiled shells with polygonal subepidermal exoskeleton, multiple cribrate apertures, and lacking endoskeletal elements. (1988) and also (2014) included the Subfamily Amijiellinae in the Family Haurinidae (, 1988), which is characterized by planispiral to peneropliform shells with a polygonal subepidermal (hypodermal in , 1980) network. However, such characteristics are shared with the Family Spirocyclinidae (see above), which has priority. However, further studies are needed to clarify the relationship between the two families: Haurinidae used mainly for the Jurassic taxa (see the excellent work of , 1980, 1988), and Spirocyclinidae for the Cretaceous taxa.
In addition, (2014) placed the Subfamily Amijiellinae (and the Family Haurinidae) in the Suborder Orbitolinina , 2004, that was defined as "trochospiral to conical, later stage may have reduced number of chambers per whorl, or may become uniserial and rectilinear". However, neither the Subfamily Amijiellinae nor the Family Haurinidae possesses such characteristics and, in the current state of knowledge, they should be removed from Orbitolinina and included in Loftusiina. They are presented in this paper this way.
Pseudocyclammina & , 1926
(Type-species: Cyclammina lituus , 1890)
Description: Agglutinated shell with subcylindrical-to-subspherical morphology with multiple cribrate apertures piercing the apertural face. The arrangement is planispiral-involute, but may become uncoiled in the late stage of growth. Pseudocyclammina presents exoskeleton composed of beams and rafters resulting in a polygonal subepidermal network. It lacks endoskeleton.
Age: After & (1988), the age of Pseudocyclammina ranges from the Early Jurassic to Coniacian, but the presence of P. sphaeroidea in the Santonian (, 1968) and in the Early Campanian (this paper) indicates a younger age for this genus.
Remarks: Although the "plexus" Pseudocyclammina (, 1980; 1988) is mentioned as having a long stratigraphic extension, the idea of a polyphyletic group is recurring in the literature (see , 1980, and the papers cited therein). (1970) placed the genus Pseudocyclammina together with Choffatella in the Choffatellinae; however, both genera differ in the foraminal pattern: aligned in Choffatella and cribrate in Pseudocyclammina.
Pseudocyclammina sphaeroidea , 1968
(Fig. 12 A-L ![]() )
)
Synonymies:
1968 Pseudocyclammina sphaeroidea, ,
Figs. 1-5.
2013 Pseudocyclammina sphaeroidea, et al.,
Fig. 7g.
Description: Small and globular Pseudocyclammina (the observed diameter varies from 0.6 mm to 1.1 mm). Dimorphism is not observed in our material from the Font de les Bagasses: all the specimens have similar size at the same growth stage. Where visible, a small and spherical protoconch is followed by a small hemispherical deuteroconch. The post-embryonic chambers exhibit a planispiral-involute arrangment, resulting in two and half to three whorls of spire, rarely three and half whorls. Specimens showing uncoiled stage are not frequent in our material. The earliest chambers are quadrangular in shape, but in the latest stage the chambers become low and elongated with peneropliform morphology. The number of chambers in the last whorl varies from 10-12 in the small forms to 13-14 in the largest ones. The external wall (epidermis) of our specimens is diagenetically altered. Moreover, the specimens incorporate angular, coarse quartz grains in the external wall and in the septa, frequently masking the exoskeleton structures. Where they are visible; these structures show short beams and rafters forming a poorly developed subepidermal network. The chamber lumen is free of endoskeletal elements. The septa are massive and pierced by small, cribrate intercameral foramina.
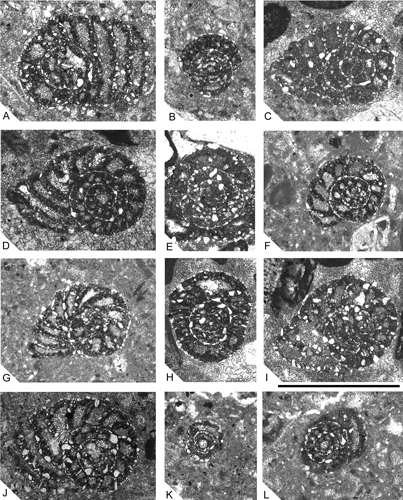
Click on thumbnail to enlarge the image.
Figure 12: Pseudocyclammina sphaeroidea. A. Oblique section showing three spiral whorls in a large specimen. B, E, H, K. Axial and subaxial sections. C-D, F-J. Equatorial and subequatorial sections. L. Centred oblique section. A, B, J from CO-30; C-E, H, I from CO-1; F, G, K, L from M-125. Scale bar 1 mm.
Similarities and differences: The Pyrenean specimens are similar to those described by (1968) from the Martigues deposits. However, the French specimens do not present grains of quartz in their shells, suggesting that the presence of this type of agglutination in our specimens has an environmental significance.
Specimens attributed to P. aff. sphaeroidea also occur in the Coniacian deposits of the Pyrenees ( et al., 2013). The specimens designed as P. sphaeroidea in the Turonian of the Tethyan deposits are smaller than the Pyrenean ones (see for instance, & , 2008; et al., 2012; et al., 2007; et al., in press).
Stratigraphical distribution: This small and globular Pseudocyclammina characterizes all deposits of the Font de les Bagasses Unit, although it is most frequent in its lower part. At certain levels it occurs as isolated specimens, but is more abundant in hard rocks. In the Pyrenees, the species is known from Coniacian to Lower Campanian. However, it is also mentioned from the Turonian ( & , 2008; et al., 2012; et al., 2007; among others).
Geographic distribution: P. sphaeroidea type is reported from the Pyrenean basin, while the small forms have a wider occurrence (see for instance: , 1992; et al., 2007).
Suborder Ataxophragmiina , 1958
Remarks: (2014) included in this suborder "agglutinated high trochospiral foraminifera with an asymmetrical spire that may reduce to a biserial or uniserial arrangement. The chambers may have exoskeletal and endoskeletal structures. The aperture forms a wide horizontal slit at the base of the apertural face". However, some genera included in the suborder have multiple apertures.
Superfamily Ataxophragmioidea , 1877
Remarks: After & (1988), this Superfamily is characterized by its early trochospiral growth that may be reduced to biserial or uniserial in later stages. The chambers have skeletal elements partially dividing the chamber lumen.
Family Dicyclinidae & , 1964, emended
Remarks: et al. (in press) considered the Family Dicyclinidae & , 1964, synonymous with Cuneolinidae , 1981. & 1964, 1988) defined the Family Dicyclinidae as "test free, discoidal, chambers cyclical, biserially added, partially subdivided by transverse or radial partitions with imperforate epidermal layer; peripheral multiple aperture". Whereas, the Family Cuneolinidae , 1981, was described as "conical to subflabelliform, trochospiral in early stage, with up to five chambers per whorl, rapidly reduced to biserial; chambers subdivided by many septula extending from the outer wall toward the junction of the two series of chambers¸ agglutinated wall with imperforate outer layer and reticulate subepidermal layer; aperture simple to multiple".
According to (1992), & (1995) and the present work, Cuneolina , 1839 (Type-species: C. pavonia , 1846) shows a large and structured deuteroconch followed by chambers biserially arranged, and not trochospirally arranged in the early stages of growth as defined by & (1964). Other characteristics, like a deep subepidermal network exoskeleton, the multiple apertures and the lack of endoskeleton are identical to Dicyclina characteristics. Thus, in accordance with et al. (in press), the genus Cuneolina needs to be placed in the Family Dicyclinidae.
Therefore, the Family Dicyclinidae is emended here to include the non-cyclical forms as follows: test free, conical to fan-shaped or discoidal, chamber biserially arranged, exoskeleton composed of beams and rafters forming a regular subepidermal network covered by a fine epidermis. Multiple apertures are placed at the base of the chamber. It lacks endoskeletal structures.
Subfamily Cuneolininae , 1981
Remarks: et al. (in press) maintained the Subfamily Cuneolininae for the conical to fan-shaped morphotypes. However, further studies are needed to prove the validity of this subfamily, comprising also small and less complex forms from the Lower Cretaceous.
Cuneolina (, 1839) & , 1995
(Type-species: Cuneolina pavonia , 1846)
Description: Conical to fan-shaped foraminifera with agglutinated wall. The chambers are biserially arranged. They are generally broad and low, and commonly compressed in the plan of the biseriality. They have a complex exoskeleton composed of beams and rafters forming a regular subepidermal polygonal network covered by a fine epidermis. The beams frequently penetrate deep into the chamber lumen leaving only a narrow passage near the apertural face. It lacks endoskeleton elements. Multiple intercameral foramina are aligned at the base of the septa. For more details see & (1995).
Age: Cuneolina is known from the Aptian to Maastrichtian ( & , 1988).
Cuneolina conica , 1850
(Fig. 13 H, O ![]() ; Fig. 14 C-D
; Fig. 14 C-D ![]() )
)
Synonymies:
1850 Cuneolina conica ,
Pl. 1, figs. 7-9; Pl. 2, fig. 1.
1899 Cuneolina conica , ,
Pl. 8, figs. 8-10
1968 Cuneolina conica , ,
Pl. 4, fig. 16
Description: Fan-shaped shell, biserially arranged, with low and broad chambers occupying an opening angle of about 70º. The number of chambers is about 20. The shell varies from 0.7mm to 1.2 mm in length. The exoskeleton consists of well-developed beams and rafters. The number of beams per chamber can reach about 20 in the last chamber.
Click on thumbnail to enlarge the image.
Figure 13: A, C. Hemicyclammina cf. chalmasi: subaxial sections; from SG-8 and SG-10. B. oblique section not far from the equatorial section; from SG-8 and SG-10. D, E. Hemicyclammina cf. chalmasi:Oblique sections; from SG-8 and SG-10. F. Hemicyclammina cf. chalmasi: slightly oblique near the subaxial section; note in all of the pictures the coarse agglutinating quartz grains masking the wall structure; from SG-8 and SG-10. G, J, K, M. Cuneolina cyclindrica: axial sections; G, from M-130; K, from M-125; J, M from M-151. I. Cuneolina cyclindrica: slightly oblique section almost subaxial; from M-149. L. Cuneolina cyclindrica: basal section showing the beams penetrating deep into the chamber lumen and leaving a narrow passage; from M-130. N. Cuneolina cyclindrica: oblique section showing fifth chambers; from M-130. H, O. Cuneolina conica: longitudinal-oblique sections; respectively from M-143 and from M-130. Scale bar 1 mm.
Similarities and differences: Our specimens from the Font de les Bagasses are identical to those described by (1899) from Tragó de Noguera.
Stratigraphical distribution: It is known only from the Upper Cretaceous.
Geographical distribution: Cuneolina conica has been mentioned in the Pyrenean deposits.
Cuneolina cylindrica , 1948
(Fig. 13 G, I-N ![]() ; Fig. 14 A-B
; Fig. 14 A-B ![]() )
)
Synonymies:
1948 Cuneolina cylindrica ,
Pl. 14. fig. 7.
1968 Cuneolina cylindrica ,
Pl. 4, fig. 14.
Description: Agglutinated, cylindrical, small shell with biserially arranged chambers. The chambers grow regularly and have a subglobular to slightly elongated morphology, opening with an angle of 10º-15º (7º to 14º in , 1948). The shell varies from 0.5mm to 1.1 mm in length. The wall consists of a fine epidermis (recrystallized) and few developed beams and rafters. A maximum of six beams per chamber have been measured. The external wall and septa agglutinate coarse and angular quartz grains, as occurs in many other foraminifera located in the Les Bagasses deposits (see for instance, Pseudocyclammina sphaeroidea).
Click on thumbnail to enlarge the image.
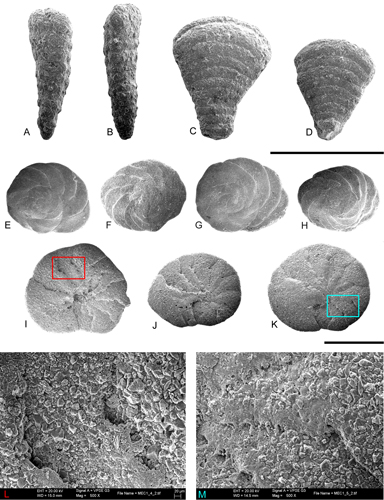
Figure 14: A-B. Cuneolina cylindrica: external view of isolated specimens (scanning electron microphotographs); note the cylindrical morphology of this species. C-D. Cuneolina conica: lateral view of well preserved specimens. E-I. Dictyopsella kiliani: external view of the dorsal side showing the spiral half-moon shaped chambers (scanning microphotographies). J-L. Dictyopsella kiliani: ventral side with the chambers of the last whorl showing the main aperture and small septal openings. M-N. Details of J and L specimens. All from SG-36. Scale bar 1 mm.
Stratigraphical distribution: This species in known from the Santonian and Campanian deposits.
Geographical distribution Cuneolina cylindrica was described from the Middle East, but is a poorly known species. It is common in the Pyrenees.
Family Dictyopsellidae et al., 1983
Remarks: Low trochospiral shell with polygonal subepidermal network. The adjacent chambers communicate by a main aperture in an interiomarginal position.
Dictyopsella , 1899
(Type-species: Dictyopsella kiliani in 1899)
Description: Finely agglutinated shell, with low conical shape showing a convex dorsal side and a plane or plano-convex umbilical side. The chambers are arranged in a low trochospire. In the dorsal side, the chambers have the typical half-moon shape, while in the ventral side they are triangular leaving an open umbilicus in the center of the shell. The main aperture is arched and placed at the base of the apertural face. Small supplementary openings are developed in the septal sutures. The interior of the chambers is occupied by a deep exoskeleton, composed of numerous beams and rafters forming a fine subepidermal polygonal network covered by the epidermis. They lack endoskeleton.
Age: The age attributed by and (1988) to this genus is Cenomanian-to-Maastrichtian.
Dictyopsella kiliani in , 1899
(Fig. 14 E-M ![]() ; Fig. 15 A-F
; Fig. 15 A-F ![]() )
)
Synonymies:
1899 Dictyopsella kiliani in ,
Pl. 8, figs. 5, 7; Pl. 11, fig. 20.
1967 Dictyopsella kiliani in , ,
Pl. 10, figs. 7-10.
1968 Dictyopsella kiliani in , ,
Pl. 5, figs. 7-10.
1981 Dictyopsella kiliani in , ,
Pl. 9, figs. 7-10; Pl. 10, figs. 1-3
1985 Dictyopsella kiliani in , & ,
Pl. 4, figs. 1-7; Fig. 3.
1992 Dictyopsella kiliani in , ,
Figs. 7-10.
Description: All the specimens found in the Font de les Bagasses unit are attributed to the type-species of the genus: D. kiliani. The main characteristics of this species are reflected in the generic description of (1899) from the deposits of the Tragó de Noguera. The shell forms a low cone with its base plane or slightly convex. The periphery is circular and slightly lobulated. The maximum diameter of the measured specimens varies from 0.9 to 2.2 mm (A-forms). In the dorsal side, they show 20-21 half-moon-shaped chambers arranged in two and half whorls, with the last whorl having 8-10 chambers. The dorsal sutures are arched and slightly depressed. In the ventral side, the chambers are triangular and the sutures appear precisely at 2/3 of its inner part of the shell and slightly curved at the 1/3 of the external margin. The embryo consists of a spherical protoconch and a reniform to subglobular deuteroconch with exoskeleton.
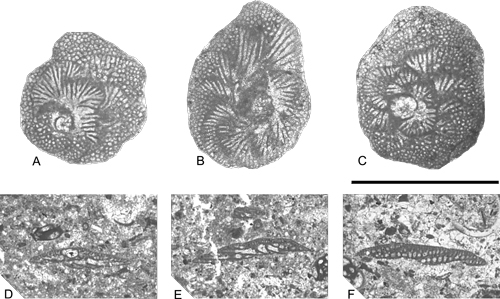
Click on thumbnail to enlarge the image.
Figure 15: A-C. Dictyopsella kiliani from SG-36. A, C. traverse sections showing the disposition of chambers in the spiral side. B. traverse section showing the disposition of chambers in the last whorl.D. axial section of a small specimen showing the spherical protoconch and a reniform deuteroconch, from M-148. E- F. Subaxial sections, from M-150. Scale bar 1 mm.
Remarks: Although (1968), in the material from Martigues, attributed the small specimens (with a diameter of about 1.3 mm) to the A-generation and the large ones (up to 2 mm) to the B-generation, all the measured specimens from the Font de les Bagasses Unit are A- forms
Similarities and differences: (1967), using material from Tragó de Noguera, described the species D. muretae. This species, according to the author, differs from D. kiliani in its larger number of chambers in the last whorl (about 15) and the bigger size of the embryo compared with the embryo of D. kiliani. Moreover, (1967) indicated that the walls of the ventral side of the successive chambers are superposed, forming a columellar plate, suggesting a higher degree of evolution for this last species. However, in the studied specimens collected from the Font de les Bagasses Unit, such characteristics have not been observed.
(1968) described from Martigues, under the name of D. cuvillieri, a small, thin and flat species with circular outline, which was used by & (1985) to erect their new genus Dictyopselloides. & (1985) based the new genus on the presence of an exoskeleton formed only by beams or, if they exist, a few rafters (see & , 1988, p. 150), while D. kiliani has well developed beams and rafters. However, the pictures presented by (1968, Pl. 5, fig. 5) clearly indicate the presence of rafters; therefore, the validity of the genus Dictyopselloides is questioned here. (1968) differentiated D. cuvillieri from D. kiliani by its smaller size, flatter morphology, and the number of chambers in the last whorl, four to five. In our material, morphotypes similar to those described by (1968) as D. cuvillieri occur only sporadically in thin-sections, not permitting a detailed study.
Stratigraphical distribution: Coniacian-Lower Campanian.
Geographical distribution: Dictyopsella kiliani is mentioned frequently from western and central Europe.
In the Font de les Bagasses Unit the agglutinated larger foraminifera are frequently associated with porcellaneous and lamellar-perforate foraminifera. Among the former group, lacazinids and meandropsinids are dominant, while the latter group is dominated by rotaliids and orbitoidids.
The lacazinids are represented by two species that have replaced each other in time: Pseudofabularia cantabrica et al., 1989, and Lacazina elongata , 1899 (generic and specific characteristics of these two taxa are treated in et al., 1989). Meandropsinids are represented clearly by two small, apparently nondimorphic forms: Nummofallotia cretacea (, 1899) and Eofallotia cf. simplex , 2009, which are present within complete unit. The large and extremely dimorphic species Alexina payracea & , 2009, is frequent only in the lower part of the sections, whereas the larger B forms of Larrazetia larrazeti ( in , 1899) occur only in the upper part. The specimens designated in the charts as Fascispira cf. schlumbergeri & , 2009, need to be further studied. Meandropsina vidali , 1899, described from the facies of the Tragó de Noguera, has been identified only in some washed samples collected in the middle part of the unit, and is omitted in the chart range (for more information see & , 2009).
Among the lamellar-perforate foraminifera, the small Orbitoides hottingeri and O. santae-pelagiae are frequent, and they replace each other in time ( et al., 2014). The Family Rotaliidae (sensu , 2014) is represented by Rotorbinella spp., and Pyrenerotalia longifolia et al., 2009. The very small, heavily ornamented forms with areal foramina, umbilical flaps and tooth-plate are designated in this work as Pararotalia tuberculifera (, 1861), but the ensemble should be further studied. The list of hyaline foraminifera ends with the well-known Praestorsella roestae (, 1951), Praesiderolites praevidali (, 1987), and probably several species of the genus Goupillaudina.
The detailed architectural study of the foraminifera collected in the Font de les Bagasses Unit, in the Montsec Mountains, has allowed the following agglutinated foraminifera to be identified: Choffatella aff. rugoretis, Hemicyclammina chalmasi, Pseudocyclammina sphaeroidea, Cuneolina conica, C. cylindrica and Dictyopsella kiliani. All of these share the same type of exoskeleton composed of beams and rafters with a fine epidermis (polygonal subepidermal network) that is combined with different arrangements of chambers to conform:
1. Planispiral to uncoiled, peneropliform or cyclical (Spirocyclinidae);
2. Biserial (Dicyclinidae);
3. Low trochospiral (Dictyopsellidae).
In the suprageneric classification Choffatella is included in Choffatelinae , Hemicyclammina in Hemicyclammininae , and Pseudocyclammina in Amijiellinae within the Spirocyclinidae. Cuneolina is included in the Family Dicyclinidae.
We wish to express our sincere gratitude to Prof. Joan for his help in the field work. Particular thanks are due to the reviewers Dr. Annie , Dr. Felix and to Prof. Bruno (editor) whose comments considerably improved this work. This research was funded by the Spanish Ministry of Science and Innovation (project CGL2009-08371) and Economy and Competitivity (project CGL2012-33160).
S., G., e M. & E. (2014).- The evolution of the earliest representatives of the genus Orbitoides: implications for Upper Cretaceous biostratigraphy.- Cretaceous Research, vol. 51, p. 22-34.
A. & W.V. (1995).- Early Cretaceous shallow-water benthic foraminifers and fecal pellets from leg 143 compared with coeval faunas from the Pacific basin, Central America, and the Tethys.- Proceeding of the Ocean Drilling Program, Scientific Results, College Station, vol. 143, p. 537-564.
M.E., , G., M. & E. (in press).- Benthic foraminifera in the aftermath of the Cenomanian-Turonian boundary extinction event in the carbonate platform facies of the southern Apennines (Italy).- Journal of Foraminiferal Research, Lawrence.
F.T. (1970).- A synopsis of the Spirocyclinidae.- Revista Española de Micropaleontologia, Madrid, vol. 2, nº 3, p. 243-290.
C., G., V., J.M., M., M. & E. (2011).- Larger foraminifera distribution and strontium isotope stratigraphy of the La Cova limestones (Coniacian - Santonian, Serra del Montsec, Pyrenees, NE Spain).- Cretaceous Research, vol. 32, p. 806-822.
C., R., E. & L. (2009).- Late Cretaceous rotaliids (Foraminiferida) from the Western Tethys.- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 253, nº 2-3, p. 197-227.
E. (1981).- Structural analysis of larger foraminifera in random section as an instrument for rapid determination of rock age and environment. In: J. (ed.), International Symposium on Concept and Method in Palaeontology.- Departament de Paleontologia, Universitat de Barcelona, p. 223-232.
E. & A. (1983).- Macroforaminifères du Crétacé supérieur du Bassin Sud-Pyrénéen.- Géologie Méditerranéenne, Marseille, vol. 13, nº 4, p. 137-142.
E., A. & J.M. (1978).- Foraminíferos bentónicos del Santoniense surpirenaico (Montsec de Rubies, prov. de Lerida, España). Nueva adscripción genérica de Dictyopsella chalmasi .- Revista Española de Micropaleontología, Madrid, vol. 10, nº 3, p. 453-460.
E. & A. (1989a).- Correlation of larger benthic and planktonic foraminifera of the Late Cretaceous in the South-Central Pyrenees. In: J. (ed.), Cretaceous of the western Tethys. Proceedings of the 3rd International Cretaceous Symposium (Tübingen).- E. Schweizerbart'sche Verlagsbucchandlung, Stuttgart, p. 231-238.
E. & A. (1989b).- Upper Cretaceous bioestratigraphy of the south-central Pyrenees (Lleida, Spain).- Geodinamica Acta, Paris, (Series 3), vol. 3, p. 221-228.
E., C., J. & J.M. (1999).- El Coniaciense superior-Santoniense inferior de la Sierra del Montsec (Pirineos, NE de España).- Revista de la Sociedad Geológica de España, Salamanca, vol. 12, nº 2, p. 269-280.
E., M., V., G. & R. (2013).- Broeckina gassoensis sp. nov., a larger foraminiferal index fossil for the middle Coniacian shallow-water deposits of the Pyrenean Basin (NE Spain).- Cretaceous Research, vol. 45, p. 76-90.
M., M.L. & R.M. (2012).- Microfacies e microfossili delle successioni carbonatiche mesozoiche del Lazio e dell'Abruzzo (Italia centrale) - Cretacico.- Memorie per servire alla descrizione della Carta Geologica d'Italia, Roma, p. 269.
S. (1997, unpublished).- Els nivells de gresos santonians de les Serres Marginals (Prepirineu català).- MSc Thesis, Universitat Autònoma de Barcelona, Bellaterra, 151 p. [in Catalan].
A. (1977).- Foraminíferos bentónicos del Santoniense del Barranco de la Font de la Plata, Montsec de Rúbies, prov. de Lérida.- Publicaciones de Geología de la Universidad Autónoma de Barcelona, Bellaterra, nº 8, 45 p.
J., L.P., L.E., V.G., X., A.L., C., J.M., J., O., J., C., B., J.C., L.A., M. & J.P. (1985).- Présentation de 9 cartes paléogéographiques au 1/20 000 000, s'étendant de l'Atlantique au Pamir pour la période du Lias à l'Actuel.- Bulletin de la Société géologique de France, Paris, (8e Série), t. I, nº 5, p. 637-652.
H. (1906).- Évolution et enchaînement des Foraminifères.- Bulletin de la Société géologique de France, Paris, (4e Série), t. IV, p. 588-602 (Pl. XVIII).
G. & M. (2008).- Reticulinella kaeveri , and : a marker for the middle upper Turonian in the shallow-water carbonate facies of the peri-adriatic area.- Bollettino della Società Geologica Italiana, Roma, vol. 127, p. 275–284.
G., M., M. & M. (2007).- Carbon and strontium isotope stratigraphy of the Upper Cretaceous (Cenomanian-Campanian) shallow-water carbonates of southern Italy: Chronostratigraphic calibration of larger foraminifera biostratigraphy.- Cretaceous Research, vol. 53, p. 110-139.
C. (1968).- Stratigraphie et micropaléontologie du Sénonien de la région des Martigues près Marseille (Bouches-du-Rhône).- Eclogae Geologicae Helvetiae, Basel, vol. 61, nº 2, p. 657-705.
F.M., J.G., K. & G. (2012).- The Geologic Time Scale 2012.- Elsevier, 2 volumes, 1144 p.
W.W., R.M., C.N., K.M., S., M., A.R., W.C., A.B., A.N. & E. (1999).- Alternative global Cretaceous paleogeography.- Geological Society of America, Special Paper, Washington D.C., nº 332, 47 p.
F.R.S. (1948).- Larger imperforate Foraminifera of South-Western Asia.- British Museum (Natural History), London, 127 p.
J.E. van. (1966).- Orbitoides hottingeri n. sp. from Northern Spain.- Proceedings of the Koninklijke nederlandse Akademie van Wetenschappen, Amsterdam, (Series B), vol. 69, nº 3, p. 388-402.
L. (1967).- Foraminifères imperforés du Mésozoïque marocain.- Notes et Mémoires du Service géologique, Rabat, vol. 209, p. 5-168.
L. (1978).- Comparative anatomy of elementary shell structure in selected larger Foraminifera. In: R.H. & C.G. (eds.), Foraminifera.- Academic Press, London, vol. 3, p. 204-266.
L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, nº M02 (CG2006_M02), 125 p.
L. (2014).- Paleogene larger Rotaliid Foraminifera from the Western and Central Neothetys.- Springer, p. 3-191.
L. & E. (2009).- Meandropsinidae, an ophtalmidid family of Late Cretaceous K-strategist foraminifera endemic in the Pyrenean Gulf.- Neues Jahrbuch für Geologie und Paläontologie, Stuttgart, vol. 253, nº 2-3, p. 249-279.
L., K. & E. (1989).- Late Cretaceous, Larger, Complex Miliolids (Foraminifera) endemic in the Pyrenean Faunal Province.- Facies, Erlangen, vol. 21, p. 99-134.
M.A. (2004).- The year 2000 classification of the agglutinated foraminifera. In: M. & M.A. (eds.), Proceedings of the fifth international workshop on agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 8, p. 237-255.
M.A. (2014).- The year 2010 classification of the agglutinated foraminifera.- Micropaleontology, New York, vol. 60, nº 1, p. 89-108.
A.R. & H. (1964).- Sarcodina, chiefly "Thecameobians" and Foraminiferida. In: R.C. (ed.), Treatise on Invertebrate Palaeontology, Part C, Protist 2.- Geological Society of America, Boulder, and University of Kansas, Lawrence, 900 p.
A.R. & H. (1985).- Some new and redefined genera and families of agglutinated foraminifera; I.- Journal of Foraminiferal Research, Lawrence, vol. 15, p. 91-104.
A.R. & H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, 970 p.
A.R. & H. (1992).- Present status of Foraminiferal classification. In: Y. & T. (eds.), Studies in benthic Foraminifera.- Tokai University, Tokyo, p. 93-102.
A.R. & H. (1994).- Foraminifera of the Sahul Shelf and Timor Sea.- Cushman Foundation for Foraminiferal Research, Special Publication, Lawrence, vol. 31, 661 p.
W. (1950).- The foraminiferal genus Choffatella in the Lower Cretaceous (Urgonian) of the Caribbean region (Venezuela, Cuba, Mexico and Florida).- Eclogae geologicae Helvetiae, Basel, vol. 42, nº 2, p. 529-547.
W. (1953).- Hemicyclamina sigali, n. gen. nov. sp. from Cenomanian of Algeria.- Contributions from the Cushman Foundation for Foraminiferal Research, Lawrence, vol. 4, nº 9, p. 148-149.
W. (1958).- Feurtillia frequens, n. gen., n. sp., a new genus of Lituolid foraminifera.- Contributions from the Cushman Foundation for Foraminiferal Research, Lawrence, vol. 9, part. 1, p. 1-3.
W. (1972).- Gendrotella n. gen. and Choffatella caronae n. sp. from the Lower Senonian of Southern France.- Eclogae geologicae Helvetiae, Basel, vol. 65, p. 355-359.
K.G., M.A., J.V., J.D., G.S., M.E., P.J., B.S., N. & S.F. (2005).- The Phanerozoic Record of Global Sea-Level Change.- Science, Washington D.C., vol. 310, p. 1293-1298.
E. (1887).- Sur la Cyclolina et trois nouveaux genres de foraminifères des couches à rudistes: Cyclopsina, Dicyclina et Spirocyclina.- Compte Rendu des Séances de la Société géologique de France, Paris, vol. 4, nº 7, p. 30-31.
J.A. (1992).- Evolution of a continental collision belt: ECORS-Pyrenees crustal balanced cross-section. In: K.R. (ed.), Thrust tectonics.- Chapman & Hall, London, p. 235-246.
A. (2000, unpublished).- Biostromas de corales, rudistas y esponjas coralinas del Santoniense del Montsec d'Ares: caracterización estratigràfica, sedimentológica y paleontológica.- MSc Thesis, Universitat Autónoma de Barcelona, Bellaterra, 216 p.
A. d' (1850).- Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson, Paris, vol. 2, 428 p.
J., M. & J. (2013).- New supraordinal classification of Foraminifera: Molecules meet morphology.- Marine Micropaleontology, vol. 100, p. 1-10.
S. (1999).- On some little known foraminifers from the Senonian of the Vardar Zone.- Acta Palaeontology Romaniae, Cluj-Napoca, vol. 2, p. 373-375.
J.M. (1977).- Estudio estratigráfico y paleontológico de los yacimientos de Rudistas del Cretácico Superior del Prepirineo de la provincia de Lérida.- Publicaciones de Geología, Universitat Autònoma de Barcelona, Bellaterra, nº 3, 105 p.
E., C., S.M.F., G., S. & P. (2003).- Thermal evolution of Cretaceous Tethyan marine waters inferred from oxygen isotope composition of fish tooth enamels.- Paleoceanography, Washington D.C., vol. 18, nº 2, p. 1-12.
R., V., D. & B. (2010).- The age of the brachiopod limestones from Guča, western Serbia.- Annales géologiques de la Péninsule Balkanique, vol. 71, p. 73-93.
F. (1992).- Benthonische Foraminiferen aus Flachwasserkarbonaten der Oberkreide der Nördlichen Kalkalpen (Gosauschichtgruppe, Österreich).- Mitteilungen der Österreichischen geologischen Gesellschaft, Wien, vol. 84, p. 327-353.
F. & M. (2004).- Hemicyclammina chalmasi (, 1899) - Larger benthic Foraminifera from the Upper Cretaceous Gosau Group of the Northern Calcareous Alps (Gosau, Austria).- Jahrbuch der geologischen Bundesanstalt, Wien, vol. 144, p. 367-372.
C. (1899).- Note sur quelques foraminifères nouveaux ou peu connus du Crétacé d'Espagne.- Bulletin de la Société géologique de France, Paris, (3e Série), t. XXVII, p. 456-465 (Pls. VIII-XI).
M. (1980).- Les Foraminifères imperforés des milieux de plate-forme au Mésozoïque : détermination pratique, interprétation phylogénétique et utilisation biostratigraphique.- Revue de Micropaléontologie, Paris, vol. 23, nº 3-4, p. 169-203.
M. (1988).- Vers une classification évolutive des Lituolidés (Foraminifères) jurassiques en milieu de plate-forme carbonatée.- Revue de Paléobiologie, Genève, vol. spec. nº 2 (Benthos '86), p. 229-256.
A. (1986).- Carbonate platform depositional sequences, Upper Cretaceous, south-central Pyrenees (Spain).- Tectonophysics, vol. 129, p. 205-231.
A. & J.A. (2000).- Evolución tectonosedimentaria del Pirineo meridional durante el Terciario: una síntesis basada en la transversal del río Noguera- Ribagorçana.- Revista de la Sociedad geológica de España, Salamanca, vol. 13, nº 2, p. 251-264.
G. (1981).- Les foraminifères crétacés de Provence (Aptien-Santonien). Systématique, biostratigraphie, paléoécologie, paléogéographie.- PhD Thesis, Université de Provence ; Travaux du Laboratoire de Géologie Historique et de Paléontologie, Marseille, vol. 12, nº 3, 559 p.
R. (2009, unpublished).- Los macroforaminíferos de la plataforma Campaniense del margen sur de la cuenca pirenaica (Unidad Surpirenaica central, NE de la Península Ibérica).- PhD Thesis, Universitat Autònoma de Barcelona, Bellaterra, 216 p.
M. (1983).- Évolution, biostratigraphie et systématique des Siderolitinae (Foraminifères).- Revista Española de Micropaleontologia, Madrid, vol. 15, nº 1, p. 5-37.