◄ Carnets Geol. 16 (17) ►
![]()
Contents
[Editorial note] [Introduction]
[Secondary comments]
[Testing Hardie's (1996) model]
[Alternative stratigraphic model and its implication] and ...
[Bibliographic references]
Dépt. STU, Fac. Sci. Tech., UBO, 6 avenue Le Gorgeu, CS
93837, F-29238 Brest (France)
Department
of Ecology and Evolutionary Biology, The University of Kansas, 1200 Sunnyside
Avenue, Lawrence, Kansas 66045 (USA)
24 ch. des Champs d'Amot, F-74140 Messery (France)
Département de Géologie et Paléontologie, Université
de Genčve, 13 rue des Maraîchers, 1211 Genčve 4 (Switzerland)
Published online in final form (pdf) on August 5, 2016
[Editor:
Maurice Tucker; technical editor: Christian Emig;
language editor: Phil Salvador]
![]()
• In
this study we reiterate that there is a reliable alternative, well-documented
bio- and sequence-stratigraphic model in addition to the one used by Godet
et al.
•
Application of our model leads to different conclusions concerning stratal
correlations.
• We
question the sequence and timing of diagenetic events presented by Godet
et al.
• We
elaborate on the dissolution of calcitic ooid cortices.
• We
discuss Sandberg's and Hardie's models of calcite and aragonite
seas.
A recent paper by Godet et al. on Urgonian carbonates from the Swiss Jura concluded that diagenesis may hinder chemostratigraphic correlations of deep- to shallow-water facies. Although we agree with this conclusion we question their arguments and interpretations. These authors correctly identified diagenesis as the key factor, but we question the timing of events in their paragenetic sequence. In particular, they reported the leaching of originally calcitic oolitic cortices but failed to discuss this puzzling feature properly. In addition, the backbone of their argument is the stratigraphic correlation of a specific sequence from three dominantly shallow-water sections together with a basinal reference section. However, a reliable alternative biostratigraphic framework shows that this attempt of long-distance correlation might be erroneous.
• Paragenetic
sequence;
• Calcite
and aragonite seas;
• Chemostratigraphy;
• Carbon
isotopes;
• Early
Cretaceous.
Granier B., Clavel B. & Charollais J. (2016).- Comments on "Estimating the impact of early diagenesis on isotope records in shallow-marine carbonates: A case study from the Urgonian platform in western Swiss Jura" by A. Godet et al. [Palaeogeography Palaeoclimatology Palaeoecology 454 (2016) 125-138].- Carnets Geol., Madrid, vol. 16, no. 17, p. 417-429.
Commentaires sur "Estimating the impact of early diagenesis on isotope records in shallow-marine carbonates: A case study from the Urgonian platform in western Swiss Jura" par A. Godet et al. [Palaeogeography Palaeoclimatology Palaeoecology 454 (2016) 125-138].- Une publication récente de Godet et al. sur les carbonates urgoniens du Jura suisse concluait que la diagenčse pouvait compromettre les corrélations chimiostratigraphiques des facičs distaux vers les proximaux. Bien que nous soyons d'accord avec cette conclusion, nous nous interrogeons sur les arguments des auteurs et leurs interprétations. Ils ont bien identifié la diagénčse comme étant le facteur déterminant, mais nous mettons en doute la chronologie des événements de leur séquence paragénétique. Ils ont notamment signalé la dissolution de cortex d'ooďdes originellement calcitiques mais n'ont pas abordé sous un bon angle cet élément étonnant. De plus, leur argumentaire s'appuie sur la corrélation stratigraphique d'une séquence spécifique identifiée dans trois coupes aux facičs essentiellement peu profonds avec une coupe de bassin utilisée comme référence. Toutefois, un autre cadre biostratigraphique plus robuste que celui utilisé par Godet et al. suggčre que leur tentative de corrélations sur de longues distances est erronée.
• Séquence paragénétique ;
• Mers ŕ calcite ou ŕ
aragonite ;
• Chimiostratigraphie ;
• Isotopes du carbone ;
• Crétacé inférieur.
Editorial note |
| This 'comment manuscript' was submitted for publication in Palaeogeography, Palaeoclimatology, Palaeoecology and was rejected on the basis of the Editor's recommendation (personal communication to Bruno Granier, July 10, 2016) and the following anonymous review. We (the Editor-in-chief, Bruno Granier, and the Senior Editors, Christian Emig and Michel Moullade), at Carnets Geol., agreed to publish it because we think our readers should be informed and have the chance to reach their own opinions: |
|
"After having carefully examined the Comment by Bruno Granier
et al. on the paper by Alexis Godet et al. ("Estimating the
impact of early diagenesis on isotope records in shallow-marine
carbonates: A case study from the Urgonian platform in western Swiss
Jura") as well as the Godet et al. paper, I came to the
conclusion that the publication of this comment is not justified. The motivations for my appraisal are the following: 1. Does the Comment raise important scientific issues? The Comment of Granier et al. (hereafter GEA) is not focused on the core dataset, interpretations and conclusion raised by Godet et al. On the contrary, GEA acknowledge that the conclusion of this paper is correct. As such, this comment fails at carrying any substantial criticism of the Godet et al. study, but seems more related to another debate between the two groups of authors, namely the biostratigraphy of the Urgonian limestones in Eclepens. 2. Are the criticisms of the Godet et al. study contained in the Comment justified? There are numerous criticisms raised by Granier et al. that are of little importance, some concerning vocabulary, that are not of primordial interest for the reader. Some are even useless and off-topic such as the first one in the text. There are however two main criticisms raised by Granier et al: Firstly, about the leaching of oolitic cortices and its interpretation, secondly about the correlation of the Eclepens section with basinal reference section. For the first point, GEA bring forward original materials derived from an unpublished MSc study (Grundman, 2007) to discuss the interpretation of Godet et al. Although it might be interesting to hear further about this, it is certainly not appropriate to present and discuss these original results in a Comment of the Godet et al. paper, acknowledging that these authors couldn't be reasonably aware of this unpublished MSc thesis. Furthermore, the fact that Grundman (2007) hasn't seen some of the features observed by Godet et al in the Urgonian Jaune doesn't prove anything. The absence of evidence is not the evidence for the absence! Concerning the second point, this is absolutely not central to the discussion and conclusion of the Godet et al. paper which focuses on the Eclepens section, and for which the observation and interpretation with regards to the diagenetic history are independent of the biostratigraphic scheme used. The correlation with the basinal reference section (namely the Angles section) made by Godet et al. aims at showing that the negative d13C isotope shift observed in Eclepens has no equivalent in the Angles section, and is therefore related to diagenetic overprint. This conclusion doesn't change even if the alternative biostratigraphic scheme of GEA is applied. 3. To what extent the "alternative model" presented in the Comment represents a viable alternative stratigraphic interpretation of the study area? The alternative model presented by GEA is not about diagenetic model, but an alternative age model, derived from a different biostratigraphic interpretation, mostly based on differential appreciation of orbitolinids taxonomy between GEA and the work of Godet et al. (2010, 2011) and references therein. This part of GEA's comment is therefore not related to the Godet et al. (2016) paper s.s. The dating of the Urgonian limestone is a long-lived debate, which has so far not been settled and awaits further independent studies. GEA model however implies the absence of any major hiatus (time gap) within the Urgonian limestone, leading these authors (Clavel et al., 2014; their Fig. 9) to attribute part of the proximal Urgonian sequence as Lowstand System Tract deposits, which is at odd with conventional sequence stratigraphic schemes. This is a major weakness of GEA's stratigraphic model, questioning its viability as a robust alternative scheme. In summary, GEA comments are mostly off topic or of only minor importance, undermining any justification for their publication as a Comment in Paleo3." |
Recently, while revisiting the correlations of the Shu'aiba and Bab lithostratigraphic units in Abu Dhabi (United Arab Emirates) based on peaks and trends observed in stable isotope curves (Vahrenkamp, 1996, 2010; Grötsch et al., 1998), Granier (2012a, 2014a; Granier & Busnardo, 2013) was able to demonstrate by means of ammonite occurrences that Vahrenkamp's (1996) seminal publication had reached erroneous conclusions regarding basin to platform correlations. Thus, it would have been gratifying to read another publication concluding that "chemostratigraphy can only be applied to shallow-marine successions with great caution" (Godet et al., 2016). However, in this last case (Godet et al., 2016), the application is not valid.
In their study, Godet et al. (2016, p. 127) use a sequence stratigraphic framework with age "controlled by orbitolinid and ammonite findings" that was developed by Arnaud and Arnaud-Vanneau in the 1990s (Arnaud et al., 1998, inter alia). Godet et al. do not discuss or even mention the existence of an alternative stratigraphic framework developed by another research team (Clavel et al., 2007, 2009, 2010, 2014; Conrad et al., 2012; Charollais et al., 2013; Granier et al., 2013). Here we present a summary of the alternative biostratigraphic scheme suggesting that the B3 (Barremian no. 3) sequence(s) correlated from four sections by Godet et al. (2016: Fig. 7) might not be coeval, a point that would significantly undermine their conclusions regarding correlation and diagenesis. However, before that, we address several inconsistencies present in their publication.
First, using an irrelevant or imprecise vocabulary to describe features in sedimentary petrography can lead to confusion and ambiguities:
the title of the paper refers to "early diagenesis" and the text to "eogenesis" but it is hard to get a precise idea of how long this "early" period lasted. From their paragenetic sequence (Godet et al., 2016: Fig. 4), we understand it ends with the first phase of fracturing that is associated with a blocky calcite subzone C2. This is a smart idea as fracturing probably induced a renewal of the brine circulating within the porous network (as observed by Granier & Staffelbach, 2009, in the Paris Basin). However, the first fractures could be as early as Late Cretaceous (Eo-Alpine phase) or as late as Oligocene (Pyrenean-Provence phase), i.e., at least the duration of the Aptian and Albian stages, which is at least 25 m.y. after the end of Urgonian times. This is a rather long time-span for "early" diagenesis;
on page 127, Godet et al. (2016) state that a specific "surface is (...) heavily bored by Skolithos-like perforations". Skolithos is a tubular burrow, not a boring, indicative of a soft or firm ground. A tubular boring would likely have been Trypanites (as mentioned elsewhere in their text), indicative of a hardground (see, for instance, the recent list of ichnogenera provided by Taylor & Wilson, 2003);
on page 130, they state that "Former aragonitic bioclasts are systematically dissolved and replaced by calcite". Being metastable aragonite is commonly leached, leaving a mouldic cavity (commonly outlined by a micritic envelope) that might later be filled in part or in full by a drusy cement. These molds were cemented by calcite, but the bioclasts themselves were not "replaced by calcite". Replacement of aragonite by calcite (i.e., calcitisation) commonly results in a mosaic of crystals of a similar size, with a brownish pseudopleiochroism due to the preservation of the original organic matter of the bioclast. In the same paragraph Godet et al. (2016) point out that "centripetal subhedral growth bands indicate crystal growth in a cavity rather than progressive replacement of aragonite by calcite". In our opinion, the competitive growth of calcite crystals that results in a drusy fabric provides better primary evidence of cementation, as opposed to calcitisation;
on page 132, Godet et al. (2016) write that "The micrite-rich part of 56 whole-rock samples were microdrilled and analysed for their carbon and oxygen stable isotope composition". In her recent "critical evaluation of carbon isotope stratigraphy", Wendler (2013) described some limitations of the method. Regarding "micrite matrix", she suggested sampling mudstones or mud-supported fabrics only. However, in the current case, the log of Godet et al. (2016: Fig. 5) displays almost no mud-dominated or mud-supported fabrics, i.e., no mudstone and wackestone textures with a significant amount of micrite. On the contrary, this log displays mostly grain-dominated and grain-supported fabrics, with a limited amount of micrite in intergranular position or no micrite at all, i.e., respectively packstone and grainstone textures. Of course it is possible that their definition for packstone may cover some genuine wackestone. However, because the end results of their measurements do not differ significantly from Schootbrugge's (2001; Godet et al., 2016: Fig. 5) for the bulk rock volume of similar samples, one could legitimately question the authors' ability to pick microsamples with micrite in such unfavorable facies. Another issue is the nature of the micrite, a word that results from the contraction of "micr"-ocrystalline and calc-"ite". In shallow-water sections the main contributors to calcareous mud are benthic "calcareous" Chlorophyta, Bryopsidales and Dasycladales, that produce aragonite needles, whereas in basinal facies the main producers are various planktonic Haptophyta (coccolithophorids, Nannoconus sp., etc.). It is hard to imagine that a diagenetically modified, originally aragonitic mud (the future micrite) will have a final geochemical signature similar to a coeval low-Mg calcite ooze;
in the figure caption on page 129, Godet et al. (2016: Fig. 3.E) note "An intense micro-stylolitization (...) developed between ooids". Such features are usually referred to as intergranular pressure dissolution seams, typical of fitted fabrics with sutured grains. They represent an early stage of chemical compaction (as opposed to mechanical compaction) and they should not be called stylolites (which, by definition, are visible to the naked eye), nor even microstylolites.
Second, we question parts of the diagenetic sequence of events:
in the figure caption on page 129, Godet et al. (2016: Fig. 3.F-G) report "Dolomite rhombs with two successive subzones (D1a and D1b) developed in the intragranular [sic: the authors were probably thinking of intergranular] pore space, before precipitation of C1b and C2 blocky calcite subzones". However, in our opinion, the edges of the dolomite rhombs clearly cross cut the luminescent growth zones of their C2 calcite, suggesting that dolomitic replacement postdates this calcite cement;
on page 130, Godet et
al. (2016) write
that "Calcite dissolution first partially and locally affects oolitic
cortices. Ooids are very common in this section, especially in the F6 and
F5 microfacies associations". In an earlier publication Godet et
al. (2005) stated that "F6
microfacies with oomoldic porosity were observed by Blanc-Alétru,
so these surfaces are interpreted as sequence boundaries". Actually Blanc-Alétru
(1995) reported it from three intervals only, all sited in the Urgonien
Jaune. These intervals, which are two metres thick on average, are located
below marker discontinuities labelled D, I and H from top to bottom, and
that correspond roughly to 80 m, 66 m and 59 m on Godet et
al.'s log (2016: Fig. 5). However, in the new interpretation by Godet
et al. (2016),
none of these surfaces coincide with a sequence boundary. Furthermore, D,
the uppermost one, which marks the top of the Urgonien Jaune, is close to
a maximum flooding surface! Godet et
al. (2016: Fig. 5) add new occurrences of intervals with dissolution
of oolitic cortices, at the bottom of the Urgonien Jaune above a sequence
boundary, and at the top of both the upper and the lower Pierre Jaune
below sequence boundaries. Föllmi and Godet
(2013: Fig.
7.4, bottom right corner) illustrate one ooid with a partly dissolved
cortex from "Heterozoan [sic], oolitic carbonates" of the "Lower Pierre Jaune de Neuchâtel (early Hauterivian). Eclépens,
western Switzerland". Thanks to a MSc student,
Gaëlle Grundman (2007), we examined some one hundred thin-sections
from the working Eclépens quarry. However, in this case, ooids
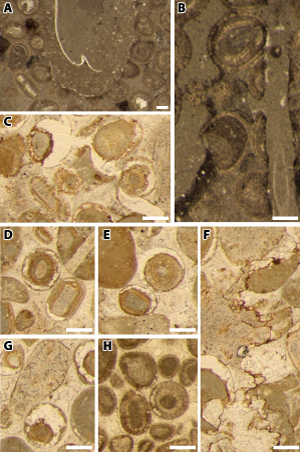
with leached cortex are found only in the upper Pierre Jaune (Fig. 1 ![]() ).
There are two options, either 1) the student failed to pick such oolitic
samples in the Urgonien Jaune, or 2) because the quarry walls are
regularly rejuvenated and material removed, leaching may be a local
phenomenon, not related to stratiform discontinuities (including possible
sequence boundaries) but to heterogeneous zones possibly determined by a
fracture or karstic network (as interpreted by Granier et
al., 2014). In the upper Pierre Jaune at Montricher, the interval with
partly leached ooids is almost 14 metres thick (Granier et
al., 2014). At Eclépens, this interval is roughly two metres thick.
It is found near the top of the upper Pierre Jaune (Fig. 1
).
There are two options, either 1) the student failed to pick such oolitic
samples in the Urgonien Jaune, or 2) because the quarry walls are
regularly rejuvenated and material removed, leaching may be a local
phenomenon, not related to stratiform discontinuities (including possible
sequence boundaries) but to heterogeneous zones possibly determined by a
fracture or karstic network (as interpreted by Granier et
al., 2014). In the upper Pierre Jaune at Montricher, the interval with
partly leached ooids is almost 14 metres thick (Granier et
al., 2014). At Eclépens, this interval is roughly two metres thick.
It is found near the top of the upper Pierre Jaune (Fig. 1 ![]() ), almost one
metre below the discontinuity, a feature that pleads in favour of the
second option;
), almost one
metre below the discontinuity, a feature that pleads in favour of the
second option;
|
|
Figure 1: A) Gastrochaenolites macroboring with the borer shell preserved; B) Trypanites mesoboring with the endolith foraminifer Bullopora tuberculata (Sollas, 1877). A-B) Thin-section EC55, hardground at the top of the Pierre Jaune; C) dissolved cortices and fractures cemented by white calcite whereas the intergranular porosity is cemented by a slightly yellowish calcite. Thin-section EC53; D-E & G) as above, but without fracture. Thin-section EC44; F) late tectonic (not stratiform) stylolite. Thin section EC53; A-G) Eclépens, upper Pierre Jaune, Hauterivian; H) dissolved cortices. Thin-section -127 m, Montricher, Urgonien Jaune, Upper Hauterivian. All scale bars = 250 µm. |
earlier records of oolitic cortices leached in full (Wherry,
1915; Sandberg, 1975,
1983; Richter, 1983; Zen et al.,
1983; Tucker, 1984; Bates & Brand,
1990; Algeo
& Watson, 1995) or in part, i.e.,
only some cortical layers (Tucker, 1984; Algeo & Watson,
1995) came to the conclusion that they are dealing respectively with
either aragonitic or "bimineralic" (aragonite-calcite) cortices.
Ancient occurrences of aragonitic ooids may be subject to interpretation. Strasser
(1986) who was more cautious suggested that the metastable mineral could
have been either aragonite or high-Mg calcite. However, he also stated
that the latter "converts faster to low-Mg calcite than aragonite
when exposed to freshwater diagenesis, without textural changes of the
affected crystals", as it is the case with high-Mg calcite echinoderm
remains accompanying our ooids. The cortices of some ooids from Eclépens
and from the neighboring locality of Montricher were partly leached.
However they correspond to a new category (Granier et
al., 2014), first because some cortical layers are not fully leached
and second because the remaining parts display a typical radial fabric
that corresponds to a primary calcite (Fig. 1 ![]() ), not to a primary
aragonite. Leaching of the calcite from oolitic cortices is a challenging
question that was not addressed by Godet
et al. (2016). There are very
few records in the literature that advocate either leaching by acidic
hydrothermal fluids, but not eogenetic dissolution by meteoric waters, or
the dual, organic and mineral nature of the ooids (Granier,
2014b; Granier
et al., 2014);
), not to a primary
aragonite. Leaching of the calcite from oolitic cortices is a challenging
question that was not addressed by Godet
et al. (2016). There are very
few records in the literature that advocate either leaching by acidic
hydrothermal fluids, but not eogenetic dissolution by meteoric waters, or
the dual, organic and mineral nature of the ooids (Granier,
2014b; Granier
et al., 2014);
on the same page (130), same paragraph, they state that "When fully
or partially preserved, their cortex shows structures indicative of an
initial precipitation of calcite, as it is expected in the context of a
calcite sea period
(Wilkinson et al., 1985; Morse
et al., 1997)". In writing this, Godet et
al. ignore the starting point of this concept and its authors, namely Sandberg
(1975) and Hardie (1996). While investigating marine ooids in order
to identify their primary mineralogy Sandberg (1975) was the first
to recognise an oscillating trend of periods dominated by aragonitic ooids
alternating with periods dominated by calcitic ooids (Sandberg,
1983; Wilkinson
et al., 1985). Hardie
(1996) used a "simple quantitative model for calculation of the
chemistry of ancient seawater based on the MOR" (mid-ocean ridge) / "RW" (river water)
"flux ratio". Best predictions were obtained
with a MOR/RW equal to 1.25 and a molar ratio Mg++/Ca++ equal to 2
(Fig. 2 ![]() ). To test this model, he
plotted the resulting curves for Ca++, Mg++, and Mg++/Ca++ ratio against the stratigraphic ranges of some
marine non-skeletal precipitates,
more specifically "aragonite cements" and "aragonite oolites".
However, the
stratigraphic inventory of both aragonitic ooids and aragonite cements is
incomplete.
). To test this model, he
plotted the resulting curves for Ca++, Mg++, and Mg++/Ca++ ratio against the stratigraphic ranges of some
marine non-skeletal precipitates,
more specifically "aragonite cements" and "aragonite oolites".
However, the
stratigraphic inventory of both aragonitic ooids and aragonite cements is
incomplete.
|
|
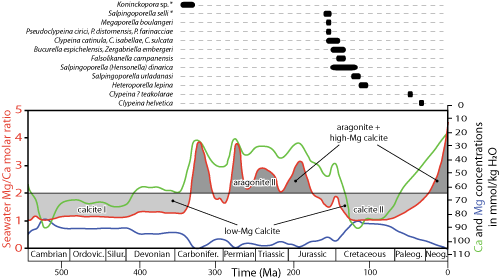
Figure 2: Stratigraphic distribution of the supposedly calcitic Dasycladales (* starred) and of the originally calcitic Dasycladales (modified from Granier, 2012b) plotted against the predicted molar Mg/Ca ratio of seawater for the Phanerozoic (adapted from Hardie, 1996). The horizontal line is the approximate divide between low-Mg calcite seas (Mg++/Ca ++ < 2) and aragonite seas (Mg++/Ca ++ > 2). |
Ooids, which are by definition non-skeletal grains, are
not entirely of "abiotic origin". There is evidence that microbial
processes are actively involved at various stages in their mineralization (Brehm
et al., 2004; Edgcomb
et al., 2013; Diaz et al.,
2013, 2014, 2015; O'Reilly et al.,
2016). For instance,
some microbes may promote mineralization of ooids that takes place in
extracellular polymeric substances (EPS) whereas others may alter the original
mineralization of ooid cortices (Duguid et al.,
2010). A parallel may be drawn with the mineralization of
the "calcareous" green algae that also takes place within an
extracellular mucilage. Accordingly Bryopsidales and Dasycladales, which are
giant unicells with
a relatively fair stratigraphic record, have been used as an alternative to
the marine ooids (and the marine cements) to test the validity of Hardie's
(1996) model. Thus, looking for some
supportive evidence, Stanley and Hardie
(1998) drew attention to
the "aragonitic dasycladaceans" which are "dominant rock
formers" in the Aragonite II interval that spans the latest Carboniferous
through to the Early Jurassic. However, that view does not match the data of
palaeophycologists. For example, Barattolo (1991: Figs. 1 & 4)
documented the scarcity of the Dasycladales at the transition from the
Triassic to the Jurassic (end of Aragonite II) and noted their abundance
during the Paleogene (end of Calcite II), in clear contradiction with Stanley
and Hardie's (1998) conclusion. Similarly Wilkinson
(1979)
provided a short summary of his views on "calcareous" green algae stating
that "Modern marine chlorophytes are exclusively aragonite" and that "Paleozoic occurrences are sufficiently rare and petrologically nondescript
that their original mineralogy cannot be determined with certainty". Stanley
and Hardie (1998) noted that Wilkinson
(1979)
"failed to
include data for (...) pre-Jurassic dasycladacean algae" and even that "some" of his data
"were in error". As a matter of fact, although
Bryopsidales never mineralized as calcite, some Dasycladalean species do, but
there are very few calcitic species in comparison to the coeval aragonitic
species (Granier, 2012b). Plotting their
stratigraphic ranges against the predicted molar Mg/Ca ratio of seawater for the Phanerozoic
(Fig. 2 ![]() )
shows that they occur both at the beginning and the end of Calcite II, but
that they are surprisingly missing at the height of this interval, which was
not "expected in the context of a calcite sea
period".
Revisiting the publication of Stanley and Hardie
(1998), Aguirre and Riding
(2005) also reached a conclusion similar to ours, noting that there is no "straightforward relationship between dasycladalean diversity and calcite-
and aragonite- sea episodes". Does this invalidate the Hardie
(1996) model? The answer must be
in the negative because, as
documented above, biological processes involved in CaCO3
mineralization may mask or prevail over physicochemical (purely abiotic)
processes.
)
shows that they occur both at the beginning and the end of Calcite II, but
that they are surprisingly missing at the height of this interval, which was
not "expected in the context of a calcite sea
period".
Revisiting the publication of Stanley and Hardie
(1998), Aguirre and Riding
(2005) also reached a conclusion similar to ours, noting that there is no "straightforward relationship between dasycladalean diversity and calcite-
and aragonite- sea episodes". Does this invalidate the Hardie
(1996) model? The answer must be
in the negative because, as
documented above, biological processes involved in CaCO3
mineralization may mask or prevail over physicochemical (purely abiotic)
processes.
As mentioned at the beginning of this comment paper, this last section will present an alternative stratigraphic model that has implications for the relative ages and geometric arrangement of the various stratal units. On page 127, Godet et al. (2016) state that "Their age is controlled by orbitolinid and ammonite findings" with orbitolinid ranges from Arnaud et al. (1998). The alternative biostratigraphic model (Clavel et al., 2002, 2007, 2009, 2010, 2014, inter alia) is based on the same fossils but the interpreted ranges of the orbitolinids differ materially. For instance, let us consider Palorbitolina lenticularis (Blumenbach, 1805), which is an emblematic, cosmopolitan orbitolinid for the Barremian - Aptian interval (Clavel et al., 2002). According to Arnaud et al. (1998) it first appears in the Sartousiana Zone of the Late Barremian but according to Clavel et al. (2014, inter alia) this species is already known from the Pulchella or even the Nicklesi Zone of the Early Barremian.
Godet
et al. (2016: Fig. 7) provide a correlation panel for their
SB B3 (Figs. 3 ![]() - 4
- 4 ![]() ), which is "close to" the Lower-Upper
Barremian boundary, as a reference horizon for four sections ranging from the
basin, on the left, to the platform, on the right (Fig. 3
), which is "close to" the Lower-Upper
Barremian boundary, as a reference horizon for four sections ranging from the
basin, on the left, to the platform, on the right (Fig. 3 ![]() ). In the first column, i.e., a composite section for
the Vocontian basin, Palorbitolina
lenticularis would theoretically first
occur some 20 metres above their SB B3 according to Arnaud's scale, but
25 metres below it according to Clavel's scale. Actually, in a
reference section of the Vocontian basin, namely L'Estellon (Drôme), the
first P. lenticularis occurs some 70 metres below the Lower-Upper
Barremian boundary in turbidites that are directly dated by ammonites of the
Pulchella Zone of the Early Barremian (Granier et
al., 2013). In the second and third columns, i.e.,
in the Gorges du Nant section and in the Cluses section, this foraminifer
first occurs some 190 and some 175 metres (Clavel et al.,
2014), respectively, above the reference horizon. Finally,
in the last column, i.e., at Eclépens,
it occurs in karstic cavity fills of the Urgonien Blanc (Godet,
2006; Clavel
et al., 2014). Actually, in the Swiss Jura,
this species is also found in a similar karstic cavity setting from La Lance (Charollais
et al., 1994; Blanc-Alétru,
1995) but it is not reported from the borehole at Montricher (Granier
et al., 2014; Jan du Chęne et al.,
2016).
). In the first column, i.e., a composite section for
the Vocontian basin, Palorbitolina
lenticularis would theoretically first
occur some 20 metres above their SB B3 according to Arnaud's scale, but
25 metres below it according to Clavel's scale. Actually, in a
reference section of the Vocontian basin, namely L'Estellon (Drôme), the
first P. lenticularis occurs some 70 metres below the Lower-Upper
Barremian boundary in turbidites that are directly dated by ammonites of the
Pulchella Zone of the Early Barremian (Granier et
al., 2013). In the second and third columns, i.e.,
in the Gorges du Nant section and in the Cluses section, this foraminifer
first occurs some 190 and some 175 metres (Clavel et al.,
2014), respectively, above the reference horizon. Finally,
in the last column, i.e., at Eclépens,
it occurs in karstic cavity fills of the Urgonien Blanc (Godet,
2006; Clavel
et al., 2014). Actually, in the Swiss Jura,
this species is also found in a similar karstic cavity setting from La Lance (Charollais
et al., 1994; Blanc-Alétru,
1995) but it is not reported from the borehole at Montricher (Granier
et al., 2014; Jan du Chęne et al.,
2016).
|
|
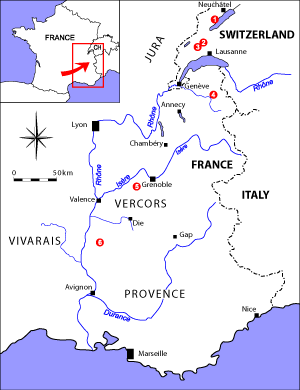
Figure 3: Location map. Swiss Jura: 1) La Lance, 2) Eclépens, 3) Montricher; Subalpine Chains: 4) Cluses; Vercors: 5) Gorges du Nant; Vocontian Basin: 6) L'Estellon. |
|
|
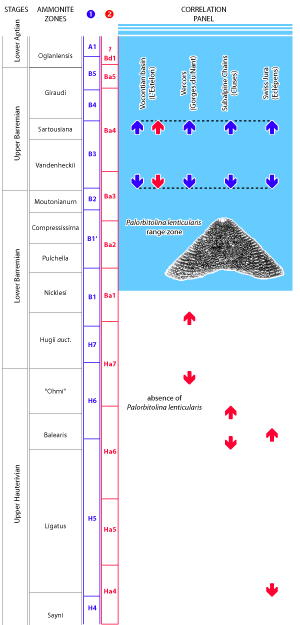
Figure 4:
Column 1) in blue colour, sequences according to Arnaud et
al. (1998) and correlation (arrows) according to Godet et
al. (2016); column 2) in red colour, sequences and holostratigraphic
(bio-, litho-, and sequence stratigraphy) correlations (arrows) according to Clavel
et al. (2014, inter alia). The light blue background corresponds to
the lower part of the Range Zone of Palorbitolina lenticularis (Blumenbach,
1805). See locations on Fig. 3 |
According
to Godet et al. (2016), their
whole sequence B3, equivalent to the upper Urgonien Jaune and the Urgonien
Blanc at Eclépens, is Late Barremian in age (Fig. 4 ![]() , in blue colour).
However, they never questioned the critical lack of the cosmopolitan Palorbitolina lenticularis in most of their
supposed Upper Barremian shallow-water facies (although it is found reworked
in coeval or even older deep-water facies, e.g.,
at L'Estellon in the Vocontian basin). On the contrary, according to Clavel
et al. (2014), this interval B3 is
Late Hauterivian in age (Fig. 4
, in blue colour).
However, they never questioned the critical lack of the cosmopolitan Palorbitolina lenticularis in most of their
supposed Upper Barremian shallow-water facies (although it is found reworked
in coeval or even older deep-water facies, e.g.,
at L'Estellon in the Vocontian basin). On the contrary, according to Clavel
et al. (2014), this interval B3 is
Late Hauterivian in age (Fig. 4 ![]() , in red colour), older than the first record
of Palorbitolina lenticularis, a simple and logical explanation for its
absence. In the Table 1 below, we summarize the stratigraphic interpretations
of the latter authors.
, in red colour), older than the first record
of Palorbitolina lenticularis, a simple and logical explanation for its
absence. In the Table 1 below, we summarize the stratigraphic interpretations
of the latter authors.
Table
1: Sequences and ages of the Godet et
al.'s B3 sequence (supposedly Late Barremian in age) according to the
alternative stratigraphic framework put forward herein (Clavel et
al., 2014). Ha for Hauterivian, Ba for Barremian, LST for Lowstand Systems
Tract, TST for Transgressive Systems Tract, HST for Highstand Systems Tract.
See locations on Fig. 3 ![]() and ranges on Fig. 4
and ranges on Fig. 4 ![]() .
.
| Sections |
Sequences
equivalent to Godet et al.'s B3 (2016) |
Corresponding ammonite zones |
Ages |
| Vocontian basin (L'Estellon) |
Ba3
HST, and Ba4 LST and TST pars |
Vandenheckii - Giraudi | Late Barremian |
| Vercors (Gorges du Nant) |
Ha7
TST and HST, and Ba1 LST pars |
"Ohmi" pars - Hugii auct. | transition of the Hauterivian to the Barremian |
| Subalpine Chains (Cluses) | Ha6 LST pars, TST and HST | Balearis - "Ohmi" pars | Late Hauterivian |
|
Swiss
Jura
(Eclépens) = Urgonien Jaune + Urgonien Blanc |
Ha4
TST and HST, Ha5 LST, TST and HST, and Ha6 LST |
Sayni pars - Balearis pars | Late Hauterivian |
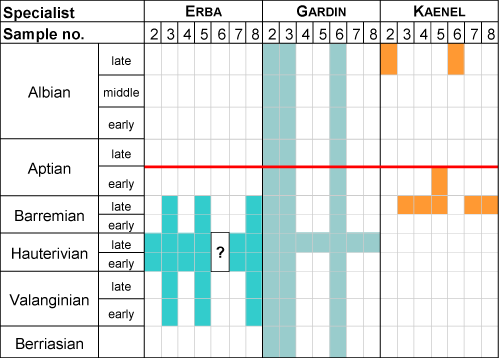
Godet et al. (2016, p. 135) use not only orbitolinids and ammonites but nannofossils. We shall refer here to a "blind test" undertaken under the supervision of Strasser (see Godet, 2006, p. 373-386). Three specialists: E. Erba, Universitŕ degli Studi di Milano, S. Gardin, Université Pierre-et-Marie-Curie, and E. de Kaenel, Consulting Geologist, examined the same set of samples from the lower part of the Urgonien Jaune, immediately below the SB B3, and their results are shown in Table 2 below.
|
Table 2: Ages of a set of seven samples based on nannofossils according to E. Erba, S. Gardin and E. de Kaenel (Godet, 2006). There are no "Urgonian" platforms in the area above the red horizon. |
|
The dating confidence is the narrower range common to the seven discrete samples: Hauterivian for Erba, Late Hauterivian for Gardin, possibly "Late Barremian" for Kaenel with two odd Late Albian dates (the studied interval cannot be younger than the Late Aptian!). Although both academic experts pointed to an Hauterivian age, Godet (2006) and Godet et al. (2016) still refer to this interval as (?) Upper Hauterivian-Lower Barremian, and the overlying interval as the Upper Barremian.
Over more than two decades our team has released a large amount of information regarding the dating of the Urgonian and coeval facies. For instance, we reported the discovery of the L'Estellon section, a "Rosetta stone" for Urgonian biostratigraphy because this section allows calibration of the first appearance of several orbitolinids directly on Barremian ammonite zones. However, against all odds (e.g., the above discussion on Palorbitolina lenticularis; Conrad et al., 2012; Charollais et al., 2013; Clavel et al., 2014; Jan du Chęne et al., 2016), some authors, e.g., Godet et al. (2016), keep on referring to the obsolete bio- and litho- stratigraphic framework of Arnaud et al. (1998).
In conclusion, although we agree with Godet et al. (2016) that diagenesis "hampers long-distance chemostratigraphic correlation", we suggest that they have failed to illustrate this with their case study because, in our opinion, they have compared non-coeval intervals; this error undermines the credibility of their research.
We gratefully acknowledge the help of Gaëlle Grundman for sharing information on her unpublished material (Grundman, 2007), the assistance of Phil Salvador in revising the English text of the original manuscript, the legal advises from Michelle Bergadaŕ, Nadine Eck and Pierre Hoffmeyer of the "Institut International de Recherche et d'Action sur la Fraude et le Plagiat Académiques". The first author (BG) also thanks Trevor Burchette and Bruce Railsback, as well as Robert Burne, Stjepko Golubic and Robert Riding, for having shared their views on the paragenetic sequence and the nature of ooids, respectively. Last but not least, we sincerely thank the editor, Maurice E. Tucker, for his insightful comments and discussion that helped to improve the original manuscript.
Aguirre J. & Riding R. (2005).- Dasycladalean algal biodiversity compared with global variations in temperature and sea level over the past 350 myr.- Palaios, Lawrence, vol. 20, p. 581-588.
Algeo T.J. & Watson B.A. (1995).- Calcite, aragonite, and bimineralic ooids in Missourian (Upper Pennsylvanian) strata of Kansas: Stratigraphic and geographic patterns of variation. In: Pausé P.H. & Candelaria M.P. (eds.), Carbonate facies and sequence stratigraphy: Practical applications of carbonate models.- Society of Economic Paleontologists and Mineralogists, Permian Basin Section, Midland, vol. 36, p. 141-173.
Arnaud H., Arnaud-Vanneau A., Blanc-Alétru M.-C., Adatte T., Argot M., Delanoy G., Thieuloy J.-P., Vermeulen J., Virgone A., Virlouvet B. & Wermeille S. (1998).- Répartition stratigraphique des orbitolinidés de la plate-forme urgonienne subalpine et jurassienne (SE de la France).- Géologie alpine, Grenoble, t. 74, p. 3-89.
Barattolo F. (1991).- Mesozoic and Cenozoic marine benthic calcareous algae with Particular regard to Mesozoic Dasycladaleans. In: Riding R. (ed.), Calcareous algae and stromatolites.- Springer-Verlag, Berlin, p. 504-540.
Bates N.R. & Brand U. (1990).- Secular variation of calcium carbonate mineralogy: An evaluation of oöid and micrite chemistries.- Geologische Rundschau, Band 79, p. 27-46.
Blanc-Alétru M.-C. (1995).- Importance des discontinuités dans l'enregistrement sédimentaire de l'Urgonien jurassien. Micropaléontologie, sédimentologie, minéralogie et stratigraphie séquentielle.- Géologie alpine, Grenoble, Mémoire HS 24, 299 p.
Brehm U., Palinska K.A. & Krumbein W.E. (2004).- Laboratory cultures of calcifying biomicrospheres generate ooids - A contribution to the origin of oolites.- Carnets Geol., Madrid, vol. 4, no. L03 (CG2004_L03), 6 p.
Charollais J., Clavel B., Granier B., Busnardo R. & Conrad M.A. (2013).- Discussion of the paper by Godet et al. 2011, entitled "Reconciling strontium-isotope and K-Ar ages with biostratigraphy: the case of the Urgonian platform, Early Cretaceous of the Jura Mountains, Western Switzerland" (Swiss Journal of Geosciences, 104, 147-160).- Swiss Journal of Geosciences, vol. 106, no. 3, p. 559-567.
Charollais J., Clavel B., Schroeder R., Busnardo R. & Horisberger P. (1994).- Mise en évidence de l'émersion post-urgonienne et de la lacune du Barrémien-Bédoulien inférieur/moyen dans le Jura neuchâtelois.- Publications du Département de Géologie et Paléontologie, Genčve, no. 14, p. 57-80.
Clavel B., Busnardo R., Charollais J., Conrad M.A. & Granier B. (2009).- Nouvelles données sur la répartition biostratigraphique des orbitolinidés ŕ l'Hauterivien supérieur, au Barrémien et ŕ l'Aptien inférieur.- Archives des Sciences, Genčve, vol. 62, p. 125-146.
Clavel B., Busnardo R., Charollais J., Conrad M.A. & Granier B. (2010).- Répartition biostratigraphique des orbitolinidés dans la biozonation ŕ ammonites (plate-forme urgonienne du Sud-Est de la France). Partie 1 : Hauterivien supérieur - Barrémien basal.- Carnets Geol., Madrid, vol. 10, no. A06 (CG2010_A06), 53 p.
Clavel B., Charollais J., Busnardo R., Granier B., Conrad M., Desjacques P. & Metzger J. (2014).- La plate-forme carbonatée urgonienne (Hauterivien supérieur - Aptien inférieur) dans le Sud-Est de la France et en Suisse : Synthčse.- Archives des Sciences, Genčve, vol. 67, no. 1, p. 1-97.
Clavel B., Charollais J., Conrad M.A., Jan du Chęne R., Busnardo R., Gardin S., Erba E., Schroeder R., Cherchi A., Decrouez D., Granier B., Sauvagnat J. & Weidmann M. (2007).- Dating and progradation of the Urgonian limestone from the Swiss Jura to South East France.- Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, Hannover, Band 158, Heft 4, p. 1025-1063.
Clavel B., Schroeder R., Charollais J., Busnardo R., Martin Closas C., Decrouez D., Sauvagnat J. & Cherchi A. (2002).- Les "Couches inférieures ŕ orbitolines" (Chaînes subalpines septentrionales) : Mythe ou réalité ?- Revue de Paléobiologie, Genčve, vol. 21, no. 2, p. 865-871.
Conrad M., Clavel B., Granier B., Charollais J., Busnardo R., Erba E., Gardin S., Jan du Chęne R., Decrouez D., Cherchi A., Schroeder R., Sauvagnat J. & Weidmann M. (2012).- "Stratigraphic, sedimentological and palaeoenvironmental constraints on the rise of the Urgonian platform in the western Swiss Jura", by A. Godet et al., Sedimentology (2010) 57, 1088-1125: Discussion.- Sedimentology, vol. 59, no. 3, p. 1121-1125.
Diaz M.R., Piggot A.M., Eberli G.P. & Klaus J.S. (2013).- Bacterial community of oolitic carbonate sediments of the Bahamas Archipelago.- Marine Ecology Progress Series, 485, p. 9-24.
Diaz M.R., Swart P.K., Eberli G.P., Oehlert A.M., Devlin Q., Saeid A. & Altabet M.A. (2015).- Geochemical evidence of microbial activity within ooids.- Sedimentology, vol. 62, p. 2090-2112.
Diaz M.R., Van Nordstrand J.D., Eberli G.P., Piggot A.M., Zhou J. & Klaus J.S. (2014).- Functional gene diversity of oolitic sands from Great Bahama Bank.- Geobiology, vol. 12, p. 231-249.
Duguid S.M.A., Kyser T.K., James N.P. & Rankey E.C. (2010).- Microbes and ooids.- Journal of Sedimentary Research, vol. 80, p. 236-251.
Edgcomb V.P., Bernhard J.M., Beaudoin D., Pruss S., Welander P.V., Schubotz F., Mehay S., Gillespie A.L. & Summons R.E. (2013).- Molecular indicators of microbial diversity in oolitic sands of Highborne Cay, Bahamas.- Geobiology, vol. 11, p. 234-251.
Föllmi K.B. & Godet A. (2013).- Palaeoceanography of Lower Cretaceous Alpine platform carbonates.- Sedimentology, vol. 60, p. 131-151.
Godet A. (2006).- The evolution of the Urgonian platform in the western Swiss Jura realm and its interactions with palaeoclimatic and palaeoceanographic change along the Northern Tethyan Margin (Hauterivian-earliest Aptian).- PhD Thesis, Université de Neuchâtel, 405 p.
Godet A., Blanc-Alétru M.-C., Bodin S., Adatte T. & Föllmi K.B. (2005).- Day 1 The Hauterivian-Barremian of the western Swiss Jura around Neuchâtel. In: Adatte T., Arnaud-Vanneau A., Arnaud H., Blanc-Alétru M.-C., Bodin S., Carrio-Schaffhauser E., Föllmi K.B., Godet A., Raddadi M.C. & Vermeulen J. (eds.), The Hauterivian-Lower Aptian sequence stratigraphy from Jura Platform to Vocontian Basin: A multidisciplinary approach.- Géologie alpine, Grenoble, Série spéciale "Colloques et Excursions" 7, p. 79-96.
Godet A., Durlet C., Spangenberg J.E. & Föllmi K.B. (2016).- Estimating the impact of early diagenesis on isotope records in shallow-marine carbonates: A case study from the Urgonian platform in western Swiss Jura.- Palæogeography, Palæoclimatology, Palæoecology, vol. 454, p. 125-138.
Granier B. (2012a).- Geochemical correlations of Shu'aiba sections: Stop trusting the trends! In: Gawlick H.J. & Missoni S. (eds.), Sedimentology in the hearth of the Alps.- CD-ROM, 29th IAS Meeting of Sedimentology, Schladming, p. 361.
Granier B. (2012b).- The contribution of calcareous green algae to the production of limestones: a review. In: Basso D. & Granier B. (eds.), Calcareous algae and the global change: from identification to quantification.- Geodiversitas, Paris, vol. 34, no. 1, p. 35-60.
Granier B. (2014a).- Comment on "Early Aptian paleoenvironmental evolution of the Bab Basin at the southern Neo-Tethys margin: Response to global-carbon cycle perturbations across Ocean Anoxic Event 1a" by K. Yamamoto et al.- Geochemistry, Geophysics, Geosystems, vol. 15, no. 5, p. 2086-2090.
Granier B. (2014b).- Borings and etchings in the Upper Bathonian-Lower Callovian oolite of the Paris Basin (France).- Carnets Geol., Madrid, vol. 14, no. 21, p. 461-469.
Granier B., Barbin V. & Charollais J. (2014).- Significance of partial leaching in calcareous ooids: The case study of Hauterivian oolites in Switzerland.- Carnets Geol., Madrid, vol. 14, no. 22, p. 471-481.
Granier B. & Busnardo R. (2013).- New stratigraphic data on the Aptian of the Persian Gulf. In: Skelton P., Granier B. & Moullade M. (eds.), Special issue: Spatial patterns of change in Aptian carbonate platforms and related events.- Cretaceous Research, vol. 39, p. 170-182.
Granier B., Clavel B., Moullade M., Busnardo R., Charollais J., Tronchetti G. & Desjacques P. (2013).- L'Estellon (Baronnies, France), a "Rosetta Stone" for the Urgonian biostratigraphy.- Carnets Geol., Madrid, vol. 13, no. A04 (CG2013_A04), p. 163-207.
Granier B. & Staffelbach C. (2009).- Quick look cathodoluminescence analyses and their impact on the interpretation of carbonate reservoirs. Case study of mid-Jurassic oolitic reservoirs in the Paris Basin.- Carnets Geol., Madrid, vol. 9, no. A07 (CG2009_A07), 14 p.
Grötsch J., Billing I. & Vahrenkamp V.C. (1998).- Carbon-isotope stratigraphy in shallow-water carbonates: implications for Cretaceous black-shale deposition.- Sedimentology, vol. 45, no. 4, p. 623-634.
Grundman G. (2007).- Étude sédimentologique et stratigraphique de la carričre d'Eclépens (Vaud, Suisse, Crétacé inférieur).- MSc Thesis, Université Pierre et Marie Curie, Paris, 46 p.
Hardie L.A. (1996).- Secular variation in seawater chemistry: an explanation for the coupled secular variation in the mineralogies of marine limestones and potash evaporites over the past 600 m.y.- Geology, vol. 24, p. 279-283.
Jan du Chęne R., Monteil É., Charollais J., Granier B. & Clavel B. (2016).- Étude palynologique du sondage du Morand (commune de Montricher, canton de Vaud, Suisse). Lithologie, inventaire paléontologique et biostratigraphie du Crétacé inférieur du Jura vaudois.- Archives des Sciences, Genčve, vol. 68, p. 117-162.
Morse J.W., Wang Q., Tsio M.Y. (1997).- Influences of temperature and Mg:Ca ratio on CaCO3 precipitates from seawater.- Geology, vol. 25, no. 1, p. 85-87.
O'Reilly S.S., Mariotti G., Winter A.R., Newman S.A., Matys E.D., McDermott F., Pruss S.B., Bosak T., Summons R.E. & Klepac-Ceraj V. (2016).- Molecular biosignatures reveal common benthic microbial sources of organic matter in ooids and grapestones from Pigeon Cay, The Bahamas.- Geobiology. doi:10.1111/gbi.12196
Richter D.K. (1983).- II.1 Calcareous ooids: A synopsis. In: Peryt T.M. (ed.), Coated grains.- Springer-Verlag, Berlin, p. 71-99.
Sandberg P.A. (1975).- New interpretations of Great Salt Lake ooids and of ancient non-skeletal carbonate mineralogy.- Sedimentology, vol. 22, p. 497-537.
Sandberg P.A. (1983).- An oscillating trend in Phanerozoic non-skeletal carbonate mineralogy.- Nature, vol. 305, p. 19-22.
Schootbrugge B. van de (2001).- Influence of paleo-environmental changes during the Hauterivian (early Cretaceous) on carbonate deposition along the northern margin of the Tethys: Evidence from geochemical records (C, O and Sr-isotopes, P, Fe, Mn).- Ph.D. thesis, Institut de Géologie, Université de Neuchâtel, Switzerland, 291 p.
Stanley S.M. & Hardie L.A. (1998).- Secular oscillations in the carbonate mineralogy of reef-building and sediment-producing organisms driven by tectonically forced shifts in seawater chemistry.- Palæogeography, Palæoclimatology, Palæoecology, vol. 144, p. 3-19.
Strasser A. (1986).- Ooids in Purbeck limestones (lowermost Cretaceous) of the Swiss and French Jura.- Sedimentology, vol. 33, no. 5, p. 711-727.
Taylor P.D. & Wilson M.A. (2003).- Palaeoecology and evolution of marine hard substrate communities.- Earth-Science Reviews, vol. 62, p. 1-103.
Tucker M.E. (1984).- Calcitic, aragonitic and mixed calcitic-aragonitic ooids from the mid-Proterozoic Belt Supergroup, Montana.- Sedimentology, vol. 31, no. 5, p. 627-644.
Vahrenkamp V.C. (1996).- Chemostratigraphy on the Lower Cretaceous Shu'aiba Formation: a δ13C reference profile for the Aptian stage from the southern Neo-Tethys Ocean.- American Association of Petroleum Geologists, Bulletin, vol. 80, no. 5, p. 647-662.
Vahrenkamp V.C. (2010).- Chemostratigraphy on the Lower Cretaceous Shu'aiba Formation: a δ13C reference profile for the Aptian stage from the southern Neo-Tethys Ocean.- GeoArabia Special Publication 4, vol. 1, p. 107-137.
Wendler I. (2013).- A critical evaluation of carbon isotope stratigraphy and biostratigraphic implications for Late Cretaceous global correlations.- Earth-Science Reviews, vol. 126, p. 116-146.
Wherry E.T. (1915).- A peculiar oolite from Bethlehem, Pennsylvania.- Proceedings of the United States National Museum, vol. 49, no. 2012, p. 153-156 (Pls. 40-41).
Wilkinson B.H. (1979).- Biomineralization, paleoceanography, and the evolution of calcareous marine organisms.- Geology, vol. 7, p. 524-527.
Wilkinson B.H., Owen R.M. Carroll A.R. (1985).- Submarine hydrothermal weathering, global eustasy, and carbonate polymorphism in Phanerozoic marine oolites.- Journal of Sedimentary Petrology, vol. 55, p. 171-183.
Zen Y.-F., Lee N.H. & Huang Y.Z. (1983).- II.10 Sedimentary characteristics of oolitic carbonates from the Jialing-Jiang Formation [Lower Triassic (T12J1)], South Sichuan Basin, China. In: Peryt T.M. (ed.), Coated grains.- Springer-Verlag, Berlin, p. 176-187.