◄ Carnets Geol. 16 (16) ►
![]()
Contents
[1. Introduction]
[2. Material]
[3. Methods]
[4. Systematic paleontology]
[5. Discussion]
[Bibliographic references] and ... [Plates]
Universidad Nacional Autónoma de México (UNAM),
Instituto de Geología (IGL),
Estación Regional del Noroeste (ERNO),
Hermosillo, Sonora (México)
Published online in final form (pdf) on July 7, 2016
[Editor: Bruno Granier;
language editor: Donald Owen]

The new genus Eosiderastrea Löser, 2016, was established for Cretaceous corals formerly assigned to the extant coral genus Diploastrea. It represents the oldest member of the Family Siderastraeidae. The genus is characterised by an astreoid arrangement, occasionally although some have a plocoid or a cerioid calicular arrangement, and a sub-regular hexameral, heptameral, octameral or decameral septal symmetry. The septal blades are made of medium-sized trabeculae. Synapticulae are present. Almost a hundred specimens were examined, among them numerous type specimens. In total, 26 Cretaceous species were separated. Fifteen species were assigned to formally established species, three species were compared to one of these species, two species were described as new and the remaining six species were reported in open nomenclature. The genus occurs in the Cretaceous worldwide and ranges from the late Valanginian to the Santonian. It shows its greatest diversity in the earliest Aptian and early to middle Cenomanian.
Corals; Scleractinia; Cretaceous; taxonomy; new taxa.
Löser H. (2016).- Taxonomy and distribution of the Cretaceous coral genus Eosiderastrea.- Carnets Geol., Madrid, vol. 16, no. 16, p. 383-416.
Taxinomie et répartition du genre de coraux crétacés Eosiderastrea.- Le nouveau genre Eosiderastrea Löser, 2016, a été érigé pour des coraux crétacés attribués auparavant au genre actuel Diploastrea. Il constitue le plus ancien membre de la famille des Siderastraeidae. Ce genre est caractérisé par un arrangement astréoïde, bien que de temps en temps quelques-uns montrent un arrangement caliculaire plocoïde ou cérioïde, et une symétrie septale d'ordre 6, 7, 8 ou 10 presque régulière. Les lames septales sont constituées de trabécules de taille moyenne. Des synapticules sont présents. Près d'une centaine de spécimens furent examinés, parmi lesquels des spécimens-types. Au total, 26 espèces crétacées sont distinguées. Quinze espèces sont attribuées à des espèces déjà connues, trois sont comparées à l'une d'elles, deux sont décrites comme nouvelles, et les six autres restantes sont laissées en nomenclature ouverte. Le genre est présent dans le Crétacé du monde entier et existe depuis le Valanginien supérieur jusqu'au Santonien. Il monte sa plus grande diversité à l'Aptien basal et au Cénomanien inférieur et moyen.
Coralliaires ; Scléractiniaires ; Crétacé ; taxinomie ; nouveaux taxons.
Wells (1932) was the first to identify the extant coral genus Diploastrea from the Lower Cretaceous, followed by numerous authors (Wells, 1933; Baron-Szabo, 1993, 1997; Morycowa & Masse, 1998; Baron-Szabo & González León, 2003; summarised in Löser et al., 2002). As shown by Löser (2016) this material cannot be assigned to Diploastrea and requires the creation of a new genus, Eosiderastrea, within the Family Siderastraeidae. In addition, it was found that numerous Cretaceous species formerly assigned to genera such as Confusastrea, Diplocoenia, Montastraea, Phyllocoenia, Placocoenia, and Siderastrea belong to this genus. The sum of all this material, mainly from localities in France, Germany, Greece, Italy, Poland, Spain and the USA, makes the genus relatively common and palaeogeographically widely distributed.
The material comes from various localities. Most of them are listed, commented and provided with additional references in Löser et al. (2005). Only details not reported in this publication are mentioned here. If no sample number is given, the material from the locality concerned was not available for study. Each number refers to only one specimen.
Austria
• Oberösterreich, Gosau basin (A.399); Gosau Gp;
Coniacian to Santonian. NHMW 1864/0040/1352.
• Oberösterreich, Gosau, Wegscheidgraben (A.576);
Hochmoos Fm; late Santonian. GPSL FLX 8059.
• Salzburg, Rußbach, Neffgraben (A.575); Hochmoos Fm; late Santonian. GPSL FLX 3126, FLX 5385.
• Salzburg, Rußbach, Zimmergraben (A.969); Gosau Gp,
Streideck Fm; Santonian. MHE A0601.
France
• Ardèche, St.Remèze, Pont de Laval; late
Barremian. BSPG 2003 XX 5908. - The locality and coral fauna was described by Löser & Ferry
(2006).
• Aude, Les Corbières, Col de Escudiés; latest
Cenomanian, Juddi Zone. UPS HL 025. - The stratigraphy and outcrop situation is
described in Bilotte
(2007); the coral fauna is under preparation for its publication.
• Aude, Les Corbières, Sougraigne, Prat-Périé (F.275); uppermost Cenomanian, Juddi
Zone. BSPG 2011 XXVI 46.
• Bouches-du-Rhône, Marseille, Calanque de la Mounine; early Hauterivian. ERNO L-5185, L-5186, L-5193. - The locality and coral
fauna was described by Löser & Ferry
(2006).
• Charente-Maritime, Fouras (F.429); early
Cenomanian. ERNO L-5601, ERNO L-5614.
• Charente-Maritime, Ile d'Aix (F.52); early
Cenomanian. MNHN A26591.
• Sarthe, Le Mans (F.283); Sable du Perche Fm; middle to late Cenomanian. MHNLM 2003.1.7593, 7593b.
• Vaucluse, Ventoux Mts, combe des Glacières (F.3167); early Aptian. UJ 137P3608.
• Yonne, Chenay (F.446); Calcaire à Spatangues; early Hauterivian. MNHN A25815.
• Yonne, Gy-l'Evêque, fields SW Gy-l'Evêque, 3 (F.2732);
Calcaire à Spatangues; early Hauterivian. BSPG 2003 XX 6455, 6458,
6486, 6539, 6602, 6603.
Germany
• Bayern, Allgäuer Helvetikum, Falkenberg (D.2295);
Schrattenkalk; late Barremian to early Aptian. BSPG 1994 XI
292, 53; BSPG 1997 V 113-2.
• Bayern, Allgäuer Helvetikum, NE Hoher Ifen, Kühberg;
Schrattenkalk; late Barremian to early Aptian. BSPG 1996 VI
2. - The outcrop area belongs to the same unit described by Scholz
(1984).
• Bayern, Allgäuer Helvetikum, Tiefenbach (D.256);
Schrattenkalk; late Barremian to early Aptian. BSPG 1994 XI
13, 293, 294.
• Bayern, Roßstein-Almen; Branderfleck Fm; middle
Cenomanian. BSPG 1947 XVI 47; BSPG 1991 X 97. - The geology, stratigraphy and
coral fauna was described by Löser
et al. (2013).
• Nordrhein/Westfalen, Mülheim/Ruhr, Kassenberg (D.510);
Rotkalke; early Cenomanian. BSPG 2003 XX 1086, 1147.
• Sachsen, Meißen-Zscheila, Trinitatis church (D.697);
Meissen Fm; early Cenomanian. SNSD-MMG SaKL253, SaKL542; BSPG 2003 XX
0955.
Greece
• Fokída, Mariolada, Kria Vrissi spring, east of
(GR.975); early Cenomanian. ERNO L-5992.
• Kozani, Kozani, Nea Nikopolis, c4; early
Cenomanian. BSPG 2003 XX 5812, 5829, 5846, 5868, 5900, 6145.
Iran
• Esfahan, Esfahan Basin, Dizlu; late Aptian to Albian. PIUEN b45. - Geology, stratigraphy and the coral fauna was described by Baron-Szabo et al. (2003).
Israel
• Palestine, Mulergat Well; Lower Cretaceous. NHM R43455. - No data are available about this outcrop since it deals with an isolated collection sample.
Italy
• Abruzzi, L'Aquila, Monti d'Ocre, Fossa Cerasetti
(I.1734); early Aptian. PU 17965, 17993, 18099.
• Abruzzi, L'Aquila, Monti d'Ocre, Fossa Mezza Spada
(I.1732); early Aptian. PU 17963, 17964; BSPG 2003 XX 6834.
Japan
• Iwate-ken, Shimohei-gun, Iwaizumi-cho, Moshi,
Matsushima (J.154); Miyako Gr, Hiraiga Fm, Hiraiga sandstone; uppermost
Aptian. TUM 65480.
• Iwate-ken, Shimohei-gun, Tanohata-mura, Haipe,
southern cliff; Miyako Gr, Hiraiga Fm, Hiraiga sandstone; uppermost
Aptian. TUM 65317.
• Iwate-ken, Shimohei-gun, Tanohata-mura, Koikorobe
(J.3326); Miyako Gr, Tanohata Fm, Moshi sandstone; late Aptian. TUM
65367, 65481.
Mexico
• Baja California, Eréndira, N Playa Blanca; Alisitos
Fm; early Albian. ERNO L-134915. - The section was described by Payne
et al. (2004). The coral fauna is
under preparation for publication.
• Oaxaca, Tepelmeme, El Rodeo Ranch (MEX.115);
Teposcolula Fm; early Albian. IGM 2734.
• Puebla, Tehuacán, San Antonio Texcala (MEX.764);
Zapotitlán; Barremian. GPSL FLX 2050.
• Sonora, Municipio Opodepe, Tuape, Cerro de la Espina
(MEX.3202); Mural Limestone, Cerro La Espina Mbr; early Albian. ERNO
3155.
• Sonora, Municipio San Pedro de la Cueva, Tepache,
Lampazos area, Espinazo de Diablo (MEX.3025); Espinazo del Diablo Fm; middle
Albian. ERNO L-130206, L-130135.
Poland
• Malopolskie, Wadowice, Lanckorona, Jastrzebia (PL.1170); Grodziszcze Sandstones; early Aptian. ERNO L-5428.
Spain
• Cantabria, Santander, Cobreces, Luaña playa (E.2088);
Altamira Fm; early Cenomanian. BSPG 2007 V 061, 199, 309, 329.
• Cataluña, Lérida, Com. Alt Urgell, Mun. Coll de Nargó,
Set Comelles, El Caso section (E.2040); Font Bordonera Fm; uppermost
Aptian. BSPG 2003 XX 4027.
• Cataluña, Lérida, Com. La Noguera, Mun. Vilanova de
Meià, Montsec de Rubies, section NW La Cabrua quarry (E.1766); early late Aptian. ERNO L-7368.
• Cataluña, Tarragona, Com. Baix Penedés, Mun. Olivella,
Can Grau (E.834); Montmell Fm; earliest Albian. MV 13539.
• Murcia, Sierra Larga; late Valanginian. ERNO
L-121509. - The Sierra Larga with sediments of the almost entire Cretaceous in
the Prebetic zone encompasses various coral faunas that are partly under
investigation. See Vilas et al.
(2003) for details on the geology.
• Valencia, Castellón, La Avellà, Catí; Hauterivian.
FGUB AV-155-B4. - The sedimentology and coral fauna was described by Götz
et al. (2005).
USA
• Texas, Bell County, Santa Fe Railroad quarry
(USA.1266); Fredericksburg Gp, Comanche Peak Limestone; middle Albian. NMNH
I-313352.
• Texas, Blanco and Hays Counties, Blanco River, narrows
(USA.1211); Trinity Gp, Lower Glen Rose Limestone; earliest Albian.
TMM UT-11482.
• Texas, Buda Hill; early Cenomanian. TMM
UT-10642.
• Texas, Burnet County, Cox's Crossing, Austin-Marble
Falls road (USA.1218); Trinity Gp, Cow Creek Limestone; early late Aptian. TMM UT-11474.
• Texas, Dawson County, 5 mi. S O'donnell, John Earle
ranch, Lone Star Materials Crushing Plant; Fredericksburg Gp; middle Albian.
TMM 1449TX6. - The outcrop is not formally described since it is only known
from collection material.
• Texas, Kendall County, Guadalupe River Ranch; Trinity
Gp, Lower Glen Rose Limestone; early Albian. ERNO L-4789. - The
outcrop area is not yet described.
• Texas, Kerr County, Kerrville, Hiram Hall Ranch
(USA.1252); Fredericksburg Gp, Lower Segovia Limestone; middle Albian. NMNH
I-75128.
• Texas, Kerr County, SE part; Fredericksburg Gp, Edwards
Limestone; middle Albian. TMM 1448TX1. - There are no details known about the
exact spot.
• Texas, Pecos County, Leon springs (USA.1276);
Fredericksburg Gp, Fort Terrell Limestone; middle Albian. TMM UT-14180B.
• Texas, Travis County, Travis Peak, Cow Creek
(USA.1217); Trinity Gp, Cow Creek Limestone; early late
Aptian. NMNH I-75126.
Ukraine
• Sakarpatskaya, Lushanka river basin (UA.1003); Kamenelina Fm; late Barremian to early Aptian.
Corals were cut and polished and thin sections were prepared where possible. Microphotographs for measurements and illustrative purposes were obtained using a transparency scanner, Epson Perfection V750-M Pro, with an optical resolution of 6400 dots per inch. Their quality was improved using histogram contrast manipulation (contrast stretching) where possible. The measurements and septal counts in the descriptions are based on systematic measurements of the calicular dimensions and counts of the septa, carried out using the computer program PaleoTax/Measure (http://www.paleotax.de/measure). The first interval for septal counts is only provided when the number of counts is higher than nine.
Species separation was carried out using distance analysis of metric data. The unweighted euclidean distance was applied as the data were very homogenous (Mucha, 1992). Data visualisation was done through agglomerative cluster analysis using the single-linkage method (Shi, 1993; Löser, 2002). The computer program PaleoTax/Graph was applied for data processing and visualisation. Because the morphological data are not normally distributed (Löser, 2012), an analysis of variance (ANOVA) would not allow distinguishing specimens or taxa. Non-parametric statistical methods would demand more data for each single coral colony that are simply not available.
The synonymy lists encompass - with a few exceptions - only samples that were available for observation. The distribution data are almost entirely based on well-examined material. Material only mentioned in the literature and material not available or insufficiently described and illustrated in the literature has not been taken into account. To obtain better insight into the distribution patterns of the genus, numerous yet-unpublished material - indicated under 'Material' by a collection acronym and sample number in parenthesis - has been included. Therefore distribution data indicated under 'Occurrence' are also provided for species remaining in open nomenclature.
Institutional abbreviations: BSPG, Bayerische Staatssammlung für Paläontologie und Geologie, München, Germany; ERNO, Universidad Nacional Autónoma de México, Instituto de Geología, Estación Regional de Noroeste, Hermosillo, Mexico; FGUB,Facultad de Geología de la Universidad de Barcelona, Spain; GPSL, Geologische und Paläontologische Sammlung der Universität Leipzig, Germany; MGSB, Museo Geológico del Seminario de Barcelona, Spain; MHE, Matthias Heinrich, Eckental, Germany; MHNLM, Musée Vert, Le Mans, France; MNHN, Muséum National d'Histoire Naturelle, Paris, France; NHM, The Natural History Museum, London, England; NHMW, Naturhistorisches Museum, Wien, Austria; MV, Vinseum, Vilafrance del Penedés, Spain; NHM, The Natural History Museum, London, UK; NHMW, Naturhistorisches Museum, Wien, Austria; NMNH, National Museum of Natural History, Washington, D.C., USA; PIUEN, Paläontologisches Institut, Erlangen, Germany; PU, Museo di Geologia e Paleontologia dell' Università di Torino, Italy; TMM, Texas Memorial Museum, Austin, Texas, USA; TUM, The Tohoku University Museum, Sendai, Japan; UJ, Jagiellonian University, Instytut Nauk Geologicznych, Kraków, Poland; UPS, Université Paul Sabatier, Laboratoire de Géologie Sédimentaire et Paléontologie, Toulouse, France.
Abbreviations of measurements: Ccd, distance between calicular centres; clmax, larger calicular diameter (calicular pit); clmin, smaller calicular diameter (calicular pit); cmax, larger outer calicular diameter; cmin, smaller outer calicular diameter; s, number of radial elements in adult calices.
For each type of measurement (calicular diameter and distance, number of septa) in one thin section the following values were obtained and/or calculated: n, number of measurements; min-max, lowest and highest measured value; µ: arithmetic mean (average); s, standard deviation; cv, coefficient of variation; µ±s, first interval.
Abbreviations in synonymy lists: The abbreviations used in the synonymy lists follow Matthews (1973): *, earliest valid publication of the species name; p, the described material belongs only in part to the species concerned; v, the specimen was observed by the author.
Order Scleractinia Bourne, 1900
?Suborder Fungiina Verrill, 1870
Remarks: The position of the family within the Suborder Fungiina is provisional. The suborder was, for a long time (Vaughan & Wells, 1943; Alloiteau, 1952; Wells, 1956), considered to gather all corals that have perforated septa and presence of synapticulae. Gill (1981) proposed that Fungiina sensu stricto shows synapticular bars or compound synapticulae (fulturae). Former fungiid corals with pennulae (see Gill, 1967) were later separated into the Suborder Microsolenina (Morycowa & Roniewicz, 1995), and highly perforated non-pennular corals into the Suborder Poritina (Veron, 1995). Presently the Suborder Fungiina includes various families that differ much in their morphology but show synapticulae (not even all have perforated septa). As already mentioned by Morycowa & Roniewicz (1995), the suborder needs profound revision. The Superfamily Thamnasterioidea is characterised by the presence of synapticulae and remains, for the time being, in the Suborder Fungiina, but a profound modification of the whole classification system of the Mesozoic Scleractinia is required to gain more consistency.
Superfamily Thamnasterioidea Reuss, 1864
Family Siderastraeidae Vaughan & Wells, 1943
Remarks: Details on morphology, evolution and genera were provided by Löser (2016).
Genus Eosiderastrea Löser, 2016
Type species: Siderastrea cuyleri Wells, 1932, by original designation.
Diagnosis: Plocoid, sometimes also cerioid or astreoid colonies. Septa made of medium-large trabeculae with rare pores at their inner margin. Septal symmetry subregular, in varying systems. Septal lateral face with thorns, septal upper border granulated. Synapticulae present. Columella styliform or by septal fusion. Endotheca and wall absent. Budding extracalicinal.
Description: Cerioid, astreoid or plocoid colony. Calicular outline circular to polygonal, often slightly elongated. Septa can have pores at the inner margins. Microstructure of medium-sized trabeculae. In cross section septa (and costae) are thick at the wall and thinner towards the centre. Symmetry of septa radial, presenting hexameral, heptameral, octameral, and decameral symmetries. Cycles of septa sub-regular. Septal cycles differ in length. Generally, two septal cycles reach the centre of the calice, further cycles are shorter. Septa of the younger cycles are often attached to those of the older cycles. Septal upper border granulated, lateral face has numerous, irregularly distributed fine thorns, the inner margin is slightly swollen in places. Some septa may be attached to the columella. Costae present, non-confluent to confluent. Synapticulae present, mainly in the wall region. Columella styliform and by septal fusion in the centre of the calice. Endotheca consists of numerous thin tabulae. Wall subcompact or absent, septothecal, with synapticulae. Coenosteum narrow in some species with isolated trabeculae from the external margins of the costae. Budding extracalicinal.
Remarks: The calicular arrangement varies within the genus. Depending on the compactness of the wall and the confluence of the costae, there exist species with a cerioid arrangement (e.g., E. cuyleri), with an astreoid arrangement (e.g., E. paragrandipora) or plocoid arrangement (e.g., E. harrisi). The amount of synapticulae, which defines whether a wall exists or not, and so characterises the organisation form, varies from species to species but is constant within one specimen.
The microstructure of the septa is made of medium-sized trabeculae in the sense that the trabeculae are easily recognisable but do not give shape to the sepum (as for instance in members of the Montlivaltiidae and Synastraeidae). Small trabeculae are not visible or only as a medium dark line. The absolute size of the trabeculae depends on the thickness of septa. The septa are thicker in the Siderastraeidae than, for instance, in the very similar Thamnasteriidae because the structures are generally larger, e.g., the calices are bigger and the septa thicker in the Siderastraeidae and the individuals are smaller and the septa are thinner in the Thamnasteriidae. The trabeculae have, therefore, a differing absolute size but are in both families medium-sized, related to the thickness of septa.
Species separation: The distinction of species in Mesozoic scleractinian corals is almost entirely based on (1) the dimensions of the corallites, the distance of the corallites and the width of calicular rows in meandrinoid corals respectively, (2) the number of regular septal systems (if present) and (3) the number and/or density of the septa (Löser, 2012). The specimens assigned to Eosiderastrea offer a wide range of numerical morphometric data: The larger and smaller inner and outer calicular diameters, the calicular centre distances, septal symmetry, absolute number of septa and the number of septal cycles. To find out which of these characteristics are most suitable for the separation of taxa, the calicular dimensions were systematically measured in all available samples and the septa counted. The highest available number of measurements and counts were obtained. The results (for details refer to the species) show that the most constant character within a colony is septal symmetry, which is the same for all individuals in a colony. Hence, the number of septa also shows a certain variation, as the number of septa increases constantly with increasing calicular diameter. So the septal cycles are not regular. Among the calicular dimensions the distance of calicular centres shows the highest variation. The inner (larger and smaller) calicular diameter can only be measured when a wall is clearly marked, which was not the case for all the specimens. However, the outer calicular diameter (smaller and larger) can be obtained for all specimens. Generally, smaller and larger calicular diameters were distinguished in all colonies. The smaller calicular diameter has a lower variation. It was also found that the absence or presence of even an incomplete wall, which defines whether the colony is astreoid or plocoid, is constant within one colony and was included in the analysis. For species separation, the septal symmetry, the number of septal cycles, the smaller outer calicular diameter, the distance of calicular centres and the absence or presence of a wall were used.
Other differences, such as the fine structure of the corallite wall, the formation of the columella, the coenosteum, or differences in the number and arrangement of synapticulae and dissepiments vary within one colony considerably, and are often difficult to observe and to quantify. The variation of the thickness of the septa is low.
Species: In the present study 26 species were distinguished, six of them remain in open nomenclature as the poor number of available samples does not allow a new species to be established for the moment. For the distinction of the species see Table 1.
Range: Late Valanginian to Santonian.
Table 1: Distinction of species. (1) The stratigraphy of the Iran locality is not well constrained. (2) The occurrence of the species in the Campanian is in doubt. (3) The stratigraphy of the Palestine locality is not well constrained.
| Septa | Smaller outer diameter | Wall | Name | Stratigraphy | |||
| sym. | cycles | min | max | average | |||
| 6 | 3 | 2.3 | 3.2 | 2.8 | absent | cuyleri | late Aptian early Cenomanian |
| 2.5 | 4.1 | 3.3 | present | nagaoi | Hauterivian early Albian | ||
| 3.9 | 5.8 | 4.9 | present | hilli | middle Albian | ||
| 4 | 3.5 | 4.3 | 3.9 | present | sp. 1 | late Valanginian | |
| 4 | 6 | 5.0 | present | dollfusi | Aptian early Albian | ||
| 3.8 | 6.3 | 5.1 | absent | paragrandipora | Cenomanian | ||
| 4.9 | 7 | 6.0 | absent | sp. 2 | early Hauterivian | ||
| 5.6 | 7.2 | 6.4 | absent | grandipora | middle Albian early Cenoman. | ||
| 7 | 9.5 | 8.3 | present | crassa | late Barremian middle Albian | ||
| 7 | 3 | 3.6 | 4 | 3.8 | present | cotteaui | early Hauterivian |
| 4.1 | 4.7 | 4.4 | present | cf. cotteaui | early Hauterivian | ||
| 8 | 3 | 2.7 | 3.4 | 3.1 | present | cyclops | Hauterivian Barremian |
| 3.1 | 4.5 | 3.8 | absent | aff. cuyleri | late Aptian early Cenomanian | ||
| 4 | 4.2 | 4.1 | present | cf. cyclops | Hauterivian | ||
| 4.5 | 6.2 | 5.4 | present | harrisi | Hauterivian early Albian | ||
| 4 | 7.2 | 5.6 | absent | villaltai | late Barremian early Albian (1) | ||
| 4 | 3.3 | 4.4 | 3.9 | absent | sp. 3 | early Cenomanian | |
| 5.5 | 7.3 | 6.4 | present | splendida | early Aptian | ||
| 10 | 3 | 1.4 | 3.2 | 2.3 | present | vaughani | middle Albian (2) |
| 3.5 | 5.4 | 4.5 | present | felixi | Aptian | ||
| 3.5 | 6.3 | 4.9 | absent | stefani | Cenomanian (3) | ||
| 5 | 7.9 | 6.5 | present | crassicosta | Hauterivian early Aptian | ||
| 5.7 | 8.2 | 7.0 | absent | sp. 4 | latest Aptian Cenomanian | ||
| 4 | 4 | 5.1 | 4.6 | absent | sp. 5 | early Cenomanian | |
| 6.2 | 8.2 | 7.2 | absent | sp. 6 | early middle Cenomanian | ||
| 7.4 | 10.4 | 8.9 | absent | reussi | Santonian | ||
Eosiderastrea cotteaui (Orbigny, 1850)
(Pl. 1 ![]() , fig. 1)
, fig. 1)
Material: MNHN A25815; 1 thin section.
Synonymy:
|
*v |
Phyllocoenia Cottaldina - Orbigny, (2), p. 91 |
Dimensions:
| (MNHN A25815) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 7 | 3.58-4.06 | 3.78 | 0.16 | 4.3 | 3.62-3.95 |
| ccd | 7 | 4.26-6.17 | 5.48 | 0.62 | 11.3 | 4.86-6.10 |
| s | 5 | 29-31 | 30.0 | 0.70 | 2.3 | |
Remarks: The material provides a very rare case of heptameral symmetry. The calices are in a plocoid arrangement. The costae show all types of confluence. The septa appear in three septal cycles. A wall is present but incomplete. The material encompasses three syntypes that seem to be conspecific. There is one thin section that is poorly preserved.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Chenay.
Eosiderastrea cf. cotteaui (Orbigny, 1850)
(Pl. 1 ![]() ,
figs. 2-3)
,
figs. 2-3)
Material: BSPG 2003 XX 6458; 1 thin section.
Synonymy:
|
v |
Placocoenia cotteaui (Orbigny, 1850) - Löser, p. 44, Pl. 2, fig. 5 |
Dimensions:
| (BSPG 2003 XX 6458) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 2.31-3.59 | 3.02 | 0.44 | 14.5 | 2.58-3.46 |
| cmin | 5 | 3.99-4.75 | 4.41 | 0.27 | 6.2 | 4.14-4.68 |
| ccd | 14 | 2.52-4.92 | 3.65 | 0.66 | 18.1 | 2.99-4.31 |
| s | 6 | 24-30 | 27.3 | 2.1 | 7.5 | |
Remarks: The only specimen has smaller dimensions than E. cotteaui.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque.
Eosiderastrea crassa (Kuzmicheva, 1980)
(Pl. 1 ![]() ,
figs. 4-5)
,
figs. 4-5)
Material: TMM 1449TX6; 6 thin sections.
Synonymy:
|
Diploastraea crassa Kuzmicheva, p. 105, Pl. 39, fig. 2 |
Dimensions:
| (TMM 1449TX6) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 3 | 7.42-8.02 | 7.64 | 0.33 | 4.3 | 7.30-7.97 |
| cmin | 3 | 9.07-9.48 | 9.28 | 0.20 | 2.2 | 9.07-9.48 |
| ccd | 10 | 6.14-12.52 | 9.12 | 2.01 | 22.0 | 7.11-11.14 |
| s | 3 | 43-48 | 46.33 | 2.88 | 6.2 | |
Remarks: Based on the illustrations of Kuzmicheva (1980), the species seems to be mainly astreoid with confluent or sub-confluent septa and an incomplete wall. The symmetry seems to be hexameral with four cycles. The material from Texas is not well preserved and the thin section is very small, which makes systematic measurement almost impossible. In terms of its dimensions it is slightly larger than the material described by Kuzmicheva.
Occurrence: Upper Barremian to lower Aptian of Ukraine (Sakarpatskaya) Lushanka river basin; middle Albian of USA (Texas) Dawson County, 5 mi. S O'donnell, John Earle ranch, Lone Star Materials crushing Plant.
Eosiderastrea crassicostata (Morycowa & Masse, 1998)
(Pl. 1 ![]() ,
fig. 6)
,
fig. 6)
Material: BSPG 2003 XX 6603, ERNO L-5428, UJ 137P3608; 5 thin sections.
Synonymy:
|
v |
?Diploastrea crassicostata Morycowa & Masse, p. 755, Fig. 19.1-2 |
Dimensions:
| (BSPG 2003 XX 4027) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 10 | 6.20-8.06 | 7.00 | 0.55 | 7.9 | 6.44-7.56 |
| ccd | 7 | 5.84-9.62 | 7.72 | 1.13 | 14.6 | 6.58-8.85 |
| s | 6 | 39-47 | 43.66 | 2.8 | 6.4 | |
| (BSPG 2003 XX 5428) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 9 | 4.39-5.72 | 4.94 | 0.47 | 9.5 | 4.47-5.41 |
| cmin | 7 | 5.19-8.00 | 6.81 | 1.06 | 15.6 | 5.75-7.88 |
| ccd | 10 | 6.67-9.74 | 8.27 | 0.83 | 10.1 | 7.43-9.10 |
| s | 3 | 41-44 | 42.33 | 1.52 | 3.6 | |
| (BSPG 2003 XX 5846) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 20 | 3.80-5.55 | 4.77 | 0.44 | 9.3 | 4.32-5.21 |
| cmin | 15 | 5.91-8.04 | 6.96 | 0.64 | 9.2 | 6.32-7.61 |
| ccd | 20 | 5.11-7.71 | 6.55 | 0.85 | 12.9 | 5.70-7.40 |
| s | 12 | 39-47 | 41.83 | 2.4 | 5.7 | 39-44 |
| (UJ 137P3608) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 5 | 5.76-6.87 | 6.27 | 0.52 | 8.4 | 5.74-6.79 |
| cmax | 3 | 7.04-7.92 | 7.40 | 0.46 | 6.2 | 6.94-7.86 |
| ccd | 10 | 4.96-8.74 | 6.94 | 1.22 | 17.6 | 5.71-8.16 |
| s | 5 | 39-43 | 41.0 | 1.58 | 3.8 | |
Remarks: The species shows an astreoid calicular arrangement with mainly non-confluent costae. The septa show decameral symmetry and three cycles. A wall does not exist. The species is well documented; therefore, it is not necessary to reiterate the type material.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque; lower Aptian of France (Vaucluse) Ventoux Mts, combe des Glacières; Poland (Malopolskie, Wadowice) Lanckorona, Jastrzebia.
Eosiderastrea cuyleri (Wells, 1932)
(Pl. 1 ![]() ,
figs. 7-9)
,
figs. 7-9)
Material: ERNO L-5992, TMM UT-11474; 3 thin sections.
Synonymy:
|
*v |
Siderastraea cuyleri Wells, p. 243, Pl. 37, fig. 9; Pl. 39, fig. 1 |
|
|
v |
Eoiderastraea cuyleri (Wells, 1932) - Löser |
Dimensions:
| (TMM UT-11474) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 30 | 2.35-3.41 | 2.87 | 0.30 | 10.6 | 2.56-3.17 |
| clmax | 30 | 3.05-4.29 | 3.50 | 0.35 | 10.0 | 3.15-3.86 |
| ccd | 40 | 2.46-3.92 | 3.12 | 0.35 | 11.4 | 2.77-3.48 |
| s | 20 | 24-36 | 30.25 | 4.25 | 14 | 26-35 |
| (BSPG 2003 XX 5992) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 2.34-2.90 | 2.52 | 0.20 | 8.1 | 2.32-2.73 |
| cmin | 10 | 2.07-2.76 | 2.50 | 0.21 | 8.6 | 2.29-2.72 |
| ccd | 20 | 1.93-3.45 | 2.74 | 0.39 | 14.3 | 2.34-3.13 |
| s | 5 | 24-25 | 24.4 | 0.54 | 2.2 | |
Remarks: The astreoid species has very small dimensions. The septal symmetry is hexameral with three complete cycles and the beginnings of a fourth one. A wall is practically absent, and the costae are mainly non-confluent.
Occurrence: Lower upper Aptian of USA (Texas) Burnet County, Cox's Crossing, Austin-Marble Falls road; lower Cenomanian of Greece (Fokída) Mariolada, Kria Vrissi spring.
Eosiderastrea aff. cuyleri (Wells, 1932)
(Pl. 1 ![]() ,
fig. 10)
,
fig. 10)
Material: ERNO L-134915, NMNH I-75126, TMM UT-10642; 3 thin sections.
Synonymy:
|
vp |
Siderastraea cuyleri Wells, p. 243, Pl. 37, fig. 9; Pl. 39, fig. 1 |
Dimensions:
| (TMM UT-10642) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 20 | 2.66-4.53 | 3.59 | 0.46 | 12.9 | 3.13-4.06 |
| cmax | 15 | 3.04-5.00 | 3.97 | 0.54 | 13.6 | 3.42-4.51 |
| ccd | 20 | 2.66-4.72 | 3.59 | 0.57 | 15.9 | 3.01-4.16 |
| s | 15 | 31-43 | 36.86 | 3.66 | 9.9 | 33-41 |
Remarks: This species encompasses the paratype of E. cuyleri, which shows a different septal symmetry and has larger dimensions than the holotype. The material is clearly astreoid; a wall is absent. The symmetry is octameral with three cycles and the beginnings of a fourth.
Occurrence: Lower upper Aptian of USA (Texas) Travis County, Travis Peak, Cow Creek; lower Albian of Mexico (Baja California) Eréndira, N Playa Blanca; lower Cenomanian of USA (Texas) Buda Hill.
Eosiderastrea cyclops (Felix, 1891)
(Pl. 3 ![]() ,
fig. 3)
,
fig. 3)
Material: BSPG 2003 XX 6455, GPSL FLX 2050; 1 thin section.
Synonymy:
|
*v |
Phyllocoenia cyclops Felix, p. 155, Pl. 23, figs. 3, 3.a, 5 |
|
|
v |
?Diploastrea cyclops (Felix, 1891) - Löser, p. 30, fig. 2.I |
Dimensions:
| (GPSL FLX 2050) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 20 | 2.56-3.51 | 3.05 | 0.30 | 10.1 | 2.74-3.36 |
| ccd | 20 | 2.83-4.84 | 3.89 | 0.63 | 16.3 | 3.25-4.52 |
| s | ca. 32 | |||||
| (BSPG 2003 XX 6602) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 6 | 3.98-4.19 | 4.08 | 0.07 | 1.9 | 4.00-4.16 |
| cmin | 6 | 5.22-5.92 | 5.72 | 0.25 | 4.4 | 5.47-5.98 |
| ccd | 9 | 4.04-7.10 | 5.98 | 0.83 | 13.8 | 5.15-6.81 |
| s | 3 | 31-34 | ||||
Remarks: The lectotype of the species is poorly preserved and does not allow for the preparation of thin sections (see Löser, 2006, for details). Therefore, the species is not well defined. The Hauterivian material shows a more plocoid calicular arrangement with sub-confluent costae. The wall is incomplete. The septal symmetry is octameral.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque. Barremian of Mexico (Puebla) Tehuacán, San Antonio Texcala.
Eosiderastrea cf. cyclops (Felix, 1891)
(Pl. 1 ![]() ,
figs. 11-12)
,
figs. 11-12)
Material: BSPG 2003 XX 6602; 1 thin section.
Dimensions:
| (BSPG 2003 XX 6602) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 6 | 3.98-4.19 | 4.08 | 0.07 | 1.9 | 4.00-4.16 |
| cmin | 6 | 5.22-5.92 | 5.72 | 0.25 | 4.4 | 5.47-5.98 |
| ccd | 9 | 4.04-7.10 | 5.98 | 0.83 | 13.8 | 5.15-6.81 |
| s | 3 | 31-34 | ||||
Remarks: The material has larger dimensions than E. cyclops. The calicular arrangement is plocoid, the costae show varying stages of confluence and the septal symmetry is octameral.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque.
Eosiderastrea dollfusi (Prever, 1909)
(Pl. 2 ![]() ,
fig. 6)
,
fig. 6)
Material: IGM 2734, PU 17963, 17964.
Synonymy:
|
*v |
Confusastraea Dollfusi Prever, p. 89, Pl. 5, figs. 2, 2.a & 3 |
|
|
v |
Diploastrea harrisi Wells, 1932 - Reyeros de Castillo, p. 27, Pl. 18, fig. 1 |
Dimensions:
| (PU 17964) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 3.71-4.97 | 4.13 | 0.39 | 9.5 | 3.73-4.53 |
| cmin | 8 | 4.24-5.37 | 4.85 | 0.45 | 9.3 | 4.39-5.30 |
| ccd | 15 | 3.90-5.39 | 4.56 | 0.47 | 10.4 | 4.09-5.04 |
| s | 4 | 42-50 | 46.5 | 3.69 | 7.9 | |
Remarks: PU 17964 is here designated as the lectotype. PU 17963 becomes the paralectotype. The species forms astreoid colonies with sub or non-confluent septa. The septa appear in six systems and four septal cycles, which are often not quite complete. A wall does not exist. The lectotype of the species is small and no thin sections or topotypical material exists.
Occurrence: Lower Aptian of Italy (Abruzzi, L'Aquila) Monti d'Ocre, Fossa Mezza Spada; lower Albian of Mexico (Oaxaca) Tepelmeme, El Rodeo Ranch.
Eosiderastrea felixi (Prever, 1909)
(Pl. 2 ![]() ,
figs. 1-5)
,
figs. 1-5)
Material: BSPG 2003 XX 6834, PU 17965, TUM 65481; 4 thin sections.
Synonymy:
|
*v |
Confusastraea Felixi Prever, p. 88, Pl. 5, fig. 1 |
|
|
v |
Placocoenia tanohataensis Eguchi, p. 26, Pl. 3, figs. 6-8 |
Dimensions:
| (PU 17965) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 7 | 3.65-5.07 | 4.27 | 0.46 | 10.9 | 3.81-4.74 |
| cmax | 5 | 4.63-6.42 | 5.18 | 0.74 | 14.3 | 4.44-5.92 |
| ccd | 6 | 4.20-5.85 | 5.05 | 0.52 | 10.4 | 4.52-5.57 |
| s | 38-43 | |||||
| (BSPG 2003 XX 6834) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 25 | 2.07-3.25 | 2.71 | 0.37 | 13.6 | 2.34-3.08 |
| cmax | 20 | 3.69-5.86 | 4.59 | 0.57 | 12.4 | 4.02-5.16 |
| cmin | 20 | 3.16-4.71 | 3.95 | 0.42 | 10.7 | 3.53-4.38 |
| ccd | 20 | 3.42-5.57 | 4.45 | 0.76 | 17.0 | 3.69-5.21 |
| s | 6 | 38-44 | 41.16 | 2.56 | 6.2 | 39-44 |
| (TUM 65481) 4.5-5.4 | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 20 | 3.39-4.33 | 3.78 | 0.29 | 7.7 | 3.48-4.07 |
| cmin | 20 | 4.14-5.76 | 4.93 | 0.44 | 9.0 | 4.49-5.38 |
| ccd | 20 | 4.01-5.71 | 4.95 | 0.48 | 9.8 | 4.47-5.44 |
| s | 15 | 34-43 | 38.9 | 2.7 | 6.9 | 36-42 |
Remarks: The type specimen is too small to provide significant morphometric values. The topotypic specimen, BSPG 2003 XX 6834, has slightly smaller dimensions.
Occurrence: Lower Albian of Italy (Abruzzi, L'Aquila) Monti d'Ocre, Fossa Cerasetti; Fossa Mezza Spada; upper Aptian of Japan (Iwate-ken) Shimohei-gun, Tanohata-mura, Koikorobe.
Eosiderastrea grandipora (Orbigny, 1850)
(Pl. 3 ![]() ,
figs. 1-2)
,
figs. 1-2)
Material: BSPG 2003 XX 1086, 5900, BSPG 2007 V 199, MNHN A26591, NMNH I-313352; 4 thin sections.
Synonymy:
|
*v |
Stephanocoenia grandipora Orbigny, (2), p. 182 |
|
|
vp |
Favoidioseris fredericksburgensis Wells, p. 98, Pl. 8, figs. 15-18 |
|
|
vp |
Baryphyllia haimei Fromentel, 1857 - Löser, p. 38, Figs. 26-27; Pl. 8, fig. 1; Pl. 12, figs. 10-11 |
Dimensions:
| (MNHN A26591) | |
| cmin | 6-7 |
| ccd | 7.5-8 |
| s | 48-52 |
| (BSPG 2003 XX 5900) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 14 | 5.77-7.37 | 6.67 | 0.54 | 8.1 | 6.13-7.22 |
| ccd | 20 | 6.23-9.27 | 7.62 | 0.92 | 12.0 | 6.70-8.55 |
| s | 5 | 50-61 | 53.8 | 4.26 | 7.9 | |
Remarks: The astreoid species has large dimensions. The septal symmetry is hexameral with four complete cycles and the beginnings of a fifth one. A wall is practically absent, and the costae are mainly non-confluent.
Occurrence: Middle Albian (Lautus Zone) of USA (Texas) Bell County, Santa Fe Railroad quarry; lower Cenomanian (Mantelli Zone) of Germany (Nordrhein/Westfalen) Mülheim/Ruhr, Kassenberg; lower Cenomanian of France (Charente-Maritime) Ile d'Aix; Greece (Kozani) Kozani, Nea Nikopolis; lower Cenomanian (Dixoni Zone) of Spain (Cantabria, Santander) Cobreces, Luaña playa.
Eosiderastrea harrisi (Wells, 1932)
(Pl. 3 ![]() ,
figs. 4-9)
,
figs. 4-9)
Material: BSPG 2003 XX 6486, FGUB AV-155-B4, PU 18099, TMM UT-11482, TUM 65367, 65480; 12 thin sections.
Synonymy:
|
v |
Orbicella Simonyi - Prever, p. 87, Pl. 4, fig. 6 |
|
|
*v |
Diploastrea harrisi Wells, p. 248, Pl. 30, fig. 9; Pl. 37, fig. 6; Pl. 38, figs. 5-6 |
|
|
v |
Placocoenia orbitoides Eguchi, p. 26, Pl. 3, figs. 3-5 |
|
|
v |
(no name) - Budd & Coates, fig. 2.B |
Dimensions:
| (PU 18099) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 14 | 3.32-4.03 | 3.65 | 0.22 | 6.0 | 3.43-3.87 |
| cmin | 10 | 5.01-5.88 | 5.54 | 0.27 | 5.0 | 5.26-5.82 |
| ccd | 20 | 4.56-6.02 | 5.39 | 0.40 | 7.4 | 4.98-5.79 |
| s | 5 | 39-44 | 41.2 | 2.16 | 5.2 | |
| (TMM UT-11482) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 4.20-4.91 | 4.58 | 0.24 | 5.4 | 4.33-4.83 |
| clmax | 10 | 3.84-4.85 | 4.52 | 0.35 | 7.8 | 4.16-4.87 |
| cmin | 10 | 4.92-6.49 | 5.54 | 0.49 | 8.8 | 5.04-6.03 |
| cmax | 10 | 5.42-6.56 | 6.14 | 0.43 | 7.0 | 5.71-6.58 |
| ccd | 20 | 4.48-6.75 | 5.58 | 0.64 | 11.5 | 4.94-6.23 |
| s | 7 | 32-36 | 34.4 | 1.6 | 4.7 | |
| (TUM 65367) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 20 | 3.53-4.53 | 4.03 | 0.26 | 6.5 | 3.76-4.29 |
| cmin | 20 | 4.76-6.03 | 5.34 | 0.36 | 6.8 | 4.98-5.71 |
| ccd | 20 | 4.31-6.93 | 5.35 | 0.65 | 12.2 | 4.69-6.00 |
| s | 10 | 29-34 | 30.6 | 1.42 | 4.6 | 29-32 |
| (BSPG 2003 XX 6486) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 7 | 3.30-4.12 | 3.74 | 0.34 | 9.1 | 3.40-4.08 |
| cmin | 5 | 4.42-5.39 | 4.83 | 0.36 | 7.5 | 4.47-5.20 |
| ccd | 10 | 3.24-5.74 | 4.36 | 0.82 | 18.9 | 3.53-5.19 |
| s | 32 | |||||
Remarks: E. harrisi is an astreoid to plocoid species with a more or less compact wall. The septa are generally sub to non-confluent. They appear in eight systems and three cycles. The material shows some variation in the number of septa. Some samples have a larger or smaller number of septa compared to the type material of the type specimen. The Hauterivian material (BSPG 2003 XX 6486; FGUB AV 155 B 4) has slightly smaller dimensions.
Occurrence: Hauterivian of Spain (Valencia, Castellón) La Avellà, Catí; lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque; lower Aptian of Italy (Abruzzi, L'Aquila) Monti d'Ocre, Fossa Cerasetti; upper Aptian of Japan (Iwate-ken) Shimohei-gun, Tanohata-mura, Koikorobe; uppermost Aptian of Japan (Iwate-ken) Shimohei-gun, Iwaizumi-cho, Moshi, Matsushima; lowermost Albian (Tardefurcata Zone) of USA (Texas) Blanco and Hays Counties, Blanco River narrows.
Eosiderastrea hilli (Wells, 1933)
(Pl. 3 ![]() ,
figs. 10-12)
,
figs. 10-12)
Material: TMM 1448TX1, UT-14180B; 4 thin sections.
Synonymy:
|
*v |
Diploastrea hilli Wells, p. 93, Pl. 2, figs. 13-14; Pl. 8, fig. 12 |
Dimensions:
| (TMM UT-14180) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 8 | 2.65-3.35 | 3.00 | 0.24 | 8.1 | 2.76-3.25 |
| cmin | 8 | 3.78-5.10 | 4.41 | 0.47 | 10.6 | 3.94-4.88 |
| ccd | 9 | 3.67-5.24 | 4.47 | 0.52 | 11.8 | 3.94-5.00 |
| s | 3 | 24-26 | 24.7 | 1.15 | 4.6 | |
| (TMM 1448TX1) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 3.32-4.49 | 3.84 | 0.38 | 10.0 | 3.45-4.23 |
| cmin | 5 | 4.56-6.09 | 5.26 | 0.58 | 11.1 | 4.67-5.85 |
| ccd | 8 | 4.53-6.98 | 5.55 | 0.79 | 14.2 | 4.76-6.34 |
| s | 24 | |||||
Remarks: The holotype of E. hilli is a small specimen that almost does not allow for the measurement of significant morphometric values. The species has six septal system with three septal cycles. It is clearly plocoid, occasionally with confluent costae. A wall is present. Only the type specimen is known. Specimen TMM 1448 TX1 is very similar to the holotype of E. hilli but clearly has larger dimensions. It is, as with E. hilli, a plocoid form with confluent to sub-confluent septa. A wall is present. The septa show hexameral symmetry in three cycles.
Occurrence: Middle Albian of USA (Texas) Kerr County, SE part; middle Albian (Lautus Zone) of USA (Texas) Pecos County, Leon springs.
Eosiderastrea nagaoi (Eguchi, 1951)
(Pl. 4 ![]() ,
figs. 1-6)
,
figs. 1-6)
Material: BSPG 2003 XX 5908, ERNO 3155, L-5185, L-5186, L-5193, TUM 65317; 12 thin sections.
Synonymy:
|
*v |
Montastrea nagaoi Eguchi, p. 25, Pl. 3, figs. 1-2 |
|
|
v |
Diploastrea harrisi Wells, 1932 - Baron-Szabo & González León, p. 212, Fig. 8.B & 8.E |
|
|
v |
Kobyastraea sp. - Löser & Ferry, p. 489, Fig. 5.3 |
Dimensions:
| (TUM 65317) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 30 | 1.62-2.19 | 1.90 | 0.17 | 8.9 | 1.73-2.07 |
| clmax | 30 | 1.72-2.64 | 2.33 | 0.19 | 8.2 | 2.13-2.52 |
| cmin | 25 | 2.86-4.02 | 3.28 | 0.31 | 9.7 | 2.96-3.60 |
| cmax | 25 | 3.10-4.22 | 3.66 | 0.31 | 8.6 | 3.35-3.98 |
| ccd | 30 | 2.39-3.62 | 3.01 | 0.33 | 11.1 | 2.68-3.34 |
| s | 10 | 30-40 | 33.9 | 3.0 | 8.9 | 31-37 |
| (ERNO 3155) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 25 | 3.02-4.49 | 3.75 | 0.35 | 9.5 | 3.39-4.10 |
| ccd | 25 | 3.16-4.25 | 3.70 | 0.28 | 7.6 | 3.42-3.99 |
| s | 10 | 23-30 | 26.1 | 2.28 | 8.7 | 24-28 |
| (BSPG 2003 XX 5186) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 15 | 2.26-3.20 | 2.78 | 0.28 | 10.0 | 2.50-3.06 |
| cmin | 10 | 3.21-4.10 | 3.60 | 0.23 | 6.4 | 3.37-3.84 |
| ccd | 20 | 2.65-4.13 | 3.39 | 0.44 | 12.9 | 2.95-3.83 |
| s | 10 | 25-30 | 28.4 | 2.0 | 7.0 | 27-30 |
| (BSPG 2003 XX 5193) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 20 | 2.72-3.70 | 3.12 | 0.27 | 8.9 | 2.84-3.40 |
| ccd | 30 | 2.47-3.89 | 3.31 | 0.38 | 11.4 | 2.93-3.69 |
| s | 20 | 26-34 | 28.15 | 1.87 | 6.6 | 26-30 |
| (BSPG 2003 XX 5908) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 20 | 2.44-3.49 | 2.91 | 0.36 | 12.4 | 2.55-3.27 |
| ccd | 25 | 2.17-3.47 | 3.03 | 0.36 | 11.9 | 2.66-3.39 |
| s | 8 | 28-34 | 30.5 | 2.0 | 6.5 | |
Remarks: The species has a plocoid calicular arrangement. The costae are sub or non-confluent. The wall is nearly complete. The septa appear in six systems and three cycles with the beginnings of a fourth cycle.
Occurrence: Lower Hauterivian of France (Bouches-du-Rhône) Marseille, Calanque de la Mounine; upper Barremian of France (Ardèche) St.Remèze, Pont de Laval; uppermost Aptian of Japan (Iwate-ken) Shimohei-gun, Tanohata-mura, Haipe, southern cliff; lower Albian of Mexico (Sonora) Municipio Opodepe, Tuape, Cerro de la Espina.
Eosiderastrea paragrandipora n. sp.
(Pl. 4 ![]() ,
figs. 7-9)
,
figs. 7-9)
Synonymy:
|
vp |
Baryphyllia haimei Fromentel, 1857 - Löser, p. 38, Figs. 26-27; Pl. 8, fig. 1; Pl. 12, figs. 10-11 |
Origin of the name: In relation to the very similar species E. grandipora.
Holotype: BSPG 2007 V 309.
Paratype: BSPG 2007 V 329.
Material studied: Holotype, paratype, four further specimens (BSPG 2003 XX 1147, 5812, 5868, BSPG 2011 XXVI 46) from different localities.
Type locality: Spain, Cantabria, Santander, Cobreces, Luaña playa. 43°23'53"N 4°12'50"W (WGS 84).
Type level: Cretaceous, lower Cenomanian, Dixoni Zone.
Diagnosis: Eosiderastrea with an astreoid calicular arrangement, non to sub-confluent costae and septa in hexameral symmetry with four septal cycles (43 to 51 septa in the holotype).
Description: Astreoid colony. Calicular outline circular to polygonal. Septa are compact. In cross section, the septa (and costae) in the wall are thick, thinning towards the centre. Symmetry of septa radial is hexameral with sub-regular cycles. Septal cycles differ in length. Generally, two septal cycles reach the centre of the calice, further cycles are shorter. Septa of the last cycle are often attached to those of the third cycle. Septal upper border granulated, lateral face with numerous, irregularly distributed fine thorns, inner margin slightly swollen in places. Costae present, non-confluent to confluent. Synapticulae present, but not common, mainly in the wall region. Columella by septal fusion. Endotheca consists of numerous thin tabulae. Wall absent. Coenosteum narrow. Budding extracalicinal.
Dimensions:
| (BSPG 2007 V 309) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 20 | 54.70-6.5 | 5.53 | 0.50 | 9.1 | 5.02-6.03 |
| clmax | 20 | 4.88-7.00 | 5.80 | 0.60 | 10.4 | 5.20-6.41 |
| ccd | 40 | 4.10-6.65 | 5.47 | 0.72 | 13.1 | 4.75-6.19 |
| s | 20 | 43-51 | 46.3 | 2.5 | 5.3 | 44-49 |
| (BSPG 2007 V 329) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 3.65-5.18 | 4.52 | 0.50 | 11.1 | 4.01-5.02 |
| clmax | 10 | 4.38-6.82 | 5.59 | 0.90 | 16.1 | 4.68-6.50 |
| ccd | 10 | 4.15-5.84 | 4.82 | 0.47 | 9.8 | 4.35-5.29 |
| s | 10 | 35-47 | 40.30 | 3.59 | 8.9 | 37-44 |
| (BSPG 2003 XX 1147) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 6 | 4.20-5.67 | 5.17 | 0.54 | 10.4 | 4.63-5.72 |
| clmax | 6 | 5.32-5.85 | 5.66 | 0.21 | 3.7 | 5.45-5.87 |
| ccd | 8 | 4.65-6.06 | 5.37 | 0.58 | 10.8 | 4.78-5.95 |
| s | 3 | 51-64 | 59.33 | 7.23 | 12.1 | |
| (BSPG 2003 XX 5812) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 15 | 4.66-6.21 | 5.50 | 0.43 | 7.9 | 5.06-5.94 |
| ccd | 20 | 3.92-6.91 | 5.40 | 0.86 | 16.0 | 4.53-6.27 |
| s | 7 | 42-44 | 42.85 | 0.69 | 1.6 | |
Comparisons: In its calicular dimensions the new species is comparable to E. dollfusi, which clearly shows a wall. It is similar to E. grandipora but has smaller dimensions.
Occurrence: Lower Cenomanian (Mantelli Zone) of Germany (Nordrhein/Westfalen) Mülheim/Ruhr, Kassenberg; lower Cenomanian of Greece (Kozani) Kozani, Nea Nikopolis; lower Cenomanian (Dixoni Zone) of Spain (Cantabria, Santander) Cobreces, Luaña playa; uppermost Cenomanian (Juddi Zone) of France (Aude) Les Corbières, Sougraigne, Prat-Périé.
Eosiderastrea reussi (Milne Edwards, 1857)
(Pl. 5 ![]() ,
figs. 1-5)
,
figs. 1-5)
Material: GPSL FLX 3126, FLX 5385, FLX 8059, MHE A0601, NHMW 1864/0040/1352; 6 thin sections.
Synonymy:
|
v |
Pleurocora Haueri M. Edw. et H. - Reuss, p. 112, Pl. 6, figs. 26-27 |
|
|
*v |
Pleurocora Reussi Milne Edwards, (2), p. 602 |
|
|
Pleurocora reussi Milne-Edwards - Oppenheim, p. 365, Pl. 35, fig. 12; Pl. 45, fig. 9 |
||
|
Neocaeniopsis reussi (H. Milne-Edwards) 1857 - Beauvais, (2), p. 109, Pl. 36, fig. 5 |
Dimensions:
| (NHMW 1864/0040/1352) | |
| cmin | 9.62 |
| s | 90 |
| (MHE A0601) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 4 | 7.11-9.19 | 8.30 | 0.86 | 10.4 | 7.44-9.17 |
| cmax | 4 | 9.55-12.33 | 10.98 | 1.18 | 10.8 | 9.80-12.17 |
| ccd | 7 | 6.46-9.73 | 7.87 | 1.08 | 13.7 | 6.79-8.95 |
| s | 4 | 58-74 | 68.25 | 7.32 | 10.7 | |
Remarks: E. reussi is a problematic species because it has never been properly recognised. The holotype is small and has a polished surface on only one calice. Additional material has been difficult to obtain. The species seems to be extremely rare in the Santonian of the Gosau Valley. All the specimens from the GPSL Felix collection are small, only the MHE sample allowed some modest statistical analysis of the species. E. reussi is the species with the largest dimensions. The calicular arrangement is astreoid, the costae are non-confluent and the septal symmetry is hexameral with four septal cycles.
Occurrence: Coniacian to Santonian of Austria (Oberösterreich) Gosau basin. Santonian of Austria (Salzburg) Rußbach, Zimmergraben and Neffgraben; Austria (Oberösterreich) Gosau, Wegscheidgraben.
Eosiderastrea splendida (Prever, 1909)
(Pl. 5 ![]() ,
figs. 6-7)
,
figs. 6-7)
Material: PU 17993.
Synonymy:
|
*v |
Diplocoenia splendida Prever, p. 127, Pl. 13, fig. 19 |
Dimensions:
| (PU 17993) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 4 | 3.40-5.13 | 4.28 | 0.75 | 17.7 | 3.52-5.04 |
| cmin | 4 | 5.38-6.35 | 5.88 | 0.43 | 7.3 | 5.45-6.31 |
| ccd | 9 | 5.59-7.32 | 6.20 | 0.56 | 9.1 | 5.63-6.76 |
| s | 60-65 | |||||
Remarks: There is only the small and poorly preserved type specimen known. It is a plocoid colony with non-confluent costae. The septal are arranged in octameral symmetry forming four septal cycles with about 60 to 65 septa. Due to the poor state of the preservation, precise septal counts are difficult to make.
Occurrence: Lower Aptian of Italy (Abruzzi, L'Aquila) Monti d'Ocre, Fossa Cerasetti
Eosiderastrea stefani n. sp.
(Pl. 2 ![]() ,
figs. 7-9)
,
figs. 7-9)
Synonymy:
|
v |
?Diploastrea tanohataensis (Eguchi, 1951) - Löser, Werner & Darga, p. 46, Pl. 2, figs. 7-9 |
|
|
v |
"Diploastrea" tanohataensis Eguchi, 1951 - Löser, p. 281, Fig. 193.B |
Dimensions:
| (BSPG 2007 V 61) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 17 | 4.35-6.31 | 5.33 | 0.63 | 11.9 | 4.69-5.97 |
| cmax | 17 | 5.04-8.17 | 6.44 | 0.89 | 13.8 | 5.55-7.34 |
| ccd | 20 | 4.05-8.26 | 5.90 | 1.39 | 23.6 | 4.50-7.30 |
| s | 6 | 41-46 | 44.5 | 2.34 | 5.2 | |
| (MHNLM 2003.1.7593b) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| ccd | 15 | 4.38-7.03 | 5.58 | 0.71 | 12.7 | 4.87-6.30 |
| cmax | 15 | 4.72-7.75 | 5.90 | 0.80 | 13.6 | 5.09-6.70 |
| cmin | 15 | 4.71-6.76 | 5.61 | 0.66 | 11.7 | 4.95-6.27 |
| s | 5 | 34-44 | 40.80 | 4.14 | 10.1 | 36.65-44.94 |
Origin of the name: Named after Stefan Götz (1965-2012) who in 2007 initiated the massive re-collection of fossils (mainly corals) at the type locality and who invited me to revise the corals (which are the subject of a paper in progress).
Holotype: BSPG 2007 V 61.
Material studied: Holotype, six further specimens (BSPG 1947 XVI 47, 2003 XX 6145, ERNO L-5614, MHNLM 2003.1.7593, 2003.1.7593b, NHM R43455; 7 thin sections.) from different localities.
Type locality: Spain, Cantabria, Santander, Cobreces, Luaña playa. 43°23'46"N 4°12'56"W (WGS 84).
Type level: Cretaceous, lower Cenomanian, Dixoni Zone.
Diagnosis: Eosiderastrea with an astreoid calicular arrangement, sub-confluent costae and septa in decameral symmetry with three septal cycles (41 to 46 septa in the holotype).
Description: Astreoid colony. Calicular outline circular. Septa have a few perforations at the inner margins. In cross section the septa (and costae) in the wall are thick, thinning towards the centre. Symmetry of septa radial is decameral with sub-regular cycles. Septal cycles differ in length. Generally, two septal cycles reach the centre of the calice, the third cycle is shorter. Septa of the third cycle are attached to those of the second cycle in places. Septal upper border granulated, lateral face with numerous, irregularly distributed fine thorns, inner margin smooth. Costae present, non-confluent to confluent, but rather non-confluent. Synapticulae not common. Columella by septal fusion. Endotheca consists of a few thin dissepiments. Wall absent. Coenosteum narrow. Budding extracalicinal.
Comparisons: The new species is only comparable to E. felixi; however, it differs due to the absence of a wall. E. tanohatoensis, to which the material was formerly assigned, is now assigned to E. felixi.
Occurrence: Lower Cretaceous of Palestine, Mulergat Well; lower Cenomanian of France (Charente-Maritime) Fouras; Greece (Kozani) Kozani, Nea Nikopolis; lower Cenomanian (Dixoni Zone) of Spain (Cantabria, Santander) Cobreces, Luaña playa; middle Cenomanian of Germany (Bayern) Roßstein-Almen; middle to upper Cenomanian (Rhotomagense - Naviculare Zone) of France (Sarthe) Le Mans.
Eosiderastrea vaughani (Wells, 1933)
(Pl. 5 ![]() ,
figs. 9-11)
,
figs. 9-11)
Material: NMNH I-75128, TMM 1454TX1; 5 thin sections.
Synonymy:
|
*v |
Diploastraea vaughani Wells, p. 94, Pl. 8, figs. 13-14 |
Dimensions:
| (USNM I-75128) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 30 | 2.07-2.70 | 2.38 | 0.18 | 7.7 | 2.19-2.56 |
| clmax | 30 | 2.18-3.47 | 2.88 | 0.27 | 9.4 | 2.61-3.16 |
| ccd | 30 | 2.76-4.32 | 3.43 | 0.43 | 12.7 | 3.00-3.87 |
| s | 10 | 37-46 | 41.30 | 2.62 | 6.3 | 39-44 |
| (TMM 1454TX1) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 30 | 1.10-2.02 | 1.65 | 0.21 | 13.2 | 1.43-1.87 |
| clmax | 30 | 1.65-2.42 | 2.01 | 0.22 | 11.0 | 1.79-2.24 |
| ccd | 30 | 2.32-3.50 | 2.86 | 0.30 | 10.4 | 2.56-3.16 |
| s | 15 | 34-46 | 39.1 | 3.7 | 9.5 | 35-43 |
Remarks: E. vaughani is the species with the smallest dimensions. The species is characterised by an extremely extended coenosteum and a high amount of synapticulae in the coenosteum. The specimen 1454TX1 is very similar to the type of E. vaughani but has smaller dimensions. It also shows a more extended coenosteum and more synapticulae than other specimens. The locality of the latter specimen is indicated as 'Texas, Travis County, Pilot Knob, north rim, with an early late Campanian age. This locality is doubted. Material from the Pilot Knob was available for study, also in thin sections, and all the coral specimens were much more poorly preserved. The lithology of the TMM 1454TX1 is more similar to corals from the upper Aptian or lower Albian outcrops in Texas.
Occurrence: Middle Albian (Lautus Zone) of USA (Texas) Kerr County, Kerrville, Hiram Hall Ranch.
Eosiderastrea villaltai (Reig Oriol, 1991)
(Pl. 6 ![]() ,
figs. 1-3)
,
figs. 1-3)
Material: BSPG 1994 XI 13, 292, 293, 294, 53, BSPG 1996 VI 2, BSPG 1997 V 113-2, ERNO L-4789, L-7368, MV 13539, PIUEN b45; 9 thin sections.
Synonymy:
|
*v |
Plesiofavia villaltai Reig Oriol, p. 19, Pl. 4, fig. 9 |
|
|
v |
Isastrea neocomiensis Fromentel, 1857 - Baron-Szabo, p. 70, Pl. 7, figs. 2 & 4 |
|
|
v |
Diploastrea harrisi Wells, 1932 - Baron-Szabo et al., p. 210, Pl. 37, fig. 3; Pl. 38, fig. 2; Pl. 39, fig. 4 |
Dimensions:
| (BSPG 1994 XI 53) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 30 | 3.34-4.46 | 3.93 | 0.29 | 7.6 | 3.63-4.23 |
| clmax | 30 | 4.13-5.43 | 4.70 | 0.36 | 7.7 | 4.33-5.06 |
| cmin | 30 | 4.74-6.18 | 5.50 | 0.43 | 7.9 | 5.06-5.93 |
| cmax | 30 | 5.31-7.35 | 6.16 | 0.59 | 9.7 | 5.56-6.76 |
| ccd | 40 | 4.54-6.72 | 5.67 | 0.60 | 10.7 | 5.06-6.27 |
| s | 25 | 34-44 | 39.1 | 3.1 | 7.9 | 36-42 |
Remarks: The species forms astreoid colonies that possess no wall. The septa are non-confluent. The symmetry is octameral with three to four septal cycles. In its dimensions, it is comparable to E. splendida but differs by the absence of a wall.
Occurrence: Upper Barremian to lower Aptian (Sartousi - Weissi Zone) of Germany (Bayern) Allgäuer Helvetikum, Falkenberg; NE Hoher Ifen, Kühberg; Tiefenbach; lower upper Aptian of Spain (Cataluña, Lérida) Com. La Noguera, Mun. Vilanova de Meià, Montsec de Rubies, section NW La Cabrua quarry; upper Aptian to Albian of Iran (Esfahan) Esfahan Basin, Dizlu; lowermost Albian (Tardefurcata Zone) of Spain (Cataluña, Tarragona) Com. Baix Penedés, Mun. Olivella, Can Grau; lower Albian of USA (Texas) Kendall County, Guadalupe River Ranch.
Eosiderastrea sp. 1
(Pl. 6 ![]() ,
figs. 4-6)
,
figs. 4-6)
Material: ERNO L-121509; 2 thin sections.
Dimensions:
| (ERNO L-121509) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 13 | 2.84-4.47 | 3.73 | 0.56 | 15.1 | 3.16-4.29 |
| ccd | 16 | 5.68-7.92 | 6.76 | 0.60 | 8.9 | 6.15-7.36 |
| s | 9 | 46-61 | 51.55 | 5.3 | 10.2 | 46-57 |
Remarks: This astreoid species with mainly non-confluent septa and an incomplete wall shows hexameral symmetry with four septal cycles. It is the oldest known species.
Occurrence: Upper Valanginian of Spain (Murcia) Sierra Larga.
Eosiderastrea sp. 2
(Pl. 5 ![]() ,
fig. 8)
,
fig. 8)
Material: BSPG 2003 XX 6539; 1 thin section.
Synonymy:
|
v |
"Diploastrea" sp. - Löser, Fig. 2.3 |
Dimensions:
| (BSPG 2003 XX 6539) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 7 | 3.63-5.59 | 4.64 | 0.63 | 13.6 | 4.01-5.28 |
| cmin | 7 | 4.68-7.39 | 5.97 | 1.05 | 17.6 | 4.92-7.03 |
| ccd | 11 | 5.71-8.83 | 7.16 | 1.10 | 15.4 | 6.05-8.26 |
| s | 3 | 43-60 | ||||
Remarks: This species shows an astreoid calicular arrangement. The wall is absent. The septa are non to sub-confluent. The septal symmetry is hexameral.
Occurrence: Lower Hauterivian (Radiatus Zone) of France (Yonne) Gy-l'Evêque, fields SW Gy-l'Evêque.
Eosiderastrea sp. 3
(Pl. 6 ![]() ,
figs. 7-9)
,
figs. 7-9)
Material: SNSD-MMG SaKL542; 2 thin sections.
Synonymy:
|
v |
"Diploastrea" sp. 2 - Löser, p. 28, Fig. 4.b |
Dimensions:
| (SNSD-MMG SaKL542) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 10 | 2.12-2.74 | 2.34 | 0.19 | 8.3 | 2.15-2.54 |
| clmax | 10 | 2.54-3.35 | 2.78 | 0.26 | 9.5 | 2.52-3.05 |
| cmin | 10 | 2.92-4.08 | 3.65 | 0.40 | 11.0 | 3.24-4.05 |
| cmax | 10 | 3.75-4.54 | 4.15 | 0.29 | 7.0 | 3.86-4.44 |
| ccd | 10 | 3.26-4.21 | 3.70 | 0.33 | 9.1 | 3.36-4.04 |
| s | 6 | 50-63 | 53.83 | 4.66 | 8.6 | |
Remarks: This astreoid form shows octameral symmetry with a strong bilateral component. It is most similar to Siderastraea, but the wall is incomplete and very weak. The septal microstructure is partly preserved. The columella is made up of a group of isolated trabeculae, which is rarely visible in other species.
Occurrence: Lower Cenomanian (Dixoni Zone) of Germany (Sachsen) Meißen-Zscheila, Trinitatis church.
Eosiderastrea sp. 4
(Pl. 7 ![]() , figs. 4-6)
, figs. 4-6)
Material: BSPG 2003 XX 4027, 2003 XX 5846, ERNO L-130135, L-130206, L-5601, UPS HL-025; 8 thin sections.
Dimensions:
| (BSPG 003 XX 5846) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| clmin | 20 | 3.80-5.55 | 4.77 | 0.44 | 9.3 | 4.32-5.21 |
| cmin | 15 | 5.91-8.04 | 6.96 | 0.64 | 9.2 | 6.32-7.61 |
| ccd | 20 | 5.11-7.71 | 6.55 | 0.85 | 12.9 | 5.70-7.40 |
| s | 12 | 39-47 | 41.83 | 2.40 | 5.7 | 39-44 |
Remarks: The astreoid species shows non to sub-confluent septa. A wall is absent. The septal symmetry is decameral.
Occurrence: Uppermost Aptian of Spain (Cataluña, Lérida) Com. Alt Urgell, Mun. Coll de Nargó, Set Comelles, El Caso section; middle Albian of Mexico (Sonora) Municipio San Pedro de la Cueva, Tepache, Lampazos area, Espinazo de Diablo; lower Cenomanian of France (Charente-Maritime) Fouras; Greece (Kozani) Kozani, Nea Nikopolis; uppermost Cenomanian (Juddi Zone) of France (Aude) Col de Escudiés.
Eosiderastrea sp. 5
(Pl. 7 ![]() , figs. 1-3)
, figs. 1-3)
Material: BSPG 1991 X 97, 2003 XX 5829; 4 thin sections.
Synonymy:
|
v |
?Diploastrea sp. - Löser et al., p. 48, Pl. 1, figs. 5-6 |
Dimensions:
| (BSPG 1991 X 97) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 3 | 7.50-7.56 | 7.53 | 0.03 | 0.4 | 7.50-7.56 |
| ccd | 7 | 5.16-8.26 | 6.84 | 1.25 | 18.2 | 5.59-8.09 |
| s | 60-70 | |||||
| (BSPG 2003 XX 5829) | ||||||
| n | min-max | µ | s | cv | µ±s | |
| cmin | 6 | 5.82-8.42 | 7.11 | 1.05 | 14.8 | 6.06-8.17 |
| ccd | 12 | 6.05-8.37 | 7.09 | 0.84 | 11.9 | 6.24-7.93 |
| s | 5 | 51-63 | 57.8 | 5.8 | ||
Remarks: This species forms astreoid colonies with sub to non-confluent septa. The septal symmetry is difficult to decipher, most likely it is decameral with three to four cycles. A wall does not exist. Both specimens show higher septal perforations than any other species.
Occurrence: Lower Cenomanian of Greece (Kozani) Kozani, Nea Nikopolis; middle Cenomanian of Germany (Bayern) Roßstein-Almen.
Eosiderastrea sp. 6
(Pl. 6 ![]() ,
figs. 10-11)
,
figs. 10-11)
Material: ERNO L-0955, SNSD-MMG SaKL253; 3 thin sections.
Synonymy:
|
v |
1989 |
Brachyphyllia sp. - Löser, p. 138, Figs. 39-40; Pl. 27, figs. 1-2 |
|
v |
2014 |
"Diploastrea" sp. 1 - Löser, p. 28, Fig. 4.a |
Dimensions:
| (SNSD-MMG SaKL253) | |
| cmin | 4.8-5.1 |
| ccd | 4.3-4.9 |
| s | ca. 70 |
Remarks: This species shows an astreoid calicular arrangement with non-confluent septa. A wall does not exist. The septal symmetry is probably decameral with almost four cycles.
Occurrence: Lower Cenomanian (Dixoni Zone) of Germany (Sachsen) Meißen-Zscheila, Trinitatis church.
Kuzmicheva (2002, Pl. 27, fig. 5) depicts a specimen from the upper Aptian of Uzbekistan, Hemiporites jacobi, that clearly belongs to Eosiderastrea but cannot be assigned to any species. The indicated magnification factor (2x) would indicate a smaller outer calicular diameter of more than 10mm. The symmetry is hexameral with three cycles of septa. Since the number of septa generally increases with the calicular diameter (see Table 1), the indicated magnification is in doubt. The same applies to the material presented by Baron-Szabo (1993, Pl. 4, fig. 6) as Diploastrea harrisi; the measured small calicular diameter of seven to eight millimetres does not coincide with the comparably low number of septa (32). Baryphyllia gasseri Reig Oriol, 1995, from the lowermost Albian of East Iberia may belong to Eosiderastrea, but the type is small and has neither a polished section nor thin sections.
Species
separation: To cross check the taxonomic assignments and to
identify possible synonyms, the co-occurrences of the taxa in palaeo-provinces
(see Löser
& Minor, 2007) were correlated resulting in a correlation dendrogram (Fig. 1 ![]() ). Highly
correlated species with comparable morphological characteristics (septal
systems, calicular dimensions) could be synonymous. A high palaeobiogeographic
correlation was seen in the following species:
). Highly
correlated species with comparable morphological characteristics (septal
systems, calicular dimensions) could be synonymous. A high palaeobiogeographic
correlation was seen in the following species:
E. grandipora and the new species E. paragrandipora: both species differ clearly in their calicular dimensions.
E. cotteaui and E. cf. cotteaui: the species differ only in their calicular dimensions. It is possible that both species are synonymous but more and better preserved material is necessary to understand the variability of the complex.
E. cyclops and E. cf. cyclops: the species differ only by their calicular dimensions. E. cyclops is not well defined. Material, aside from the type material, consists of small colonies that do not allow the estimation of intraspecific variability.
E. cuyleriand E. cf. cuyleri (holotype and paratype of the species cuyleri): the species differ clearly in their calicular dimensions and the septal symmetry.

Click on thumbnail to enlarge the image.
Figure 1: Correlation of the Eosiderastrea species based on their occurrence in the palaeo-provinces. The Correlation Ratio coefficient was applied. The graph is logarithmic.
Stratigraphic
distribution: The
genus diversified during the earliest Hauterivian but is not
present from there to the early Barremian (Fig. 2 ![]() ). From the
early Barremian onwards, the genus was rich in species, surprisingly also in
the Albian. Coral faunas from this age are poorly represented in the Tethys but
common in the Western Atlantic, where most species of this age are derived.
There is a gap again in the late Albian, but the genus reached its
highest diversity in the early to middle Cenomanian, with a drop in
the late Cenomanian. One last species is known from the Santonian.
The gap in the Turonian and Coniacian is probably due to a lack of available
collections. The genus is widely distributed throughout clastic and carbonatic
sediments. It seems to be more common in clastic sediments but this may be also
due to a sample bias because corals are more difficult to obtain from massive
limestones and can be more easily collected from clastic sediments.
). From the
early Barremian onwards, the genus was rich in species, surprisingly also in
the Albian. Coral faunas from this age are poorly represented in the Tethys but
common in the Western Atlantic, where most species of this age are derived.
There is a gap again in the late Albian, but the genus reached its
highest diversity in the early to middle Cenomanian, with a drop in
the late Cenomanian. One last species is known from the Santonian.
The gap in the Turonian and Coniacian is probably due to a lack of available
collections. The genus is widely distributed throughout clastic and carbonatic
sediments. It seems to be more common in clastic sediments but this may be also
due to a sample bias because corals are more difficult to obtain from massive
limestones and can be more easily collected from clastic sediments.

Click on thumbnail to enlarge the image.
Figure 2: Cumulative stratigraphic distribution of the Eosiderastrea species. The thickness of the bars corresponds to the number of specimens from the various time intervals. The bar at the bottom summarises the occurrences and indicates diversity peaks in the earliest Aptian and early to middle Cenomanian.
Palaeobiogeography: The
genus shows an almost worldwide distribution, being most abundant in the Tethys
and less abundant in the Western Atlantic and Western Pacific, except for the
Albian when coral faunas were more common in the Western Atlantic than in the
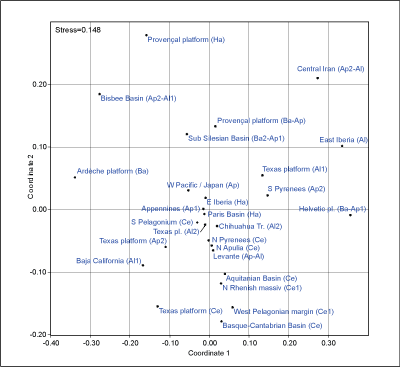
Tethys. No clear palaeobiogeographic relationships exist (Fig. 3 ![]() ). The number
of species is high, but the number of specimens (and therefore occurrences) is
comparatively low, which makes a correlation difficult. A certain correlation
of provinces with the same age can be observed. A multidimensional scaling
analysis (based on a correlation of the absence or presence in
palaeo-provinces) for all occurrences (Fig. 4
). The number
of species is high, but the number of specimens (and therefore occurrences) is
comparatively low, which makes a correlation difficult. A certain correlation
of provinces with the same age can be observed. A multidimensional scaling
analysis (based on a correlation of the absence or presence in
palaeo-provinces) for all occurrences (Fig. 4 ![]() ) also shows a correlation between
species that co-occur in the same stages, in general in the Cenomanian and
Barremian to Aptian. A more detailed palaeobiogeographic analysis is difficult
to provide because of the low amount of data.
) also shows a correlation between
species that co-occur in the same stages, in general in the Cenomanian and
Barremian to Aptian. A more detailed palaeobiogeographic analysis is difficult
to provide because of the low amount of data.

Click on thumbnail to enlarge the image.
Figure 3: Correlation of provinces with joint species in the study area. The Correlation Ratio coefficient was applied. For details of the calculation see Löser & Minor (2007).
Click on thumbnail to enlarge the image.
Figure 4: Multidimensional scaling (Euclidean), based on the correlation in figure 3. The STRESS value is relatively high due to the low number of occurrences in comparison to the high number of species.
Evolution: There is an interesting tendency for an increase in calicular diameter during the Cretaceous. The species with small diameters occurred predominantly during the Hauterivian to Aptian; while species wither larger dimensions occurred mainly during the Cenomanian and the Santonian. With the increasing size of the corallites, the number of septa also increased, so the species with the highest number of septa are mainly found in the early Late Cretaceous.
I am grateful to Steven Cairns (Washington D.C.), Jill Darrell (London), Matthias Heinrich (Eckental), Ann Molineux (Austin), Nicolas Morel (Le Mans), Kei Mori (Sendai), Arnold Müller (Leipzig), Daniele Ormezzano (Torino), Baba Senowbari-Daryan (Erlangen), and Winfried Werner (München) who allowed me to examine the (type) material of their collections. Field work, the examination of type material in the USA, and the preparation of thin sections were partly covered by PAPIIT-DGAPA project IN101111. Additional thin sections were carefully prepared by Aimée Orcí (Hermosillo). English language correction by Proof-Reading-Service (Letchworth Garden City, England). Two anonymous reviews helped to improve the manuscript.
Alloiteau J. (1952).- Embranchement des coelentérés. In: Piveteau J. (ed.), Traité de Paléontologie (1).- Masson, Paris, p. 376-684.
Baron-Szabo R.C. (1993).- Korallen der höheren Unterkreide ("Urgon") von Nordspanien (Playa de Laga, Prov. Guernica).- Berliner geowissenschaftliche Abhandlungen, Berlin, (E), vol. 9, p. 147-181.
Baron-Szabo R.C. (1997).- Die Korallenfazies der ostalpinen Kreide (Helvetikum: Allgäuer Schrattenkalk; Nördliche Kalkalpen: Brandenberger Gosau). Taxonomie, Palökologie.- Zitteliana, München, vol. 21, p. 3-97.
Baron-Szabo R.C. & González León C.M. (2003).- Late Aptian-early Albian corals from the Mural Limestone of the Bisbee Group (Tuape and Cerro de Oro areas), Sonora, Mexico. In: Scott R.W. (ed.), Bob F. Perkins Memorial Volume.- Special Publications in Geology, Houston, p. 187-225.
Baron-Szabo R.C., Hamedani A. & Senowbari-Daryan B. (2003).- Scleractinian corals from Lower Cretaceous deposits north of Esfahan (Central Iran).- Facies, Erlangen, vol. 48, p. 199-216.
Beauvais M. (1982).- Révision systématique des Madréporaires des couches de Gosau.- Comptoir géologique, Paris, 5 vols.
Bilotte M. (2007).- Permanence, au Crétacé supérieur, de la position de la limite plate-forme/bassin dans la zone sous-pyrénéenne orientale (Aude, France). Implications géodynamiques.- Géologie de la France, Paris, vol. 1, p. 33-53.
Bourne G.C. (1900).- The Anthozoa. In: Lankester R. (ed.), Treatise on Zoology, Part 2, Porifera and Coelenterata.- A. & C. Black, London, p. 59-79.
Budd A.F. & Coates A.G. (1992).- Nonprogressive evolution in a clade of Cretaceous Montastrea-like corals.- Paleobiology, Lawrence, vol. 18, no. 4, p. 425-446.
Eguchi M. (1951).- Mesozoic hexacorals from Japan.- Science Reports of the Tohoku Imperial University, Sendai, (2: Geology), vol. 24, p. 1-96.
Felix J. (1891).- Versteinerungen aus der mexicanischen Jura und Kreideformation.- Palaeontographica, Stuttgart, vol. 37, p. 140-194.
Gill G.A. (1967).- Madréporaires (2:) Quelques précisions sur les septes perforés des polypiers mésozoïques.- Mémoires de la Société géologique de France, Paris, vol. 106, p. 53-83.
Gill G.A. (1981).- The fulturae ("compound synapticulae"), their structure and reconsideration of their systematic value.- Acta Palaeontologica Polonica, Warszawa, vol. 25, p. 301-310.
Götz S., Löser H. & Schmid D.U. (2005).- Reef development on a deepening platform: two Early Cretaceous coralgal patch reefs (Catí, Llàcova Formation, eastern Spain) compared.- Cretaceous Research, vol. 26, no. 6, p. 864-881.
Kuzmicheva E.I. (1980).- Corals. In: Chernov V.G., Yanin B.T. & Golovinova M.A. (eds.), Urgonian sediments of the Soviet Carpathes.- Nauka, Moskva, p. 90-108 [in Russian].
Kuzmicheva E.I. (2002).- Skeletal morphology, systematics and evolution of the Scleractinia.- Trudy Paleontologicheskogo instituta, Moskva, vol. 286, 211 p. [in Russian].
Löser H. (1989).- Die Korallen der sächsischen Oberkreide (1:) Hexacorallia aus dem Cenoman.- Abhandlungen des Staatlichen Museums für Mineralogie und Geologie zu Dresden, Leipzig, vol. 36, p. 88-154.
Löser H. (1994).- La faune corallienne du mont Kassenberg à Mülheim-sur-la-Ruhr (Bassin crétacé de Westphalie, Nord Ouest de l'Allemagne).- Coral Research Bulletin, Dresden, vol. 3, 93 p.
Löser H. (2001).- Le site de Vallières (département de l'Aube, France) : résultats préliminaires sur des coraux de l'Hauterivien inférieur (Crétacé).- Bulletin annuel de l'Association géologique de l'Aube, Sainte Savine, vol. 22, p. 39-53.
Löser H. (2002).- Verfahren einer von der Taxonomie unabhängigen paläobiogeographischen Analyse post-paläozoischer Korallen.- Mathematische Geologie, Dresden, vol. 6, p. 15-43.
Löser H. (2006).- Barremian corals from San Antonio Texcala, Puebla, Mexico - A review of the type material of Felix 1891.- Boletín del Instituto Geológico de México, Mexico City, vol. 114, p. 1-68.
Löser H. (2012).- Intraspecific variation in the genus Stelidioseris (Family Actinastraeidae, Suborder Archeocaeniina, Order Scleractinia; Jurassic-Cretaceous).- Geologica Belgica, Brussels, vol. 15, no. 4, p. 382-387.
Löser H. (2013).- Revision of the Hauterivian (Early Cretaceous) corals of the Paris Basin, France: a work in progress.- Bulletin d'Information des Géologues du Bassin de Paris, vol. 50, no. 1, p. 17-24.
Löser H. (2014).- 3. Korallen / 3. Corals. In: Niebuhr B. & Wilmsen M. (eds.), Kreide-Fossilien in Sachsen, Teil 1.- Geologica Saxonica, Dresden, vol. 60, no. 1, p. 17-56.
Löser H. (2015).- Les coraux. In: Morel N. (ed.), Stratotype Cénomanien.- Muséum national d'Histoire naturelle, Paris, p. 280-282.
Löser H. (2016).- Early evolution of the coral Family Siderastraeidae (Scleractinia; Cretaceous-extant).- Paläontologische Zeitschrift, Berlin, vol. 90, no. 1, p. 1-17.
Löser H. & Ferry S. (2006).- Coraux du Barrémien du Sud de la France (Ardèche et Drôme).- Geobios, Villeurbanne, vol. 39, no. 4, p. 469-489.
Löser H. & Minor K. (2007).- Palaeobiogeographic aspects of late Barremian to late Albian coral faunas from Northern Mexico (Sonora) and the southern USA (Arizona, Texas).- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 245, no. 2, p. 193-218.
Löser H., Werner W. & Darga R. (2013).- A Middle Cenomanian coral fauna from the Northern Calcareous Alps (Bavaria, Southern Germany) - new insights into the evolution of Mid-Cretaceous corals.- Zitteliana, München, vol. A53, p. 37-76.
Löser H. (ed.), Barattolo F., Badia S., Chikhi-Aouimeur F., Dhondt A., Erlich R.N., Fözy I., Geister J., Hiss M., Kolodziej B., Leloux J., Lewy Z., Minor K.P., Mitchell S., Moosleitner G., Peza L., Remane J., Romano R., Sikharulidze G.Y., Sinnyovski D., Steuber T., Tröger K.-A., Turnsek D., Vecchio E., Vilella i Puig J. & Zítt J. (2002).- List of Citations.- Catalogue of Cretaceous Corals, Dresden, vol. 2, 784 p.
Löser H. (ed.), Barattolo F., Badia S., Chikhi-Aouimeur F., Dhondt A., Erlich R.N., Fözy I., Geister J., Hiss M., Kolodziej B., Leloux J., Lewy Z., Madhavaraju J., Minor K.P., Mitchell S., Moosleitner G., Niebuhr B., Peza L., Remane J., Romano R., Sanders D., Sikharulidze G.Y., Sinnyovski D., Steuber T., Tröger K.-A., Turnsek D., Vecchio E., Vilella i Puig J. & Zítt J. (2005).- List of Localities.- Catalogue of Cretaceous Corals, Dresden, vol. 3, 366 p.
Matthews S.C. (1973).- Notes on open nomenclature and on synonymy lists.- Palaeontology, London, vol. 16, no. 4, p. 713-719.
Milne Edwards H. (1857).- Histoire naturelle des coralliaires ou polypes proprement dits (1+2).- Librairie encyclopédique de Roret, Paris, 2 vols.
Morycowa E. & Masse J.P. (1998).- Les Scléractiniaires du Barrémien-Aptien inférieur de Provence (SE de la France).- Geobios, Villeurbanne, vol. 31, no. 6, p. 725-766.
Morycowa E. & Roniewicz E. (1995).- Microstructural disparity between Recent fungiine and Mesozoic microsolenine scleractinians.- Acta Palaeontologica Polonica, Warszawa, vol. 40, p. 361-385.
Mucha H.-J. (1992).- Clusteranalyse mit Mikrocomputern.- Akademie-Verlag, Berlin, 206 p.
Oppenheim L.P. (1930).- Die Anthozoen der Gosauschichten in den Ostalpen.- Privately published, Berlin, 604 p.
Orbigny A. d' (1849).- Note sur les polypiers fossiles.- Masson, Paris, p. 1-12.
Orbigny A. d' (1850).- Prodrôme de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés (1-2).- Masson, Paris, 2 vols.
Payne J.L., Johnson M.E. & Ledesma-Vázquez J. (2004).- Formación Alisitos del Cretácico temprano en Punta San Isidro: Sedimentación costera y vulcanismo.- Ciencias Marinas, Ensenada, vol. 30, no. 2, p. 365-380.
Prever P.L. (1909).- Anthozoa. In: Parona C.F. (ed.), La fauna coralligena del Cretaceo dei Monti d'Ocre nell'Abruzzo Aquilano.- Memorie descrittive della carta geologica d'Italia, Firenze, vol. 5, no. 1, p. 51-147.
Reig Oriol J. (1991).- Fauna coralina cretácica del nordeste de España.- Privately published, Barcelona, 53 p.
Reuss A.E. (1854).- Beiträge zur Charakteristik der Kreideschichten in den Ostalpen, besonders im Gosauthale und am Wolfgangsee.- Denkschriften der Kaiserlichen Akademie der Wissenschaften, Mathematisch-Physikalische Classe, Wien, vol. 7, p. 1-157.
Reuss A.E. (1864).- Die fossilen Foraminiferen, Anthozoen und Bryozoen von Oberburg in Steiermark.- Denkschriften der Akademie der Wissenschaften, Mathemathisch-naturwissenschaftliche Klasse, Wien, vol. 23, p. 1-38.
Reyeros de Castillo M.M. (1983).- Corales de algunas formaciones cretacicas del estado de Oaxaca.- Paleontología mexicana, Mexico City, vol. 47, p. 1-67.
Scholz H. (1984).- Bioherme und Biostrome im Allgäuer Schrattenkalk (Helvetikum, Unterkreide).- Jahrbuch der Geologischen Bundesanstalt, Wien, vol. 127, no. 3, p. 471-499.
Shi G.R. (1993).- Multivariate data analysis in palaeoecology and palaeobiogeography - a review.- Palæogeography, Palæoclimatology, Palæoecology, vol. 105, p. 199-234.
Vaughan T.W. & Wells J.W. (1943).- Revision of the suborders, families and genera of Scleractinia.- Special Papers, Geological Society of America, Washington, D.C., vol. 44, 363 p.
Veron J.E.N. (1995).- Corals in space and time. The biogeography and evolution of the Scleractinia.- UNSW Press, Sydney, 321 p.
Verrill A.E. (1870).- Notes on the Radiata in the museum of Yale College, with descriptions of new genera and species. No. 6. Review of the corals and polyps of the West Coast of America.- Transactions of the Connecticut Academy of Arts and Sciences, New Haven, vol. 1, no. 2, p. 377-567.
Vilas L., Martín-Chivelet J. & Arias C. (2003).- Integration of subsidence and sequence stratigraphic analyses in the Cretaceous carbonate platforms of the Prebetic (Jumilla-Yecla Region), Spain.- Palæogeography, Palæoclimatology, Palæoecology, vol. 200, p. 107-129.
Wells J.W. (1932).- Corals of the Trinity Group of the Comanchean of central Texas.- Journal of Paleontology, Tulsa, vol. 6, no. 3, p. 225-256.
Wells J.W. (1933).- Corals of the Cretaceous of the Atlantic and Gulf Coastal Plains and Western Interior of the United States.- Bulletins of American Paleontology, Ithaca, N.Y., vol. 18, no. 67, p. 83-292.
Wells J.W. (1956).- Scleractinia. In: Moore R.C. (ed.), Treatise on Invertebrate Paleontology.- Geological Society of America, Boulder; University of Kansas Press, Lawrence, Part F, p. 328-444.
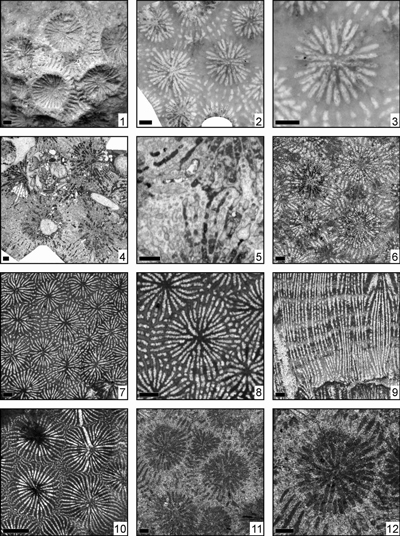
Click on thumbnail to enlarge the image.
Plate 1:
1) Eosiderastrea cotteaui
(Orbigny, 1850), syntype MNHN A25815, colony surface;
2-3) Eosiderastrea cf. cotteaui
(Orbigny, 1850), BSPG 2003 XX 6458; 2) transversal slab; 3) transversal
slab, detail;
4-5) Eosiderastrea crassa
(Kuzmicheva, 1980), TMM 1449TX6; 4) transversal thin section; 5)
transversal thin section, detail with microstructures;
6) Eosiderastrea crassicostata
(Morycowa & Masse, 1998), BSPG 2003 XX 6603, transversal thin
section;
7-9) Eosiderastrea cuyleri
(Wells, 1932), holotype TMM UT-11474; 7) transversal thin section; 8)
transversal thin section, detail; 9) longitudinal thin section;
10) Eosiderastrea aff. cuyleri
(Wells, 1932), TMM UT-10642, transversal thin section;
11-12) Eosiderastrea cf. cyclops
(Felix, 1891), BSPG 2003 XX 6602; 11) transversal thin section; 12)
transversal thin section, detail.
[Scale bar 1mm]
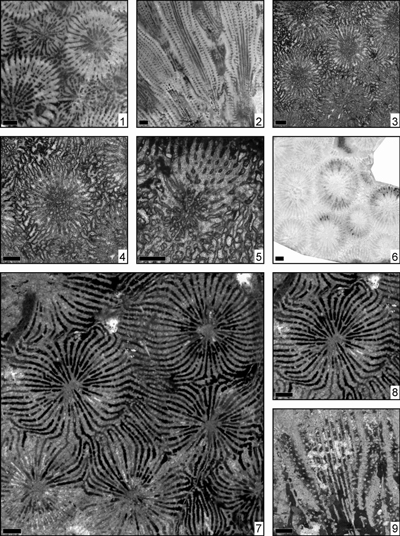
Click on thumbnail to enlarge the image.
Plate 2:
1-5) Eosiderastrea felixi
(Prever, 1909), holotype PU 17965; 1) transversal slab; 2) longitudinal
slab; 3) BSPG 2003 XX 6834, transversal thin section; 4) transversal thin
section, detail; 5) transversal thin section, detail;
6) Eosiderastrea dollfusi
(Prever, 1909), lectotype PU 17964, transversal slab;
7-9) Eosiderastrea stefani
n.sp., holotype BSPG 2007 V 061; 7) transversal thin section; 8) transversal
thin section, detail; 9) longitudinal thin section.
[Scale bar 1mm]
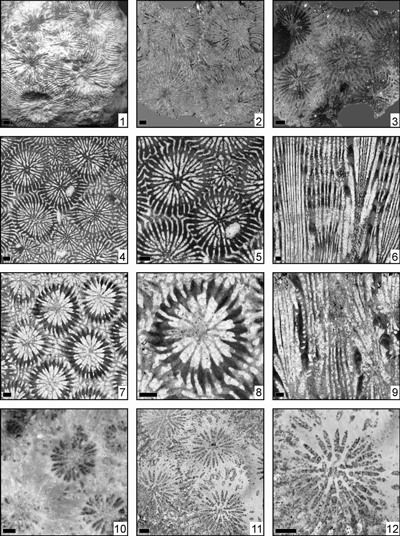
Click on thumbnail to enlarge the image.
Plate 3:
1-2) Eosiderastrea grandipora
(Orbigny, 1849), syntype MNHN A26591; 1) coral surface; 2) BSPG 2007 V
199, transversal thin section;
3) Eosiderastrea cyclops
(Felix, 1891), BSPG 2003 XX 6455, transversal thin section;
4-6) Eosiderastrea harrisi
(Wells, 1932), holotype TMM UT-11482; 4) transversal thin section; 5)
transversal thin section, detail; 6) longitudinal thin section;
7-9) Eosiderastrea harrisi
(Wells, 1932), holotype of Placocoenia orbitoides TUM
65367; 7) transversal thin section; 8) transversal thin section, detail; 9)
longitudinal thin section;
10-12) Eosiderastrea hilli
(Wells, 1933), holotype TMM UT-14180B; 10) transversal section; 11) TMM
1448TX1, transversal thin section; 12) transversal thin section, detail.
[Scale bar 1mm]
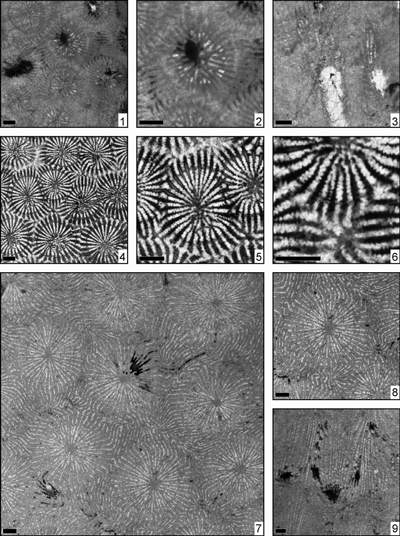
Click on thumbnail to enlarge the image.
Plate 4:
1-6) Eosiderastrea nagaoi
(Eguchi, 1951), holotype TUM 65317; 1) transversal thin section; 2)
transversal thin section, detail; 3) longitudinal thin section; 4) ERNO L-5193,
transversal thin section; 5) transversal thin section, detail; 6) transversal
thin section, detail;
7-9) Eosiderastrea paragrandipora
n. sp., holotype BSPG 2007 V 309; 7) transversal thin section; 8)
transversal thin section, detail; 9) longitudinal thin section.
[Scale bar 1mm]
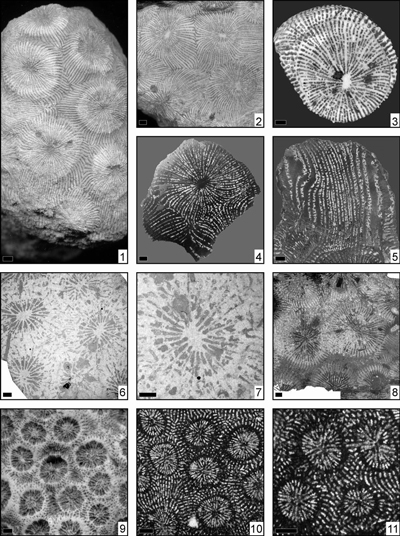
Click on thumbnail to enlarge the image.
Plate 5:
1-5) Eosiderastrea reussi
(Milne Edwards, 1857), MHE A0601; 1) coral surface; 2)
transversal section; 3) holotype NHMW 1864/0040/1352, transversal polished
section; 4) GPSL FLX 8059, transversal thin section; 5) longitudinal thin
section;
6-7) Eosiderastrea splendida
(Prever, 1909), holotype PU 17993; 6) transversal acetate peel; 7)
transversal acetate peel, detail; 8) Eosiderastrea sp. 2, BSPG 2003
XX 6539, transversal thin section;
9-11) Eosiderastrea vaughani
(Wells, 1933), holotype NMNH I-75128; 9) coral surface; 10) TMM 1454TX1,
transversal thin section; 11) transversal thin section, detail.
[Scale bar 1mm]
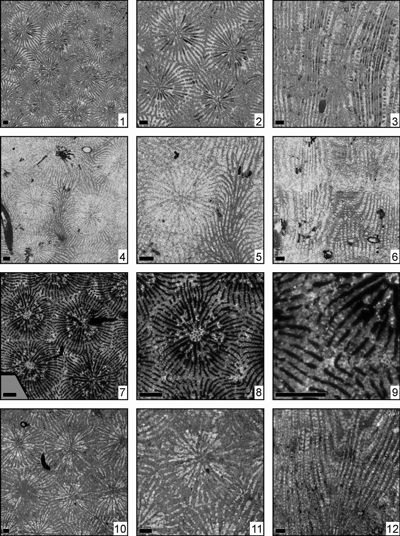
Click on thumbnail to enlarge the image.
Plate 6:
1-3) Eosiderastrea villaltai
(Reig Oriol, 1991), BSPG 1994 XI 53; 1) transversal thin section;
2) transversal thin section, detail; 3) longitudinal thin section;
4-6) Eosiderastrea sp. 1, ERNO
L-121509; 4) transversal thin section; 5) transversal thin section, detail; 6)
longitudinal thin section;
7-9) Eosiderastrea sp. 3, SNSD-MMG
SaKL542; 7) transversal thin section; 8) transversal thin section, detail; 9)
transversal thin section, detail of the microstructures;
10-12) Eosiderastrea sp. 4, BSPG 2003
XX 5846; 10) transversal thin section; 11) transversal thin section, detail;
12) longitudinal thin section.
[Scale bar 1mm]
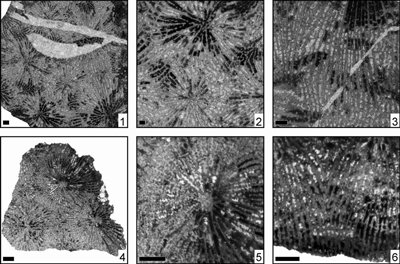
Click on thumbnail to enlarge the image.
Plate 7:
1-3) Eosiderastrea sp. 5, BSPG 2003
XX 5829; 1) transversal thin section; 2) transversal thin section, detail; 3)
longitudinal thin section;
4-6) Eosiderastrea sp. 6, SNSD-MMG
SaKL253; 4) transversal thin section; 5) transversal thin section, detail; 6)
longitudinal thin section.
[Scale bar 1mm]