◄ Carnets Geol. 16 (20) ►
![]()
Contents
[1. Introduction]
[2. Geological framework]
[3. Materials and methods]
[4. Lithostratigraphy]
[5. Micropaleontology]
[6. Biostratigraphy of the Upper Jurassic-Lower Cretaceous transition]
[7. Paleoenvironmental implications of benthic foraminifera and calcareous algae]
[8. Conclusions] and ...
[Bibliographic references]
Babeş-Bolyai University, Department of Geology and Center for Integrated Geological Studies, M. Kogălniceanu str., 1, 400084 Cluj-Napoca (Romania)
Babeş-Bolyai University, Department of Geology and Center for Integrated Geological Studies, M. Kogălniceanu str., 1, 400084 Cluj-Napoca (Romania)
Babeş-Bolyai
University, Department of Geology and Center for Integrated Geological Studies,
M. Kogălniceanu str., 1, 400084 Cluj-Napoca (Romania)
Dépt. STU, Fac. Sci.
Tech., UBO, 6 avenue Le Gorgeu, CS 93837, F-29238 Brest (France)
Department of Ecology and Evolutionary Biology, The
University of Kansas, 1200 Sunnyside Avenue, Lawrence, Kansas 66045 (USA)
Published online in final form (pdf) on October 24, 2016
[Editor:
Robert W. Scott; technical editor: Bruno Granier]
![]()
The carbonate succession of the Piatra Craiului Massif, i.e., the eastern part of the Getic carbonate platform (Southern Carpathians, Romania), comprises reefal and peritidal limestones that contain a diverse microfossil assemblage composed mainly of calcareous green algae and benthic foraminifera. The biostratigraphically most significant benthic foraminifera found in the several sections studied are described; some (Anchispirocyclina lusitanica, Neokilianina rahonensis, Bramkampella arabica, Everticyclammina praekelleri) are reported for the first time in this area. Hence, assemblages of both foraminifera and calcareous algae characterize three biostratigraphic intervals in the studied succession, the Kimmeridgian - lower Tithonian, the upper Tithonian - lower Berriasian, and the upper Berriasian - ? lower Valanginian intervals. The main microfacies types (bioclastic rudstone, coral-microbial boundstone, bioclastic grainstone, interbedded mudstones and wackestones with cyanobacteria nodules) as well as both the foraminifera and the calcareous algae are paleoecological indicators that may contribute to the decipherment of the depositional environments and to building a depositional model for the eastern part of the Getic carbonate platform at the Late Jurassic-Early Cretaceous transition.
• Upper Jurassic;
• Lower Cretaceous;
• Getic carbonate platform;
• Southern
Carpathians;
• biostratigraphy;
• foraminifera;
• paleoenvironment.
Mircescu C.V., Pleş G., Bucur I.I. & Granier B. (2016).- Jurassic-Cretaceous transition on the Getic carbonate platform (Southern Carpathians, Romania): Benthic foraminifera and algae.- Carnets Geol., Madrid, vol. 16, no. 20, p. 491-512.
Passage Jurassique-Crétacé sur la plate-forme carbonatée gétique (Carpathes méridionales, Roumanie) : Foraminifčres et algues benthiques.- La série carbonatée du Massif de Piatra Craiului, c'est-ŕ-dire de la partie orientale de la plate-forme carbonatée gétique (Carpathes méridionales, Roumanie), est constituée de calcaires récifaux et péritidaux qui recčlent des associations variées de microfossiles constituées d'algues vertes calcaires et de foraminifčres benthiques essentiellement. Parmi les foraminifčres benthiques identifiés dans les nombreuses coupes étudiées, nous décrivons ceux auxquels on attribue une certaine valeur biostratigraphique. Quelques-uns (Anchispirocyclina lusitanica, Neokilianina rahonensis, Bramkampella arabica, Everticyclammina praekelleri) sont signalés pour la premičre fois dans ce secteur. De ce fait, dans la série étudiée, grâce aux associations combinées de foraminifčres et d'algues calcaires, nous caractérisons trois intervalles biostratigraphiques : le Kimméridgien - Tithonien inférieur, le Tithonien supérieur - Berriasien inférieur et le Berriasien supérieur - ? Valanginien inférieur. Les principaux types de microfacičs (rudstone bioclastique, boundstone corallien-microbien, grainstone bioclastique, alternances de mudstones et de wackestones ŕ nodules cyanobactériens) sont des indicateurs paléoécologiques qui, avec les foraminifčres et les algues calcaires, peuvent contribuer au décryptage des environnements de dépôt et ŕ la construction d'un modčle dépositionnel pour la partie orientale de la plate-forme carbonatée gétique ŕ la transition du Jurassique supérieur au Crétacé inférieur.
• Jurassique supérieur ;
• Crétacé inférieur ;
• plate-forme carbonatée gétique ;
• Carpathes méridionales ;
• biostratigraphie ;
• foraminifčres ;
• paléoenvironnement.
This paper is part of a larger integrated study that aims to describe the geological evolution of the Piatra Craiului Massif by using sedimentological and stratigraphic techniques. The geomorphological characteristics of the Piatra Craiului Massif with extensive outcrops represent an optimal feature for studying the entire carbonate succession.
Several sections comprising the Jurassic-Cretaceous transition were studied. Earlier authors (Jekelius, 1923; Oncescu, 1943; Popescu, 1966) assigned a Late Jurassic age to the studied limestones. However, other biostratigraphic studies (Bucur, 1978; Patrulius et al., 1980; Bucur et al., 2009; Dragastan, 2010) revealed the presence of Lower Cretaceous deposits in the upper part of the carbonate succession. The present paper brings new biostratigraphical information regarding the Upper Jurassic-Lower Cretaceous transition in the studied area. Three biostratigraphic intervals (Kimmeridgian-lower Tithonian, upper Tithonian-lower Berriasian and upper Berriasian-lower Valanginian) are identified.
The
Piatra Craiului Massif forms the western flank of a larger syncline unit, which
was defined in the geological literature as the Piatra Craiului Syncline. This
structural unit is considered an integral part of the Dâmbovicioara Couloir (Patrulius,
1969) (Fig. 1 ![]() ) and it represents the eastern part of the Getic Carbonate
Platform (Patrulius, 1976).
) and it represents the eastern part of the Getic Carbonate
Platform (Patrulius, 1976).
|
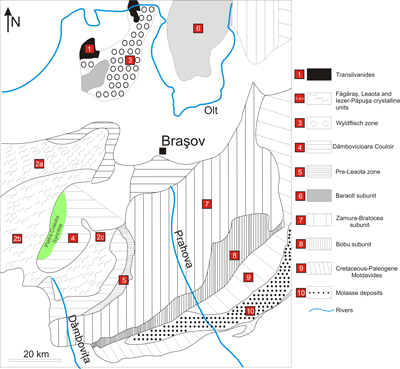
Figure 1: Paleogeographic map indicating the position of the Piatra Craiului Syncline unit within the Dâmbovicioara Couloir (modified from Patrulius, 1969). |
The lower part of the
sedimentary succession is Bajocian-early Callovian in age
(Fig. 2 ![]() ). It is
characterized by the upward transition from sandstones and marly limestones to
limestones and radiolarites (Popescu, 1966). These deposits contain
ammonite fragments and pelecypods [e.g., Bositra buchi (Roemer, 1836)]. Patrulius
(1969) and Grădinaru
(2011) described similar Bajocian deposits from the Dâmbovicioara Couloir. The
age ascription is mainly based on pelecypod fragments, gastropods and solitary
zoantharians. In the Bajocian-lower Callovian succession hardground levels and
condensed intervals rich in macro-oncoids and stromatolitic structures are
present (Lazăr & Grădinaru,
2013). Upper
Callovian-Oxfordian radiolarites overlie the Bajocian-lower Callovian succession
(Fig. 2
). It is
characterized by the upward transition from sandstones and marly limestones to
limestones and radiolarites (Popescu, 1966). These deposits contain
ammonite fragments and pelecypods [e.g., Bositra buchi (Roemer, 1836)]. Patrulius
(1969) and Grădinaru
(2011) described similar Bajocian deposits from the Dâmbovicioara Couloir. The
age ascription is mainly based on pelecypod fragments, gastropods and solitary
zoantharians. In the Bajocian-lower Callovian succession hardground levels and
condensed intervals rich in macro-oncoids and stromatolitic structures are
present (Lazăr & Grădinaru,
2013). Upper
Callovian-Oxfordian radiolarites overlie the Bajocian-lower Callovian succession
(Fig. 2 ![]() ). Detailed information about these deposits can be found in Bucur
(1980), Mészáros and Bucur
(1980), and Beccaro and Lazăr
(2007).
). Detailed information about these deposits can be found in Bucur
(1980), Mészáros and Bucur
(1980), and Beccaro and Lazăr
(2007).
The mid-Jurassic deposits
are in turn overlain by thick units of Kimmeridgian - Berriasian (- ? lower
Valanginian) shallow-water carbonates, i.e., Štramberk-like limestones.
They reach a total thickness of 1200 m (Bucur et
al., 2011; Pleş et al.,
2013) near the La Om summit
(Fig. 2 ![]() ). This succession represents an overall
shallowing-upward megasequence that is defined by the shift from coral-microbial
bioconstructions to peritidal carbonates (Mircescu et
al., 2014). Bucur (1978) first reported the presence of Berriasian
deposits in the northern part of the Piatra Craiului Massif. Recent studies (Bucur
et al., 2009; Săsăran
et al., 2013; Pleş et
al., 2013; Mircescu et al.,
2014) have confirmed the presence of lowermost Cretaceous deposits in that area
(Fig. 2
). This succession represents an overall
shallowing-upward megasequence that is defined by the shift from coral-microbial
bioconstructions to peritidal carbonates (Mircescu et
al., 2014). Bucur (1978) first reported the presence of Berriasian
deposits in the northern part of the Piatra Craiului Massif. Recent studies (Bucur
et al., 2009; Săsăran
et al., 2013; Pleş et
al., 2013; Mircescu et al.,
2014) have confirmed the presence of lowermost Cretaceous deposits in that area
(Fig. 2 ![]() ). Overlying these Štramberk limestones are either Barremian-Aptian breccias/conglomerates (Ungureanu
et al., 2015) or the Dâmbovicioara Formation the lowermost transgressive strata which are
Early Valanginian in age (Grădinaru et al., 2016).
). Overlying these Štramberk limestones are either Barremian-Aptian breccias/conglomerates (Ungureanu
et al., 2015) or the Dâmbovicioara Formation the lowermost transgressive strata which are
Early Valanginian in age (Grădinaru et al., 2016).
|
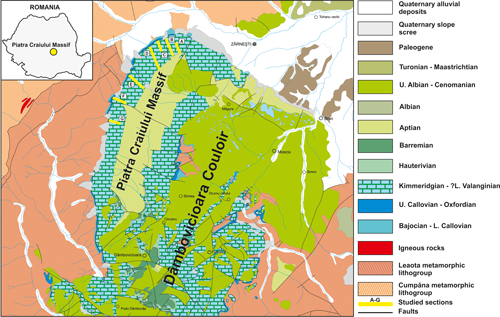
Figure 2: Location of studied sections on the geological map of the Piatra Craiului Massif (modified from Dimitrescu et al., 1971, 1974; Patrulius et al., 1971; Săndulescu et al., 1972) (A-Curmătura-Turnu; B-Padina Închisă-Drumul lui Lehmann; C-Padina Popii; D-Ciorânga Mare-Vf. Ascuţit-Padinile Frumoase; E-Vlăduşca de Vest-Vlăduşca de Est; F-Zaplaz-Lanţuri; G-Padina Lăncii). |
Seven sections were
analysed for microfacies and micropaleontological content (A-G)
(Fig. 2 ![]() ): Curmătura-Turnu
(A), Padina Închisă-Drumul lui Lehmann (B), Padina Popii (C), Ciorânga
Mare-Vf. Ascuţit-Padinile Frumoase (D), Vlăduşca de Vest-Vlăduşca
de Est (E), Zaplaz-Lanţuri (F), Padina Lăncii (G). The total thickness
of the sampled sections ranges from 700 m in the north to approximately 1200 m
in the central part of the massif. Each section is defined by a gradual
transition from massive limestones in their lower part towards
metre/decimeter-thick carbonate beds in their upper part. Sections A-E were
sampled at a resolution of 4 to 5 m and sections F and G were sampled at a
resolution of 7 to 8 m. Nine hundred samples were evaluated for their
microfacies and microfossil content. However, 120 samples were specifically used
to describe the microfossil assemblages (Fig. 3
): Curmătura-Turnu
(A), Padina Închisă-Drumul lui Lehmann (B), Padina Popii (C), Ciorânga
Mare-Vf. Ascuţit-Padinile Frumoase (D), Vlăduşca de Vest-Vlăduşca
de Est (E), Zaplaz-Lanţuri (F), Padina Lăncii (G). The total thickness
of the sampled sections ranges from 700 m in the north to approximately 1200 m
in the central part of the massif. Each section is defined by a gradual
transition from massive limestones in their lower part towards
metre/decimeter-thick carbonate beds in their upper part. Sections A-E were
sampled at a resolution of 4 to 5 m and sections F and G were sampled at a
resolution of 7 to 8 m. Nine hundred samples were evaluated for their
microfacies and microfossil content. However, 120 samples were specifically used
to describe the microfossil assemblages (Fig. 3 ![]() ). These sections were correlated
by analysing repetitive patterns of microfossil content and microfacies
characteristics. As a result, a composite bio-lithostratigraphic table was
generated (Fig. 3
). These sections were correlated
by analysing repetitive patterns of microfossil content and microfacies
characteristics. As a result, a composite bio-lithostratigraphic table was
generated (Fig. 3 ![]() ).
).
Remark: In Tethyan basinal sections stratigraphers refer to two-fold divisions of both the Kimmeridgian and the Valanginian and three-fold divisions of both the Tithonian and the Berriasian based of ammonites. Because there is no record of ammonite find in the studied shallow-water limestones, we lack direct calibration to the ammonite zones. Therefore, we do not refer to the formal subdivisions of the above stages into substages (that come with the qualifying labels: Lower/Early, Middle and Upper/Late) but to informal two-fold subdvisions, i.e., subdivisions into "lower/early" and "upper/late" standing for "lower/earlier part of" and "upper/later part of", respectively.
|
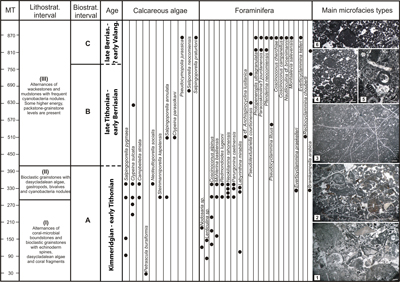
Figure 3: Lithostratigraphic and micropaleontological characteristics of the carbonate succession from the Piatra Craiului Massif [1-Bioclastic rudstone with coral fragments, echinoderm spines, dasycladalean algae (Salpingoporella pygmaea) and encrusting organisms (Crescentiella morronensis); 2-Coarse bioclastic intraclastic grainstone with cyanobacteria nodules, dasycladalean algae (Neoteutloporella socialis; Campbeliella striata) and gastropods. Black pebbles consist of blackened cyanobacteria nodules; 3-Peloidal fenestral packstone with cyanobacteria nodules; 4-Peloidal intraclastic grainstone with cyanobacteria nodules and angular/subangular micritic intraclasts; 5-Peloidal bioclastic intraclastic grainstone. Bioclasts: foraminifera (Bramkampella arabica), dasycladalean algae (Pseudocymopolia jurassica); 6-Peloidal grainstone with cyanobacteria nodules] (Scale bar: 1 mm). |
Sedimentological and
textural features were used to define three lithostratigraphic intervals
labelled I to III from bottom to top of the studied succession (Fig. 3 ![]() ).
).
Lithostratigraphic interval I comprises the lowermost 290 m of the
carbonate succesion
(Fig. 3 ![]() ). This unit consists of alternating coral-microbial
boundstones and bioclastic intraclastic rudstones. The rudstone levels contain
encrusting organisms [Crescentiella
morronensis (Crescenti, 1969)], dasycladalean algae and echinoderm
fragments (Fig. 3
). This unit consists of alternating coral-microbial
boundstones and bioclastic intraclastic rudstones. The rudstone levels contain
encrusting organisms [Crescentiella
morronensis (Crescenti, 1969)], dasycladalean algae and echinoderm
fragments (Fig. 3 ![]() ). Corals are heavily encrusted by Lithocodium/Bacinella-type
structures. The sedimentology of this lithostratigraphic interval is fully
documented by Pleş et al.
(2013).
). Corals are heavily encrusted by Lithocodium/Bacinella-type
structures. The sedimentology of this lithostratigraphic interval is fully
documented by Pleş et al.
(2013).
Lithostratigraphic interval II includes the entire package of carbonate
deposits between 290 and 408 meters of stratigraphic thickness (Fig. 3 ![]() ). The
main facies type is coarse bio-intraclastic grainstones with gastropods,
dasycladalean algae, sponges, echinoderm fragments, and foraminifera (Fig. 3
). The
main facies type is coarse bio-intraclastic grainstones with gastropods,
dasycladalean algae, sponges, echinoderm fragments, and foraminifera (Fig. 3 ![]() ).
Intraclasts are represented at some levels by various-sized black pebbles (mm to
cm). Some have a brecciated structure consisisting of blackened bioclasts
encased in a muddy matrix which is pigmented with organic matter. In some cases,
they consist of darkened bioclasts (cyanobacteria nodules, dasycladalean algae).
).
Intraclasts are represented at some levels by various-sized black pebbles (mm to
cm). Some have a brecciated structure consisisting of blackened bioclasts
encased in a muddy matrix which is pigmented with organic matter. In some cases,
they consist of darkened bioclasts (cyanobacteria nodules, dasycladalean algae).
Lithostratigraphic interval III comprises the middle and upper parts of
the carbonate deposits from the Piatra Craiului Massif (between 408 and 880 m) (Fig. 3 ![]() ). Peloidal wackestone-packstone facies (Fig. 3
). Peloidal wackestone-packstone facies (Fig. 3 ![]() )
alternate with
homogeneous mudstones with cyanobacteria and floatstones of cyanobacteria
nodules. However, some levels of bioclastic packstone/grainstone with
dasycladalean algae and foraminifera were also identified in the uppermost part
of this interval (Fig. 3
)
alternate with
homogeneous mudstones with cyanobacteria and floatstones of cyanobacteria
nodules. However, some levels of bioclastic packstone/grainstone with
dasycladalean algae and foraminifera were also identified in the uppermost part
of this interval (Fig. 3 ![]() ).
).
This
chapter focuses on the systematic description of the most abundant and
biostratigraphically important foraminiferal species, identified in 120 samples.
Apart from these, other associated species are represented including Nodosaria sp. (Fig. 3 ![]() ), Lenticulina sp. (Fig. 3
), Lenticulina sp. (Fig. 3 ![]() ), Lituola
baculiformis Schlagintweit & Gawlick, 2007 (Fig.
4.E
), Lituola
baculiformis Schlagintweit & Gawlick, 2007 (Fig.
4.E ![]() ), Frentzenella
involuta (Mantsurova & Gorbatchik, 1982) (Fig.
4.B
), Frentzenella
involuta (Mantsurova & Gorbatchik, 1982) (Fig.
4.B ![]() ), Bulbobaculites
sp. (Fig. 5.A
), Bulbobaculites
sp. (Fig. 5.A ![]() ), Scythiolina sp. (Fig.
6.J
), Scythiolina sp. (Fig.
6.J ![]() ), and Freixialina planispiralis
Ramalho, 1969 (Fig. 6.O
), and Freixialina planispiralis
Ramalho, 1969 (Fig. 6.O ![]() ). Taxonomy of benthic foraminifera follows Septfontaine
(1988), Rigaud et
al. (2013) and Kaminski (2014). Taxonomy of calcareous algae follows that of Bassoullet et
al. (1978).
). Taxonomy of benthic foraminifera follows Septfontaine
(1988), Rigaud et
al. (2013) and Kaminski (2014). Taxonomy of calcareous algae follows that of Bassoullet et
al. (1978).
|
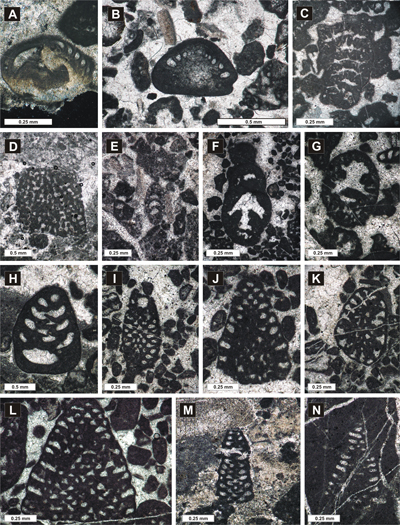
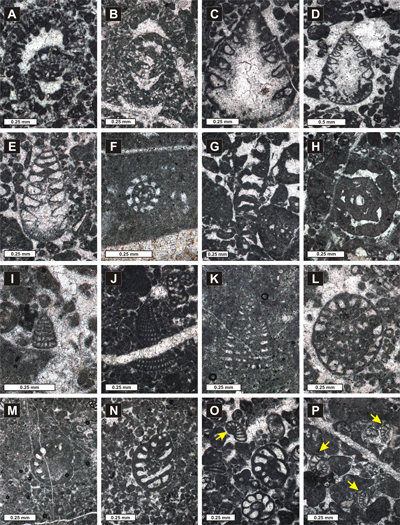
Figure 4: Foraminiferal assemblage identified in biostratigraphic interval A (A: Mohlerina basiliensis; B: Frentzenella involuta; C: Bramkampella arabica; D: Labyrinthina mirabilis; E: Lituola baculiformis; F-G: Everticyclamina praekelleri; H: Redmondoides lugeoni; I-L: Neokilianina rahonensis; M-N: Parurgonina caelinensis) (A: Sample 216, Vlăduşca de Vest-Vlăduşca de Est section; B: Sample 17, Zaplaz-Lanţuri section; C: Sample 92, Padina Popii section; D: Sample 743, Padina Lăncii section; E: Sample 18, Zaplaz-Lanţuri section; F: Sample 634, Zaplaz-Lanţuri section; H: Sample 630, Zaplaz-Lanţuri section; I-J, L: Sample 633, Zaplaz-Lanţuri section; K: Sample 628, Zaplaz-Lanţuri section; M: Sample 650, Zaplaz-Lanţuri section; N: Sample 652, Zaplaz-Lanţuri section). |
|
Figure 5: Foraminiferal assemblage identified in biostratigraphic interval B (A: Bulbobaculites sp.; B: Pseudocyclammina lituus; C: Pseudotextulariella courtionensis; D: Rectocyclammina sp.; E-F: cf. Anchispirocyclina lusitanica) (A: Sample 394, Padina Închisă-Drumul lui Lehmann section; B: Sample 269, Vlăduşca de Vest-Vlăduşca de Est section; C: Sample 464, Padina Închisă-Drumul lui Lehmann section; D: Sample 464, Padina Închisă-Drumul lui Lehmann section; E-F: Sample 46, Zaplaz-Lanţuri section). |
|
Figure 6: Foraminiferal assemblage identified in biostratigraphic interval C [(A-B: Pseudocyclammina lituus; C-D: Coscinoconus campanellus; E: Coscinoconus cherchiae; F: Nautiloculina cf. broennimanni; G: Rectocyclammina sp.; H: Everticyclammina kelleri; I: Montsalevia salevensis; J: Scythiolina sp.; K-L: Paracoskinolina ? jourdanensis; M-O: Pfenderina neocomiensis; O: Freixialina planispiralis (arrow); P: Protopeneroplis ultragranulata (arrows)] (A: Sample 11828, Ciorânga Mare-Vf. Ascuţit-Padinile Frumoase section; B: Sample 11879, Ciorânga Mare-Vf. Ascuţit-Padinile Frumoase section; C-D: Sample 11867, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; E: Sample 11868, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; F: Sample 11821, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; G: Sample 11843, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; H: Sample 11867, Zaplaz-Lanţuri section; Sample 11867, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; I: Sample 11867, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; J: Sample 287, Vlăduşca de Vest-Vlăduşca de Est section; K: Sample 331, Vlăduşca de Vest-Vlăduşca de Est section; L: Sample 11830, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; M: Sample 9, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; N: Sample 11830, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; O: Sample 12831, Ciorânga Mare- Vf. Ascuţit -Padinile Frumoase section; P: Sample 411, Padina Închisă-Drumul lui Lehmann section). |
Class Foraminifera Orbigny, 1826
Order Lituolida Lankester, 1885
Suborder Nezzazatina Kaminski, 2004
Superfamily Nezzazatoidea Hamaoui & Saint-Marc, 1970
Family Nautiloculinidae Loeblich & Tappan, 1985
Genus Nautiloculina Mohler, 1938
Nautiloculina cf. broennimanni Arnaud-Vanneau & Peybernčs, 1978
(Fig. 6.F ![]() )
)
1978 - Nautiloculina broennimanni n. sp. - Arnaud-Vanneau & Peybernčs, p. 81, Pl. 2, figs. 4-11.
1991 - Nautiloculina broennimanni Arnaud-Vanneau & Peybernčs, 1978 - Altiner, p. 194, Pl. 7, figs. 15-16.
1997 - Nautiloculina broennimanni Arnaud-Vanneau & Peybernčs, 1978 - Kołodziej & Decrouez, p. 149, Pl. 1, fig. 3.
Description: The test is lenticular in shape, planispirally coiled, made of agglutinated carbonate particles. The inner structure is simple, characterized by numerous chambers that expand in a very short distance from the proloculus to the end of the last whorl. An additional micritic layer coats the septa as a new chamber develops. The aperture is equatorial/low interiomarginal.
Remarks: N. broennimanni differs from Charentia cuvillieri in not having a developed pseudoalveolar layer and canaliculated wall structures. The identified specimens are smaller than the original specimens described by Arnaud-Vanneau & Peybernčs, 1978.
Stratigraphic range: Tithonian-?Albian.
Order Loftusiida Kaminski & Mikhalevic, 2004
Suborder Loftusiina Kaminski & Mikhalevic, 2004
Superfamily Loftusioidea Brady, 1884
Family Mesoendothyridae Voloshinova, 1958
Subfamily Labyrinthininae Septfontaine, 1988
Genus Labyrinthina Weynschenk, 1951
Labyrinthina mirabilis Weynschenk, 1951
(Fig. 4.D ![]() )
)
1951 - Labyrinthina mirabilis n. sp. - Weynschenk, p. 798, Pl. 112, figs. 4, 6-7, 9.
2005 - Labyrinthina mirabilis Weynschenk, 1951 - Schlagintweit et al., p. 31, Fig. 13.a-b.
Description: The test is characterized by a planispiral development juvenile stage and an uncoiled rectilinear adult stage. Wall is fine agglutinated, imperforated. The interseptal pillars and vertical beams of the uncoiled part form a particular labyrinthic inner structure, best observed in longitudinally sectioned specimens. Aperture is simple in early ontogenetic stages, later becoming multiple.
Stratigraphic range: uppermost Oxfordian-lower Tithonian.
Family Everticyclamminidae Septfontaine, 1988
Genus Everticyclammina Redmond, 1964
Everticyclammina praekelleri Banner & Highton, 1990
(Fig. 4.F-G ![]() )
)
1990 - Everticyclammina praekelleri n. sp. - Banner & Highton, p. 8, 10; Pl. 1, fig. 1; Pl. 3, fig. 5; Pl. 4, figs. 1-11.
2015 - Everticyclammina praekelleri Banner & Highton, 1990 - Pleş et al., p. 46, Fig. 3.f-h.
Description: The test is planispirally coiled in early ontogenetic stages, later starting to uncoil reaching terminal rectilinearity in most specimens. Wall is alveolar, imperforate made of agglutinated carbonate grains. In the adult stage, the hypodermis is characterized by the presence of elongated broadened alveoli in the posterior-lateral area and widely spaced alveoli in the anterior-peripheral area. The aperture is single, terminal.
Remarks: The differences between Everticyclammina praekelleri and E. kelleri are related mainly to the development of the alveolar structures. E. praekelleri has much enlarged lateral alveoli compared with E. kelleri. The presence of this foraminifer in the Piatra Craiului Massif is mentioned for the first time in this study.
Stratigraphic range: lower Kimmeridgian-upper Tithonian.
Everticyclammina kelleri (Henson, 1948)
(Fig. 6.H ![]() )
)
1948 - Pseudocyclammina kelleri n. sp. - Henson, p. 16-17, Pl. 9, figs. 4-5, 7.
1990 - Everticyclammina kelleri (Henson, 1948) - Banner & Highton, p. 5, Pl. 1, figs. 2-6; p. 7, Pl. 2, figs. 1-4; p. 9, Pl. 3, figs. 1-2.
Description: Planispirally enrolled test in early ontogenetic stages, involute. In several specimens, in the adult stage, the last two chambers can be uncoiled. Wall is thick, alveolar, with agglutinated particles. The inner structure of the foraminifer is composed of networks of bifurcated alveoli, mostly in the lateral parts of the hypodermis. The chambers are flattened; the septa are thick, non alveolar, pierced by a simple-type aperture.
Remarks: In comparison with E. praekelleri, E. kelleri specimens possess a more developed planispiral stage.
Stratigraphic range: ?Tithonian-Valanginian.
Genus Rectocyclammina Hottinger, 1967
? Rectocyclammina sp.
Description: The test is conical, tall, elongated with a relatively short planispiral early stage. The adult stage consists of inflated chambers that rapidly increase in height as they successively develop. The chambers are separated by thick septa. Wall is alveolar (alveoli network), agglutinated. The aperture seems to be circular, in the center of the apertural face.
Family Cyclamminidae Marie, 1941
Subfamily Choffatellinae Maync, 1958
Genus Bramkampella Redmond, 1964
Bramkampella arabica Redmond, 1964
(Fig. 4.C ![]() )
)
1964 - Bramkampella arabica n. sp. - Redmond, p. 410, Pl. 1, figs. 26-29.
1991 - Bramkampella arabica Redmond, 1964 - Banner & Whittaker, p. 45, Pl. 2, figs. 1-7.
2005 - Bramkampella arabica Redmond, 1964 - Bucur & Săsăran, Pl. 2, figs. 6-7.
Description: Medium-sized robust subconical test, planspirally enrolled in early stage. The test uncoils in a short distance and progressively gains size in the adult stage. Wall is alveolar, agglutinated. A network of elongated interseptal alveoli radially developed defines the inner structure of the foraminifer. The septa are high-arched, pierced by many openings (multiple aperture type).
Remarks: The highly curved septa and the multiple aperture type, differentiate this species from Rectocyclammina chouberti. The presence of this foraminifer in the Piatra Craiului Massif is mentioned for the first time in this study.
Stratigraphic range: Kimmeridgian-lower Valanginian.
Suborder Ataxophragmiina Fursenko, 1958
Superfamily Ataxophragmioidea Schwager, 1877
Family Montsaleviidae Zaninetti et al., 1987
Genus Montsalevia Zaninetti et al., 1987
Montsalevia salevensis (Charollais et al., 1966)
(Fig. 6.I ![]() )
)
1966 - Pseudotextulariella salevensis n. sp. - Charollais et al., p. 28, Pl. 1, figs. 1-5.
1966 - Pseudotextulariella salevensis - Brönnimann, Pl. III, fig. 4.
1987 - "Montsalevia" salevensis - Zaninetti et al., p. 166
1988 - Pseudotextulariella salevensis Charollais et al., 1966 - Bucur, p. 387, Pl. 2, figs. 11-12.
1991 - Montsalevia salevensis - Altiner, p. 170, Fig. 3; p. 173-177, Pl. 11, figs. 1-23.
2010 - Montsalevia salevensis (Charollais et al., 1966) - Ivanova & Kołodziej, p. 25, Pl. 2, fig. 10.
2016 - Montsalevia salevensis (Charollais et al., 1966) - Grădinaru et al., Fig. 14.G-K.
Description: Small-sized conical test that is trochospirally enrolled in the initial stage, later becoming biserial. The inner-structure is divided by radial partitions in numerous small flatten chamberlets. The wall is microgranular (finely agglutinated), imperforated. Aperture is interiomarginal.
Stratigraphic range: Berriasian-Hauterivian.
Family Cuneolinidae Sadoiva, 1981
Subfamily Cuneolininae Sadoiva, 1981
Genus Pseudotextulariella Barnard, 1953
Pseudotextulariella courtionensis Brönnimann, 1966
(Fig. 5.C ![]() )
)
1966 - Pseudotextulariella courtionensis n. sp. - Brönnimann, p. 267-276, Figs. 1-8; Pl. I, figs. 1-5; Pl. II, figs. 1-2; Pl. III, fig. 3.
1977 - Pseudotextulariella courtionensis Brönnimann, 1966 - Azéma et al., p. 137, Pl. 3, figs. 13-14.
1983 - Pseudotextulariella courtionensis Brönnimann, 1966 - Darsac, Pl. 5, figs. 1-8.
2016 - Pseudotextulariella courtionensis Brönnimann, 1966 - Grădinaru et al., Fig. 14.B.
Description: The initial stage of the test is trochospirally developed, subsequently becoming triserial and biserial in the adult stage. Shape of the test is low to high conical. The internal structure is composed of horizontal and vertical partitions. Wall is finely agglutinated. Aperture face is flattened with a slit-type opening at the base.
Stratigraphic range: Berriasian.
Suborder Orbitolinina Kaminski, 2004
Superfamily Pfenderinoidea Smout & Sugden, 1962
Family Pfenderinidae Smout & Sugden, 1962
Subfamily Pfenderininae Smout & Sugden, 1962
Genus Pfenderina Henson, 1948
Pfenderina neocomiensis (Pfender, 1938)
(Fig. 6.M-O ![]() )
)
1938 - Eorupertia neocomiensis n. sp. - Pfender - p. 236, Pl. XVI, figs. 1-7.
1961 - Pfenderina neocomiensis (Pfender, 1938) - Smout & Sugden, p. 585-588, Pl. 73, figs. 1-9; Pl. 74, figs. 1-3; Pl. 75, fig. 1.
1995 - Pfenderina neocomiensis (Pfender, 1938) - Bucur et al., p. 369, Pl. 6, figs. 4-6.
Description: The test is high trochospiral with a thickened (columellar) central zone. The chambers are small and numerous separated by oblique septa. The wall is agglutinated/microgranular imperforate. Primary aperture is multiple, secondary one being represented by an intercameral passage (groove), spiraling around the thick axial zone.
Stratigraphic range: upper Berriasian-Valanginian.
Family Hauraniidae Septfontaine, 1988
Subfamily Amijellinae Septfontaine, 1988
Genus Anchispirocyclina Jordan & Applin, 1952
cf. Anchispirocyclina lusitanica (Egger, 1902)
(Fig. 5.E-F ![]() )
)
1902 - Dicyclina lusitanica n. sp. - Egger, p. 585, Pl. 6, figs. 3-5.
1967 - Anchispirocyclina lusitanica lusitanica (Egger, 1902) - Hottinger, p. 74, Pl. 13, figs. 6-8.
1987 - Anchispirocyclina lusitanica (Egger, 1902) - Granier, Pl. 48, fig. k.
2005 - Anchispirocyclina lusitanica (Egger, 1902) - Schlagintweit et al., p. 25, Fig. 5.a-c.
Description: Test large, compressed, planispirally to asymmetrically coiled in juvenile stage, in adult stage spreading becoming peneropliform or circular. The wall is imperforate, finely agglutinated. Reticulate layers of beams and rafters characterize the hypodermis. The central part of the test is represented by a multitude of radial interseptal pillars. The chambers are elongated, especially in adult stages, separated by highly curved septa. Aperture is cribate, extending across the apertural face.
Remarks: The specimens identified in Piatra Craiului are poorly preserved, and the above-described characteristics are difficult to observe. The presence of this foraminifer in the Piatra Craiului Massif is mentioned for the first time in this study.
Stratigraphic range: Tithonian-lower Berriasian.
Genus Pseudocyclammina Yabe & Hanzawa, 1926
Pseudocyclammina lituus (Yokoyama, 1890)
1890 - Cyclammina lituus n. sp. - Yokoyama, p. 26, Pl. 5, fig. 7.
1926 - Pseudocyclammina lituus (Yokoyama, 1890) - Yabe & Hanzawa, p. 10, Pl. 2, figs. 3-6.
1995 - Pseudocyclammina lituus (Yokoyama, 1890) - Bucur et al., p. 358, Pl. 1, figs. 3-4.
2006 - Pseudocyclammina lituus (Yokoyama, 1890) - Kobayashi & Vuks, p. 840, Figs. 5, 7-14.
Description: Planispirally enrolled medium to large-sized test tending to uncoil in advanced ontogenetic stages. Wall alveolar, coarsely agglutinated with layers of thick alveoli in the hypodermis; septa are very thick. Aperture is areal, multiple, across the apertural face.
Stratigraphic range: Kimmeridgian-lower Valanginian.
Family Parurgoninidae Septfontaine, 1988
Genus Parurgonina Cuvillier et al., 1968
Parurgonina caelinensis Cuvillier et al., 1968
(Fig. 4.M-N ![]() )
)
1968 - Urgonina (Parurgonina) caelinensis n. sp. - Cuvillier et al., p. 151, Pl. 2, figs. 1-12.
1975 - Parurgonina caelinensis Cuvillier et al., 1968 - Schroeder et al., p. 320-325, Pl. 1, figs. 1-4; Pl. 2, figs. 3-5.
2014 - Parurgonina caelinensis Cuvillier et al., 1968 - Mircescu et al., p. 13, Pl. 1, fig. 1.
Description: Test is conical, trochospirally developed in the initial part, later becoming uniserial. The chambers are cylindrical/semi-lunar in shape, separated by vertical pillars and low-arched septa. The wall is pseudo-keriothecal, made of fine fibrous microstructural elements. Aperture is multiple.
Remarks: P. caelinensis differs from Neokilianina rahonensis by internal structural features (pillar development and the shape of the chamber lumen).
Stratigraphic range: uppermost Oxfordian-lower Tithonian.
Genus Neokilianina Septfontaine, 1988
Neokilianina rahonensis (Foury & Vincent, 1967)
(Fig. 4.I-L ![]() )
)
1967 - Kilianina rahonensis n. sp. - Foury & Vincent, Pl. 2, figs. 1-14.
1988 - Neokilianina rahonensis (Foury & Vincent, 1967) - Septfontaine, p. 249.
2005 - "Kilianina" rahonensis Foury & Vincent, 1967 - Schlagintweit et al., p. 29, Fig. 11.a-d.
Description: Medium sized conical test, with numerous chambers in the adult stage. The shape of the chamber lumen in longitudinal sections is more or less triangular (oblique stolons/low arched septa). Closely spaced interseptal pillars are developed in the central part of the test. Wall is finely agglutinated with canaliculate structure; aperture multiple.
Remarks: The presence of this foraminifer in the Piatra Craiului Massif is mentioned for the first time in this study.
Stratigraphic range: uppermost Oxfordian-lower Tithonian.
Superfamily Orbitolinoidea Martin, 1890
Subfamily Praedictyorbitolininae Schroeder, 1990
Genus Paracoskinolina Moullade, 1965
Paracoskinolina ? jourdanensis (Foury & Moullade, 1966)
(Fig. 6.K-L ![]() )
)
1966 - Meyendorffina (Paracoskinolina) jourdanensis n. sp. - Foury & Moullade, p. 252, Pl. 1, figs. 1-6.
1980 - Paracoskinolina ? jourdanensis (Foury & Moullade, 1966) - Arnaud-Vanneau, Pl. 102, figs. 1-4.
2008 - Paracoskinolina ? jourdanensis (Foury & Moullade, 1966) - Michetiuc et al., p. 226, Pl. 3, fig. 3.
2014 - Paracoskinolina ? jourdanensis (Foury & Moullade, 1966) - Bruchental et al., p. 36, Fig. 3.a.
2016 - Paracoskinolina ? jourdanensis (Foury & Moullade, 1966) - Grădinaru et al., Fig. 14.A.
Description: High conical test, some specimens with an apical bulb (a short juvenile spire), followed by rectilinear development. Long intercameral vertical pillars characterize the internal structure of the foraminifer. Also two sets of radial beams are present in the subepidermal region. Wall is microgranular, imperforated. Aperture is composed of multiple pores on the basal side.
Stratigraphic range: upper Berriasian-lower Barremian.
Order Textulariida Delage & Hérouard, 1896
Suborder Textulariina Delage & Hérouard, 1896
Superfamily Chrysalidinoidea Neagu, 1968
Family Paravalvulinidae Banner et al., 1991
Subfamily Paravalvulininae Banner et al., 1991
Genus Redmondoides Banner et al., 1991
Redmondoides lugeoni (Septfontaine, 1977)
(Fig. 4.H ![]() )
)
1977 - Valvulina lugeoni n. sp. - Septfontaine, p. 612-613, Pl. 2, figs. 2-5.
1987 - Valvulina lugeoni Septfontaine, 1977 - Granier, Pl. 14, fig. g.
1991 - Redmondoides lugeoni (Septfontaine, 1977) - Banner et al., p. 127, figs. 46-54.
2016 - Redmondoides lugeoni (Septfontaine, 1977) - Granier et al., p. 259, Pl. 1, fig. 17.
Description:
Large-sized conical, trochospiral test with thick microgranular/agglutinated
walls and septa. The test is quadriserial throughout most ontogenetic stages.
The microgranular material that forms the test walls and septa may develop
canaliculated features in some specimens. Aperture is interiomarginal
(slit-type) enclosed by a flat lip (Fig. 4.G ![]() ).
).
Stratigraphic range: upper Bathonian-lower Tithonian.
Order Involutinida Hohenegger & Piller, 1977
Suborder Involutinina Hohenegger & Piller, 1977
Superfamily Involutinoidea Bütschli, 1880
Family Trocholinidae Kristan-Tollmann, 1963
Subfamily Trocholininae Kristan-Tollmann, 1963
Genus Coscinoconus Leupold, 1936
Coscinoconus campanellus (Arnaud-Vanneau et al., 1988)
(Fig. 6.C-D ![]() )
)
1988 - Trocholina campanella n. sp. - Arnaud-Vanneau et al., p. 371, Pl. 3, figs. 9-15.
1994 - Andersenolina campanella (Arnaud-Vanneau et al., 1988) - Neagu, p. 143, Pl. 13, figs. 19-20.
2013 - Coscinoconus campanellus (Arnaud-Vanneau et al., 1988) - Rigaud et al., p. 330.
2016 - Coscinoconus campanellus (Arnaud-Vanneau et al., 1988) - Grădinaru et al., Fig. 14.U.
Description: Large-sized Coscinoconus representative with a bell-shaped (or piriform), trochospirally coiled test and a convex base. The chambers are tubular, undivided. Apical angle ranges between 60 and 70 degrees. Wall (originally aragonitic) is hyaline, perforated. Aperture located at the end of the tubular chambers.
Remarks: This species differs from the other Coscinoconus representatives by the apical angle and by the pyriform shape of the test.
Stratigraphic range: upper Berriasian-Valanginian.
Coscinoconus cherchiae (Arnaud-Vanneau et al., 1988)
(Fig. 6.E ![]() )
)
1988 - Trocholina cherchiae n. sp. - Arnaud-Vanneau et al., p. 369, Pl. 2, figs. 9-21.
1994 - Andersenolina cherchiae (Arnaud-Vanneau et al., 1988) - Neagu, p. 126, Pl. 5, figs. 1-14.
2013 - Coscinoconus cherchiae (Arnaud-Vanneau et al., 1988) - Rigaud et al., p. 330.
2016 - Coscinoconus cherchiae (Arnaud-Vanneau et al., 1988) - Grădinaru et al., Fig. 14.V.
Description: The test is medium in size characterized by variations of shape during ontogeny. In the initial stage, the test is low to high conical in shape (40-50 degrees apical angle), and in advanced ontogenetic stages, the test becomes more cylindrical and elongated. Chambers are broader than those of C. campanellus and triangular in shape in lungitudinal sections.
Stratigraphic range: upper Berriasian-Valanginian.
Family Ventrolaminidae Weynschenk, 1950
Genus Protopeneroplis Weynschenk, 1950
Protopeneroplis ultragranulata (Gorbatchik, 1971)
(Fig. 6.P ![]() )
)
1971 - Hoeglundina ? ultragranulata n. sp. - Gorbatchik, p. 135, Pl. 5, fig. 2.a-c.
1974 - Protopeneroplis trochangulata n. sp. - Septfontaine., p. 608, Pl. 1, figs. 1-18.
1987 - Protopeneroplis trochangulata Septfontaine, 1974 - Granier, Pl. 3, figs. g-j; Pl. 44, figs. a, d.
1993 - Protopeneroplis ultragranulata (Gorbatchik, 1971) - Bucur, p. 221, Pl. 2, figs. 1-2, 5, 8, 11-12.
1997 - Protopeneroplis ultragranulata (Gorbatchik, 1971) - Bucur, Pl. 6, figs. 1-14.
2016 - Protopeneroplis ultragranulata (Gorbatchik, 1971) - Grădinaru et al., Fig. 14.S-T.
Description: Small-sized trochospirally enrolled, lenticular test, involute, with a two layered calcareous wall. The inner layer is microgranular, protected by a hyaline outer layer. Aperture is areal.
Remarks: The trochospiral model of coiling of P. ultragranulata differentiates it from P. striata Weynschenk.
Stratigraphic range: middle Tithonian-Barremian (acme in Berriasian-Valanginian).
Order Rotaliida Lankester, 1885
Suborder Rotaliina Delage & Hérouard, 1986
?Family Rosalinidae Reiss, 1963
Genus Mohlerina Bucur et al., 1996
Mohlerina basiliensis (Mohler, 1938)
(Fig. 4.A ![]() )
)
1938 - Conicospirillina basiliensis n. sp. - Mohler, p. 27, Pl. 4, figs. 4-5.
1987 - "Conicospirillina" basiliensis Mohler, 1938 - Granier, Pl. 3, figs. g-j; Pl. 44, fig. e.
1996 - Mohlerina basiliensis (Mohler, 1938) - Bucur et al., p. 74, Pl. 3, figs. 3-6.
2012 - Mohlerina basiliensis (Mohler, 1938) - Schlagintweit, p. 639, Fig. 2.a-j.
Description: The test is generally low conical in shape, trochospirally coiled. The test wall is calcareous bilamellar (microcrystalline layer and radial-fibrous calcitic layer). Aperture is possibly slit-type.
Remarks: M. basiliensis is considered to be the single species of the genus Mohlerina.
Stratigraphic range: middle Bathonian-lower Valanginian.
Three biostratigraphic
intervals (A-C) were differentiated in the Upper Jurassic-Lower Cretaceous
carbonate succession from Piatra Craiului Massif based on the occurrence of
several species of foraminifera and dasycladalean algae (Fig. 3 ![]() ).
).
The first interval
contains the following algae and foraminifera (Fig. 3 ![]() ): Campbeliella striata (Carozzi, 1954), Clypeina sulcata (Alth, 1882), Neoteutloporella socialis (Praturlon, 1963), Petrascula
bursiformis (Etallon, 1859), Salpingoporella
annulata Carozzi, 1953, S.
pygmaea (Gümbel, 1891), Steinmanniporella
kapelensis (Sokač & Nikler,
1973), Bramkampella arabica Redmond,
1964 (Fig. 4.C
): Campbeliella striata (Carozzi, 1954), Clypeina sulcata (Alth, 1882), Neoteutloporella socialis (Praturlon, 1963), Petrascula
bursiformis (Etallon, 1859), Salpingoporella
annulata Carozzi, 1953, S.
pygmaea (Gümbel, 1891), Steinmanniporella
kapelensis (Sokač & Nikler,
1973), Bramkampella arabica Redmond,
1964 (Fig. 4.C ![]() ), Everticyclammina
praekelleri Banner & Highton,
1990 (Fig.
4.F-G
), Everticyclammina
praekelleri Banner & Highton,
1990 (Fig.
4.F-G ![]() ), Frentzenella
involuta (Mantsurova & Gorbatchik, 1982)
(Fig. 4.B
), Frentzenella
involuta (Mantsurova & Gorbatchik, 1982)
(Fig. 4.B ![]() ), Labyrinthina mirabilis Weynschenk,
1951 (Fig.
4.D
), Labyrinthina mirabilis Weynschenk,
1951 (Fig.
4.D ![]() ), Lituola
baculiformis Schlagintweit & Gawlick, 2007
(Fig. 4.E
), Lituola
baculiformis Schlagintweit & Gawlick, 2007
(Fig. 4.E ![]() ), Lenticulina
sp., Mohlerina basiliensis (Mohler, 1938) (Fig. 4.A
), Lenticulina
sp., Mohlerina basiliensis (Mohler, 1938) (Fig. 4.A ![]() ), Neokilianina
rahonensis (Foury & Vincent,
1967)
(Fig. 4.I-L
), Neokilianina
rahonensis (Foury & Vincent,
1967)
(Fig. 4.I-L ![]() ), Nodosaria
sp., Parurgonina caelinensis Cuvillier et al.,
1968 (Fig. 4.M-N
), Nodosaria
sp., Parurgonina caelinensis Cuvillier et al.,
1968 (Fig. 4.M-N ![]() ), and Redmondoides lugeoni (Septfontaine,
1977)
(Fig. 4.H
), and Redmondoides lugeoni (Septfontaine,
1977)
(Fig. 4.H ![]() ). The
microfossil assemblage identified in this interval (0-410 m) is characteristic
of the Kimmeridgian-lower Tithonian interval.
). The
microfossil assemblage identified in this interval (0-410 m) is characteristic
of the Kimmeridgian-lower Tithonian interval.
Although some species of algae (e.g., Salpingoporella pygmaea or Clypeina sulcata) have longer stratigraphic distributions, most taxa provide valuable biostratigraphic information:
Regarding the foraminiferal assemblage, Neokilianina rahonensis, Parurgonina caelinensis and Labyrinthina mirabilis represent the most biostratigraphically important taxa for this interval. They were reported mainly from Kimmeridgian-Tithonian strata (Cuvillier et al., 1968; Septfontaine, 1988; Tasli, 1993; Pop & Bucur, 2001; Velić, 2007; Pleş et al., 2015).
Considering
this, the whole micropaleontological assemblage identified in this
biostratigraphic interval (Fig. 3 ![]() )
represents the Kimmeridgian-lower Tithonian
interval. Although some species of foraminifera (Labyrinthina mirabilis, Neokilianina
rahonensis,
and Parurgonina caelinensis)
appear lower in the geological record in the uppermost Oxfordian (Septfontaine,
1988; Bassoullet, 1997; Velić,
2007; Pleş et
al., 2015), well-dated Oxfordian radiolarites (Mészáros & Bucur,
1980; Beccaro & Lazăr,
2007) are directly below the
limestones of interval A. Moreover, many microfossils of the assemblage (Clypeina
sulcata, Petrascula bursiformis, Salpingoporella
pygmaea, Coscinoconus alpinus, Everticyclammina praekelleri,
Mohlerina basiliensis,
Redmondoides lugeoni)
represent typical Kimmeridgian-Tithonian biota (Bucur, 1999; Schlagintweit
et al., 2005), and most of the
mentioned taxa are known from carbonates no older than lower Kimmeridgian (Bassoullet,
1997). In addition, the presence of Steinmanniporella
kapelensis and several sclerosponge species (Calcistella jachenhausenensis Reitner,
1992, Neuropora
lusitanica Termier,
1985,
and Thalamopora lusitanica Termier
et al., 1985)
confirms the Tithonian age of the upper part of biostratigraphic interval A.
)
represents the Kimmeridgian-lower Tithonian
interval. Although some species of foraminifera (Labyrinthina mirabilis, Neokilianina
rahonensis,
and Parurgonina caelinensis)
appear lower in the geological record in the uppermost Oxfordian (Septfontaine,
1988; Bassoullet, 1997; Velić,
2007; Pleş et
al., 2015), well-dated Oxfordian radiolarites (Mészáros & Bucur,
1980; Beccaro & Lazăr,
2007) are directly below the
limestones of interval A. Moreover, many microfossils of the assemblage (Clypeina
sulcata, Petrascula bursiformis, Salpingoporella
pygmaea, Coscinoconus alpinus, Everticyclammina praekelleri,
Mohlerina basiliensis,
Redmondoides lugeoni)
represent typical Kimmeridgian-Tithonian biota (Bucur, 1999; Schlagintweit
et al., 2005), and most of the
mentioned taxa are known from carbonates no older than lower Kimmeridgian (Bassoullet,
1997). In addition, the presence of Steinmanniporella
kapelensis and several sclerosponge species (Calcistella jachenhausenensis Reitner,
1992, Neuropora
lusitanica Termier,
1985,
and Thalamopora lusitanica Termier
et al., 1985)
confirms the Tithonian age of the upper part of biostratigraphic interval A.
In
the second biostratigraphic interval (B), the total number of species of
dasycladalean algae decreases compared to that of the foraminifera species (Fig. 3 ![]() ). The following species have been identified in this interval: Clypeina parasolkani Farinacci & Radoičić,
1991, Salpingoporella annulata Carozzi, 1953, Selliporella
neocomiensis
(Radoičić, 1963), cf. Anchispirocyclina
lusitanica (Egger, 1902) (Fig. 5.E-F
). The following species have been identified in this interval: Clypeina parasolkani Farinacci & Radoičić,
1991, Salpingoporella annulata Carozzi, 1953, Selliporella
neocomiensis
(Radoičić, 1963), cf. Anchispirocyclina
lusitanica (Egger, 1902) (Fig. 5.E-F ![]() ), Bulbobaculites sp.
(Fig. 5.A
), Bulbobaculites sp.
(Fig. 5.A ![]() ), Pseudocyclammina
lituus (Yokoyama, 1890) (Fig. 5.B
), Pseudocyclammina
lituus (Yokoyama, 1890) (Fig. 5.B ![]() , Pseudotextulariella courtionensis Brönnimann,
1966
(Fig. 5.C
, Pseudotextulariella courtionensis Brönnimann,
1966
(Fig. 5.C ![]() ), and Rectocyclammina sp. Hottinger,
1967 (Figs. 5.D
), and Rectocyclammina sp. Hottinger,
1967 (Figs. 5.D ![]() & 6.G
& 6.G ![]() ).
).
The
lower part of the biostratigraphic interval B is probably still Tithonian in
age. The first occurrence of "cf. Anchispirocyclina
lusitanica" is recorded near the base of
biostratigraphic interval B (Fig. 3 ![]() )
where this foraminifer is associated with
the alga Clypeina parasolkani. Pseudocyclammina
lituus appears slightly higher in the same interval of the sections studied.
The first primary evidence for a Berriasian age is brought by the first
occurrence of Selliporella neocomiensis (Fig. 3
)
where this foraminifer is associated with
the alga Clypeina parasolkani. Pseudocyclammina
lituus appears slightly higher in the same interval of the sections studied.
The first primary evidence for a Berriasian age is brought by the first
occurrence of Selliporella neocomiensis (Fig. 3 ![]() ), followed by that of Pseudotextulariella
courtionensis. Thus the
upper part of the biostratigraphic interval B can definitely be ascribed to the
lower Berriasian (Granier & Bucur,
2011).
), followed by that of Pseudotextulariella
courtionensis. Thus the
upper part of the biostratigraphic interval B can definitely be ascribed to the
lower Berriasian (Granier & Bucur,
2011).
It is impossible to identify the Tithonian-Berriasian
boundary in interval B, because none of the markers, either primary or
secondary, or proxies are present in our material. The micropaleontological
assemblage of this stratigraphic interval (Fig. 3 ![]() )
merely indicates the Upper
Jurassic-Lower Cretaceous transition.
)
merely indicates the Upper
Jurassic-Lower Cretaceous transition.
As
in interval B (Fig. 3 ![]() ), the total number of foraminiferal species in interval C
exceeds that of other taxa such as dasycladalean algae. The main microfossils
are represented by: Pseudocymopolia
jurassica (Dragastan, 1968), Salpingoporella
praturloni (Dragastan, 1978) (Fig. 3
), the total number of foraminiferal species in interval C
exceeds that of other taxa such as dasycladalean algae. The main microfossils
are represented by: Pseudocymopolia
jurassica (Dragastan, 1968), Salpingoporella
praturloni (Dragastan, 1978) (Fig. 3 ![]() ), Coscinoconus campanellus (Arnaud-Vanneau et al.,
1988) (Figs. 3
), Coscinoconus campanellus (Arnaud-Vanneau et al.,
1988) (Figs. 3 ![]() & 6.C-D
& 6.C-D ![]() ), C. cherchiae
(Arnaud-Vanneau et al., 1988) (Figs. 3
), C. cherchiae
(Arnaud-Vanneau et al., 1988) (Figs. 3 ![]() & 6.E
& 6.E ![]() ), Everticyclammina
kelleri (Henson, 1948) (Fig.
6.H
), Everticyclammina
kelleri (Henson, 1948) (Fig.
6.H ![]() ), Freixialina planispiralis Ramalho, 1969
(Fig. 6.O
), Freixialina planispiralis Ramalho, 1969
(Fig. 6.O ![]() , arrow), Montsalevia
salevensis (Charollais et al., 1966) (Figs. 3
, arrow), Montsalevia
salevensis (Charollais et al., 1966) (Figs. 3 ![]() & 6.I
& 6.I ![]() ), Nautiloculina
cf. broennimanni (Arnaud-Vanneau & Peybernčs,
1978) (Figs. 3
), Nautiloculina
cf. broennimanni (Arnaud-Vanneau & Peybernčs,
1978) (Figs. 3 ![]() & 6.F
& 6.F ![]() ), Paracoskinolina ?
jourdanensis Foury & Moullade,
1966 (Figs. 3
), Paracoskinolina ?
jourdanensis Foury & Moullade,
1966 (Figs. 3 ![]() & 6.K-L
& 6.K-L ![]() ), Pfenderina
neocomiensis (Pfender, 1938) (Figs. 3
), Pfenderina
neocomiensis (Pfender, 1938) (Figs. 3 ![]() & 6.M-O
& 6.M-O ![]() ), Protopeneroplis
ultragranulata (Gorbatchik, 1971) (Figs. 3
), Protopeneroplis
ultragranulata (Gorbatchik, 1971) (Figs. 3 ![]() & 6.P
& 6.P ![]() ), Pseudocyclammina
lituus (Yokoyama, 1890) (Fig. 6.A-B
), Pseudocyclammina
lituus (Yokoyama, 1890) (Fig. 6.A-B ![]() ), and Scythiolina
sp. (Fig. 6.J
), and Scythiolina
sp. (Fig. 6.J ![]() ).
).
Dasycladalean
algae (Pseudocymopolia jurassica, Salpingoporella
praturloni) are rare. They were identified in a stratigraphic level in the
uppermost part of this interval. Foraminifera (Coscinoconus
campanellus, C. cherchiae, Montsalevia
salevensis, Nautiloculina cf. broennimanni, Paracoskinolina ?
jourdanensis, Pfenderina neocomiensis, Protopeneroplis
ultragranulata) are abundant in the same level (Fig. 3 ![]() ).
).
Some foraminifera found in this assemblage (e.g., Protopeneroplis ultragranulata) extend up into the Barremian (Bucur, 1997; Granier & Bucur, 2011). However, most algae or foraminifera listed above do not extend higher than the Valanginian and some have never been reported from Upper Valanginian strata. In conclusion, the above mentioned assemblage indicates a late Berriasian-early Valanginian age for biostratigraphic unit C.
The exact position of the Berriasian-Valanginian boundary is difficult to identify on the sole basis of the micropaleontological assemblage. However, based on ammonite and calpionellid finds, the lowermost strata of the transgressive Dâmbovicioara Formation that directly overlie limestones of interval C are dated as Early Valanginian (Grădinaru et al., 2016). At this point, only two options are considered: 1) the uppermost strata of lithostratigraphic unit III and biostratigraphic unit C could be earliest Valanginian in age, or 2) the hiatus at the boundary with the overlying Dâmbovicioara Formation spans the stage boundary and the uppermost strata of lithostratigraphic unit III and biostratigraphic unit C are Late Berriasian in age.
The Upper Jurassic-Lower Cretaceous succession from Piatra Craiului contains mainly benthic foraminifera. Various factors, such as salinity, water temperature, and nutrients (Reiss & Hottinger, 1984; Hughes, 2000), play an essential role in the diversity and abundance of the benthic microfauna and the associated calcareous algae. Therefore, paleoecological information can be derived from their assemblages and their analysis may contribute to improving the depositional model of the carbonate succession.
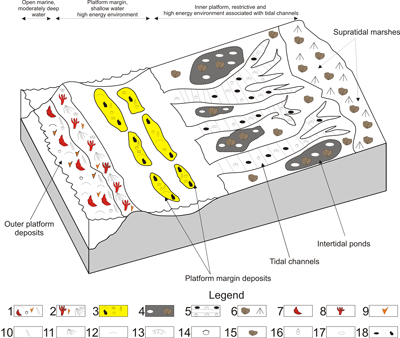
The
lowermost part of the carbonate succession corresponding to lithostratigraphic
interval I (Fig. 3 ![]() )
consists of interbedded reef rudstones and coral microbial
boundstones, i.e., a gradual transition from reef slope areas to reef
margin environments with bioconstructions becoming dominant towards the top.
Carbonate material was transported from the bioconstructions and reworked on the
deeper fore-reef slope where coral fragments and echinoderm plates are the most
important bioclasts (Fig. 7
)
consists of interbedded reef rudstones and coral microbial
boundstones, i.e., a gradual transition from reef slope areas to reef
margin environments with bioconstructions becoming dominant towards the top.
Carbonate material was transported from the bioconstructions and reworked on the
deeper fore-reef slope where coral fragments and echinoderm plates are the most
important bioclasts (Fig. 7 ![]() )
(Pleş et al., 2013). Lithostratigraphic interval I (0-289 m) (Fig. 3
)
(Pleş et al., 2013). Lithostratigraphic interval I (0-289 m) (Fig. 3 ![]() )
contains Lenticulina sp., Nodosaria
sp. (Fig. 3
)
contains Lenticulina sp., Nodosaria
sp. (Fig. 3 ![]() )
and other foraminiferal species characteristic of the lower and
middle parts of biostratigraphic interval A (Fig. 3
)
and other foraminiferal species characteristic of the lower and
middle parts of biostratigraphic interval A (Fig. 3 ![]() ). They are associated with
fragments of juvenile ammonites and encrusting organisms (mainly Crescentiella morronensis) (Fig. 7
). They are associated with
fragments of juvenile ammonites and encrusting organisms (mainly Crescentiella morronensis) (Fig. 7 ![]() ). The presence of Lenticulina
sp. in the assemblage with other bioclasts points to a moderately deep,
open-marine environment (Hughes, 2000; Reolid et
al., 2008a, 2008b; Nikitenko et
al., 2013).
). The presence of Lenticulina
sp. in the assemblage with other bioclasts points to a moderately deep,
open-marine environment (Hughes, 2000; Reolid et
al., 2008a, 2008b; Nikitenko et
al., 2013).
Lithostratigraphic
interval II (290-408 m) (Fig. 3 ![]() )
contains foraminifera from the upper part of
biostratigraphic interval A (Fig. 3
)
contains foraminifera from the upper part of
biostratigraphic interval A (Fig. 3 ![]() ). The number of specimens of Neokilianina
rahonensis and Parurgonina caelinensis
gradually increases between 290-340 m. Their abundance is higher at 340 m
compared with their isolated occurrence 50 meters below. According to Hughes
(2000), "the progressively ascending appearance of certain species" and the
gradual increase of "their vertical extent and abundance until a consistent
presence" up section reflects a "possible gradual shallowing" of the
depositional environments. These foraminifera are in coarse bioclastic
grainstones (lithostratigraphic interval II) associated with reworked
black-pebbles and dasycladalean algae (Steinmanniporella
kapelensis and
Neoteutloporella socialis) (Fig. 7
). The number of specimens of Neokilianina
rahonensis and Parurgonina caelinensis
gradually increases between 290-340 m. Their abundance is higher at 340 m
compared with their isolated occurrence 50 meters below. According to Hughes
(2000), "the progressively ascending appearance of certain species" and the
gradual increase of "their vertical extent and abundance until a consistent
presence" up section reflects a "possible gradual shallowing" of the
depositional environments. These foraminifera are in coarse bioclastic
grainstones (lithostratigraphic interval II) associated with reworked
black-pebbles and dasycladalean algae (Steinmanniporella
kapelensis and
Neoteutloporella socialis) (Fig. 7 ![]() ), which define high-energy, shallow-water
platform margin environments. Redmondoides
lugeoni, Coscinoconus alpinus and Everticyclammina
praekelleri are also present in lithostratigraphic interval II (Fig. 3
), which define high-energy, shallow-water
platform margin environments. Redmondoides
lugeoni, Coscinoconus alpinus and Everticyclammina
praekelleri are also present in lithostratigraphic interval II (Fig. 3 ![]() ).
Several authors (Pélissié et al., 1984; Tyszka, 1994; Savelieva et
al., 2014) have described these species from similar high-energy facies
(bioclastic grainstones) and depositional settings (Fig. 7
).
Several authors (Pélissié et al., 1984; Tyszka, 1994; Savelieva et
al., 2014) have described these species from similar high-energy facies
(bioclastic grainstones) and depositional settings (Fig. 7 ![]() ). Further discussion
regarding the lithostratigraphy of these deposits can be found in Mircescu et al.
(2014). The facies identified in lithostratigraphic
interval II point to the presence of elevated carbonate margin deposits where
coarser sediments were accumulating under high-energy conditions within a
shallow-water environment (Fig. 7
). Further discussion
regarding the lithostratigraphy of these deposits can be found in Mircescu et al.
(2014). The facies identified in lithostratigraphic
interval II point to the presence of elevated carbonate margin deposits where
coarser sediments were accumulating under high-energy conditions within a
shallow-water environment (Fig. 7 ![]() ). Coral fragments are common which suggests
that reworking has occurred either from the underlying bioconstructions or from
adjacent and coeval bioconstructions. The presence of micritic-rimmed bioclasts
indicates that the calcareous sand bars were probably adjacent to a lagoonal
area where the micritisation occurred under more restrictive conditions.
Subsequently, the micritised bioclasts were reworked in high-energy deposits.
). Coral fragments are common which suggests
that reworking has occurred either from the underlying bioconstructions or from
adjacent and coeval bioconstructions. The presence of micritic-rimmed bioclasts
indicates that the calcareous sand bars were probably adjacent to a lagoonal
area where the micritisation occurred under more restrictive conditions.
Subsequently, the micritised bioclasts were reworked in high-energy deposits.
Lithostratigraphic
interval III (Fig. 3 ![]() )
is composed of interbedded intertidal/supratidal peloidal
wackestone-packstone and homogeneous mudstone with cyanobacteria nodules.
Cyanobacteria are forming the main bioclasts. The intertidal conditions are
indicated by abundant laminoid fenestral structures (Fig. 7
)
is composed of interbedded intertidal/supratidal peloidal
wackestone-packstone and homogeneous mudstone with cyanobacteria nodules.
Cyanobacteria are forming the main bioclasts. The intertidal conditions are
indicated by abundant laminoid fenestral structures (Fig. 7 ![]() ). Carbonate sediment
was accumulating in intertidal ponds where cyanobacteria were the main sediment
producers. The supratidal environment is indicated by the presence of abundant
cyanobacteria, plant roots and fine, micritic sediment (Fig. 7
). Carbonate sediment
was accumulating in intertidal ponds where cyanobacteria were the main sediment
producers. The supratidal environment is indicated by the presence of abundant
cyanobacteria, plant roots and fine, micritic sediment (Fig. 7 ![]() ). As the
carbonate platform was prograding, the accomodation space was reduced and
cyanobacteria became the main carbonate producers (Săsăran et
al., 2013). However, high-energy bioclastic packstones-grainstones in the
uppermost part of the interval at 865 m contain foraminifera and dasycladalean
algae (Fig. 3
). As the
carbonate platform was prograding, the accomodation space was reduced and
cyanobacteria became the main carbonate producers (Săsăran et
al., 2013). However, high-energy bioclastic packstones-grainstones in the
uppermost part of the interval at 865 m contain foraminifera and dasycladalean
algae (Fig. 3 ![]() ). Pseudocymopolia jurassica and Salpingoporella
praturloni are commonly associated with well oxygenated, shallow-water, subtidal environments (Bucur & Săsăran,
2005). Other species (Coscinoconus
campanellus, C. cherchiae and Nautiloculina
cf. broennimanni) usually also reflect similar palaeoecological
conditions (Arnaud-Vanneau & Peybernčs,
1978; Arnaud-Vanneau,
1980; Simmons 1990). In addition, presence of large dasycladalean
algae (Pseudocymopolia jurassica, Salpingoporella
praturloni) in this
lithostratigraphic interval indicates a shallow-water environment. These
sediments were probably carried through a network of tidal channels crossing the
intertidal and supratidal areas where cyanobacteria were growing in restricted
conditions (Fig. 7
). Pseudocymopolia jurassica and Salpingoporella
praturloni are commonly associated with well oxygenated, shallow-water, subtidal environments (Bucur & Săsăran,
2005). Other species (Coscinoconus
campanellus, C. cherchiae and Nautiloculina
cf. broennimanni) usually also reflect similar palaeoecological
conditions (Arnaud-Vanneau & Peybernčs,
1978; Arnaud-Vanneau,
1980; Simmons 1990). In addition, presence of large dasycladalean
algae (Pseudocymopolia jurassica, Salpingoporella
praturloni) in this
lithostratigraphic interval indicates a shallow-water environment. These
sediments were probably carried through a network of tidal channels crossing the
intertidal and supratidal areas where cyanobacteria were growing in restricted
conditions (Fig. 7 ![]() ). The Upper Jurassic-Lower Cretaceous foraminiferal
assemblage from Piatra Craiului Massif comprises mainly subtropical forms (Everticyclammina,
Labyrinthina, Nautiloculina, Protopeneroplis,
and Pseudocyclammina) with very few
tropical exceptions (Bramkampella, Paracoskinolina,
and Redmondoides) (Kuznetsova et
al., 1996).
). The Upper Jurassic-Lower Cretaceous foraminiferal
assemblage from Piatra Craiului Massif comprises mainly subtropical forms (Everticyclammina,
Labyrinthina, Nautiloculina, Protopeneroplis,
and Pseudocyclammina) with very few
tropical exceptions (Bramkampella, Paracoskinolina,
and Redmondoides) (Kuznetsova et
al., 1996).
|
Figure 7: Conceptual depositional model of the Upper Jurassic-Lower Cretaceous carbonate succession from the Piatra Craiului Massif (1: Fore-reef rudstone facies; 2: Platform margin coral-microbial bioconstructions; 3: Platform margin sand-bar grainstone facies; 4: Intertidal pond fenestral wackestone facies; 5: Tidal channel grainstone facies; 6: Supratidal marsh, mudstone-wackestone facies; 7: Coral fragments; 8: Corals; 9: Echinoderm plates; 10: Microbial crust fragments; 11: Microbial crusts; 12: Pelecypods; 13: Dasycladalean algae; 14: Benthic foraminifera; 15: Cyanobacteria nodules; 16: Gastropods; 17: Fenestral structures; 18: Peloids and black pebbles). |
This study is a contribution to the CNCS research project financed by the PN-II-ID-PCE-2011-3-0025 grant. The authors are grateful to Mr. Mircea Vergheleţ (Director) and the scientific council of the Piatra Craiului National Park for granting permission to work in the studied area. Last but not least, R.W. Scott, F. Schlagintweit and an anonymous reviewer are thanked for their constructive comments on the original manuscript.
Altiner D. (1991).- Microfossil biostratigraphy (mainly foraminifers) of the Jurassic-Lower Cretaceous successions in north-western Anatolia (Turkey).- Geologica Romana, Roma, vol. 27, p. 167-213.
Arnaud-Vanneau A. (1980).- Micropaléontologie, paléoécologie et sédimentologie d'une plate-forme carbonatée de la marge passive de la Téthys : L'Urgonien du Vercors septentrional et de la Chartreuse.- Thčse Doctorat es Sciences; Geologie Alpine, Grenoble, 3 volumes, 876 p.
Arnaud-Vanneau A., Boisseau T. & Darsac C. (1988).- Le genre Trocholina Paalzow 1922 et ses espčces au Crétacé.- Revue de Paléobiologie, Genčve, Volume Spécial no. 2 (Benthos '86), p. 353-377.
Arnaud-Vanneau A. & Peybernčs B. (1978).- Les représentants éocrétacés du genre Nautiloculina Mohler, 1938 (Foraminifera, Fam. Lituolidae ?) dans les chaînes subalpines septentrionales (Vercors) et les Pyrénées franco-espagnoles.- Géobios, Lyon, vol. 11, p. 67-81.
Azéma J., Chabrier G., Fourcade É. & Jaffrezo M. (1977).- Nouvelles données micropaléontologiques, stratigraphiques et paléogéographiques sur le Portlandien et le Néocomien de Sardaigne.- Revue de Micropaléontologie, Paris, vol. 20, no. 3, p. 125-139.
Banner F.T. & Highton J. (1990).- On Everticyclammina Redmond (foraminifera), especially E. kelleri.- Journal of Micropalaeontology, London, vol. 9, no. 1, p. 1-14.
Banner F.T., Simmons M.D. & Whittaker J.E. (1991).- The Mesozoic Chrysalidinidae (Foraminifera, Textulariacea) of the Middle East: The Redmond (ARAMCO) taxa and their relatives.- Bulletin of the British Museum of Natural History, London, (Geology), vol. 47, p. 101-152.
Banner F.T. & Whittaker J.E. (1991).- Redmond's "new lituolid foraminifera" from the Mesozoic of Saudi Arabia.- Micropaleontology, New York, vol. 37, no. 1, p. 41-59.
Bassoullet J.-P. (1997).- Les grands Foraminifčres. In: Cariou E. & Hantzpergue P. (eds.), Biostratigraphie du Jurassique ouest-européen et méditerranéen : Zonations parallčles et distribution des microfossiles.- Bulletin des Centres de Recherches Exploration-Production elf-Aquitaine, Pau, Mémoire, vol. 17, p. 293-304.
Bassoullet J.-P., Bernier P., Conrad M.A., Deloffre R. & Jaffrezo M. (1978).- Les algues dasycladales du Jurassique et du Crétacé.- Geobios, Lyon, Mémoire Spécial no. 2, 330 p.
Beccaro P. & Lazăr I. (2007).- Oxfordian and Callovian radiolarians from the Bucegi Massif and Piatra Craiului Mountains (Southern carpathians, Romania).- Geologica Carpathica, Bratislava, vol. 58, no. 4, p. 305-320.
Bernier P. (1971).- Deux nouvelles algues Dasycladacées du Jurassique supérieur du Jura méridional.- Geobios, Lyon, vol. 4, no. 3, p. 173-184.
Boisseau T. (1987).- La plate-forme jurassique et sa bordure subalpine au Berriasien-Valanginien (Chartreuse-Vercors). Analyse et corrélation avec les séries de bassin.- Thčse de Doctorat, Université du Grenoble, 413 p.
Brönnimann P. (1966).- Pseudotextulariella courtionensis, n. sp., from the Valanginian of well Courtion 1, Courtion, Canton of Fribourg, Switzerland.- Archives des Sciences, Genčve, vol. 19, no. 3, p. 265-278.
Bruchental C., Bucur I.I., Cociuba I. & Hebriştean A.M. (2014).- Sea-level changes and sedimentary response across the Barremian-Early Aptian in Pădurea Craiului Mountains, Romania.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 9, no. 2, p. 33-46.
Bruni R., Bucur I.I. & Préat A. (2007).- Uppermost Jurassic-lowermost Cretaceous carbonate deposits from Fara San Martino (Maiella, Italy): Biostratigraphic remarks.- Studia UBB Geologia, Cluj Napoca, vol. 52, no. 2, p. 45-54.
Bucur I.I. (1978).- Microfacies of white limestones from the northern part of the Piatra Craiului Massif. Biostratigraphical remarks.- Dări de Seamă ale Şedinţelor Institutului Geologic al României, Bucureşti, vol. 64 (for 1976-1977), p. 89-105 [in Romanian].
Bucur I.I. (1980).- Rhaxella sorbyana (Blake) in Oxfordian radiolarites from the Piatra Craiului Massif.- Dări de Seamă ale Şedinţelor Institutului Geologic al României, Bucureşti, vol. 65 (for 1977-1978), p. 31-35.
Bucur I.I. (1985).- Sur la présence des cystes reproducteurs dans les ramifications de Macroporella praturloni et quelques considérations concernant l'évolution de la cladosporie chez les dasycladales fosiiles.- Evolution Adaptation, Cluj Napoca, p. 101-109.
Bucur I.I. (1988).- Les foraminifčres du Crétacé inférieur (Berriasien-Valanginien) de la zone de Reşiţa-Moldova Nouă (Carpathes Méridionales, Roumanie). Remarques biostratigraphiques.- Revue de Paléobiologie, Genčve, Volume Spécial no. 2 (Benthos '86), p. 379-389.
Bucur I.I. (1993).- Remarks on Pseudocymopolia jurassica (Dragastan, 1968). In: Barattolo F., De Castro P. & Parente M. (eds.), Studies on fossil benthic algae.- Bolletino de la Societa Paleontologica Italiana, Modena, Special Volume 1, p. 69-80.
Bucur I.I. (1997).- Representatives of the genus Protopeneroplis (Foraminifera) in the Jurassic and Lower Cretaceous deposits in Romania. Comparisons with other regions of the Tethyan Area.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 1, p. 65-74.
Bucur I.I. (1999).- Stratigraphic significance of some skeletal algae (Dasycladales, Caulerpales) of the Phanerozoic. In: Farinacci A. & Lord A.R. (eds.), Depositional episodes and bioevents.- Palaeopelagos Special Publication, Rome, vol. 2, p. 53-104.
Bucur I.I., Conrad M. & Radoičić R. (1995).- Foraminifers and calcareous algae from Valanginian limestones in the Jerma river canyon, Eastern Serbia.- Revue de Paléobiologie, Genčve, vol. 14, no. 2, p. 349-377.
Bucur I.I., Dragastan O., Lazăr I., Săsăran E. & Popa M. (2011).- Mesozoic algae bearing deposits from Hăghimaş Mountains (Bicaz Valley Area). In: Bucur I.I. & Săsăran E. (eds.), Calcareous algae from romanian Carpathians.- Field Trip Guidebook, 10th International Symposium on Fossil Algae, Cluj University Press, Cluj Napoca, p. 137.
Bucur I.I., Hoffmann M. & Kołodziej B. (2005).- Uppermost Jurassic-Lowermost Cretaceous benthic algae from Tethys and the European Platform. A case study from Poland.- Revista Espańola de Micropaleontologia, Madrid, vol. 37, no. 1, p. 105-129.
Bucur I.I. & Săsăran E. (2005).- Relationship between algae and paleoenvironment: An Early Cretaceous case study, Trascău Mountains, Romania.- Facies, Erlangen, vol. 51, p. 274-286.
Bucur I.I. & Săsăran E. (2012).- Large dasycladalean algae from Upper Jurassic limestone deposits of the Apuseni Mountains (Romania) - habitat and depositional environment.- Geodiversitas, Paris, vol. 34, no. 1, p. 219-239.
Bucur I.I., Săsăran E., Iacob R., Ichim C. & Turi V. (2009).- Upper Jurassic shallow-water carbonate deposits from some Carpathian areas: New micropaleontological results. In: Proceedings of the 8th Symposium of IGCP 506, Marine and non-marine Jurassic: Global correlation and major global events, Bucharest (Romania).- Abstracts Volume, University of Bucharest, p. 13-14.
Bucur I.I., Senowbari-Daryan B. & Abate B. (1996).- Remarks on some foraminifera from the Upper Jurassic (Tithonian) reef limestone of Madonie Mountains (Sicily).- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 35, no. 1, p. 65-80.
Carozzi A. (1954).- Survivance du genre Teutloporella dans le Jurassique supérieur du Grand Salčve.- Archives des Sciences, Genčve, vol. 7, p. 319-324.
Carras N., Conrad M.A. & Radoičić R. (2006).- Salpingoporella, a common genus of Mesozoic Dasycladales (calcareous green algae).- Revue de Paléobiologie, Genčve, vol. 25, no. 2, p. 457-517.
Charollais J., Broennimann P. & Zaninetti L. (1966).- Troisičme note sur les foraminifčres du Crétacé inférieur de la région genevoise. Remarques stratigraphiques et description de Pseudotextulariella salevensis, n. sp. ; Haplophragmoides joukowskyi, n. sp. ; Citaella? favrei, n. sp.- Archives des Sciences, Genčve, vol. 19, no. 1, p. 23-48.
Chiocchini M., Farinacci A., Mancinelli A., Molinari V. & Potetti M. (1994).- Biostratigrafia a foraminiferi, dasicladali e calpionelle delle successioni carbonatiche mesozoiche dell'Appennino centrale (Italia).- Studi Geologici Camerti, Camerino, Volume Speciale, Biostratigrafia dell'Italia centrale, p. 9-128.
Chiocchini M., Mancinelli A. & Marcucci C. (1988).- Distribution of benthic foraminifera and algae in the Lazium-Abruzzi carbonate platform facies (Central Italy) during Upper Malm-Neocomian.- Revue de Paléobiologie, Genčve, Volume Spécial no. 2 (Benthos '86), p. 219-227.
Cuvillier J., Foury G. & Pignatti-Morano A. (1968).- Foraminifčres nouveaux du Jurassique supérieur du Val Celina (Frioul occidental, Italie).- Geologica Romana, Roma, vol. 7, p. 141-156.
Darga R. & Schlagintweit F. (1991).- Mikrofazies, paläontologie und stratigraphie der Lerchkogelkalke (Tithon-Berrias) des Dietrichshorns (Salzburger Land, Nördliche Kalkalpen).- Jahrbuch der Geologischen Bundesanstalt, Wien, vol. 134, no. 2, p. 205-226.
Darsac C. (1983).- La plateforme berriaso-valanginienne du Jura méridional aux massifs subalpins (Ain, Savoie). Sédimentologie, minéralogie, stratigraphie, paléogéographie, micropaléontologie.- Thčse de Doctorat, Université du Grenoble, 319 p.
Dieni I. & Radoičić R. (1999).- Clypeina dragastani sp. nov.; Salpingoporella granieri sp. nov. and other dasycladalean algae from the Berriasian of Eastern Sardinia.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 2, p. 105-123.
Dimitrescu R., Patrulius D. & Popescu I. (1971).- Geological map of Romania, 1:50.000, sheet 110c.- Institutul Geologic şi Geofizic al României, Bucureşti [in Romanian].
Dimitrescu R., Popescu I. & Schuster C.A. (1974).- Geological map of Romania, 1:50.000, sheet 110a.- Institutul Geologic şi Geofizic al României, Bucureşti [in Romanian].
Dragastan O.N. (1975).- Upper Jurassic and Lower Cretaceous microfacies from the Bicaz Valley basin (East Carpathians).- Mémoires de l'Institut de Géologie et Géophysique, Bucarest, vol. 21, p. 1-87.
Dragastan O.N. (2010).- The Getic Carbonate Platform. Jurassic-Lower Cretaceous stratigraphy, paleogeographical reconstruction, provinces and biodiversity.- Bucharest University Press, 621 p.
Dya M. (1992).- Mikropaleontologische und fazielle Unterschungen in Oberjura zwischen Salzburg und Lofer.- PhD Thesis, University of Berlin, 137 p.
Egger J.G. (1902).- Der Bau der Orbitolinen und verwandter Formen.- Abhandlungen der Mathematisch-Physikalischen Klasse der Königlisch Bayerischen Akademie der Wissenschaften, München, Band XXI, Abh. 3, p. 577-600.
Farinacci A. & Radoičić R. (1964).- Correlazione fra serie giurese e cretacee dell'Appennino centrale e della Dinaridi externe.- La Ricerca Scientifica, Roma, vol. 2A, no. 7, p. 269-300.
Farinacci A. & Radoičić R. (1991).- Late Jurassic-Early Cretaceous dasycladales (green algae) from the Western Pontides, Turkey.- Geologica Romana, Roma, vol. 27, p. 135-165.
Fourcade É. (1970).- Le Jurassique et le Crétacé aux confins des chaînes bétiques et ibériques (Sud-Est de l'Espagne).- Thčse de Doctorat, Paris, 3 volumes, 255 p.
Foury G. & Moullade M. (1966).- Orbitolinidae nouveaux du Barrémien (facičs urgonien) des Alpilles (Bouches-du-Rhône).- Revue de Micropaléontologie, Paris, vol. 8, no. 4, p. 249-257.
Foury G. & Vincent E. (1967).- Morphologie et répartition stratigraphique du genre Kilianina Pfender (Foraminifčre).- Eclogae geologicae Helvetiae, Bâle, vol. 60, no. 1, p. 33-51.
Gorbatchik T.N. (1971).- On Early Cretaceous foraminifera of the Crimea.- Akademia Nauk, Voprosi Micropaleontologija, Moscow, vol. 14, p. 125-139.
Granier B. (1987).- Le Crétacé inférieur de la Costa Blanca entre Busot et Altea, Alicante (Espagne) : Biostratigraphie, sédimentologie, évolution tectono-sédimentaire.- Thčse, Docteur de l'Université Paris VI (nouveau régime); Mémoires des Sciences de la Terre, Université Pierre et Marie Curie, Paris, no. 87-49, vol. I: 281 p.; vol. II: 54 Pls.
Granier B. (1994).- Early Valanginian unconformities in Central Atlantic and Western Mediterranean areas.- Géologie Méditerranéenne, Marseille, t. XXI, p. 81-85.
Granier B. & Bucur I.I. (2011).- Stratigraphic ranges of some Tithonian-Berriasian benthic foraminifers and Dasycladales. Re-evaluation of their use in identifying this stage boundary in carbonate platform settings. In: Grosheny D., Granier B. & Sanders N. (eds.), Platform to basin correlations in Cretaceous times.- Boletín del Instituto de Fisiografía y Geología, Rosario, vol. 79-81, p. 9-10.
Granier B., Clavel B., Charolais J. & Weidmann M. (2014).- Latest Jurassic-Early Cretaceous dasycladalean algae (Chlorophyta) from the Morand drilling at Montricher (Canton of Vaud, Switzerland).- Acta Paleontologica Romaniae, Cluj Napoca, vol. 10, no. 1-2, p. 25-38.
Granier B. & Deloffre R. (1993).- Inventaire critique des algues dasycladales fossiles. IIş Partie - Les algues dasycladales du Jurassique et du Crétacé.- Revue de Paléobiologie, Genčve, vol. 12, no. 1, p. 19-65.
Granier B., Peebles R.G. & Sander N.J. (2006).- Discussion on the Jurassic evolution of the Arabian carbonate platform edge in the central Oman Mountains, Geological Society of London Journal, vol. 162, 2005, pp. 349-362.- Journal of the Geological Society, vol. 163, no. 6, p. 1047-1050.
Granier B., Toland C., Gčze R., Azar D. & Maksoud S. (2016).- Some steps toward a new story for the Jurassic-Cretaceous transition in Mount Lebanon.- Carnets Geol., Madrid, vol. 16, no. 8, p. 247-269.
Granier B., Virgone A., Busnardo R. & Bulot L.G. (1995).- Des calpionelles dans l'Hauterivien supérieur. Découverte exceptionnelle ŕ Busot (Alicante, Espagne).- Comptes-Rendus de l'Académie des Sciences, Paris, (IIa), t. 321, p. 1179-1186.
Grădinaru M. (2011).- Jurassic hardgrounds from Bucegi Mountains (western slope) and Almăjului Mountains (Sviniţa).- PhD Thesis (in Romanian), Universitatea din Bucureşti, 200 p.
Grădinaru M., Lazăr I., Bucur I.I., Grădinaru E., Săsăran E., Ducea M.N. & Andrăşanu A. (2016).- The Valanginian history of the eastern part of the Getic Carbonate Platform (Southern Carpathians, Romania): Evidence for emergence and drowning of the platform.- Cretaceous Research, vol. 66, p. 11-42.
Henson F.R.S. (1948).- Larger imperforate Foraminifera of south-western Asia. Families Lituolidae, Orbitolinidae and Meandropsinidae.- British Museum (Natural History), London, 126 p.
Hottinger L. (1967).- Foraminifčres imperforés du Mésozoďque marocain.- Notes et Mémoires du Service géologique, Rabat, vol. 209, 179 p.
Hughes W.G. (2000).- Saudi Arabian Late Jurassic and Early Cretaceous agglutinated foraminiferal associations and their application for age, paleoenvironmental interpretation, sequence stratigraphy and carbonate reservoir architecture. In: Hart M.B., Kaminski M.A. & Stuart C.W. (eds.), Proceedings of the Fifth International Workshop on agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 7, p. 149-165.
Ivanova D. (2000).- Middle Callovian to Valanginian microfossil biostratigraphy in the West Balkan Mountain, Bulgaria (SE Europe).- Acta Paleontologica Romaniae, Cluj Napoca, vol. 2 (for 1999), p. 231-236.
Ivanova D. & Kolodziej B. (2010).- Late Jurassic-Early Cretaceous foraminifera from Štramberk-type limestones, Polish Outer Carpathians.- Studia UBB Geologia, Cluj Napoca, vol. 55, no. 22, p. 3-31.
Jaffrezo M. (1970).- Précisions stratigraphiques et tectoniques sur le lobe de Fontjoncouse de la nappe de Corbičres orientales.- Bulletin de la Société géologique de France, Paris, (7e série), vol. 12, no. 1, p. 65-77.
Jaffrezo M. (1980).- Les formations carbonatées des Corbičres (France) du Dogger ŕ l'Aptien : Micropaléontologie, stratigraphie, biozonation, paléoécologie. Extension des résultats ŕ la Mésogée.- Thčse de Doctorat d'État, Université Pierre et Marie Curie, Paris, 655 p.
Jekelius E. (1923).- Das Gebirge von Braşov.- Anuarul Institutului Geologic al României, Bucureşti, vol. 19, p. 308-406.
Kaminski M. (2014).- The year 2010 classification of the agglutinated foraminifera. In: Alegret L. & Kaminski M. (eds.), Advances in agglutinated foraminiferal research: The Ninth International Workshop on Agglutinated Foraminifera, IWAF-9.- Micropaleontology, London, vol. 60, no. 1, p. 89-108.
Kobayashi F. & Vuks V.J.A. (2006).- Tithonian-Berriasian foraminiferal faunas from the Torinosu-type calcareous blocks of the southern Kanto Mountains, Japan: Their implications for post-accretionary tectonics of Jurassic to Cretaceous terranes.- Geobios, Lyon, vol. 39, p. 833-843.
Kołodziej B. & Decrouez D. (1997).- Foraminifera and incertae sedis from the Štramberk-type limestone (Tithonian-Berriasian). Preliminary results.- Revue de Paléobiologie, Genčve, vol. 16, no. 1, p. 145-150.
Kuznetsova K.I., Grigelis A.A., Adjamian J., Jarmakani E. & Hallaq L. (1996).- Zonal stratigraphy and Foraminifera of the Tethyan Jurassic (Eastern Mediterranean).- Gordon and Breach Publishers, Amsterdam, 255 p.
Lazăr I. & Grădinaru M. (2013).- Paleoenvironmental context and paleoecological significance of unique agglutinated polychaete worm tube-ferruginous microstromatolite assemblages from the Middle Jurassic of the Southern Carpathians (Romania).- Facies, Erlangen, vol. 60, no. 2, p. 515-540.
Loeblich A.R. & Tappan H. (1988).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, 970 p.
Luperto Sinni E. & Masse J.-P. (1986).- Données nouvelles sur la stratigraphie des calcaires de plate-forme du Crétacé inférieur du Gargano (Italie méridionale).- Revista Italiana di Paleontologia e Stratigrafia, Roma, vol. 92, no. 1, p. 33-66.
Mancinelli A. & Coccia B. (1999).- Le Trocholine dei sedimenti mesozoici di piattaforma carbonatica dell'Appennino-centro-meridionale (Abruzzo e Lazio).- Revue de Paléobiologie, Genčve, vol. 18, no. 1, p. 147-171.
Meinhold G., Kostopoulos D., Reischmann T., Frei D. & BouDagher-Fadel K.M. (2009).- Geochemistry, provenance and stratigraphic age of metasedimentary rocks from the eastern Vardar suture zone, northern Greece.- Palæogeography, Palæoclimatology, Palæoecology, vol. 277, p. 199-225.
Mészáros N. & Bucur I.I. (1980).- Oxfordian nannoplankton from the Piatra Craiului Massif.- Muzeul Bruckental Ştiinţe Naturale, vol. 24, p. 73-77 [in Romanian].
Michetiuc M., Catincuţ C. & Bucur I.I. (2008).- Microfacies and microfossils of the Lower Cretaceous limestones from the southern part of Vâlcan Mountains.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 6, p. 217-227.
Mircescu C.V., Bucur I.I. & Săsăran E. (2014).- Dasycladalean algae from Upper Jurassic-Lower Cretaceous limestones of Piatra Craiului Massif (South Carpathians, Romania) and their relationship to paleoenvironment.- Studia UBB Geologia, Cluj Napoca, vol. 59, no. 1 (for 2013), p. 5-27.
Mohler W. (1938).- Mikropaläontologische Untersuchungen in der nordschweizerischen Juraformation.- Abhandlungen der Schweizerischen Paläeontologischen Gesellschaft, vol. 60, p. 1-53.
Moshammer S. & Schlagintweit F. (1999).- The Ernstbrunn limestone (Lower Austria): New data on biostratigraphy and applied geology.- Abhandlungen der Geologischen Bundesanstalt, vol. 56, no. 2, p. 553-565.
Neagu T. (1994).- Early Cretaceous Trocholina group and some related genera from Romania. Part I.- Revista Espańola de Micropaleontologia, Madrid, vol. 26, no. 3, p. 117-143.
Nikitenko B.L., Reolid M. & Glinskikh L. (2013).- Ecostratigraphy of benthic foraminifera for interpreting Arctic record of Early Toarcian biotic crisis (northern Siberia, Russia).- Palæogeography, Palæoclimatology, Palæoecology, vol. 376, p. 200-212.
Oncescu N. (1943).- Région de Piatra Craiului-Bucegi. Étude géologique.- Anuarul Institutului Geologic al României, Bucureşti, vol. 9, p. 3-124.
Patrulius D. (1969).- Geology of Bucegi Massif and Dâmbovicioara Couloir.- Editura Academiei Republicii Socialiste România, Bucureşti, 222 p. [in Romanian].
Patrulius D. (1976).- Upper Jurassic-Lower Cretaceous carbonate rocks in the eastern part of the Getic carbonate platform and the adjacent flysch troughs. In: Patrulius D., Drăgănescu A., Baltreş A., Popescu B. & Rădan S. (eds.), Carbonates rocks and evaporates - Guidebook.- International Colloquium on Carbonates Rocks and Evaporites, Institute of Geology and Geophysics, Bucharest, Guidebook series 15, p. 71-82.
Patrulius D., Antonescu A., Avram E., Baltreş A., Dumitrică P., Iordan M., Iva M., Morariu A., Pop G., Popa E. & Popescu I. (1980).- Complex petrological and biostratigraphical study of the Jurassic and Neocomian formations from the Romanian Carpathians and Dobrogea for the evaluation of their mineral resources potential. The Leaota-Braşov-Perşani Massif sector.- Fieldwork report, Institutul Geologic şi Geofizic al României, Bucureşti, 218 p. [in Romanian].
Patrulius D., Dimitrescu R. & Popescu I. (1971).- Geological map of Romania, 1:50.000, sheet 110d.- Institutul Geologic şi Geofizic al României, Bucureşti [in Romanian].
Pélissié T., Peybernčs B. & Ray J. (1984).- Les grands foraminifčres benthiques du Jurassique moyen/supérieur du Sud-Ouest de la France (Aquitaine, Causses, Pyrénées). Intéręt biostratigraphique paléoécologique et paléobiogéographique.- Bulletin du Centre de Recherches Exploration-Production elf-Aquitaine, Pau, Mémoire, vol. 6, p. 479-489.
Peybernčs B. (1976).- Le Jurassique et le Crétacé inférieur des Pyrénées franco-espagnoles entre la Garonne et la Méditerranée.- Thčse de Doctorat d'État, Université Paul Sabatier, Toulouse, 459 p.
Pfender J. (1938).- Les foraminifčres du Valanginien provençal.- Bulletin de la Société géologique de la France, Paris, (5e série), vol. 8, no. 3-4, p. 231-242.
Pleş G., Bucur I.I. & Păcurariu A. (2015).- Foraminiferal assemblages and facies associations in the Upper Jurassic carbonates from Ardeu Unit (Metaliferi Mountains, Romania).- Acta Paleontologica Romaniae, Cluj Napoca, vol. 11, no. 2, p. 43-57.
Pleş G., Mircescu C.V., Bucur I.I. & Săsăran E. (2013).- Encrusting micro-organisms and microbial structures in Upper Jurassic limestones from the Southern Carpathians (Romania).- Facies, Erlangen, vol. 59, p. 19-48.
Pop G. & Bucur I.I. (2001).- Upper Jurassic and Lower Cretaceous sedimentary formations from the Vâlcan Mountains (South Carpathians).- Studia UBB Geologia, Cluj Napoca, vol. XLVI, no. 2, p. 77-94.
Popescu I. (1966).- Contributions to the knowledge of the stratigraphy and geological structure of the Piatra Craiului Massif.- Dări de Seamă ale Şedinţelor Institutului Geologic al României, Bucureşti, vol. 52, no. 2, p. 157-176 [in Romanian].
Redmond C.D. (1964).- Lituolid foraminifera form the Jurassic and the Cretaceous of Saudi Arabia.- Micropaleontology, New York, vol. 10, p. 405-414.
Reiss Z. & Hottinger L. (1984).- The Gulf of Aqaba: Ecological micropaleontology. Ecological studies.- Springer-Verlag, Berlin-Heidelberg, vol. 50, 534 p.
Reolid M., Rodriguez-Tovar F.J., Nagy J. & Olóriz F. (2008a).- Benthic foraminiferal morphogroups of mid to outer shelf environments of the Late Jurassic (Prebetic Zone, southern Spain): Characterization of biofacies and environmental siginificance.- Palæogeography, Palæoclimatology, Palæoecology, vol. 261, p. 280-299.
Reolid M., Rodriguez-Tovar F.J., Nagy J. & Olóriz F. (2008b).- Foraminiferal assemblages as palaeoenvironmental bioindicators in Late Jurassic epicontinental platforms: Relation with trophic conditions.- Acta Palaeontologica Polonica, Warsaw, vol. 53, p. 705-722.
Rigaud S., Blau J., Martini R. & Rettori R. (2013).- Taxonomy and phylogeny of the Trocholinidae (Involutinina).- Journal of Foraminiferal Research, Lawrence, vol. 43, no. 4, p. 317-339.
Savelieva J.N., Feodorova A.A., Shurekova O.V. & Arkadiev V. (2014).- Integrated palaeontological characteristics (ammonites, ostracods, foraminifers, dinocysts) of the Berriasian deposits of central Crimea.- Volumina Jurassica, Warsaw, vol. 12, no. 1, p. 129-162.
Săndulescu M., Popescu I., Săndulescu J., Mihăilă N. & Schuster C.A. (1972).- Geological map of Romania, 1:50.000, sheet 110b.- Institutul Geologic şi Geofizic al României, Bucureşti [in Romanian].
Săsăran E. & Bucur I.I. (2001).- Upper Jurassic-Lower Cretaceous microbialites and calcareous algae from the Štramberk like limestones in Cheile Turzii area. In: Bucur I.I., Filipescu S. & Săsăran E. (eds.), Algae and carbonate platforms in western part of Romania. Fieldtrip Guidebook, 4th Regional Meeting of IFAA.- Cluj University Press, p. 191-207.
Săsăran E., Pleș G., Mircescu C.V. & Bucur I.I. (2013).- Peritidal cyclical sequences of Kimmeridgian - Beriassian - ? Valanginian limestones from Piatra Craiului Massif (Romania); the role of microbialites and rivulariacean-type cyanobacteria. In: Proceedings of the 11 th Workshop on Alpine Geological Studies & 7th IFAA, Schladming-Dachstein (Austria).- Abstracts Volume, Berichte Geologische B.A., Vienna, p. 116-117.
Schlagintweit F. (2011).- The dasycladalean algae of the Plassen Carbonate Platform (Kimmeridgian-Early Berriasian): taxonomic inventory and palaeogeographical implications within the Northern calcareous Alps (Austria, p. p. Germany).- Geologia Croatica, Zagreb, vol. 64, no. 3, p. 185-206.
Schlagintweit F. (2012).- Mohlerina basiliensis (Mohler, 1938): A Middle Jurassic-Early Cretaceous facultative (?) epilithic benthic foraminifer.- Facies, Erlangen, vol. 58, no. 4, p. 637-647.
Schlagintweit F. & Ebli O. (1999).- New results on microfacies, biostratigraphy and sedimentology of Late Jurassic-Early Cretaceous Alps, Part I: Tressenstein Limestone, Plassen Formation.- Geologie ohne Grenzen Festschrift 150 Jahre Geologische Bundesanstalt, Vienna, vol. 56, no. 2, p. 379-418.
Schlagintweit F., Gawlick H.J. & Lein R. (2005).- Micropaleontology and biostratigraphy of the Plassen carbonate platform of the type locality (Upper Jurassic to Lower Cretaceous, Salzkammergut, Austria).- Journal of Alpine Geology, Vienna, vol. 47, p. 11-102.
Schroeder R., Clavel B., Conrad A.M., Zaninetti L., Busnardo R., Charollais J. & Cherchi A. (2000).- Corrélations biostratigraphiques entre la coupe d'Organya (Pyrénées catalanes, NE de l'Espagne) et le Sud-Est de la France pour l'intervalle Valanginien-Aptien.- Treballs del Museo de Geologia de Barcelona, vol. 9, p. 5-41.
Schroeder R., Guellal S. & Vila J.M. (1975).- Parurgonina caelinensis Cuvillier, Foury & Pignatti Morano 1968 dans le Malm du Djebel Téioualt (Constantinois, Algérie).- Eclogae geologicae Helvetiae, Bâle, vol. 68, no. 2, p. 319-326.
Senowbari-Daryan B., Bucur I.I. & Abate B. (1994).- Upper Jurassic calcareous algae from the Madonie Mountains, Sicily.- Beiträge Paläontologie, Vienna, vol. 19, p. 227-259.
Septfontaine M. (1974).- Présence de Protopeneroplis trochangulata sp. nov. (Foraminifčres) dans le Crétacé inférieur du Jura méridional et révision de Protopeneroplis Weynschenk, 1950.- Eclogae geologicae Helvetiae, Bâle, vol. 67, no. 3, p. 605-628.
Septfontaine M. (1977).- Bullopora tuberculata (Sollas) et autres foraminifčres fixés du Dogger des Préalpes Médianes - Relation avec le microfacičs.- Archives des Sciences, Genčve, vol. 30, no. 1, p. 65-76.
Septfontaine M. (1988).- Vers une classification évolutive des Lituolides (Foraminifčres) jurassiques en milieu de plate-forme carbonatée.- Revue de Paléobiologie, Genčve, Volume Spécial no. 2 (Benthos '86), p. 229-256.
Simmons M.D. (1990).- Aspects of the micropaleontology and stratigraphy of Cretaceous shelf carbonates from the Oman Mountains.- PhD Thesis, University College of Wales, Swansea, 270 p.
Smout A.H. & Sugden W. (1961).- New information on the foraminiferal genus Pfenderina.- Paleontology, vol. 4, no. 4, p. 581-591.
Sokač B. & Nikler L. (1973).- Calcareous algae from the Lower Cretaceous of the environs of Nikšić, Crna Gora (Montenegro).- Paleontologija jugoslavica, Zagreb, vol. 13, p. 1-57.
Tasli K. (1993).- Micropaléontologie, stratigraphie et environnement de dépôt des séries jurassiques ŕ faciés de la plate-forme de la région de Kale-Gümüshane (Pontides orientales, Turquie).- Revue de Micropaléontologie, Paris, vol. 36, no. 1, p. 45-65.
Tyszka J. (1994).- Response of Middle Jurassic benthic foraminiferal morphogroups to dysoxic/anoxic conditions in the Pieniny Klippen Basin, Polish Carpathians.- Palæogeography, Palæoclimatology, Palæoecology, vol. 110, p. 55-81.
Ungureanu R.R., Săsăran E., Bucur I.I., Ungur C.G. & Mircescu C.V. (2015).- The Berriasian-Valanginian and Aptian deposits from the north-western part of the Piatra Craiului Massif: Stratigraphic relationships, facies and depositional environments.- Acta Paleontologica Romaniae, Cluj Napoca, vol. 11, no. 2, p. 59-74.
Velić I. (2007).- Stratigraphy and palaeobiogeography of Mesozoic benthic Foraminifera of the Karst Dinarides (SE Europe).- Geologia Croatica, Zagreb, vol. 60, no. 1, p. 1-113.
Velić I. & Sokač B. (1983).- Stratigraphy of the Lower Cretaceous index fossils in the Karst Dinarides (Yugoslavia).- Zitteliana - Abhandlungen der Bayerischen Staatssammlung für Paläontologie, München, vol. 10, p. 485-491.
Weynschenk R. (1951).- Two new foraminifera from the Dogger and Upper Triassic of the Sonnwend Mountains of Tyrol.- Journal of Paleontology, Tulsa, vol. 25, p. 793-795.
Yabe H. & Hanzawa S. (1926).- Choffatella Schlumberger and Pseudocyclammina, a new genus of arenaceous foraminifera.- Science Reports of the Tôhoku Imperial University, Sendai, (Series 2), vol. 9, no. 1, p. 9-11.
Yokoyama M. (1890).- Foraminiferen aus dem Kalksteine von Torinosu und Kompira.- Denkschriften der Kaiserlichen Akademie der Wissenschaften, Wien, (Mathematisch-Naturwissenschaftliche Klasse), vol. 57, p. 26-27.
Zaninetti L., Salvini-Bonnard G. & Decrouez D. (1987).- Montsalevia, n. gen. (Montsaleviidae, n. fam. Foraminifčre), dans le Crétacé inférieur (Berriasien moyen-Valanginien) du Mont Salčve et du Jura Méridional (Haute-Savoie, France). Note préliminaire.- Revue de Paléobiologie, Genčve, vol. 6, no. 1, p. 165-168.