◄ Carnets Geol. 16 (12) ►
![]()
Contents
[Introduction]
[Stratigraphic data]
[Description of cactus fossil molds]
[Taphonomy] [Interpretation] [Conclusions]
and ... [Bibliographic references]
Geosciences Department, The University of Tulsa, 800
S. Tucker Ave, Tulsa, OK 74104-3189 (U.S.A.)
Department of Earth & Environmental Sciences, Tulane University, 101 Blessey Hall, New Orleans, LA 70118 (U.S.A.)
|
Erratum: The above text should read: Zexuan WangDepartment of Earth and Environmental Sciences, University of New Orleans, 2000 Lakeshore Drive, New Orleans, LA 70148 (U.S.A.) |
Research Institute of Petroleum Exploration & Development-Langfang, Hebei 065007 (China)
Oklahoma State University, School of Geology, 105 Noble Research Center, Stillwater, OK 74078 (U.S.A.)
Published online in final form (pdf) on May 19, 2016
[Editor: Bruno Granier]
![]()
Well-preserved vertical molds of a stalk-like fossil are in growth position in the upper part of the Upper Albian Fort Lancaster Formation exposed in West Texas. The Fort Lancaster Formation is the uppermost Albian carbonate unit deposited on the shallow water Barremian-early Cenomanian Comanche shelf. The Fort Lancaster represents one long-term transgressive-regressive cycle capped by a regional unconformity overlain by lower Cenomanian deeper water Del Rio Shale.
The erect stalk-like molds are up to 1 m tall and about 6 cm in diameter. The mold walls are composed of six pairs of angular ridges and grooves evenly spaced at an angle of 60° to each other. The ridge crests are dotted by paired ringed depressions with small central nodes; these negative impressions indicate the presence of oval rings with small depressions inside of each ring. The diameter of the stalk is the same at its base as at its top indicating that growth did not expand the organism but extended it upward. These stalk-like organisms appear to be rooted on disconformable bedding surfaces that were exposed to the atmosphere. Carbon isotope data shows the classic negative shift below subaerial contacts. Subsequently sea level rose and muddy carbonate sediment buried the stalk. The absence of original wall material suggests that the wall was composed of readily decayed or dissolved material such as organic matter or aragonite. Stalk-like organisms of this size and shape are rare and potential types are cacti, plant reeds, Porifera, or Scleractinian Cnidaria.
Albian; Comanche shelf; plant-like fossil.
Scott R.W., Wang X., Lai X. & Wang Y. (2016).- Late Albian cactus in shallow carbonate shelf, West Texas.- Carnets Geol., Madrid, vol. 16, no. 12, p. 337-346.
|
Erratum: The above text should read: Scott R.W., Wang Z., Lai X. & Wang Y. (2016).- Late Albian cactus in shallow carbonate shelf, West Texas.- Carnets Geol., Madrid, vol. 16, no. 12, p. 337-346. |
Un cactus de l'Albien supérieur en contexte de plate-forme carbonatée peu profonde, Ouest du Texas.- Des moules verticaux bien conservés d'un fossile en forme de tige ont été trouvés en position de croissance dans la partie supérieure de la Formation de Fort Lancaster (Albien supérieur) affleurant dans l'ouest du Texas. La Formation de Fort Lancaster est une unité carbonatée de l'Albien terminal déposée dans les eaux peu profondes de la plate-forme de Comanche (Barrémien-Cénomanien inférieur). La Formation de Fort Lancaster représente un cycle transgressif-régressif de long terme coiffé d'une discontinuité régionale, elle-męme recouverte par la Formation argileuse Del Rio déposée en eaux plus profondes du Cénomanien inférieur.
Les moules dressés en forme de tige font jusqu'ŕ 1 m de hauteur et environ 6 cm de diamčtre. Les parois des moules comportent six paires de crętes anguleuses et de sillons. Ces crętes réguličrement espacées font un angle de 60° entre elles. Les crętes des rides sont ponctuées de dépressions annulaires appariées avec de petits nśuds en leur centre ; ces impressions en négatif indiquent la présence d'anneaux ovales au centre desquels figurent de petites dépressions. Le diamčtre de la tige ŕ sa base est le męme qu'ŕ son sommet, indiquant que l'organisme ne croissait pas radialement mais s'étirait vers le haut. Ces organismes en forme de tige semblent prendre racine sur des surfaces de discordance émersives. Les données du carbone isotopique montrent une baisse négative classique sous les surfaces d'émersion. Par conséquent, ŕ la faveur d'une baisse du niveau marin, la tige a été enfouie dans des sédiments boueux carbonatés. L'absence du matériel originel de la paroi laisse penser que cette derničre était composée d'une matičre facilement putrescible ou soluble, telle que de la matičre organique ou de l'aragonite. Les organismes en forme de tige de cette taille et de cette morphologie restent rares ; les candidats envisageables sont des cactus, des roseaux, des porifčres, ou des cnidaires scléractinaires.
Albien ; plate-forme de Comanche ; fossile phytoďde.
Well-preserved vertical molds of a fossil stalk-like organism are in
growth position in the upper part of the Upper Albian Fort Lancaster Formation
exposed in U.S. 290 road cuts about 10 km (6 mi) southeast of its intersection
with I-10 and about 24 km (15 mi) east of Sheffield, Texas (Fig. 1 ![]() )
(N30°40.720', W101°40.372',
Crocket County, Texas). The steep road descends into the Pecos River Valley and
these outcrops are the type section of the Fort Lancaster (Smith et
al., 2000). During field work in 2013 Xejuan Wang noticed these
unusual molds. Subsequent study convinced the authors that these vertical molds
were fossils that deserved careful description.
)
(N30°40.720', W101°40.372',
Crocket County, Texas). The steep road descends into the Pecos River Valley and
these outcrops are the type section of the Fort Lancaster (Smith et
al., 2000). During field work in 2013 Xejuan Wang noticed these
unusual molds. Subsequent study convinced the authors that these vertical molds
were fossils that deserved careful description.
The Fort Lancaster Formation is the uppermost Albian carbonate unit deposited on the Barremian-early Cenomanian Comanche shelf that extended from Florida to Arizona on the northern margin of the proto-Gulf of Mexico (Rose, 1972; Smith et al., 2000; Scott & Kerans, 2004). The purpose of this paper is to describe the stalk-like molds, and to propose that a cactus was the most likely originator of these molds.
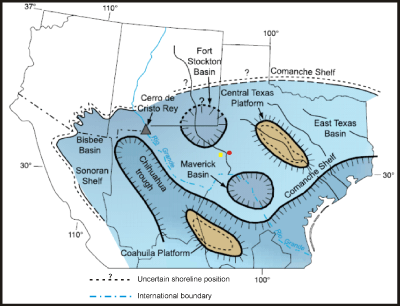
Click on thumbnail to enlarge the image.
Figure 1: Paleogeographic map of the Albian Comanche shelf showing location of outcrop of Fort Lancaster Formation on U.S. 290 and fossil molds (red dot); yellow box is site of Sheffield (after Scott, 2003).
The Fort Lancaster Formation is a dominantly limestone lithostratigraphic unit exposed north and south along the Pecos River valley in West Texas and in adjacent streams flowing to the Rio Grande River at the International border of the United States and Mexico (Smith et al., 2000). A late Albian age is defined by ammonites in the basal part and by caprinid rudists and benthic foraminifers in the upper part of the formation (Young, 1966; Scott, 2002a, 2002b; Scott & Kerans, 2004). The Fort Lancaster represents one, long-term, transgressive-regressive cycle capped by a regional unconformity overlain by the lower Cenomanian deep shelf Del Rio Shale in some places and by the lowermost Cenomanian Buda Limestone where the Del Rio has pinched out (Smith et al., 2000; Lai, 2014; Y. Wang, 2014).
The Fort Lancaster Formation is divided into a basal shale and limestone interval about 15 m thick that is correlated with the Burt Ranch Member in Central Texas (Rose, 1972; Smith et al., 2000). The base of the Burt Ranch is a regional, disconformable sequence boundary that correlates with lower-upper Albian Al SB Wa 1 in North Texas (Scott et al., 2003; Gargili, 2013; Lai, 2014; Y. Wang, 2014). In the Fort Lancaster road section the top of the underlying Fort Terrett Formation is a bored, iron-stained contact; a 2 ppm positive carbon isotope shift in the uppermost 1 m of the Fort Terrett is evidence that the contact at this site was subaerially exposed (Gargili, 2013; Lai, 2014).
The uppermost 15 m of the Fort Lancaster Formation is composed of six
m-scale shoaling-up carbonate cycles, some of which are separated by
iron-stained contacts with red clay (Smith et al.,
2000; Lai, 2014) (Figs.
2 ![]() - 3
- 3 ![]() ). These cycles were deposited during the aggrading
highstand relative sea-level. The stalk-like fossil molds are found in the upper
7 to 8 m of the Fort Lancaster in basal and upper parts of carbonate cycles 2,
3, 4, 5, and 6. Carbonate microfacies of cycle 4 grade from bioclastic packstone
with rudist and echinoderm bioclasts at the base to wackestone with miliolid and
other benthic foraminifera at the top. Cycle 5 is composed of bioclastic
wackestone with benthic foraminifera, requienid bivalves, gastropods, calcareous
algae, ostracodes, and echinoderm clasts. Carbonate facies are fully described
by Gargili (2013) and Lai
(2014). These carbonate lithologies
suggest that the cycles were deposited below local wave base in an interior
shelf lagoon and shoaled up to sea level. About 80 km south was the high-energy
Devils River shelf margin trend on the northern margin of the McKnight Basin.
The shallow shelf extended westward and eastward at least 100 km in each
direction; northward lay shoreline. Relative sea-level fall exposed these beds
to subaerial conditions, which is indicated by the positive carbon isotope shift (Fig.
2
). These cycles were deposited during the aggrading
highstand relative sea-level. The stalk-like fossil molds are found in the upper
7 to 8 m of the Fort Lancaster in basal and upper parts of carbonate cycles 2,
3, 4, 5, and 6. Carbonate microfacies of cycle 4 grade from bioclastic packstone
with rudist and echinoderm bioclasts at the base to wackestone with miliolid and
other benthic foraminifera at the top. Cycle 5 is composed of bioclastic
wackestone with benthic foraminifera, requienid bivalves, gastropods, calcareous
algae, ostracodes, and echinoderm clasts. Carbonate facies are fully described
by Gargili (2013) and Lai
(2014). These carbonate lithologies
suggest that the cycles were deposited below local wave base in an interior
shelf lagoon and shoaled up to sea level. About 80 km south was the high-energy
Devils River shelf margin trend on the northern margin of the McKnight Basin.
The shallow shelf extended westward and eastward at least 100 km in each
direction; northward lay shoreline. Relative sea-level fall exposed these beds
to subaerial conditions, which is indicated by the positive carbon isotope shift (Fig.
2 ![]() ). This muddy surface was
available to be occupied by terrestrial plants.
). This muddy surface was
available to be occupied by terrestrial plants.
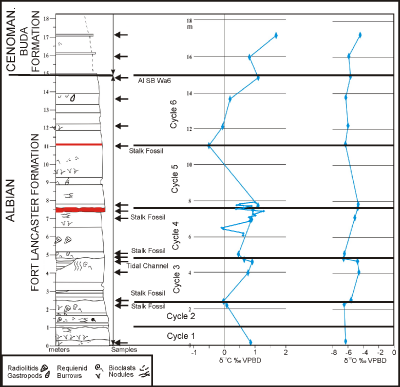
Click on thumbnail to enlarge the image.
Figure 2: Measured section of upper part of Fort Lancaster road cut showing position of stalk fossils and meter-scale depositional cycles; arrows indicate positions of isotope sample set 1; isotope sample set 2 are smaller diamonds spanning red bed (black). Contact between Fort Lancaster and Buda formations is regional Albian sequence boundary Wa 6 defined in Fort Worth section between Main Street and Grayson formations (Scott et al., 2003).

Click on thumbnail to enlarge the image.
Figure 3: East face of roadcut U.S. 290 below parking site. Note three persons in lower right for scale. Dotted lines define bedding cycles 4-7 in the uppermost 10 m of the Fort Lancaster Formation. Red arrows point to molds of fossil plants. Yellow 'Xs' mark the second set of isotope samples.
At the contact between cycles 4 and 5 is a reddish, iron-rich,
bioclastic mudstone up to 15 cm thick. The top of cycle 4 is red stained, sharp
and irregularly wavy with relief of 5-8 cm. The surface is scalloped by simple,
low-relief bowls ranging in width from 5 to 20 cm and up to 5 cm deep. These may
be footprint impressions; an alternative origin may be simple paleokarst
depressions that evolved from potholed paleokarst (Vanstone,
1998). A
penecontemporaneous fracture extends downward 1.7 m from the base of one karst
depression to the base of cycle 4 (Fig. 4 ![]() ).
This fracture is filled mainly with red clay and a few pebble- to cobble-sized
limestone clasts. Evidently the muddy carbonate lithified early and fractured
during subaerial exposure; the fracture then was filled by soil, bioclasts, and
limestone breccia. A positive carbon isotope shift of 0.64 ppm from base to top
of the cycle is overlain by a negative shift of 1.0 ppm at the base of cycle 5 (Fig.
2
).
This fracture is filled mainly with red clay and a few pebble- to cobble-sized
limestone clasts. Evidently the muddy carbonate lithified early and fractured
during subaerial exposure; the fracture then was filled by soil, bioclasts, and
limestone breccia. A positive carbon isotope shift of 0.64 ppm from base to top
of the cycle is overlain by a negative shift of 1.0 ppm at the base of cycle 5 (Fig.
2 ![]() ). These data support the interpretation that the contact between cycles 4
and 5 was subaerially exposed. This red bed has been mapped in many sections in
the Pecos River valley and farther east where the color changes to green and
black (Smith et al., 2000; Rose,
1972; Halley
& Rose, 1977; Y. Wang,
2014).
). These data support the interpretation that the contact between cycles 4
and 5 was subaerially exposed. This red bed has been mapped in many sections in
the Pecos River valley and farther east where the color changes to green and
black (Smith et al., 2000; Rose,
1972; Halley
& Rose, 1977; Y. Wang,
2014).

Click on thumbnail to enlarge the image.
Figure 4: Stratigraphic setting and morphology of cactus-like fossils in east-side road cut of U.S. 290. A, Fossil mold specimen top of cycle 4 overlain by red clay bed; note shallow depressions on top of bed 4 into which penecontemporaneous fracture ends. B, C, D. Fossil mold specimen 1 at top of cycle 5. C, D. Mold of stalk-like fossil 1 showing three faces and flat base; note outline of pores on angular ridges. E. Natural and outlined views into mold base showing poorly preserved ridges and grooves; note apparent absence of root-like structures in base. F. Measurement of mold faces in clay cast and reconstruction of circular outline of complete six-part symmetry of fossil; 'M' = mean angle. The 89.5° angle was measured on the distorted clay cast; to accommodate six facets within a 360° circle, the angle is projected at 60°.
The stalk-like molds are columnar cylindrical stems standing up to 1 m
tall and are about 6 cm in diameter (Fig. 4.A-B ![]() ); they are interpreted to be molds of a cactus. Branches are not
evident. The mold walls are composed of evenly spaced angular ribs and grooves (Fig.
4.C
); they are interpreted to be molds of a cactus. Branches are not
evident. The mold walls are composed of evenly spaced angular ribs and grooves (Fig.
4.C ![]() )
similar to the modern genus, Cereus
among others. Six pairs of ribs and grooves are arranged at an angle of 60° to
each other (Fig. 4.D-E
)
similar to the modern genus, Cereus
among others. Six pairs of ribs and grooves are arranged at an angle of 60° to
each other (Fig. 4.D-E ![]() ). The angle
between the faces was measured from the plasticene mold which was formed by
impressing the clay into the mold; however, it was slightly distorted by
extraction and subsequent handling. Thus the measurements vary on different
profiles of the cast and have been averaged ((Fig. 4.F-1
). The angle
between the faces was measured from the plasticene mold which was formed by
impressing the clay into the mold; however, it was slightly distorted by
extraction and subsequent handling. Thus the measurements vary on different
profiles of the cast and have been averaged ((Fig. 4.F-1 ![]() )). The cross section profile was then constructed from these
measurements assuming that the fossil outline was circumscribed by a circle (Fig.
4.F-2
)). The cross section profile was then constructed from these
measurements assuming that the fossil outline was circumscribed by a circle (Fig.
4.F-2 ![]() ). Ribs and flutes of some specimen molds are less well
expressed suggesting that these specimens were more inflated by water than
others as succulents may be.
). Ribs and flutes of some specimen molds are less well
expressed suggesting that these specimens were more inflated by water than
others as succulents may be.
The rib crests are dotted by paired depressions with small nodes; these
negative impressions indicate the presence of oval rings with small depressions
inside of each ring (Fig. 4.C ![]() ). The
sets of rings and pits are spaced about 1 cm apart along the rib crests. These
dimensions are approximate because the structures are preserved as negative
molds. These structures are similar to cactus tubercles and areoles from which
spines grow. The bases of two molds are flat and apparently with no indication
of rootlets or taproots (Fig. 4.D & F
). The
sets of rings and pits are spaced about 1 cm apart along the rib crests. These
dimensions are approximate because the structures are preserved as negative
molds. These structures are similar to cactus tubercles and areoles from which
spines grow. The bases of two molds are flat and apparently with no indication
of rootlets or taproots (Fig. 4.D & F ![]() ).
The diameter of the stalk is the same at its base as at its top indicating that
growth did not expand the organism but extended it upward. The absence of
original wall structure suggests that the wall was composed readily decayed or
dissolved material such as organic matter or aragonite.
).
The diameter of the stalk is the same at its base as at its top indicating that
growth did not expand the organism but extended it upward. The absence of
original wall structure suggests that the wall was composed readily decayed or
dissolved material such as organic matter or aragonite.
If these molds were formed by chlorophyll-bearing plants, they potentially would indicate shallow water depth within the photic zone. An alternative explanation is that these plants grew in the exposed marine mud proximal to the coastline between periodic inundations.

Click on thumbnail to enlarge the image.
Figure 5: Variations among stalk-like molds. A. Mold at top of cycle 4 truncated by red paleosol. B. Slightly curved specimen at basal contact of cycle 4. C. Mold in upper part of cycle 2. D-F. Closeups of mold in Figure 4; C. Base of mold buried in limestone; E. Overview of mold. F. closeup of mold facet with node impressions [B-F. Scale bars are 5 cm].
The mode of preservation of these molds in shallow-water carbonate sediment is a dilemma. At least two hypotheses are reasonable. 1) The specimens grew on the exposed substrate and were subsequently buried. 2) The specimens were buried as they grew. This latter process seems untenable if the cactus-like plant was not salt-tolerant and required a dry terrestrial habitat and fresh water as modern cacti do.
The bases of many stalk-like molds are set on exposure contacts at top of meter-scale depositional cycles. Also some terminate at the red clay exposure contact at tops of cycles. These relationships suggest that the stalk-like organisms began growing on the subaerial surface of the vast exposed surface. Subsequently sea level rose and sedimentation buried the stalk, which was terminated by subaerial exposure of the concurrent cycle. The organism terminated at the subaerial surface either ending growth or it was truncated during exposure.
This first scenario requires that the stalk remained erect as sea level deepened and muddy carbonate sediment buried it. Wave energy, tides or other currents would have been very minimal so that the stalk was not toppled during the beginning of rise. The virtual absence of water movement is conceivable across the 100 km+ wide flat substrate as sea level rose slowly and water depth was on the order of centimeters. If sedimentation rate was on the order of 6000 cm/1,000 year as it has been on the leeward side of the Great Barrier Reef (Enos, 1991, p. 73), then the meter-tall plant would be buried in 16-17 years.
Stalk-like fossil organisms of this size and shape are rare; potential candidates are reeds such as cacti, and marine Porifera and Scleractinian Cnidaria. Early Cretaceous Angiosperm evolution has been documented using molecular data combined with morphology of leaves and pollen (Doyle, 2012; Doyle & Upchurch, 2014) however none have a similar stalk-like morphology. Early Albian lineages were "Nymphaeales, Austrobaileyales, Chloranthaceae, magnoliids (Canellales and Magnoliales), monocots, and eudicots" (Doyle & Upchurch, 2014). During Middle and Late Albian angiosperms diversified into modern clades. In Europe Barremian-Albian plant impressions show that angiosperms progressively diversified their ecological habitats from braided rivers and freshwater swamps to flood plains, levees, estuaries, and salt marshes (Coiffard et al., 2007).
Early Cretaceous gymnosperms and angiosperms were widespread and diverse on the Comanche shelf on the northern Gulf of Mexico. The fossil plant record of the Comanche shelf is documented primarily by spore and pollen and sparse leaves and wood in strand plain and alluvial plain sandstone (Retallack & Dilcher, 1986). The conifers Weichselia and Frenelopsis, which grew in brackish swamps (Coiffard et al., 2007), together with seed ferns and angiosperms are preserved in chert of the middle-upper Albian Edwards Formation (Serlin, 1982). Pollen and spore microfossils in marine shale-marl of the upper Albian Washita Group indicate that a diverse plant assemblage of Lycophytea, Pterophyta, Gymnospermae, and Angiospermae populated the northern shoreline of the Comanche shelf (Hedlund, 1966; Hedlund & Norris, 1968; Wingate, 1980; Srivastava, 1981). Gymnosperm pollen was most abundant in the upper Albian Denton Formation, Washita Group, in southern Oklahoma and angiosperms were about ten percent of the assemblage. Westward in the Fort Stockton area land-derived spores and pollen were abundant in the Middle Albian Fort Terrett Formation and in the upper part of the Fort Lancaster Formation, Washita Group (Scott & Kidson, 1977), but cactus pollen has not been reported. The marine shelf supported diverse calcareous algae (Johnson, 1968), none of which has a columnar morphology.
A modern fresh-water, aquatic plant that may be analogous is Ceratophyllum or hornwort of the Family Ceratophyllaceae, which has a cosmopolitan Paleocene to Miocene fossil record (Herendeen et al., 1990). The stems are much narrower than the Fort Lancaster mold and have narrow spiny leaves originating from nodes on the stem (Herendeen et al., 1990, Figs. 19-20; Dilcher & H. Wang, 2009). Fossil species are differentiated by their ovate spiny fruits. Modern species grow up to 3 m in length and float below the water surface. The stems do not have roots and leaf whorls attach to nodes on the stem. Phylogenetic analyses suggest that herbaceous groups such as Ceratophyllum are near the base of the angiosperm clade (Doyle, 1996, 2012; Doyle & Upchurch, 2014). Doyle and Upchurch suggested that these molds could be trace fossils rather than plant molds (personal communication, December 21, 2015). However the repetition of morphology in multiple molds and the uniform structure indicates that these are molds of an organism and not traces made by the variable behavior of an animal.
The morphology of these molds and habitat setting on an exposed carbonate substrate during low sea level are consistent with a cactus plant. However that these were molds of cactus plants seems improbable because no fossils or undisputed cactus pollen are known (Nyffeler, 2002; Nyffeler & Eggli, 2010). Two current hypotheses propose that Cactaceae evolved either in the Late Cretaceous or more likely in the younger part of the Paleogene about 30 myr ago. An Eocene Opuntoid cactus, about 55-34 Ma old, was described from Utah (Chaney, 1944). If these molds were formed around cacti, these would be evidence of the oldest cactus, which would alter the presumed evolutionary history. Alternatively, these plants may have been members of an early clade that became extinct and unrelated to Cactacea as understood today.
An origin by a stalk-shaped marine animal is unlikely because this morphology is uncommon. Although Early Cretaceous Porifera species are poorly known on the Comanche shelf, known species are small, nodular or encrusting; none are erect, six-sided structures. Spicules are in chert nodules of the Albian Edwards Formation (Merrill, 1895), and sponges have been reported in the lower Cenomanian Buda Formation (Adkins, 1928). Two small nodular sponge species were described from the Lower Albian Mural Formation in Arizona and Sonora, one of which was noted also in the Fredericksburg shelf margin facies in Texas (Rigby & Scott, 1981). Encrusting stromatoporoids have been noted in shelf margin buildups but not fully described (Scott, 1990).
Scleractinian corals are diverse and widespread in Comanchean strata of North America (Wells, 1933; Filkorn, 2003). Both colonial and solitary species are known. The six-part radial symmetry is the diagnostic feature and skeletons were composed of aragonite. However none have tall, stalk-like morphologies and most are preserved as secondary calcite not as molds.
Well preserved vertical molds of a fossil reed-like plant are in growth position in the upper aggrading part of the Upper Albian Fort Lancaster Formation exposed in the Pecos River valley, West Texas. The Fort Lancaster Formation is the uppermost Albian carbonate unit deposited on the Barremian-early Cenomanian Comanche shelf. The Fort Lancaster Formation is dominantly limestone of late Albian age defined by ammonites in its basal part and by caprinid rudists and benthic foraminifers in its upper part. The Fort Lancaster represents one long-term transgressive-regressive cycle capped by a regional unconformity overlain by the lower Cenomanian Limestone. The upper 15 m interval is composed of six aggrading cycles separated by subaerial contacts representing times when sea level fell exposing the substrate to the air. These vast supratidal surfaces were populated by primitive cacti and possibly other plants.
The stalk-like molds are erect standing up to 1 m tall and about 6 cm in diameter. The mold walls are composed of six evenly spaced angular pairs of ridges and grooves arranged at an angle of 60° to each other. The ridge crests are dotted by paired depressions with small nodes; these negative impressions indicate the presence of oval rings at the nodes with small depressions inside of each ring. Roots are not preserved and the diameter of the molds is the same at the base as at the top. Apparently the organism began to grow on the exposed substrate and subsequently was buried as carbonate mud accumulated around it. The absence of original wall structure suggests that the wall was composed of readily decayed or dissolved material such as organic matter or aragonite. The most likely candidate as the original organism is a cactus.
The University of Tulsa Office Of Research and Sponsored Programs and the Geosciences Department generously supported research by Damla Gargili, Xin Lai, Yulun Wang, and Robert Scott. Field expenses were partly supported by Precision Stratigraphy Associates. Email discussions with J.A. Doyle and G.R. Upchurch, Jr, prompted multiple candidates. I am grateful to the reviewers, Bruno Ferré, Christopher Baldwin and Steve Hasiotis, who noted omissions and offered ways to improve the manuscript. Hasiotis noted the cactus similarity.
Adkins W.W. (1928).- Handbook of Texas Cretaceous fossils.- University of Texas Bulletin, Austin, no. 2838, p. 1-385.
Arakaki M., Christin P.-A., Nyffeler R., Lendel A., Eggli U., Ogburn R.M., Spriggs E., Moore M.J. & Edwards E. (2011).- Contemporaneous and recent radiations of the world's major succulent plant lineages.- Proceedings of the National Academy of Sciences, vol. 108, p. 8379-8384. URL: http://www.pnas.org/content/108/20/8379
Chaney R.W. (1944).- A fossil cactus from the Eocene of Utah.- American Journal of Botany, St. Louis, vol. 31, p. 507-528.
Coiffard C., Gomez B. & Thévenard F. (2007).- Early Cretaceous Angiosperm invasion of western Europe and major environmental changes.- Annals of Botany, Oxford, vol. 100, p. 545–553.
Dilcher D.L. & Wang H. (2009).- An Early Cretaceous fruit with affinities to Ceratophyllaceae.- American Journal of Botany, St. Louis, vol. 96, p. 2256–2269.
Doyle J.A. (1996).- Seed plant phylogeny and the relationships of Gnetales.- International Journal of Plant Sciences, Chicago, vol. 156, (6 supplement), p. S3-S39.
Doyle J.A. (2012).- Molecular and fossil evidence on the origin of Angiosperms.- Annual Review of Earth and Planetary Sciences, vol. 40, p. 301-326.
Doyle J.A. & Upchurch G.R. Jr (2014).- Angiosperm clades in the Potomac Group: What have we learned since 1977?- Bulletin of the Peabody Museum of Natural History, New Haven, vol. 55, p. 111-134.
Enos P. (1991).- Sedimentary parameters for computer modeling. In: Franseen E.K., Watney W.L., Kendall C.C.St.C. & Ross W. (eds.), Sedimentary modeling: computer simulations and methods for improved parameter definition.- Kansas Geological Survey, Bulletin 233, p. 63-99.
Filkorn H.F. (2003).- The Cretaceous corals of Mexico: Occurrences and history of research.- Revista Mexicana de Ciencias Geológicas, Querétaro, vol. 20, p. 52-78.
Gargili D. (2013, unpublished).- Middle-Upper Albian Comanche shelf sequence stratigraphy, Fredericksburg Group, Texas.- MSc. Thesis, University of Tulsa, 154 p.
Halley R.B. & Rose P.R. (1977).- Significance of fresh-water limestones in marine carbonate succession of Pleistocene and Cretaceous age. In: Bebout D.G. & Loucks R.G. (eds.), Cretaceous carbonates of Texas and Mexico: Applications to subsurface exploration.- Report of Investigations, Bureau of Economic Geology, University of Texas Austin, no. 89, p. 206-215.
Hedlund R.W. (1966).- Palynology of the Red Branch Member of the Woodbine formation (Cenomanian), Bryan County, Oklahoma.- Bulletin of the Oklahoma Geological Survey, Norman, vol. 112, p. 1-69.
Hedlund R.W. & Norris G. (1968).- Spores and pollen grains from Fredericksburg (Albian) strata, Marshall County, Oklahoma.- Pollen et Spores, vol. 10, p. 129-159.
Herendeen P.S., Les D.H. & Dilcher D.L. (1990).- Fossil Ceratophyllum (Ceratophyllaceae) from the Tertiary of North America.- American Journal of Botany, St. Louis, vol. 77, p. 7-16.
Johnson J.H. (1968).- Lower Cretaceous algae from Texas: Part 2, Pre-Buda Lower Cretaceous algae from Texas.- Colorado School of Mines Professional Contributions, Boulder, no. 4, p. 29-66.
Lai X. (2014, unpublished).- Upper Albian sequence stratigraphy and geochemical events of Comanche shelf interior Washita Group, southwest Texas.- MSc. Thesis, University of Tulsa, 93 p.
Merrill J.A. (1895).- Fossil sponges of the flint nodules in the Lower Cretaceous of Texas.- Bulletin Museum of Comparative Zoology at Harvard College, Cambridge, vol. 28, p. 1-26.
Nyffeler R. (2010).- Phylogenetic relationships in the cactus family (Cactaceae) based on evidence from trnK/matK and trnL-trnF sequences.- American Journal of Botany, St. Louis, vol. 89, p. 312-326.
Nyffeler R. & Eggli U. (2010).- A farewell to dated ideas and concepts: Molecular phylogenetics and a revised supra generic classification of the Family Cactaceae.- Schumannia, Bonn, vol. 6, p. 109-149.
Retallack G.J. & Dilcher D.L. (1986).- Cretaceous Angiosperm invasion of North America.- Cretaceous Research, vol. 7, p. 227-252.
Rigby J.K. & Scott R.W. (1981).- Sponges from the Lower Cretaceous Mural Limestone in Arizona and northern Mexico.- Journal of Paleontology, vol. 55, p. 552-562.
Rose P.R. (1972).- Edwards Group, surface and subsurface, Central Texas.- Report of Investigations, Bureau of Economic Geology, The University of Texas at Austin, no. 74, 198 p.
Scott R.W. (1990).- Models and stratigraphy of Mid-Cretaceous reef communities, Gulf of Mexico.- Concepts in Sedimentology and Paleontology, SEPM (Society for Sedimentary Geology), vol. 2, 102 p.
Scott R.W. (2002a).- Upper Albian benthic foraminifers new in West Texas.- Journal of Foraminiferal Research, Lawrence, vol. 32, no. 1, p. 43-50.
Scott R.W. (2002b).- Albian caprinid rudists from Texas re-evaluated.- Journal of Paleontology, vol. 76, p. 408-423.
Scott R.W. (2003).- Introduction. In: Scott R.W. (ed.), Cretaceous stratigraphy and paleoecology, Texas and Mexico.- Special Publications in Geology, Perkins Memorial Volume, GCSSEPM Foundation, no. 1, CD book, p. viii-xiv.
Scott R.W., Benson D.G., Morin R.W., Shaffer B.L. & Oboh-Ikuenobe F.E. (2003).- Integrated Albian-lower Cenomanian Chronostratigraphy and Paleoecology, Texas and Mexico. In: Scott R.W. (ed.), Cretaceous stratigraphy and paleoecology, Texas and Mexico.- Special Publications in Geology, Perkins Memorial Volume, GCSSEPM Foundation, no. 1, CD book, p. 277-334.
Scott R.W. & Kerans C. (2004).- Late Albian carbonate platform chronostratigraphy, Devils River Formation cycles, West Texas.- Courier Forschungs-Institut Senckenberg, Frankfurt, vol. 247, p. 129-148.
Scott R.W. & Kidson E.J. (1977).- Lower Cretaceous depositional systems, West Texas. In: Bebout D.G. & Loucks R.G. (eds.), Cretaceous carbonates of Texas and Mexico: Applications to subsurface exploration.- Report of Investigations, Bureau of Economic Geology, University of Texas Austin, no. 89, p. 169-181.
Serlin B.S. (1982).- An Early Cretaceous fossil flora from northwest Texas: its composition and implications.- Palaeontographica, vol B182, p. 52-86.
Smith C.I., Brown J.B. & Lozo F.E. (2000).- Regional stratigraphic cross sections, Comanche Cretaceous (Fredericksburg – Washita Division), Edwards and Stockton plateaus, West Texas: Interpretation of sedimentary facies, depositional cycles, and tectonics.- Bureau of Economic Geology, The University of Texas at Austin, 39 p.
Srivastava S.K. (1981).- Stratigraphic ranges of selected spores and pollen from the Fredericksburg Group (Albian) of southern United States.- Palynology, vol. 5, p. 1-26.
Vanstone S. (1998).- Late Dinantian palaeokarst of England and Wales: implications for exposure surface development.- Sedimentology, vol. 45, p. 19-37.
Wang Y. (2014, unpublished).- Chemostratigraphy of Upper Albian (Lower Cretaceous) Comanche shelf margin.- MSc. Thesis, University of Tulsa, 169 p.
Wells J.W. (1933).- Corals of the Cretaceous of the Atlantic and Gulf Coastal Plain and Western Interior of the United States.- Bulletins of American Paleontology, vol. 18, no. 67, p. 83-288.
Wingate F.H. (1980).- Plant microfossils from the Denton shale Member of the Bokchito Formation (Lower Cretaceous, Albian) in southern Oklahoma.- Oklahoma Geological Survey, Bulletin 130, p. 1-93.
Young K. (1966).- Texas Mojsisovicziinae (Ammonoidea) and the zonation of the Fredericksburg.- Geological Society of America Memoir 100, p. 5-119.