◄ Carnets Geol. 16 (6) ►
![]()
Contents
[Introduction]
[Material studied]
[Taxonomic affinities]
[Systematics]
[Comparisons] [Conclusions]
[Bibliographic references] and ... [Plates]
"Cátedra Franco-Brasileira no Estado de São Paulo 2015", UNESP - Universidade Estadual Paulista, Center for
Geosciences Applied to Petroleum (UNESPetro), Caixa Postal 178, Av. 24 A, no. 1515, Bela Vista, CEP13506-900 - Rio
Claro - SP (Brazil)
Dépt. STU, Fac. Sci. Tech., UBO, CS 93837, F-29238 Brest (France)
Department of Ecology and Evolutionary Biology, The University of Kansas, 1200 Sunnyside
Avenue, Lawrence, Kansas 66045 (USA)
UNESP - Universidade Estadual Paulista, Center for
Geosciences Applied to Petroleum (UNESPetro) & Departamento de Geologia
Aplicada, Caixa Postal 178, Av. 24 A, no. 1515, Bela Vista, CEP13506-900 - Rio
Claro - SP (Brazil)
Published online in final form (pdf) on April 14, 2016
[Editor:
Michel ; technical editor: Bruno ;
language editor: Phil ]

According to some authors, the three fossil algal forms discussed in this paper, Cordilites cretosus (, 1846), Elianella elegans & , 1948, and Parachaetetes asvapatii in & , 1936, are synonyms. However, we document that they represent three discrete species. The genus Elianella & , 1948, gets unique features that make it easy to discriminate; we describe a new species from Albian limestones of Brazil. The other two genera, Cordilites , 1887, and Parachaetetes , 1906, share some common features, such as having a thallus made of a lattice-network with columns (filaments) and rows. However, on the basis of their diagnostic criteria, the species Cordilites cretosus and Parachaetetes asvapatii should not be confused.
Rhodogorgonales; Elianellaceae; Solenoporacea; Cordilites; Elianella; Parachaetetes.
B. & D. (2016).- On the fossil alga Elianella elegans & , 1948, and its so-called lookalikes, with description of Elianella brasiliana n.sp. Revision of the Juliette Collection. Part 1.- Carnets Geol., Madrid, vol. 16, no. 6, p. 213-229.
À propos de l'algue Elianella elegans & , 1948, et ses soit-disants semblables, avec la description de Elianella brasiliana n.sp. Révision de la Collection Juliette . 1e partie.- D'après certains auteurs, les trois formes fossiles algaires discutées dans cette publication, Cordilites cretosus (, 1846), Elianella elegans & , 1948, et Parachaetetes asvapatii in & , 1936, sont synonymes. Nous démontrons cependant qu'elles constituent trois espèces distinctes. Le genre Elianella & , 1948, a des caractéristiques uniques qui le rendent facile à identifier. Nous décrivons d'ailleurs une nouvelle espèce provenant de calcaires albiens du Brésil. Les deux autres genres, Cordilites , 1887, et Parachaetetes , 1906, partagent quelques caractères communs, notamment celui d'avoir un thalle constitué par un réseau de cellules ayant l'aspect d'une grille avec des colonnes (filaments) et des rangées. Toutefois, sur la base de leurs critères diagnostiques, les espèces Cordilites cretosus et Parachaetetes asvapatii ne devraient normalement pas être confondues.
Rhodogorgonales ; Elianellaceae ; Solenoporacea ; Cordilites ; Elianella ; Parachaetetes.
The present paper focuses on Elianella elegans & , 1948, and two "lookalikes": Cordilites cretosus (, 1846), and Parachaetetes asvapatii in & , 1936. Paleophycologists, i.e., those people assuming they were dealing with fossil algae, and other paleontologists, i.e., those considering they were dealing with Cnidaria or Porifera, have treated these three taxa either as individual species (e.g., & du , 1970) or as synonyms. In the latest case they have treated either the three of them or only a pair as synonyms. For instance, regarding Elianella elegans and Cordilites cretosus, (1980a) stated that "il s'agit en fait de la même forme" [as a matter of fact it is the same form]; the same year, (1980b) again concluded that "ces trois formes sont synonymes" [these three forms are synonymous]. According to (1989) and (2000), Elianella elegans is "a younger synonym of Parachaetetes asvapatii". However, in our opinion, they represent three discrete species that have often been misidentified. For instance, the Elianella elegans reported by (1967) from the Danian interval in the SNPA exploration well Bastennes-Lonné l (FB1, Aquitaine, SW France) is a genuine Parachaetetes , 1906 (cf. Parachaetetes asvapatii), as documented hereafter. We report and illustrate below the main features of these three species in order to prevent them being confused again in the future.
Ideally, this kind study would have required access to all the type materials, which was not feasible in the present case. However, the material studied here comprises thin sections from several researchers' collections: (topotypic material), , and - , as well as Brazilian material collected more recently, which display well-preserved specimens documenting diagnostic features.
Considering Elianella elegans & ,
1948, the first
author (B.G.) found only one thin section in the Juliette
Collection (Fig. 1.A ![]() ). It is not one of the thin sections with the specimens
originally illustrated by the authors ( & ,
1948,
Pl. XII, figs. 1-3), because this set is probably lost, but this thin section is
marked "37 EB16", "NW de Lohaosy" (Ampanihy, Toliary), and
"Calc. à Algues", i.e., it is a topotypic material from the
Paleocene of S Madagascar. This specimen (Fig. 1.B
). It is not one of the thin sections with the specimens
originally illustrated by the authors ( & ,
1948,
Pl. XII, figs. 1-3), because this set is probably lost, but this thin section is
marked "37 EB16", "NW de Lohaosy" (Ampanihy, Toliary), and
"Calc. à Algues", i.e., it is a topotypic material from the
Paleocene of S Madagascar. This specimen (Fig. 1.B ![]() ) is selected here as the
neotype.
) is selected here as the
neotype.

Click on thumbnail to enlarge the image.
Figure 1: A) Scan of the thin section marked "37 EB16", "NW de Lohaosy", and "Calc. à Algues", Juliette Collection; B) photomicrograph of the neotype; C) detail with longitudinal sections of filaments. [B scale bar 5 mm ; C scale bar 500 µm]
While investigating the Collection at the Smithsonian National Museum of Natural History, the first author identified within a small set of thin sections nice specimens of Cordilites cretosus (, 1846) in one thin section marked D619-a-629 and bearing "Puerto Rico" as geographical label. The remaining thin sections yield "mid" Cretaceous (=Aptian-Cenomanian) foraminifera, i.e., undetermined orbitolinidids.
The material from the Collection was combined with that of the Collection as both consist of 27 thin sections of similar origin (core samples from the intervals 222.36-233.75 with a BGh1 label and 223.03-233.53 with a Bastennes 1 label, Danian in age, and both from the SNPA exploration well Bastennes-Lonné l - FB1, Aquitaine, SW France).
Most of the studied Brazilian Albian-Cenomanian material is registered in the collection of the "UNESPetro - Centro de Geociências Aplicadas ao Petróleo, UNESP - Universidade Estadual Paulista, Rio Claro" (São Paulo, Brazil); another part, consisting of a limited number of thin sections, is deposited with LPB ("Laboratoire de Paléontologie de Brest") numbers in the collections of the "Département des Sciences de la Terre et de l'Univers, Université de Bretagne Occidentale, Brest" (France).
The three forms studied were formerly ascribed to the Family Solenoporaceae , 1927, a group of extinct "calcareous algae", because they share some morphological features: A) they are either encrusting or forming free structures; B) their framework is made of erected filaments, more or less densely juxtaposed, with common to rare horizontal partitions (i.e., they are pluricellular algae), defining a more or less preserved lattice-network with columns and rows; C) their mineralization is intracellular, centripetal (starting from the cell walls inwards), and consists of elongated fibrous calcite (probably high-Mg calcite) crystals, another feature that justifies their ascription to the red algae. However, D) reproductive organs (possibly external) are unknown contrary to the records in its sister families, e.g., Hapalidiaceae , Sporolithaceae or Graticulaceae & .
After having reviewed earlier records of suspected
reproductive structures in the Family Solenoporaceae and concluding that there
were none, and
(2001) describe structures that
they interpret as "nemathecia" on a Parachaetetes asvapatii
in & , 1936. Unfortunately, this unique specimen proves
to be a Cidaroida spine. The stereom of the echinoid spine was misinterpreted as
the algal structure and the longitudinal crests forming the ornamentation of the
spine were misinterpreted as nemathecia ( et al.,
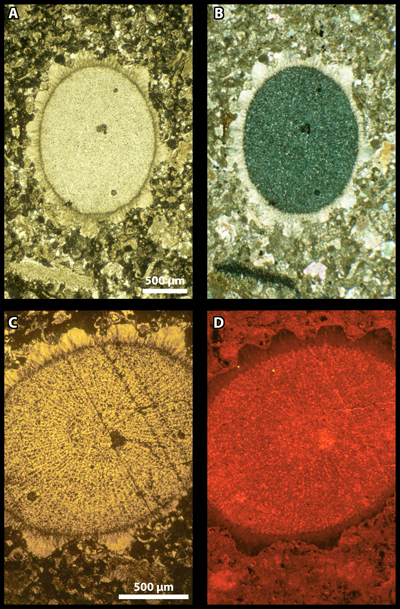
2009): see Fig. 2 ![]() , for example.
, for example.
Click on thumbnail to enlarge the image.
Figure 2: Section of a Cidaroida spine. A & C) conventional transmitted light/plane polarized light (C is an enlargement of A); B) cross polarized light; D) cathodoluminescence. The view under cross nicols (B) illustrates the spine core with a single-crystal behaviour (dark) and its fibrous rim with a radial behaviour (as documented by a faint extinction cross). Under cathodoluminescence, the crests forming the rim record various shades of colour suggesting that they grew slowly enough to record slight changes in the chemical composition of the spine calcite. Sample A10-3660, Paleocene, Libya. Similar material was illustrated by (1966, Pl. LXXXIII, fig. b) and et al. (1972, Pl. CLI, figs. 1-2). [A-B scale bar 500 µm ; C-D scale bar 500 µm]
Last but not least, (2004) demonstrated that the type of the genus Solenopora, S. spongioides , 1877, is actually "a chaetetid sponge, not an alga". The first author (B.G.) introduces hereafter the Family Elianellaceae nov. fam. in replacement of the Family Solenoporaceae because, further to 's (2004) report, the type of the latter is currently ascribed to the Animal Kingdom. This new family is tentatively assigned to the Order Rhodogorgonales & , 1995, a sister group to the Sporolithales, which to date includes few living representatives with two genera only arranged into one family, the Rhodogorgonaceae & , 1995.
Preliminary remark: The "synonymy lists" below are not as comprehensive as one could expect. The main difficulty stands in the magnification of the original photomicrographs that is often insufficient to allow discrimination of Elianella elegans from Parachaetetes asvapatii, for instance. Actually, in such photomicrographs, it is easier to say which one is not displaying diagnostic features rather than to accurately identify a species. Another issue is that the type-material of Parachaetetes asvapatii might be composite, consisting of pieces of both Parachaetetes and ? Elianella. Both taxa are partly coeval and share the same living environment: for instance, in our Albian Brazilian material, representatives of Elianella and Parachaetetes are commonly found together. Largely for this reason, our standard for Parachaetetes asvapatii will be limited to the sole photomicrograph referred to as representing the "type-specimen" by its author ( in & , 1936: Pl. III, fig. 3), which illustrates a genuine representative of the genus Parachaetetes (, 1906), but it will not comprise any of the accompanying figures ( in & , 1936: Figs. 35-38; Pl. III, figs. 1-2 & 4-5).
Division Rhodophyta
Class Florideophyceae , 1960
Subclass Corallinophycidae & , 2007
Order ? Rhodogorgonales & , 1995
Family Elianellaceae nov. fam. (B.G.)
(formerly Family Solenoporaceae , 1927)
Genus Elianella & , 1948
Type: Elianella elegans & , 1948, p. 275-276, Pl. XII, fig. 1-3.
Elianella elegans & , 1948
(Fig. 1.B-C ![]() ;
Pl. 1
;
Pl. 1 ![]() , figs. A-F;
Pl. 2
, figs. A-F;
Pl. 2 ![]() , figs. A, D, F & H-I)
, figs. A, D, F & H-I)
Selected references:
|
Elianella elegans n.sp.- & , p. 275-276, Pl. XII, fig. 1-3 |
||
|
non |
Elianella elegans.- , p. 47-48, Pl. 1, fig. 15 pars; Pl. 2, figs. 1-3 [cf. Parachaetetes asvapatii] |
|
|
non |
Elianella elegans.- & du , p. 208, Pl. 2, fig. 11 [= ? Cordilites sp.] |
|
|
non |
Elianella elegans.- et al., p. 499-500, Pl. 48, figs. 1-2 [= Parachaetetes sp.] |
|
|
Elianella elegans.- , p. 324-327, Pl. I, figs. 1-2; Pl. II, figs. 1-2 |
||
|
Elianella elegans.- & , p. 36-37, Pl. 1, figs. 1 & 4 |
||
|
Elianella elegans.- & , Pl. 4, figs. 3-4 |
||
|
non |
Elianella elegans.- & , Pl. 4, fig. 1 [= ? Parachaetetes sp.] |
|
|
non |
Elianella elegans.- & , Pl. 4, fig. 1 [= ? Parachaetetes sp.] |
|
|
non |
Elianella elegans.- et al., p. 433, Pl. II, fig. 3 [= Parachaetetes sp.] |
|
|
non |
Elianella elegans.- , p. 428, Pl. II, fig. 9 [= Parachaetetes sp.] |
|
|
? |
Baratangia densituba gen. et sp. nov.- & , p. 151, Figs. 3 (holotype) & 4-5 (paratypes) |
|
|
? |
Parachaetetes asvapatii.- , p. 70-71, Pl. 4, figs. 2-4 & 6; Pl. 5, figs. 1-3 |
|
|
Elianella elegans.- et al., p. 177, Pl. 5, figs. 5-6 & 8 pars |
||
|
Elianella elegans.- , p. 47, Pl. 17, figs. 4-6 |
||
|
Parachaetetes asvapatii.- , p. 428-432, Figs. 3.a-c-4.a-d |
||
|
non |
Elianella elegans.- & , p. 1117-1118, Pl. I, figs. 5-6 [= Parachaetetes sp.] |
|
|
? |
Elianella elegans.- & , p. 303, Pl. III, figs. 1-2; Pl. IV, figs. 1-4 |
|
|
Elianella elegans.- et al., p. 313, Pl. 2, fig. E |
||
|
Elianella elegans.- , p. 527, Pl. 6 |
Neotype
(defined here): Fig. 1.B ![]() , details in
Fig. 1.C
, details in
Fig. 1.C ![]() ; Pl. 1
; Pl. 1 ![]() , figs. A-F. Thin section
"37 EB16", "NW de Lohaosy", "Calc. à Algues",
Juliette Collection; Paleocene; S Madagascar.
, figs. A-F. Thin section
"37 EB16", "NW de Lohaosy", "Calc. à Algues",
Juliette Collection; Paleocene; S Madagascar.
Original description ( () in & , 1948, p. 275-276): "À l'extérieur, dans la roche, Elianella se montre sous forme de taches claires, jaunâtres, d'aspect porcelané, sur lesquelles la structure est généralement indiscernable macroscopiquement.
Il s'agit de thalles arrondis ou lobés, composés de files cellulaires, juxtaposées plus ou moins étroitement suivant les points, et qui, au lieu de présenter des parois rectilignes, sont moniliformes ; ce sont comme de petits tonnelets disposés bout à bout, les cloisons sont très arquées, en forme de verre de montre. Elles peuvent arriver, suivant l'orientation de la coupe, à se montrer superposées comme une pile d'écuelles ; elles sont disposées sans ordre, et non pas linéairement. Cependant, lorsqu'un thalle, qui débute toujours avec un contour circulaire, arrive à former une sorte de rameau ou de digitation, les tonnelets s'allongent, dans l'axe de la poussée, et tendent à se disposer en rangées concentriques. On voit parfois subsister ces lignes d'accroissement du thalle, alors même que tout le tissu a été détruit par cristallisation, au cours de la fossilisation. En section longitudinale, les tonnelets ont 45 µ de large, avec 50 à 60 µ de long ; ils atteignent toutefois 100 x 50 µ sur le rachis (axe des digitations). La section transversale des filaments est arrondie et devient polygonale quand ils sont plus serrés. Parfois on voit les sections rondes des tubes paraître en sombre au centre d'hexagones hyalins, quand la cristallisation envahit les parois."
["From outside, in the rocks, Elianella shows up as bright spots, yellowish, porcelain-like, on which the structure is usually macroscopically invisible.
It forms rounded or lobed thalli, consisting of lines of cells, more or less closely juxtaposed depending on the area, and that, instead of having rectilinear walls, are moniliform; they look like small drums placed end-to-end, the cross partitions are highly-arched, with a watchglass shape. Depending on the angle of the section they may look stacked like a pile of bowls; they are arranged without order, non-linearly. However, because a thallus, which always begins with a circular outline, comes to form some sort of branch or digitation, the drums get longer along the growth axis and tend to form a set of concentric rows. Sometimes such growth lines of the thallus remain visible, even though the tissue has been destroyed by crystallization in the course of the fossilization process. In longitudinal sections, the drums are 45 µm wide and 50 to 60 µm long; however they reach 100 x 50 µm in the rachis (digitation axis). Transverse sections of the filaments are round-shaped, becoming polygonal-shaped as they tighten. Sometimes, when crystallization invades the walls, the rounded sections of the tubes appear as dark spots at the center of hyaline hexagons."]
Additional remarks: A key diagnostic feature, i.e., the round outline of the transverse sections of the erected filaments, is directly correlated to the loose arrangement of the filaments. Cells may vary in length (and growth rates may have varied accordingly) within a single filament or from one filament to the neighbouring ones; filaments grew independently from each other.
Elianella brasiliana n.sp.
(Pl. 2 ![]() , figs. B-C, E & G)
, figs. B-C, E & G)
|
Elianella sp.- , p. 527, Pl. 7 |
Holotype: Pl. 2 ![]() , fig. B, detail in
Pl. 2
, fig. B, detail in
Pl. 2 ![]() , fig. C
(longitudinal section); paratype: Pl. 2
, fig. C
(longitudinal section); paratype: Pl. 2 ![]() , fig. G (transverse section). Thin
section UPAFSE 049a.
, fig. G (transverse section). Thin
section UPAFSE 049a.
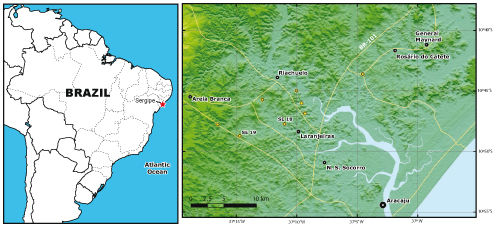
Locus typicus (Fig. 3 ![]() ): locality SE18 (latitude: 10°47'44.017"S;
longitude: 37°11'0.290"W),
2.5 km NW of Laranjeiras, some 20 km NW of Aracaju (Sergipe, Brazil). It
is also present in SE19 (latitude: 10°48'43.033"S; longitude: 37°14'42.330"W),
9 km W of Laranjeiras
and 10 km SE of Areia Branca, some 25 km NW of Aracaju.
): locality SE18 (latitude: 10°47'44.017"S;
longitude: 37°11'0.290"W),
2.5 km NW of Laranjeiras, some 20 km NW of Aracaju (Sergipe, Brazil). It
is also present in SE19 (latitude: 10°48'43.033"S; longitude: 37°14'42.330"W),
9 km W of Laranjeiras
and 10 km SE of Areia Branca, some 25 km NW of Aracaju.
Stratum typicum: Lower part of the Riachuelo Formation, Albian.
Description: It is a typical Elianella with rather large diameter filaments (100 µm in average, up to 150 µm, twice those of E. elegans) and cell lengths of 40 µm in average (and up to 160 µm). The filaments are separated and the open space between them was later filled by a hyaline calcitic drusy cement. As for the type-species, the transverse cell-walls are upward-concave (downward-convex) and the files of cells may branch dichotomously. Both features can be used to identify the thallus growth direction.
Paleophycological association: The limestones studied are either algal framestones or algal floatstones with grainstone matrices, resulting from the breakage and erosion of former framestones. This new species is found there is associated with Elianella elegans & , 1948, Lithothamnion sp., Parachaetetes cf. tornquisti (, 1906), Pycnoporidium sinuosum & , 1960, Marinella lugeoni , 1939, the problematic Bacinella, and Neomeris cretacea , 1899.
Click on thumbnail to enlarge the image.
Figure 3: Elianella brasiliana collection localities in the Sergipe Basin, Brazil. SE18 = 2.5 km NW of Laranjeiras; SE19 = 9 km W of Laranjeiras and 10 km SE of Areia Branca; both localities NW of Aracaju, State of Sergipe, Brazil.
Genus Parachaetetes (, 1906)
Type: Parachaetetes tornquisti (, 1906), p. 65-66, Pl. VI, fig. 6.
Parachaetetes cf. asvapatii in & , 1936
(Pl. 3 ![]() , figs. A-D)
, figs. A-D)
Selected references (for Parachaetetes asvapatii in & , 1936):
|
Parachaetetes
asvapatii sp. nov.- in & , p.
32-34, Pl. III, fig. 3
("Type-specimen") [herein Fig. 4 |
||
|
? |
Parachaetetes asvapatii sp. nov.- in & , p. 32-34, Figs. 35-38; Pl. III, figs. 1-2 & 4-5 |
|
|
cf. |
Elianella elegans.- , p. 47-48, Pl. 1, fig. 15 pars; Pl. 2, figs. 1-3 |
|
|
? |
Parachaetetes asvapatii.- , p. 70-71, Pl. 5, fig. 4 |
|
|
non |
Parachaetetes asvapatii.- , Pl. 4, figs. 2-4 & 6; Pl. 5, figs. 1-3 [= ? Elianella elegans] |
|
|
non |
Parachaetetes asvapatii.- , p. 428-432, Figs. 3.a-c-4.a-d [= Elianella elegans] |
|
|
non |
Parachaetetes asvapatii.- & , p. 1117-1118, Pl. I, figs. 1-4 [= a Cidaroida spine according to et al., 2009] |
|
|
? |
Parachaetetes asvapatii.- & , p. 305, Pl. II, figs. 1-2 |
Description: The definition that follows is an excerpt from (in & , 1936, p. 34):
"Parachaetetes asvapatii builds small calcareous nodules measuring a few millimetres only, composed of several more or less broad and blunt lobes. The tissue is fairly well differentiated into a hypothallium and a perithallium. The latter shows in longitudinal sections a "lattice-work" formed by the transverse cell-walls (situated at identical height in adjoining cell-rows) and by the longitudinal ones. The cells measure 0.04 to 0.06 mm. across and are one to three times as long as broad.
Type-specimen G.S.I. Type Number 15,769 and Plate III, fig. 3."
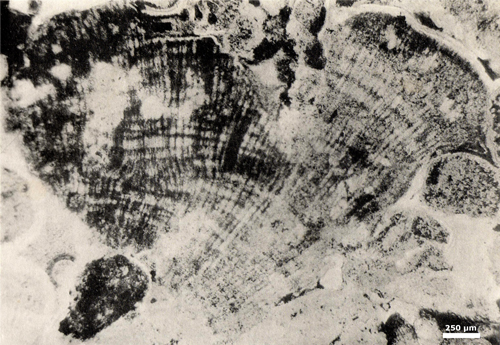
Click on thumbnail to enlarge the image.
Figure 4: Holotype of Parachaetetes asvapatii in & , 1936 (excerpt from & , 1936: Pl. III, fig. 3), G.S.I. = Geological Survey of India, Calcutta, India. Type Number 15,769. Niniyur Group, Upper Cretaceous; Trichinopoly, S India. [scale bar 250 µm]
Additional remarks: The specimens found in the - Collections show "transverse cell-walls" ( ) "situated at identical height in adjoining cell-rows", which ascribes them to a genuine representative of the genus Parachaetetes (, 1906). In longitudinal section, the side-walls of the cells were either poorly calcified or may have suffered some dissolution; consequently, the rows of the alga "lattice-work" are commonly better expressed than the columns. In transverse section, the thalli of these Parachaetetes cf. asvapatii show wavy concentric zones identical to those observed in the generotype, P. tornquisti (, 1906: Pl. VI, fig. 6a; , 1929: Pl. IV, fig. 1).
Genus Cordilites , 1887
Type: Chaetites cretosus.- , 1846, p. 63, Pl. XLIII = VIIL, fig. 4.a-d.
Cordilites cretosus (, 1846)
(Pl. 4 ![]() , figs. A-I)
, figs. A-I)
Synonymy list:
|
Chaetites cretosus.- , p. 63, Pl. XLIII = VIIL, fig. 4.a-d |
||
|
Cordilites nov. gen. cretosus.- , p. 24-25, Figs. 7-8; Pl. I, fig. 3.a-d |
||
|
Petrophyton miyakoense gen. et sp. nov.- , p. 6-8, Fig. 4.a-c ; Pl. II, figs. 1-8 |
||
|
Cordilites cretosus.- , p. 104-110, Pl. I, figs. 1-2; Pl. II, figs. 3-6; Pl. III, figs. 7-10; Pl. IV, figs. 11-13 |
||
|
Cordilites cretosus.- , p. 234, Pl. 1, figs. 1-6 |
||
|
Cordilites cretosus.- , p. 70, Pl. 3, fig. 5 |
Description: The description that follows is an excerpt from (1912, p. 6-7), the first author to treat this fossil as an alga under "Petrophyton miyakoense gen. et sp. nov.":
"Die manchmal 12 cm. erreichenden Knollen zeichnen sich durch die auf der frischen Bruchfläche deutlich hervortretenden Radialfasern aus, welche gegen die concentrischen Schichten weit hervortreten; auf der verwitterten Oberfläche aber ist dieses Verhältnis ganz umgekehrt. Die meisten mir vorliegenden Exemplare sind abgerundet und deshalb ist die Gestalt der vollständigen Stöcke sowie die Anfangsstelle ihres Wachstums nicht leicht zu erkennen. Es scheint mir aber höchst wahrscheinlich dass die Stöcke welche mehr oder weniger birnförmig sind, gewöhnlich zu einer grösseren Masse mit der nierenförmigen Oberfläche, zusammenwachsen. (Textfig. 4, a-c).
Die Stöcke sind aus feinen, im Durchschnitt 0.04 mm. breiten, prismatischen, dicht aneinander liegenden Zellröhren von wesentlich gleichartiger Beschaffenheit zusammengesetzt. Die Zellröhrenwände sind doppelt und nicht einfach bei der vorhergehenden Art. Da ist eine Medianlamelle, welche erst bei starker Vergrösserung als wirklich vorhanden wahrnehmbar ist; sie ist gleichmässig sehr dünn und scheinbar undurchbohrt. An beiden Seiten dieser Lamelle findet immer eine Verdickung statt, welche in, bestimmten Abständen sehr regelmässig sich verengen und sich dann wieder verbreitep. Die erstere Kalkschichte scheint ein Absatz ausserhalb der Zellmembran zu sein, während die zweite eine innerhalb der Zellmembran stattgefundene Verkalkung representiert. Die Querböden sind zahlreich, aber sehr schwach entwickelt; sie sind etwas nach oben concav und finden sich gewöhnlich dort, wo die Wandverdickung beinahe ganz verschwunden ist. Zumeist tretten sie in einem Abstand von 0.055 bis 0.035 mm. nicht selten aber auch ganz dicht aufeinander auf (Fig. 2). In den gesammten Zellröhren erscheinen sie in derselben Höhe und infolgedessen bilden sich die zahlreichen concentrischen Schichten welche wie oben gesagt, auf den ausgewitterten Bruchflächen deutlich sichtbar werden. Es kommt auch bei dieser Form die Diskordanzlagerung der neuen Zellröhrenreihen auf den alten vor, gerade so wie bei Lithothamnium, Solenopora und Metasolenopora (Fig. 2).
Die Vermehrung der Zellröhren findet statt: erstens durch die Einschaltung neuer Zellen an der Ecke, wo drei oder vier Röhren zusammenstossen, und zweitens durch die Teilung, welche ihre Entstehung der neuen Bildung einer Wand in der Mitte eines Rohres verdankt.
Die sehr feinen Zellröhren dieser Form schliessen ihre Zugehörigkeit zu einer Bryozoen- oder Tabulaten-Gattung aus, obwohl die Verdickung der Wände stark an Stenopora erinnert. Ueberhaupt wird die Stock einem Kalkgebilde von einem Tier nicht angehören; vielmehr scheint ihr gesammter Habitus sowie. Struktur den Kalkalgen wie Lithothamnium und Solenopora sich näher anzuschliessen. Die Wandverdickung ausgenommen, stimmt die Wandstruktur unserer Form mit der von Solenopora besonders gut überein; bei beiden sind die Zellröhren langgestreckt und prismatisch, und mit schwach entwickelten Querböden versehen; sie sind auch gerade so breit bei unserer Form wie bei Solenoporella jurassica und Metasolenopora Rothpletzi.
Da diese Form einen ganz neuen Kalkalgentypus zu representieren scheint, werde ich diese Petrophyton nennen."
["The tubers are as large as 12 cm in size. On the freshly broken surface clearly visible radial fibers stand out against concentric layers. On the weathered surface this shape is reversed. Most of the samples available to me are rounded. Therefore, the shape of the original samples as well as the initial place of their growth cannot be easily discerned. However, I conclude that the original specimens were probably pear-shaped fused into a larger mass with a kidney-shaped surface (text-fig. 4, a-c).
The samples are composed of fine, 0.04 mm diameter, prismatic, tightly packed homogenous cellular tubes. The walls of the tubes are double compared to the single ones in the previous species. A median lamella can be observed only at high magnification because it is very thin throughout; it is apparently continuously without holes. A thickening on both sides of the lamella gets alternately narrower and wider in certain distances. The first layer of lime seems to be deposited outside the cell membrane while the second layer of calcification has been deposited inside. The numerous cross walls are very thin; they are sometimes upwardly concave and are usually located where the wall is thinnest. Most frequently they appear at a distance of 0.055 to 0.035 mm, but usually not close to each other (Fig. 2). They are observed at the same level in the cell tubes, and because of this the numerous concentric layers are clearly recognizable on the weathered surfaces, as mentioned above. As in Lithothamnium, Solenopora and Metasolenopora there is also in this sample unequal disposition of the new rows of cell tubes upon the old ones (Fig. 2).
Reproduction of the cell tubes occurs: first by insertion of new cells at the junction where three or four tubes meet, and then followed by division to form a wall in the middle of a tube.
The very fine cell tubes of this specimen exclude it being Bryozoa or Tabulata although the thickening of the walls is characteristic of the genus Stenopora. In any case, the sample is different from the calcified construction of an animal; the basic habit and the structure are similar to calcareous algae such as Lithothamnium and Solenopora. Except for the wall structure, our specimen corresponds especially well to the genus Solenopora; both have long, prismatic cell tubes and are supported by thin cross walls. Also they have exactly the same width as those in Solenoporella jurassica and Metasolenopora Rothpletzi.
Since the specimen seems to represent a completely new type of calcareous algae I will name it Petrophyton." translated from the German by Sigrid ]
Additional
remarks: The thallus pattern is similar to that of a genuine Parachaetetes,
except for the thicker calcification. In addition, "septal
projection" sensu (2004) or incomplete walls (Pl.
4 ![]() ,
figs. B-D) are more common than in Elianella elegans.
,
figs. B-D) are more common than in Elianella elegans.
Thalli of Cordilites cretosus and Parachaetetes asvapatii, as the peripheral parts of the vegetative thalli of both the Sporolithaceae and the Hapalidiaceae, display a lattice-pattern defined by columns made of files of cells and rows resulting from the concurrent growth of these cells in adjacent filaments.

Click on thumbnail to enlarge the image.
Figure 5: A-E) transverse sections, F-J) longitudinal sections. A & F) Parachaetetes cf. asvapatii in & , 1936, Paleocene, SW France; B & G) Parachaetetes cf. tornquisti (, 1906), "Middle" Cretaceous, Brazil; C & H) Cordilites cretosus (, 1846), "Middle" Cretaceous, Puerto Rico; D) densely packed filaments of Elianella elegans & , 1948, "Middle" Cretaceous, Brazil; E) loosely arranged filaments of Elianella elegans & , 1948, "Middle" Cretaceous, Brazil ; I) densely packed filaments of Elianella brasiliana n.sp., "Middle" Cretaceous, Brazil; J) loosely arranged filaments of Elianella brasiliana n.sp., "Middle" Cretaceous, Brazil. [All photos with 100 µm scale bar]
In transverse section, the thallus of Parachaetetes tornquisti displays wavy concentric zones (, 1906: Pl. VI, fig. 6a; , 1929: Pl. IV, fig. 1). A darker band corresponds to random cuts of cells at or near the junction of two successive rows and the next lighter bands to random cuts of cells above and below the junction, in the median part of the upper or lower rows. Similar features are also documented in our Parachaetetes cf. asvapatii, as well as in Cordilites cretosus and in Elianella elegans (in Elianella, the zoning is related to changes in the width and density of the cells). Transverse partitions in our Parachaetetes cf. asvapatii are commonly upward-concave, as it is the case for many Elianellaceae (see Solenoporella jurassica (, 1894) documented by & , 1982, for instance); accordingly, this feature is not exclusive of Elianella elegans.
In longitudinal sections, the side walls of the cells in Cordilites cretosus are heavily calcified. On the opposite, secondary pit connections, i.e., connections of the cells of adjacent filaments, or diagenetic alteration of their side walls are commonly observed in our Parachaetetes cf. asvapatii.
Transverse
and longitudinal sections of Cordilites cretosus show the common
occurrence of "septal projection" sensu
(2004) or incomplete walls, tending to split individual cells
longitudinally (Pl. 4 ![]() , figs. B-D). We could not observe any "septal projection" in our
Parachaetetes cf.
asvapatii (but there are possibly some in Elianella elegans at the
level of filament dichotomous branchings: Pl. 2
, figs. B-D). We could not observe any "septal projection" in our
Parachaetetes cf.
asvapatii (but there are possibly some in Elianella elegans at the
level of filament dichotomous branchings: Pl. 2 ![]() , fig. I).
, fig. I).
In contrast, the pattern of the thallus of Elianella elegans is unique. As already stated by its authors ( & , 1948), the filaments of Elianella elegans are moniliform, whereas those of Cordilites cretosus and those of our Parachaetetes cf. asvapatii, when they are well preserved, look like straight-sided polygonal tubes. In addition, most of the time the files of cells of Elianella are not adjacent and consequently their transverse sections are commonly (but not always) rounded. Filaments of the new species, E. brasiliana n.sp., are much thicker with a diameter double that of the type-species, E. elegans. Because cells belonging to close-set filaments and located at the same level do rarely or never share any side wall in both species, and because their thicknesses/lengths significantly vary, their growths were not necessarily simultaneous and the growth rates of neighbouring filaments might have varied too. A filament grew independently from its neighbouring ones. As a consequence, even if the column pattern is still visible in Elianella, a row pattern may be detected only on some occasions.
(2004) pointed out the need for a "comprehensive review" of the artificial
group labelled "Solenoporaceae". Our study, which focuses on some Cretaceous
and Paleocene algal forms, represents just one more step toward a better
knowledge of these fossil organisms ascribed to the Family Elianellaceae nov.
fam. and including Marinella , 1939 ( & ,
2016) and Pycnoporidium & ,
1928, among
other forms. We demonstrate that the three forms considered in this paper, Cordilites
cretosus (, 1846), Elianella elegans & ,
1948, and Parachaetetes cf. asvapatii in
& , 1936, correspond to three discrete species
(Fig.
5 ![]() ). The
genera Cordilites and Parachaetetes share some architectural
patterns with distinct columns and rows whereas rows are present only on
rare occasions in the genus Elianella, which gives it its unique
structure.
). The
genera Cordilites and Parachaetetes share some architectural
patterns with distinct columns and rows whereas rows are present only on
rare occasions in the genus Elianella, which gives it its unique
structure.
In 2015, the first author (B.G.) was the laureate of a Franco-Brazilian professorship at the UNESPetro in Rio Claro (São Paulo State, Brazil) that enabled him to work on Brazilian material. Earlier, in 2012, he also benefited from a Smithsonian Fellowship allowing him to investigate the J. Harlan Collection stored in the premises of the Smithsonian Institution. He would like to thank the staff of the Department of Paleobiology at the Smithsonian National Museum of Natural History, particularly William A. and Jonathan G. for their hospitality and having facilitated his work there. He is also grateful for the support provided by the successive curators of paleontological collections at the Université Pierre et Marie , first Jean-Pierre , then Isabelle . Special thanks go to Sigrid who kindly translated the original German text on Petrophyton miyakoense by (1912). Phil helped polishing the English text. This research is associated with the "Carbonatos do Brasil Project" linked to the Brazilian Sedimentology/Stratigraphy Net sponsored by Petrobras; it was also partly sponsored by the Foundation "Carnets de Géologie".
J. & F. (2001).- Presence of nemathecia in Parachaetetes asvapatii 1936 (Rhodophyta, Gigartinales?): reproduction in 'solenoporaceans' revisited.- Palaeontology, London, vol. 44, no. 6, p. 1113-1125.
R.M. & P. (1986).- Marine Cretaceous algae from the Baratang Formation, Andaman Islands, India.- Geological, Mining and Metallurgical Society of India, Bulletin, Calcuta, no. 54, p. 149-158 (Pls. I-VI).
P. & P.Y. (1982).- Microfossiles et échinodermes incertae sedis des dépôts albiens à coniaciens du bassin de Sergipe-Alagoas, Brésil.- Cahiers de Micropaléontologie, Paris, vol. 3, p. 13-22.
G., J. & A.-F. (1984).- The limestone with Corallinaceae of the Upper Cretaceous of Sümeg (Transdanubia, Hungary): paleogeographic implications.- Acta Geologica Hungarica, Budapest, vol. 27, no. 3-4, p. 429-440 (Pls. I-II).
S. & E. (2005).- Contribution to the recognition of the algal genus Elianella.- Slovak geological Magazine, Bratislava, Roč. 11, č. 4, p. 301-310 (IV Pls.).
R. (1967).- Les algues du Danien de Bastennes-Lonné l (Aquitaine, France Sud-Ouest).- Bulletin du Centre de Recherche Pau - SNPA, Pau, vol. l, no. 1, p. 43-53 (2 Pls.).
K. (1906).- Einige neue Tabulaten und Hydrozoen aus mesozoischen Ablagerungen.- Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Stuttgart, Band I, p. 61-70 (Pls. V-VII).
I., F. & A.-F. (1979).- Testimonianze di Paleocene marino in Sardegna.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 85, no. 2, p. 481-516 (Pls. 45-50).
S. & J.N. (1995).- A new Order (Rhodogorgonales) and Family (Rhodogorgonaceae) of red algae composed of two tropical calciferous genera, Renouxia gen. nov. and Rhodogorgon.- Cryptogamic Botany, Stuttgart, vol. 5, p. 316-331.
B. (2015).- Algas calcárias marinhas bentônicas no Cretáceo do Brasil. In: D. & P. (eds.), Atlas dos Calcários do Cretáceo do Brasil: um Atlas.- IGCE/UNESP/Rio Claro, UNESPetro, Obra 1, p. 518-567.
B., P.-Y. & A.-F. (1991).- Constructions bio-sédimentaires laminées, Lithothamnium et Parachaetetes de la Formation Riachuelo (Albien) du bassin de Sergipe (Nord-Est du Brésil).- Geociências, São Paulo, vol. 10, p. 169-181 (5 Pls.).
B. & D. (2016).- A short note on the fossil alga Marinella lugeoni , 1939, nom. cons., and its seven unfortunate avatars. Revision of the Juliette Collection. Part 2. Revision of the Jesse Harlan Collection. Part 2.- Carnets Geol., Madrid, vol. 16, no. 7, p. 231-245 (4 Pls.).
B., D. & I.I. (2008).- Calcareous algae from Upper Albian Cenomanian strata of the Potiguar basin (NE Brazil).- Geologia Croatica, Zagreb, vol. 61, no. 2-3, p. 139-147 (2 Pls.).
B., P. & L. (2009).- Calcareous algae (and problematica) from the Paleocene of Libya - Part 1. In: 6th Regional Symposium of the International "Fossil Algae" Association, Milan (July 1st-5th).- Museologia Scientifica e Naturalistica, Ferrara, Volume Speciale 2009, p. 33-34.
T.L. & H.S. (1981).- A redescription of the Bathonian red alga Solenopora jurassica from Gloucestershire, with remarks on its preservation.- Palaeontology, London, vol. 25, no. 4, p. 905-912 (Pl. 100).
M. (1966).- Microfacies of the Mesozoic and Tertiary limestones of the West Carpathians.- Slovenská Akadémia Vied, Bratislava, 280 p. (CI Pls.).
E. (1989).- Über die systematische Stellung und die Besystematiche der Solenoporaceen (Rhodophyceae).- Courier Forschungsinstitut Senckenberg, Frankfurt am Main, Band 109, p. 51-91 (6 Pls.).
E. (1929).- Étude de l'algue jurassique Parachaetetes.- Eclogae geologicæ Helvetiæ, Bâle, vol. 22, no. 1, p. 41-43 (Pl. IV).
J. & E. (1948).- Elianella nov. gen. elegans nov. sp., organisme constructeur de calcaires typiquement développé dans le Paléocène du SW malgache.- Bulletin de la Société géologique de France, Paris, (série 5), t. XVII, p. 275-278 (Pl. XII).
J. (1927).- 1. Abteilung: Thallophyta. In: M. (ed.), Handbuch der Paläobotanik.- R. Oldenbourg, München & Berlin, p. 31-136.
J. (1943).- Zur Kenntnis der kalkalgen der sächsisch-böhmischen Kreide.- Sitzungsberichte der Akademie der Wissenschaften, Wien, (Mathematisch-Naturwissenschaftliche Klasse), Abt. I, Band 152, p. 103-114 (Pls. I-IV).
Ph. (1887).- Die Anthozoen der böhmischen Kreideformation.- Abhandlungen des königlich böhmischen Gesellschaft der Wissenschaften, Prag, (Mathematisch-naturwissenschaftliche Classe Nr. 1), Folge VII, Band 2, 60 p. (Pls. I-II).
A.-F. (1969).- Les algues du toit des bauxites des environs de Mazaugues (Var).- Revue de Micropaléontologie, Paris, vol. 11, no. 4, p. 233-240.
A.-F. (1980a).- Sur deux algues calcaires du Crétacé supérieur du Nord de l'Espagne.- C.R. 105e Congrès National des Sociétés savantes, Caen (1980), Section des Sciences, fasc. II, p. 323-328 (Pls. I-II).
A.-F. (1980b).- Réflexions sur le genre Elianella (algue rouge).- 8e RAST, Marseille (1980), p. 286.
A.-F. (1984).- Les algues du Crétacé de Sumeg (Hongrie).- C.R. 111e Congrès National des Sociétés savantes, Dijon (1984), Section des Sciences, fasc. I, p. 427-434.
A.-F. & du S. (1970).- Les algues des formations transgressives maestrichtiennes, paléocènes et yprésiennes de la côte sud-orientale de la Corse.- Revue de Micropaléontologie, Paris, vol. 12, no. 4, p. 202-208 (2 Pls.).
A.-F. & H. (1982).- Les algues de l'Albien supérieur du Nigéria.- Cahiers de Micropaléontologie, Paris, vol. 2, p. 35-40 (1 Pl.).
L.R. & J. (1936).- Fossil algae from the uppermost Cretaceous beds (the Niniyur Group) of the Trichinopoly district, S. India.- Memoirs of the Geological Survey of India, Palæontologia Indica, Calcuta, (New Series), vol. XXI, no. 4, 49 p. (Pls. I-VI).
A.E. (1846).- Die Verteinerungen der böhmischen Kreideformation.- E. Schweizerbart,
Stuttgart, iv + 148 p. (Pls. I-LI).
URL: https://archive.org/stream/dieversteinerung00reus#page/n3/mode/2up
R. (2004).- Solenopora is a chaetetid sponge, not an alga.- Palaeontology, London, vol. 47, no. 1, p. 117-122.
O., K. & E. (1972).- Microfauna and lithostratigraphy of the Paleogene and adjacent Cretaceous of the Middle Váh Valley (West Carpathians).- Geologický ústav Dionýza túra, Bratislava, 246 p. (CLXXX Pls.).
F. (1991).- Allochtone Urgonkalke im Mittleren Abschnitt der Nördlichen Kalkalpen: Fazies, Paläontologie und Paläogeographie.- Münchner Geowissenschaftliche Abhandlungen, Reihe A Geologie und Paläontologie, 20, 120 p. (19 Pls.).
R. (2000).- On the occurrence of Parachaetetes asvapatii , 1936 (Solenoporaceae) in the Montorfano Member type-section (Tabiago Formation, Como, northern Italy).- Revue de Paléobiologie, Genève, vol. 19, no. 2, p. 427-434.
H. (1912).- Ueber Einige Gesteinbildende Kalkalgen von Japan und China.- The Science Reports of the Tohoku Imperial University, Sendai, (2nd Series), vol. 1, no. 1, p. 1-8 (Pls. I-II).
H. & S. (1928).- On some rock-forming algae from the younger Mesozoic of Japan.- The Science Reports of the Tohoku Imperial University, Sendai, (2nd Series), vol. 12, no. 1, p. 141-152 (Pls. XVIII-XXIII).

Click on thumbnail to enlarge the image.
Plate 1:
A-F) Elianella
elegans & ,
1948. Thin section "37
EB16", "NW de Lohaosy", "Calc. à Algues", Juliette
Collection; Paleocene; S Madagascar.
A-D)
details of the neotype (Fig. 1.B) with longitudinal sections of filaments
(1990's analog photography); E-F) details of the neotype (Fig. 1.B) with
longitudinal sections of filaments. B is a detail of Fig. 1.C; C is a detail of
E; D is a detail of F.
[All
photos same scale bar 500 µm]

Click on thumbnail to enlarge the image.
Plate 2:
A, D, F
& H-I) Elianella elegans & ,
1948.
Albian, Riachuelo Formation; Sergipe, Brazil.
A) longitudinal sections (Pl. 6, upper left figure in ,
2015);
(b) worm boring; (arrow) filament branching. Thin section UPAFSE 047a, SE 18;
D)
longitudinal sections with filament branchings (arrows). Thin section UPAFSE
037, SE 18;
F)
oblique to transverse sections. Thin section UPAFSE_069o, SE 19;
H-I)
transverse section displaying a zoned pattern; I is a detail of H. Thin section
UPAFSE_046, SE 18;
I)
Longitudinal parting of some cells; (paired arrows) longitudinal partitions are
a right angle with the cell side walls.
B-C, E
& G) Elianella brasiliana n.sp. Albian, Riachuelo Formation;
Sergipe, Brazil.
B-C)
holotype with longitudinal sections; C is a detail of B; (arrow) filament
branching;
E)
longitudinal sections with variable cell's heights (Pl. 7, lower left figure in
, 2015);
G)
paratype with transverse sections (Pl. 7, lower left figure in ,
2015);
B-C
& G) Thin section UPAFSE 049a, SE 7; G) thin section UPAFSE 067c, SE 19.
[A-B,
D-F & H scale bar 500 µm; C, G & I scale bar 250 µm]
Click on thumbnail to enlarge the image.
Plate 3:
A-D) Parachaetetes
cf. asvapatii in & ,
1936.
Danian; interval 223.03-233.53 m, SNPA exploration well Bastennes-Lonné l - FB1
(Aquitaine, SW France).
A) longitudinal sections with variably preserved
filaments (long cells) in the core of a thallus;
B)
longitudinal sections with variably preserved filaments (short cells) near the
surface of a thallus;
C)
transverse sections with polygonal arrangement of the cells;
D)
transverse section displaying a zoned pattern.
A) Thin
section Bastennes 1 (3) 223.03-233.53 m; B-C) thin section Bastennes 1 (5)
223.03-233.53 m ; D) thin section Bastennes 1 223.03-233.53 m.
[A-C
scale bar 250 µm; D scale bar 500 µm]

Click on thumbnail to enlarge the image.
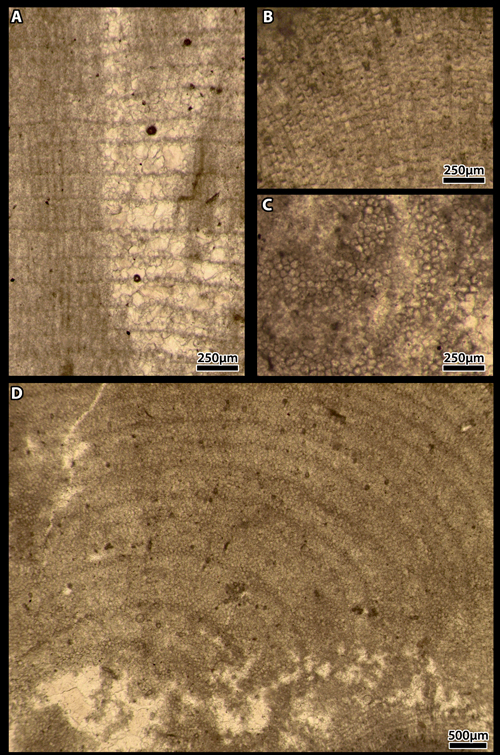
Plate 4:
A-I) Cordilites
cretosus (, 1846). "Middle" Cretaceous; Puerto Rico.
A-C) transverse sections with polygonal arrangement of the
cells. B-C are details of A;
B-C)
(paired arrows) double-headed arrows are the mark of the longitudinal parting of
cells. The longitudinal partition forms a right angle with the side walls;
D)
longitudinal section with incomplete longitudinal partings of some cells;
E &
G) longitudinal section with one row (arrows) varying laterally in thickness. G
is a detail of E;
F)
longitudinal section with thicker calcification of cell's walls in some rows;
H)
transverse section displaying a zoned pattern;
I)
abrupt transition from transverse to longitudinal section, corresponding to the
superposition of two thalli.
A-D
& H-I) Thin section D619a630a, Smithsonian; E-G) thin section D619a630a,
Smithsonian.
[E &
H-I scale bar 250 µm; A & F-G scale bar 100 µm; B-C scale bar 50 µm]