◄ Carnets Geol. 16 (21) ►
![]()
Contents
[Introduction]
[Paleogeography]
[Lithostratigraphy]
[Biostratigraphy]
[Regional correlations]
[Conclusions]
[Systematic paleontology] and ...
[Bibliographic references]
Precision Stratigraphy Associates, 149 West Ridge Road, Cleveland OK
74020 (U.S.A.)
The University of Tulsa, 800 South Tucker Drive, Tulsa OK 74104 (U.S.A.)
The University of Tulsa, 800 South Tucker Drive, Tulsa OK 74104 (U.S.A.)
Oklahoma State University, School of Geology, 105 Noble Research Center, Stillwater OK 74078 (U.S.A.)
Research Institute of Petroleum Exploration & Development-Langfang, Hebei 065007 (P.R. China)
Published online in final form (pdf) on November 11, 2016
[Editor: Bruno Granier]
![]()
� Barremian-Albian caprinids biostratigraphic zones are revised and integrated with ammonites and benthic foraminifers.
� New caprinid rudist species are the key to revising long-held correlations of Albian strata on the Comanche shelf, Texas.
� On the San Marcos Arch, central Texas, the shallow shelf Person Formation is the upper unit of the Fredericksburg Group.
� The Person underlies the basal Washita Group sequence boundary Al Sb Wa1 and the Georgetown Formation.
Rudists were widespread and locally abundant carbonate producers on the Early Cretaceous Comanche Shelf from Florida to Texas, and on Mexican atolls. As members of the Caribbean Biogeographic Province, their early ancestors emigrated from the Mediterranean Province and subsequently evolved independently. Comanchean rudists formed biostromes and bioherms on the shelf interior and at the shelf margin. Carbonate stratigraphic units of the Comanche Shelf record rudist evolution during the Barremian through the Albian ages and an established zonal scheme is expanded.
This study documents new Albian rudist occurrences from the Middle-Upper Albian Fredericksburg and Washita groups in Central and West Texas. Rudists in cores at and directly behind the shelf margin southeast of Austin and San Antonio, Texas, complement the rudist zonation that is integrated with ammonites and foraminifers. These new rudist data test long-held correlations of the Edwards Group with both the Fredericksburg and Washita groups based solely on lithologies. Rudist and foraminifer biostratigraphy indicate that the Edwards Group is coeval with the Fredericksburg not the Washita Group.
In West Texas sections Caprinuloidea romeri occurs at the top of the Fredericksburg Group in the Fort Terrett Formation approximately 3 meters below the iron-stained hardground subaerial exposure surface. It is overlain by the Washita Group with the ammonite Eopachydiscus marcianus Zone. Caprinuloidea romeri is also documented from the type cored section of the Person Formation in the upper part of the Edwards Group. These species allow a precise correlation of the subsurface Person Formation in Central Texas with the Ft. Terrett Formation cropping out in West Texas. This evidence together with foraminifer and ammonite zones demonstrates that the Person Formation is coeval with Fredericksburg units underlying the basal Washita sequence boundary Al Sb Wa1. The upper Albian Washita Group Kimbleia and Mexicaprina caprinid zones overly the Caprinuloidea Zone and the successive Washita mortonicerid ammonite zones.
� Albian;
� caprinuloid biostratigraphy;
�
Comanche shelf;
� Texas.
Scott R.W., Campbell W., Hojnacki R., Wang Y. & Lai X. (2016).- Albian rudist biostratigraphy (Bivalvia), Comanche shelf to shelf margin, Texas.- Carnets Geol., Madrid, vol. 16, no. 21, p. 513-541.
Biostratigraphie de l'Albien au moyen des rudistes (Bivalves), de la plate-forme de Comanche � sa bordure (Texas).- Les rudistes furent des producteurs de calcaires largement r�pandus et localement abondants depuis la Floride jusqu'au Texas sur la plate-forme de Comanche au Cr�tac� inf�rieur, ainsi que dans les atolls mexicains. En tant que repr�sentants de la province biog�ographique carib�enne, leurs anc�tres lointains ont migr� depuis la province m�diterran�enne avant d'�voluer ind�pendamment par la suite. Les rudistes de Comanche ont form� des biostromes et des biohermes � l'int�rieur de la plate-forme et � sa bordure. Les unit�s stratigraphiques carbonat�es de la plate-forme de Comanche enregistrent l'�volution des rudistes depuis le Barr�mien jusqu'� l'Albien. La zonation pr�existante est compl�t�e.
Cette �tude figure de nouvelles occurrences albiennes de rudistes dans les groupes de Fredericksburg et de Washita � l'Albien moyen et sup�rieur au Texas central et occidental. Les rudistes identifi�s dans les sondages � la bordure de plate-forme et juste derri�re, au sud-est d'Austin et de San Antonio (Texas), compl�tent la zonation des rudistes qui est int�gr�e � celles des ammonites et des foraminif�res. Ces nouvelles donn�es concernant les rudistes permettent de tester la robustesse des corr�lations, depuis longtemps �tablies mais bas�es uniquement sur les lithologies, du Groupe d'Edwards avec les groupes de Fredericksburg et de Washita. La biostratigraphie des rudistes et celle des foraminif�res indiquent que le Groupe d'Edwards est �quivalent au Groupe de Fredericksburg, et non � celui de Washita.
Dans les coupes du Texas occidental, Caprinuloidea romeri est pr�sente au sommet du Groupe de Fredericksburg dans la Formation de Fort Terrett, environ 3 m�tres sous la surface d'�mersion sommitale. Cette surface durcie et ferruginis�e est surmont�e par le Groupe de Washita avec la Zone � ammonite Eopachydiscus marcianus. Caprinuloidea romeri est aussi repr�sent�e dans la section carott�e de r�f�rence pour la Formation de Person dans la partie sup�rieure du Groupe d'Edwards. Ces esp�ces permettent une corr�lation pr�cise entre la Formation de Person reconnue en subsurface dans le Texas central et la Formation de Fort Terrett identifi�e � l'affleurement dans le Texas occidental. Cet argument, ainsi que les zones d'ammonites et de foraminif�res, d�montre que la Formation de Person est �quivalente aux unit�s de Fredericksburg sous-jacentes � la limite de s�quence Al Sb Wa1 � la base du Groupe de Washita. Les zones de caprinid�s � Kimbleia et Mexicaprina de l'Albien sup�rieur du Groupe de Washita surmontent la Zone � Caprinuloidea ainsi que les zones successives � ammonites (Mortonic�ratid�s) du Groupe de Washita.
� Albien ;
� biostratigraphie des caprinulo�d�s ;
� plate-forme de Comanche ;
� Texas.
Early Cretaceous was a time of major diversification of the specialized bivalves of the Order Hippuritida Newell (1965) (Skelton, 2013; Steuber et al., 2016). During this time span foraminifers and nannofossils diversified and ocean-water oxygen composition experienced repetitive oceanic anoxic events (OAEs) (Leckie et al., 2002). Repeated shelf flooding stressed shallow shelf communities of colonial corals and rudist bivalves accompanied by rudist iterative evolution (Scott, 1995).
The Barremian-Early Cenomanian Comanche Shelf was a mixed
carbonate-siliciclastic sedimentary province along the southern margin of the
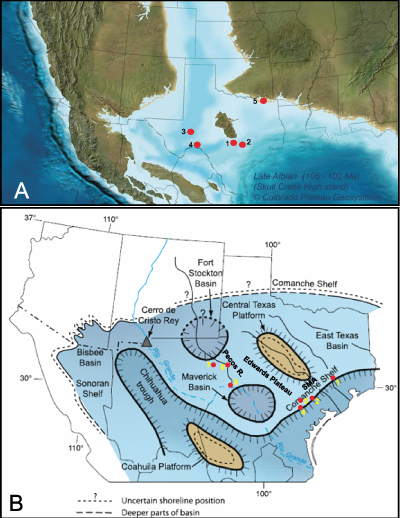
North American continent (Fig. 1.A ![]() ). It extended from Florida westward to
southeastern Arizona (Scott, 1993; Yurewitz et al.,
1993; Mancini and Scott, 2006). Paleoclimate
modeling suggests that Tethyan sea-surface currents flowed westward across the
Comanche Shelf, and surface winds varied seasonally from the southwest in summer
to the west and northwest in the winter (Glancy et
al., 1993).
). It extended from Florida westward to
southeastern Arizona (Scott, 1993; Yurewitz et al.,
1993; Mancini and Scott, 2006). Paleoclimate
modeling suggests that Tethyan sea-surface currents flowed westward across the
Comanche Shelf, and surface winds varied seasonally from the southwest in summer
to the west and northwest in the winter (Glancy et
al., 1993).
Rudists were widespread and locally abundant carbonate producers on the Early Cretaceous Comanche Shelf from Florida to Texas, and they occupied the atolls in Mexico. As members of the Caribbean Biogeographic Province (Kauffman, 1973), their early ancestors emigrated from the Mediterranean Province and subsequently evolved independently (Masse et al., 2007). Comanchean rudists formed biostromes and bioherms on the shelf interior and at the shelf margin and were abundant members of reefal paleocommunities (Scott, 1990). Carbonate stratigraphic units of the Comanche Shelf record rudist evolution during the Hauterivian through the Albian ages (Coogan, 1977) and a zonal scheme has been developed (Scott and Filkorn, 2007).
Rudist
species are relatively well known from Lower Cretaceous outcrops in Central
Texas (Coogan, 1977; Davis-Strickland,
1988; Scott and Filkorn,
2007; Mitchell, 2013a), from shelf-margin cores (Scott,
1990; Mancini
and Scott, 2006), and on the outer Devils River platform along the
southern reaches of the Pecos River in West Texas (Fig. 1.B ![]() )
(Kerans et
al., 1999; Scott and Kerans,
2004). Rudists are less abundant
and less diverse northward/landward on the interior of the Devils River Platform
and eastward across the Edwards Plateau. Rudists are common in the subsurface of
central Texas on the San Marcos Arch carbonate shelf landward of the shelf
margin (Rose, 1972) but unstudied taxonomically.
)
(Kerans et
al., 1999; Scott and Kerans,
2004). Rudists are less abundant
and less diverse northward/landward on the interior of the Devils River Platform
and eastward across the Edwards Plateau. Rudists are common in the subsurface of
central Texas on the San Marcos Arch carbonate shelf landward of the shelf
margin (Rose, 1972) but unstudied taxonomically.
This
study documents new Albian rudist occurrences from the middle-upper Albian Fort
Terrett and Fort Lancaster formations on the upper reaches of the Pecos River,
West Texas, and in the Kainer and Person formations on the San Marcos Arch
(Fig. 2 ![]() ). Rudists are in cores at and directly behind the shelf margin from
south Texas northeastward to offshore Louisiana (Coogan, 1977; Scott,
1990). Landward of the shelf margin in Karnes County southwest of Austin, Texas,
rudist taxa are reported here for the first time from the Edwards Group, which
is comprised of the older Kainer and the younger Person formations (Rose,
1972; Smith et al., 2000) (Fig.
1.B
). Rudists are in cores at and directly behind the shelf margin from
south Texas northeastward to offshore Louisiana (Coogan, 1977; Scott,
1990). Landward of the shelf margin in Karnes County southwest of Austin, Texas,
rudist taxa are reported here for the first time from the Edwards Group, which
is comprised of the older Kainer and the younger Person formations (Rose,
1972; Smith et al., 2000) (Fig.
1.B ![]() ).
).
|
Figure 1: A. Late Albian paleogeographic map in south-central U.S. (Blakey, 2010; � Ron Blakey, Colorado Plateau Geosystems); red dots indicate locations of rudist collections: 1) Karnes County wells and Travis County outcrops; 2) shelf margin cores; 3) Crockett County roadcuts and roadcut in Pecos County; 4) Pecos River outcrops in Val Verde County at Pandale and south to U.S. 90 (Kerans et al., 1999); and 5) Fort Towson, Choctaw County, southeast Oklahoma. B. Middle-Late Albian paleogeographic map of Comanche shelf depositional features and measured sections (orange dots) (map from Scott, 2007). Measured cores and sections: 1. Shell Chapman core, Waller County; 2. Shell Tomasek core, Bee County; 3. Pioneer Myra Kelley core, Dewitt County; 4. Pioneer Schroeder core, Bee County; 5. U.S. 90 Pecos River Bridge, Val Verde County; 6. East Iraan U.S. 190 section, Crockett County; 7. Fort Stockton, Pecos County. |
During
the middle to early late Albian the Comanche Shelf was a wide shallow carbonate
shelf on which were deposited units of the Fredericksburg Group and the
equivalent Edwards Group. During this phase the carbonate shelf prograded into
the ancestral Gulf of Mexico (Bay, 1977; Salvador,
1991) (Fig.
1.B ![]() ). This phase was followed by a widespread late Albian sea-level rise that
flooded the shallow shelf and intrashelf basins and deposited the mixed
carbonate-siliciclastic Washita Group. The later Albian was a time of carbonate
shelf retrograding and drowning. The Georgetown Formation limestone and marl
overly shallow-water facies of the Edwards Group (Rose,
1972; Kerans, 2002) and document this sea-level rise. During multiple episodes of shelf
retrogradation and progradation extensive rudist buildups developed in
depositional cycles (Scott; 1993; Kerans,
2002). The episodes are
documented in well-exposed sections along the Pecos River and in spectacular
road cuts through the steep banks of the Pecos River. Kerans
(2002)
subdivided measured sections in the Pecos River Canyon into high frequency
cycles that document multiple shoaling-deepening cycles.
). This phase was followed by a widespread late Albian sea-level rise that
flooded the shallow shelf and intrashelf basins and deposited the mixed
carbonate-siliciclastic Washita Group. The later Albian was a time of carbonate
shelf retrograding and drowning. The Georgetown Formation limestone and marl
overly shallow-water facies of the Edwards Group (Rose,
1972; Kerans, 2002) and document this sea-level rise. During multiple episodes of shelf
retrogradation and progradation extensive rudist buildups developed in
depositional cycles (Scott; 1993; Kerans,
2002). The episodes are
documented in well-exposed sections along the Pecos River and in spectacular
road cuts through the steep banks of the Pecos River. Kerans
(2002)
subdivided measured sections in the Pecos River Canyon into high frequency
cycles that document multiple shoaling-deepening cycles.
Barremian
to Lower Cenomanian strata on the Comanchean shelf are grouped as the Comanchean
Series, which is defined by unconformity-bounding surfaces
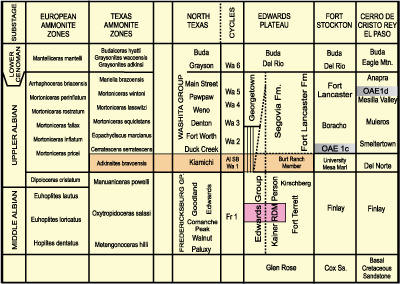
(Fig. 2 ![]() )
(Hill, 1901; reviewed by Scott et al.,
2003). The Comanchean Series is comprised of three
disconformity-bounded groups in north central Texas: Trinity, Fredericksburg and
Washita. The Trinity Group comprises the Hensel and Glen Rose formations; the
Fredericksburg Group comprises the Walnut and Edwards/Comanche Peak and Goodland
formations; and the Washita Group includes seven shale and limestone formations
in the East Texas Basin (Scott et al., 2003).
)
(Hill, 1901; reviewed by Scott et al.,
2003). The Comanchean Series is comprised of three
disconformity-bounded groups in north central Texas: Trinity, Fredericksburg and
Washita. The Trinity Group comprises the Hensel and Glen Rose formations; the
Fredericksburg Group comprises the Walnut and Edwards/Comanche Peak and Goodland
formations; and the Washita Group includes seven shale and limestone formations
in the East Texas Basin (Scott et al., 2003).
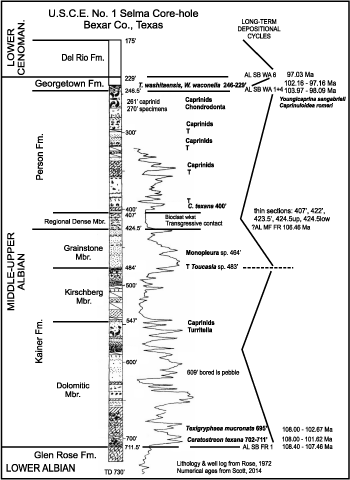
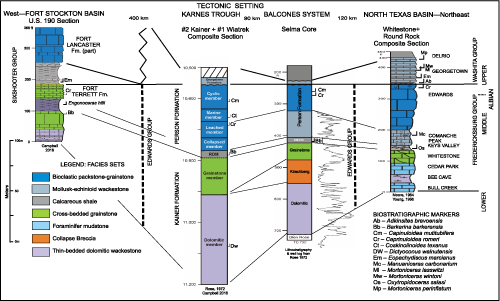
Southward in central Texas on the San Marcos Arch the formations become dominantly carbonate and comprise a different lithostratigraphy. The Kainer and Person formations make up the Edwards Group (Rose, 1972), which Rose correlated with the Fredericksburg Group and the lower part of the Washita Group. Newly discovered caprinid rudists in the Person challenge this correlation. The overlying Georgetown, Del Rio and Buda formations are part of the Washita Group, which is bounded by disconformities (Rose, 1972).
Westward on the Edwards Plateau the Fort Terrett and Segovia formations are mapped as the Edwards Group (Rose, 1972). The Fort Terrett Formation correlates by mapping and biostratigraphy with the Fredericksburg Group. The Segovia correlates with the Washita Group. Westward the Segovia grades into the carbonate shelf Fort Lancaster Formation (Smith et al., 2000). Each of these units is bounded by disconformities and each hosts biostratigraphically significant caprinids.
|
Figure 2: Stratigraphic chart correlating European ammonite zones (Reboulet et al., 2014) with U.S. Gulf Coast zones (Young, 1986) and Comanchean stratigraphy of North Texas (Scott et al., 2003), of the Edwards Plateau (modified from Rose, 1972), and of the Fort Stockton and El Paso sections (Scott and Kidson, 1977; Lucas et al., 2010). |
Edwards Formation/Group. The Edwards Formation is
mapped north of the Central Texas Platform (Fig. 1.B ![]() )
as the uppermost unit of
the Fredericksburg Group. This medium to thick bedded limestone underlies the
Washita Group
(Fig. 2
)
as the uppermost unit of
the Fredericksburg Group. This medium to thick bedded limestone underlies the
Washita Group
(Fig. 2 ![]() )
(Moore, 1964; Amsbury,
2003). Here the
Edwards overlies and grades down into the Comanche Peak and Walnut formations (Fig. 3
)
(Moore, 1964; Amsbury,
2003). Here the
Edwards overlies and grades down into the Comanche Peak and Walnut formations (Fig. 3 ![]() ). Middle and basal Upper Albian ammonites in these units define widely
correlated zones (Young, 1957,
1966; Kennedy et al., 1998). Caprinids and other rudists are common in the Edwards
Formation in this area and are identified as Caprinuloidea romeri (formerly C.
perfecta Palmer in Scott, 2002, and revised by Mitchell,
2013a) and Youngicaprina sangabrieli Mitchell,
2013a (Fig. 4
). Middle and basal Upper Albian ammonites in these units define widely
correlated zones (Young, 1957,
1966; Kennedy et al., 1998). Caprinids and other rudists are common in the Edwards
Formation in this area and are identified as Caprinuloidea romeri (formerly C.
perfecta Palmer in Scott, 2002, and revised by Mitchell,
2013a) and Youngicaprina sangabrieli Mitchell,
2013a (Fig. 4 ![]() ). Eoradiolites davidsoni
Hill is the common radiolitid.
). Eoradiolites davidsoni
Hill is the common radiolitid.
In central Texas the Edwards Group is comprised of two limestone/marl formations, the Kainer and Person formations. These units are separated by a thin marl-limestone unit, the "Regional Dense member" (RDM) (Rose, 1972, p. 25). The Kainer and Person are mapped in outcrops between Austin and San Antonio and down dip in the subsurface towards the Comanche Shelf margin Stuart City trend. Westward across the Edwards Plateau the Fort Terrett and Segovia formations compose the Edwards Group. Farther west the Segovia grades into the Fort Lancaster Formation, which disconformably overlies the Fort Terrett. Southward at the northern margin of the Maverick Basin and the Devils River trend the Edwards Group grades into the Devils River Formation (Rose, 1972; Smith et al., 2000).
|
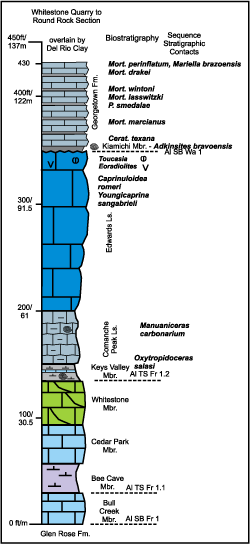
Figure 3: Composited section of middle-upper Albian strata in northeastern Travis and Williamson counties north of the San Marcos Arch. Lithostratigraphy from Moore (1964), Feray in Hazzard (1949), Wilbert (1967), and in Scott and Weaver (2010, Fig. 2). Ammonite biostratigraphy from Young (1957) and Kennedy et al. (1998). Rudist biostratigraphy from Mitchell (2013a). Sequence stratigraphic notation from Scott et al. (2003). |
|
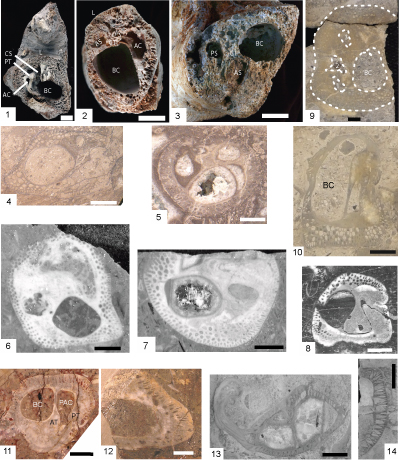
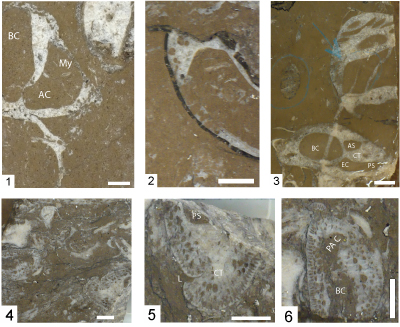
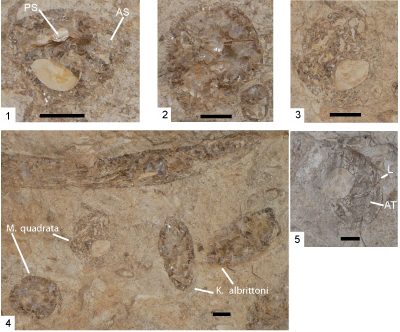
Figure 4: Caprinid specimens from Edwards Formation and Mural Formation outcrops and Stuart City Formation cores. Scale bar is 1 cm. 1-3. Youngicaprina sangabrieli Mitchell; Edwards Formation, Travis County, Texas; silicified specimens. 1, LV NPL 2381; 2, RV UT 11276; 3) RV NPL 15739; Scale bar 1 cm. Accessory cavity (AC); anterior socket (AS); anterior tooth (AT); body cavity (BC); central tooth (CT); central socket (CS); ligament (L); posterior tooth (PT); posterior socket (PS). Photos from Scott and Weaver (2010, Fig. 3). 4-5. Caprinuloidea romeri Mitchell; Stuart City Formation, Pioneer Myra Kelley Gas Unit 3-4, Sawfish Field, Dewitt County, Texas; 4, RV view into valve at 14,789.5 ft and 5, RV view into valve at 14,863 ft. 6-8. Texicaprina kugleri Bouwman; Stuart City Formation, Pioneer No. 1 Schroeder, Pawnee Field, Bee County, Texas; 6, view into RV at 14,063 ft; 7, view into RV at 14,243; and 8, articulated specimen at 14,296 ft. 9-10. Youngicaprina sangabrieli Mitchell; Stuart City Formation, Pioneer Burns Gas Unit No. 1-2, Moray Field, Dewitt County, Texas; 9, view into RV at 14,812.5 ft, BC = body cavity, 1 is anterior myophore, 2 is anterior socket, 3 is posterior socket; 10, view out of LV at 14,827 ft. 11- 12. Caprinuloidea perfecta Palmer. Lower Albian Mural Formation, Los Coyotes Member, Santa Ana section, Sonora, Mexico (Gonz�les-Le�n et al., 2008, Fig. 4G-H); 11, LV view into valve (op. cit., Fig. 4H); 12, fragment of RV (op. cit., Fig. 4G). 13- 14. Coalcomana ramosa (Boehm), Stuart City Formation, Pioneer No. 1 Schroeder, Pawnee Field, Bee County, Texas; 13, RV view into valve, 14,735.6 ft; 14, ventral margin of valve fragment, 14,620.2 ft. |
Kainer Formation. The Kainer Formation is the lower
unit of the Edwards Group on the San Marcos Arch at the southeast nose of the
Central Texas Platform and in up-dip outcrops
(Figs. 2 ![]() & 5
& 5 ![]() )
(Rose, 1972,
p. 18). In its type well, Shell No. 2 Kainer, the Kainer is 122.6 m thick and in
other wells it ranges up to 137 m. The Kainer is comprised of an informal lower
dolomite interval and an upper grainstone interval. The Kainer overlies the
lower Albian dolomitic Glen Rose Formation with a sharp contact and is overlain
by a sharp, locally iron-stained contact with the basal Regional Dense member
(RDM) of the Person Formation
(Fig. 5
)
(Rose, 1972,
p. 18). In its type well, Shell No. 2 Kainer, the Kainer is 122.6 m thick and in
other wells it ranges up to 137 m. The Kainer is comprised of an informal lower
dolomite interval and an upper grainstone interval. The Kainer overlies the
lower Albian dolomitic Glen Rose Formation with a sharp contact and is overlain
by a sharp, locally iron-stained contact with the basal Regional Dense member
(RDM) of the Person Formation
(Fig. 5 ![]() )
(Rose, 1972; Campbell,
2016). The Kainer yields diverse shallow-water fossils: miliolids, Dictyoconus
walnutensis (Carsey), Ceratostreon
texana (Roemer), texigryphaeid oysters, toucasiids, indeterminate
caprinids, and gastropods. Because the Kainer is at the base of the Edwards
Group (Rose, 1972), it correlates with the Walnut Formation in nearby
updip outcrops, which are composed of similar lithologies (Moore,
1964).
)
(Rose, 1972; Campbell,
2016). The Kainer yields diverse shallow-water fossils: miliolids, Dictyoconus
walnutensis (Carsey), Ceratostreon
texana (Roemer), texigryphaeid oysters, toucasiids, indeterminate
caprinids, and gastropods. Because the Kainer is at the base of the Edwards
Group (Rose, 1972), it correlates with the Walnut Formation in nearby
updip outcrops, which are composed of similar lithologies (Moore,
1964).
Person Formation. The Person Formation is the upper
unit of the Edwards Group in Central Texas wells and outcrops
(Figs. 2 ![]() & 5
& 5 ![]() ) (Rose,
1972, p. 19). In its type section, Standard of Texas No. 1 Wiatrek core, the
Person is 69.2 m thick and in other wells it ranges up to 95 m thick. The Person
is comprised of the basal RDM, which is overlain by a collapsed member, a
leached member, a marine member, and a cyclic member. The Person is overlain
disconformably by the Georgetown Formation; locally a soil is preserved at the
contact and in places borings penetrate into the uppermost bed up to 20 cm; some
borings are filled with pelagic mudstone (Campbell, 2016).
) (Rose,
1972, p. 19). In its type section, Standard of Texas No. 1 Wiatrek core, the
Person is 69.2 m thick and in other wells it ranges up to 95 m thick. The Person
is comprised of the basal RDM, which is overlain by a collapsed member, a
leached member, a marine member, and a cyclic member. The Person is overlain
disconformably by the Georgetown Formation; locally a soil is preserved at the
contact and in places borings penetrate into the uppermost bed up to 20 cm; some
borings are filled with pelagic mudstone (Campbell, 2016).
Common
rudists in parts of the Person Formation are Caprinuloidea romeri, Neokimbleia
planata (Conrad), Toucasia
texana (Roemer), and Sellaea
sp. (Figs.
6 ![]() - 7
- 7 ![]() ). The benthic foraminifers are Barkerina barkerensis Frizzell and Schwartz, Buccicrenata
subgoodlandensis (Vanderpool), Coskinolinoides
texanus Keijzer, Cuneolina parva Henson,
Dictyoconus walnutensis, and
Pseudonummoloculina heimi (Bonet).
). The benthic foraminifers are Barkerina barkerensis Frizzell and Schwartz, Buccicrenata
subgoodlandensis (Vanderpool), Coskinolinoides
texanus Keijzer, Cuneolina parva Henson,
Dictyoconus walnutensis, and
Pseudonummoloculina heimi (Bonet).
The RDM is a bioclastic, peloidal, bioturbated wackestone to lime mudstone. In places miliolids are common together with the larger benthic foraminifers Barkerina and Cuneolina, texigryphaeid oysters, echinoderms, and ostracodes. Up dip in outcrop the RDM is a moderate yellow gray, argillic lime mudstone with small Planolites burrows and the rare infaunal bivalve, Pleuromya knowltoni (Hill), which is preserved in life position. In the uppermost Person cyclic interval caprinids are moderately preserved and identifiable as members of the Caprinuloidea Zone (Scott and Filkorn, 2007).
|
Figure 5: Lithostratigraphy of the Edwards Group in the U.S.C.E. Selma No. 1 (Rose, 1972, Pl. 4, cross section Z-Z'). The contact between the Georgetown and Del Rio formations is a widespread regional disconformity Al SB Wa 6 (Scott et al., 2003). The Washita Group is comprised of the Georgetown, Del Rio and Buda formations. |
|
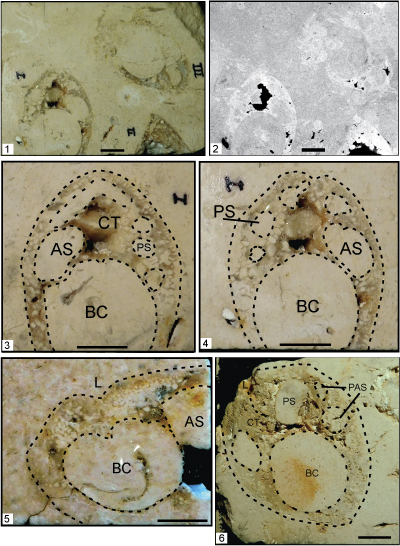
Figure 6: Rudists from interior shelf cores, Karnes County, Texas (Campbell, 2016). Scale bar is 1 cm. 1. Sellaea sp., LV with body cavity (BC), accessory cavity (AC) and myophore (MY); Standard of Texas No. 1 Wiatrek, 10,870 ft, Person Formation, marine member. 2. Toucasia texana (Roemer) whorl cross section showing two-layer wall of outer dark brown fibrous calcite and inner sparry calcite; Standard of Texas No. 1 Wiatrek, 10,865 ft, Person Formation, marine member. 3. Neokimbleia planata (Conrad), RV with body cavity (BC), anterior tooth socket (AS), central tooth (CT), posterior socket (PS), and endomyophoral cavity (EC); Shell No. 2 L. Urbanczyk, 10,738 ft, Person Formation, cyclic member. Note solitary coral in left margin. 4-6. Caprinuloidea romeri Mitchell; Shell No. 2 L. Urbanczyk, 10,818.5 ft, Person Formation, marine member; 5. Apertural view into RV showing ligament groove (L), central tooth (CT) and posterior socket (PS). |
|
Figure
7: Edwards Group rudists, Person Formation in
U.S.C.E. Selma No. 1 core, Bexar County, Texas
(Fig. 5 1-4, Neokimbleia acutus Mitchell, 270.5 ft; 1, Core slab photograph view into RV; 2, CT-x-ray scan; 3- 4, RV, umbonal and abumbonal views, obverse sides of same core slab illustrates ligament groove; posterior accessory cavity (PAC), posterior tooth socket (PS), ligament cavity (L), central tooth dissolved (CT), anterior tooth socket (AS), anterior myophore (AM); specimen II is the acute ventral margin; specimen III is partly etched but shows main interior structures; arrow indicates stratigraphic up. 5-6, Caprinuloidea romeri Mitchell; 5, RV abumbonal view view, depth 261.5 ft. 6, RV abumbonal view at depth of 263.5 ft; anterior socket (AS); body cavity (BC); central tooth (CT); ligament groove (L); posterior accessory cavity (PAS); posterior socket (PS). |
Fort Terrett Formation. The Fort Terrett Formation (Fig. 8 ![]() )
is the lower unit of the Edwards Group exposed on the Edwards Plateau
(Fig. 2
)
is the lower unit of the Edwards Group exposed on the Edwards Plateau
(Fig. 2 ![]() )
(Rose, 1972). The Fort Terrett overlies the Glen Rose Formation
or the Hensel Formation, and it disconformably underlies the Segovia Formation
and its basal Burt Ranch Member (Rose, 1972). The Fort Terrett thickens
southward towards the Maverick Basin from about 49 m to over 90 m. The formation
is comprised of four informal lithological units: a basal nodular limestone and
marl, the burrowed resistant limestone unit, an upper dolomitic member, and the
uppermost Kirschberg evaporate and breccia member (Rose,
1972). Fossil
abundances vary within the Fort Terrett. Common fossils are echinoid clasts,
texigryphaeids, other bivalves, gastropods, miliolids, Cuneolina,
Barkerina barkerensis, Pseudonummoloculina
heimi, and ostracodes (Gargili, 2013). A road cut section of the Fort
Terrett at the western margin of the Edwards Plateau exposes marl with
the ammonite, Engonoceras hilli B�hm, which is overlain by limestone with
caprinids (Fig. 9.3-4, 9.7-8
)
(Rose, 1972). The Fort Terrett overlies the Glen Rose Formation
or the Hensel Formation, and it disconformably underlies the Segovia Formation
and its basal Burt Ranch Member (Rose, 1972). The Fort Terrett thickens
southward towards the Maverick Basin from about 49 m to over 90 m. The formation
is comprised of four informal lithological units: a basal nodular limestone and
marl, the burrowed resistant limestone unit, an upper dolomitic member, and the
uppermost Kirschberg evaporate and breccia member (Rose,
1972). Fossil
abundances vary within the Fort Terrett. Common fossils are echinoid clasts,
texigryphaeids, other bivalves, gastropods, miliolids, Cuneolina,
Barkerina barkerensis, Pseudonummoloculina
heimi, and ostracodes (Gargili, 2013). A road cut section of the Fort
Terrett at the western margin of the Edwards Plateau exposes marl with
the ammonite, Engonoceras hilli B�hm, which is overlain by limestone with
caprinids (Fig. 9.3-4, 9.7-8 ![]() )
and colonial corals (Campbell, 2016). At
multiple localities the carbon isotope profiles at the iron-stained top of the
Fort Terrett indicate subaerial exposure (Gargili, 2013; Wang,
2014; Lai, 2014; Campbell, 2016).
)
and colonial corals (Campbell, 2016). At
multiple localities the carbon isotope profiles at the iron-stained top of the
Fort Terrett indicate subaerial exposure (Gargili, 2013; Wang,
2014; Lai, 2014; Campbell, 2016).
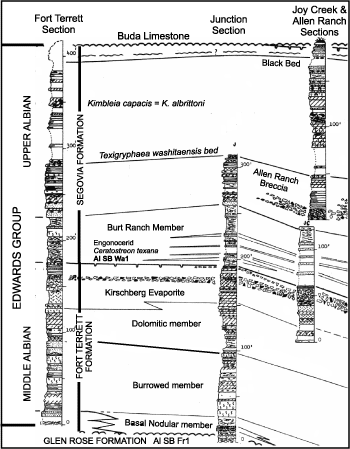
Segovia Formation. The Segovia Formation
disconformably overlies the Fort Terrett Formation on the Edwards Plateau and is
overlain disconformably by the lower Cenomanian Del Rio or Buda formations (Fig. 8 ![]() )
(Rose, 1972; Young,
1979; Smith et
al., 2000). Lithologically the Segovia is a heterogenous unit of limestone,
marl, dolomite, and collapse breccia. In the upper part of the Segovia oyster
beds of Texigryphaea washitaensis (Hill) and two lithologically
distinct beds are widely mapped (Smith et
al., 2000). The Kimbleia albrittoni (Perkins) caprinid zone
characterizes the upper Segovia, which is the type horizon of Kimbleia
capacis Coogan, 1973, the junior synonym of K.
albrittoni (Perkins, 1961; Mitchell, 2013a)
(Fig. 10
)
(Rose, 1972; Young,
1979; Smith et
al., 2000). Lithologically the Segovia is a heterogenous unit of limestone,
marl, dolomite, and collapse breccia. In the upper part of the Segovia oyster
beds of Texigryphaea washitaensis (Hill) and two lithologically
distinct beds are widely mapped (Smith et
al., 2000). The Kimbleia albrittoni (Perkins) caprinid zone
characterizes the upper Segovia, which is the type horizon of Kimbleia
capacis Coogan, 1973, the junior synonym of K.
albrittoni (Perkins, 1961; Mitchell, 2013a)
(Fig. 10 ![]() ). The
Segovia thickens southward from 70 m to 110 m where it grades into the Devils
River Formation at the shelf margin of the Maverick Basin (Rose,
1972).
). The
Segovia thickens southward from 70 m to 110 m where it grades into the Devils
River Formation at the shelf margin of the Maverick Basin (Rose,
1972).
The basal interval of the Segovia Formation consists of interbedded marl and thin bedded limestone mapped as the Burt Ranch Member (Rose, 1972). Rose designated the base at the contact between the lowest marl overlying "porcellaneous micrite breccia" and the top at the base of a miliolid bed overlain by dolomite (Rose, 1972, p. 35). In many sections the basal contact is a bored, iron-stained surface, and in other sections the top surfaces of several limestone beds are bored, iron-stained and identification of sequence boundary Al SB Washita 1 is equivocal. Carbon isotope analyses of the basal contact suggest that at some sections it has been subaerially exposed whereas others show no isotopic signal of subaerial exposure (Gargili, 2013; Lai, 2014; Wang, 2014). Ammonites from the Burt Ranch interval represent the Adkinsites bravoensis (B�se) ammonite Zone (Young, 1966), which indicates that this unit correlates with the Kiamichi Formation in North Texas and represents flooding of the Comanche shelf (Rose, 1972). Therefore Kimbleia albrittoni in overlying beds correlates with some part of the upper Washita Group and is in the upper Albian Stage.
|
Figure 8: Lithostratigraphy of the Fort Terrett and Segovia formations in Kimble County, Texas (Rose, 1972, Pl. 4). The outcrop section at the abandon site of Fort Terrett is the type section of the unit. The Joy Creek composited section is the type section of the Segovia Formation (Rose, 1972) and is composited with the Allen Ranch section of Rose (1972). Biostratigraphic data from Perkins (1961), Coogan (1977) and Gargili (2013). |
|
Figure 9: Rudist specimens from bed FT16 in the upper part of the Fort Terrett Formation exposed in the north roadcut, U.S. 190, east of Iraan, Crockett County, Texas north roadcut beneath picnic area (Campbell, 2016). Scale bar is 1 cm. 1, Caprinuloidea romeri Mitchell; Specimen 2B, view into RV; body cavity (BC), ligament groove (L), anterior tooth socket (AS), central tooth (CT), posterior tooth socket (PS), ectomyophoral cavity (EC). 2, Youngicaprina sangabrieli Mitchell; Specimen 4A, view into RV highly recrystallized cardinal platform and abraded margins; anterior myophore (AM), anterior socket (AS), posterior socket (PS), posterior myophore (PM). 3, Indeterminate Caprinid; Specimen 5A, LV with abraded ventral and dorsal margins. 4, Indeterminate Caprinid; Specimen 4B, RV highly abraded dorsal margin. 5, Caprinuloidea romeri Mitchell?; Sample 2A, upper specimen cut through coiled LV, associated with fragment of solitary coral and chondrodontid bivalve shells. 6, Eoradiolites davidsoni (Hill, 1893); Specimen 1B, oblique section through. 7, Sample A with broken shells of Caprinuloidea, Eoradiolites and Chondrodonta. 8, Sample 3 with broken subparallel rudists and other bioclasts. 9-10, Caprinuloidea romeri Mitchell; two cross sections of same specimen in upper part, Fort Terrett Formation, U.S. 67/375, 5.8 mi northeast of I-10, 15 mi east of Fort Stockton, Pecos County, Texas (Hojnacki, 2016). 11-12, Caprinuloidea romeri Mitchell, LV body cavity (BC) and posterior accessory cavity (PAC). |
Fort Lancaster Formation. The Fort Lancaster
Formation is mapped in the western part of the Edwards Plateau and grades east
into the Segovia Formation, south into the Devils River Formation and west into
the Boracho Formation
(Fig. 2 ![]() )
(Smith et al., 2000). It is comprised mainly of resistant bedded carbonates
and at its base is the Burt Ranch Member comprised of interbedded marl and
limestone (Rose, 1972). The Fort Lancaster disconformably overlies the
Fort Terrett Formation and underlies the West Prong, Del Rio or Buda formations.
)
(Smith et al., 2000). It is comprised mainly of resistant bedded carbonates
and at its base is the Burt Ranch Member comprised of interbedded marl and
limestone (Rose, 1972). The Fort Lancaster disconformably overlies the
Fort Terrett Formation and underlies the West Prong, Del Rio or Buda formations.
Here
the Burt Ranch Member yields the ammonites, Adkinsites
and Metengonoceras, and the oysters Texigryphaea
navia (Hall) and Ceratostreon
texana, and the bivalve Scabrotrigonia
emoryi (Conrad). These taxa correlate the Burt Ranch with the
Kiamichi Formation in North Texas, with the University Mesa Marl and with the
Del Norte Formation at Cerro de Cristo Rey near El Paso. The upper seven meters
of the Fort Lancaster are thin to medium bedded bioclastic limestone with the
rudists Kimbleia albrittoni (Perkins) and Mexicaprina quadrata Alenc�ster & Oviedo-Garc�a
(Fig. 10 ![]() ). These rudists are characteristic of the upper Albian Mexicaprina Zone
(Scott and Filkorn, 2007). The Del Rio and Buda formations
disconformably overlie the Fort Lancaster here.
). These rudists are characteristic of the upper Albian Mexicaprina Zone
(Scott and Filkorn, 2007). The Del Rio and Buda formations
disconformably overlie the Fort Lancaster here.
|
Figure 10: Rudist specimens from upper part of Fort Lancaster Formation exposed in the north roadcut, U.S. 190, east of Iraan, Crockett County, Texas (Lai, 2014). Scale bar is 1 cm. 1, 2, 3, Mexicaprina quadrata Alenc�ster & Oviedo-Garc�a, 1, view out of RV, anterior socket (AS), posterior socket (PS); 2, 3, recrystallized RVs. 4, Subhorizontal specimens of Mexicaprina quadrata and Kimbleia albrittoni; 5, Mexicaprina quadrata, view into LV, anterior tooth (AT), ligament (L). |
Devils River Formation. South of the Edwards Plateau and the Fort Stockton Basin the Devils River Formation encircles the northern margin of the Maverick Basin (Lozo and Smith, 1964; Rose, 1972; Miller, 1984). It is an unconformity-bounded unit overlying the Glen Rose Formation and underlying the West Prong, Del Rio or Buda formations (Smith et al., 2000). At the Pecos River bridge on U.S. 90 the Devils River is about 36 m thick and grades down into the Salmon Peak Formation, a peloid-planktic foraminifer wackestone and fine grained bioclastic wackestone (Smith and Brown, 1983; Scott, 1990, Fig. 50). In the upper 5 meters multiple caprinid biostromes are separated by hardground contacts and host Kimbleia albrittoni and Mexicaprina minuta (Scott, 1990; Wang, 2014).
Stuart City Formation. The Stuart City Formation
forms the narrow Comanche Shelf margin trend in south Texas subsurface (Winter,
1961; Cook, 1979; Wilson,
1986). In its type area of LaSalle
County, Texas, the Stuart City overlies the Aptian-Albian Tamaulipas Formation
and underlies the Georgetown Formation or Upper Cretaceous units
(Fig. 2 ![]() ). The
carbonate shelf sections cored in the Shell No. 1 Chapman, Waller County, and
the Shell No. 1 Tomasek, Bee County, serve as lectostratotypes (Figs.
1.A-B
). The
carbonate shelf sections cored in the Shell No. 1 Chapman, Waller County, and
the Shell No. 1 Tomasek, Bee County, serve as lectostratotypes (Figs.
1.A-B ![]() - 2
- 2 ![]() ) (Cook,
1979, Fig. 18; Scott, 1990). The Stuart City rudist-coral facies grades
down into pelagic lime mudstone/wackestone of the Tamaulipas Formation (Scott,
1990, Fig. 25) and is unconformably overlain by upper Albian Georgetown, or by
Cenomanian shale of the Eagle Ford Formation, or even the Coniacian Austin Chalk
(Cook, 1979; Scott,
1990; Waite et
al., 2007). The Stuart City Formation in the Chapman and Tomasek cores
correlates with the Edwards Formation of the Fredericksburg Group in North
Texas. The benthic foraminifers, Dictyoconus
walnutensis and Coskinolinoides
texanus, and caprinids range to within about 10 meters of the top
unconformity (Scott, 1990). The caprinids, Caprinuloidea
romeri Mitchell and Texicaprina
vivari (Palmer), now T. kugleri
(Bouwman) according to Mitchell (2013a), and Youngicaprina sangabrieli Mitchell are the common diagnostic species.
) (Cook,
1979, Fig. 18; Scott, 1990). The Stuart City rudist-coral facies grades
down into pelagic lime mudstone/wackestone of the Tamaulipas Formation (Scott,
1990, Fig. 25) and is unconformably overlain by upper Albian Georgetown, or by
Cenomanian shale of the Eagle Ford Formation, or even the Coniacian Austin Chalk
(Cook, 1979; Scott,
1990; Waite et
al., 2007). The Stuart City Formation in the Chapman and Tomasek cores
correlates with the Edwards Formation of the Fredericksburg Group in North
Texas. The benthic foraminifers, Dictyoconus
walnutensis and Coskinolinoides
texanus, and caprinids range to within about 10 meters of the top
unconformity (Scott, 1990). The caprinids, Caprinuloidea
romeri Mitchell and Texicaprina
vivari (Palmer), now T. kugleri
(Bouwman) according to Mitchell (2013a), and Youngicaprina sangabrieli Mitchell are the common diagnostic species.
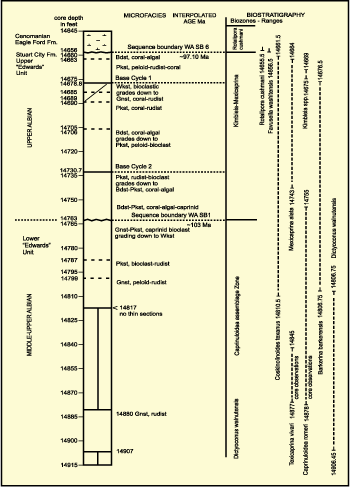
The
Stuart City Formation cored in the Pioneer Myra Kelley Gas Unit 3-4, Sawfish
Field, DeWitt County, Texas (Figs. 1.A ![]() & 3
& 3 ![]() ), is 79 m thick and is divided into
lower and upper parts by an intraformational unconformity (Fig. 11
), is 79 m thick and is divided into
lower and upper parts by an intraformational unconformity (Fig. 11 ![]() )
(Waite
et al., 2007; Phelps et
al., 2014). The lower part of the Stuart City below 4500 m (14,763 ft) is
mainly comprised of peloid-rudist grainstone that grades up into rudist
bioclastic packstone and wackestone. Caprinuloidea
romeri and Texicaprina kugleri
characterize this interval. Dictyoconus
walnutensis correlates the lower part of the Stuart City with the
Fredericksburg Group (Coogan, 1977).
)
(Waite
et al., 2007; Phelps et
al., 2014). The lower part of the Stuart City below 4500 m (14,763 ft) is
mainly comprised of peloid-rudist grainstone that grades up into rudist
bioclastic packstone and wackestone. Caprinuloidea
romeri and Texicaprina kugleri
characterize this interval. Dictyoconus
walnutensis correlates the lower part of the Stuart City with the
Fredericksburg Group (Coogan, 1977).
The
upper part of the Stuart City interval is comprised of deeper-water
peloid-bioclast packstone that grades up into coral-algal boundstone and
overlies the shallower rudist-dominated assemblage, which indicates a
transgressive depositional cycle. The upper interval is unconformably overlain
by the Cenomanian-Turonian Eagle Ford Formation. Upper Albian rudist species Ichthyosarcolites [Mexicaprina] alatus (Aguilar et
al., 2008; Mitchell, 2013b) and Kimbleia
albrittoni (Fig. 12 ![]() )
range to the top of this upper unit. Two agglutinate
benthic foraminifers in this interval are Coskinolinoides
texanus and Barkerina barkerensis,
both of which range from the middle Albian Fredericksburg Group into the upper
Albian Washita Group (Scott, 2014).
)
range to the top of this upper unit. Two agglutinate
benthic foraminifers in this interval are Coskinolinoides
texanus and Barkerina barkerensis,
both of which range from the middle Albian Fredericksburg Group into the upper
Albian Washita Group (Scott, 2014).
|
Figure 11: Carbonate microfacies and biostratigraphic log of Pioneer Natural Resources Myra Sue Kelley Gas Unit 3-4, DeWitt County, Texas. Abbreviations: grainstone (Gnst), packstone (Pkst), boundstone (Bdst), wackestone (Wkst). |
|
1-5, Rudists in the Stuart City Formation in DeWitt County, Texas. 1-3 Pioneer Natural Resources Myra Sue Kelly Gas Unit No. 3-4. 1, Ichthyosarcolites [Mexicaprina] alatus at 14,743ft. 2, 3, Kimbleia albrittoni at 14,789ft and 14,845.5ft. 4, 5, Pioneer Natural Resources Burns 1-2 at 14,812.5ft, 14,827ft. 6-11, Comanche rudists in Chandeleur Sound offshore Louisiana Chevron 4898 #2; scale bar = 1cm. 6, 10, Mexicaprina quadrata at 12.810ft and 12,831ft. 7, Ichthyosarcolites alatus and Kimbleia albrittoni at 12,815.5ft. 8, 9, K. albrittoni at 12,820ft and 12,829ft. 11, Caprinuloidea romeri at 13,644ft. |
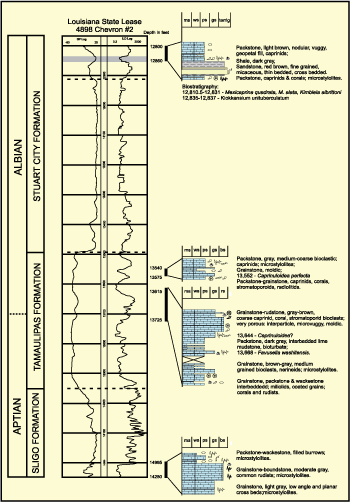
Offshore Louisiana Lower Cretaceous Carbonates. The
Albian section is drilled in numerous wells and cored in a few wells onshore and
offshore Louisiana (Tyrrell and Scott,
1989; Yurewitz et
al., 1993; Scott, 1993; Mancini et
al., 2005). The Chevron State Lease 4894 #2 well was cored in several
intervals and rudists were found in two intervals (Fig. 13 ![]() ). The caprinids
correlate this interval with the upper Albian Mexicaprina Zone and the upper
part of the Washita Group.
). The caprinids
correlate this interval with the upper Albian Mexicaprina Zone and the upper
part of the Washita Group.
|
Figure 13: Chevron State Lease 4898 No. 2 Chandeleur Sound Upper Albian; facies (E.A. Mancini, personal communication), caprinids (R.W. Scott, personal communication) and palynomorphs (D.G. Benson, personal communication). |
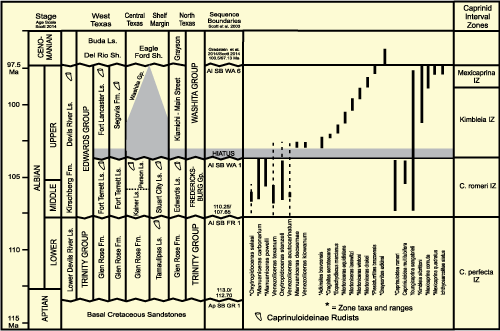
The
Fredericksburg and Washita groups host a diverse assemblage of Brancoceratid
ammonites that serve to define precise zones (Fig. 14 ![]() )
(Young, 1966, 1986; Kennedy et al.,
1998, 1999; Scott et al.,
2003).
Numerous middle to lower-upper Albian species of the Subfamily Mojsisovicziinae Hyatt,
1903, characterize the Fredericksburg: Venezoliceras Spath,
1925, Oxytropidoceras Stieler, 1920, and Manuaniceras Spath,
1925. These genera cross the lower-upper
Albian sequence boundary Washita 1 (Al SB Wa1) into the basal Washita Group
having survived the sea-level flooding of North America. One genus of this group
is new, Adkinsites Spath, 1931,
and defines the lowermost Washita zone in the Kiamichi Formation and equivalent
units in the Western Interior seaway (Young, 1966,
1986; Scott,
1970). The younger stratigraphic units of the Washita Group are
characterized by ammonites of the Subfamily Mortoniceratinae H. Douvill�
(Kennedy et al., 1998,
1999): Mortoniceras
Meek, 1876, and its several subgenera. Seven upper Albian mortonicerid
zones overlie the Adkinsites bravoensis Range Zone in the Kiamichi Formation (Fig. 14
)
(Young, 1966, 1986; Kennedy et al.,
1998, 1999; Scott et al.,
2003).
Numerous middle to lower-upper Albian species of the Subfamily Mojsisovicziinae Hyatt,
1903, characterize the Fredericksburg: Venezoliceras Spath,
1925, Oxytropidoceras Stieler, 1920, and Manuaniceras Spath,
1925. These genera cross the lower-upper
Albian sequence boundary Washita 1 (Al SB Wa1) into the basal Washita Group
having survived the sea-level flooding of North America. One genus of this group
is new, Adkinsites Spath, 1931,
and defines the lowermost Washita zone in the Kiamichi Formation and equivalent
units in the Western Interior seaway (Young, 1966,
1986; Scott,
1970). The younger stratigraphic units of the Washita Group are
characterized by ammonites of the Subfamily Mortoniceratinae H. Douvill�
(Kennedy et al., 1998,
1999): Mortoniceras
Meek, 1876, and its several subgenera. Seven upper Albian mortonicerid
zones overlie the Adkinsites bravoensis Range Zone in the Kiamichi Formation (Fig. 14 ![]() ) (Young,
1959,
1986). Co-occurring European species correlate
the middle-upper Albian boundary in the upper part of the Fredericksburg Group (Kennedy
et al., 1998, 1999; Scott et
al., 2003).
) (Young,
1959,
1986). Co-occurring European species correlate
the middle-upper Albian boundary in the upper part of the Fredericksburg Group (Kennedy
et al., 1998, 1999; Scott et
al., 2003).
Rudist
bivalves have undergone multiple Early Cretaceous episodes of extinction and the
group rapidly diversified throughout the Early Cretaceous making them an
important tool for biostratigraphic correlation (Scott and Filkorn,
2007; Skelton, 2013; Steuber et
al., 2016). Although rudists do not occur in the same beds as ammonites,
they are in facies superposed with ammonites. The lower parts of the
Fredericksburg Group, the Walnut Formation yields numerous middle Albian
ammonite species (Fig. 15 ![]() )
(Young, 1966). The upper part of the Edwards
Formation spans the middle-upper Albian zones, and hosts common caprinid rudists
among other rudist groups. In shelf carbonates of the West Texas Pecos River
region rudist genera overlie upper Albian ammonite facies (Scott and Kidson,
1977; Scott and Kerans,
2004).
)
(Young, 1966). The upper part of the Edwards
Formation spans the middle-upper Albian zones, and hosts common caprinid rudists
among other rudist groups. In shelf carbonates of the West Texas Pecos River
region rudist genera overlie upper Albian ammonite facies (Scott and Kidson,
1977; Scott and Kerans,
2004).
Five
rudist zones in the Comanchean Series are defined by the ranges of sixteen
rudist taxa (Scott and Filkorn,
2007). These zones are integrated
with Barremian to Albian ammonite zones and foraminiferal ranges in the Trinity,
Fredericksburg and Washita groups (Coogan, 1973, 1977; Scott,
1990, 2002). The Caprinuloidea Zone of
Scott and Filkorn (2007) is redefined here as the interval between
the last appearance of Coalcomana ramosa
(Fig. 4.13-14 ![]() ) and the last appearance of Caprinuloidea perfecta. Mitchell (2013a) revised this group
and restricted C. perfecta to the
Trinity Group. He redefined Fredericksburg specimens previously identified as Caprinuloidea
perfecta by Scott (2002) as
Caprinuloidea romeri. This species is diagnostic of the Fredericksburg Group
and its zone is the interval between the last appearance of C.
perfecta (Fig. 4.11-12
) and the last appearance of Caprinuloidea perfecta. Mitchell (2013a) revised this group
and restricted C. perfecta to the
Trinity Group. He redefined Fredericksburg specimens previously identified as Caprinuloidea
perfecta by Scott (2002) as
Caprinuloidea romeri. This species is diagnostic of the Fredericksburg Group
and its zone is the interval between the last appearance of C.
perfecta (Fig. 4.11-12 ![]() ) and the last appearance of C. romeri. This Caprinuloidea romeri Interval Zone spans the middle
to lower upper Albian Fredericksburg Group; this zone includes an assemblage of
other bivalves and benthic foraminifers that are diagnostic of the Texas
Fredericksburg Group. The Caprinuloidea
romeri IZ is present in the shelf facies of the Fredericksburg Group and is
overlain by the regionally correlated sequence boundary Al SB WA1 located at the
base of the overlying Washita Group (Scott and Filkorn,
2007).
) and the last appearance of C. romeri. This Caprinuloidea romeri Interval Zone spans the middle
to lower upper Albian Fredericksburg Group; this zone includes an assemblage of
other bivalves and benthic foraminifers that are diagnostic of the Texas
Fredericksburg Group. The Caprinuloidea
romeri IZ is present in the shelf facies of the Fredericksburg Group and is
overlain by the regionally correlated sequence boundary Al SB WA1 located at the
base of the overlying Washita Group (Scott and Filkorn,
2007).
In
the Pecos River Valley outcrops Caprinuloidea
romeri occurs in the Fort Terrett Formation approximately 3 meters below the
iron-stained hardground subaerial exposure surface that marks the contact at the
top of the Fredericksburg Group
(Fig. 2 ![]() ). It is overlain by the Eopachydiscus
marcianus Zone in gray shale at the base of the Fort Lancaster Formation (Campbell,
2016).
). It is overlain by the Eopachydiscus
marcianus Zone in gray shale at the base of the Fort Lancaster Formation (Campbell,
2016).
Caprinuloideid
rudists, including Caprinuloidea romeri,
are here documented from the cored type section of the Person Formation in the
upper part of the Edwards Group (Rose,
1972) (Figs.
6 ![]() - 7
- 7 ![]() ,
9
,
9 ![]() & 14
& 14 ![]() ). These
specimens enable a precise correlation of the Person Formation with the Fort
Terrett Formation cropping out on the Pecos River, and with the Fredericksburg
Group. This evidence together with foraminifer and ammonite zones (Scott
and Kerans, 2002) demonstrates that the Person Formation is coeval with
Fredericksburg units and underlies sequence boundary Al SB Wa1.
). These
specimens enable a precise correlation of the Person Formation with the Fort
Terrett Formation cropping out on the Pecos River, and with the Fredericksburg
Group. This evidence together with foraminifer and ammonite zones (Scott
and Kerans, 2002) demonstrates that the Person Formation is coeval with
Fredericksburg units and underlies sequence boundary Al SB Wa1.
The
upper Albian Kimbleia albrittoni and Ichthyosarcolites [Mexicaprina] alatus
caprinid zones are stratigraphically above the Caprinuloidea romeri Interval Zone
(IZ) (Fig. 14 ![]() )
(Scott and Filkorn, 2007). The Kimbleia IZ
correlates with three ammonite zones in the lower Washita Group of north-central
Texas: the Adkinsites bravoensis IZ, the Eopachydiscus marcianus IZ, and the
Drakeoceras wintoni IZ. On the Pecos River in West Texas the Kimbleia IZ spans
the lower part of the Upper Devils River Limestone (Scott and Filkorn,
2007). The upper Albian Mexicaprina IZ is the interval of the total range of Mexicaprina
in the uppermost part of the Upper Devils River Limestone and in the El Abra
Limestone in Mexico (Filkorn, 2002; Scott and Filkorn,
2007). This interval correlates with the uppermost Albian Plesioturrilites
brazoensis ammonite IZ in the Main Street Formation and directly underlies the
Lower Cenomanian Del Rio Shale, which contact is sequence boundary WA SB 6 in
the North Texas section (Scott et
al., 2003). In the Fort Stockton Basin these rudist zones overly the Boracho
Formation, which hosts several mortonicerid ammonite zones (Scott and Kidson,
1977).
)
(Scott and Filkorn, 2007). The Kimbleia IZ
correlates with three ammonite zones in the lower Washita Group of north-central
Texas: the Adkinsites bravoensis IZ, the Eopachydiscus marcianus IZ, and the
Drakeoceras wintoni IZ. On the Pecos River in West Texas the Kimbleia IZ spans
the lower part of the Upper Devils River Limestone (Scott and Filkorn,
2007). The upper Albian Mexicaprina IZ is the interval of the total range of Mexicaprina
in the uppermost part of the Upper Devils River Limestone and in the El Abra
Limestone in Mexico (Filkorn, 2002; Scott and Filkorn,
2007). This interval correlates with the uppermost Albian Plesioturrilites
brazoensis ammonite IZ in the Main Street Formation and directly underlies the
Lower Cenomanian Del Rio Shale, which contact is sequence boundary WA SB 6 in
the North Texas section (Scott et
al., 2003). In the Fort Stockton Basin these rudist zones overly the Boracho
Formation, which hosts several mortonicerid ammonite zones (Scott and Kidson,
1977).
|
Figure 14: Biostratigraphy of ammonites and rudists. Lithostratigraphy from Rose (1972) and Smith et al., 2000). Ammonite biostratigraphy from Young (1966, 1984) and Kennedy et al. (1998, 1999). Rudist biostratigraphy revised from Scott and Filkorn (2007). |
The revised Caprinuloidea romeri Interval Zone is
recognized in outcrops on the eastern margin of the Fort Stockton Basin and in
the type section of the Person Formation on the subsurface San Marcos Arch (Campbell,
2016; Hojnacki, 2016) (Fig. 15 ![]() ). This species is widespread in the
Edwards Formation on the western outcrop margin of the East Texas Basin (Mitchell,
2013a). The C. romeri Zone overlies the Manuaniceras carbonarium Zone in the
underlying Comanche Peak Formation (Young, 1986). The C. romeri IZ
underlies the Adkinsites bravoensis Zone in the East Texas Basin (Figs. 14
). This species is widespread in the
Edwards Formation on the western outcrop margin of the East Texas Basin (Mitchell,
2013a). The C. romeri Zone overlies the Manuaniceras carbonarium Zone in the
underlying Comanche Peak Formation (Young, 1986). The C. romeri IZ
underlies the Adkinsites bravoensis Zone in the East Texas Basin (Figs. 14 ![]() - 15
- 15 ![]() ).
Thus the Person Formation correlates both physically and biostratigraphically
with the Edwards Formation to the northeast and with the Fort Terrett Formation
westwards across the Edwards Plateau. The Regional Dense member underlies the C.
romeri IZ and is a flooding facies that correlates with the middle part of the
Walnut Formation. The lower part of the Edwards Group, which consists of the
Fort Terrett, Kainer and Person formations is physically continuous with and
biostratigraphically equivalent with the Fredericksburg Group in the East Texas
Basin. The Segovia Formation and the Burt Ranch Member at its base correlate
with the upper Albian part of the Washita Group in the East Texas Basin.
).
Thus the Person Formation correlates both physically and biostratigraphically
with the Edwards Formation to the northeast and with the Fort Terrett Formation
westwards across the Edwards Plateau. The Regional Dense member underlies the C.
romeri IZ and is a flooding facies that correlates with the middle part of the
Walnut Formation. The lower part of the Edwards Group, which consists of the
Fort Terrett, Kainer and Person formations is physically continuous with and
biostratigraphically equivalent with the Fredericksburg Group in the East Texas
Basin. The Segovia Formation and the Burt Ranch Member at its base correlate
with the upper Albian part of the Washita Group in the East Texas Basin.
|
Figure 15: Stratigraphic correlation of Middle-Upper Albian strata west to east from the Fort Stockton Basin to North Texas Basin. Rudist biostratigraphy defined herein; Ammonite biostratigraphy defined by Young (1986). Sequence stratigraphic units defined in North Texas Basin (Scott et al. 2003). |
The Albian Caprinuloidea IZ of Scott and Filkorn (2007) is revised to consist of Caprinuloidea perfecta in the lower Albian part of the Trinity Group. The Caprinuloidea romeri IZ is defined as its total range in the Fredericksburg Group (Mitchell, 2013a). The C. romeri IZ is widespread in the Fredericksburg in northern Texas and in the Fort Terrett Formation in West Texas. It is also recognized in the Person Formation in the Edwards Group in the subsurface of the San Marcos Arch. Thus the upper part of the Edwards Group correlates with the Fredericksburg Group and not with the lower part of the Washita Group as postulated for many years. The disconformity between the Person and Georgetown formations can be traced northward with Albian Sequence Boundary Washita 1 and westward with the flooding disconformity in or at the base of the Burt Ranch Member of the Segovia Formation. In many sections this contact was subaerially exposed in early late Albian Age prior to the sea-level rise that flooded North America. The upper part of the Edwards Group on the Edwards Plateau, the Segovia Formation, does correlate with the Washita Group. In some sections on the Edwards Plateau an interval of multiple iron-stained, bored hardgrounds separate the Fort Terrett from the basal Segovia (Rose, 1972). We suggest that these hardgrounds formed during the final late highstand stage and only the uppermost is the candidate Albian Sequence Boundary Washita 1.
Order Hippuritida Newell, 1965
Suborder Requieniidina Skelton, 2013
Family Requieniidae Kutassy, 1934
Subfamily Requieniinae Kutassy, 1934
Genus Toucasia Munier-Chalmas, 1873
(Fig. 6.2 ![]() )
)
Type species.- Requienia carinata Matheron, 1842
Discussion.- Two-dimensional cross sections of requieniids are common in cores and outcrops of middle and upper Albian strata on the Comanche shelf. These shells have two layered walls: a dark calcite outer layer and a light sparry calcite inner layer that replaces the original aragonitic layer. Cross sections oriented normal to the whorl spiral axis have flat and gently curved sides. Three species have been identified in outcrop by whole valves, Toucasia hancockensis Whitney, 1952, in the Glen Rose Formation, and in the Fredericksburg and Washita groups, Toucasia texana (Roemer, 1852) and Toucasia patagiata (White, 1884) (Coogan, 1977; Garc�a-Barrera, 1995; Aguilar P�rez, 2008). In Mexico T. texana ranges from the Barremian-Aptian Cupido throughout the Albian El Abra formations (Aguilar P�rez, 2008).
Suborder Radiolitidina Skelton, 2013
Superfamily Radiolitoidea Orbigny, 1847
Family Radiolitidae Orbigny, 1847
Genus Eoradiolites Douvill�, 1909
(Fig. 9.6 ![]() )
)
Discussion.- The type species of this genus, Eoradiolites davidsoni (Hill, 1893), recently has been described and illustrated (Alenc�ster and Garc�a-Barrera, 2008). This species is highly variable and most subsequent Comanchean species have been synonymized by these authors. A new species, Eoradiolites corrugatus Davis-Strickland, 1988, is somewhat larger than E. davidsoni and may be a junior synonym. In core slabs and thin sections Eoradiolites is recognized by its distinctive thick reticulated outer shell wall comprised of thin vertical and horizontal laminae. Cross sections parallel to the commissure may intersect the three prominent radial ridges separated by two wide grooves.
Occurrence.- Eoradiolites davidsoni is widespread in Texas, Louisiana, Mississippi, and Mexico in middle and upper Albian strata (Coogan, 1977). It is documented in the upper parts of the Fort Terrett and Fort Lancaster formations (Smith et al., 2000).
Family Caprinulidae Yanin, 1990
Genus Sellaea Di Stefano, 1889
(Fig. 6.1 ![]() )
)
Discussion.- Attached valve-right valve (AV-RV) is divided into three cavities by anterior and posterior myophore plates extending from a large central tooth to anterior and posterior shell walls; anterior shell wall of RV divided into multiple ovate canals by short radial plates; dorsal to central tooth an arcuate ridge connects to posterior cavity/tooth socket; deep ligament groove. Free valve-left valve (FV-LV) has two large teeth separated by a dorsal arcuate socket connected to posterior myophore pit; anterior myophore is an ovate pit ventral to anterior tooth (Cox et al., 1969, p. N787).
Occurrence.- In Texas this genus is present in outcrops of the Edwards Formation; it also occurs in Aptian to Cenomanian strata in Italy (Coogan, 1977; Davis-Strickland, 1980). In Oman this genus occurs in Albian inner platform facies (Skelton and Masse, 2000).
Superfamily Caprinoidea Orbigny, 1847
Family Caprinuloideidae Damestoy, 1971
Subfamily Caprinuloideinae Damestoy, 1971
Genus Caprinuloidea Palmer, 1928
Type species.- Caprinuloidea perfecta Palmer, 1928, in the area of Soyatlan Adentro, Jalisco, Mexico; presumably lower Albian strata (Mitchell, 2013a).
Diagnosis.- Right valve (RV) profile with flat anterior margin and convex dorsal margin; body cavity and interior canals tabulate; external ligament groove connected to an inner curved or hooked shaped ridge; ventral shell wall with two or more rows of pallial canals; in left valve (LV) flat anterior and posterior myophore plates project into RV, the anterior plate fits onto a ledge and the posterior plate fits into a cavity (Mitchell, 2013a).
Discussion.- Mitchell (2013a) includes four species under this genus: C. perfecta, C. septata Palmer, 1928, C. multitubifera Palmer, 1928, and C. romeri Mitchell, 2013a. Mitchell narrowed the concept of C. perfecta to lower Albian specimens having small pallial canals in the central tooth and in myophoral structures in some RV specimens.
Caprinuloidea perfecta Palmer, 1928
(Fig. 4.11-12 ![]() )
)
Caprinuloidea perfecta Palmer, 1928, p. 59-60, Fig. 6; Pl. VIII, fig. 8; Pl. IX, figs. 1-2
Caprinuloidea costata Palmer, 1928, p. 62-63, Pl. XI, figs. 2-5; Scott and Gonz�les-Le�n, 1991, p. 62; Scott, 2002, p.410.
Mitchell (2013a) provides additional synonymy
Type specimens.- Holotype CAS.66651-10 (California Academy of Sciences), Palmer No. 2168 from Soyatlan Adentro, Jalisco, Mexico; housed at California Academy of Science.
Diagnosis.- RV long, straight to curved, LV curved to partly coiled; valve cross section ovate to rectangular with rounded antero-ventral keel; distinct external ligament groove; RV with tooth large, with few small pallial canals and locally canals in myocardinal areas of RVs; LV with two large teeth, triangular to oval in cross section; shell wall with an outer row of ellipsoidal canals and one to three inner rows of ovate canals.
Discussion.- Mitchell (2013a) considered that the types
of Caprinuloidea perfecta gracilis Palmer, 1928, to be smaller specimens of C.
perfecta. Palmer differentiated C. perfecta gracilis
by its narrower cross section in relation to its great length, which Mitchell
(2013a) includes as a subspecies of C. perfecta. This species is also
from the Lower Albian Mural Formation, Sonora, Mexico (Fig.
4.11-12 ![]() ;
Gonz�les-Le�n
et al., 2008). Palmer (1928) erected Caprinuloidea
costata for specimens having distinct growth rings. However Scott and
Gonz�les-Le�n
(1991) considered these features to be within the range of
variation of C. perfecta because the
type material of each species was from the same locality.
;
Gonz�les-Le�n
et al., 2008). Palmer (1928) erected Caprinuloidea
costata for specimens having distinct growth rings. However Scott and
Gonz�les-Le�n
(1991) considered these features to be within the range of
variation of C. perfecta because the
type material of each species was from the same locality.
Occurrence.- C. perfecta is from lower Albian strata at Soyatlan de Adentro, Jalisco and El Abra Formation in Tamaulipas, Mexico (Aguilar P�rez, 2008).
Caprinuloidea romeri Mitchell, 2013a
(Figs. 4.4-5 ![]() ,
6.4-6
,
6.4-6 ![]() ,
7.5-6
,
7.5-6 ![]() ,
9.1, 9.5, 9.9-12
,
9.1, 9.5, 9.9-12 ![]() ,
12.11
,
12.11 ![]() )
)
Caprinuloidea romeri Mitchell, 2013a, p. 52-60, Figs. 3A-E, 4A-D
Type specimens.- Holotype at Texas Museum of Natural History TMM.UT10932; paratypes TMM.UT10930.1, 11268, 33867, and 10922; Edwards Limestone, Austin, Texas area.
Diagnosis.- RV long, straight to torted, LV coiled, external ligament groove, pallial canals in RV central tooth and myocardinal areas, LV myophores partly invaded by pallial canals, outer valve wall with narrow tear-drop pallial canals, inner wall with three to six rows of ovate canals (Mitchell, 2013a).
Discussion.- Until 2013 C. perfecta included specimens from both the Trinity and Fredericksburg groups (Coogan, 1977; Scott, 2002; Scott and Filkorn, 2007). However Mitchell (2013a) distinguished C. romeri from C. perfecta by internal properties. C. perfecta has small pallial canals in the central tooth of the RV and rarely in the myocardinal area as opposed to C. romeri, which has large pallial canals in the myocardinal area. Palmer (1928) noted that C. perfecta has canals in the large central tooth of the RV and in the ridge connecting it to the outer wall.
Mitchell (2013a) assigned most specimens in the Edwards Formation to Caprinuloidea romeri Mitchell, "because their pallial canals are regularly developed and some pallial canals invade the myophores." He included in C. romeri the silicified Edwards specimens illustrated by Scott and Weaver (2010). Because, many specimens in the Edwards are partly recrystallized and pallial canals are not clearly visible in the central tooth and myocardinal area, a broad concept of C. perfecta was practical and useful for biostratigraphy. However, Edwards specimens having an external ligament groove are now identified as C. romeri and can be distinguished from those without a groove, which are now placed in the species Youngicaprina sangabrielensis Mitchell, 2013a.
The outcrop specimens in the uppermost beds of the
Fort Terrett Formation (Fig. 6.1,6.5, 6.9-12 ![]() ) have an ovate shape with a
distinct anterior-ventral rounded keel. Therefore the specimens from the Fort
Terrett Formation are placed in C. romeri.
) have an ovate shape with a
distinct anterior-ventral rounded keel. Therefore the specimens from the Fort
Terrett Formation are placed in C. romeri.
The two specimens in the marine member of the Person
Formation in the Shell No. 2 L. Urbanczyk at 10,818.5 ft (Fig.
6.4-6 ![]() ) show the
external groove and curved ligament and the characteristic pallial canals in the
shell wall, so they are identified as C.
romeri. However pallial canals in the myocardinal area are partially
obscured by recrystallization.
) show the
external groove and curved ligament and the characteristic pallial canals in the
shell wall, so they are identified as C.
romeri. However pallial canals in the myocardinal area are partially
obscured by recrystallization.
Occurrence.- Type specimens of C. romeri are from limestones of the Edwards Group in the Austin area and in central Texas (Mitchell, 2013a). Here this species is reported in the Fort Terrett Formation in Crocket County and in the Person Formation in the subsurface of Karnes County. In contrast, C. perfecta is from lower Albian outcrops at Soyatlan de Adentro, Jalisco, Mexico; the species has been reported widely in the Caribbean Province including in the Edwards Limestone (Coogan, 1977) and in the Los Picachos and Espinazo del Diablo formations in Sonora, Mexico (Scott and Gonz�les-Le�n, 1991). C. perfecta was included as a member of the Caprinuloidea Interval Zone of the middle to basal upper Albian, which characterizes the Fredericksburg Group in Texas (Scott and Filkorn, 2007). However, specimens reported from strata of the Fredericksburg and Edwards groups are now re-assigned to C. romeri, which becomes the diagnostic taxon of the Caprinuloidea Zone and C. perfecta characterizes a zone of the Glen Rose Formation.
Genus Mexicaprina Coogan, 1973
Type species.- Mexicaprina cornuta Coogan, 1973.
Mexicaprina quadrata Alenc�ster & Oviedo-Garc�a, 1998
(Figs. 10.1-5 ![]() & 12.6-10
& 12.6-10 ![]() )
)
Mexicaprina quadrata Alenc�ster & Oviedo-Garc�a, 1998, p. 172, Fig. 6.2
Holotype.- RV IGM-4581.
Type locality.- El Madro�o, Queretaro, Mexico.
Diagnosis.- Cross section ovate, valve margins flat, keeled at corners; LV conical with acute apex (Alenc�ster and Oviedo-Garc�a, 1998, p. 172).
Discussion.- Specimens in the
uppermost part of the Fort Lancaster Formation are somewhat larger in diameter
than most of the Mexican specimens, 30 to 35 mm compared to 22 mm. The
recrystallized cardinal area precludes detailed comparisons of these specimens
with the free specimens from El Madro�o. M.
quadrata lacks the ridges characteristic of Mexicaprina
cornuta Coogan (1973) and the long spine-like keels of Ichthyosarcolites
[Mexicaprina] alatus (Filkorn, 2002) (Mitchell, 2013b)
(Fig. 12.7 ![]() ).
).
Occurrence.- Mexicaprina species were first described from the El Abra Formation in the Taninul quarry east of Valles, San Luis Potosi, Mexico. In Texas M. cornuta is in uppermost Albian strata (Scott, 2002). The genus characterizes the uppermost upper Albian Mexicaprina Interval Zone (Scott and Filkorn, 2007).
Genus Kimbleia Coogan, 1973
(Figs. 10.4 ![]() & 12.2-5
& 12.2-5 ![]() )
)
Type species.- Kimbleia capacis Coogan, 1973, p. 58, Pl. 1, figs. 1-3.
Type specimens.- Deposited in Instituto de Geolog�a, Universidad de Mexico, Mexico.
Kimbleia albrittoni (Perkins, 1961)
Caprinuloidea? albrittoni Perkins, 1961, p. 78-81, Pl. 22, figs. 14-17; Pl. 23, figs. 1-5.
Type specimens.- Deposited University of Michigan Museum of Paleontology (U.M.M.P.) 32847, 32848-32850.
Discussion.- Coogan (1977) differentiated Kimbleia capacis from Kimbleia albrittoni (Perkins, 1961) by the former's "much larger, suboval to subrectangular canals next to the body cavity and tooth plus accessory cavity in the inner row of marginal canals, and in having two to three rows of polygonal canals" (Coogan, 1977, p. 64). K. albrittoni is indistinguishable from K. capacis and has priority (Scott, 2002; Mitchell, 2013a).
Occurrence.- Kimbleia albrittoni was described from the upper 38 m (125 ft) of the Aurora Limestone in Sierra de Tlahualilo, Coahuila (Perkins, 1961). This species occurs with other taxa that correlate the upper Aurora section with the Washita Group in Texas (Perkins, 1961), which Coogan (1973) considered to be upper Albian. Other locales with Kimbleia are in the upper Albian Segovia, Fort Lancaster and Devils River formations (Coogan, 1973; Scott, 2002). The genus characterizes the late Albian Kimbleia Interval Zone overlying the basal Washita sequence boundary Al SB WA 1 (Scott and Filkorn, 2007).
Genus Neokimbleia Mitchell, 2013a
Type species.- Neokimbleia acutus Mitchell, 2013a.
Diagnosis.- The LV anterior tooth (AT) is larger than the posterior tooth (PT), which has a posterior ridge; the anterior myophore is connected to the anterior tooth; the posterior myophore is long, blade-like inserted into the RV into an elongate cavity. No external ligament groove; ligament is a pear-shaped cavity (Mitchell, 2013a, p. 57).
Neokimbleia acutus Mitchell, 2013a
(Fig. 7.1-4 ![]() )
)
Neokimbleia acutus Mitchell, 2013a, p. 57, 59, Fig. 6A.
Holotype.- RV IGM-4586.
Type locality.- El Abra Formation, El Madro�o, Quer�taro, Mexico.
Diagnosis.- "A species of Neokimbleia with a bifid AT and a triangular flange on the ventral shell margin" (Mitchell, 2013a, p. 57.
Discussion.- The
diagnostic ventral flange is illustrated by specimen II (Fig.
7.1 ![]() ). The genus
Neokimbleia is characterized by the presence of the internal ligament in a "pear-shaped cavity" and the absence of an external ligamental groove. The
posterior tooth cavity is triangular and the myophore cavity is elongate. The
anterior myophore is connected to the anterior tooth. Mitchell (2013a)
proposed that Neokimbleia evolved from
Kimbleia by loss of the external
ligament groove.
). The genus
Neokimbleia is characterized by the presence of the internal ligament in a "pear-shaped cavity" and the absence of an external ligamental groove. The
posterior tooth cavity is triangular and the myophore cavity is elongate. The
anterior myophore is connected to the anterior tooth. Mitchell (2013a)
proposed that Neokimbleia evolved from
Kimbleia by loss of the external
ligament groove.
Occurrence.- The holotype was collected from the upper Albian part of the El Abra Formation at El Madro�o, Quer�taro, Mexico.
Neokimbleia planata (Conrad, 1855)
(Fig. 6.3 ![]() )
)
Caprina planata Conrad, 1855, p. 268.
Neokimbleia planata (Conrad, 1855) Mitchell, 2013a, p. 59-60, Figs. 6B-E; provides complete synonymy.
Holotype.- RV USNM 9891.
Type locality.- Fort Lancaster reference section on U.S. Highway 290, east of Fort Lancaster State Park, Crockett County, Texas.
Diagnosis.- "A species of Neokimbleia with a single, non-bifid anterior tooth, the shell flattened in an anterio-posteriorly direction, with the shell smoothly rounded, without flanges" (Mitchell, 2013a, p. 59).
Discussion.- The specimen in the Person Formation has the same oval, smoothly rounded valve outline characteristic of the holotype. It has a large oval body cavity, an arcuate elongate endomyophoral cavity ending in a narrow tooth socket and an ovate small anterior tooth socket. No external ligament groove is present. The marginal ellipsoidal pallial canals are cut obliquely and are largely recrystallized. Because the original inner shell is recrystallized, no other canals are visible.
Occurrence.- The presence of this species in the Person Formation with C. romeri and Y. sangabrieli indicates that its range is in the Fredericksburg Group. Mitchell (2013a) reported Scott's opinion that the holotype was collected near the Fort Lancaster reference section in Crockett County, Texas, near the site of Fort Lancaster State Park. Subsequent study of this section has failed to identify similar specimens in either the Fort Terrett or the Fort Lancaster formations, although in the Fort Lancaster specimens of Kimbleia and Mexicaprina have been found at this locality (Gargili, 2013; Lai, 2014).
Subfamily Youngicaprininae Mitchell, 2013a
Discussion.- The myocardinal arrangement of this subfamily is distinct from that of the Caprinuloideinae. The LV anterior myophore is a low platform that connects to the anterior tooth and the posterior LV myophore is tooth-like and not connected to the posterior tooth (Mitchell, 2013a). Whereas the LV myophores of the Caprinuloideinae are in cavities separate from the body cavity wall and not connected to the teeth. Mitchell (2013a) included three genera in this subfamily, Texicaprina Coogan, 1973, Jalpania Alenc�ster & Aguilar-P�rez, 1995, and Youngicaprina Mitchell, 2013a. The subfamily ranges from lower to upper Albian in Texas and Mexico.
Genus Texicaprina Coogan, 1973
Type species.- Sabinia kugleri Bouwman, 1937, p. 58-59; not Sabinia vivari Palmer, 1928; see discussion in Mitchell (2013a, p. 61).
Texicaprina kugleri (Bouwman, 1937)
(Fig. 4.6-8 ![]() )
)
Synonymy.- See Mitchell 2013a, p. 63.
Lectotype.- NHMB.G7220 (Natural History Museum Basel, Switzerland); Bouwman, 1937, Figs. 1, 8-9 (designated by Mitchell, 2013a, p. 63).
Type locality.- Point-a-Pierre, Trinidad.
Diagnosis.- "A species of Texicaprina with a weakly tooth-like elongate, posterior myophore separated from the posterior tooth by a gutter filled with pallial canals, and an anterior myophore that envelops the anterior-side of anterior tooth and is ribbed with the ribs comprised of compact shell material and the furrows filled with pallial canals" (Mitchell, 2013a, p. 63).
Occurrence.- The lectotype
is from a limestone block displaced out of stratigraphic position in Trinidad so
its age is uncertain; it has been presumed to be upper Albian (Mitchell,
2013a). The illustrated specimens are from the upper part of the Stuart City
Formation, which is overlain by Upper Albian marl. Associated fossils are Caprinuloidea
romeri, and the foraminifers Barkerina
barkerensis and Dictyoconus
walnutensis. This species is reported from the upper Albian Devils River
Formation on the Pecos River in West Texas (Scott and Kerans,
2004). The Devils River here overlies the Fort Terrett Formation; the species is
in the uppermost part of local high frequency cycle AL 19, which correlates with
the upper part of the Duck Creek Formation in North Texas
(Fig. 2 ![]() ).
).
Genus Youngicaprina Mitchell, 2013a
Type species.- Youngicaprina gloria Mitchell, 2013a, p. 61, from upper Albian carbonates, El Madro�o, Quer�taro.
Youngicaprina sangabrieli Mitchell, 2013a
Youngicaprina sangabrieli Mitchell, 2013a, p. 69-71, Figs. 10, 11A-F.
Holotype.- TMM.UT-36135.5; paratypes TMM.UT-36135.1-4, BEG.2523.1, UT.3388, UT.33871.
Type locality.- Roy Gunn Ranch on North Fork San Gabriel River is now flooded by Lake Georgetown; Edwards Limestone overlies the Walnut Formation and underlies Georgetown Formation. Although Mitchell (2013a, p. 69) reported that the Roy Gunn ranch was on the South San Gabriel River, historical records indicate that it was on the North San Gabriel River (Linda Scarbrough, "A Tale of The Gunns," The Sunday Sun, March 23, 1986, Georgetown).
Discussion.- This species is characterized by a coiled LV, in which the anterior myophore has four or five ribs at an angle to the margin; also it has five to seven rows of pallial canals. The type specimens are from a locality in the Edwards Limestone now drowned by Lake Georgetown. Silicified specimens in the Texas Museum of Non-Vertebrate paleontology are from the Edwards Group in Travis County. It is also found in the Segovia Formation with Kimbleia. Its complete range is from the upper part of the Fredericksburg into the Washita. Rare specimens also are in the Person Formation in the USCE No. 1 Selma core at -261 ft.
Occurrence.- The holotype is from the Edwards Formation. It also occurs in the Segovia Formation with Kimbleia albrittoni (Mitchell, 2013a, p. 71).
This study was financed in part by The University of Tulsa Office of Research and Sponsored Programs to Scott, Campbell, Hojnacki, Wang, and Lai. The Department of Geosciences and Precision Stratigraphy Associates supported field expenses.
Many people supported and promoted this research. Peter R. Rose provided insightful, stimulating scientific discussion of relevant hypotheses and generous hospitality during field work. Ann Molineux and her staff at the Non-Vertebrate Paleontology Collections, Texas Natural Science Center, the University of Texas at Austin, gave ready access to collections and type material. Staff of the Austin Core Research Center, Bureau of Economic Geology, provided assistance with core layout and sampling. Ryan Phelps and Charlie Kerans, Jackson School of Geosciences, the University of Texas at Austin, and Lowell Waite, Pioneer Natural Resources, shared pertinent data and discussions. Simon Mitchell, Department of Geography and Geology, the University of the West Indies, Jamaica, published important taxonomic revisions of Edwards Group rudists and offered constructive suggestions to improve this manuscript. An anonymous second reviewer also suggested critical improvements. Discussions with Peter Skelton, Thomas Steuber, Javier Aguilar P�rez, and Gloria Al�ncaster were seminal to the results of this study.
Aguilar-P�rez J. (2008).- Rudistas del Cretacico inferior y medio, Barremiano-Cenomaniano, Noreste, Centro y Oeste de M�xico.- Universitat Aut�noma de Barcelona, Tesis Doctoral, 139 p.
Aguilar J., Alenc�ster G. & Pons J.M. (2008).- Review of the genus Ichthyosarcolites in America. In: Eighth International Congress on Rudists, Cretaceous Rudists and Carbonate Platforms Abstracts, June 23-25, 2008.- Dokuz Eyl�l University İzmir - Turkey, p. 61.
Alenc�ster G. & Aguilar-P�rez J. (1995).- Jalpania queretana, a new rudist (Bivalvia-Hippuritacea) from the mid-Cretaceous of central Mexico.- Revista Mexicana de Ciencias Geol�gicas, Mexico City, vol. 12, p. 117-122.
Alenc�ster G. & Garc�a-Barrera P. (2008).- Albian radiolitid rudists (Mollusca Bivalvia) from east-central Mexico.- Geobios, Villeurbanne, vol. 41, p. 571-587.
Alenc�ster G. & Oviedo-Garcia A. (1998).- Re-examination of the genera Texicaprina Coogan, Mexicaprina Coogan, and Kimbleia Coogan (Caprinid rudists) from the Albian of central Mexico.- Revista de la Sociedad Mexicana de Paleonotog�a, Mexico City, vol. 8, p. 163-179.
Amsbury D.L. (2003).- Stratigraphy of Fredericksburg Group (Middle-Upper Albian Cretaceous), North-Central Texas. In: Scott R.W. (ed.), Cretaceous stratigraphy and paleoecology, Texas and Mexico.- GCS SEPM Foundation, Houston, Special Publications in Geology no. 1, CD book, Perkins Memorial Volume, p. 227-276.
Bay T.A. Jr (1977).- Lower Cretaceous stratigraphic models from Texas and Mexico. In: Bebout D.G. & Loucks R.G. (eds.), Cretaceous carbonates of Texas and Mexico.- Bureau of Economic Geology, Austin, Report of Investigations, no. 89, p. 12-30.
Blakey R.C. (2010).- Paleogeographic maps.- Colorado Plateau Geosystems, Phoenix, AZ. URL: http://cpgeosystems.com/
Bouwman L.A.H. (1937).- Sur une esp�ce nouvelle du genere Sabinia (Caprinin�s).- Koninklijke Akademie van Wetenschappen te Amsterdam, Proceedings of the Section of Sciences, vol. 40, p. 449-453.
Campbell W.R. (2016, unpublished).- Middle-upper Albian sequence stratigraphy of the interior shelf to shelf margin of the Comanche shelf.- The University of Tulsa, Master's thesis, 128 p.
Conrad T.A. (1855).- Descriptions of one Tertiary and eight new Cretaceous fossils from Texas, in the collection of Major Emory.- Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 7, p. 268-269.
Coogan A.H. (1973).- Nuevos rudistas del Albiano y Cenomaniano de Mexic� y del sur de Texas.- Instituto Mexicano del Petr�leo, Revista, Mexico City, vol. 5, p. 51-74.
Coogan A.H. (1977).- Early and middle Cretaceous Hippuritacea (rudists) of the Gulf Coast. In: Bebout D.G. & Loucks R.G. (eds.), Cretaceous carbonates of Texas and Mexico.- The University of Texas, Bureau of Economic Geology, Report of Investigations, no. 89, p. 32-70.
Cook T.D. (1979).- Exploration history of south Texas Lower Cretaceous carbonate platform.- American Association of Petroleum Geologist, Tulsa, Bulletin vol. 63, p. 32-49.
Cox L.R. et al. (1969).- Mollusca 6 Bivalvia, Part N, Vol. 2. In: Moore R.C. (ed.), Treatise on Invertebrate Paleontology.- The Geological Society of America, Boulder & the University of Kansas, Lawrence, p. N1-N952.
Damestoy G. (1971).- Essai de classification phylog�nique des caprinid�s (Lamellibranches).- Bulletin du Mus�um National d'Histoire Naturelle, Paris, (s�rie 2), vol. 42, p. 1003-1008.
Davis-Strickland E.R. (1980).- A new species of caprotinid rudists from the Fredericksburg Group (Albian) of north central Texas.- Texas Journal of Science, Austin, vol. 32, p. 115-128.
Davis-Strickland E.R. (1988).- A new species of rudist bivalve from the Albian Cretaceous of central Texas, with a review of the genus Eoradiolites Douvill�, 1909.- Bulletin of Paleomalacology, Charlottesville, vol. 1, p. 59-65.
Feray D.E. (1947).- Brusy Creek, Round rock, Williamson County, Texas. In: Hazzard R.T., Feray D.E., Lozo F.E., Durham C.O. & Nunnally J.D. (eds.), Field Conference of the Shreveport Geological Society, September 2-4, 1949, Austin, Texas, p. 30-33.
Filkorn H.F. (2002).- A new species of Mexicaprina (Caprinidae, Coalcomaninae) and review of the age and paleobiogeography of the genus.- Journal of Paleontology, vol. 76, p. 672-691.
Garc�a-Barrera P. (1995).- Toucasia hancockensis (Hippuritacea-Requienidae) in southwestern Mexico.- Revista Mexicana de Ciencias Geol�gicas, vol. 12, p. 191-194.
Gargili D. (2013, unpublished).- Middle-Upper Albian Comanche shelf sequence stratigraphy, Fredericksburg Group, Texas.- The University of Tulsa, Master's thesis, 154 p.
Glancy T.J. Jr, Arthur M.A., Barron E.J. & Kauffman E.G. (1993).- A paleoclimate model for the North American Cretaceous (Cenomanian-Turonian) epicontinental sea. In: Caldwell W.G.E. & Kauffman E.G. (eds.), Evolution of the Western Interior Basin.- Geological Association of Canada, St. John's, Newfoundland, Special Paper 39, p. 219-241.
Gonz�les-Le�n C.M., Scott R.W., L�ser H., Lawton T.F., Robert E. & Valencia V.A. (2008).- Upper Aptian-Lower Albian Mural Formation: Stratigraphy, biostratigraphy and depositional cycles on the Sonoran Shelf, northern Mexico.- Cretaceous Research, vol. 29, p. 249-266.
Hill R.T. (1893).- The paleontology of the Cretaceous formations of Texas: The invertebrate fossils of the Caprina limestone beds.- Washington Biological Society Proceedings, St. Louis, vol. 8, p. 97-108.
Hill R.T. (1901).- Geography and geology of the Black and Grand Prairies, Texas.- U.S Geological Survey, Annual Report, vol. 21, Part 7, 666 p.
Hojnacki R.E. (2016, unpublished).- Middle-Upper Albian (Cretaceous) sequence stratigraphy and geochemical events of the Fort Stockton Basin, Texas.- The University of Tulsa, Master's thesis, 68 p.
Kauffman E.G. (1973).- Cretaceous Bivalvia. In: Hallam A. (ed.), Atlas of palaeobiogeography.- Elsevier Scientific Publishing Company, Amsterdam, p. 353-383.
Kennedy W.J., Cobban W.A., Gale A.S., Hancock J.M. & Landman N.H. (1998).- Ammonites from the Weno Limestone (Albian) in northeast Texas.- American Museum Novitates, New York City, no. 3236, 37 p.
Kennedy W.J., Gale A.S., Hancock J.M., Crampton J.S. & Cobban W.A. (1999).- Ammonites and inoceramid bivalves from close to the Middle-Upper Albian boundary around Fort Worth, Texas.- Journal of Paleontology, Lawrence, vol. 73, p. 1101-1125.
Kerans C. (2002).- Styles of rudist buildup development along the northern margin of the Maverick Basin, Pecos River Canyon, southwest Texas.- Gulf Coast Association of Geological Societies, Houston, Transactions, vol. 52, p. 501-516.
Kerans C., Kempter K., Fitchen B. & Zahm L.C. (1999).- Sequence framework and facies architecture of a Cretaceous carbonate platform: Late Albian of the Pecos River, West Texas. SEPM Field Trip Guidebook, Annual AAPG/SEPM Convention, San Antonio, Texas, April 14-18, 1999.- The University of Texas at Austin, Bureau of Economic Geology, 98 p.
Lai Xin (2014, unpublished).- Upper Albian sequence stratigraphy and geochemical events of Comanche shelf interior Washita Group, southwest Texas.- The University of Tulsa, Master's thesis, 93 p.
Leckie R.M., Bralower T.J. & Cashman R. (2002).- Oceanic anoxic events and plankton evolution: Biotic response to tectonic forcing during the mid-Cretaceous.- Paleoceanography, Washington, vol. 17, p. 13-29.
Lozo F.E. & Smith C.I. (1964).- Revision of Comanche Cretaceous stratigraphic nomenclature, southern Edwards Plateau, southwest Texas.- Gulf Coast Association of Geological Societies Transactions, Houston, vol. 14, p. 285-306.
Lucas S. (2010).- Cretaceous stratigraphy, paleontology, petrography, depositional environments, and cycle stratigraphy at Cerro de Cristo Rey, Do�a Ana County, New Mexico.- New Mexico Geology, Albuquerque, New Mexico, vol. 32, no. 4, p. 103-130.
Mancini E.A., Llin�s J.C., Scott, R.W. & Llin�s R. (2005).- Potential reef-reservoir facies: Lower Cretaceous deep-water thrombolites, onshore ventral Gulf of Mexico.- Gulf Coast Association of Geological Societies Transactions, Houston, vol. 55, p. 505-515.
Mancini E.A. & Scott R.W. (2006).- Sequence stratigraphy of Comanchean Cretaceous outcrop strata of northeast and south central Texas: Implications for enhanced petroleum exploration.- Gulf Coast Association of Geological Societies Transactions, Houston, vol. 56, p. 539-550.
Masse J.-P., Beltramo J., Martinez-Reyes J. & Arnaud-Vanneau A. (2007).- Revision of Albian polyconitid and monopleurid rudist bivalves from the New World. In: Scott R.W. (ed.), Cretaceous rudists and carbonate platforms: Environmental feedback.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication no. 87, p. 221-230.
Matheron P. (1842).- Catalogue m�thodique et descriptif des corps organis�s fossiles du d�partement des Bouches-du-Rh�ne et lieux circonvoisins.- R�pertoire des Travaux de la Soci�t� de Statistique de Marseille, vol. 6, p. 269 p.
Miller B.C. (1984).- Physical stratigraphy and facies analysis, Lower Cretaceous, Maverick Basin and Devils River Trend, Uvalde and Real counties, Texas. In: Smith C.I. (ed.), Stratigraphy and structure of the Maverick Basin and Devils River Trend, Lower Cretaceous, southwest Texas- a field guide and related papers.- San Antonio Geological society, Field Trip Guidebook, Annual Meeting of the AAPG and SEPM, p. 2-33.
Mitchell S.F. (2013a).- A revision of selected Lower Cretaceous American caprinoid rudists: Implications for phylogeny and biostratigraphy.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 45, p. 47-75.
Mitchell S.F. (2013b).- A new rudist bivalve Curtocaprina clabaughikinsorum gen. et sp. nov. from the Middle Albian of Texas and its bearing on the origin of the Ichthyosarcolitidae Douvill�.- Caribbean Journal of Earth Science, vol. 45, p. 85-89.
Moore C.H. Jr (1964).- Stratigraphy of the Fredericksburg Division, south-central Texas.- The University of Texas at Austin, Bureau of Economic Geology, Report of Investigations, no. 52, 48 p.
Newell N.D. (1965).- Classification of the Bivalvia.- American Museum Novitates, New York City, no. 2206, 25 p.
Orbigny A. d' (1847).- Sur les Brachiopodes ou Palliobranches (deuxi�me m�moire).- Comptes Rendus hebdomadaires des S�ances de l'Acad�mie des Sciences, Paris, vol. 25, no. 7, p. 266-269.
Palmer R.H. (1928).- The rudistids of southern Mexico.- California Academy of Sciences Occasional Papers, San Francisco, no. 14, 137 p.
Perkins B.F. (1961).- Biostratigraphic studies in the Comanche (Cretaceous) Series of northern Mexico and Texas.- Geological Society of America, Boulder, Colorado, Memoir 83, 138 p.
Phelps R.M., Kerans C., Loucks R.G., Da Gama Rui O.B.P., Jeremiah J. & Hull D. (2014).- Oceanographic and eustatic control of carbonate platform evolution and sequence stratigraphy on the Cretaceous (Valanginian-Campanian) passive margin, northern Gulf of Mexico.- Sedimentology, vol. 61, p. 461-496.
Reboulet S., Szives O., Aguirre-Urreta B., Barag�n R., Company M., Idakieva V., Ivanov M., Kakabadze M.V., Moreno-Bedmar J.A., Sandoval J. Baraboshkin E., �ağlar M.K., F�zy I., Gonz�lez-Arreola C., Kenjo S., Lukeneder A., Raisossadat S.N., Rawson P.F. & Tavera J.M. (2009).- Report on the 5th International meeting of the IUGS Lower Cretaceous ammonite working group, the Kilian Group (Ankara, Turkey, 31st August, 2013).- Cretaceous Research, vol. 50, p. 126-137.
Roemer F. (1852).- Der Kreidebildungen von Texas und ihre organischein Einschl�sse.- Adolph Marcus, Bonn, 100 p. (11 Pls.).
Rose P.R. (1972).- Edwards Group, surface and subsurface, Central Texas.- The University of Texas at Austin, Bureau of Economic Geology, Report of Investigations, no. 74, 198 p.
Salvador A. (1991).- Chapter 14. Origin and development of the Gulf of Mexico basin.- In: Salvador A. (ed.), The geology of North America, the Gulf of Mexico Basin.- The Geological Society of America, Boulder, vol. J, p. 389-444.
Scott R.W. (1970).- Stratigraphy and sedimentary environments of Lower Cretaceous rocks, southern Western Interior.- American Association of Petroleum Geologists, Tulsa, Bulletin, vol. 54, p. 1225-1244.
Scott R.W. (1990).- Models and stratigraphy of Mid-Cretaceous reef communities, Gulf of Mexico.- SEPM (Society for Sedimentary Geology), Tulsa, Concepts in Sedimentology and Paleontology, vol. 2, 102 p.
Scott R.W. (1993).- Cretaceous carbonate platform, U.S. Gulf Coast. In: Simo J.A.T., Scott R.W. & Masse J.-P. (eds.), Cretaceous carbonate platforms.- American Association of Petroleum Geologists, Tulsa, Memoir 56, p. 97-109.
Scott R.W. (1995).- Global environmental controls on Cretaceous reefal ecosystems.- Palæogeography, Palæoclimatology, Palæoecology, vol. 119, p. 187-199.
Scott R.W. (2002).- Albian caprinids re-evaluated.- Journal of Paleontology, Lawrence, vol. 76, p. 408-423.
Scott R.W. (2007).- Introduction, In: Scott R.W. (ed.), Cretaceous rudists and carbonate platforms: Environmental feedback.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication no. 87, p. 5-8.
Scott R.W. (2014).- Cretaceous chronostratigraphic database: Construction and applications.- Carnets Geol., Madrid, vol. 14, no. 1, p. 15-37. DOI: 10.4267/2042/53522
Scott R.W. & Filkorn H.F. (2007).- Barremian-Albian rudist zones, U.S. Gulf Coast. In: Scott, R.W. (ed.), Cretaceous rudists and carbonate platforms: Environmental feedback.- SEPM (Society for Sedimentary Geology), Tulsa, Special Publication no. 87, p. 167-180.
Scott R.W., Benson D.G., Morin R.W., Shaffer B.L. & Oboh-Ikuenobe F.E. (2003).- Integrated Albian-Lower Cenomanian chronostratigraphy standard, Trinity River Section, Texas. U.S. Gulf Coast Cretaceous Stratigraphy and Paleoecology. In: Scott R.W. (ed.), Cretaceous stratigraphy and paleoecology, Texas and Mexico.- GCS SEPM Foundation, Houston, Special Publications in Geology no. 1, CD book, Perkins Memorial Volume, p. 277-332.
Scott R.W. & Gonz�les-Le�n C. (1991).- Paleontology and biostratigraphy of Cretaceous rocks, Lampazos area, Sonora, Mexico.- Geological Society of America, Special Paper 254, p. 51-67.
ScottKerans R.W. & Kerans C. (2004).- Late Albian carbonate platform chronostratigraphy, Devils River Formation cycles, west Texas.- Courier Forschungs-Institut Senckenberg, vol. 247, p. 129-148.
Scott R.W. & Kidson E.J. (1977).- Lower Cretaceous depositional systems, West Texas. In: Bebout D.G. & Loucks R.G. (eds.), Cretaceous carbonates of Texas and Mexico: Applications to subsurface exploration.- Bureau of Economic Geology, The University of Texas at Austin, Report of Investigations, no. 89, p. 169-181.
Scott R.W. & Weaver M. (2010).- Ontogeny and functional morphology of a Lower Cretaceous Caprinid rudist (Bivalvia, Hippuritoida).- Turkish Journal of Earth Sciences, Ankara, vol. 19, p. 527-542.
Skelton P.W. (2013).- Rudist classification for the revised Bivalvia volumes of the 'Treatise on Invertebrate Paleontology'.- Caribbean Journal of Earth Science, Kingston, vol. 45, p. 9-33.
Skelton P.W. & Masse J.-P. (2000).- Synoptic guide to Lower Cretaceous rudist bivalves of Arabia. In: Alsharhan A.S. & Scott R.W. (eds.), Middle East models of Jurassic/Cretaceous carbonate systems.- SEPM (Society for Sedimentology), Tulsa, Special Publication no. 69, p. 89-99.
Small T.A., Hanson J.A. & Hauwert N.M. (1996).- Geologic framework and hydrologic characteristics of the Edwards aquifer outcrop (Barton Springs segment), northeastern Hays and southwestern Travis counties, Texas.- U.S. Geological Survey, Washington, Water-Resources Investigations Report 96-4306, 15 p.
Smith C.I., Brown J.B. & Lozo F.E. (2000).- Regional stratigraphic cross sections, Comanche Cretaceous (Fredericksburg - Washita Division), Edwards and Stockton plateaus, West Texas: Interpretation of sedimentary facies, depositional cycles, and tectonics.- The University of Texas at Austin, Bureau of Economic Geology, 39 p.
Smith C.I. & Brown J.B. (1983).- Introduction to road log; Cretaceous stratigraphy and first day road log. In: Ketternbrink E.C. Jr (ed.), Structure and stratigraphy of the Val Verde Basin-Devils River uplift, Texas.- West Texas Geological Society, Midland, Publication no. 83-77, 26 p.
Steuber T., Scott R.W., Mitchell S.F. & Skelton P.W. (in press).- Stratigraphy and diversity dynamics of Jurassic-Cretaceous genera of the Hippuritida Newell 1965 (rudist bivalves).- Treatise of Invertebrate Paleontology, Lawrence.
Tyrrell W. & Scott R.W. (1989).- Early Cretaceous shelf margins, Vernon Parish, Louisiana.- American Association of Petroleum Geologists, Tulsa, Special Publication no. 27, p. 11-17.
Waite L.E., Scott, R.W. & Kerans C. (2007).- Middle Albian age of the regional dense marker bed of the Edwards Group, Pawnee Field, south-central Texas.- Gulf Coast Association of Geological Societies, Houston, Transactions, vol. 57, p. 759-774.
Wang Y. (2014, unpublished).- Chemostratigraphy of Upper Albian (Lower Cretaceous) Comanche shelf margin.- The University of Tulsa, Master's thesis, 169 p.
White C.A. (1884).- On Mesozoic fossils.- U.S. Geological Survey, Washington, Bulletin 4, 36 p.
Whitney M. (1952).- Some new Pelecypoda from the Glen Rose Formation of Texas.- Journal of Paleontology, Lawrence, vol. 26, p. 697-707.
Wilbert W.P. (1967).- Stratigraphy of the Georgetown Formation, Central Texas. In: Hendricks L. (ed.), Comanchean (Lower Cretaceous) stratigraphy and paleontology of Texas.- Permian Basin Section, Tulsa, SEPM, Publication no. 67-8, p. 257-285.
Wilson W.F. (1986).- The Stuart City reef: Problems with stratigraphic nomenclature and exploration. In: Stapp W.L. (ed.), Contributions to the geology of south Texas.- South Texas Geological Society, San Antonio, p. 422.
Winter J.A. (1961).- The Atascosa Group of the Lower Cretaceous of south Texas.- South Texas Geological Society, San Antonio, Bulletin, vol. 1, no. 11, p. 15-21.
Yurewitz D.A., Marler T.B., Meyerholtz K.A. & Siroky F.X. (1993).- Early Cretaceous carbonate platform, north rim of the Gulf of Mexico, Mississippi and Louisiana. In: Simo J.A.T., Scott R.W. & Masse J.-P. (eds.), Cretaceous carbonate platforms.- American Association of Petroleum Geologists, Tulsa, Memoir 56, p. 81-96.
Young K. (1957).- Upper Albian (Cretaceous) Ammonoidea from Texas.- Journal of Paleontology, Lawrence, vol. 31, p. 1-33.
Young K. (1959).- Techniques of mollusk zonation in Texas Cretaceous.- American Journal of Science, New Haven, Connecticut, vol. 257, p. 752-769.
Young K. (1966).- Texas Mojsisovicziinae (Ammonoidea) and the zonation of the Fredericksburg.- Geological Society of America, Boulder, Memoir 100, p. 5-119.
Young K. (1979).- Lower Cenomanian and Late Albian (Cretaceous) ammonites, especially Lyelliceridae, of Texas and Mexico.- The Texas Memorial Museum, Austin, Bulletin 26, 99 p.
Young K. (1986).- Cretaceous marine inundations of the San Marcos platform, Texas.- Cretaceous Research, vol. 7, p. 117-140.