◄ Carnets Geol. 16 (5) ►
![]()
Contents
[I. Introduction]
[II. Phylogeny]
[III. Systematic paleontology]
[IV. Phylogenetic relationships ...] [V. Stratigraphical and paleogeographical distribution] [VI. Conclusions]
[Bibliographic references] [Plates]
and ... [Appendix]
16, rue Frédéric Mistral, 26200 Montélimar (France)
Centre d'Études Méditerranéennes, 04170 St-André les Alpes (France)
Département des Sciences de la Terre, Université de Nice-Sophia-Antipolis,
Faculté des Sciences, 28 avenue Valrose, 06108 Nice Cedex 2 (France)
Centre d'Études Méditerranéennes, 04170 St-André les Alpes (France)
Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad
Universitaria, Coyoacán, 04510 México, D.F. (Mexico)
Institute of Earth Sciences, Bâtiment Géopolis, 1015 Lausanne (Suisse)
Grand rue, 04330 Barręme (France)
"Lou Seren", 04300 Forcalquier (France)
Chemin de Dardčne, La Barthelasse, 84000 Avignon (France)
Centre d'Études Méditerranéennes, 04170 St-André les Alpes (France)
57 bis, avenue des Patriotes, 26300 Bourg-de-Péage (France)
Centre d'Études Méditerranéennes, 04170 St-André les Alpes (France)
57 bis, avenue des Patriotes, 26300 Bourg-de-Péage (France)
Centre d'Études Méditerranéennes, 04170 St-André les Alpes (France)
Published online in final form (pdf) on April 14, 2016
[Editor: Bruno ;
language editor: Stephen ]
![]()
In spite of recent interest, the various species belonging to the genera Heminautilus , 1927, and Josanautilus & , 2006, have never been the object of a general revision. More than 160 specimens belonging to various species of these two genera were studied, with the aim of identifying the specific characteristics and the stratigraphical and paleogeographical distribution of the various species which compose them, as well as to propose a phylogeny for the group. This study confirms the non-synonymy of Heminautilus saxbii (, 1848) and H. lallierianus (, 1841), establishes that H. tejeriensis & , 2006, and H. verneuilli (, 1870) represent junior synonyms of H. saxbii () and suggests the creation of the species H. ? japonicus sp. nov. The origin of the genus Heminautilus from Pseudocenoceras , 1927, proposed by and the origin of Josanautilus & from Heminautilus are both confirmed.
Nautiloidea; Cenoceratidae; Heminautilus; Josanautilus; Early Cretaceous; Barremian; Aptian.
C., G., J.A., A., J., G. , R., P. & M. (2016).- Revision of the Early Cretaceous genera Heminautilus , 1927, and Josanautilus & , 2006 (Nautilida, Cenoceratidae).- Carnets Geol., Madrid, vol. 16, no. 5, p. 61-212.
Révision des genres Heminautilus , 1927, et Josanautilus & , 2006 (Nautilida, Cenoceratidae), du Crétacé inférieur.- Malgré des travaux récents, les différentes espčces appartenant aux genres Heminautilus , 1927, et Josanautilus & , 2006, n'ont jamais fait l'objet d'une révision d'ensemble. Plus de 160 spécimens appartenant ŕ différentes espčces de ces deux genres ont été étudiés dans le but de cerner les caractčres spécifiques et la répartition stratigraphique et paléogéographique des différentes espčces qui les composent, ainsi que de proposer un schéma phylogénétique de ce groupe. Cette étude a permis en particulier de confirmer la non-synonymie d'Heminautilus saxbii (, 1848) et d'H. lallierianus (, 1841), d'établir qu'H. tejeriensis & , 2006, et H. verneuilli (, 1870) représentent des synonymes juniors d'H. saxbii (), et de créer l'espčce H. ? japonicus sp. nov. L'origine du genre Heminautilus chez Pseudocenoceras , 1927, proposée par , a pu ętre confortée et l'origine de Josanautilus & chez Heminautilus a également pu ętre confirmée.
Nautiloidea ; Cenoceratidae ; Heminautilus ; Josanautilus ; Crétacé inférieur ; Barrémien ; Aptien.
The genus Heminautilus , 1927, includes forms present in the Early Cretaceous (Early Barremian to Early Aptian), with very distinctive and easily recognizable morphology within the Nautiloidea , 1825. Its various representatives share a relatively compressed section, a strongly differentiated ventral region and a sinuous suture line, with a particularly well developed ventral lobe. For a long time the genus was poorly known and, in numerous works (F.-J. & , 1858; , 1916; , 1967; & , 1976; et al., 1984; & , 1995; , 2006; , 2008), its European and North African representatives were attributed without distinction to the species H. lallierianus (, 1841) and H. saxbii (, 1848), sometimes in spite of characteristics that clearly separate them from the type specimens. The recent works of (1980, 1985, 1989, 2007, 2010), & (2006) and et al. (2012) contributed to a better knowledge of the genus, in particular with the creation of the species H. sanctaecrucis , 1980, H. tejeriensis & , 2006, and H. boselliorum et al., 2012. Furthermore, & recently erected the monospecific genus Josanautilus & , 2006, for the species Nautilus lacerdae , 1870, phylogenetically very close to the genus Heminautilus . However, no global revision of these two genera has been undertaken until now. The object of the present work is to clarify the diagnostic characteristics of the various species and to summarise their geographical and stratigraphical distribution as well as the phyletic links between them.
We also look at the possible synonymy between the species H. lallierianus (, 1841) and H. saxbii (, 1848), that has been suggested by numerous authors (, 1850; F.-J. & , 1858; , 1916; , 1943).
(1927) is the first author to propose a classification of Nautiloidea, based purely on morphological criteria. Thus he classifies the genus Heminautilus , 1927, in the Family Paracenoceratidae , 1927, which gathers all the forms with a differentiated ventral region and a suture line containing a well developed ventral lobe. However, (1927, p. 25) himself recognizes the polyphyletic character of this family. The works of ( & , 1983; , 1993) will later show that this classification does not actually take into account phyletic data and that numerous identical morphological characteristics appear in an iterative way in various lineages. The classification of was used, however, by (1984, p. 181) who presented a phylogenetic outline of the Family Paracenoceratidae .
In 1956, briefly returns to the classification proposed by (1927). He keeps most of the families proposed by the latter but reducing them to the rank of subfamily. However, he moves the genus Heminautilus , 1927, into the Subfamily Cymatoceratinae , 1927. Indeed, he notes (p. 435) that the suture line of the genus Heminautilus is more sinuous than that of the other representatives of the Paracenoceratidae , 1927, and that some specimens of Heminautilus [specimens attributed to Nautilus lallieri , 1841, by , 1916, Pl. 17, figs. 2-4, 6; Heminautilus rangei (, 1922)] possess sinuous ribbing close to that observed on the genus Cymatoceras , 1884, while other representatives of the Paracenoceratidae show no ribbing.
However, in a recent work, & (2006, p. 62) showed that the structure of the shell as well as the ribbing observed on the specimens of Heminautilus illustrated by (1916, Pl. 17, figs. 2-4, 6) are very different from that observed on Cymatoceras , and that consequently there is no direct phyletic link between these two genera.
In various papers ( & , 1983, 1985; , 1980, 1989, 1993), established a new classification based on phyletic data rather than strictly morphological. Thus, he established (1993) that characteristics such as the presence of ribbing, a flat or concave ventral region, or even a sinuous suture line, appear in an iterative way in very different lineages. These features are interpreted as an adaptive answer to a particular environment and are not systematically considered as the consequence of a phyletic link. To establish his new classification, used the usual specific characters (ornamentation, suture line, section and ventral region shape, position of the siphon) by interpreting them either as the indication of a phyletic link or as an iterative homeomorphy, in particular for stratigraphically distant species. For & (1983), the genus Heminautilus , 1927, derives from the genus Pseudocenoceras , 1927, by the appearance of a sinuous suture line and a concave ventral region. However, this last genus, in the conception of (1956), seems polyphyletic: it contains various species distributed from the Berriasian to the Maastrichtian. In the Early Cretaceous, the only known species are Pseudocenoceras berriasensis (F.-J. , 1867) from the Berriasian of southeast France, Pseudocenoceras campichei (, 1907) from the Hauterivian? of Crimea and Pseudocenoceras picteti (, 1907) from the Early Barremian? of Crimea. No representative of the genus Pseudocenoceras seems to be known in the Valanginian, Aptian and Albian; in the Cenomanian the genus is again present, particularly in the form of its type species P. largilliertianus (, 1841). It seems clear that for the genus Heminautilus derives from the group formed by the Early Cretaceous taxa P. berriasensis (F.-J. ), P. campichei () and P. picteti (), even if the phyletic links amongst these three species remain to be specified.
By considering the morphological characteristics and the stratigraphical position of the various representatives of the genus Heminautilus , 1927, another hypothesis on the phyletic position of this genus can be envisaged. Indeed, the genus Xenocheilus & , 1955, present from the Berriasian to the Hauterivian, even to the Early Barremian, shows morphological characteristics close to Heminautilus . According to (1975), followed by & (1983), this genus belongs to the Family Pseudonautilidae & , 1955, close to the genera Pseudonautilus , 1876, and Aulaconautilus , 1927, which would indicate that all three derive from the Jurassic genus Pseudaganides , 1927.
The
genus Xenocheilus & , and in
particular the taxon Xenocheilus ulixis
& , 1955, from the Hauterivien of Crimea
(see Pl. I ![]() , fig. 1), have a very involute coiling with a flat or slightly
concave ventral region, and a suture line with a deep lateral lobe and a well
indented ventral lobe. As observed by (1984, p. 180-181), it would
be possible to postulate here the origin of the first representatives of the
genus Heminautilus , which
could derive from Xenocheilus ulixis
& by a slight modification of the sutural line (lateral lobe
shallower, wider and round in Heminautilus
) and a reduction of the width of the ventral region. However, the
position of the siphuncle is a major obstacle to the hypothesis of a direct
ancestor-descendant relationship between Xenocheilus
and Heminautilus, because in Heminautilus
the siphuncle is always situated in the lower half of the section
while it is close to the ventral region in Xenocheilus
& and in all the Pseudonautilidae
& . These ornamental and sutural similarities were previously
noted by (1975) who considered them due to morphological
convergence.
, fig. 1), have a very involute coiling with a flat or slightly
concave ventral region, and a suture line with a deep lateral lobe and a well
indented ventral lobe. As observed by (1984, p. 180-181), it would
be possible to postulate here the origin of the first representatives of the
genus Heminautilus , which
could derive from Xenocheilus ulixis
& by a slight modification of the sutural line (lateral lobe
shallower, wider and round in Heminautilus
) and a reduction of the width of the ventral region. However, the
position of the siphuncle is a major obstacle to the hypothesis of a direct
ancestor-descendant relationship between Xenocheilus
and Heminautilus, because in Heminautilus
the siphuncle is always situated in the lower half of the section
while it is close to the ventral region in Xenocheilus
& and in all the Pseudonautilidae
& . These ornamental and sutural similarities were previously
noted by (1975) who considered them due to morphological
convergence.
The
systematic position adopted by ( & ,
1983; , 1993), who considers that the genus Heminautilus
, 1927, comes from Pseudocenoceras
, 1927, corresponds best with the current data, and consequently the
genus Heminautilus is
here placed within the Family Cenoceratidae
& , 1983. The main characteristics of the genus
Pseudocenoceras , 1927, are
a wide and flattened ventral region, a subrectangular section, a slightly
sinuous suture line and the siphuncle in a subcentral position, closer to the
dorsal face than to the ventral region. In the hypothesis of , the
genus Heminautilus would
thus derive from Pseudocenoceras
by the appearance of a sinuous suture line developing a strong lateral lobe, and
of a ventral region with angular edges, as well as by a slight lower position of
the siphuncle on the section. From this perspective, the taxon Pseudocenoceras
campichei (, 1907), from the Hauterivian? of Crimea
(Pl. I ![]() ,
fig. 2), seems to be the best candidate for the origin of the genus Heminautilus
, both morphologically and stratigraphically. However, the current
data on these two genera and, in particular, on possible specimens from the Late
Hauterivian - Early Barremian remain too fragmentary to provide confirmation,
and the hypothesis of needs to be supported by a more plentiful
material.
,
fig. 2), seems to be the best candidate for the origin of the genus Heminautilus
, both morphologically and stratigraphically. However, the current
data on these two genera and, in particular, on possible specimens from the Late
Hauterivian - Early Barremian remain too fragmentary to provide confirmation,
and the hypothesis of needs to be supported by a more plentiful
material.
The monospecific genus Josanautilus & , 2006, based on its type species Nautilus lacerdae , 1870, is a taxon known only from the Early Aptian of Spain and southeast France. Its ornamental and sutural characteristics, as well as its stratigraphical position, show that this species is very close to the genus Heminautilus , 1927, as noted by (1975, p. 124) and & (2006, p. 17). Furthermore, both genera seem to have similar environmental requirements and are generally present only in the outer platform domain; the genus Heminautilus is generally associated with Josanautilus & in deposits where the latter is present. They have in common a similar sutural line, a siphuncle placed in the lower half of the section, a flat ventral region, a trapezoidal section and very involute coiling. The ribbing on the flanks of the body chamber in Josanautilus & is also similar to that observable on some ornamented forms of the genus Heminautilus . The essential difference between the genera is the presence in Josanautilus & of a wider and slightly convex ventral region, with a strong central keel surrounded by one or two longitudinal ribs. These characteristics show clearly that Josanautilus & comes from the genus Heminautilus , probably by way of the taxon Heminautilus sanctaecrucis , 1980, whose ventral region shows a median keel.
On
the other hand, Josanautilus lacerdae
(, 1870) shows morphological similarities with the Berriasian
species Aulaconautilus sexcarinatus
(F.-J. , 1867), the cast of whose holotype is shown here
(Pl. II ![]() ,
fig. 1). Indeed, this last taxon is characterized by a sinuous suture line, as
well as by the presence of six or eight longitudinal ribs on a wide and
flattened ventral region. However Aulaconautilus sexcarinatus
(F.-J. ) differs from Josanautilus
lacerdae () by clear differences in the suture line
which shows a far less developed lateral saddle and a narrower and less round
lateral lobe. The ornamentation of the ventral region of J. lacerdae ()
also presents only superficial similarities to that of A. sexcarinatus
(F.-J. ) and can be distinguished easily by the presence of a
median keel and the absence of longitudinal ribs on the phragmocone.
Furthermore, no ribbing is visible on the flanks in A. sexcarinatus
(F.-J. ); the section of the latter is clearly narrower, and
subrectangular instead of trapezoidal as in J.
lacerdae (); and the
whorls' height increases more slowly than in J. lacerdae ().
All of these characteristics, associated with a very different stratigraphical
position [Berriasian for A. sexcarinatus (F.-J. ) and Early Aptian for J. lacerdae ()],
show that the similarities between these taxa represent only homeomorphic
characteristics and not the consequence of a phyletic link, as noted by
& (2006, p. 17).
,
fig. 1). Indeed, this last taxon is characterized by a sinuous suture line, as
well as by the presence of six or eight longitudinal ribs on a wide and
flattened ventral region. However Aulaconautilus sexcarinatus
(F.-J. ) differs from Josanautilus
lacerdae () by clear differences in the suture line
which shows a far less developed lateral saddle and a narrower and less round
lateral lobe. The ornamentation of the ventral region of J. lacerdae ()
also presents only superficial similarities to that of A. sexcarinatus
(F.-J. ) and can be distinguished easily by the presence of a
median keel and the absence of longitudinal ribs on the phragmocone.
Furthermore, no ribbing is visible on the flanks in A. sexcarinatus
(F.-J. ); the section of the latter is clearly narrower, and
subrectangular instead of trapezoidal as in J.
lacerdae (); and the
whorls' height increases more slowly than in J. lacerdae ().
All of these characteristics, associated with a very different stratigraphical
position [Berriasian for A. sexcarinatus (F.-J. ) and Early Aptian for J. lacerdae ()],
show that the similarities between these taxa represent only homeomorphic
characteristics and not the consequence of a phyletic link, as noted by
& (2006, p. 17).
Methodology
and conventions:
the measurements of height (H),
thickness of the whorls (E) and width
of the ventral region (M) were made at
various diameters (D) (Fig. 1 ![]() ). The
measurements of the variation of the lateral lobe of the suture line were made
according to the method of &
(1971): L
indicates the width of the lateral lobe and P
indicates its depth (Fig. 1
). The
measurements of the variation of the lateral lobe of the suture line were made
according to the method of &
(1971): L
indicates the width of the lateral lobe and P
indicates its depth (Fig. 1 ![]() ). The measurements indicated in bold and italic type
were made from photos and are thus subject to some uncertainty (Tables 2-14). To
simplify the text, repositories of the studied or quoted specimens are
abbreviated (Table 1). The biozonation of the Early Cretaceous used here (Fig.
2
). The measurements indicated in bold and italic type
were made from photos and are thus subject to some uncertainty (Tables 2-14). To
simplify the text, repositories of the studied or quoted specimens are
abbreviated (Table 1). The biozonation of the Early Cretaceous used here (Fig.
2 ![]() )
is the one developed by the IUGS Lower Cretaceous Ammonite Working Group (
et al., 2011, 2014), for the
North-Tethyan Domain, with the exception of the Imerites giraudi Zone which is split into three subzones at the top
of the Barremian ( et al., 2008, and et al.,
2011). It is correlated for the end of the Early Aptian with the biozonation
developed by
et al. (2015) for Mexico and southern North America and the one
developed by
(1979) for Colombia.
)
is the one developed by the IUGS Lower Cretaceous Ammonite Working Group (
et al., 2011, 2014), for the
North-Tethyan Domain, with the exception of the Imerites giraudi Zone which is split into three subzones at the top
of the Barremian ( et al., 2008, and et al.,
2011). It is correlated for the end of the Early Aptian with the biozonation
developed by
et al. (2015) for Mexico and southern North America and the one
developed by
(1979) for Colombia.
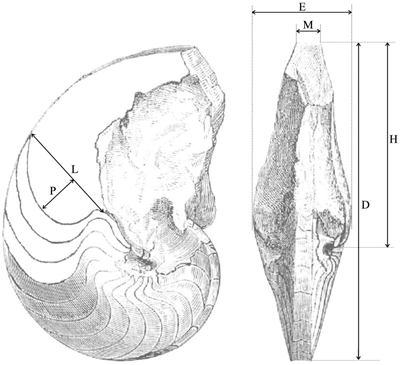
Click on thumbnail to enlarge the image.
Figure 1: Explanatory scheme for the measurements made on the studied specimens.
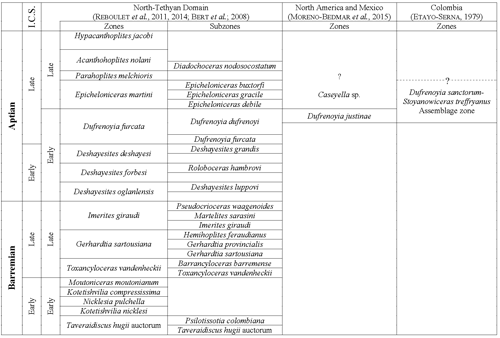
Click on thumbnail to enlarge the image.
Figure 2: Biozonation of the Barremian and Aptian (Early Cretaceous) used in this work.
Table 1: List of abbreviations used in the text.
| AMNH | American Museum of Natural History, New York, United States |
| BEG | Bureau of Economic Geology, Austin, Texas, United States |
| BMNH | British Museum of National History, England |
| EM | École des Mines, Claude University, Lyon, France |
| FSM | Faculté des Sciences of Marseille, France |
| GK | Kyushu University, Japan |
| GMMU | Geological Museum of Mansoura University, Egypt |
| GSUB | Geosciences Collection of the University of Bremen, Germany |
| HNHM | Hungarian Natural history Museum |
| IGM | Instituto de Geología, Universidad Nacional Autónoma de México |
| MB | Natural History Museum of Bâle, Switzerland |
| MCGL | Muséum Géologique Cantonal of Lausanne, Switzerland |
| MGB | Museu de Geologia de Barcelona, Spain |
| MGNR | Museo Geológico Nacional J. Royo y Gómez, Bogotá, Colombia |
| MHNA | Natural History Museum of Auxerre, France |
| MHNG | Natural History Museum of Geneva, Switzerland |
| MHNN | Natural History Museum of Neuchâtel, Switzerland |
| MNCN | Museo Nacional de Ciencias Naturales, Madrid, Spain |
| MNHN | Muséum National d’Histoire Naturelle of Paris, France |
| MPL | Museo Paleontológico Laguna, Torreón, Coahuila State, Mexico |
| MPUC | Museum of Paleontology, California University, United States |
| MPUS | Museum of Paleontology, Sofia University, Bulgaria |
| MV | Museum of Valltorta, Spain |
| NMNS | National Science Museum, Tokyo, Japan |
| PNRL | Parc Naturel Régional du Lubéron, France |
| PUAB | Universitat Autňnoma de Barcelona, Spain |
| UNCB | Colecciones Paleontológicas, Departamento de Geociencias, Universidad Nacional de Colombia, Bogotá |
| UPMC | Pierre et Marie University, Paris, France |
| USNM | United States National Museum, Washington, United States |
Order Nautilida , 1847
Superfamily Nautiloidea , 1825
Family Cenoceratidae & , 1983
Genus Heminautilus , 1927
(= Vorticoceras , 1940; ? Platynautilus & , 1953)
Type species: Nautilus saxbii , 1848 [by original designation of (1927, p. 22)].
Discussion: the genus Vorticoceras , 1940, is a synonym of Heminautilus , 1927, as established previously (1943) and (1956). The genus Platynautilus & , 1953, is probably also a synonym of Heminautilus (see discussion in section V). The genus Heminautilus includes the species H. boselliorum et al., 2012, H. rangei (, 1922), H. sanctaecrucis , 1980, H. saxbii (, 1848) [= H. verneuilli (, 1870); H. tejeriensis & , 2006], H. lallierianus (, 1841), H. etheringtoni , 1946, H. stantoni (, 1940), H. ? japonicus sp. nov. and H. ? tyosiensis ( & , 1953); its known stratigraphical distribution extends from the Early Barremian, Kotetishvilia nicklesi Zone, to the top of the Early Aptian, Dufrenoyia furcata Zone. It is present in Europe (Bulgaria, England, France, Hungary, Spain and Switzerland), North Africa (Egypt and Tunisia), Ethiopia, Japan, North America (USA [Arkansas], Mexico [Puebla and Durango states]), and northern South America (Colombia and Venezuela).
The
taxon Heminautilus akatsui ,
1980 (in et
al.,
1980), from the Early Albian of Japan, shows an unusual stratigraphical
position and ornamental and sutural characteristics very different from those
classically present in the genus Heminautilus
and in particular in its type species; it certainly belongs to a
different genus. Its holotype is featured in this work (Pl. II ![]() , fig. 2).
, fig. 2).
Heminautilus boselliorum et al., 2012
Pl. II ![]() , fig. 3;
Pl. III
, fig. 3;
Pl. III ![]() ,
figs. 1-3;
Pl. IV
,
figs. 1-3;
Pl. IV ![]() ,
figs. 1-3;
,
figs. 1-3;
Pl. V ![]() ,
figs. 1-2;
Pl. VI
,
figs. 1-2;
Pl. VI ![]() ,
figs. 1-2;
Pl. VII
,
figs. 1-2;
Pl. VII ![]() ,
figs. 1-2, 4;
,
figs. 1-2, 4;
Pl. VIII ![]() ,
figs. 1-3;
Pl. IX
,
figs. 1-3;
Pl. IX ![]() ,
figs. 1-3
,
figs. 1-3
|
? |
Heminautilus sp. - , Pl. 27, fig. 4. |
|
|
pars |
Heminautilus saxbii (, 1848) - & , p. 65: no. K1 1693, K1 1694, non no. K1 6005, Pl. 5, fig. 1, 1a (= ? H. sanctaecrucis , 1980). |
|
|
Heminautilus aff. saxbii (, 1848) - , p. 37. |
||
|
Heminautilus boselliorum sp. nov. - et al., p. 157, Pl. 1, fig. 1; Pl. 2, figs. 1-2; Pl. 3, figs. 1-2; Pl. 4, fig. 2; Pl. 8, fig. 2; Pl. 9, fig. 2; Pl. 11, fig. 2; Pl. 12, fig. 1. |
||
|
Heminautilus sp. - & , p. 183. |
Material studied (N = 13): specimens no. Mej01a, Mej02, Mej03, Mej11, coll. , no. RG2000, RG2001, RG2002a, b, coll. , no. LUS01, LUS02, coll. , no. MNHN.F.A52065, Early Barremian, Nicklesia pulchella Zone, La Lčque, Lussan (Gard, France); specimen no. MHNG GEPI 15935, Nicklesia pulchella Zone?, Escragnolles (Alpes-Maritimes, France); specimen no. 415950, coll. , upper part of the Kotetishvilia nicklesi Zone, section no. 833, Comps-sur-Artuby (Var).
Type: the holotype (by original designation) is the specimen no. Mej01a, coll. , presented by et al. (2012, Pl. 1, fig. 1a-c); a cast is deposited in the paleontology collections of the Muséum National d'Histoire Naturelle of Paris with the no. MNHN.F.A52064.
Geographical distribution: the species is known from France (departments of the Gard, the Var and the Alpes-Maritimes), Bulgaria and Hungary.
Stratigraphical
distribution:
Heminautilus boselliorum et
al., 2012, is present in the Early Barremian, upper part of the Kotetishvilia nicklesi Zone and Nicklesia
pulchella Zone of southeast France, and in the lower part of the Kotetishvilia
compressissima Zone in Hungary (specimen no. M 2002.554, coll. HNHM,
Pl. IX ![]() ,
fig. 2, bed 126 of the Bersek quarry, Gerecse Mountains;
& , 2009). The
Bulgarian specimens (coll. MPUS) no. K1 1693 (Pl. VIII
,
fig. 2, bed 126 of the Bersek quarry, Gerecse Mountains;
& , 2009). The
Bulgarian specimens (coll. MPUS) no. K1 1693 (Pl. VIII ![]() , fig. 1;
Pl. IX
, fig. 1;
Pl. IX ![]() ,
fig. 1) and K1 1694 (Pl. VIII
,
fig. 1) and K1 1694 (Pl. VIII ![]() , fig. 2) (studied by & ,
1995, p. 65) come from the Early Barremian without more precision. The specimen
no. MHNG
GEPI 15935 (Pl. VII
, fig. 2) (studied by & ,
1995, p. 65) come from the Early Barremian without more precision. The specimen
no. MHNG
GEPI 15935 (Pl. VII ![]() , fig. 1;
Pl. IX
, fig. 1;
Pl. IX ![]() , fig. 3) from Escragnolles (Alpes-Maritimes,
France) is assumed to come from the Nicklesia pulchella Zone based upon the character of its matrix.
, fig. 3) from Escragnolles (Alpes-Maritimes,
France) is assumed to come from the Nicklesia pulchella Zone based upon the character of its matrix.
Dimensions (in mm): see Table 2.
Table 2: Measurements of Heminautilus boselliorum et al., 2012.
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. Mej01a | 156 | 89.4 | 56.3 | 0.57 | 0.36 | 0.63 | 15 | 0.1 | 0.17 | - | - | - | - |
| 141.9 | 85.5 | 48.7 | 0.6 | 0.34 | 0.57 | 14 | 0.1 | 0.16 | - | - | - | - | |
| 122.2 | 73.3 | 36.5 | 0.6 | 0.3 | 0.5 | 12.3 | 0.1 | 0.17 | 16 | 56 | 0.29 | 0.76 | |
| 113.1 | 67 | 33.2 | 0.59 | 0.29 | 0.5 | 10.7 | 0.09 | 0.16 | 16 | 51 | 0.31 | 0.76 | |
| - | 50 | 26.1 | - | - | 0.52 | 8.6 | - | 0.17 | 14 | 35 | 0.4 | 0.7 | |
| - | 43.5 | 24.1 | - | - | 0.55 | 8.5 | - | 0.2 | 9.4 | 33 | 0.28 | 0.76 | |
| no. Mej02 | 139.3 | 74.4 | 44.5 | 0.53 | 0.32 | 0.6 | 12.4 | 0.09 | 0.17 | - | - | - | - |
| - | 54 | - | - | - | - | 8 | - | 0.15 | 12 | 41 | 0.29 | 0.76 | |
| no. Mej03 | 133.5 | c80 | 34 | 0.6 | 0.25 | 0.43 | 12.5 | 0.09 | 0.16 | - | - | - | - |
| 102.8 | c58 | 29.5 | 0.56 | 0.29 | 0.51 | 8.7 | 0.08 | 0.15 | 16.3 | 47 | 0.35 | 0.81 | |
| 95.2 | 53.8 | 29.3 | 0.57 | 0.31 | 0.54 | 8.2 | 0.09 | 0.15 | 15.5 | 45 | 0.34 | 0.84 | |
| no. Mej11 | - | 89.7 | - | - | - | - | 16.3 | - | 0.18 | - | - | - | - |
| - | 70.1 | 37.9 | - | - | 0.54 | 13.1 | - | 0.19 | 11.1 | 58.3 | 0.19 | 0.83 | |
| no. RG2000 | 131.3 | 78.2 | - | 0.6 | - | - | 12.4 | 0.09 | 0.16 | - | - | - | - |
| 112.3 | 71.2 | - | 0.63 | - | - | 11.3 | 0.1 | 0.16 | 14.9 | 52.2 | 0.29 | 0.73 | |
| 98.2 | 61.9 | - | 0.63 | - | - | 8.7 | 0.09 | 0.14 | 16.6 | 48.2 | 0.34 | 0.78 | |
| no. RG2001 | 148.8 | 87.4 | 46.6 | 0.59 | 0.31 | 0.53 | - | - | - | - | - | - | - |
| 134.3 | 82.1 | c32.6 | 0.61 | 0.24 | 0.4 | c10.2 | 0.08 | 0.12 | 18.8 | 59.6 | 0.32 | 0.73 | |
| - | 52.4 | c22.8 | - | - | 0.44 | c4.5 | - | 0.09 | 9.4 | 40.4 | 0.23 | 0.77 | |
| no. RG2002a | 155.8 | 92.8 | 55.5 | 0.6 | 0.36 | 0.6 | 14.2 | 0.09 | 0.15 | - | - | - | - |
| - | 71 | 40.9 | - | - | 0.58 | 12.3 | - | 0.17 | 19.2 | 59.7 | 0.32 | 0.84 | |
| - | 63.3 | 33.2 | - | - | 0.52 | 11 | - | 0.17 | 18.2 | 51.4 | 0.35 | 0.81 | |
| no. RG2002b | 153 | 82.2 | c54.7 | 0.54 | 0.36 | 0.67 | 21.1 | 0.14 | 0.26 | - | - | - | - |
| 132 | 82.1 | c41.2 | 0.62 | 0.31 | 0.5 | 14.2 | 0.11 | 0.17 | 15.8 | 60.7 | 0.26 | 0.74 | |
| no. MNHN.F.A52065 | 65.7 | c41 | 24.5 | 0.62 | 0.37 | 0.6 | 5 | 0.08 | 0.12 | - | - | - | - |
| no. LUS01 | 148.7 | 83.8 | 45.9 | 0.56 | 0.31 | 0.55 | 18.5 | 0.12 | 0.22 | - | - | - | - |
| 108.9 | 69 | 30.4 | 0.63 | 0.28 | 0.44 | 9.7 | 0.09 | 0.14 | 11.7 | 52.8 | 0.22 | 0.77 | |
| 92 | 58.9 | - | 0.64 | - | - | 7.6 | 0.08 | 0.13 | 11.8 | 44.7 | 0.26 | 0.76 | |
| no. LUS02 | 68.3 | 41.2 | 21.7 | 0.6 | 0.32 | 0.53 | 7.6 | 0.11 | 0.18 | 9.1 | 30.1 | 0.3 | 0.73 |
| - | 26.5 | 16 | - | - | 0.6 | 4.4 | - | 0.17 | 5.1 | 20.5 | 0.25 | 0.77 | |
| no. MHNG GEPI 15935 | 128 | 81 | - | 0.63 | - | - | 15 | 0.12 | 0.19 | - | - | - | - |
| - | 78 | - | - | - | - | 13.5 | - | 0.17 | 15 | 54 | 0.28 | 0.69 | |
| - | 65 | - | - | - | - | 9 | - | 0.14 | 14 | 46.5 | 0.3 | 0.72 | |
| no. 415950 | 83.5 | 47.3 | 26.1 | 0.57 | 0.31 | 0.55 | 7.4 | 0.09 | 0.16 | - | - | - | - |
| 74.5 | 43.5 | 22.1 | 0.58 | 0.3 | 0.51 | 6.5 | 0.09 | 0.15 | 11.3 | 34.1 | 0.33 | 0.78 | |
| - | 28.6 | 19.6 | - | - | 0.69 | 5.3 | - | 0.19 | 8.8 | 22.6 | 0.39 | 0.79 | |
| no. K1 1693 | - | 85.6 | 43.4 | - | - | 0.51 | 13.5 | - | 0.16 | - | - | - | - |
| - | 74.4 | - | - | - | - | 11.5 | - | 0.15 | 14.9 | 54.9 | 0.27 | 0.74 | |
| - | 54.1 | - | - | - | - | - | - | - | 13.3 | 43.3 | 0.31 | 0.8 | |
| no. K1 1694 | - | 30.4 | 17.9 | - | - | 0.59 | 6.3 | - | 0.21 | 5.9 | 22.7 | 0.26 | 0.75 |
| - | 27.9 | - | - | - | - | 5.2 | - | 0.19 | 6 | 20.8 | 0.29 | 0.75 | |
| - | 23.3 | - | - | - | - | - | - | - | 5.5 | 18.7 | 0.29 | 0.8 | |
| no. M 2002.554 | 104.1 | 60.9 | - | 0.59 | - | - | - | - | - | 14.9 | 44.5 | 0.33 | 0.73 |
| 79.1 | 45.4 | - | 0.57 | - | - | - | - | - | 10.5 | 32.6 | 0.32 | 0.72 |
Description: the studied specimens show a compressed section (E/H between 0.40 and 0.69) and a very involute conch, with convex flanks, with maximum thickness situated approximately at the bottom third. Until D = 35 mm the ventral region is slightly convex, becoming then clearly flat, with two angular edges. On the body chamber, the angular edges heighten as weak keels and give a clearly concave shape to the ventral region. In well preserved specimens, we also observe the presence of a very slight keel, wide and rounded, especially visible at the end of the phragmocone and the beginning of the body chamber.
Virtually none of the shells displays ornamentation. However, in the adult specimens, some weak rursiradiate ribs appear at the end of the body chamber, in the upper third of the flanks. Specimen no. MHNG GEPI 15935, from the Nicklesia pulchella Zone? of Escragnolles (Alpes-Maritimes), shows similar ornamentation which appears earlier on the body chamber. Its characteristics and stratigraphical position permits its assignment to H. boselliorum et al.
The
suture line (Fig. 3 ![]() )
contains a very wide, relatively shallow lateral lobe (P/L
ranges between 0.19 and 0.40), a reduced umbilical saddle and an indented
ventral lobe. The siphuncle, highly visible on specimens no. Mej11 (Pl. III
)
contains a very wide, relatively shallow lateral lobe (P/L
ranges between 0.19 and 0.40), a reduced umbilical saddle and an indented
ventral lobe. The siphuncle, highly visible on specimens no. Mej11 (Pl. III ![]() ,
fig. 3c) and no. LUS02 (Pl. IV
,
fig. 3c) and no. LUS02 (Pl. IV ![]() , fig. 3b) is located very low, near the dorsal
edge of the section.
, fig. 3b) is located very low, near the dorsal
edge of the section.
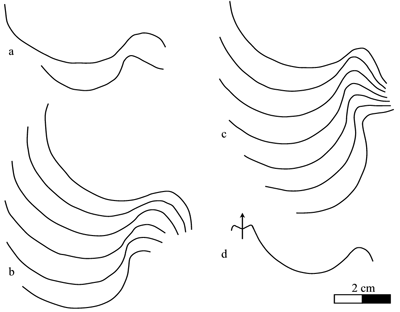
Click on thumbnail to enlarge the image.
Figure 3: Heminautilus boselliorum et al., 2012. Suture line of specimens no. Mej01, left side (a), MHNG GEPI 15935, right side (b), RG2001, left side (c) and Mej02, right side (d).
Variability: the sample (N = 13) studied here consists of sufficient well stratigraphically located specimens to allow a statistical analysis. The sample constitutes a quite homogeneous group, permitting us to estimate the intraspecific variability. The variability is minor and involves only the wideness of the conch and the morphology of the ventral region. The thickness of the shell varies from thin, in which the ratio E/H is between 0.40 and 0.50, to clearly thicker in which E/H can exceed 0.60. The width of the ventral region is directly correlated to the thickness of the shell: in the thin forms M/H is between 0.10 and 0.15, while in thick morphs this ratio can exceed 0.25. Independently, the shape of the ventral region varies on the phragmocone from flat to slightly concave, with the presence or absence of a very slight central keel, generally towards the end of the phragmocone or the beginning of the body chamber. To a lesser extent, we also observe a certain variability in the the suture line, with a more or less deep lateral lobe, whose ratio P/L can vary from 0.19 to 0.40. Rather significant variations in its depth are sometimes observed in the same specimen at various diameters, without an obvious link to an ontogenic stage.
Discussion and comparisons: Heminautilus boselliorum et al., 2012, is a species very close to H. saxbii (, 1848). However, it can be distinguished from H. saxbii () by a slightly less developed lateral lobe, with the average ratio P/L of 0.30 in H. boselliorum et al., and of 0.35 in H. saxbii (). In reference to the ornamentation and form of the shell, it can be distinguished by the presence of a very slight keel on the ventral region of the best preserved specimens of H. boselliorum et al. and a slightly differing section, as well as by ribbing which appears earlier on the body chamber of H. saxbii (). H. boselliorum et al. also occupies a stratigraphical position very different from that of H. saxbii (, 1848); this last taxon is known only in the Early Aptian, Deshayesites forbesi Zone.
H. boselliorum et al. is also similar to H. sanctaecrucis , 1980, from the latest Barremian - Early Aptian, Deshayesites forbesi Zone. H. sanctaecrucis can be easily distinguished by the presence of a strong median keel on the ventral region, as well as by shoulders lining the ventral region which are clearly stronger on the body chamber of adult specimens.
Finally, H. boselliorum et al. can be distinguished readily from H. lallierianus (, 1841) from the Early Aptian by its far less developed lateral lobe [P/L between 0.35 and 0.50 in H. lallierianus ()], its far less concave ventral region and its ontogenetically much later ribbing on the body chamber.
Heminautilus rangei (, 1922)
Pl. VII ![]() , fig. 3;
Pl. IX
, fig. 3;
Pl. IX ![]() ,
figs. 4-5;
,
figs. 4-5;
Pl. X ![]() ,
figs. 1-3;
Pl. XI
,
figs. 1-3;
Pl. XI ![]() ,
figs. 1-2
,
figs. 1-2
|
pars |
Nautilus lallieri , 1841 - , p. 129, Pl. 17, figs. 2-4, 6, non fig. 5 [= H. saxbii (, 1848)]. |
|
|
Nautilus rangeď nov. sp. - , p. 144, Pl. 4, figs. 2-3. |
||
|
Heminautilus lallierianus (, 1841) - , fig. 3, A-C. |
Material studied (N = 4): specimens no. EM696, EM697, EM698, EM699, Late Barremian?, Bir Lagama (north Sinai, Egypt).
Type:
(1922) figured two specimens (Pl. 4, figs. 2-3) from "Ledschime" (north Sinai, Egypt;
Fig. 4 ![]() ), without nomination of a holotype; the
illustration of (1922) is reproduced here on Pl. XI
), without nomination of a holotype; the
illustration of (1922) is reproduced here on Pl. XI ![]() ,
figs. 1-2.
These two specimens belonged to the collection, stored in the Geologisch-paläontologischen
Institut der Universität Leipzig (Germany), but could not be found and can be considered as lost. The original
figures of (1922, reproduced here Pl. XI
,
figs. 1-2.
These two specimens belonged to the collection, stored in the Geologisch-paläontologischen
Institut der Universität Leipzig (Germany), but could not be found and can be considered as lost. The original
figures of (1922, reproduced here Pl. XI ![]() ,
figs. 1-2) prevent the
development of a complete picture of the species. In particular, the shape of
the ventral region is difficult to construe and gives the impression of being
strongly concave. The specimens no. EM696, EM697, EM698 and EM699, as well as
specimen no. TK25/15 (Pl. X
,
figs. 1-2) prevent the
development of a complete picture of the species. In particular, the shape of
the ventral region is difficult to construe and gives the impression of being
strongly concave. The specimens no. EM696, EM697, EM698 and EM699, as well as
specimen no. TK25/15 (Pl. X ![]() , fig. 3) illustrated by
(2008), show that the ventral region of this species is not strongly concave.
Furthermore, in the original depictions the specimen in lateral view is
fragmentary, so that only the ornamentation of the upper part of the flanks is
visible. In the absence of specimens attributable to this species in the
collection, we erect as a neotype the specimen no. EM699 (Pl. X
, fig. 3) illustrated by
(2008), show that the ventral region of this species is not strongly concave.
Furthermore, in the original depictions the specimen in lateral view is
fragmentary, so that only the ornamentation of the upper part of the flanks is
visible. In the absence of specimens attributable to this species in the
collection, we erect as a neotype the specimen no. EM699 (Pl. X ![]() ,
fig. 1),
illustrated by (1916, Pl. 17, fig.
6a-b) and stored in the
collection of the École
des Mines (Université Claude , Lyon, France). It comes from Bir Lagama, a locality very close to that of the type
specimens displayed in (1922) (Fig. 4
,
fig. 1),
illustrated by (1916, Pl. 17, fig.
6a-b) and stored in the
collection of the École
des Mines (Université Claude , Lyon, France). It comes from Bir Lagama, a locality very close to that of the type
specimens displayed in (1922) (Fig. 4 ![]() ), and is probably from the
Late Barremian, Gerhardtia sartousiana
or Imerites giraudi zones (cf. section "stratigraphical
distribution").
), and is probably from the
Late Barremian, Gerhardtia sartousiana
or Imerites giraudi zones (cf. section "stratigraphical
distribution").
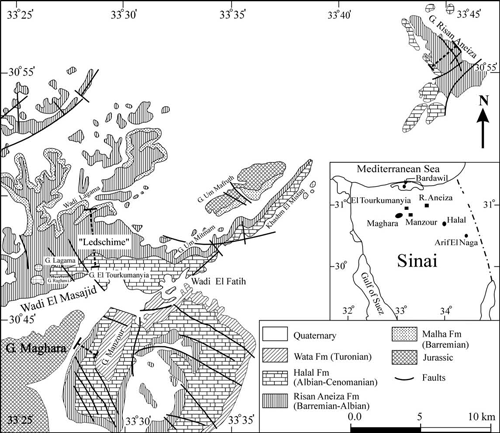
Click on thumbnail to enlarge the image.
Figure 4: Geological map of the north Sinai (Egypt). Modified from (2008).
By means of this neotype, completed by the paratypes no. EM696, EM697 and EM698 showing the various ontogenetic stages of the species, we can define the specific characteristics of H. rangei , 1922, more accurately than using the original depiction of , 1922.
Geographical distribution: the species is known only in the north of the Sinai (Egypt). All the known
specimens come from a very restricted area (Fig. 4 ![]() ): those presented by
(1922) come from "Ledschime", a location corresponding approximately (,
1920, Pl. 8) to the section
of El Tourkumanyia described by (2008), the source of the
specimen no. TK25/15 (coll. GMMU); the neotype no. EM699 and the three paratypes
no. EM696, EM697 and EM698 come from Bir Lagama, situated slightly further west
in the same massif.
): those presented by
(1922) come from "Ledschime", a location corresponding approximately (,
1920, Pl. 8) to the section
of El Tourkumanyia described by (2008), the source of the
specimen no. TK25/15 (coll. GMMU); the neotype no. EM699 and the three paratypes
no. EM696, EM697 and EM698 come from Bir Lagama, situated slightly further west
in the same massif.
Stratigraphical distribution: both specimens shown in (1922) were dated, according to him,
from the Vraconian (= latest Albian). However, the presence in the same
formation ("Ledschime-Horizont") of representatives of the genus Peudohaploceras
, 1900, close to P. douvillei
, 1920) (= Puzosia kossmati
sp. nov. in , 1922, p.
140, Pl. 3, fig. 10)
indicates a very likely Late Barremian age. The specimen no. TK25/15 (Pl. X ![]() ,
fig. 3) illustrated by (2008) comes from the Late
Barremian, Subpulchellia oehlerti Zone
[= Mogharaeceras priscum (,
1916)]. According to et al.
(2011), Mogharaeceras priscum ()
is a Late Barremian species, probably present in an interval included in Gerhardtia
sartousiana to Imerites giraudi
zones. (2008) also indicates, without figures, the
presence of Heminautilus lallierianus (, 1841) [= Heminautilus
rangei (, 1922) or Heminautilus saxbii (, 1848) ?] in the Early
Aptian. Specimens depicted by are attributed by him to the
Aptian without more precision. However, compared with the specimen shown by
(2008), which presents exactly the same ornamental and morphological
characteristics, such a stratigraphical position seems doubtful and these
specimens are more likely from the Late Barremian. These specimens were also
reallocated to the Barremian by (1955, p. 12). Thus
Heminautilus rangei (, 1922) is clearly present in
the Late Barremian, in the Gerhardtia
sartousiana Zone or Imerites giraudi
Zone; its supposed presence in the Early Aptian is unproven and might stem from
the confusion with another species collected in higher levels as attested by the
presence of Heminautilus
saxbii (, 1848)
among the material collected by (1916).
,
fig. 3) illustrated by (2008) comes from the Late
Barremian, Subpulchellia oehlerti Zone
[= Mogharaeceras priscum (,
1916)]. According to et al.
(2011), Mogharaeceras priscum ()
is a Late Barremian species, probably present in an interval included in Gerhardtia
sartousiana to Imerites giraudi
zones. (2008) also indicates, without figures, the
presence of Heminautilus lallierianus (, 1841) [= Heminautilus
rangei (, 1922) or Heminautilus saxbii (, 1848) ?] in the Early
Aptian. Specimens depicted by are attributed by him to the
Aptian without more precision. However, compared with the specimen shown by
(2008), which presents exactly the same ornamental and morphological
characteristics, such a stratigraphical position seems doubtful and these
specimens are more likely from the Late Barremian. These specimens were also
reallocated to the Barremian by (1955, p. 12). Thus
Heminautilus rangei (, 1922) is clearly present in
the Late Barremian, in the Gerhardtia
sartousiana Zone or Imerites giraudi
Zone; its supposed presence in the Early Aptian is unproven and might stem from
the confusion with another species collected in higher levels as attested by the
presence of Heminautilus
saxbii (, 1848)
among the material collected by (1916).
Dimensions (in mm): see Table 3.
Table 3: Measurements of Heminautilus rangei (, 1922).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Type specimen (, 1922) | 125 | 80 | 50 | 0.64 | 0.4 | 0.63 | - | - | - | - | - | - | - |
| Neotype no. EM699 | 135.8 | 87.8 | - | 0.65 | - | - | 16.1 | 0.12 | 0.18 | - | - | - | - |
| 94.1 | 55.5 | 38.4 | 0.59 | 0.41 | 0.69 | 10.4 | 0.11 | 0.19 | - | - | - | - | |
| no. EM698 | 72.5 | 43.6 | 28.7 | 0.6 | 0.4 | 0.66 | 6.3 | 0.09 | 0.14 | 9.6 | 35 | 0.27 | 0.8 |
| 52.8 | 34.8 | 22.5 | 0.66 | 0.43 | 0.65 | 5.3 | 0.1 | 0.15 | 7.5 | 28 | 0.27 | 0.8 | |
| no. EM697 | 59 | 38 | 24.6 | 0.64 | 0.42 | 0.65 | 5.6 | 0.09 | 0.15 | - | - | - | - |
| 46.3 | 28.8 | 18.5 | 0.62 | 0.4 | 0.64 | 4.7 | 0.1 | 0.16 | 6.1 | 23.5 | 0.26 | 0.82 | |
| 32.3 | 19.8 | 13.9 | 0.61 | 0.43 | 0.7 | 3.5 | 0.11 | 0.18 | 4.1 | 16.4 | 0.25 | 0.83 | |
| no. EM696 | 35 | 24.7 | 15.6 | 0.71 | 0.45 | 0.63 | 3.8 | 0.11 | 0.15 | - | - | - | - |
| no. TK25/15 (, 2008) | 125 | 74 | 41 | 0.59 | 0.33 | 0.55 | 12 | 0.1 | 0.16 | - | - | - | - |
Description: the four specimens of the collection of the École des Mines, all illustrated by (1916, Pl. 17) as Nautilus lallierianus , 1841, provide precise knowledge about the ontogeny and the characteristics of this species. The neotype no. EM699, preserved with at least a part of its body chamber, probably represents the adult stage of the species.
With
a diameter of up to about 20 mm, no ornamentation is visible with the exception
of fine longitudinal striae on the flanks and the ventral region, visible on the
shell of the paratype no. EM696 (Pl. IX, fig. 4). The
section is oval, with rounded flanks and ventral region. Beyond this diameter,
the section changes and becomes trapezoidal; flanks flatten and converge on the
ventral region which very gradually becomes flat, with the appearance of a
slight ventro-lateral shoulder. At the same time, the ribbing appears,
consisting of broad ribs arising near the umbilical border and whose aboral edge
follows a flat slope while the adoral edge is clearly more abrupt. Ribs are
radial or slightly rursiradiate in the lower half of the flanks, becoming
strongly rursiradiate beyond, and then fading approaching the ventral region.
With growth, these ribs become broader and their point of appearance moves
higher on the flanks. From approximately D = 70 mm, the ventral region is
clearly flat (specimen no. EM698, Pl. IX ![]() , fig. 5;
Pl. X
, fig. 5;
Pl. X ![]() , fig. 2). Finally, in the adult specimens
(no. EM699, Pl. X
, fig. 2). Finally, in the adult specimens
(no. EM699, Pl. X ![]() , fig. 1; no. TK25/15,
Pl. X
, fig. 1; no. TK25/15,
Pl. X ![]() , fig. 3), ribs appear slightly
before the middle of flanks; they are very broad, flat, and barely separated by
a narrow groove. They become irregular in their spacing, delineation and point
of appearance on the flanks. The ventral region is lined by strong shoulders,
giving to the ventral region a slightly to clearly concave shape (for example in
the specimen illustrated by (1922) Pl. 4, fig. 3; reproduced here
Pl. XI
, fig. 3), ribs appear slightly
before the middle of flanks; they are very broad, flat, and barely separated by
a narrow groove. They become irregular in their spacing, delineation and point
of appearance on the flanks. The ventral region is lined by strong shoulders,
giving to the ventral region a slightly to clearly concave shape (for example in
the specimen illustrated by (1922) Pl. 4, fig. 3; reproduced here
Pl. XI ![]() , fig. 2). We also observe fine growth lines there forming a rounded
chevron (specimen no. EM699, Pl. X
, fig. 2). We also observe fine growth lines there forming a rounded
chevron (specimen no. EM699, Pl. X ![]() , fig. 1b). Finally,
at the end of the preserved part of the neotype no. EM699, a wide and slightly
raised keel appears on the ventral region.
, fig. 1b). Finally,
at the end of the preserved part of the neotype no. EM699, a wide and slightly
raised keel appears on the ventral region.
The
suture line (Fig. 5 ![]() ), easily observable on specimens no. EM697 and no. EM698,
shows a relatively undeveloped, rather narrow lateral saddle and a wide and
shallow lateral lobe (P/L between 0.25
and 0.27). It crosses the ventral region straight, without forming a ventral
lobe. The siphuncle, visible on specimens no. EM696 and no. EM697, is situated
near the dorsal edge of the section.
), easily observable on specimens no. EM697 and no. EM698,
shows a relatively undeveloped, rather narrow lateral saddle and a wide and
shallow lateral lobe (P/L between 0.25
and 0.27). It crosses the ventral region straight, without forming a ventral
lobe. The siphuncle, visible on specimens no. EM696 and no. EM697, is situated
near the dorsal edge of the section.
Click on thumbnail to enlarge the image.
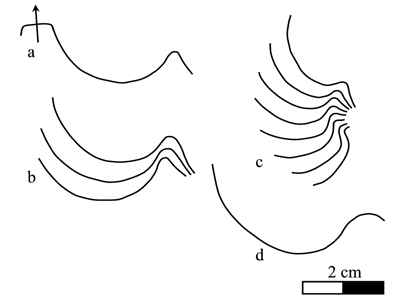
Figure 5: Heminautilus rangei (, 1922). Suture line of specimens no. EM698, right side (a-b), EM697, left side (c) and of the unregistered specimen, right side (d) figured by (1922; Pl. 4, fig. 2).
Discussion and comparisons: the very characteristic ornamentation of Heminautilus rangei (, 1922) differentiates it immediately from most of the other species of the genus Heminautilus , 1927, where the ornamentation is absent or only present on the adoral part of the body chamber, with much narrower rounded ribs.
However, some characteristics of the shell of H. rangei (), in particular the pattern of its suture line and the shape of its ventral region, indicate that this species is close to H. boselliorum et al., 2012, and H. sanctaecrucis , 1980. In H. sanctaecrucis the median keel is consistently well developed in all specimens, whereas in H. rangei () it is observable in one specimen, the neotype no. EM699, but less developed than H. sanctaecrucis .
H. rangei () is also similar to H. ? japonicus sp. nov., in particular its slightly concave ventral region, ornamented with fine growth lines, as well as its very similar stratigraphical position in the Late Barremian. However the suture line and the ribbing of H. ? japonicus sp. nov. are very different, which distinguishes it easily from H. rangei ().
H. rangei () also shows some affinities with the Japanese species H. ? tyosiensis ( & , 1953), because of its slightly concave ventral region lined with two strong shoulders, and the presence of comparable ribbing on a part of the phragmocone. However, H. ? tyosiensis ( & ) can be easily distinguished from H. rangei () by its much wider ventral region, its quickly fading ribbing and a different suture line, with a clearly deeper lateral lobe. H. ? tyosiensis ( & ) also occupies a very different stratigraphical and geographical position (= Dufrenoyia furcata Zone, Japan).
Heminautilus sanctaecrucis , 1980
Pl. XI ![]() ,
figs. 3-5;
Pl. XII
,
figs. 3-5;
Pl. XII ![]() ,
figs. 1-3;
Pl. XIII
,
figs. 1-3;
Pl. XIII ![]() ,
figs. 1-3;
,
figs. 1-3;
Pl. XIV ![]() ,
figs. 1-2;
Pl. XV
,
figs. 1-2;
Pl. XV ![]() ,
fig. 1;
Pl. XVI
,
fig. 1;
Pl. XVI ![]() , fig. 1
, fig. 1
|
Nautilus lallierianus , 1841 - F.-J. & , p. 148, Pl. 19, fig. 6a-c. |
||
|
Xenocheilus lallierianus (, 1841) - , p. 18, Pl. 4, fig. 1. |
||
|
Heminautilus sanctaecrucis sp. nov. - , p. 137, Pl. 1. figs. 1-2, 4a. |
||
|
Heminautilus cf. sanctaecrucis , 1980 - , p. 30, Pl. 1, fig. 6. |
||
|
Heminautilus sanctaecrucis , 1980 - , p. 30, Figs. 5-7. |
||
|
pars |
Heminautilus sanctaecrucis , 1980 - & , p. 63, Pl. 1, fig. 1?; Pl. 2, fig. 1?; Pl. 3, figs. 1?, 2, 2a; Pl. 4, figs. 1, 1a, 2, 2a. |
|
|
? |
Heminautilus saxbii (, 1848) - & , p. 65, Pl. 5, fig. 1, 1a. |
|
|
Heminautilus sanctaecrucis , 1980 - et al., p. 904, Pl. 1, figs. 6. |
||
|
Heminautilus sanctaecrucis , 1980 - , p. 118, Figs. 6-7. |
Material studied (N = 4): specimen no. MCGL 20747, Early Aptian, Deshayesites forbesi Zone, Sainte-Croix (Switzerland); specimens no. MHNG GEPI 15936, MHNN 26904, Early Aptian, Deshayesites forbesi Zone, La Presta, Val-de-Travers (Switzerland); specimen no. Bw599, coll. FSM, Late Barremian, Imerites giraudi Zone, Pseudocrioceras waagenoides Subzone, bed 78, Les Caniers, La Bédoule (Bouches du Rhône, France).
Type: the holotype (by original designation of , 1980, p. 138) is the specimen no. MCGL 20747 illustrated by F.-J. & (1858) Pl. 19, fig. 6, as Nautilus lallierianus , 1841. Stored in the Muséum Géologique Cantonal of Lausanne (Switzerland), it is depicted anew in this work Pl. XI , fig. 3.
Geographical distribution: the species is known in Switzerland, Bulgaria, southeast France and Tunisia.
Stratigraphical distribution: the holotype comes from the base of the "Marnes
jaunes de la Vraconne" (F.-J. & , 1858) at
Sainte-Croix (Switzerland), immediately above the Urgonian Limestone. The bottom
third of these Jura marls corresponds to the Grünten Member defined in the
Helvetic Domain, whose base is attributed to the Deshayesites forbesi Zone
(A. et al., 2009, p.
492-493). The Bulgarian
specimens (coll. MPUS) studied by (1967) and
& (1995) are attributed to the Late Barremian
(specimens no. K1 6001, K1 6002, K1 6003, K1 6004) and to the Early Aptian
(specimen no. K1 1692) without more precision. In Tunisia the species is present
in the Early Aptian, Deshayesites
oglanlensis Zone (specimen no. GSUB C4035,
et al., 2009).
In southeast France, the range of Heminautilus
sanctaecrucis , 1980, seems to be latest Barremian/earliest Aptian. The unregistered specimen of the
collection (Pl. XV ![]() , fig. 1) comes from the Coustellet quarry (Vaucluse, France),
situated in the units 3 and 4 of the series of the southwest region of the
Vaucluse mountains and attributed to the Early Bedoulian by
(1976,
p. 38-42). The specimen no. 131H (coll. , Pl. XI, fig. 5;
Pl. XII
, fig. 1) comes from the Coustellet quarry (Vaucluse, France),
situated in the units 3 and 4 of the series of the southwest region of the
Vaucluse mountains and attributed to the Early Bedoulian by
(1976,
p. 38-42). The specimen no. 131H (coll. , Pl. XI, fig. 5;
Pl. XII ![]() ,
fig. 1) comes from the col
des Abeilles, Monieux (Vaucluse, France), and corresponds to the unit of the "Calcaires
fins des Colombičres" of the Nesque series, attributed to the latest Barremian/earliest Aptian by
(1976, p. 34-38). At La Bédoule (Bouches du Rhône, France) the
species was collected at the top of the Late Barremian, Imerites giraudi Zone, Pseudocrioceras
waagenoides Subzone (specimen no. Bw599, Pl. XII
,
fig. 1) comes from the col
des Abeilles, Monieux (Vaucluse, France), and corresponds to the unit of the "Calcaires
fins des Colombičres" of the Nesque series, attributed to the latest Barremian/earliest Aptian by
(1976, p. 34-38). At La Bédoule (Bouches du Rhône, France) the
species was collected at the top of the Late Barremian, Imerites giraudi Zone, Pseudocrioceras
waagenoides Subzone (specimen no. Bw599, Pl. XII ![]() , fig. 2;
Pl. XIII
, fig. 2;
Pl. XIII ![]() , fig. 1).
Heminautilus sanctaecrucis , 1980, is thus present from the Late Barremian, top of the Imerites
giraudi Zone, to the Early Aptian, Deshayesites
forbesi Zone.
, fig. 1).
Heminautilus sanctaecrucis , 1980, is thus present from the Late Barremian, top of the Imerites
giraudi Zone, to the Early Aptian, Deshayesites
forbesi Zone.
Dimensions (in mm): see Table 4.
Table 4: Measurements of Heminautilus sanctaecrucis , 1980.
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. MCGL 20747 | 96.3 | 55.4 | 34 | 0.58 | 0.35 | 0.61 | 9.8 | 0.1 | 0.18 | - | - | - | - |
| 86.3 | 51 | 26.8 | 0.59 | 0.31 | 0.53 | 8.7 | 0.10 | 0.17 | - | - | - | - | |
| 69.2 | 41.3 | 21.9 | 0.6 | 0.32 | 0.53 | 7.2 | 0.10 | 0.17 | - | - | - | - | |
| - | - | - | - | - | - | - | - | - | 7 | 34 | 0.21 | - | |
| - | - | - | - | - | - | - | - | - | 9.5 | 34 | 0.28 | - | |
| no. BW599 | 126.1 | 72.4 | - | 0.57 | - | - | 9.5 | 0.08 | 0.13 | - | - | - | - |
| 104.3 | 60.1 | - | 0.58 | - | - | 8.2 | 0.08 | 0.14 | 11 | 48.2 | 0.23 | 0.8 | |
| - | 48.2 | - | - | - | - | 6.8 | - | 0.14 | 7.3 | 37.7 | 0.19 | 0.78 | |
| Unregistered specimen (coll. ) | 154.1 | 92.9 | 45.4 | 0.6 | 0.29 | 0.49 | 17.9 | 0.12 | 0.19 | - | - | - | - |
| - | - | - | - | - | - | - | - | - | 7.2 | 49.9 | 0.14 | - | |
| no. 131H | 170 | 90 | - | 0.53 | - | - | - | - | - | - | - | - | - |
| - | 78 | - | - | - | - | - | - | - | 20 | 52 | 0.38 | 0.67 | |
| - | 60 | - | - | - | - | - | - | - | 20 | 42 | 0.48 | 0.7 | |
| no. K1 1692 ( & , 1995) | 133 | 80 | 53 | 0.6 | 0.4 | 0.66 | 12 | 0.09 | 0.15 | 16 | 47 | 0.34 | 0.59 |
| - | 58 | - | - | - | - | - | - | - | 14 | 39 | 0.36 | 0.67 | |
| no. K1 6001 ( & , 1995) | 147 | 97 | 59 | 0.66 | 0.4 | 0.61 | 13 | 0.09 | 0.13 | - | - | - | - |
| no. K1 6003 ( & , 1995) | 109 | 67 | - | 0.61 | - | - | - | - | - | - | - | - | - |
| - | 55 | - | - | - | - | - | - | - | 13 | 37 | 0.35 | 0.67 | |
| no. K1 6005 ( & , 1995) | 53 | 35 | 24 | 0.66 | 0.45 | 0.69 | 4 | 0.08 | 0.11 | - | - | - | - |
Description:
the studied specimens show a relatively compressed section, with slightly
rounded flanks, where maximum thickness is in the lower third. The ventral
region is rather narrow and flat or slightly concave. It is lined by two more or
less strongly angular ventro-lateral shoulders and shows a clear median keel,
especially visible in the last whorl. On the specimen no. Bw599, this median
keel seems to appear where the diameter is about 85 mm, later than on the
holotype where it is visible from approximately D = 60 mm. The ornamentation is
absent on almost all the shell; we observe however some sinuous ribs present
only at the adoral part of the body chamber of adult specimens (specimen no.
131H, Pl. XII ![]() , fig. 1). The body chamber is only partially preserved on the
holotype and
probably for this reason there are no ribs present.
, fig. 1). The body chamber is only partially preserved on the
holotype and
probably for this reason there are no ribs present.
The
suture line is slightly sinuous, with a shallow lateral lobe (P/L most of the time between 0.20 and 0.35). It crosses the ventral
region without forming a ventral lobe, except when the ventral region shows
strong shoulders, where it forms a shallow ventral lobe (Fig. 6 ![]() ). The siphuncle,
visible on specimen no. MHNN 26904 (Pl. XIII
). The siphuncle,
visible on specimen no. MHNN 26904 (Pl. XIII ![]() , fig. 3), is situated in the dorsal half of the
section.
, fig. 3), is situated in the dorsal half of the
section.
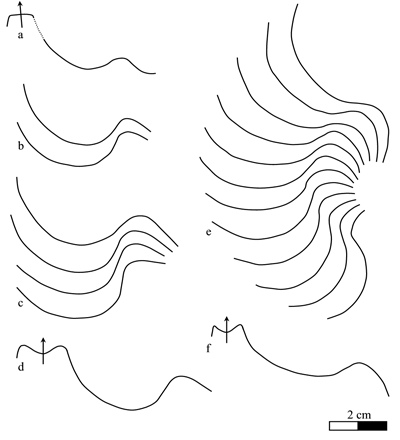
Click on thumbnail to enlarge the image.
Figure 6: Heminautilus sanctaecrucis , 1980. Suture line of specimens no. MCGL 20747, right side (a), 131H, right side (b), K1 1692, left side (c, d) and Bw599, left side (e, f).
The
specimen no. K1 6001, shown by &
(1995)
and depicted anew (Pl. XIV ![]() , fig. 1;
Pl. XVI
, fig. 1;
Pl. XVI ![]() ,
fig. 1), is tentatively attributed to Heminautilus sanctaecrucis
, 1980. It differs from other representatives of the species
by the presence of clear rursiradiate ribbing on the top of the flanks of the
body chamber and by the shape of its ventral region, lined by two very strong
ventro-lateral ridges. This last characteristic may be a consequence of the
different mode of preservation of this specimen, where the shell is very well
preserved on the ventral region because it is recrystallized, while the other
specimens are preserved as internal molds.
,
fig. 1), is tentatively attributed to Heminautilus sanctaecrucis
, 1980. It differs from other representatives of the species
by the presence of clear rursiradiate ribbing on the top of the flanks of the
body chamber and by the shape of its ventral region, lined by two very strong
ventro-lateral ridges. This last characteristic may be a consequence of the
different mode of preservation of this specimen, where the shell is very well
preserved on the ventral region because it is recrystallized, while the other
specimens are preserved as internal molds.
Discussion and comparisons: Heminautilus sanctaecrucis , 1980, is a species close to H. lallierianus (, 1841) and H. saxbii (, 1848). It can be distinguished from both taxa by its suture line, since the lateral lobe is clearly shallower [P/L is mostly between 0.20 and 0.35 while it is generally between 0.30 and 0.40 for H. saxbii () and between 0.35 and 0.50 for H. lallierianus ()], as well as by the appearance of a keel on the ventral region proximal to the body chamber. H. sanctaecrucis can also be distinguished from H. lallierianus () by a less concave ventral region and by the almost total absence of ornamentation.
H. sanctaecrucis also shows numerous affinities with H. boselliorum et al., 2012, with a similar suture line, an identical section and the same type of ornamentation visible at the adoral part of the body chamber. The two can be distinguished by the ventral keel, well defined on the main part of the shell in H. sanctaecrucis while it is very discreet or absent in H. boselliorum et al.
The shape of the ventral region of H. sanctaecrucis is also similar to H. rangei (, 1922), but can readily be distinguished from it by the absence of ornamentation on the flanks of the main part of the shell.
Heminautilus aff. sanctaecrucis , 1980
Pl. XII ![]() , fig. 4
, fig. 4
Material studied (N = 1): specimen no. 415951, coll. (donated by ), Late Barremian, Gerhardtia sartousiana Zone, base of the Hemihoplites feraudianus Subzone, bed 165/041, Angles (Alpes-de-Haute-Provence, France).
Dimensions (in mm): see Table 5.
Table 5: Measurements of Heminautilus aff. sanctaecrucis , 1980.
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| no. 415951 | 58.1 | 35 | - | 0.6 | - | - | 4.3 | 0.07 | 0.12 | - | - | - | - |
| - | 28.6 | 14.7 | - | - | 0.51 | 4.5 | - | 0.16 | 6.2 | 21.1 | 0.29 | 0.74 |
Description: small incomplete specimen, consisting of part of the phragmocone and the beginning of the body chamber. The section is narrow, with maximum thickness in the lower third of the flanks, with a flat ventral region with angular edges, and slightly rounded flanks where no ornamentation is visible. On the ventral region, there is a visible yet relatively undeveloped median keel.
The
suture line is sinuous, with a wide, round and relatively shallow lateral lobe (Fig. 7 ![]() ). The ventral lobe, difficult to observe, seems shallow or absent. The
position of the siphuncle is not visible.
). The ventral lobe, difficult to observe, seems shallow or absent. The
position of the siphuncle is not visible.
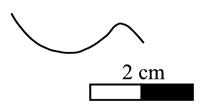
Click on thumbnail to enlarge the image.
Figure 7: Heminautilus aff. sanctaecrucis , 1980. Suture line of specimen no. 415951, right side.
Discussion and comparisons: this specimen has a stratigraphical position intermediate between Heminautilus boselliorum et al., 2012, and H. sanctaecrucis , 1980. The suture line and the section of H. aff. sanctaecrucis are identical to those observed in H. boselliorum et al. and H. sanctaecrucis ; H. aff. sanctaecrucis can be distinguished from H. boselliorum et al. by its clearly visible ventral keel, but it is weaker than the one observed in H. sanctaecrucis . Its fragmentary state does not allow us to be more precise on its taxonomic identification.
Heminautilus saxbii (, 1848)
Pl. XV ![]() , fig. 2;
Pl. XVI
, fig. 2;
Pl. XVI ![]() ,
figs. 2-3;
Pl. XVII
,
figs. 2-3;
Pl. XVII ![]() ,
figs. 1-3;
,
figs. 1-3;
Pl. XVIII ![]() ,
figs. 1-3;
Pl. XIX
,
figs. 1-3;
Pl. XIX ![]() ,
figs. 1-3;
Pl. XX
,
figs. 1-3;
Pl. XX ![]() ,
figs. 1-3;
,
figs. 1-3;
Pl. XXI ![]() ,
figs. 1-4;
Pl. XXII
,
figs. 1-4;
Pl. XXII ![]() ,
figs. 1-3;
Pl. XXIII
,
figs. 1-3;
Pl. XXIII ![]() ,
figs. 1-4;
,
figs. 1-4;
Pl. XXIV ![]() ,
figs. 1-3;
Pl. XXV
,
figs. 1-3;
Pl. XXV ![]() ,
figs. 1-4;
,
figs. 1-4;
Pl. XXVI ![]() ,
figs. 1-3;
Pl. XXXI
,
figs. 1-3;
Pl. XXXI ![]() ,
fig. 3
,
fig. 3
|
Nautilus saxbii nov. sp. - , p. 106, unnumbered fig. |
||
|
Nautilus verneuilli nov. sp. - , Pl. 3, fig. 4. |
||
|
pars |
Nautilus lallieri , 1841 - , p. 129, Pl. 17, fig. 5, non figs. 2-4, 6 [= H. rangei (, 1922)]. |
|
|
Heminautilus saxbii (, 1848) - , p. 434, Pl. 10, figs. 1-2. |
||
|
Heminautilus saxbii (, 1848) - , p. K454, Fig. 335, 2a-b. |
||
|
Heminautilus saxbii (, 1848) - , p. 139, Pl. 1, fig. 4b. |
||
|
Heminautilus saxbii (, 1848) - , p. 30, Fig. 4. |
||
|
Heminautilus lallierianus (, 1841) - , p. 356, Pl. 175, fig. 3. |
||
|
non |
Heminautilus saxbii (, 1848) - & , p. 65, Pl. 5, fig. 1, 1a (= ? H. sanctaecrucis , 1980). |
|
|
non |
Heminautilus aff. saxbii (, 1848) - , p. 37 (= H. boselliorum et al., 2012). |
|
|
Heminautilus saxbii (, 1848) - & , Fig. 3b, f. |
||
|
? |
Heminautilus lallierianus (, 1841) - , p. 101, Pl. 1, figs. 1-3. |
|
|
Heminautilus saxbii (, 1848) - & , p. 22, Figs. 3b, 5a-d. |
||
|
Heminautilus tejeriensis nov. sp. - & , p. 24, Figs. 3c, 4a, 6a, 7a-b. |
||
|
pars |
Heminautilus saxbii (, 1848) - , p. 11, photos 1-2, 11, 12?. |
|
|
Heminautilus saxbii (, 1848) - et al., Figs. 16-17. |
||
|
Heminautilus saxbii (, 1848) - , p. 117, Figs. 1-2, 4-5. |
||
|
Heminautilus saxbii (, 1848) - , Fig. 3.12A, B. |
Material studied (N = 101): specimens no. EM700, Early Aptian?, Bir Lagama, Gabal Maghara (Egypt); no. PNRL 1189, Early Aptian, Deshayesites forbesi Zone, Murs (Vaucluse, France); no. PM1, PM2, PM3, PM4, PM5, PM6, PM13, PM3N1, PM3N2, PM3N11, PMBb2, coll. MNHN, no. A-114, A-117, A-119, A-122, A-127, A-131, A-140, A-144, A-145, A-146, AG-3, AG-13, JM-26, JM-34, JM-36, JM-37, JM-38, M-8, PUAB 5905, PUAB 48011, PUAB 48013, PUAB 48018, PUAB 48027, PUAB 48035, PUAB 48037, PUAB 48040, PUAB 48042, PUAB 48043, PUAB 48044, PUAB 48046, PUAB 48064, PUAB 48074, PUAB 48080, PUAB 48092, PUAB 48098, PUAB 48106, PUAB 48125, PUAB 48127, PUAB 48128, PUAB 48133, PUAB 48143, PUAB 48158, PUAB 48163, PUAB 48164, PUAB 48165, PUAB 48169, PUAB 48202, PUAB 48205, PUAB 48215, PUAB 48223, PUAB 48232, PUAB 48242, PUAB 48246, PUAB 48254, PUAB 48255, PUAB 48256, PUAB 48261, PUAB 48275, PUAB 48277, PUAB 48278, PUAB 48282, PUAB 48288, PUAB 48292, PUAB 48295, PUAB 48304, PUAB 48314, PUAB 48316, PUAB 48318, PUAB 48321, PUAB 48322, PUAB 48328, PUAB 48366, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain); no. PUAB 48177, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Alacón, Province of Teruel (Spain); no. PUAB 89786, PUAB 89787, PUAB 89788, PUAB 89790, PUAB 89791, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Galve, Province of Teruel (Spain); no. PUAB 88767, PUAB 88770, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Tolodella, Maestrat Basin (Spain); no. MV 423.1, MV 428.2, MV 428.3, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mola Murada, Maestrat Basin (Spain); no. PUAB 88762, PUAB 88763, Early Aptian, Cabo Marls Formation, Deshayesites forbesi Zone, Lo Pui, Organyŕ Basin (Spain); no. PUAB 88372, PUAB 88373, Early Aptian, Patrocinio Formation, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Cuchía, Basque Cantabrian Basin (Spain); no. MB RE1962A, Early Aptian, Tibú Formation, Seboruco section (east of Mérida, Venezuela).
Type: the holotype
(by monotypy) is the specimen illustrated by (1848) p. 106, stored
in the collection at the British Museum of National History with
the no. BMNH 47019. It comes from the Early Aptian of the Isle of Wight
(England) and is depicted anew here
Pl. XVI ![]() , fig. 2.
, fig. 2.
Geographical distribution: the species is known from England, France, Spain and Egypt. It may be
present in Venezuela (specimen no. MB RE1962A,
Pl. XXIV ![]() , fig. 3).
, fig. 3).
Stratigraphical distribution: the type of comes from the Lower Greensand,
"Crackers'
group" of Atherfield, Isle of Wight and is dated as the Deshayesites forbesi Zone; it most certainly comes from the Lower
Lobster Beds Formation (, written communication, 2013),
corresponding to the bottom of the Roloboceras
hambrovi Subzone (, 1985; et al.,
1998).
All the Spanish specimens studied here come from levels dated as the Deshayesites
forbesi Zone, Roloboceras hambrovi
Subzone ( et al., 2007).
The Egyptian specimen no. EM700 is assigned by (1916) to the
Early Aptian, as well as the specimens displayed
in (2006); however, a stratigraphical position in the top of the Late
Barremian cannot be excluded. The
specimens quoted by &
(1995; coll. MPUS)
in the Early Barremian (no. K1 1693 and no. K1 1694, displayed anew Pl.
VII ![]() , figs.
1-2; Pl.
IX
, figs.
1-2; Pl.
IX ![]() , fig. 1) and the Late Barremian (no. K1 6005) belong to taxa
different from Heminautilus
saxbii (, 1848).
The specimen no. K1 6005 probably belongs to H.
sanctaecrucis , 1980;
however, without a visible suture line and with a very weak ventral keel
confirmation is not possible. The specimens no. K1 1693 and no. K1 1694 belong
to H.
boselliorum
et al., 2012,
as does the specimen of H. aff. saxbii () of
(2002, p.
37; figured here Pl. VI
, fig. 1) and the Late Barremian (no. K1 6005) belong to taxa
different from Heminautilus
saxbii (, 1848).
The specimen no. K1 6005 probably belongs to H.
sanctaecrucis , 1980;
however, without a visible suture line and with a very weak ventral keel
confirmation is not possible. The specimens no. K1 1693 and no. K1 1694 belong
to H.
boselliorum
et al., 2012,
as does the specimen of H. aff. saxbii () of
(2002, p.
37; figured here Pl. VI ![]() , fig. 2; Pl.
VII
, fig. 2; Pl.
VII ![]() , fig. 4) in the Kotetishvilia
nicklesi Zone. The specimens of H. saxbii () of
southeast France [specimens displayed by (2007, 2010) and specimen
no. 12576, coll. FSM, shown Pl. XXIV
, fig. 4) in the Kotetishvilia
nicklesi Zone. The specimens of H. saxbii () of
southeast France [specimens displayed by (2007, 2010) and specimen
no. 12576, coll. FSM, shown Pl. XXIV ![]() , fig. 1;
Pl. XXV
, fig. 1;
Pl. XXV ![]() , fig. 1] come from the
Early Aptian without more precision. Finally, specimen no. MB RE1962A,
tentatively attributed to H. saxbii (),
comes from the Seboruco section (Venezuela). This section also yielded ammonites
attributed to the genus Roloboceras ,
1954, by (1982, p. 19), but which more likely belong to the genus Cheloniceras
, 1903; the
stratigraphical position of this specimen can thus be situated approximately in
the top of the Early Aptian.
, fig. 1] come from the
Early Aptian without more precision. Finally, specimen no. MB RE1962A,
tentatively attributed to H. saxbii (),
comes from the Seboruco section (Venezuela). This section also yielded ammonites
attributed to the genus Roloboceras ,
1954, by (1982, p. 19), but which more likely belong to the genus Cheloniceras
, 1903; the
stratigraphical position of this specimen can thus be situated approximately in
the top of the Early Aptian.
Heminautilus saxbii (, 1848) is thus a species present in the Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, in particular in the bottom of the subzone. The existence of older or more recent representatives of the species remains doubtful and remains to be confirmed.
Dimensions (in mm): see Table 6.
Table 6: Measurements of Heminautilus saxbii (, 1848).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. BMNH 47019 | 86 | 56 | 32 | 0.65 | 0.37 | 0.57 | 7 | 0.08 | 0.13 | - | - | - | - |
| 69 | 42 | - | 0.61 | - | - | 8 | 0.12 | 0.19 | 13 | 31 | 0.42 | 0.74 | |
| 52 | 31 | - | 0.60 | - | - | 6 | 0.12 | 0.19 | 8 | 23 | 0.35 | 0.74 | |
| no. EM700 | 128.6 | 83.6 | 49.2 | 0.65 | 0.38 | 0.59 | 12.2 | 0.09 | 0.15 | - | - | - | - |
| 90.6 | 57.8 | 35.1 | 0.64 | 0.39 | 0.61 | 9.8 | 0.11 | 0.17 | 14.5 | 42.4 | 0.34 | 0.73 | |
| no. PNRL 1189 | - | 87.2 | 43.3 | - | - | 0.50 | 13.7 | - | 0.16 | - | - | - | - |
| - | 77.5 | 30.5 | - | - | 0.39 | 9.7 | - | 0.13 | 14.7 | 57.1 | 0.26 | 0.74 | |
| no. 12576 | 197 | 118 | 50 | 0.60 | 0.25 | 0.42 | - | - | - | - | - | - | - |
| 145 | 100 | - | 0.69 | - | - | - | - | - | 32 | 70 | 0.46 | 0.70 | |
| - | 89 | - | - | - | - | - | - | - | 25 | 67 | 0.37 | 0.74 | |
| no. PM1 | 138 | 85 | 51 | 0.62 | 0.37 | 0.60 | 20 | 0.14 | 0.24 | - | - | - | - |
| - | 63 | 36 | - | - | 0.57 | 9 | - | 0.14 | 17 | 49 | 0.35 | 0.78 | |
| - | 46 | 30 | - | - | 0.65 | 8 | - | 0.17 | 11 | 36 | 0.31 | 0.78 | |
| no. PM2 | 126 | - | 45 | - | 0.36 | - | 16 | 0.13 | - | - | - | - | - |
| 98 | - | 33 | - | 0.34 | - | 7 | 0.07 | - | 14 | 45 | 0.31 | - | |
| no. PM3 | 129 | 81 | 54 | 0.63 | 0.42 | 0.67 | 19 | 0.15 | 0.23 | - | - | - | - |
| no. PM4 | 81 | 53 | 29 | 0.65 | 0.36 | 0.55 | 9 | 0.11 | 0.17 | 13 | 38 | 0.34 | 0.72 |
| 67 | 43 | 25 | 0.64 | 0.37 | 0.58 | 8 | 0.12 | 0.19 | 10 | 31 | 0.32 | 0.72 | |
| no. PM5 | 144 | c87 | 55 | 0.60 | 0.38 | 0.63 | 20 | 0.14 | 0.23 | - | - | - | - |
| - | c58 | 35 | - | - | 0.60 | 8 | - | 0.14 | 15 | 40 | 0.38 | 0.69 | |
| no. PM6 | 125 | 80 | - | 0.64 | - | - | 11 | 0.09 | 0.14 | - | - | - | - |
| 106 | 67 | 36 | 0.63 | 0.34 | 0.54 | 8 | 0.08 | 0.12 | 17 | 50 | 0.34 | 0.75 | |
| 81 | 50 | 31 | 0.62 | 0.38 | 0.62 | 7 | 0.09 | 0.14 | 14 | 40 | 0.35 | 0.80 | |
| no. PM13 | 146 | 87 | - | 0.60 | - | - | 18 | 0.12 | 0.21 | - | - | - | - |
| 109 | 65 | 38 | 0.60 | 0.35 | 0.58 | 8 | 0.07 | 0.12 | 18 | 49 | 0.37 | 0.75 | |
| - | 52 | 32 | - | - | 0.62 | 7 | - | 0.13 | 12 | 37 | 0.32 | 0.71 | |
| no. PM3N1 | - | 74 | 39 | - | - | 0.53 | 11 | - | 0.15 | 20 | 54 | 0.37 | 0.73 |
| - | 53 | 31 | - | - | 0.58 | 8 | - | 0.15 | 14 | 37 | 0.38 | 0.70 | |
| no. PM3N2 | 118 | 75 | 38 | 0.64 | 0.32 | 0.51 | 10 | 0.08 | 0.13 | - | - | - | - |
| 114 | 72 | 38 | 0.63 | 0.33 | 0.53 | 9 | 0.08 | 0.13 | 20 | 54 | 0.37 | 0.75 | |
| 74 | 48 | 29 | 0.65 | 0.39 | 0.60 | 7 | 0.09 | 0.15 | 12 | 35 | 0.34 | 0.73 | |
| no. PM3N11 | 97 | 60 | 36 | 0.62 | 0.37 | 0.60 | 11 | 0.11 | 0.18 | - | - | - | - |
| 72 | 44 | 29 | 0.61 | 0.40 | 0.66 | 8 | 0.11 | 0.18 | 9 | 34 | 0.26 | 0.77 | |
| no. PMBb2 | - | 51 | 29 | - | - | 0.57 | 7 | - | 0.14 | 11 | 41 | 0.27 | 0.80 |
| - | 41 | 26 | - | - | 0.63 | 6 | - | 0.15 | 10 | 33 | 0.30 | 0.80 | |
| no. A-114 | 106.3 | 66.2 | - | 0.62 | - | - | 10.5 | 0.10 | 0.16 | 16.6 | 49.2 | 0.34 | 0.74 |
| - | 43.7 | 33.3 | - | - | 0.76 | 9.5 | - | 0.22 | 9.4 | 29.9 | 0.31 | 0.68 | |
| no. A-145 | 116.7 | 70.0 | 43.5 | 0.6 | 0.37 | 0.62 | 11.6 | 0.10 | 0.17 | 14.6 | 49.9 | 0.29 | 0.71 |
| 85.5 | 49.9 | - | 0.58 | - | - | 10.8 | 0.13 | 0.22 | 12.9 | 37.1 | 0.35 | 0.74 | |
| no. A-146 | 84.9 | 50.1 | - | 0.59 | - | - | 8.1 | 0.10 | 0.16 | 14.4 | 38.9 | 0.37 | 0.78 |
| 72.5 | 41.8 | - | 0.58 | - | - | - | - | - | 11.8 | 33.3 | 0.35 | 0.8 | |
| - | 26.2 | - | - | - | - | - | - | - | 6.7 | 20.5 | 0.33 | 0.78 | |
| no. AG-3 | 99.8 | 61.9 | - | 0.62 | - | - | 8.0 | 0.08 | 0.13 | 17.2 | 45.7 | 0.38 | 0.74 |
| no. AG-13 | 52.7 | 30.8 | 23.1 | 0.58 | 0.44 | 0.75 | 7.2 | 0.14 | 0.23 | 8.2 | 22.8 | 0.36 | 0.74 |
| - | 21.2 | - | - | - | - | 5.7 | - | 0.27 | 5.0 | 15.8 | 0.32 | 0.74 | |
| no. JM-34 | 117.1 | 73.1 | 47.8 | 0.62 | 0.41 | 0.65 | 12.3 | 0.10 | 0.17 | 22.5 | 51.3 | 0.44 | 0.7 |
| - | 32.9 | - | - | - | - | 7.5 | - | 0.23 | 9.1 | 25.4 | 0.36 | 0.77 | |
| no. JM-36 | 127 | 74 | 50 | 0.58 | 0.39 | 0.68 | - | - | - | - | - | - | - |
| no. M-8 | - | 67.0 | 39.1 | - | - | 0.58 | 10.0 | - | 0.15 | 15.7 | 48.0 | 0.33 | 0.72 |
| no. PUAB 5905 | - | 56.2 | - | - | - | - | 8.7 | - | 0.15 | 12.5 | 38.6 | 0.32 | 0.69 |
| - | 33.6 | - | - | - | - | 5.4 | - | 0.16 | 6.6 | 25.0 | 0.26 | 0.74 | |
| no. PUAB 48011 | 99.8 | 59.3 | - | 0.59 | - | - | 9.4 | 0.09 | 0.16 | 16.2 | 43.8 | 0.37 | 0.74 |
| no. PUAB 48013 | 113.0 | 73.0 | - | 0.65 | - | - | - | - | - | 16.5 | 51.4 | 0.32 | 0.7 |
| - | 44.9 | 30.4 | - | - | 0.68 | 8.4 | - | 0.19 | 11.1 | 33.8 | 0.33 | 0.75 | |
| - | 36.9 | - | - | - | - | 7.7 | - | 0.21 | 8.5 | 28.0 | 0.3 | 0.76 | |
| no. PUAB 48027 | 94.1 | 59.8 | - | 0.64 | - | - | 8.2 | 0.09 | 0.14 | 15.1 | 44.1 | 0.34 | 0.74 |
| no. PUAB 48040 | - | 48.9 | 32.3 | - | - | 0.66 | 8.5 | - | 0.17 | 13.9 | 36.5 | 0.38 | 0.75 |
| - | 39.2 | 27.6 | - | - | 0.7 | 8.1 | - | 0.21 | 10.8 | 30.5 | 0.35 | 0.78 | |
| - | 34.2 | 25.4 | - | - | 0.74 | 7.5 | - | 0.22 | 7.9 | 25.1 | 0.32 | 0.73 | |
| no. PUAB 48042 | 126 | 69 | 49 | 0.55 | 0.39 | 0.71 | 17 | 0.13 | 0.25 | - | - | - | - |
| c99 | 59 | - | 0.60 | - | - | 12 | 0.12 | 0.20 | 17 | 45 | 0.38 | 0.76 | |
| - | 44 | - | 0.44 | - | - | - | - | - | 13 | 33 | 0.39 | 0.75 | |
| no. PUAB 48044 | 120 | 68 | 45 | 0.57 | 0.38 | 0.66 | - | - | - | - | - | - | - |
| 99.2 | 61.7 | 35.1 | 0.62 | 0.35 | 0.57 | 8.3 | 0.08 | 0.13 | 18.7 | 44.6 | 0.42 | 0.72 | |
| 85.6 | 49.8 | 34.3 | 0.58 | 0.4 | 0.69 | 9.5 | 0.11 | 0.19 | 14.7 | 35.1 | 0.42 | 0.71 | |
| 119.3 | 40.9 | 31.4 | 0.34 | 0.26 | 0.77 | 8.4 | 0.07 | 0.21 | 11.0 | 29.6 | 0.37 | 0.72 | |
| no. PUAB 48046 | 136 | 80 | 60 | 0.59 | 0.44 | 0.75 | - | - | - | - | - | - | - |
| no. PUAB 48098 | - | 47.4 | 31.3 | - | - | 0.66 | 9.4 | - | 0.20 | 13.5 | 39.3 | 0.34 | 0.83 |
| no. PUAB 48128 | 153 | 90 | 49 | 0.59 | 0.32 | 0.54 | - | - | - | - | - | - | - |
| no. PUAB 48143 | - | 27.6 | 17.5 | - | - | 0.63 | 4.7 | - | 0.17 | 8.4 | 19.6 | 0.43 | 0.71 |
| no. PUAB 48163 | 60.8 | 35.1 | - | 0.58 | - | - | 7.6 | 0.13 | 0.22 | 8.0 | 25.5 | 0.31 | 0.73 |
| no. PUAB 48164 | 81.5 | 52.9 | 34.3 | 0.65 | 0.42 | 0.65 | 7.7 | 0.09 | 0.15 | 14.0 | 36.1 | 0.39 | 0.68 |
| 76.7 | 47.9 | 29.9 | 0.62 | 0.39 | 0.62 | 6.7 | 0.09 | 0.14 | 11.6 | 34.7 | 0.33 | 0.72 | |
| no. PUAB 48205 | - | 54.6 | 35.4 | - | - | 0.65 | 8.5 | - | 0.16 | 14.8 | 33.5 | 0.44 | 0.61 |
| no. PUAB 48215 | 109.8 | 69.5 | 37.2 | 0.63 | 0.34 | 0.53 | 8.9 | 0.08 | 0.13 | 14.7 | 51.6 | 0.28 | 0.74 |
| - | 51.0 | - | - | - | - | 8.9 | - | 0.17 | 11.3 | 37.2 | 0.3 | 0.73 | |
| - | 37.9 | - | - | - | - | 8.2 | - | 0.22 | 8.6 | 29.8 | 0.29 | 0.78 | |
| no. PUAB 48202 | - | 51.9 | 36.1 | - | - | 0.7 | 7.8 | - | 0.15 | 15.3 | 36.1 | 0.43 | 0.7 |
| - | 36.1 | 27.3 | - | - | 0.76 | - | - | - | 11.7 | 30.1 | 0.39 | 0.83 | |
| no. PUAB 48246 | - | 49.4 | 34.4 | - | - | 0.7 | - | - | - | 14.0 | 36.5 | 0.38 | 0.74 |
| no. PUAB 48254 | - | 63.9 | - | - | - | - | 9.6 | - | 0.15 | 12.4 | 46.1 | 0.27 | 0.72 |
| no. PUAB 48255 | - | 63.5 | 37.8 | - | - | 0.59 | 9.0 | - | 0.14 | 18.2 | 43.5 | 0.42 | 0.68 |
| - | 42.0 | 26.4 | - | - | 0.63 | 7.5 | - | 0.18 | 11.8 | 30.9 | 0.38 | 0.74 | |
| no. PUAB 48256 | - | 49.4 | 31.5 | - | - | 0.64 | 8.1 | - | 0.16 | 13.9 | 38.7 | 0.36 | 0.78 |
| no. PUAB 48261 | - | 71.9 | 44.0 | - | - | 0.61 | 9.2 | - | 0.13 | 19.1 | 46.6 | 0.41 | 0.65 |
| - | 65.3 | 40.6 | - | - | 0.62 | 8.8 | - | 0.13 | 17.6 | 42.2 | 0.42 | 0.65 | |
| - | 56.8 | 35.0 | - | - | 0.62 | 8.2 | - | 0.14 | 16.2 | 39.2 | 0.41 | 0.69 | |
| no. PUAB 48275 | - | 76.7 | - | - | - | - | 12.4 | - | 0.16 | 16.5 | 52.3 | 0.32 | 0.68 |
| no. PUAB 48292 | 128 | 76 | 56 | 0.59 | 0.44 | 0.74 | 17 | 0.13 | 0.22 | - | - | - | - |
| 111.8 | 68.6 | 38.5 | 0.61 | 0.34 | 0.56 | 10.5 | 0.09 | 0.15 | 14.6 | 49.6 | 0.29 | 0.72 | |
| 109.0 | 63.5 | 37.5 | 0.58 | 0.34 | 0.59 | 9.9 | 0.09 | 0.16 | 15.2 | 49.6 | 0.31 | 0.78 | |
| no. PUAB 48295 | - | 53.8 | - | - | - | - | 8.5 | - | 0.16 | 16 | 42 | 0.38 | 0.78 |
| no. PUAB 48366 | - | 65.2 | - | - | - | - | 11.8 | - | 0.18 | 15.3 | 48.3 | 0.32 | 0.74 |
| - | 61.4 | - | - | - | - | 9.4 | - | 0.15 | 14.2 | 47.1 | 0.3 | 0.77 | |
| no. PUAB 89788 | - | 66.0 | - | - | - | - | 10.3 | - | 0.16 | 16.1 | 45.5 | 0.35 | 0.69 |
| - | 60.0 | 34.4 | - | - | 0.57 | 9.9 | - | 0.16 | 15.1 | 42.8 | 0.35 | 0.71 | |
| - | 46.5 | 29.3 | - | - | 0.63 | 19.1 | - | 0.41 | 13.0 | 35.9 | 0.36 | 0.77 | |
| no. PUAB 89791 | - | 62.2 | 31.6 | - | - | 0.51 | 9.7 | - | 0.16 | 16.7 | 47.0 | 0.35 | 0.76 |
| no. PUAB 48009 ( & , 2006) | 85 | 52 | 33 | 0.61 | 0.39 | 0.63 | - | - | - | - | - | - | - |
| no. PUAB 48093 ( & , 2006) | 147 | 80 | 56 | 0.54 | 0.38 | 0.70 | - | - | - | - | - | - | - |
| no. PUAB 48095 ( & , 2006) | 65 | 42 | 26 | 0.65 | 0.40 | 0.62 | - | - | - | - | - | - | - |
| Specimen photo 11 (, 2007) | - | 73 | - | - | - | - | - | - | - | 20 | 58 | 0.34 | 0.79 |
| Specimen Fig. 3.12 ( , 2011) | - | 58 | - | - | - | - | 12 | - | 0.21 | 16 | 48 | 0.33 | 0.83 |
| Holotype of Nautilus verneuilli , 1870 | 118 | 76 | 38 | 0.64 | 0.32 | 0.50 | 2 | 0.02 | 0.03 | - | - | - | - |
| 112 | 71 | - | 0.63 | - | - | 4 | 0.04 | 0.06 | 18 | 54 | 0.33 | 0.76 | |
| 89 | 57 | - | 0.64 | - | - | 5 | 0.06 | 0.09 | 14 | 47 | 0.30 | 0.82 | |
| no. MB RE1962A | - | 47 | - | - | - | - | - | - | - | 10 | 32 | 0.31 | 0.68 |
| - | 45 | - | - | - | - | 7 | - | 0.16 | 12 | 32 | 0.38 | 0.71 |
Description:
the studied specimens show a relatively compressed section (E/H generally between 0.50 and 0.70), with slightly convex flanks,
with maximum thickness located in the lower third of the flanks. The ventral
region is relatively narrow; convex in the inner whorls (specimen no. PUAB
48158, Pl. XXIII ![]() , fig. 3), it quickly becomes slightly concave or flat, with
angular edges, or very slightly convex on the body chamber of some specimens
(holotype of Heminautilus tejeriensis
& , 2006, no. PUAB 48292; Pl. XXVI
, fig. 3), it quickly becomes slightly concave or flat, with
angular edges, or very slightly convex on the body chamber of some specimens
(holotype of Heminautilus tejeriensis
& , 2006, no. PUAB 48292; Pl. XXVI ![]() , fig. 1).
In the most internal whorls, rursiradiate fine growth riblets are visible on the
flanks and the ventral region where they form an angular chevron (no. PUAB
specimen 48158, Pl. XXXI
, fig. 1).
In the most internal whorls, rursiradiate fine growth riblets are visible on the
flanks and the ventral region where they form an angular chevron (no. PUAB
specimen 48158, Pl. XXXI ![]() , fig. 3); no other ornamentation is visible on the
phragmocone. On the body chamber of adult specimens, strongly rursiradiate ribs
appear in the upper half of the flanks, more or less broad and high (specimens
no. A-127, Pl. XVII
, fig. 3); no other ornamentation is visible on the
phragmocone. On the body chamber of adult specimens, strongly rursiradiate ribs
appear in the upper half of the flanks, more or less broad and high (specimens
no. A-127, Pl. XVII ![]() , fig. 2; no. MV 428.3,
Pl. XIX
, fig. 2; no. MV 428.3,
Pl. XIX ![]() , fig. 1; no. MV
428.2, Pl. XXI
, fig. 1; no. MV
428.2, Pl. XXI ![]() , fig. 1; no. PUAB 48042,
Pl. XXIII
, fig. 1; no. PUAB 48042,
Pl. XXIII ![]() , fig. 1).
, fig. 1).
The
suture line is sinuous (Fig. 8 ![]() ), with a wide and relatively deep lateral lobe (P/L
generally between 0.30 and 0.40) and a fairly narrow and angular lateral saddle,
becoming round with growth. The ventral lobe is shallow or absent.
), with a wide and relatively deep lateral lobe (P/L
generally between 0.30 and 0.40) and a fairly narrow and angular lateral saddle,
becoming round with growth. The ventral lobe is shallow or absent.
Click on thumbnail to enlarge the image.
Figure 8: Heminautilus saxbii (, 1848). Suture line of specimens no. BMNH 47019, right side (a-c), EM700, left side (d) and PM1, right side (e).
The
siphuncle, particularly visible in the specimen no. PMBb2 (Pl. XVII ![]() , fig. 3), is
located between the dorsum and the center of the section, near the dorsal
region.
, fig. 3), is
located between the dorsum and the center of the section, near the dorsal
region.
Variability:
the number of specimens of Heminautilus saxbii (, 1848) studied here, with a particularly
important sample from Josa, Province of Teruel (Spain), allows estimation of the
variability of the species. The variability essentially concerns the width of
the conch (Fig. 9 ![]() ), the shape of the ventral region, the ornamentation and the
plan of the suture line. The thickness of the shell varies from compressed forms
where E/H is about 0.50 (specimen no.
PUAB 89791) to wide forms where E/H reaches 0.76 (specimen no. PUAB 48044,
Pl. XXVI
), the shape of the ventral region, the ornamentation and the
plan of the suture line. The thickness of the shell varies from compressed forms
where E/H is about 0.50 (specimen no.
PUAB 89791) to wide forms where E/H reaches 0.76 (specimen no. PUAB 48044,
Pl. XXVI ![]() ,
fig. 2). The
shape of the ventral region varies from slightly concave to flat, even very
slightly convex. The ornamentation of the body chamber of the adult specimens is
also rather variable, being almost smooth (specimen no. PM13, Pl. XXII
,
fig. 2). The
shape of the ventral region varies from slightly concave to flat, even very
slightly convex. The ornamentation of the body chamber of the adult specimens is
also rather variable, being almost smooth (specimen no. PM13, Pl. XXII ![]() , fig. 1),
or with broad and well defined ribs (specimens no. MV 428.3,
Pl. XIX
, fig. 1),
or with broad and well defined ribs (specimens no. MV 428.3,
Pl. XIX ![]() , fig. 1;
no. MV 428.2, Pl. XXI , fig. 1), with intermediate forms with very visible but
narrower and weaker ribs (specimen no. PUAB 48042, Pl. XXIII
, fig. 1;
no. MV 428.2, Pl. XXI , fig. 1), with intermediate forms with very visible but
narrower and weaker ribs (specimen no. PUAB 48042, Pl. XXIII ![]() , fig. 1). Finally,
the suture line shows a lateral lobe of variable depth, with P/L
between 0.26 (specimen no. PM3N11, Pl. XXI , fig. 2) and 0.44 (specimen no.
JM-34, Pl. XIX
, fig. 1). Finally,
the suture line shows a lateral lobe of variable depth, with P/L
between 0.26 (specimen no. PM3N11, Pl. XXI , fig. 2) and 0.44 (specimen no.
JM-34, Pl. XIX ![]() , fig. 3), and a shallow to absent ventral lobe.
, fig. 3), and a shallow to absent ventral lobe.
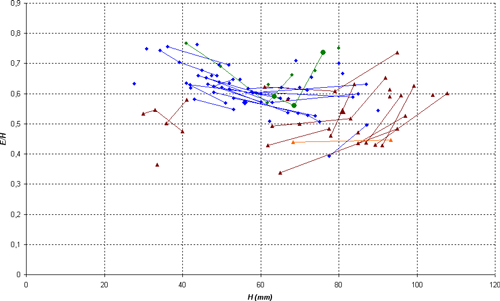
Click on thumbnail to enlarge the image.
Figure 9: E/H in function of H. In red, Heminautilus lallierianus (, 1841); in orange, Heminautilus aff. lallierianus (, 1841); in blue, Heminautilus saxbii (, 1848); in green, specimens attributed to Heminautilus tejeriensis & , 2006 [= H. saxbii ()] by & (2006). When taken at various diameters, measurements of the same specimen are connected by a line. The enlarged symbols represent the respective type specimens.
Discussion
and comparisons:
Heminautilus saxbii (,
1848) is particularly close to Heminautilus lallierianus
(, 1841). The latter can be distinguished from H. saxbii () by its clearly more sinuous suture line, showing
a deeper and narrower lateral lobe (P/L
generally near 0.35 - 0.50; Fig. 10 ![]() ; L/H
generally between 0.70 and 0.80 in H. saxbii (), between 0.60 and 0.75 in H. lallierianus
(); Fig. 11
; L/H
generally between 0.70 and 0.80 in H. saxbii (), between 0.60 and 0.75 in H. lallierianus
(); Fig. 11 ![]() )
and a wider and rounded lateral saddle, as
well as by the shape of the strongly concave ventral region which contains a
deep ventral lobe. The shell of H. lallierianus
() is also more compressed (Fig. 12
)
and a wider and rounded lateral saddle, as
well as by the shape of the strongly concave ventral region which contains a
deep ventral lobe. The shell of H. lallierianus
() is also more compressed (Fig. 12 ![]() )
than that of H.
saxbii (), with
E/H generally between 0.40 and 0.60 in
H. lallierianus ()
and between 0.50 and 0.70 in H. saxbii (). According to numerous authors these species are
synonyms (, 1850; F.-J.
& , 1858; , 1916; ,
1943)
while other later authors consider them distinct (, 1980, 1985, 2007; & ,
2006). In Europe both taxa occupy very close stratigraphical positions
within the Deshayesites forbesi Zone, Roloboceras
hambrovi Subzone. This leads us to two possible hypotheses: either the taxa
represent two morphological poles of the same relatively variable species, or
they represent two distinct species.
)
than that of H.
saxbii (), with
E/H generally between 0.40 and 0.60 in
H. lallierianus ()
and between 0.50 and 0.70 in H. saxbii (). According to numerous authors these species are
synonyms (, 1850; F.-J.
& , 1858; , 1916; ,
1943)
while other later authors consider them distinct (, 1980, 1985, 2007; & ,
2006). In Europe both taxa occupy very close stratigraphical positions
within the Deshayesites forbesi Zone, Roloboceras
hambrovi Subzone. This leads us to two possible hypotheses: either the taxa
represent two morphological poles of the same relatively variable species, or
they represent two distinct species.
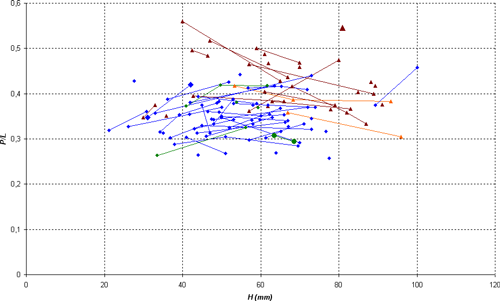
Click on thumbnail to enlarge the image.
Figure 10: P/L in function of H. In red, Heminautilus lallierianus (, 1841); in orange, Heminautilus aff. lallierianus (, 1841); in blue, Heminautilus saxbii (, 1848); in green, specimens attributed to Heminautilus tejeriensis & , 2006 [= H. saxbii ()] by & (2006). When taken at various diameters, measurements of the same specimen are connected by a line. The enlarged symbols represent the respective type specimens.
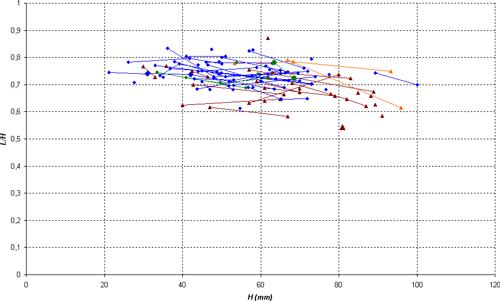
Click on thumbnail to enlarge the image.
Figure 11: L/H in function of H. In red, Heminautilus lallierianus (, 1841); in orange, Heminautilus aff. lallierianus (, 1841); in blue, Heminautilus saxbii (, 1848); in green, specimens attributed to Heminautilus tejeriensis & , 2006 [= H. saxbii ()] by & (2006). When taken at various diameters, measurements of the same specimen are connected by a line. The enlarged symbols represent the respective type specimens.
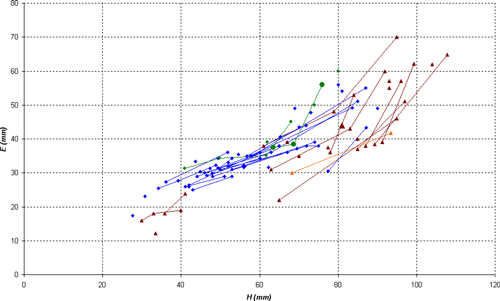
Click on thumbnail to enlarge the image.
Figure 12: E in function of H. In red, Heminautilus lallierianus (, 1841); in orange, Heminautilus aff. lallierianus (, 1841); in blue, Heminautilus saxbii (, 1848); in green, specimens attributed to Heminautilus tejeriensis & , 2006 [= H. saxbii ()] by & (2006). When taken at various diameters, measurements of the same specimen are connected by a line. The enlarged symbols represent the respective type specimens.
The study of numerous specimens coming from the section of Josa (Province of Teruel, Spain) establishes that, of these specimens, almost all are unambiguously morphologically attributable to H. saxbii () and that the morphologies of lallierianus () type are almost absent. These taxa do not appear to coexist and occupy slightly different stratigraphic positions in Europe within the Roloboceras hambrovi Subzone. On the Isle of Wight, the holotype and an important population of H. saxbii () were collected in the Lower Lobster Beds Formation (, written communication, 2013) and are thus situated in the bottom of the Roloboceras hambrovi Subzone (, 1985; et al., 1998). The population of H. saxbii () from the section of Josa comes from a slightly condensed level attributed to the Roloboceras hambrovi Subzone ( et al., 2007); however, the dominance of the genus Roloboceras , 1954, and the scarcity of the genus Megatyloceras , 1949, among Roloboceratinae of this level indicate a stratigraphical position in the bottom of the Roloboceras hambrovi Subzone (equivalent to the Lower Lobster Beds Formation of the Isle of Wight). On the other hand, the small population of H. lallierianus () of Le Teil and Viviers (Ardčche, France), where no typical specimen of H. saxbii () is known, is associated with numerous Megatyloceras and is probably situated at the top of the Roloboceras hambrovi Subzone (equivalent of the Upper Lobster Beds Formation of the Isle of Wight; et al., 1998).
So, although the two taxa are undoubtedly phyletically very close, they never coexist in the localities studied here. Consequently, H. lallierianus () represents a species separate from H. saxbii (), which it probably replaces within the Roloboceras hambrovi Subzone in Europe.
On
the other hand, the important sample from Josa allows us to consider Heminautilus
tejeriensis & ,
2006, as a junior synonym of H. saxbii (, 1848). In the original description, H.
tejeriensis &
was characterized essentially by the shape of its ventral region, flat or very
slightly convex, as well as by the absence of a ventral lobe, while H. saxbii
() shows a slightly concave to flat ventral region, with a
shallow ventral lobe. Other than the ventral region, the suture lines of both
taxa are identical. However, these slight differences appear to represent only
minor intraspecific variability within the one species. Indeed, we frequently
observe, on the same specimen [in particular in the holotype of H. saxbii ()], some suture lines forming a lobe at the
passage of the ventral region while others cross it straight, or even form a
slight convexity (specimen no. PUAB 48044,
Pl. XXV ![]() , fig. 4;
Pl. XXVI
, fig. 4;
Pl. XXVI ![]() ,
fig. 2). Most of the specimens attributed to H. tejeriensis
& show a flat ventral region
identical to that of the holotype of H. saxbii
(). Only the holotype of H. tejeriensis
& shows a very slight convexity that
solely affects the ventral region of the body chamber; this singular
characteristic does not appear to justify a differentiation at the species
level. Furthermore, all specimens attributed to H. tejeriensis & come from the Province of
Teruel (Spain) where they are associated with numerous clearly coeval specimens
of H. saxbii ().
The contemporaneity of these two taxa and minimal morphological differences lead
us to consider them as the expression of a slight intraspecific variability
within the species H. saxbii ().
,
fig. 2). Most of the specimens attributed to H. tejeriensis
& show a flat ventral region
identical to that of the holotype of H. saxbii
(). Only the holotype of H. tejeriensis
& shows a very slight convexity that
solely affects the ventral region of the body chamber; this singular
characteristic does not appear to justify a differentiation at the species
level. Furthermore, all specimens attributed to H. tejeriensis & come from the Province of
Teruel (Spain) where they are associated with numerous clearly coeval specimens
of H. saxbii ().
The contemporaneity of these two taxa and minimal morphological differences lead
us to consider them as the expression of a slight intraspecific variability
within the species H. saxbii ().
Also, Heminautilus
verneuilli (,
1870), whose holotype also comes from Josa, Province of Teruel (Spain), shows
characteristics identical to those of H. saxbii (). Following &
(2006), we consider this taxon as a junior synonym of H. saxbii
(). The original illustration of (1870) is
reproduced here (Pl. XXVI ![]() , fig. 3).
, fig. 3).
Heminautilus saxbii () is also close to H. boselliorum et al., 2012, and H. sanctaecrucis , 1980. The differences with these two species were highlighted in the pertinent sections.
Remarks:
the specimen no. MB RE1962A
(Seboruco section, Venezuela; Pl. XXIV ![]() ,
fig. 3) shows morphological and sutural characteristics that lead us to
attribute it to H. saxbii ().
However, the presence of Heminautilus etheringtoni
, 1946 (specimen no. MB RE1962a, Pl. XXXII
,
fig. 3) shows morphological and sutural characteristics that lead us to
attribute it to H. saxbii ().
However, the presence of Heminautilus etheringtoni
, 1946 (specimen no. MB RE1962a, Pl. XXXII ![]() , fig. 2), in the
same section makes this attribution tentative; this specimen may also represent
a slender variant of H. etheringtoni
. Only the study of a more representative population of this
last taxon would provide more certainty about its variability.
, fig. 2), in the
same section makes this attribution tentative; this specimen may also represent
a slender variant of H. etheringtoni
. Only the study of a more representative population of this
last taxon would provide more certainty about its variability.
Heminautilus lallierianus (, 1841)
Pl. XXVII ![]() ,
figs. 1-2;
Pl. XXVIII
,
figs. 1-2;
Pl. XXVIII ![]() ,
figs. 1-3;
Pl. XXIX
,
figs. 1-3;
Pl. XXIX ![]() ,
figs. 1-3;
,
figs. 1-3;
Pl. XXX ![]() ,
figs. 1-2;
Pl. XXXI
,
figs. 1-2;
Pl. XXXI ![]() ,
figs. 1-2;
Pl. XXXII
,
figs. 1-2;
Pl. XXXII ![]() , fig. 1;
, fig. 1;
Pl. XXXIII ![]() , fig. 1;
Pl. XXXIV
, fig. 1;
Pl. XXXIV ![]() , fig. 1;
Pl. XXXV
, fig. 1;
Pl. XXXV ![]() ,
figs. 1-2;
,
figs. 1-2;
Pl. XXXVI ![]() ,
fig. 1; Pl. XXXVII
,
fig. 1; Pl. XXXVII ![]() , fig. 1;
Pl. XXXVIII
, fig. 1;
Pl. XXXVIII ![]() , fig. 1;
, fig. 1;
Pl. XL ![]() , fig. 2;
Pl. XLI
, fig. 2;
Pl. XLI ![]() , fig. 1
, fig. 1
|
Nautilus lallierianus nov. sp. - , p. 318 |
||
|
non |
Nautilus lallierianus , 1841 - F.-J. & , p. 148, Pl. 19, fig. 6 (= H. sanctaecrucis , 1980). |
|
|
non |
Nautilus lallieri , 1841 - , p. 129, Pl. 17, figs. 2-6 [figs. 2-4, 6 = H. rangei (, 1922); fig. 5 = H. saxbii (, 1848)]. |
|
|
Heminautilus lallieri (, 1841) - , p. 90, Pl. 25, fig. 2. |
||
|
non |
Xenocheilus lallierianus (, 1841) - , p. 18, Pl. 4, fig. 1 (= H. sanctaecrucis , 1980). |
|
|
non |
Heminautilus lallierianus (, 1841) - & , p. 103, Fig. 7a-c; Pl. 2, fig. 3 (= H. ? japonicus sp. nov.). |
|
|
Heminautilus lallierianus (, 1841) - , Pl. 1, Figs. 3, 4c. |
||
|
non |
Heminautilus cf. H. lallierianus (, 1841) - et al., p. 30, Fig. 3; Pl. 5, fig. 1a-b; Pl. 6, fig. 1 (= H. ? japonicus sp. nov.). |
|
|
Heminautilus lallierianus (, 1841) - , p. 30, Pl. 1, figs. 1-5. |
||
|
non |
Heminautilus lallierianus (, 1841) - , p. 101, Pl. 1, figs. 1-3 [= ? H. saxbii (, 1848)]. |
|
|
Heminautilus lallierianus (, 1841) - , p. 10, photos 1-7, 9, 10. |
||
|
non |
Heminautilus lallierianus (, 1841) - , Fig. 3, A-C [= H. rangei (, 1922)]. |
|
|
Heminautilus lallierianus (, 1841) - et al., Fig. 4. |
||
|
Heminautilus lallierianus (, 1841) - et al., Pl. 1, fig. 2. |
Material studied (N = 17): lectotype no. MNHN.F.A32800, Early Aptian, top of the Deshayesites forbesi Zone?, Gurgy (Yonne, France); specimen no. UPMC-250, Early Aptian, top of the Deshayesites forbesi Zone?, Gurgy (Yonne, France); specimens no. tl028, tl029, tl056, tl069, tl078, tl079, tl080, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France); no. BRG098, BRG103, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, bed no. 12, Pierrelatte section (Drôme, France; cf. A. et al., 2009, Fig. 2); no. VI009, VI012, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France); no. MPP.B-EM.69, coll. FSM, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, bed no. 148, La Bédoule (Bouches du Rhône, France); no. PM95, coll. MNHN, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain); no. PUAB 89780, Early Aptian, probably Deshayesites forbesi Zone, El Vendrell, Garraf (Spain); no. MGB 22607, Early Aptian, Garraf (Spain).
Type:
the species was created by (1841) with no illustration and
without designation of holotype; a specimen was shown by (1980,
2007) with the description "type";
the same specimen was depicted anew by et al. (2012) with the description
"holotype".
It is the syntype no. MNHN.F.A32800,
coll. (no. 5573) stored in the Muséum
d'Histoire Naturelle of
Paris, which we therefore designate as the lectotype. It is shown in this work
as Pl. XXIX ![]() , fig. 1 and comes from the Early Aptian of Gurgy (Yonne, France).
, fig. 1 and comes from the Early Aptian of Gurgy (Yonne, France).
Geographical distribution: the species seems to be known with certainty only from France, Spain and Ethiopia.
Stratigraphical distribution: the lectotype and the topotypes no. 77-543, no. 85-3 (coll. MHNA) and no. UPMC-250 come from the Argiles ŕ Plicatules Formation of Gurgy (Yonne), where the accompanying ammonite fauna indicates a stratigraphical position between the top of the Deshayesites forbesi Zone and the Deshayesites deshayesi Zone (, 1925; & , 2004). However, the preservation of these four specimens, as calcareous moulds of relatively large size, differs from that typically encountered in the form of pyritic nucleus and could be characteristic of a particular level within the Argiles ŕ Plicatules. The holotype of Roloboceras transiens , 1961, with the same preservation and from the same locality, suggests a stratigraphical position of this level in the top of the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone. Specimens no. tl028, tl029, tl056, tl069, tl078, tl079 and VI009, collected in association with numerous representatives of the genus Megatyloceras , 1949, come from the top of the Roloboceras hambrovi Subzone ( et al., 1998). Specimens no. BRG098 and BRG103, from bed no. 12 of the Pierrelatte section, would come from the lower part of the Roloboceras hambrovi Subzone (A. et al., 2009, 2015). Specimen no. AMNH 25455 depicted by (1943) comes from the Harrar region (Ethiopia). The local geology was studied by et al. (1999), who show that in this region there is only a very short Aptian marine incursion, with sandy fluvial deposition before and after; the presence of Palorbitolina lenticularis (, 1805) and Praeorbitolina cormyi , 1964, allows us to determine a middle Early Aptian age for this formation ( et al., 2010). Heminautilus lallierianus (, 1841) is thus present in Europe in the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, in particular at the top of the subzone.
Dimensions (in mm): see Table 7.
Table 7: Measurements of Heminautilus lallierianus (, 1841).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Lectotype no. MNHN.F.A32800 | 124 | 81 | 44 | 0.55 | 0.30 | 0.54 | 9 | 0.06 | 0.11 | 24 | 44 | 0.55 | 0.54 |
| - | 68 | - | - | - | - | - | - | - | 20 | 40 | 0.50 | 0.59 | |
| - | 48 | - | - | - | - | - | - | - | 15 | 30 | 0.50 | 0.63 | |
| no. 77-543 | 176 | 104 | 62 | 0.59 | 0.35 | 0.60 | - | - | - | - | - | - | - |
| 112 | 70 | - | 0.63 | - | - | 10 | 0.09 | 0.14 | 22 | 48 | 0.46 | 0.69 | |
| no. 85-3 | - | 67 | 39 | - | - | 0.58 | 9 | - | 0.13 | 17 | 39 | 0.44 | 0.58 |
| - | 47 | - | - | - | - | 8 | - | 0.17 | 15 | 29 | 0.52 | 0.62 | |
| no. UPMC-250 | - | 42.7 | - | - | - | - | 6.8 | - | 0.16 | 11.8 | 29.9 | 0.39 | 0.70 |
| - | 66 | - | - | - | - | 9.6 | - | 0.15 | 16.8 | 43.8 | 0.38 | 0.66 | |
| no. tl028 | 67 | 40 | 19 | 0.60 | 0.28 | 0.48 | 12 | 0.18 | 0.30 | - | - | - | - |
| 55 | 33 | 18 | 0.60 | 0.33 | 0.55 | 6 | 0.11 | 0.18 | 9 | 24 | 0.38 | 0.73 | |
| 49 | 30 | 16 | 0.61 | 0.33 | 0.53 | 6 | 0.12 | 0.20 | 8 | 23 | 0.35 | 0.77 | |
| no. tl029 | 154 | 96 | 57 | 0.62 | 0.37 | 0.59 | 13 | 0.08 | 0.14 | - | - | - | - |
| 122 | 87 | 38 | 0.71 | 0.31 | 0.44 | 8 | 0.07 | 0.09 | 18 | 54 | 0.33 | 0.62 | |
| 119 | 82 | - | 0.69 | - | - | 6 | 0.05 | 0.07 | 19 | 53 | 0.36 | 0.65 | |
| - | 68 | - | - | - | - | - | - | - | 20 | 48 | 0.42 | 0.71 | |
| no. tl056 | 153 | 93 | 55 | 0.61 | 0.36 | 0.59 | 13 | 0.08 | 0.14 | - | - | - | - |
| 135 | 89 | - | 0.66 | - | - | 10 | 0.07 | 0.11 | 24 | 60 | 0.40 | 0.67 | |
| - | 57 | - | - | - | - | c6 | - | 0.11 | 20 | 43 | 0.47 | 0.75 | |
| no. tl069 | 159 | 97 | c51 | 0.61 | 0.32 | 0.53 | 11 | 0.07 | 0.11 | - | - | - | - |
| c132 | 85 | c37 | 0.64 | 0.28 | 0.44 | 6 | 0.05 | 0.07 | 23 | 57 | 0.40 | 0.67 | |
| no. tl078 | 173.9 | 108 | 64.9 | 0.62 | 0.37 | 0.60 | 11.3 | 0.06 | 0.10 | - | - | - | - |
| 128.9 | 89.3 | 38.4 | 0.69 | 0.30 | 0.43 | 7.4 | 0.06 | 0.08 | 23.4 | 56 | 0.42 | 0.63 | |
| no. tl079 | - | 88.3 | - | - | - | - | 7.4 | - | 0.08 | 24.7 | 58.1 | 0.43 | 0.66 |
| no. tl080 | - | 99.3 | 62.1 | - | - | 0.63 | 15.3 | - | 0.15 | - | - | - | - |
| 139.1 | 91.2 | 39.1 | 0.66 | 0.28 | 0.43 | 11.6 | 0.08 | 0.13 | 20.1 | 53.4 | 0.38 | 0.59 | |
| no. BRG098 | - | 41.1 | 23.8 | - | - | 0.58 | 6.6 | - | 0.16 | - | - | - | - |
| - | 35.9 | 18 | - | - | 0.50 | 4.8 | - | 0.13 | 9.7 | 27.6 | 0.35 | 0.77 | |
| no. BRG103 | 134.3 | 77.5 | 37.5 | 0.58 | 0.28 | 0.48 | 7.5 | 0.06 | 0.10 | - | - | - | - |
| - | 61.8 | 26.5 | - | - | 0.43 | 6.2 | - | 0.10 | 20.1 | 53.9 | 0.37 | 0.87 | |
| no. VI009 | 148 | 84 | 53 | 0.57 | 0.36 | 0.63 | 15 | 0.10 | 0.18 | - | - | - | - |
| 122 | 78 | c36 | 0.64 | 0.30 | 0.46 | 8 | 0.07 | 0.10 | 19 | 52 | 0.37 | 0.67 | |
| no. VI012 | 153 | 92 | 60 | 0.60 | 0.39 | 0.65 | 9 | 0.06 | 0.10 | - | - | - | - |
| 128 | 83 | 43 | 0.65 | 0.34 | 0.52 | 5 | 0.04 | 0.06 | 22 | 60 | 0.37 | 0.72 | |
| - | 63 | 31 | - | - | 0.49 | - | - | - | 18 | 47 | 0.38 | 0.75 | |
| no. MPP.B-EM.69 | 58 | 33.5 | 12.2 | 0.58 | 0.21 | 0.36 | 4.9 | 0.08 | 0.15 | - | - | - | - |
| no. PM95 | 159 | 95 | 70 | 0.60 | 0.44 | 0.74 | 17 | 0.11 | 0.18 | - | - | - | - |
| 128 | 79 | 48 | 0.62 | 0.38 | 0.61 | 13 | 0.10 | 0.16 | 19.5 | 52 | 0.38 | 0.66 | |
| - | 61 | 38 | - | - | 0.62 | 10 | - | 0.16 | 17 | 42 | 0.40 | 0.69 | |
| no. PUAB 89780 | - | 46.5 | - | - | - | - | - | - | - | 16.4 | 33.9 | 0.48 | 0.73 |
| - | 42.5 | - | - | - | - | - | - | - | 15.8 | 31.8 | 0.5 | 0.75 | |
| no. AMNH 25455 (, 1943) | 146 | 93 | 57 | 0.64 | 0.39 | 0.61 | - | - | - | - | - | - | - |
| 116 | 80 | - | 0.69 | - | - | - | - | - | 28 | 59 | 0.47 | 0.74 | |
| - | 57 | - | - | - | - | - | - | - | 13 | 36 | 0.36 | 0.63 | |
| no. 12572 | 145 | 85 | 40 | 0.59 | 0.28 | 0.47 | - | - | - | - | - | - | - |
| 90 | 61 | - | 0.68 | - | - | - | - | - | 19 | 39 | 0.49 | 0.64 | |
| - | 35 | - | - | - | - | 4 | - | 0.11 | - | - | - | - | |
| no. 12573 | 160 | 95 | 46 | 0.59 | 0.29 | 0.48 | - | - | - | - | - | - | - |
| c95 | 65 | 22 | 0.68 | 0.23 | 0.34 | - | - | - | 18 | 42 | 0.43 | 0.65 | |
| - | 40 | - | - | - | - | 6.5 | - | 0.16 | 14 | 25 | 0.56 | 0.63 | |
| Specimen photo 7 (, 2007) | c121 | 69 | - | 0.57 | - | - | - | - | - | - | - | - | - |
| - | 62 | - | - | - | - | - | - | - | 21 | 45 | 0.47 | 0.73 | |
| no. MGB 22607 ( et al., 2008) | c113 | 70 | 35 | 0.62 | 0.31 | 0.50 | 6 | 0.05 | 0.09 | 22 | 47 | 0.47 | 0.67 |
| - | 59 | - | - | - | - | - | - | - | 21 | 42 | 0.50 | 0.71 |
Description: Heminautilus
lallierianus (, 1841) has a large shell (D = 176 mm, specimen no. 77-543,
Pl. XXX ![]() , fig. 1), compressed (E/H
generally between 0.40 and 0.60; Fig. 9
, fig. 1), compressed (E/H
generally between 0.40 and 0.60; Fig. 9 ![]() ), with slightly rounded flanks, of
maximum thickness in the lower third of the flanks. The initial whorls are
unknown; from a small diameter, the ventral region is narrow, strongly concave
(specimen no. MPP.B-EM.69, Pl. XXXVIII
), with slightly rounded flanks, of
maximum thickness in the lower third of the flanks. The initial whorls are
unknown; from a small diameter, the ventral region is narrow, strongly concave
(specimen no. MPP.B-EM.69, Pl. XXXVIII ![]() , fig. 3), lined with angular ventro-lateral
edges. On the body chamber of adult specimens, the concavity of the ventral
region frequently becomes less pronounced (no. 77-543, Pl. XXIX
, fig. 3), lined with angular ventro-lateral
edges. On the body chamber of adult specimens, the concavity of the ventral
region frequently becomes less pronounced (no. 77-543, Pl. XXIX ![]() , fig. 2; no. tl078,
Pl. XXXV
, fig. 2; no. tl078,
Pl. XXXV ![]() , fig. 2; no. tl029,
Pl. XXXVI
, fig. 2; no. tl029,
Pl. XXXVI ![]() ,
fig. 1). No ornamentation is present on the phragmocone. On the body chamber,
the ornamentation shows some variability. Some complete adult specimens are
completely smooth (no. BRG103, Pl. XXXI
,
fig. 1). No ornamentation is present on the phragmocone. On the body chamber,
the ornamentation shows some variability. Some complete adult specimens are
completely smooth (no. BRG103, Pl. XXXI ![]() ,
fig. 2; no. tl029, Pl. XXXV
,
fig. 2; no. tl029, Pl. XXXV ![]() , fig. 1; no. tl056,
Pl. XXXI
, fig. 1; no. tl056,
Pl. XXXI ![]() ,
fig. 1; no. VI009, Pl. XXXII
,
fig. 1; no. VI009, Pl. XXXII ![]() , fig. 1; no. tl069,
Pl. XXXIII
, fig. 1; no. tl069,
Pl. XXXIII ![]() ,
fig. 1), while others show at the end of the body chamber broad and strongly
rursiradiate ribs, restricted to the upper part of the flanks (no. PUAB
89780, Pl. XXVII
,
fig. 1), while others show at the end of the body chamber broad and strongly
rursiradiate ribs, restricted to the upper part of the flanks (no. PUAB
89780, Pl. XXVII ![]() , fig. 1; no. PM95,
Pl. XXXVIII
, fig. 1; no. PM95,
Pl. XXXVIII ![]() , fig. 1; no. 77-543,
Pl. XXX
, fig. 1; no. 77-543,
Pl. XXX ![]() ,
fig. 1; no. tl078, Pl. XXXIV
,
fig. 1; no. tl078, Pl. XXXIV ![]() , fig. 1).
, fig. 1).
The suture line is strongly sinuous (Fig. 13 ![]() ), with a
fairly wide and deep lateral lobe (Fig. 11
), with a
fairly wide and deep lateral lobe (Fig. 11 ![]() ; P/L generally between 0.35 and 0.50,
Fig. 10
; P/L generally between 0.35 and 0.50,
Fig. 10 ![]() ), a wide and rounded
lateral saddle and a well defined, angular ventral lobe. In some specimens, we
observe the space between the last suture lines narrowing, characteristic of the
adult stage (no. PM95, Pl. XXXVIII
), a wide and rounded
lateral saddle and a well defined, angular ventral lobe. In some specimens, we
observe the space between the last suture lines narrowing, characteristic of the
adult stage (no. PM95, Pl. XXXVIII ![]() , fig. 1; no. tl056,
Pl. XXXI
, fig. 1; no. tl056,
Pl. XXXI ![]() , fig. 1). The
siphuncle, very visible in the lectotype no. MNHN.F.A32800 (Pl. XXIX
, fig. 1). The
siphuncle, very visible in the lectotype no. MNHN.F.A32800 (Pl. XXIX ![]() ,
fig. 1) and the specimen no. 85-3 (Pl. XXVII
,
fig. 1) and the specimen no. 85-3 (Pl. XXVII ![]() , fig. 2), is placed very low on the
section, near the dorsal edge.
, fig. 2), is placed very low on the
section, near the dorsal edge.
Click on thumbnail to enlarge the image.
Figure 13: Heminautilus lallierianus (, 1841). Suture line of specimens no. MNHN.F.A32800, left side (a-b), PM95, right side (c) and tl056, left side (d).
Discussion and comparisons: Heminautilus lallierianus (, 1841) is very similar to H. saxbii (, 1848). Relationships between these two taxa were discussed in the section on H. saxbii (, 1848).
The specific characteristics of H. lallierianus (), in particular its strongly concave and narrow ventral region, as well as its very sinuous suture line with a deep lateral lobe, readily distinguish it from other species of the genus Heminautilus , 1927.
Heminautilus aff. lallierianus (, 1841)
Pl. XXXIX ![]() , fig. 1;
Pl. XL
, fig. 1;
Pl. XL ![]() , fig. 1
, fig. 1
|
2015 Heminautilus cf. lallierianus (, 1841) - et al., p. 209. |
Material studied (N = 2): specimen no. IGM 10050, Early Aptian, Dufrenoyia justinae Zone, bed no. 15, Mina Texali, Santa Isabel Atenayuca, Puebla State (Mexico); specimen no. MPL1, Early Aptian, Dufrenoyia justinae Zone, Sierra del Rosario, Durango State (Mexico).
Geographical distribution: Heminautilus aff. lallierianus (, 1841) is known only in Mexico.
Stratigraphical distribution: the specimen no. IGM 10050 comes from the Dufrenoyia justinae Zone (= top of the Dufrenoyia furcata Zone, et al., 2012, 2015). The specimen no. MPL1 comes from the La Peńa Formation, corresponding to a stratigraphical interval ranging from the Dufrenoyia justinae Zone to the lowermost part of the Caseyella aguilerae Zone [personal observations of one of us (J.A. ); cf. , 1989; et al., 2013]. It was found ex-situ in a section that can be attributed to the Dufrenoyia justinae Zone.
Dimensions (in mm): see Table 8.
Table 8: Measurements of Heminautilus aff. lallierianus (, 1841).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| no. IGM 10050 | 146.8 | 93.9 | 47.6 | 0.64 | 0.32 | 0.51 | c7.8 | 0.05 | 0.08 | 27.1 | 61.8 | 0.44 | 0.66 |
| - | 68.3 | 30 | - | - | 0.44 | 7.4 | - | 0.11 | 20.9 | 49.5 | 0.42 | 0.72 | |
| - | 53.3 | - | - | - | - | - | - | - | 16.7 | 39.9 | 0.42 | 0.75 | |
| no. MPL1 | 180.0 | 95.0 | - | - | - | - | - | - | - | - | - | - | - |
| - | 96.0 | - | - | - | - | - | - | - | 18.0 | 59.0 | 0.31 | 0.61 | |
| - | 67.0 | - | - | - | - | - | - | - | 19.0 | 53.0 | 0.36 | 0.79 |
Description: Heminautilus
aff. lallierianus (, 1841)
has a relatively compressed section, with rounded flanks, of maximum thickness
close to the umbilical edge. The ventral region is narrow, clearly concave and
lined with two angular ventro-lateral edges. No ornamentation is visible in the
specimen no. IGM 10050; in the specimen no. MPL1 some broad and strongly
rursiradiate ribs are observable on the top of the flanks at the end of the body
chamber. Heminautilus
aff. lallierianus ()
can reach a large size (D = 180 mm in specimen no. MPL1, Pl. XL ![]() , fig. 1).
, fig. 1).
The
suture line is very sinuous (Fig. 14 ![]() ), with a rounded and well developed lateral
saddle and a deep lateral lobe (P/L
between 0.31 and 0.44). The ventral lobe is well defined and angular in shape.
The position of the siphuncle is not visible.
), with a rounded and well developed lateral
saddle and a deep lateral lobe (P/L
between 0.31 and 0.44). The ventral lobe is well defined and angular in shape.
The position of the siphuncle is not visible.
Click on thumbnail to enlarge the image.
Figure 14: Heminautilus aff. lallierianus (, 1841). Suture line of specimen no. IGM 10050, left side.
Discussion and comparisons: Heminautilus aff. lallierianus (, 1841) is very close to H. lallierianus () and can be distinguished from it only by a slightly wider lateral saddle and a ventral region which is less concave. However, its stratigraphical position is clearly higher, in the Dufrenoyia justinae Zone (= top of the Dufrenoyia furcata Zone, et al., 2012, 2015), while H. lallierianus () is known only in the Deshayesites forbesi Zone, Roloboceras hambrovi Subzone. This unusual stratigraphical position, as well as its very distant geographical distribution, makes its incorporation into 's species uncertain, with additional data on specimens of intermediate stratigraphical position needed.
H. aff. lallierianus () can be easily distinguished from the other American species of the genus, H. etheringtoni , 1946, and H. stantoni (, 1940). It differs from H. etheringtoni by its narrower section and ventral region, from H. stantoni (, 1940) by the absence of ribbing on the phragmocone, and from both taxa by its clearly more concave ventral region and its different suture line, with a deep lateral lobe and a well developed lateral saddle.
Heminautilus etheringtoni , 1946
Pl. XXX ![]() , fig. 3;
Pl. XXXII
, fig. 3;
Pl. XXXII ![]() , fig.
2; Pl. XLII
, fig.
2; Pl. XLII ![]() , fig. 1;
, fig. 1;
Pl. XLIII ![]() , fig. 1;
Pl. XLIV
, fig. 1;
Pl. XLIV ![]() ,
figs. 1-2;
Pl. XLV
,
figs. 1-2;
Pl. XLV ![]() ,
figs. 1-2
,
figs. 1-2
|
Heminautilus etheringtoni nov. sp. - , p. 432, Pl. 62, fig. 4; Pl. 63, figs. 3, 5-7; Pl. 64, figs. 1-3. |
||
|
Heminautilus etheringtoni , 1946 - , p. 576, Figs. 1-3. |
||
|
Heminautilus etheringtoni , 1946 - , p. 104, Pl. 4, fig. 8; Pl. 5, fig. 9. |
||
|
? |
Heminautilus etheringtoni , 1946 - et al., Fig. 3c. |
Material studied (N = 4): specimens no. M3, 124, coll. UNCB, Early Aptian, Quebrada el Cobre, Payandé, Tolima department (Colombia); no. UN-DG-NT-001, coll. UNCB, Early Aptian, Firavitoba, Boyacá department (Colombia); no. MB RE1962a, Early Aptian, Tibú Formation, Seboruco section, east of Mérida (Venezuela).
Type: the holotype
(by original designation) is specimen no. 35731 (coll. MPUC) shown in
(1946: Pl. 62, fig. 4; Pl. 63, fig. 6; Pl. 64, fig. 3), stored in the
collections of the Museum of Paleontology, University of California. It comes
from locality A of (1946), situated between El Valle and Payandé
in Tolima department (Colombia)
and is depicted anew in this work Pl. XLII ![]() , fig. 1;
Pl. XLIII
, fig. 1;
Pl. XLIII ![]() , fig. 1.
, fig. 1.
Geographical distribution: the species is known in Colombia and Venezuela.
Stratigraphical distribution: according to , 1946, the holotype comes from locality A, associated with representatives of the genus Cheloniceras , 1903. It is consequently possible to attribute it a stratigraphical position near the top of the Early Aptian. Also, specimen no. MB RE1962a comes from the Seboruco section (Venezuela) which, according to (1982, p. 19), yielded ammonites of the genus Roloboceras , 1954, but which more likely belong to the genus Cheloniceras , 1903; its stratigraphical position is also near the top of the Early Aptian. Specimen no. F-15 (coll. MGNR) illustrated by (1979) comes from the uppermost part of the Early Aptian, Dufrenoyia sanctorum-Stoyanowiceras treffryanus Assemblage zone. The stratigraphical position of the specimens no. M3, 124 and UN-DG-NT-001 is not known.
Heminautilus etheringtoni , 1946, is thus a species present at the top of the Early Aptian (= Dufrenoyia furcata Zone); its presence at a lower stratigraphical level in the Early Aptian is possible but remains hypothetical with the data in our possession.
Dimensions (in mm): see Table 9.
Table 9: Measurements of Heminautilus etheringtoni , 1946.
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. 35731 | 153 | 86 | 77 | 0.56 | 0.5 | 0.9 | 32 | 0.21 | 0.37 | - | - | - | - |
| 116 | 65 | - | 0.56 | - | - | - | - | - | 17 | 45 | 0.38 | 0.69 | |
| - | 53 | 59 | - | - | 1.11 | - | - | - | 12 | 38 | 0.32 | 0.72 | |
| no. 124 | 105 | - | - | - | - | - | - | - | - | - | - | - | - |
| - | 50.2 | - | - | - | - | - | - | - | 9.2 | 33.2 | 0.28 | 0.66 | |
| - | 32 | - | - | - | - | - | - | - | 5.8 | 23.2 | 0.25 | 0.73 | |
| no. M3 | 80 | - | - | - | - | - | - | - | - | - | - | - | - |
| - | 48.9 | - | - | - | - | - | - | - | 6.6 | 33.1 | 0.2 | 0.68 | |
| - | 42.2 | - | - | - | - | - | - | - | 8.3 | 32.3 | 0.26 | 0.77 | |
| no. UN-DG-NT-001 | 154 | - | - | - | - | - | - | - | - | - | - | - | - |
| - | 72.3 | - | - | - | - | - | - | - | 15.3 | 53.8 | 0.28 | 0.74 | |
| - | 65.1 | - | - | - | - | - | - | - | 13.6 | 49.5 | 0.27 | 0.76 | |
| no. 35733 (, 1946) | 53 | 31 | 25 | 0.58 | 0.47 | 0.81 | - | - | - | - | - | - | - |
| - | - | - | - | - | - | - | - | - | 3.5 | 12 | 0.29 | - | |
| no. F-15 (, 1979) | 115 | 75 | 52 | 0.65 | 0.45 | 0.69 | 14 | 0.12 | 0.19 | - | - | - | - |
| - | - | - | - | - | - | - | - | - | 6 | 20 | 0.30 | - | |
| no. MB RE1962a | - | 78 | 49 | - | - | 0.63 | 18 | - | 0.23 | - | - | - | - |
| - | 60 | - | - | - | - | 14 | - | 0.23 | 12 | 53 | 0.23 | 0.88 |
Description:
specimens illustrated by (1946) show a fairly compressed section
in the internal whorls, which widens through its ontogeny, particularly on the
adult body chamber (holotype no. 35731, Pl. XLIII ![]() , fig. 1; specimen no. MB
RE1962a, Pl. XXXII
, fig. 1; specimen no. MB
RE1962a, Pl. XXXII ![]() , fig. 2). The flanks, rounded in the internal whorls, quickly
become flat and convergent towards the ventral region, with maximum thickness
situated near the umbilical edge. The ventral region is fairly wide and rounded
in the internal whorls; it widens and becomes gradually flat with a relatively
angular ventro-lateral edge on the body chamber.
, fig. 2). The flanks, rounded in the internal whorls, quickly
become flat and convergent towards the ventral region, with maximum thickness
situated near the umbilical edge. The ventral region is fairly wide and rounded
in the internal whorls; it widens and becomes gradually flat with a relatively
angular ventro-lateral edge on the body chamber.
The ornamentation is formed by ribs and growth lines. Ribs, rounded, wide and weak, are present on the top of the flanks of the holotype's body chamber, where the shell is preserved. They are convex and become gradually strongly rursiradiate near the ventral region. Growth lines are visible on the ventral region; they are weak and form an angular sinus.
The
suture line is fairly sinuous (Fig. 15 ![]() ), with a very wide and relatively deep
lateral lobe (P/L between 0.32 and
0.38 in the holotype). The ventral lobe is shallow. The siphuncle is placed near
the dorsal edge of the section [specimen no. M3 (coll. UNCB), Pl. XLIV
), with a very wide and relatively deep
lateral lobe (P/L between 0.32 and
0.38 in the holotype). The ventral lobe is shallow. The siphuncle is placed near
the dorsal edge of the section [specimen no. M3 (coll. UNCB), Pl. XLIV ![]() , fig. 2; no.
124
(coll. UNCB), Pl. XLV
, fig. 2; no.
124
(coll. UNCB), Pl. XLV ![]() , fig. 2].
, fig. 2].
Click on thumbnail to enlarge the image.
Figure 15: Heminautilus etheringtoni , 1946. Suture line of specimens no. 35731, right side (a-b) and 35733, left side (c).
Discussion and comparisons: Heminautilus etheringtoni , 1946, presents affinities with H. ? tyosiensis ( & , 1953), a species with a comparable shape of the body chamber section, associated with a wide ventral region, ornamented by growth lines forming an angular sinus. However, H. ? tyosiensis ( & ) can be distinguished by its clearly more concave ventral region and its different ribbing, well defined on the phragmocone and absent on the body chamber. Its suture line is also different, with a more developed lateral saddle and a clearly deeper lateral lobe [P/L between 0.40 and 0.50 in H. ? tyosiensis ( & )].
H. etheringtoni is also rather close to H. saxbii (, 1848), particularly in its similar ornamentation and suture line, but can be distinguished by its section and its much wider ventral region, as well as by a much more rounded ventro-lateral shoulder, especially on the phragmocone.
H. etheringtoni can also be compared to Josanautilus lacerdae (, 1870) by its section and the form of its ventral region, but it can readily be distinguished from it by the absence of a median keel and of longitudinal ornamentation on the ventral region.
Heminautilus stantoni (, 1940)
Pl. XXXVI ![]() ,
figs. 2-3
,
figs. 2-3
|
Vorticoceras stantoni nov. sp. - , p. 1076, Pl. 68, figs. 4-5. |
Type: the holotype
(by original designation) is specimen no. USNM103260, shown in
(1940) Pl. 68, figs. 4-5, stored in the collections of the United States
National Museum, Washington, and coming from the locality M15 (,
1940, p. 988), near Murfreesboro, Arkansas (USA). It is depicted anew in this
work in Pl. XXXVI ![]() , fig. 3. A plaster cast of it (no. 35688, coll. BEG) is also shown in
Pl. XXXVI
, fig. 3. A plaster cast of it (no. 35688, coll. BEG) is also shown in
Pl. XXXVI ![]() , fig. 2.
, fig. 2.
Geographical distribution: the species is known only by its holotype, coming from Arkansas (USA).
Stratigraphical distribution: the holotype comes from the Dierks Limestone, Trinity Group of Arkansas. (1940) indicates in this formation the presence of representatives of the genus Pseudosaynella , 1923. In Mexico and in the south of the USA, the genus Pseudosaynella appears to occupy a restricted stratigraphical position, limited to the uppermost part of the Early Aptian (, 1940, p. 976; et al., 2015). Furthermore, (1945) reported the presence of the genus Pseudosaynella , associated with Dufrenoyia & , 1915, at the top of the Pine Island Shale (lateral equivalent in Louisiana of the Dierks Limestone). These observations allow us to attribute to Heminautilus stantoni (, 1940) a stratigraphical position at the top of the Early Aptian, confidently placed in the Dufrenoyia justinae Zone (= top of the Dufrenoyia furcata Zone, et al., 2012, 2013, 2015).
Dimensions (in mm): see Table 10.
Table 10: Measurements of Heminautilus stantoni (, 1940).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. 1162 | 114 | 69 | 40 | 0.61 | 0.35 | 0.58 | - | - | - | 16 | 51 | 0.31 | 0.74 |
| 98 | 60 | - | 0.61 | - | - | - | - | - | 13 | 43 | 0.3 | 0.72 |
Description: the holotype no. USNM103260 consists of an incomplete phragmocone. It shows a fairly compressed section, with flanks clearly convex and bulging in their lower half and otherwise flat, converging to the ventral region. The ventral region is fairly wide and seems slightly concave, lined with two well defined shoulders.
The ornamentation is formed of spaced, wide, rursiradiate and rounded ribs, becoming higher and thus more visible in the upper half of the flanks.
The
suture line (Fig. 16 ![]() )
is sinuous, with a wide and rounded lateral saddle, a well
developed, fairly deep lateral lobe (P/L
close to 0.30) and a narrow and well defined ventral lobe. The position of the
siphuncle is not known.
)
is sinuous, with a wide and rounded lateral saddle, a well
developed, fairly deep lateral lobe (P/L
close to 0.30) and a narrow and well defined ventral lobe. The position of the
siphuncle is not known.
Click on thumbnail to enlarge the image.
Figure 16: Heminautilus stantoni (, 1940). Suture line of specimen no. USNM103260, left side.
Discussion and comparisons: Heminautilus stantoni (, 1940) is relatively close to H. saxbii (, 1848) in its section, the shape of its ventral region and its suture line. H. stantoni () can be distinguished from H. saxbii () by its ribbing, consisting of very wide ribs and present on the phragmocone, which is only visible on the body chamber in H. saxbii (). Its suture line also contains a lateral saddle more developed than the one of H. saxbii ().
Heminautilus stantoni () also shows some affinities with H. lallierianus (, 1841), in particular in the presence of a well developed lateral saddle. It differs, however, from H. lallierianus () in the presence of well defined ribbing on the phragmocone, a clearly shallower lateral lobe and a less concave ventral region.
Heminautilus stantoni () can be distinguished immediately from other species of the genus Heminautilus , 1927, by its section, the shape of the ribbing and the presence of ribbing on the phragmocone.
Heminautilus ? japonicus sp. nov.
Pl. XXXVIII ![]() , fig. 2;
Pl. XLIX
, fig. 2;
Pl. XLIX ![]() , fig. 2;
Pl. L
, fig. 2;
Pl. L ![]() , fig. 3
, fig. 3
|
Heminautilus lallierianus (, 1841) - & , p. 103, Fig. 7a-c; Pl. 2, fig. 3. |
||
|
Heminautilus cf. H. lallierianus (, 1841) - et al., p. 30, Fig. 3; Pl. 5, fig. 1a-b; Pl. 6, fig. 1. |
Holotype: specimen
no. SH-1 (coll. E. Shimizu), illustrated by et al. (1984) and depicted anew here in
Pl. XLIX ![]() , fig. 2 and
Pl. L
, fig. 2 and
Pl. L ![]() ,
fig. 3. A cast is deposited in the National Science Museum in Tokyo, Japan under
the no. NSM-PM 9381.
,
fig. 3. A cast is deposited in the National Science Museum in Tokyo, Japan under
the no. NSM-PM 9381.
Origin of the name: the name comes from Japan, the origin of the holotype.
Type locality: according to et al. (1984), the holotype comes from locality 203 of the sector of Sanchu, Kwanto Moutains (Gunma Prefecture, Japan).
Type strata: the presence, in the horizon where the holotype comes from, of Phylloceras sp., Barremites aff. strettostoma (, 1883), Pseudohaploceras japonicum et al., 1984, and Heteroceras aff. astierianum , 1851, allows us to attribute Heminautilus ? japonicus sp. nov. to the latest Barremian (= Imerites giraudi Zone).
Geographical distribution: this taxon is known only in Japan. The holotype no. SH-1 comes from the
Kwanto Moutains (Gunma Prefecture) and the specimen studied by
& (1976) comes from the north of Yuasa-machi (Wakayama
Prefecture) (Fig. 17 ![]() ).
).
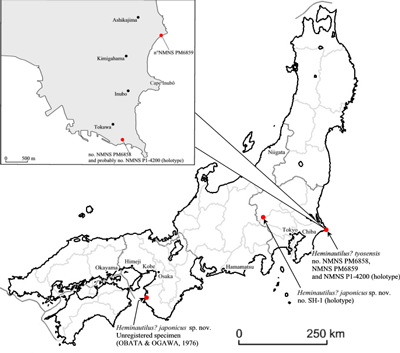
Click on thumbnail to enlarge the image.
Figure 17: Geographical position of specimens of Heminautilus ? japonicus sp. nov. and Heminautilus ? tyosiensis ( & , 1953) referred to in the text.
Stratigraphical distribution: the holotype is dated from the latest Barremian (= Imerites giraudi Zone). The specimen illustrated by & (1976) as Heminautilus lallierianus (, 1841) comes from the Arida Formation, whose uppermost part delivered Heteroceras aff. astieri , 1851. It is contemporary with specimens attributed to Anahamulina cf. subcylindrica (, 1850) ( & , 1976, Pl. 4, figs. 9a-b, 10) but which are probably fragmentary representatives of Heteroceras , 1849. Consequently this specimen comes certainly from the uppermost part of the Late Barremian (= Imerites giraudi Zone?). The species is only known with certainty in the latest Barremian (= Imerites giraudi Zone).
Dimensions (in mm): see Table 11.
Table 11: Measurements of Heminautilus ? japonicus sp. nov.
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. SH-1 | 74.4 | 42.3 | 32 | 0.57 | 0.43 | 0.76 | 7 | 0.09 | 0.17 | - | - | - | - |
| - | 46 | - | - | - | - | - | - | - | 14 | 39 | 0.36 | 0.85 | |
| Unregistered specimen ( & , 1976) | 106 | 69 | 45 | 0.65 | 0.42 | 0.65 | 17 | 0.16 | 0.25 | - | - | - | - |
| 77 | 55 | - | 0.71 | - | - | 14 | 0.18 | 0.25 | 20 | 45 | 0.44 | 0.82 |
Diagnosis: species of small size, relatively compressed, with rounded flanks, concave ventral region, ornamented with fine growth lines in chevron. Ornamentation absent or formed by fine rursiradiate ribs, only present on the top of the flanks of the body chamber. Suture line very sinuous, with a wide, deep and asymmetrical lateral lobe and a clear ventral lobe.
Description: Heminautilus ?
japonicus sp. nov.
is small-sized, with a fairly wide section and rounded flanks, of maximum
thickness near the umbilical edge. The ventral region is narrow to fairly wide,
clearly concave, with an angular ventro-lateral edge. It is ornamented with fine
growth lines, which form an angular sinus. On the phragmocone, no ornamentation
is visible on the flanks. On the body chamber, the holotype no. SH-1 shows an
ornamentation formed by fine rursiradiate ribs, tight and only visible on the
top of the flanks (Pl. L ![]() , fig. 3). No ornamentation seems to be present on the
flanks of the specimen shown by & (1976).
, fig. 3). No ornamentation seems to be present on the
flanks of the specimen shown by & (1976).
The
suture line is strongly sinuous (Fig. 18 ![]() ), with a very wide, deep and
asymmetrical lateral lobe (P/L close
to 0.40), and a very narrow and rounded lateral saddle. The ventral lobe is well
defined and angular.
), with a very wide, deep and
asymmetrical lateral lobe (P/L close
to 0.40), and a very narrow and rounded lateral saddle. The ventral lobe is well
defined and angular.
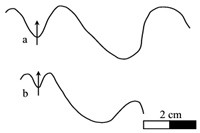
Click on thumbnail to enlarge the image.
Figure 18: Heminautilus ? japonicus sp. nov. Suture line of the unregistered specimen, right side (a) illustrated by & (1976) and of specimen no. SH-1, left side (b).
The position of the siphuncle is not known.
Discussion and comparisons: Heminautilus ? japonicus sp. nov. is close to H. ? tyosiensis ( & , 1953), possessing a similar suture line and a ventral region ornamented with fine growth lines. H. ? tyosiensis ( & ) can however be distinguished by its much wider ventral region, as well as the presence of a well defined ribbing on the flanks of the phragmocone. The stratigraphical position of H. ? tyosiensis ( & ) is also different, at the top of the Early Aptian (= Dufrenoyia furcata Zone).
Heminautilus ? japonicus sp. nov. is also relatively close to H. rangei (, 1922), having a similar shape and ornamentation of its ventral region. Relationships between both species were analyzed in the section concerning H. rangei (); the similarities observed could indicate a phyletic link between these taxa.
Heminautilus ? japonicus sp. nov. also presents some affinities with H. lallierianus (, 1841), with in particular its concave ventral area, but can be distinguished from it easily by its small size, the ornamentation of its ventral area, its rounded flanks, its distinct suture line and its clearly older stratigraphical position.
As in H. ? tyosiensis ( & ), the plan of the suture line of H. ? japonicus sp. nov., relatively different from that of the other representatives of the genus Heminautilus , 1927, could be compared to the one of the genus Xenocheilus & , 1955. The important stratigraphical gap as well as the misunderstanding of the position of the siphuncle in H. ? japonicus sp. nov. do not allow us to retain this hypothesis. For these reasons the attribution to the genus Heminautilus , 1927, remains conditional.
Heminautilus ? tyosiensis ( & , 1953)
Pl. XXXIII ![]() , fig. 2;
Pl. XXXIV
, fig. 2;
Pl. XXXIV ![]() , figs. 2-3;
Pl. XXXV
, figs. 2-3;
Pl. XXXV ![]() , fig. 3
, fig. 3
|
Platynautilus tyôsiensis nov. sp. - & , p. 57, Pl. 1, figs. 1-4. |
||
|
Heminautilus tyosiensis ( & , 1953) - et al., p. 28, Pl. 2, fig. 2; Pl. 4, fig. 4. |
||
|
Heminautilus aff. tyosiensis ( & , 1953) - et al., p. 28, Pl. 4, fig. 2. |
Type: the holotype
(by original designation) is the specimen no. NMNS P1-4200 illustrated by
& (1953), Pl. 1, figs. 1-4. It comes from the Tokawa quarry (Tyôsi
Peninsula, Tiba Prefecture) and is stored in the National Science Museum in
Tokyo, Japan. It is depicted anew in this work in Pl. XXXIII ![]() , fig. 2.
, fig. 2.
Geographical distribution: the species is known only in Japan, in the Choshi Peninsula (Chiba
Prefecture) (Fig. 17 ![]() ).
).
Stratigraphical distribution: the holotype comes from the Inubouzaki Formation, dated doubtfully by & (1953) as Aptian. However, & (2009) indicate for this formation the presence of the genera Australiceras , 1926, Tropaeum J. de C. , 1837, Cheloniceras , 1903, Dufrenoyia & , 1915, and Colombiceras , 1923, which allows us to attribute it without ambiguity to the top of the Early Aptian (= Dufrenoyia furcata Zone). According to these authors, specimens illustrated by et al. (1975) come from the Kimihagama Formation [likely equivalent to the major part of the Kimihagama, Inubouzaki and Toriakeura formations in the sense of & (2009)], above the Ashikajima Formation [with Dufrenoyia aff. dufrenoyi (, 1841)] and below the Nagasakihana Formation [with Cheloniceras meyendorffi (, 1845)]; these specimens can thus be attributed also to the top of the Early Aptian (= Dufrenoyia furcata Zone).
In the absence of figures and of bibliographical reference, the presence of Heminautilus ? tyosiensis ( & , 1953) indicated by et al. (1982, p. 148) and by & (2009, p. 254) in the Ashikajima Formation (in the sense of & (2009), not et al. (1975); Early Barremian) cannot be confirmed.
Heminautilus ? tyosiensis ( & ) is thus known with certainty only from the top of the Early Aptian.
Dimensions (in mm): see Table 12.
Table 12: Measurements of Heminautilus ? tyosiensis ( & , 1953).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. NMNS P1-4200 | 70 | 43 | 20 | 0.61 | 0.29 | 0.47 | 18 | 0.26 | 0.42 | - | - | - | - |
| - | 36 | - | - | - | - | - | - | - | 11 | 28 | 0.39 | 0.78 | |
| - | 26 | - | - | - | - | - | - | - | 11 | 22 | 0.5 | 0.85 |
Description:
the holotype (Pl. XXXIII ![]() , fig. 2) is a specimen of rather small size, preserved
as a partially complete phragmocone with the major part of the body chamber. It
shows a rather wide section, of maximum thickness near the umbilical edge, with
a very involute coiling. Flanks are almost flat and converge on the ventral
region. The ventral region is wide, slightly concave, surrounded by two angular
edges; it shows in its median part a clear furrow.
, fig. 2) is a specimen of rather small size, preserved
as a partially complete phragmocone with the major part of the body chamber. It
shows a rather wide section, of maximum thickness near the umbilical edge, with
a very involute coiling. Flanks are almost flat and converge on the ventral
region. The ventral region is wide, slightly concave, surrounded by two angular
edges; it shows in its median part a clear furrow.
On the adapical part of the phragmocone, four wide, weak and flexuous ribs are visible only on the outside part of the flanks. This ribbing quickly fades and then no ornamentation is visible on the flanks of the adoral part of the phragmocone and the body chamber. On the ventral region, the only discernible ornamentation consists of weak growth lines strongly retroverse and forming an angular chevron at the level of the median furrow.
The
suture line, very sinuous (Fig. 19 ![]() ), shows a well developed lateral saddle and
an asymmetrical, wide and deep lateral lobe, with a ratio P/L between 0.40 and 0.50. The depth of the lateral lobe decreases
through ontogeny. The delineation of the suture line on the ventral region is
not visible. The position of the siphuncle is unknown.
), shows a well developed lateral saddle and
an asymmetrical, wide and deep lateral lobe, with a ratio P/L between 0.40 and 0.50. The depth of the lateral lobe decreases
through ontogeny. The delineation of the suture line on the ventral region is
not visible. The position of the siphuncle is unknown.
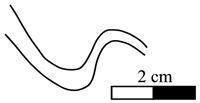
Click on thumbnail to enlarge the image.
Figure 19: Heminautilus ? tyosiensis ( & , 1953). Suture line of specimen no. NMNS P1-4200, right side.
The
specimens no. NMNS PM6858 and no. NMNS PM6859 are very fragmentary, and show
characteristics identical to those observable in the holotype. The specimen no.
NMNS PM6858 (Pl. XXXIV ![]() , fig. 2;
Pl. XXXV
, fig. 2;
Pl. XXXV ![]() , fig. 3) corresponds to a body chamber; we
observe in particular, on the ventral region, the characteristic ornamentation
visible in the holotype, constituted by fine growth lines forming an angular
chevron. The specimen no. NMNS PM6859 (Pl. XXXV
, fig. 3) corresponds to a body chamber; we
observe in particular, on the ventral region, the characteristic ornamentation
visible in the holotype, constituted by fine growth lines forming an angular
chevron. The specimen no. NMNS PM6859 (Pl. XXXV ![]() , fig. 3) is constituted by a
fragment of a small-sized whorl (young specimen or fragment of inner whorls of
an adult specimen). Flanks show an ornamentation formed by wide and radial ribs,
flexuous, similar to the ones visible on the phragmocone of the holotype.
, fig. 3) is constituted by a
fragment of a small-sized whorl (young specimen or fragment of inner whorls of
an adult specimen). Flanks show an ornamentation formed by wide and radial ribs,
flexuous, similar to the ones visible on the phragmocone of the holotype.
Discussion and comparisons: Heminautilus ? tyosiensis ( & , 1953) is close to Heminautilus ? japonicus sp. nov. The differences between both taxa were previously commented in the section of discussion and comparisons of H. ? japonicus sp. nov.
The very particular characteristics of Heminautilus ? tyosiensis ( & ) (the width of the ventral region and the presence of growth lines in the same part of the nautiloid) distinguish it from most of the other taxa of the genus Heminautilus , 1927. Furthermore, the general shape of the shell and the plan of its suture line, with an asymmetrical lateral lobe and a wide and rounded lateral saddle, could also move this taxon closer to the genus Xenocheilus & , 1955, in particular to the species Xenocheilus ulixis & , 1955, from the Hauterivian of Crimea. The stratigraphical gap as well as the unknown position of the siphuncle in Heminautilus ? tyosiensis ( & ) make it difficult to attribute this species with certainty to the genus Heminautilus .
Heminautilus etheringtoni , 1946, shows, however, a comparable ventral region, but can be distinguished by a clearly more rounded ventro-lateral edge, as well as the absence of a median furrow and lateral ornamentation on the phragmocone. The plan of the suture line is also different, with a more reduced lateral saddle and a clearly shallower lateral lobe in H. etheringtoni .
Heminautilus rangei (, 1922) also presents some affinities with Heminautilus ? tyosiensis ( & ), in particular the presence of a rather similar ribbing on the phragmocone and of the ventro-lateral edges well defined on the body chamber. H. ? tyosiensis ( & ) can be distinguished from it easily by a greater thickness and by the absence of ribbing on the body chamber.
Genus Josanautilus & , 2006
Type species: Nautilus lacerdae , 1870 (by original designation of & , 2006, p. 17).
Discussion: the genus is monospecific. Its stratigraphic distribution is very restricted, limited to the Early Aptian, upper part of the Deshayesites forbesi Zone (Roloboceras hambrovi Subzone). It is known only from France and Spain.
Josanautilus lacerdae (, 1870)
Pl. XXXIX ![]() , figs. 2-3;
Pl. XLI
, figs. 2-3;
Pl. XLI ![]() , figs. 2-3;
Pl. XLII
, figs. 2-3;
Pl. XLII ![]() , fig. 2;
, fig. 2;
Pl. XLIII ![]() ,
fig. 2;
Pl. XLVI
,
fig. 2;
Pl. XLVI ![]() , figs. 1-3;
Pl. XLVII
, figs. 1-3;
Pl. XLVII ![]() , figs. 1-2;
, figs. 1-2;
Pl. XLVIII ![]() , figs. 1-3;
Pl. XLIX
, figs. 1-3;
Pl. XLIX ![]() , fig. 1;
, fig. 1;
Pl. L ![]() , fig. 1;
Pl. LI
, fig. 1;
Pl. LI ![]() , fig.
1
, fig.
1
|
Nautilus lacerdae sp. nov. - , Pl. 3, fig. 3. |
||
|
Nautilus lacerdae , 1870 - , p. 64, Fig. 653. |
||
|
Carinonautilus ? lacerdae (, 1870) - , p. 194, Fig. 24. |
||
|
Josanautilus lacerdae (, 1870) - & , p. 18, Figs. 3a, 4b-e, 6b-c, 7c. |
||
|
? |
Nautilus cf. lacerdae , 1870 - , p. 10, photos 8- 9. |
Material studied (N = 15): specimens no. tl074, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France); no. VI011, coll. , no. CH01, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France); no. MHNG GEPI 83570, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Groumaud section, Bourg-Saint-Andéol (Ardčche, France); no. A-148, JM-35, PUAB 48003, PUAB 48057, PUAB 48082, PUAB 48086, PUAB 48178, PUAB 48222, PUAB 48276, PUAB 48302, coll. PUAB, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain); no. Se-11.1, coll. MV, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mas Segura, Province of Castellon (Spain).
Type: the holotype
(by monotypy) is the specimen no. MNCN I-14855, coll. of the Museo Nacional de
Ciencias Naturales (Madrid), drawn by (1870) and photographed by
&
(2006). It comes from the Early Aptian, Deshayesites
forbesi Zone, Roloboceras hambrovi
Subzone of Josa (Province of Teruel, Spain). The figure of
& (2006) is reproduced here Pl. XLII ![]() , fig. 2.
, fig. 2.
Geographical distribution: Josanautilus lacerdae (, 1870) is known only from France and Spain.
Stratigraphical distribution: according to et al. (2007), the species Josanautilus lacerdae (, 1870) is present in the sector of Oliete and Josa, in the Margas del Forcall Formation, dated from the Roloboceras hambrovi Subzone by its plentiful fauna. It is likely that the holotype as well as paratypes studied by & (2006) come from this formation. Specimens no. tl074, VI011, CH01 and MHNG GEPI 83570 also come from the Roloboceras hambrovi Subzone. Thus, Josanautilus lacerdae () seems restricted to the upper part of the Deshayesites forbesi Zone (Roloboceras hambrovi Subzone).
Dimensions (in mm): see Table 13.
Table 13: Measurements of Josanautilus lacerdae (, 1870).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| Holotype no. MNCN I-14855 | 126 | 75 | 56 | 0.6 | 0.44 | 0.75 | - | - | - | - | - | - | - |
| 109 | 71 | - | 0.65 | - | - | - | - | - | 22 | 49 | 0.45 | 0.69 | |
| 93 | 57 | - | 0.61 | - | - | - | - | - | 20 | 42 | 0.48 | 0.74 | |
| no. tl074 | c167 | 106 | 72 | 0.63 | 0.43 | 0.68 | c28 | 0.17 | 0.26 | - | - | - | - |
| - | 80 | 46 | - | - | - | 19 | - | 0.24 | - | - | - | - | |
| no. MHNG GEPI 83570 | c167 | 99 | c65 | 0.59 | 0.39 | 0.66 | - | - | - | - | - | - | - |
| - | 91 | - | - | - | - | 26 | - | 0.29 | - | - | - | - | |
| - | 75 | - | - | - | - | 23 | - | 0.31 | 16 | 61 | 0.26 | 0.81 | |
| no. VI011 | 148 | 95 | 72 | 0.64 | 0.49 | 0.76 | 31 | 0.21 | 0.33 | - | - | - | - |
| 98 | 62 | 43 | 0.63 | 0.44 | 0.69 | c13 | 0.13 | 0.21 | - | - | - | - | |
| no. CH01 | 156 | 110 | 92 | 0.71 | 0.59 | 0.84 | - | - | - | - | - | - | - |
| no. A-148 | 87 | 57 | 41 | 0.66 | 0.47 | 0.72 | |||||||
| 58.8 | 31.5 | 35.6 | 0.53 | 0.61 | 1.13 | 12.6 | 0.21 | 0.40 | 8.6 | 22.5 | 0.38 | 0.72 | |
| 47.4 | 26.5 | 28.3 | 0.56 | 0.6 | 1.07 | 11.1 | 0.23 | 0.42 | 8.1 | 20.5 | 0.4 | 0.77 | |
| no. JM-35 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| no. PUAB 48003 | c155 | c96 | c70 | 0.62 | 0.45 | 0.73 | |||||||
| - | 59.3 | 40.2 | - | - | 0.68 | 22.0 | - | 0.37 | 16.6 | 37.7 | 0.44 | 0.64 | |
| no. PUAB 48057 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| no. PUAB 48082 | c165 | 98 | c84 | 0.59 | 0.51 | 0.86 | - | - | - | - | - | - | - |
| no. PUAB 48086 | - | c57 | 53 | - | - | 0.93 | |||||||
| - | 45.2 | 47.1 | - | - | 1.04 | 18.7 | - | 0.41 | 11.6 | 31.2 | 0.37 | 0.69 | |
| no. PUAB 48178 | c165 | c100 | - | 0.61 | - | - | |||||||
| 135 | 88 | 57.3 | 0.65 | 0.42 | 0.65 | ||||||||
| - | 80.8 | 58.3 | - | - | 0.72 | 27.4 | - | 0.34 | 16.9 | 50.5 | 0.33 | 0.63 | |
| - | 67.9 | 54.2 | - | - | 0.8 | 18.2 | - | 0.27 | 17.6 | 43.9 | 0.4 | 0.65 | |
| no. PUAB 48222 | 103 | 72.0 | c48 | 0.7 | 0.47 | 0.67 | - | - | - | - | - | - | - |
| - | 57.3 | 47.7 | - | - | 0.83 | - | - | - | 14.4 | 36.0 | 0.4 | 0.63 | |
| no. PUAB 48276 | 88 | 56 | c46 | 0.64 | 0.52 | 0.82 | |||||||
| - | 35.3 | 42.3 | - | - | 1.2 | 14.3 | - | 0.41 | 7.4 | 23.9 | 0.31 | 0.68 | |
| no. PUAB 48302 | 78 | 53 | c38 | 0.68 | 0.49 | 0.72 | |||||||
| 74.7 | 42.8 | 40.3 | 0.57 | 0.54 | 0.94 | 15.0 | 0.20 | 0.35 | 11.8 | 30.0 | 0.39 | 0.7 | |
| 52.5 | 30.0 | 27.7 | 0.57 | 0.53 | 0.92 | 10.1 | 0.19 | 0.34 | 8.0 | 22.1 | 0.36 | 0.74 |
Description: the various studied specimens show a very involute coiling, with a wide trapezoidal section, whose maximum thickness is located near the umbilical edge. Flanks are flat, convergent towards a relatively wide ventral region.
In
the internal whorls the ventral region is rounded, with a weak median keel, and
becomes gradually flat through ontogeny, with rounded ventro-lateral edge. On
the adoral part of the phragmocone and on the body chamber, the ventral region
widens and the ventral ornamentation strengthens: the median keel becomes
strong, with round relief, lined with longitudinal ribs which appear gradually,
but always remain weaker than the median keel. One or two longitudinal ribs are
observable on each side of the keel at the beginning of the body chamber, up to
three on each side at the end of the body chamber (specimen no. Se-11.1, Pl. XLVI ![]() ,
fig. 2).
,
fig. 2).
Lateral
ornamentation is almost absent: we observe only, as in some species of the genus
Heminautilus , 1927, the appearance of convex and
rursiradiate ribs at the end of the body chamber, especially visible in the
upper half of the flanks (specimen no. Se-11.1, Pl. XLVIII ![]() fig. 3).
fig. 3).
The
suture line (Fig. 20 ![]() ) shows a well developed, rounded lateral saddle, and a wide
lateral lobe of rather variable depth, with P/L
between 0.26 and 0.48. It crosses the ventral region almost straight, without
forming a ventral lobe. The siphuncle, whose position was unknown until now, is
visible on the specimen no. CH01 (Pl. XLVII
) shows a well developed, rounded lateral saddle, and a wide
lateral lobe of rather variable depth, with P/L
between 0.26 and 0.48. It crosses the ventral region almost straight, without
forming a ventral lobe. The siphuncle, whose position was unknown until now, is
visible on the specimen no. CH01 (Pl. XLVII ![]() fig. 2): it is placed near the
dorsal region, at approximately 1/3 of the height of the section.
fig. 2): it is placed near the
dorsal region, at approximately 1/3 of the height of the section.
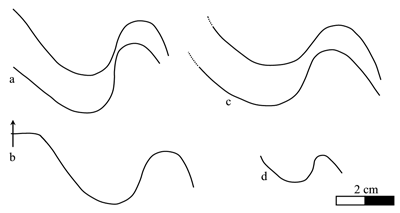
Click on thumbnail to enlarge the image.
Figure 20: Josanautilus lacerdae (, 1870). Suture line of specimen no. MNCN I-14855, left side (a-b), PUAB 48222, right side (c) and A-148, left side (d).
Discussion and comparisons: the particular characteristics of Josanautilus lacerdae (, 1870) distinguish it immediately from most of the representatives of the genus Heminautilus , 1927.
J. lacerdae () can, however, be compared to the slightly older (Late Barremian, Imerites giraudi Zone, to Early Aptian, Deshayesites forbesi Zone) species Heminautilus sanctaecrucis , 1980, which also shows a ventral keel, the same type of ribbing only present at the end of the body chamber, and a suture line which shows a comparable delineation. J. lacerdae () can be distinguished from H. sanctaecrucis by its section and its much wider ventral region, its stronger and rounded ventral keel, and by the presence of longitudinal ribs on the ventral region of the body chamber.
The wide section of J. lacerdae () is also comparable to that observed in the species H. ? tyosiensis ( & , 1953) and H. etheringtoni , 1946, but can be distinguished from them easily by the absence of longitudinal ornamentation in the latter two.
Josanautilus aff. lacerdae (, 1870)
Pl. L ![]() , fig. 2;
Pl. LI
, fig. 2;
Pl. LI ![]() , fig. 2
, fig. 2
Material studied (N = 1): specimen no. VI010, coll. , Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France).
Dimensions (in mm): see Table 14.
Table 14: Measurements of Josanautilus aff. lacerdae (, 1870).
| D | H | E | H/D | E/D | E/H | M | M/D | M/H | P | L | P/L | L/H | |
| no. VI010 | 146 | 95 | 83 | 0.65 | 0.57 | 0.87 | 27 | 0.18 | 0.28 | - | - | - | - |
| 90 | 62 | 50 | 0.69 | 0.56 | 0.81 | 23 | 0.26 | 0.37 | 17 | 46 | 0.37 | 0.74 | |
| 81 | 54 | 43 | 0.67 | 0.53 | 0.8 | 15 | 0.19 | 0.28 | 13 | 39 | 0.33 | 0.72 |
Description: adult specimen, badly preserved. The section is trapezoidal, relatively wide in the phragmocone, then widens very strongly in the body chamber. Flanks are flat or slightly convex, convergent towards the ventral region.
On the visible part of the phragmocone, the ventral region is fairly wide, flat, with a poorly defined median keel. On the body chamber, it strongly widens, where two longitudinal ribs gradually appear on each side of the median keel. On the last part of the body chamber, the longitudinal ribs and the median keel have the same strength.
No ornamentation can be seen on the major part of the flanks. On the second half of the body chamber, rursiradiate ribs appear, only visible near the ventral region.
The
suture line (Fig. 21 ![]() ) shows a wide and rounded lateral saddle and a well
developed and relatively deep lateral lobe (P/L
between 0.33 and 0.37). There is no ventral lobe.
) shows a wide and rounded lateral saddle and a well
developed and relatively deep lateral lobe (P/L
between 0.33 and 0.37). There is no ventral lobe.
Click on thumbnail to enlarge the image.
Figure 21: Josanautilus aff. lacerdae (, 1870). Suture line of specimen no. VI010, left side.
Discussion and comparisons: Josanautilus aff. lacerdae (, 1870) is very close to J. lacerdae (), but differs from it by its weaker median keel, of the same strength as the longitudinal ribs which frame it, as well as by its particularly wide section. It is not however excluded that these characteristics could represent only one slightly atypical morph of J. lacerdae ().
J. aff. lacerdae () also shows some superficial resemblance with the Berriasian species Aulaconautilus sexcarinatus (F.-J. , 1867), but can be distinguished from it by the characteristics explained in the section II. 2.: a very different section, a higher expansion rate of whorls, the presence of a median keel on the ventral region, the presence of ribbing at the end of the body chamber and a suture line with a different plan.
At present, H. boselliorum et al. is the oldest known species of the genus Heminautilus , 1927, and is present in the Early Barremian, upper part of Kotetishvilia nicklesi Zone to lower part of Kotetishvilia compressissima Zone. As explained in the section II. 1., its origin must probably be related to the genus Pseudocenoceras , 1927, and in particular to the taxon Pseudocenoceras campichei (, 1907), from the Hauterivian? However, the evolutionary relationship between these genera requires more data and for this reason remains to be developed.
The characteristics observed in H. rangei () allow us
to envisage an origin from the taxon H. boselliorum
et al., 2012 (Fig. 22 ![]() ):
a comparable section; a similar ventral region, flat or slightly concave with
the possible presence of a weak median keel; an identical suture line, with a
lateral lobe of comparable depth (P/L most of the time between 0.25 and
0.30). Only a slightly wider section and especially the early appearance of
characteristic ribbing clearly differentiate the species H. rangei ()
from H. boselliorum et
al. The very early appearance of the ribbing could be interpreted in this
case as the result of a peramorphic process of acceleration. However, the
stratigraphical gap between both taxa is large [Early Barremian, Kotetishvilia
nicklesi, Nicklesia pulchella and Kotetishvilia compressissima
zones for H. boselliorum et al., Late Barremian, Gerhardtia
sartousiana or Imerites giraudi zones for H. rangei ()];
with the absence of stratigraphically intermediate specimens, this hypothesis
needs to be confirmed.
):
a comparable section; a similar ventral region, flat or slightly concave with
the possible presence of a weak median keel; an identical suture line, with a
lateral lobe of comparable depth (P/L most of the time between 0.25 and
0.30). Only a slightly wider section and especially the early appearance of
characteristic ribbing clearly differentiate the species H. rangei ()
from H. boselliorum et
al. The very early appearance of the ribbing could be interpreted in this
case as the result of a peramorphic process of acceleration. However, the
stratigraphical gap between both taxa is large [Early Barremian, Kotetishvilia
nicklesi, Nicklesia pulchella and Kotetishvilia compressissima
zones for H. boselliorum et al., Late Barremian, Gerhardtia
sartousiana or Imerites giraudi zones for H. rangei ()];
with the absence of stratigraphically intermediate specimens, this hypothesis
needs to be confirmed.
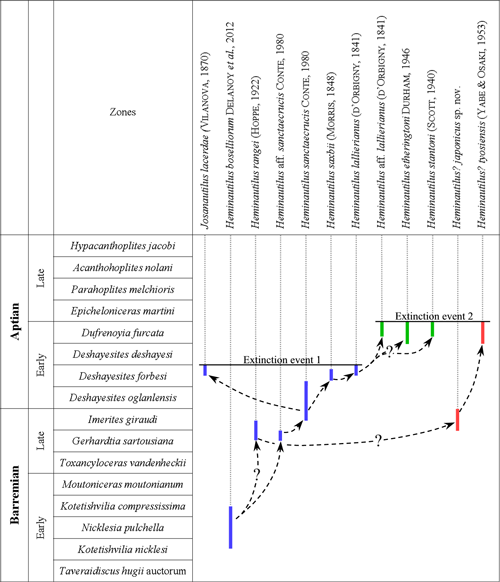
Click on thumbnail to enlarge the image.
Figure 22: Phylogenetic relationships between the various species of the genera Heminautilus , 1927, and Josanautilus & , 2006. In blue, species present only in the Tethyan Domain; in green, species present only in the North-Atlantic Domain; in red, species present only in Japan. Extinction event 1 represents the extinction of the genera Heminautilus and Josanautilus & in the Tethyan Domain whereas extinction event 2 represents the global extinction of the genus Heminautilus . Explanations in text.
As H. rangei (, 1922), H. sanctaecrucis
presents very numerous similarities with H. boselliorum
et al., 2012, in particular a similar suture line, an identical section with a
ventral region of the same profile and the same type of ribbing present only at
the adoral end of the body chamber of the adult specimens. All these
characteristics lead us to envisage a direct filiation (Fig. 22 ![]() ) between H.
boselliorum et
al. and H. sanctaecrucis ,
the major evolutionary trend being the development of a well defined ventral
keel. Here too, the stratigraphical gap between both species is important [Early
Barremian, Kotetishvilia nicklesi Zone to Kotetishvilia compressissima
Zone for H. boselliorum et
al.; Late Barremian, Imerites giraudi Zone to Early Aptian, Deshayesites
forbesi Zone for H. sanctaecrucis ]. The presence of H.
aff. sanctaecrucis in the Gerhardtia sartousiana Zone
(specimen no. 415951, Pl. XII
) between H.
boselliorum et
al. and H. sanctaecrucis ,
the major evolutionary trend being the development of a well defined ventral
keel. Here too, the stratigraphical gap between both species is important [Early
Barremian, Kotetishvilia nicklesi Zone to Kotetishvilia compressissima
Zone for H. boselliorum et
al.; Late Barremian, Imerites giraudi Zone to Early Aptian, Deshayesites
forbesi Zone for H. sanctaecrucis ]. The presence of H.
aff. sanctaecrucis in the Gerhardtia sartousiana Zone
(specimen no. 415951, Pl. XII ![]() , fig. 4), whose characteristics seem intermediate
between H. boselliorum et
al. and H. sanctaecrucis ,
confirms this hypothesis.
, fig. 4), whose characteristics seem intermediate
between H. boselliorum et
al. and H. sanctaecrucis ,
confirms this hypothesis.
H.
saxbii () replaces H. sanctaecrucis , 1980, in
the Early Aptian, Deshayesites forbesi Zone and seems to descend directly
from it (Fig. 22 ![]() ), essentially by disappearance of the typical ventral keel of H.
sanctaecrucis , development of a suture line with a slightly
deeper lateral lobe [P/L generally between 0.20 and 0.35 for H.
sanctaecrucis and between 0.30 and 0.40 for H. saxbii ()]
and a more premature appearance of the ribbing on the adult body chamber.
), essentially by disappearance of the typical ventral keel of H.
sanctaecrucis , development of a suture line with a slightly
deeper lateral lobe [P/L generally between 0.20 and 0.35 for H.
sanctaecrucis and between 0.30 and 0.40 for H. saxbii ()]
and a more premature appearance of the ribbing on the adult body chamber.
H. saxbii () has also affinities with H. boselliorum et al., 2012, but the very large stratigraphical gap between these species does not allow us to retain the hypothesis of a direct phyletic relation.
As explained in part II, H. lallierianus () is a
species very close to H. saxbii (, 1848), and both
taxa were considered as synonyms in numerous works. In reality, it seems that
both species never coexisted; we think that H. lallierianus ()
represents a species different from H. saxbii () that in
Europe, seems to have a slightly upper stratigraphical position in the Deshayesites
forbesi Zone, Roloboceras hambrovi Subzone. These data support that H.
lallierianus () descends directly from H. saxbii ()
(Fig. 22 ![]() ), with the appearance of a more concave ventral region and the
development of a more sinuous suture line (wider and rounded lateral saddle,
clearly deeper lateral and ventral lobes).
), with the appearance of a more concave ventral region and the
development of a more sinuous suture line (wider and rounded lateral saddle,
clearly deeper lateral and ventral lobes).
The presence of H. aff. lallierianus (, 1841)
in Mexico (specimen no. IGM 10050, Pl. XXXIX ![]() , fig. 1 and specimen no. MPL1,
Pl. XL
, fig. 1 and specimen no. MPL1,
Pl. XL ![]() , fig. 1), in the Dufrenoyia justinae Zone (= top of the Dufrenoyia
furcata Zone), shows that the genus Heminautilus
, 1927, was present in
Europe from the Early Barremian to the Early Aptian, Deshayesites forbesi
Zone, Roloboceras hambrovi Subzone, but later during the latest Early
Aptian the species H. lallierianus () probably migrated to
America (cf. section V). H. lallierianus () is thus
probably the ancestor (Fig. 22
, fig. 1), in the Dufrenoyia justinae Zone (= top of the Dufrenoyia
furcata Zone), shows that the genus Heminautilus
, 1927, was present in
Europe from the Early Barremian to the Early Aptian, Deshayesites forbesi
Zone, Roloboceras hambrovi Subzone, but later during the latest Early
Aptian the species H. lallierianus () probably migrated to
America (cf. section V). H. lallierianus () is thus
probably the ancestor (Fig. 22 ![]() ) of the species H. etheringtoni
and H. stantoni (), known in Colombia, Venezuela and Arkansas
(USA) in the uppermost part of the Early Aptian. The morphological differences
with H. lallierianus () are relatively important: the
appearance of a convex ventral region and a wider section in H. etheringtoni
, the development of a strong ribbing on the phragmocone in H.
stantoni (), and less sinuous suture lines in these two species.
Furthermore, H. etheringtoni and H. stantoni ()
also presents numerous similarities with H. saxbii (, 1848)
and an origin from this last species cannot be ruled out. In South America,
Central America and southern North America, the genus Heminautilus
, 1927, is only
represented by some isolated specimens of often poorly known stratigraphical
position; the evolution of the group in this paleogeographic domain remains
uncertain.
) of the species H. etheringtoni
and H. stantoni (), known in Colombia, Venezuela and Arkansas
(USA) in the uppermost part of the Early Aptian. The morphological differences
with H. lallierianus () are relatively important: the
appearance of a convex ventral region and a wider section in H. etheringtoni
, the development of a strong ribbing on the phragmocone in H.
stantoni (), and less sinuous suture lines in these two species.
Furthermore, H. etheringtoni and H. stantoni ()
also presents numerous similarities with H. saxbii (, 1848)
and an origin from this last species cannot be ruled out. In South America,
Central America and southern North America, the genus Heminautilus
, 1927, is only
represented by some isolated specimens of often poorly known stratigraphical
position; the evolution of the group in this paleogeographic domain remains
uncertain.
This new species is known only by two specimens, coming from the Late Barremian of Japan (= Imerites giraudi Zone). Its origin is difficult to specify:
Heminautilus ?
japonicus sp. nov. shows clear morphological affinities
with H. rangei (, 1922), from the Late Barremian of Egypt, explained in the paleontological
study of the Japanese taxon. It could have descended from it (Fig. 22 ![]() ), by the
disappearance of the characteristic ribbing of H.
rangei () and the development of a
clearly wider and deeper lateral lobe. However, the important geographical gap
between these taxa suppose a migration which cannot be confirmed by the current
available data.
), by the
disappearance of the characteristic ribbing of H.
rangei () and the development of a
clearly wider and deeper lateral lobe. However, the important geographical gap
between these taxa suppose a migration which cannot be confirmed by the current
available data.
As explained previously, the characteristics of Heminautilus ? japonicus sp. nov. could indicate a relationship with some representatives of the genus Xenocheilus & , 1955, but the stratigraphical gap and the uncertain position of the siphuncle in Heminautilus ? japonicus sp. nov. prevent confirmation.
H. ?
tyosiensis ( & ) is only represented by the four
specimens illustrated by & , 1953, and
et al., 1975 (three of them shown here: Pl. XXXIII ![]() , fig. 2;
Pl. XXXIV
, fig. 2;
Pl. XXXIV ![]() , figs. 2-3;
Pl. XXXV, fig. 3),
all coming from the uppermost part of the Early Aptian (= Dufrenoyia furcata
Zone) from the Choshi Peninsula (Chiba Prefecture, Japan). This species is very
close to Heminautilus ? japonicus sp. nov. from the Late Barremian of
Japan and shows a similar sutural plan and the same ornamentation on the ventral
region, formed by growth lines arranged in a chevron. H. ? tyosiensis (
& ) probably descended from Heminautilus ? japonicus sp.
nov. (Fig. 22
, figs. 2-3;
Pl. XXXV, fig. 3),
all coming from the uppermost part of the Early Aptian (= Dufrenoyia furcata
Zone) from the Choshi Peninsula (Chiba Prefecture, Japan). This species is very
close to Heminautilus ? japonicus sp. nov. from the Late Barremian of
Japan and shows a similar sutural plan and the same ornamentation on the ventral
region, formed by growth lines arranged in a chevron. H. ? tyosiensis (
& ) probably descended from Heminautilus ? japonicus sp.
nov. (Fig. 22 ![]() ), with the appearance of a much wider ventral region and a ribbing
well defined on the phragmocone.
), with the appearance of a much wider ventral region and a ribbing
well defined on the phragmocone.
The
characteristics of J.
lacerdae (),
from the Deshayesites forbesi Zone, Roloboceras
hambrovi Subzone of Spain and France, are very close to those of the genus Heminautilus
, 1927, and were discussed in the section II. 2. Josanautilus
& ,
2006, probably had its origin from Heminautilus
sanctaecrucis , 1980 (Fig. 22 ![]() ). These species follow one another
in the Deshayesites forbesi Zone and
have numerous common characteristics, in particular the presence of a ventral
median keel; the main morphological change is the appearance in J.
lacerdae () of a
wider section and a clearly different ventral region (wider and on which
longitudinal ribs are added to the median keel on the body chamber).
). These species follow one another
in the Deshayesites forbesi Zone and
have numerous common characteristics, in particular the presence of a ventral
median keel; the main morphological change is the appearance in J.
lacerdae () of a
wider section and a clearly different ventral region (wider and on which
longitudinal ribs are added to the median keel on the body chamber).
As
mentionned above, the oldest representatives of the genus Heminautilus
, 1927, whose stratigraphical positions are exactly known belong to the species H.
boselliorum et
al., 2012, from the
Early Barremian, Kotetishvilia nicklesi,
Nicklesia pulchella and Kotetishvilia
compressissima zones. In Europe, North Africa and Japan, the genus is known
at intervals up to the Early Aptian. In particular, no specimens are known with
certainty from the uppermost part of the Early Barremian or the lowermost part
of the Late Barremian, in the Moutoniceras
moutonianum and Toxancyloceras
vandenheckii zones. The genus presents its acme in the Early Aptian, Deshayesites forbesi Zone, with the species
H.
sanctaecrucis , 1980, H.
saxbii (, 1848) and H. lallierianus (,
1841), particularly frequent
in Spain, France and England (Isle of Wight; , written
communication, 2013). No younger specimens are known with certainty from Europe
and North Africa. In the uppermost part of the Early Aptian, the genus seems to
have migrated outside the Tethyan Domain (Fig. 23 ![]() ). Indeed, the taxa H. aff. lallierianus (, 1841), H. etheringtoni
, 1946, H. stantoni (,
1940) and H. ?
tyosiensis ( & , 1953)
are known only from the uppermost part of the Early Aptian of Mexico, Colombia,
Venezuela, USA (Arkansas) and Japan, respectively.
). Indeed, the taxa H. aff. lallierianus (, 1841), H. etheringtoni
, 1946, H. stantoni (,
1940) and H. ?
tyosiensis ( & , 1953)
are known only from the uppermost part of the Early Aptian of Mexico, Colombia,
Venezuela, USA (Arkansas) and Japan, respectively.
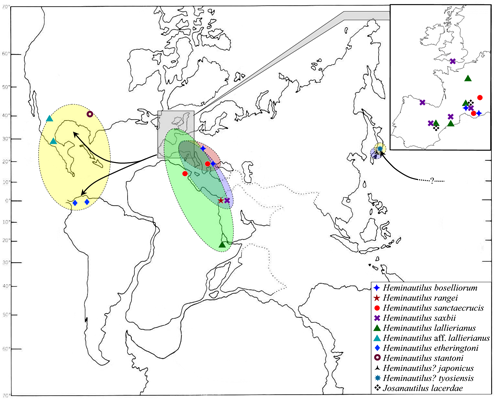
Click on thumbnail to enlarge the image.
Figure 23: Geographical distribution of the various species of the genus Heminautilus , 1927 (paleogeographical map of the Aptian from & , 1977, modified). In red, distribution of the genus Heminautilus during the Early Barremian; in blue, distribution of the genus Heminautilus during the Late Barremian; in green, distribution of the genera Heminautilus and Josanautilus & , 2006, during the early Early Aptian (Deshayesites oglanlensis and Deshayesites forbesi zones); in yellow, distribution of the genus Heminautilus during the late Early Aptian (Dufrenoyia furcata Zone).
With
the exception of the Japanese specimens, the stratigraphical and
paleogeographical distribution of the genus Heminautilus
(Fig. 23 ![]() )
can be split into two clearly different phases:
)
can be split into two clearly different phases:
A first phase, beginning in the Early Barremian, Kotetishvilia
nicklesi Zone and continuing to the Early Aptian, uppermost part of the Deshayesites forbesi Zone, where the genus is only present in the
Tethyan Domain (Europe, North Africa and certainly Ethiopia; et al.,
2012, p.
793). This phase is marked
by the progressive extension of the paleogeographic domain of distribution of
the genus Heminautilus
(Fig. 23 ![]() )
and by the appearance of numerous species, differing in their ornamentation, the
plan of their suture line and the shape of their ventral region, as well as by
the appearance of the genus Josanautilus
& ,
2006. Such a progressive extension of their distribution seems to be linked to
the progressive expansion of the Early Barremian platforms (
et al., 2003;
& , 2011), which were their main habitat. This
phase ends at the top of the Deshayesites
forbesi Zone with the disappearance of the genus Josanautilus
&
and the migration of the genus Heminautilus
outside the
Tethyan Domain (Extinction event 1, Fig. 22
)
and by the appearance of numerous species, differing in their ornamentation, the
plan of their suture line and the shape of their ventral region, as well as by
the appearance of the genus Josanautilus
& ,
2006. Such a progressive extension of their distribution seems to be linked to
the progressive expansion of the Early Barremian platforms (
et al., 2003;
& , 2011), which were their main habitat. This
phase ends at the top of the Deshayesites
forbesi Zone with the disappearance of the genus Josanautilus
&
and the migration of the genus Heminautilus
outside the
Tethyan Domain (Extinction event 1, Fig. 22 ![]() ), which coincides with the end of
the Oceanic Anoxic Event 1a ( et
al., 2009).
), which coincides with the end of
the Oceanic Anoxic Event 1a ( et
al., 2009).
A second phase, situated at the upper part of the Early Aptian, in the
Dufrenoyia furcata Zone, where the genus Heminautilus
is absent in the Tethyan Domain and present in the North-Atlantic Domain
[Mexico, USA (Arkansas), Colombia and Venezuela; et al.,
2012,
p. 793], with forms of clearly less diversified morphology. The genus Heminautilus
is here very frequently
associated with diverse species of the genera Dufrenoyia
& , 1915, and
Pseudosaynella , 1923,
and it is likely that it followed the same migratory routes as these latter (Fig. 23 ![]() ), thanks to the marine transgression recorded in the uppermost
part of the Early Aptian (Ap4 tethyan sequence;
et al., 2012). This phase ends at the top of the Early Aptian by the
disappearance of the genus Heminautilus
(Extinction
event 2, Fig. 22
), thanks to the marine transgression recorded in the uppermost
part of the Early Aptian (Ap4 tethyan sequence;
et al., 2012). This phase ends at the top of the Early Aptian by the
disappearance of the genus Heminautilus
(Extinction
event 2, Fig. 22 ![]() ). It should be noted that with the current knowledge, no specimen is
known with certainty from an interval corresponding in Europe to the Deshayesites
deshayesi Zone and the lower part of the Dufrenoyia furcata Zone,
between the top of the Deshayesites forbesi Zone of Europe and the
uppermost part of the Early Aptian (Dufrenoyia justinae Zone of North
America and Mexico (= top of the Dufrenoyia furcata Zone); Dufrenoyia sanctorum-Stoyanowiceras treffryanus
Assemblage zone
in Colombia).
). It should be noted that with the current knowledge, no specimen is
known with certainty from an interval corresponding in Europe to the Deshayesites
deshayesi Zone and the lower part of the Dufrenoyia furcata Zone,
between the top of the Deshayesites forbesi Zone of Europe and the
uppermost part of the Early Aptian (Dufrenoyia justinae Zone of North
America and Mexico (= top of the Dufrenoyia furcata Zone); Dufrenoyia sanctorum-Stoyanowiceras treffryanus
Assemblage zone
in Colombia).
The Japanese specimens studied here, belonging to the species Heminautilus ? japonicus sp. nov. and H. ? tyosiensis ( & , 1953) are present in the Late Barremian (= Imerites giraudi Zone) and the Early Aptian (= Dufrenoyia furcata Zone), perhaps even in the Early Barremian ( et al., 1982, p. 148; & , 2009, p. 254). The Japanese species seem to correspond to a local radiation of the group in a different domain (Pacific Domain; et al., 2012, p. 793) with a particular migration pattern and an endemic species evolution that remains poorly known. However, the particular morphology of this group does not allow excluding an origin in Xenocheilus & , 1955 (cf. section IV. 7.), which, if it were proved true by new data, could involve the restoration of the genus Platynautilus & , 1953 (type species Platynautilus tyosiensis & , 1953), for these species.
The genera Heminautilus and Josanautilus & seem to have particular ecological requirements and to be generally present only in the outer platform domain. They are nearly absent in deeper marine environments, for example in the Vocontian Basin (southeast France) or the Subbetic Domain (Spain), where other genera like Cymatoceras , 1884, are nevertheless present. Heminautilus and Josanautilus & are also frequently associated with ammonoids that are also characteristic of an outer platform environment:
Heminautilus boselliorum et al., 2012, mainly known in the Lussan area (Gard, France), is associated there with numerous ammonites and nautilids characteristic of an external platform environment, belonging in particular to the genera Davidiceras , 2003, Torcapella , 1970, and Eucymatoceras , 1927. This species is also present at Escragnolles (Alpes-Maritimes) and Comps-sur-Artuby (Var), where the Early Barremian contains similar ammonite fauna and is represented by strongly condensed and glauconitic deposits, characteristic of a drowned platform deposit under a relatively shallow water depth (, 2005).
Apparently, Heminautilus sanctaecrucis , 1980, is also only present in an external platform environment. Indeed the holotype
no. MCGL 20747 comes from the "Marnes
jaunes de la Vraconne"
immediately above the Urgonian Limestone. The specimen no. Bw599 comes from the Pseudocrioceras
waagenoides Subzone of La Bédoule (Bouches du Rhône, France), where it is
associated with representatives of the genus Pseudocrioceras , 1924, typical of such an environment. Both the
specimen of the collection (Pl. XV ![]() , fig.
1a-b) and the specimen
no. 131H (Pl. XI
, fig.
1a-b) and the specimen
no. 131H (Pl. XI ![]() , fig. 5;
Pl. XII
, fig. 5;
Pl. XII ![]() , fig. 1) come from the Urgonian Limestone of the Vaucluse, which
represents deposits of carbonate platform associated with a reef environment.
Finally, the specimens from Bulgaria no. K1 6001, K1 6002, K1 6003, K1 6004, K1
1692 studied by &
(1995) also come from
platform deposits. However, the presence of Heminautilus aff. sanctaecrucis
, 1980 (Pl. XII
, fig. 1) come from the Urgonian Limestone of the Vaucluse, which
represents deposits of carbonate platform associated with a reef environment.
Finally, the specimens from Bulgaria no. K1 6001, K1 6002, K1 6003, K1 6004, K1
1692 studied by &
(1995) also come from
platform deposits. However, the presence of Heminautilus aff. sanctaecrucis
, 1980 (Pl. XII ![]() , fig. 4), in the stratotypic section of Angles (Alpes
de Haute-Provence, Vocontian
Basin, France) shows that an episodic presence in basinal settings is possible.
, fig. 4), in the stratotypic section of Angles (Alpes
de Haute-Provence, Vocontian
Basin, France) shows that an episodic presence in basinal settings is possible.
In Spain, France and England, the species Heminautilus saxbii (, 1848), H. lallierianus (, 1841) and Josanautilus lacerdae (, 1870) are frequently associated with ammonites belonging to the genera Roloboceras , 1954, and Megatyloceras , 1949, also characteristic of an external platform environment ( et al., 2009, p. 70). Furthermore, the specimen of Heminautilus aff. lallierianus () no. IGM 10050, from the Dufrenoyia justinae Zone of Mexico, is associated in the same deposit with the genus Pseudosaynella , 1923, also characteristic of a platform environment. The specimen no. AMNH 25455 (illustrated by , 1943) comes from the Harrar region (Ethiopia) where, during the Aptian, only a very short marine incursion is recorded within sandstone formations of fluvial origin ( et al., 1999), very certainly representing a marine deposit under a very shallow water depth.
As previously explained about the genus Angulithes , 1808 (, 2010), the frequent appearance of ribbing on the body chamber or near the peristome can be interpreted as a strengthening of the shell, representing an adaptation to an environment of relatively high energy.
The study of more than 160 specimens of the genera Heminautilus , 1927, and Josanautilus & , 2006, allows us to identify more exactly the specific and stratigraphical characteristics of the various taxa of these two genera. In particular, this study demonstrates accurately the validity of the species H. saxbii (, 1848) and H. lallierianus (, 1841), and shows that H. tejeriensis & , 2006, is a junior synonym of H. saxbii (), of which it represents a slightly atypical morph. As well, some specimens filled, at least partially, important gaps in the knowledge of the genus Heminautilus , whose stratigraphical distribution is very intermittent between the Early Barremian and the upper part of the Early Aptian, as well as showing that the genus Heminautilus was present from the Early Barremian, Kotetishvilia nicklesi Zone.
This work also highlights the existence of an important
migration of the genus Heminautilus
, 1927, during the Early Aptian. Indeed, with the exception of the
Japanese specimens (belonging to a group with particular morphology and
attributed with doubt to the genus Heminautilus
), the genus seems to be only present in the Tethyan Domain (Europe
and Africa) up to the uppermost part of the Deshayesites
forbesi Zone, and later, it is absent in this domain, but it is present in
the North-Atlantic Domain (Colombia, Venezuela, Mexico and the USA). Due to this
paleogeographic migration and successive disappearances and appearances, we
define two successive extinction events (Fig. 22 ![]() ).
).
It was also possible to formulate hypotheses about the origin and the phylogenetic and paleogeographic evolution of these two genera. In particular, the proposal of ( & , 1983; , 1993), who suggested the origin of Heminautilus from Pseudocenoceras , 1927, seems the most likely with the current data. However, the incomplete knowledge of the genus Heminautilus , in particular during the Barremian, does not allow confirmation of this hypothesis. New specimens will be necessary for confirmation, in particular in the stratigraphical levels where the genus is unknown, in the middle part of the Barremian (Moutoniceras moutonianum and Toxancyloceras vandenheckii zones in particular), but also in the Early Aptian, between the uppermost part of the Deshayesites forbesi Zone and the uppermost part of the Early Aptian (= Dufrenoyia furcata Zone).
For their help and photos of numerous specimens, we warmly thank Masaki (Tokyo Gakugei University, Japan), Yasunari (National Museum of Nature and Science, Japan), Haruyoshi (Kyushu University Museum, Japan), Mark B. (Museum of Paleontology, University of California, Berkeley, USA), Thomas (United States National Museum, Washington, USA), Pedro C.R. (Universidad Nacional de Colombia), Steve (Nationaal Natuurhistorisch Museum, Leiden, Netherlands), Ramadan (King Abdulaziz University, Saudi Arabia), Jens (Faculty of Geosciences, Bremen University, Germany), Jesús (Director of the Museo Paleontológico de la Laguna, Torreón, Coahuila State, Mexico), Gilles (Muséum d'Histoire Naturelle of Auxerre, France), Marin and Vyara (Sofia University, Bulgaria), István (Department of Paleontology and Geology, Hungarian Natural History Museum), Frank (Geologisch-paläontologischen Institut, Leipzig University, Germany), Martin (Isle of Wight, England), Jean-Louis (Lazer, France), Camille (La Ciotat, France), Loic and Axel (Aix-Marseille University, France), Dominique and Daniel . We also thank Didier , Jean-Michel and Sylvain (Muséum National d'Histoire Naturelle of Paris, France) for their welcome and their help during our investigations in the collection, Isabelle (Pierre et Marie University, Paris, France) for examination of the collections of the Sorbonne University, and Emmanuel (co-director of the geological collections, Lyon 1 University, France) for his welcome during our visit to the collection of the École des Mines stored in the Claude University. We warmly thank our reviewers Marcela (Buenos Aires University, Argentina) and Miguel (Granada University, Spain) for their constructive remarks, and Stephen P. (Federation University, University of Ballarat, Australia) for the improvement of the English text. Part of this study was supported by grant PAPIIT IA104715 from DGAPA, Universidad Nacional Autónoma de México.
R.H. (2008).- Lithostratigraphy and biostratigraphy of some Lower Cretaceous outcrops from Northern Sinai, Egypt.- Cretaceous Research, 29, p. 603-624.
M.F. (2006).- Aptian cephalopods from Gabal Abu Ruqum, north Sinai, Egypt.- Egyptian Journal of Paleontology, Cairo, 6, p. 89-123 (7 Pls.).
F. & B. (2004).- Les ammonites aptiennes des Argiles ŕ Plicatules de la bordure orientale du bassin de Paris: un aperçu.- Bulletin de l'Association Géologique Auboise, Sainte-Savine, 24-25, p. 75-80 (1 Pl.).
H. (2005).- The South-East France Basin (SFB) and its Mesozoic evolution. In: T., A., H., M.C., S., E., K.B., A., M. & J. (eds.), The Hauterivian-Lower Aptian sequence stratigraphy from Jura platform to vocontian basin: a multidisciplinary approach. Field-trip of the 7th International symposium on the Cretaceous (September 1-4, 2005).- Géologie alpine, Grenoble, (Série spéciale "Colloques et Excursions"), 7, p. 5-28.
J.R. (1950).- Sinopsis de las especies nuevas del Cretácico de Espańa.- Anales de la Escuela de Peritos Agrícolas y de Especialidades Agropecuarias y de Servicios Técnicos de Agricultura, Barcelona, 9, p. 61-164.
D., G. & S. (2008).- Nouveaux biohorizons et propositions pour un nouveau découpage bio-zonal ammonitique, et nouveaux bio-horizons pour le Barrémien supérieur.- Carnets Geol., Madrid, vol. 8, no. A03 (CG2008_A03), 18 p.
A., A. & R. (1999).- Stratigraphic evidence for an Early Aptian sea-level fluctuation: the Graua Limestone of south-eastern Ethiopia.- Cretaceous Research, 20, p. 783-791.
L.G., J.-L., J. & M. (2011).- Mogharaeceras priscum (, 1916) a peculiar Barremian ammonite (Desmoceratoidea, Barremitinae) from Northern Sinai (Egypt).- Geologica Carpathica, Bratislava, 62, p. 575-578.
A. (1989).- La Peńa Formation (Aptian): a condensed limestone-shale sequence from the subsurface of NE Mexico.- Journal of Petroleum Geology, 12 (1), p. 69-84 (1 Pl.).
R., H.M. & M.I. (1998).- Observations on the lithostratigraphy and ammonite succession of the Aptian (Lower Cretaceous) Lower Greensand of Chale Bay, Isle of Wight, UK.- Cretaceous Research, 19, p. 511-535.
R. & H. (2006).- Shell microstructure and morphogenesis of the ornamentation in Cymatoceras , 1883, Cretaceous Nautilida. Systematic implications.- Lethaia, 39, p. 57-64.
G. (1980).- Heminautilus sanctaecrucis, nouvelle espčce de Nautiloďde crétacé.- Geobios, Villeurbanne, 13 (1), p. 137-141 (1 Pl.).
G. (1985).- Le genre Heminautilus , 1927 dans le Bédoulien (Aptien inférieur) de la région de la Bédoule (SE France).- Géologie Méditerranéenne, Marseille, XII/XIII, p. 29-35 (1 Pl.).
G. (1989).- Fossiles du plateau d'Albion.- Les Alpes de Lumičre, Forcalquier, 99, 75 p.
G. (2007).- Présence du genre Heminautilus , 1927 dans le Gard et l'Ardčche.- Bulletin de la Société d'Étude des Sciences naturelles de Nîmes et du Gard, 66, p. 9-15.
G. (2010).- Fossiles peu connus découverts en Vaucluse.- Courrier scientifique du Parc naturel régional du Luberon, Apt, 9, p. 114-119.
G. (1925).- Le Néocomien de la bordure orientale du Bassin de Paris.- Thčse, Paris, Imprimerie J. et fils, 334 p. (11 Pls.).
G., C., R., G. & C. (2012).- Sur la présence des genres Heminautilus , 1927 et Eucymatoceras , 1927 (Nautilida, Nautilaceae) dans le Barrémien inférieur du Gard (sud-est de la France).- Annales du Muséum d'Histoire naturelle de Nice, XXVII, p. 155-195 (13 Pls.).
N. (1967).- Les fossiles de Bulgarie, IV, Crétacé inférieur, Cephalopoda (Nautiloidea et Ammonoidea).- Académie Bulgare des Sciences, Sofia, 424 p. (93 Pls.).
S.K. & S. (2003).- Fossils explained 44 - Post-Palaeozoic nautiloids.- Geology Today, 19 (5), p. 181-186.
H. (1916).- Les terrains secondaires dans le massif du Moghara ŕ l'est de l'isthme de Suez d'aprčs les explorations de M. . Paléontologie.- Mémoire de l'Académie des Sciences de l'institut de France, 54 (2), 185 p. (21 Pls.).
J.W. (1946).- Upper Aptian nautiloids from Colombia.- Journal of Paleontology, 20 (5), p. 428-434 (3 Pls.).
J.W. (1947).- Nuclear whorls of Heminautilus etheringtoni .- Journal of Paleontology, 21 (6), p. 576-577.
J. (1984).- Phylogeny of the Nautiloidea.- Palaeontologia Polonica, Warszawa, 45, 219 p. (47 Pls.).
F. (1979).- Zonation of the Cretaceous of central Colombia by ammonites.- Publicaciones Geológicas Especiales del Ingeominas, Bogotá, 2, 188 p. (15 Pls.).
I. & N. (2009).- Integrated Lower Cretaceous biostratigraphy of the Bersek Quarry, Gerecse Mountains, Transdanubian Range, Hungary.- Cretaceous Research, 30, p. 78-92.
I. & I. (2013).- Fossils of the Carpathian region.- Life of the Past, Indiana University Press, Bloomington, 508 p.
J. (2010).- Taxonomy and palaeoecology of Cretaceous nautilids Angulithes galea ( in & , 1872) and Angulithes westphalicus (, 1872).- Bulletin of Geosciences, Prague, 85 (3), p. 487-496.
J.A.S. (1988).- Guía de Campo de los fósiles de Espańa y de Europa.- Ediciones Omega, Barcelona, 972 p. (237 Pls.).
W. (1922).- Jura und Kreide der Sinaihalbinsel.- Zeitschrift des Deutschen Palästina-Vereins, Wiesbaden, 45, p. 61-79, 97-219 (4 Pls.).
R. (1945).- Ammonites from the Dierks Limestone of Southern Arkansas.- Journal of Paleontology, 19 (3), p. 277-281 (2 Pls.).
N.I. (1907).- Le Crétacé inférieur de la Crimée et sa faune.- Travaux de la société impériale des naturalistes de St.-Pétersbourg, 32 (5), 482 p. (28 Pls.).
H.C. & W.J. (1977).- Cretaceous faunas from Zululand, South Africa, and southern Mozambique, the Aptian Ancyloceratidae (Ammonoidea).- Annals of the South African Museum, Cape Town, 73 (9), p. 215-359.
B. (1956).- Post-Triassic nautiloid genera.- Bulletin of the Museum of comparative Zoology, Cambridge, Mass., 114 (7), p. 319-494 (28 Pls.).
B. (1964).- Nautiloidea-Nautilida. In: R.C. (ed.), Treatise on Invertebrate Paleontology, part K, Mollusca 3.- Geological Society of America and University of Kansas Press, p. K383-466.
J., M., M. & M.E. (2009).- Aptian (Lower Cretaceous) biostratigraphy and cephalopods from north central Tunisia.- Cretaceous Research, 30, p. 895-910.
I. (1955).- Études paléontologiques sur la faune crétacique du Massif du Moghara (Sinaď - Égypte).- Publications de l'Institut du Désert d'Égypte, Le Caire, 8, 192 p. (19 Pls.).
D. & H. (1971).- Études statistiques sur Pseudaganites aganiticus () et diverses espčces voisines.- Bulletin scientifique de Bourgogne, Dijon, 28, p. 111-170 (4 Pls.).
R. & A. (2006).- Nautílidos del Aptiense Inferior (Cretácico Inferior) de la Subcuenca de Oliete, Cordillera Ibérica oriental (Teruel, Espańa).- Revista Espańola de Paleontología, Madrid, 21 (1), p. 15-27.
J.-P. (1976).- Les calcaires urgoniens de Provence (Valanginien-Aptien inférieur). Stratigraphie, paléontologie, les paléoenvironnements et leur évolution.- Thčse Université Aix-Marseille-II, 445 p. (60 Pls.).
J.-P. & M. (2011).- Drowning discontinuities and stratigraphic correlation in platform carbonates. The late Barremian-early Aptian record of southeast France.- Cretaceous Research, 32, p. 659-684.
, T., K. & Y. (1980).- Cephalopod faunule from the Cretaceous Yatsushiro Formation (Kyushu) and its implications.- Transactions and Proceedings of the Palaeontological Society of Japan, Tokyo, (N.S.), 118, p. 325-338 (1 Pl.).
J.A., M., G., A., G. & R. (2007).- Precisiones sobre la edad, mediante ammonoideos y nautiloideos, de la fm. Margas del Forcall en la subcuenca de Oliete (Cadena Ibérica, Espańa).- Geogaceta, Madrid, 42, p. 75-78 (1 Pl.).
J.A., C. & R. (2008).- El registre fňssil marí (porífers, ammonítids, nautílids i equinoďdeus) i continental (carňfits) del cretaci inferior (barremiŕ superior - aptiŕ) del Garraf.- V Trobada d'Estudiosos del Garraf, Diputació de Barcelona, p. 205-211.
J.A., R., M. & L.G. (2013).- Aptian (Lower Cretaceous) ammonite biostratigraphy of the Francisco Zarco Dam stratigraphic section (Durango State, northeast Mexico).- Journal of South American Earth Sciences, 42, p. 150-158.
J.A., T., R. & R. (2012).- Uppermost Lower Aptian transgressive records in Mexico and Spain: chronostratigraphic implications for the Tethyan sequences.- Terra Nova, 24, p. 333-338.
J.A., M., T., R., G., R., A. (2009).- Biostratigraphic characterization by means of ammonoids of the lower Aptian Oceanic Anoxic Event (OAE 1a) in the eastern Iberian Chain (Maestrat Basin, eastern Spain).- Cretaceous Research, 30, p. 864-872.
J.A., C., K.P., G., R. & O. (2015).- Towards an Aptian (Lower Cretaceous) ammonite biostratigraphy of the Mina Texali section, Central Atlantic province (Puebla State, Central Mexico).- Cretaceous Research, 54, p. 203-211.
J. (1848).- Description of a new species of Nautilus from the Lower Greensand of the Isle of Wight.- Annals and Magazine of Natural History, 1, p. 106-107.
T. & P. (1995).- Some fossil cephalopods from the Loveč Urgonian Group at the village of Puševo, Veliko Târnovo District (Lower Cretaceous, Central Fore-Balkan).- Geologica Balcanica, Sofia, 25 (1), p. 61-70 (10 Pls.).
I., S. & S. (1975).- Geological age of the Cretaceous Choshi Group.- Bulletin of the National Science Museum, Tokyo, (series C), 1 (1), p. 17-36 (4 Pls.).
I. & M. (2009).- Supplementary description of the ammonoids from the Barremian to the Albian of the Choshi Peninsula, Japan.- Cretaceous Research, 30, p. 253-269.
I., S., Y. & M. (1982).- Integrated Mega- and Micro-fossil Biostratigraphy of the Lower Cretaceous Choshi Group, Japan.- Bulletin of the National Science Museum, Tokyo, (series C), 8 (4), p. 145-179 (7 Pls.).
I., M., K., Y. & T. (1984).- Cretaceous Cephalopods from the Sanchu Area, Japan.- Bulletin of the National Science Museum, Tokyo, (series C), 10 (1), p. 9-37 (6 Pls.).
I. & Y. (1976).- Ammonites Biostratigraphy of the Cretaceous Arida Formation, Wakayama Prefecture.- Bulletin of the National Science Museum, Tokyo, (series C), 2 (2), p. 93-110 (4 Pls.).
J.G., L.A. & C. (2012).- Chapter 27: Cretaceous. In: F.M., J.G., M. & G. (eds.), The geologic time scale 2012.- Elsevier B.V., Amsterdam, p. 793-853.
A. d' (1841).- Description de quelques espčces de mollusques fossiles de France.- Revue zoologique par la Société Cuvierienne, Paris, p. 318-319.
A. d' (1850).- Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson éd., Paris, 394 p.
C. (2011).- El Aptiense del margen occidental de la Cuenca del Maestrazgo: controles tectónico, eustático y climático en la sedimentación.- Thčse de doctorat, Departamento de Estratigrafía de la Universidad Complutente de Madrid & Instituto de Geociencias (CSIC-UCM), 649 p.
A., G., C. & P. (2009).- Le genre Lithancylus , 1960 (Ammonoidea, Ancyloceratina) dans l'Aptien inférieur du Couloir rhodanien (Drôme, Sud-Est de la France).- Revue de Paléobiologie, Genčve, 28 (2), p. 491-509 (4 Pls.).
A., G., T., J. E., C., P., M., P. & K.B. (2015).- Three successive phases of platform demise during the early Aptian and their association with the oceanic anoxic Selli episode (Ardčche, France).- Palæogeography, Palæoclimatology, Palæoecology, 418, p. 101-125.
F.-J. & G. (1858-1860).- Description des fossiles du terrain Crétacé des environs de Sainte-Croix, premičre partie.- Matériaux pour la Paléontologie suisse, Genčve, 380 p. (43 Pls.).
P. (1920).- Die Geologie der Isthmuswüste.- Zeitschrift der Deutschen Geologischen Gesellschaft, Stuttgart, 72, p. 233-242 (1 Pl.).
S., P.F., J.A., M.B., R., Y., M., C., V., A., B., V., H., Z., E.J., D., S., T.N., L.G., J.-L., I.A., P. & O. (2011).- Report on the 4th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the " Group" (Dijon, France, 30th August 2010).- Cretaceous Research, 32, p. 786-793.
S., O., B., R., M., V., M., M.V., J.A., J., E.J., M.K., I., C., S., A., S.N., P.F. & J.M. (2014).- Report on the 5th International Meeting of the IUGS Lower Cretaceous Ammonite Working Group, the Group (Ankara, Turkey, 31st August 2013).- Cretaceous Research, 50, p. 126-137.
O. (1982).- The Cretaceous ammonites of Venezuela.- Maraven Editor, Caracas, 132 p. (40 Pls.).
R., F. van, A., D., B., A. & B. (2010).- Revised orbitolinid biostratigraphic zonation for the Barremian - Aptian of the eastern Arabian Plate and implications for regional stratigraphic correlations.- GeoArabia Special Publication, Manama, 4 (1), p. 49-96.
G. (1940).- Cephalopods from the Cretaceous Trinity group of the south-central United States.- The University of Texas Publication, Austin, no. 3945, p. 969-1106 (68 Pls.).
G. (1943).- Palaeontology of Harrar Province, Ethiopia. Part 4. Jurassic Cephalopoda and a Cretaceous Nautilus.- Bulletin of the American Museum of Natural History, New York, 82 (3), p. 61-93 (Pls. 10-25).
V.N. (1975).- Cretaceous Nautiloids.- Transactions of the Palaeontological Institute of the Academy of Sciences of the USSR, Moscow, 150, 208 p. (34 Pls.).
M.I. (1985).- The stratigraphy of the Atherfield Clay Formation (Lower Aptian; Lower Cretaceous) at the type and other localities in southern England.- Proceeding of the Geologists' Association, London, 96 (1), p. 23-45.
P.W., R.A., S.P. & I. (2003).- The Cretaceous World.- Cambridge University Press, 360 p.
L.F. (1927).- Revision of the Jurassic cephalopod fauna of Kachh (Cutch).- Memoirs of the Geological Survey of India, Paleontologia Indica, Lucknow, 9, 71 p. (7 Pls.).
H. (1980).- Un cas de parallélisme évolutif synchrone chez les Nautiles ŕ côtes du Jurassique.- Boletim da Societa Geologica de Portugal, Lisboa, XII, p. 63-69 (2 Pls.).
H. (1989).- Les avatars ontogénétiques et phylogénétiques de l'ornementation chez les Nautiles post-triasiques.- Geobios, Villeurbanne, Mém. spéc. 12, p. 357-367.
H. (1993).- L'évolution itérative des Nautiles post-triasiques.- Geobios, Villeurbanne, Mém. spéc. 15, p. 359-372.
H. & M. (1983).- Le Nautile, fossile vivant ou forme cryptogčne ? Essai sur l'évolution et la classification des Nautilacés.- Bulletin de la Société zoologique de France, Paris, 108, p. 569-579.
H. & M. (1985).- The role of environment in the evolution of Nautilacea. In: U. & A. (eds.), Sedimentary and evolutionary cycles.- Springer Verlag, p. 58-66.
F.-J., R.M., F., H.D. & J. (2008).- Occurrence of Meyeria magna , 1849 in Colombia: a widely distributed species during Aptian times.- Boletín de la Sociedad Geológica Mexicana, Ciudad de México, 60 (1), p. 1-10.
J. (2002).- Étude stratigraphique et paléontologique de la Famille des Pulchellidae (Ammonoidea, Ammonitina, Endemocerataceae).- Géologie Alpine, Grenoble, Mémoire Hors Série, 42, 333 p. (57 Pls.).
J. (1870).- Ensayo de descripción geognóstica de la Provincia de Teruel en sus relaciones con la agricultura de la misma.- Junta General de Estadística, 1863 y 1868, 312 p. (1 Pl.).
J. (1960).- Zur systematik jungmesozoischer nautiliden.- Palaeontographica, Abt. A, Stuttgart, 115, p. 144-206 (Pls. 17-27).
H. & H. (1953).- A new type of Cretaceous nautiloids from Tyosi Peninsula, Kwanto Region.- Bulletin of the National Sciences Museum, Tokyo, 32, p. 55-61 (1 Pl.).
Click on thumbnail to enlarge the image.
Fig. 1a-b: Xenocheilus ulixis & , 1955. Holotype no. 103/101, Early Hauterivian, Biassala (Crimea, Ukraine). Reproduction of the original illustration of (1907; Nautilus malbosi F.-J. , Pl. I, fig. 12a-b).
Fig. 2a-b: Pseudocenoceras campichei (, 1907). Holotype, Hauterivian?, Biassala (Crimea, Ukraine). Reproduction of the original illustration of (1907; Pl. VIII, fig. 13a-b).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Aulaconautilus sexcarinatus (F.-J. , 1867). Cast of the holotype no. MHNG GEPI 15856, Berriasian, Berrias (Ardčche, France). Coll. F.-J. .
Fig. 2a-b: "Heminautilus" akatsui , 1980. Holotype no. GK. H6909, Early Albian, Bishô (Kumamoto Prefecture, Japan).
Fig. 3a-b: Heminautilus boselliorum et al., 2012. Holotype no. Mej01a, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus boselliorum et al., 2012. Holotype no. Mej01a, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 2a-c: Heminautilus boselliorum et al., 2012. Specimen no. MNHN.F.A52065, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 3a-c: Heminautilus boselliorum et al., 2012. Specimen no. Mej11, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus boselliorum et al., 2012. Specimen no. Mej03, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 2: Heminautilus boselliorum et al., 2012. Specimen no. Mej02, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 3a-b: Heminautilus boselliorum et al., 2012. Specimen no. LUS02, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-c: Heminautilus boselliorum et al., 2012. Specimen no. RG2001, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 2: Heminautilus boselliorum et al., 2012. Specimen no. LUS02, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus boselliorum et al., 2012. Specimen no. RG2002b, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 2: Heminautilus boselliorum et al., 2012. Specimen no. 415950, upper part of the Kotetishvilia nicklesi Zone, section no. 833, Comps-sur-Artuby (Var). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus boselliorum et al., 2012. Specimen no. LUS01, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .
Fig. 2: Heminautilus boselliorum et al., 2012. Specimen no. MHNG GEPI 15935, Early Barremian, Nicklesia pulchella Zone?, Escragnolles (Alpes-Maritimes, France). Coll. F.-J. .
Fig. 3a-d: Heminautilus rangei (, 1922). Specimen no. EM697, Late Barremian?, Bir Lagama, Gabal Maghara (north Sinai, Egypt).
Fig. 4a-b: Heminautilus boselliorum et al., 2012. Specimen no. 415950, upper part of the Kotetishvilia nicklesi Zone, section no. 833, Comps-sur-Artuby (Var). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus boselliorum et al., 2012. Specimen no. K1 1693, Early Barremian, Kara Mihal village (Šumen District, Bulgaria). Coll. MPUS.
Fig. 2a-c: Heminautilus boselliorum et al., 2012. Specimen no. K1 1694, Early Barremian, Kriva Reka village (Šumen District, Bulgaria). Coll. MPUS.
Fig. 3a-b: Heminautilus boselliorum et al., 2012. Specimen no. RG2002a, Early Barremian, Nicklesia pulchella Zone, la Lčque, Lussan (Gard, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus boselliorum et al., 2012. Specimen no. K1 1693, Early Barremian, Kara Mihal village (Šumen District, Bulgaria). Coll. MPUS.
Fig. 2a-c: Heminautilus boselliorum et al., 2012. Specimen no. M 2002.554, Early Barremian, Kotetishvilia compressissima Zone, Bersek quarry (Gerecse Mountains, Hungary). Coll. HNHM.
Fig. 3: Heminautilus boselliorum et al., 2012. Specimen no. MHNG GEPI 15935, Early Barremian, Nicklesia pulchella Zone?, Escragnolles (Alpes-Maritimes, France). Coll. F.-J. .
Fig. 4a-c: Heminautilus rangei (, 1922). Specimen no. EM696, Late Barremian?, Bir Lagama, Gabal Maghara (north Sinai, Egypt).
Fig. 5a-b: Heminautilus rangei (, 1922). Specimen no. EM698, Late Barremian?, Bir Lagama, Gabal Maghara (north Sinai, Egypt).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus rangei (, 1922). Neotype no. EM699, Late Barremian?, Bir Lagama, Gabal Maghara (north Sinai, Egypt).
Fig. 2: Heminautilus rangei (, 1922). Specimen no. EM698, Late Barremian?, Bir Lagama (north Sinai, Egypt).
Fig. 3a-c: Heminautilus rangei (, 1922). Specimen no. TK25-15, Late Barremian, El Tourkumanyia (north Sinai, Egypt). Coll. GMMU.

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus rangei (, 1922). Late Barremian?, "Ledschime", (north Sinai, Egypt). Reproduction of the original illustration of (1922; Pl. 4, fig. 2).
Fig. 2: Heminautilus rangei (, 1922). Late Barremian?, "Ledschime", (north Sinai, Egypt). Reproduction of the original illustration of (1922; Pl. 4, fig. 3).
Fig. 3a-c: Heminautilus sanctaecrucis , 1980. Holotype no. MCGL 20747, Early Aptian, Deshayesites forbesi Zone, Sainte-Croix (Switzerland). Coll. F.-J. .
Fig. 4: Heminautilus sanctaecrucis , 1980. Specimen no. MHNG GEPI 15936, Early Aptian, Deshayesites forbesi Zone, La Presta, Val-de-Travers (Switzerland). Coll. F.-J. .
Fig. 5: Heminautilus sanctaecrucis , 1980. Specimen no. 131H, latest Barremian/earliest Aptian, col des Abeilles, Monieux (Vaucluse, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus sanctaecrucis , 1980. Specimen no. 131H, latest Barremian/earliest Aptian, col des Abeilles, Monieux (Vaucluse, France). Coll. .
Fig. 2: Heminautilus sanctaecrucis , 1980. Specimen no. Bw599, Late Barremian, Imerites giraudi Zone, Pseudocrioceras waagenoides Subzone, bed 78, Les Caniers, La Bédoule (Bouches du Rhône, France). Coll. , FSM.
Fig. 3a-b: Heminautilus sanctaecrucis , 1980. Specimen no. K1 6003, Late Barremian, nord of the Puševo village (Veliko Târnovo District, Bulgaria). Coll. MPUS.
Fig. 4a-b: Heminautilus aff. sanctaecrucis , 1980. Specimen no. 415951, Late Barremian, Gerhardtia sartousiana Zone, base of the Hemihoplites feraudianus Subzone, bed 165/041, Angles (Alpes-de-Haute-Provence, France). Coll. (donated by ).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus sanctaecrucis , 1980. Specimen no. Bw599, Late Barremian, Imerites giraudi Zone, Pseudocrioceras waagenoides Subzone, bed 78, Les Caniers, La Bédoule (Bouches du Rhône, France). Coll. , FSM.
Fig. 2a-b: Heminautilus sanctaecrucis , 1980. Specimen no. K1 1692, Late Barremian, Lipnica village (Veliko Târnovo District, Bulgaria). Coll. MPUS.
Fig. 3a-c: Heminautilus sanctaecrucis , 1980. Specimen no. MHNN 26904, Early Aptian, Deshayesites forbesi Zone, La Presta, Val-de-Travers (Switzerland). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus sanctaecrucis , 1980. Specimen no. K1 6001, Late Barremian, north of the Puševo village (Veliko Târnovo District, Bulgaria). Coll. MPUS.
Fig. 2a-b: Heminautilus sanctaecrucis , 1980. Specimen no. K1 6002, Late Barremian, north of the Puševo village (Veliko Târnovo District, Bulgaria). Coll. MPUS.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus sanctaecrucis , 1980. Unregistered specimen, Early Aptian, Coustellet quarry (Vaucluse, France). Coll. .
Fig. 2a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 88770, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Tolodella, Maestrat Basin (Spain).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus sanctaecrucis , 1980. Specimen no. K1 6001, Late Barremian, north of the Puševo village (Veliko Târnovo District, Bulgaria). Coll. MPUS.
Fig. 2a-b: Heminautilus saxbii (, 1848). Holotype no. BMNH 47019, Early Aptian, Deshayesites forbesi Zone, Isle of Wight (England). Coll. Saxby.
Fig. 3a-b: Heminautilus saxbii (, 1848). Specimen no. EM700, Early Aptian?, Bir Lagama, Gabal Maghara (Egypt).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus saxbii (, 1848). Specimen no. EM700, Early Aptian?, Bir Lagama, Gabal Maghara (Egypt).
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. A-127, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.
Fig. 3a-b: Heminautilus saxbii (, 1848). Specimen no. PMBb2, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus saxbii (, 1848). Specimen no. A-144, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.
Fig. 2a-b: Heminautilus saxbii (, 1848). Specimen no. A-145, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.
Fig. 3a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 88767, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Tolodella, Maestrat Basin (Spain).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus saxbii (, 1848). Specimen no. MV 428.3, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mola Murada, Maestrat Basin (Spain).
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. PUAB 88762, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Lo Pui, Organyŕ Basin (Spain).
Fig. 3a-b: Heminautilus saxbii (, 1848). Specimen no. JM-34, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus saxbii (, 1848). Specimen no. PNRL 1189, Early Aptian, Deshayesites forbesi Zone, Murs (Vaucluse, France). Coll. A. .
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. PUAB 89788, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Galve, Province of Teruel (Spain).
Fig. 3: Heminautilus saxbii (, 1848). Specimen no. AG-3, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.
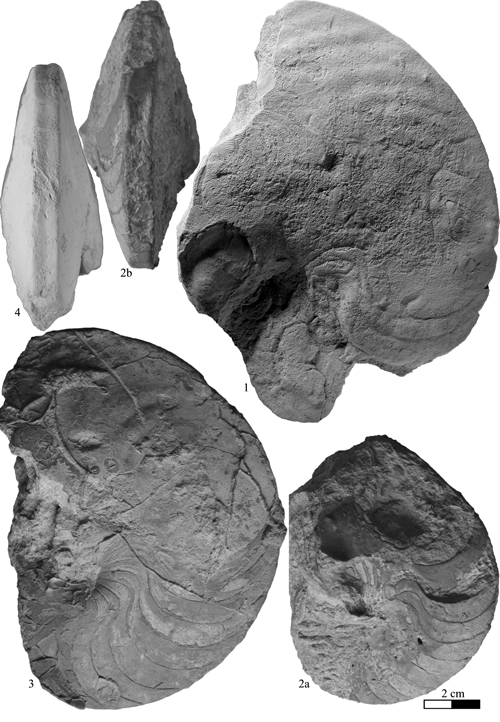
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus saxbii (, 1848). Specimen no. MV 428.2, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mola Murada, Maestrat Basin (Spain).
Fig. 2a-b: Heminautilus saxbii (, 1848). Specimen no. PM3N11, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .
Fig. 3: Heminautilus saxbii (, 1848). Specimen no. PM1, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .
Fig. 4: Heminautilus saxbii (, 1848). Specimen no. PUAB 89788, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Galve, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus saxbii (, 1848). Specimen no. PM13, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. PUAB 48013, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 3: Heminautilus saxbii (, 1848). Specimen no. PM1, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 48042, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. PUAB 48013, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 3: Heminautilus saxbii (, 1848). Specimen no. PUAB 48158, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 4a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 48255, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus saxbii (, 1848). Specimen no. 12576, Early Aptian, "Provence" (France). Coll. FSM.
Fig. 2a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 88373, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Cuchía, Basque Cantabrian Basin (Spain).
Fig. 3a-b: ? Heminautilus saxbii (, 1848). Specimen no. MB RE1962A, Early Aptian, Tibú Formation, Seboruco section, east of Mérida (Venezuela).
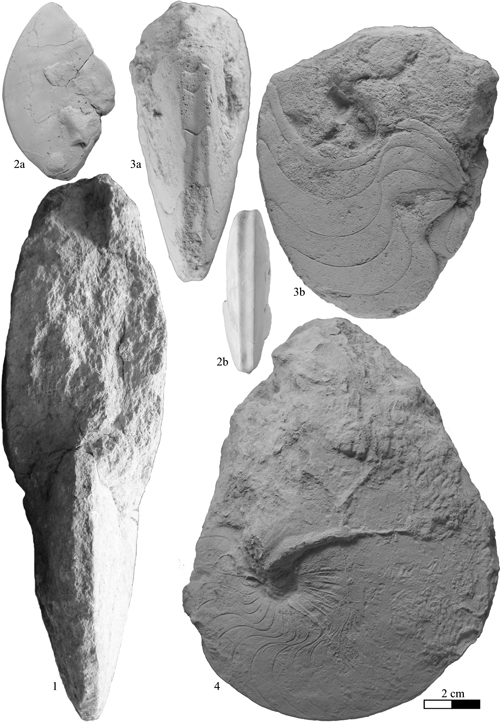
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus saxbii (, 1848). Specimen no. 12576, Early Aptian, "Provence" (France). Coll. FSM.
Fig. 2a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 88372, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Cuchía, Basque Cantabrian Basin (Spain).
Fig. 3a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 48261, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 4: Heminautilus saxbii (, 1848). Specimen no. PUAB 48044 (paratype of Heminautilus tejeriensis & , 2006), Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus saxbii (, 1848). Specimen no. PUAB 48292 (holotype of Heminautilus tejeriensis & , 2006), Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 2: Heminautilus saxbii (, 1848). Specimen no. PUAB 48044 (paratype of Heminautilus tejeriensis & , 2006), Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 3a-b: Heminautilus saxbii (, 1848). Early Aptian, Josa, Province of Teruel (Spain). Holotype of Heminautilus verneuilli , 1870, reproduction of the original illustration.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus lallierianus (, 1841). Specimen no. PUAB 89780, Early Aptian, probably Deshayesites forbesi Zone, El Vendrell, Garraf (Spain).
Fig. 2a-c: Heminautilus lallierianus (, 1841). Specimen no. 85-3, Early Aptian, upper part of the Deshayesites forbesi Zone?, Gurgy (Yonne, France). Coll. MHNA.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus lallierianus (, 1841). Specimen no. PM95, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. .
Fig. 2a-b: Heminautilus lallierianus (, 1841). Specimen no. MGB 22607, Early Aptian, Garraf (Spain).
Fig. 3a-b: Heminautilus lallierianus (, 1841). Specimen no. MPP.B-EM.69, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, banc no. 148, La Bédoule (Bouches du Rhône, France). Coll. Frau, FSM.

Click on thumbnail to enlarge the image.
Fig. 1a-c: Heminautilus lallierianus (, 1841). Lectotype no. MNHN.F.A32800, coll d'Orbigny (no. 5573), Early Aptian, upper part of the Deshayesites forbesi Zone?, Gurgy (Yonne, France). Coll. d'Orbigny.
Fig. 2: Heminautilus lallierianus (, 1841). Specimen no. 77-543, Early Aptian, upper part of the Deshayesites forbesi Zone?, Gurgy (Yonne, France). Coll. MHNA.
Fig. 3a-b: Heminautilus lallierianus (, 1841). Specimen no. tl028, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
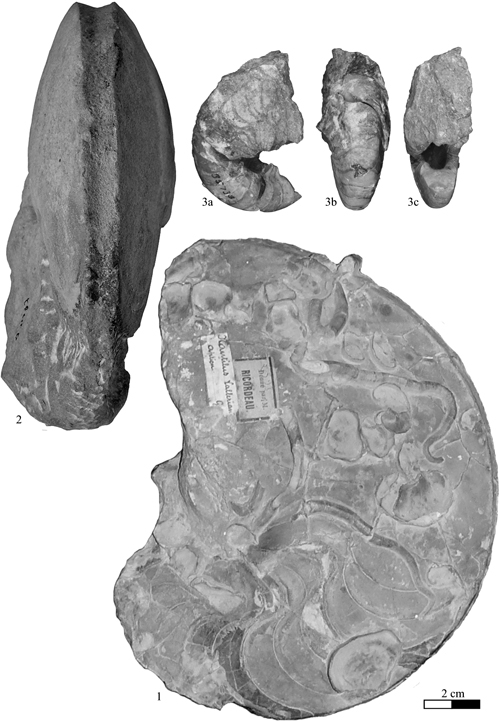
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. 77-543, Early Aptian, upper part of the Deshayesites forbesi Zone?, Gurgy (Yonne, France). Coll. MHNA.
Fig. 2: Heminautilus lallierianus (, 1841). Specimen no. tl056, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 3a-c: Heminautilus etheringtoni , 1946. Specimen no. 35733, upper part of the Early Aptian, locality A (, 1946), Tolima department (Colombia). Coll. MPUC.
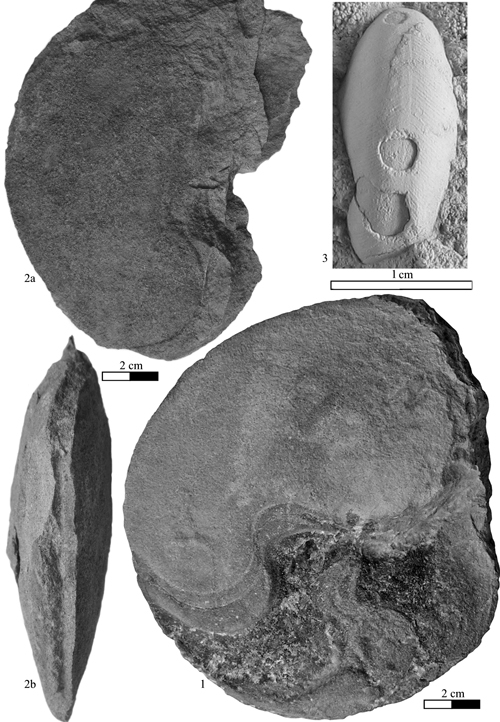
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. tl056, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2a-b: Heminautilus lallierianus (, 1841). Specimen no. BRG103, upper part of the Deshayesites forbesi Zone, bed no. 12, Pierrelatte section (Drôme, France). Coll. .
Fig. 3: Heminautilus saxbii (, 1848). Specimen no. PUAB 48158, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. VI009, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 2a-b: Heminautilus etheringtoni , 1946. Specimen no. MB RE1962a, Early Aptian, Tibú Formation, Seboruco section, east of Mérida (Venezuela).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus lallierianus (, 1841). Specimen no. tl069, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2a-c: Heminautilus ? tyosiensis ( & , 1953). Holotype no. NMNS P1-4200, upper part of the Early Aptian (= Dufrenoyia furcata Zone), Tokawa quarry, Tyôsi Peninsula, Tiba Prefecture (Japan).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. tl078, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2a-b: Heminautilus ? tyosiensis ( & , 1953). Specimen no. NMNS PM6858, upper part of the Early Aptian (= Dufrenoyia furcata Zone), Tyôsi Peninsula, Tiba Prefecture (Japan).
Fig. 3a-c: Heminautilus ? tyosiensis ( & , 1953). Specimen no. NMNS PM6859, upper part of the Early Aptian (= Dufrenoyia furcata Zone), Tyôsi Peninsula, Tiba Prefecture (Japan).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. tl029, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2: Heminautilus lallierianus (, 1841). Specimen no. tl078, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 3a-b: Heminautilus ? tyosiensis ( & , 1953). Specimen no. NMNS PM6858, upper part of the Early Aptian (= Dufrenoyia furcata Zone), Tyôsi Peninsula, Tiba Prefecture (Japan).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. tl029, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2a-b: Heminautilus stantoni (, 1940). Plastotype no. 35688, uppermost part of the Early Aptian, probably Dufrenoyia justinae Zone, locality M15, near Murfreesboro (Arkansas, USA). Coll. BEG.
Fig. 3: Heminautilus stantoni (, 1940). Holotype no. USNM103260, uppermost part of the Early Aptian, probably Dufrenoyia justinae Zone, locality M15, near Murfreesboro (Arkansas, USA).
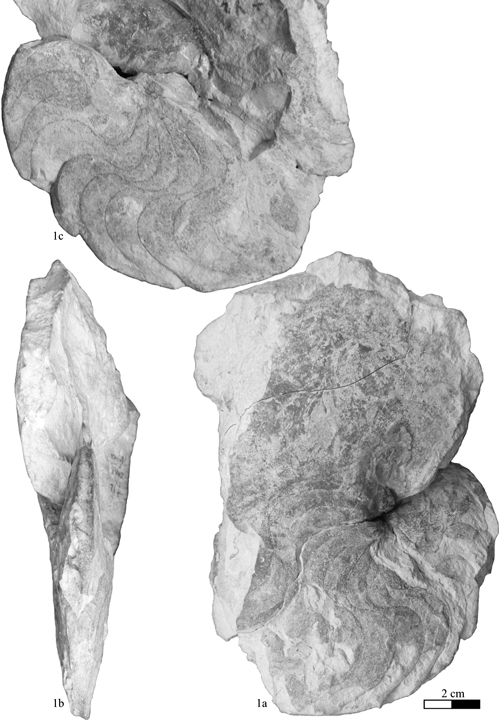
Click on thumbnail to enlarge the image.
Fig. 1a-c: Heminautilus lallierianus (, 1841). Specimen no. 12573, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, La Bédoule (Bouches du Rhône, France). Coll. FSM.
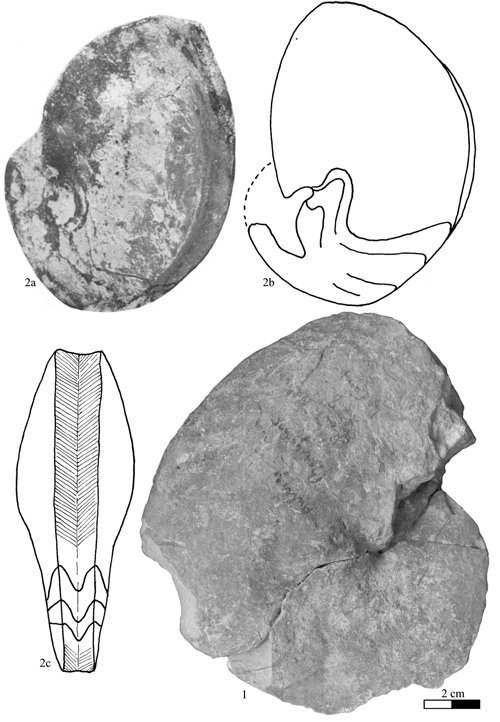
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. 12572, Early Aptian, Hameau of Les Roux, Cuges-les-Pins (Bouches du Rhône, France). Coll. FSM.
Fig. 2a-c: Heminautilus ? japonicus sp. nov. Unregistered specimen, Late Barremian (= Imerites giraudi Zone?), north of Yuasa-machi, Wakayama Prefecture (Japan). Reproduction of the illustration of & (1976).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus aff. lallierianus (, 1841). Specimen no. IGM 10050, Early Aptian, Dufrenoyia justinae Zone, bed no. 15, Mina Texali, Santa Isabel Atenayuca, Puebla State (Mexico).
Fig. 2: Josanautilus lacerdae (, 1870). Specimen no. PUAB 48057, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 3a-b: Josanautilus lacerdae (, 1870). Specimen no. PUAB 48276, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus aff. lallierianus (, 1841). Specimen no. MPL1, Early Aptian, Dufrenoyia justinae Zone, Sierra del Rosario, Durango State (Mexico).
Fig. 2a-b: Heminautilus lallierianus (, 1841). Specimen no. UPMC-250, Early Aptian, upper part of the Deshayesites forbesi Zone?, Gurgy (Yonne, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus lallierianus (, 1841). Specimen no. AMNH 25455, median part of the Early Aptian, Harrar region (Ethiopia). Reproduction of the illustration of (1943).
Fig. 2: Josanautilus lacerdae (, 1870). Specimen no. PUAB 48222, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
Fig. 3a-b: Josanautilus lacerdae (, 1870). Specimen no. PUAB 48302, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus etheringtoni , 1946. Holotype no. 35731, upper part of the Early Aptian, locality A (, 1946), Tolima department (Colombia). Coll. MPUC.
Fig. 2a-b: Josanautilus lacerdae (, 1870). Holotype no. MNCN I-14855, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Repoduction of the illustration of & (2006).
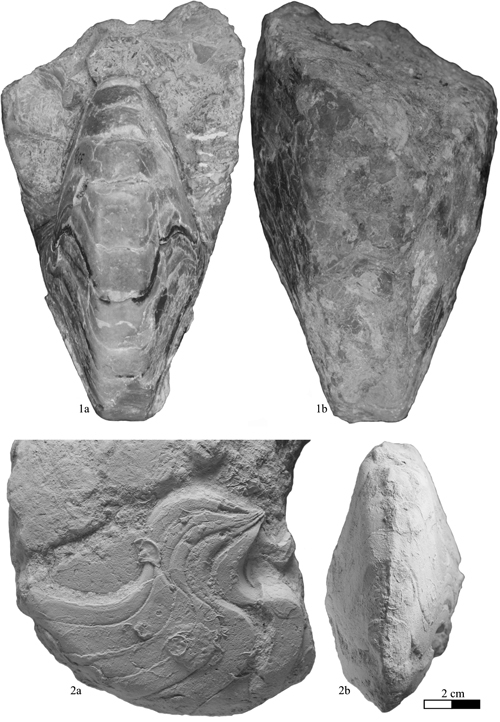
Click on thumbnail to enlarge the image.
Fig. 1a-b: Heminautilus etheringtoni , 1946. Holotype no. 35731, upper part of the Early Aptian, locality A (, 1946), Tolima department (Colombia). Coll. MPUC.
Fig. 2a-b: Josanautilus lacerdae (, 1870). Specimen no. PUAB 48178, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain).
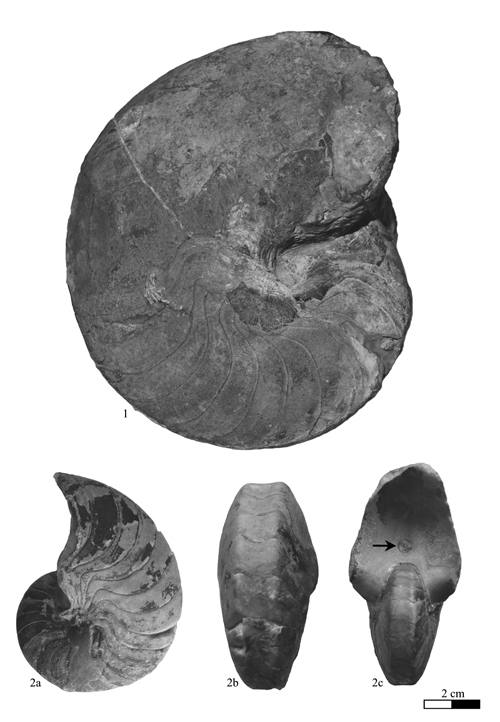
Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus etheringtoni , 1946. Specimen no. UN-DG-NT-001, Early Aptian, Firavitoba, Boyacá department (Colombia). Coll. UNCB.
Fig. 2a-c: Heminautilus etheringtoni , 1946. Specimen no. M3, Early Aptian, Quebrada el Cobre, Payandé, Tolima department (Colombia). Coll. UNCB.

Click on thumbnail to enlarge the image.
Fig. 1: Heminautilus etheringtoni , 1946. Specimen no. UN-DG-NT-001, Early Aptian, Firavitoba, Boyacá department (Colombia). Coll. UNCB.
Fig. 2a-b: Heminautilus etheringtoni , 1946. Specimen no. 124, Early Aptian, Quebrada el Cobre, Payandé, Tolima department (Colombia). Coll. UNCB.

Click on thumbnail to enlarge the image.
Fig. 1: Josanautilus lacerdae (, 1870). Specimen no. VI011, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 2: Josanautilus lacerdae (, 1870). Specimen no. Se-11.1, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mas Segura, Maestrat Basin (Spain). Coll. MV.
Fig. 3: Josanautilus lacerdae (, 1870). Specimen no. A-148, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Josa, Province of Teruel (Spain). Coll. PUAB.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Josanautilus lacerdae (, 1870). Specimen no. VI011, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 2: Josanautilus lacerdae (, 1870). Specimen no. CH01, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .

Click on thumbnail to enlarge the image.
Fig. 1: Josanautilus lacerdae (, 1870). Specimen no. VI011, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 2: Josanautilus lacerdae (, 1870). Specimen no. CH01, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 3: Josanautilus lacerdae (, 1870). Specimen no. Se-11.1, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Mas Segura, Maestrat Basin (Spain). Coll. MV.

Click on thumbnail to enlarge the image.
Fig. 1a-b: Josanautilus lacerdae (, 1870). Specimen no. tl074, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Le Teil (Ardčche, France). Coll. .
Fig. 2a-b: Heminautilus ? japonicus sp. nov. Holotype no. SH-1, Late Barremian (= Imerites giraudi Zone?), locality 203, sector of Sanchu, Kwanto Moutains, Gunma Prefecture (Japan). Coll. E. . Reproduction of the illustration of et al. (1984).

Click on thumbnail to enlarge the image.
Fig. 1a-b: Josanautilus lacerdae (, 1870). Specimen no. MHNG GEPI 83570, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Groumaud section, Bourg-Saint-Andéol (Ardčche, France). Coll. A. .
Fig. 2: Josanautilus aff. lacerdae (, 1870). Specimen no. VI010, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Fig. 3: Heminautilus ? japonicus sp. nov. Holotype no. SH-1, Late Barremian (= Imerites giraudi Zone?), locality 203, sector of Sanchu, Kwanto Moutains, Gunma Prefecture (Japan). Coll. E. Shimizu. Reproduction of the illustration of et al. (1984).

Click on thumbnail to enlarge the image.
Fig. 1: Josanautilus lacerdae (, 1870). Specimen no. MHNG GEPI 83570, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Groumaud section, Bourg-Saint-Andéol (Ardčche, France). Coll. A. .
Fig. 2a-b: Josanautilus aff. lacerdae (, 1870). Specimen no. VI010, Early Aptian, Deshayesites forbesi Zone, Roloboceras hambrovi Subzone, Viviers (Ardčche, France). Coll. .
Appended table: List of and information on the specimens of Nautiloidea , 1825, studied, referred to or depicted.
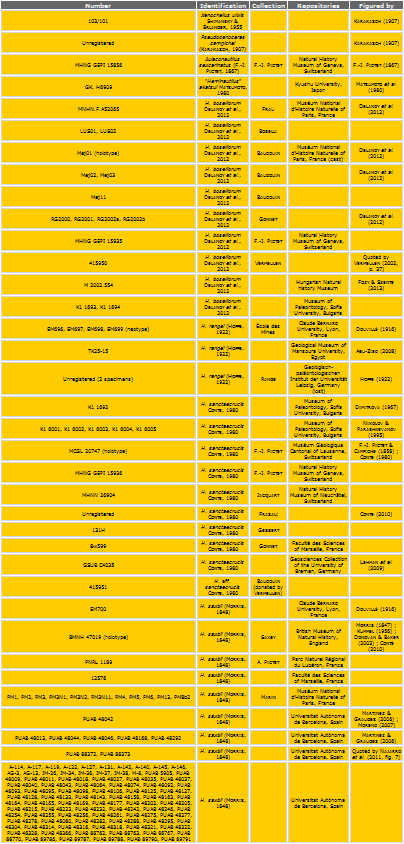
Click on thumbnail to enlarge the image.