![]()
Contents
[Introduction] [Material] [Description of "Lingula" krausei Dames] [Systematics]
[Complementary data] and ... [Bibliographic references]
Institute of Paleobiology, Polish Academy of Sciences, ul. Twarda 51/55, PL-00-818 Warszawa (Poland)
BrachNet, 20, Rue Chaix, F-13007 Marseille (France)
Published online in final form (pdf) on December 12, 2016
[Editor: Bruno Granier; technical editor:
Christian C. Emig; language editor: Stephen
Carey]
![]()
The material under study has previously been identified as Lingula krausei, collected from glacial erratics at Cape Rozewie, Poland. All specimens come from glauconitic sandstone of Cenomanian age. Dames (1874) identified his Lingula sp. as L. krausei which differs in shell outline from Lingula truncata. This species, dedicated to A. Krause, together with L. subovalis, is considered characteristic of the Cenomanian. Several new species and genera have recently been described or redescribed and compared to our specimens, here identified as Lingularia similis. By priority (ICZN, 1999), the type species L. similis is currently synonymized with Lingula krausei. A new genus Eolingularia, within the Family Lingulidae, is here described, with Lingularia siberica Biernat et Emig, 1993, as type species. This new genus ranges from the Carboniferous to the Triassic in Russia, China, and Spain. Possible synonymies are discussed.
• Brachiopoda;
• Lingulidae;
• Mesozoic;
• Cenomanian;
• new genus.
Bitner M.A. & Emig C.C. (2016).- On the type species of Lingularia, and description of Eolingularia n. gen.- Carnets Geol., Madrid, vol. 16, no. 22, p. 543-555.
Au sujet de l'espèce type de Lingularia, et description de Eolingularia n. gen.- Le matériel étudié a été précédemment identifié sous Lingula krausei, récolté dans des blocs erratiques du cap Rozewie (Poméranie, Pologne). Tous les échantillons proviennent des grès glauconieux d'âge cénomanien. Dames (1874) décrit sa Lingula sp. sous L. krausei qui, selon lui, diffère par le contour de la coquille de Lingula truncata. Cette espèce, dédiée à A. Krause, est considérée comme caractéristique du Cénomanien, de même que L. subovalis. Plusieurs espèces nouvelles et de nouveaux genres ont été décrits récemment ou redécrits et comparés à nos spécimens qui ont été finalement décrits et identifiés comme appartenant à Lingularia similis. Selon l'ICZN (1999), l'espèce type L. similis est actuellement synonyme de Lingula krausei. Au sein de la famille Lingulidae, un nouveau genre a été décrit sous Eolingularia avec Lingularia siberica Biernat et Emig, 1993 ; l'extension géologique va du Carbonifère au Trias, en Russie, Chine, Espagne. De possibles synonymies sont discutées.
• Brachiopoda ;
• Lingulidae ;
• Mésozoïque ;
• Cénomanien ;
• nouveau genre.
Lingulides, because of their very low taphonomic potential, are rare fossils and only catastrophic events allow their preservation in the fossil record. Moreover, as external shell characters have no taxonomic value, the identification of lingulides requires examination of soft body parts or their scars in fossil forms, i.e., the muscle scar pattern and the disposition of the main mantle canals (Biernat & Emig, 1993; Emig, 2003).
|
Click on thumbnail to enlarge the image. |
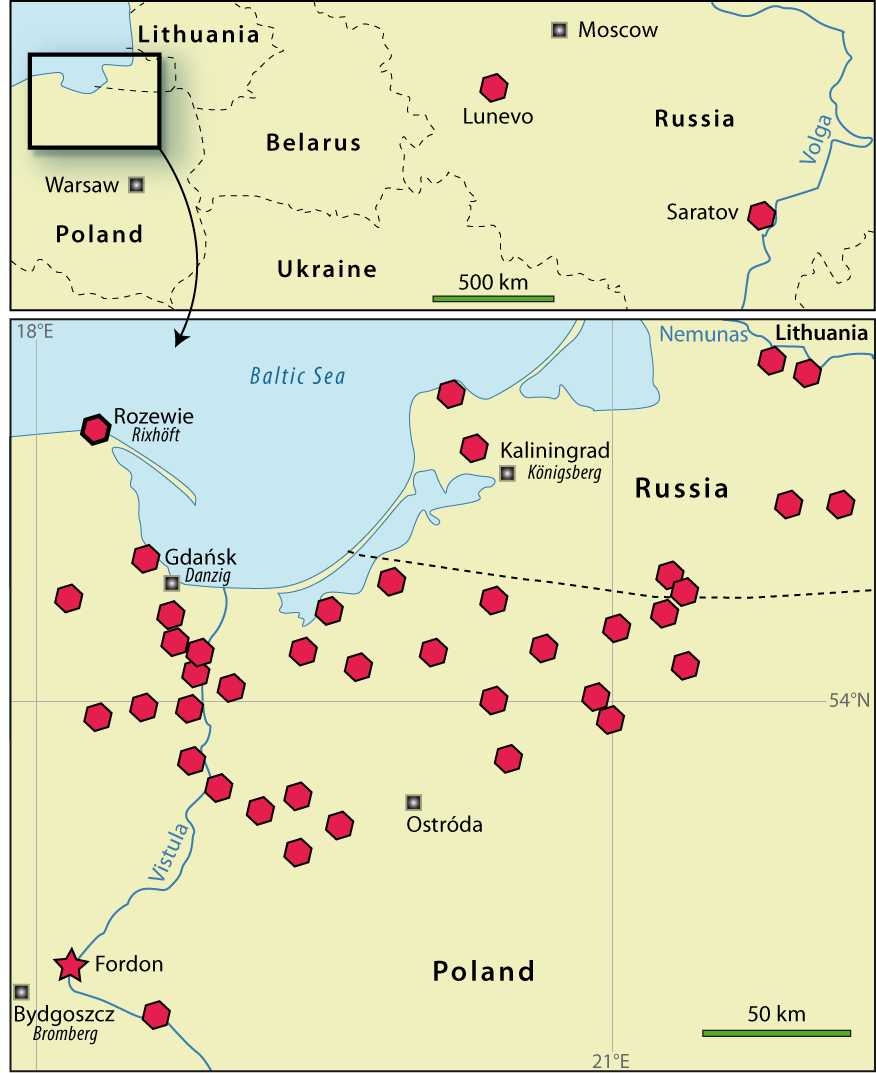
Figure 1: Distribution of "Lingula" krausei from data of Dames (1873, 1874), Kiesow
(1880, 1881), Nötling
(1885), Leonov
(1974), Kharitonov et al.
(2003), Saltykov & Pervushov
(2007). |
The first record of Lingula krausei is in Dames (1873) as Lingula sp. from near Bydgoszcz, Poland (in German: Bromberg ) (Fig. 1 ![]() ). The outcrop of diluvial Quaternary sand with Upper Cretaceous (Cenomanian) glacial erratics is located in Fordon, now a district in Bydgoszcz, near of the confluence of the Brda (in German: Brahe) River and the Vistula (in German: Weichsel) River. The valve fragments occur at two levels. Dames (1873) noted the species' similarities with Lingula rauliniana Orbigny and L. truncata Sowerby. The former species was synonymised with the latter by Davidson
(1852), who described another Cretaceous species, Lingula
subovalis, distinct from L. truncata and L. ovalis Davidson "by its more regularly oval shape, and much smaller size": from
Fig. 29a, a dorsal valve, of Davidson (1852): H/L = 0,57 and Lc = 35% (Tables 1 & 2).
). The outcrop of diluvial Quaternary sand with Upper Cretaceous (Cenomanian) glacial erratics is located in Fordon, now a district in Bydgoszcz, near of the confluence of the Brda (in German: Brahe) River and the Vistula (in German: Weichsel) River. The valve fragments occur at two levels. Dames (1873) noted the species' similarities with Lingula rauliniana Orbigny and L. truncata Sowerby. The former species was synonymised with the latter by Davidson
(1852), who described another Cretaceous species, Lingula
subovalis, distinct from L. truncata and L. ovalis Davidson "by its more regularly oval shape, and much smaller size": from
Fig. 29a, a dorsal valve, of Davidson (1852): H/L = 0,57 and Lc = 35% (Tables 1 & 2).
Dames (1874) assigned his Lingula sp. to Lingula krausei which differs in its shell outline from Lingula truncata. This new species, dedicated to A. Krause, and L. subovalis are considered characteristic of the Cenomanian. According to Nötling (1881) L. krausei occurs in middle and upper Cenomanian erratics found in the valley of the Vistula River and in the lower part of the Nemunas River (formerly Memel), located on the northern border of the Russian enclave of Kaliningrad.
Kiesow (1880, 1881) pointed out that L. krausei had been recorded in abundance in the vicinity of Gdańsk (formerly Danzig), where the rocks are in places composed solely of lingulides, in particular in Łęgowo (in German: Langenau or Lang Heinersdorf).
Nötling (1885) stated that "wenn auch die Lage der Muskelmale im Grossen und Ganzen dieselbe ist, wie bei der lebenden Lingula anatina, so ergeben sich doch bei genauerer Betrachtung kleine Abweichungen, die eine Besprechung verdienen." His figures (Nötling,
1885:
Pl. XXVI) of a ventral and dorsal valve of Lingula krausei are corroborated by our figures (Figs. 2 ![]() - 3
- 3 ![]() ).
).
According to Nötling (1885), the number of valves and fragments may be very important in the "Lingula-Sandstein". This may be the consequence of the extreme events responsible for massive fossilization of the lingulides (see Emig, 1986). In the upper layers of the outcrops, species diversity is strongly reduced while the number of fossilized individuals markedly increases, till Lingula krausei becomes the sole species preserved in the sandstone (Nötling, 1885). Such events took place in very shallow environments, called "Strandfacies" by Nötling (1885). A second brachiopod species Rhynchonella depressa (Sowerby, 1825) has been reported but not in the same layers and is rather uncommon (Nötling, 1885).
Russian occurrences of Lingula krausei with associated fauna are reported in the vicinity of Saratov by Leonov
(1974) and Kharitonov et al.
(2003) and in Cenomanian terrigenous beds of the Tersa depression in the Kaluzhskaya region by Saltykov & Pervushov
(2007) (Fig. 1 ![]() ).
).
In this paper, on the basis of a new, excellently preserved material we re-describe and re-illustrate "Lingula" krausei, and establish its systematic position. Recently, several new species and new genera have been described or redescribed, in particular by Smirnova & Ushatinskaya (2001), Peng & Shi (2008), Holmer & Bengtson (2009), Smirnova et al., 2015), Posenato (2016). Their results will be discussed herein, where the validity of several taxa is considered. Consequently, we propose a new genus Eolingularia with Lingularia siberica Biernat et Emig, 1993, as type species.
Table 1: Width/Length, Height/L and H/W ratios "Lingula" krausei, compared to specimens of Lingularia similis and other species of the genera Lingularia, Eolingularia, Sinolingularia, Sinoglottida, and to the extant genera Lingula and Glottidia. Data of Sinolingularia and Sinoglottidia from Peng & Shi (2008).
| species | n | W/L | n | H/L | H/W | L | |||
| range | mean | range | mean | range | mean | range | |||
| "L." krausei | 10 | 0.44 - 0.57 | 0.51 | 9 | 0.11 - 0.31 | 0.18 | 0.22 - 0.60 | 0.36 | 12.6 - 22.0 |
| L. similis | 66 | 0.44 - 0.75 | 0.62 | 12 | 0.19 - 0.23 | 0.23 | 0.36 - 0.44 | 0.39 | 7.9 - 29.2 |
| L. smirnovae | 17 | 0.47 - 0.55 | 0.51 | 1 | 0.12 | 0.12 | 0.23 | 0.23 | 11.4 - 23.3 |
| L. sp. | 43 | 0.62 - 0.75 | 0.67 | 5.7 - 15.4 | |||||
| E. siberica | 9 | 0.45 - 0.56 | 0.50 | 4.3 - 8.5 | |||||
| S. huananensis | 17 | 0.47 - 0.72 | 0.58 | 2.8 - 5.3 | |||||
| S. yini | 20 | 0.45 - 0.72 | 0.63 | 2.4 - 4.6 | |||||
| S. archiboldi | 17 | 0.53 - 0.75 | 0.66 | 1.4 - 4.3 | |||||
| Lingula | 135 | 0.44 - 0.53 | 0.43 - 0.48 | 95 | 0.06 - 0.12 | 0.10 | 0.14 - 0.25 | 0.20 | |
| except L. adamsi | 31 | 0.54 - 0.70 | 0.63 | 19 | 0.11 - 0.18 | 0.13 | 0.16 - 0.26 | 0.20 | |
| and L. tumidula | 7 | 0.58 - 0.69 | 0.63 | ||||||
| Glottidia | 35 | 0.32 - 0.44 | 0.33 - 0.40 | 35 | 0.09 - 0.11 | 0.09 | 0.15 - 0.24 | 0.20 | |
The material studied comes from glacial erratics collected at Cape Rozewie, Poland (Fig. 1 ![]() ), and consists of several whole valves and tens of fragments of different sizes, all specimens occurring in glauconitic sandstone of Cenomanian age. The material is housed in the Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland, under the number ZPAL Bp.78.
), and consists of several whole valves and tens of fragments of different sizes, all specimens occurring in glauconitic sandstone of Cenomanian age. The material is housed in the Institute of Paleobiology, Polish Academy of Sciences, Warsaw, Poland, under the number ZPAL Bp.78.
Table 2: Variations in the length of the lophophoral cavity and the anterior mantle canals in "Lingula" krausei, compared to the previously reported specimens belonging to the genus Lingularia, Eolingularia, Sinolingularia, Sinoglottidia, and to the extant Glottidia and Lingula. Length of the lophophoral cavity (expressed as a percentage) is the distance between the distal limit of the muscle scars (i.e., anterior adductors on the ventral valve and anterior obliques on the dorsal valve) and the anterior margin of the valve relative to the whole valve length. Mantle canal tip (expressed as a percentage) is the distance between the tip of the canals and the anterior margin of the valve relative to the whole valve length. Data of Sinolingularia and Sinoglottidia from Peng & Shi (2008).
| species | nV/nD | Lophophoral cavity (Lc) | Mantle canal tip (Mct) | ||||||
| Ventral % | Dorsal % | Ventral % | Dorsal % | ||||||
| range | mean | range | mean | range | mean | range | mean | ||
| L. krausei | 3/3 | 43 - 46 | 45 | 29 - 38 | 34 | 27 - 39 | 32 | 36 - 41 | 39 |
| L. similis | 10/13 | 37 - 52 | 45 | 27 - 38 | 33 | 10 - 21 | 14 | 10 - 22 | 17 |
| L. smirnovae | 0/1 | 22 | 22 | ||||||
| E. siberica | 6/6 | 41 - 50 | 46 | 29 - 35 | 32 | 41 - 50 | 46 | 28 - 31 | 30 |
| S. huananensis | 2/8 | 36 - 52 | 39 | 36 - 52 | 45 | 17 - 26 | 22 | 17 - 26 | 20 |
| S. yini | 3/8 | 35 - 58 | 47 | 35 - 58 | 46 | 17 - 28 | 21 | 17 - 28 | 21 |
| S. archiboldi | 8/4 | 35 - 52 | 46 | 35 - 52 | 40 | 15 - 24 | 22 | 15 - 24 | 15 |
| Glottidia | 35/35 | 33 - 43 | 38 | 19 - 33 | 26 | 9 - 15 | 13 | 12 - 15 | 14 |
| Lingula | 44/44 | 35 - 46 | 41 | 24 - 35 | 31 | 17 - 22* | 20 | 17 - 23* | 20 |
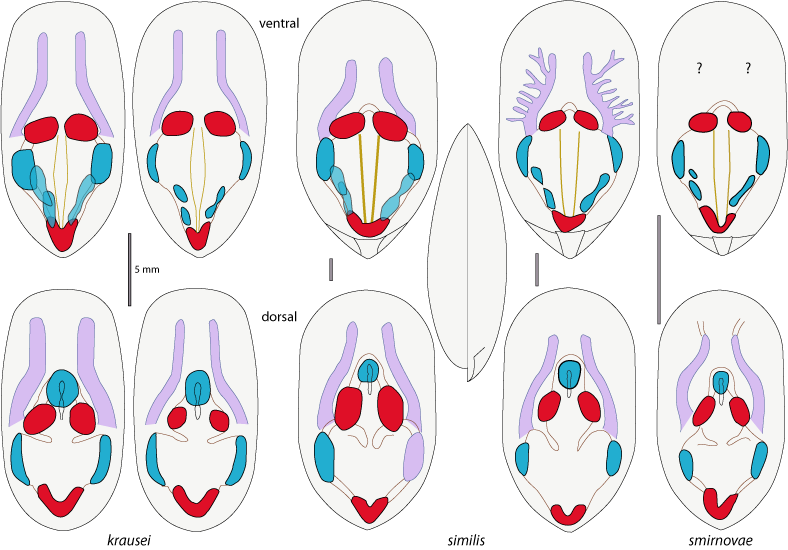
Umbonal region rounded with median beak, rostrated on ventral valve and weak on dorsal valve; large propareas separated by a broad pedicle groove. Body muscles characterized by median internal oblique narrowing near the middle. Mantle canals with S-shaped curves on both sides (Figs. 2 ![]() - 3
- 3 ![]() ).
).
Shell. Elongate oval in outline, lateral margins subparallel; anterior margin generally rounded (Figs. 2 ![]() - 3
- 3 ![]() ); shell surface bears only subconcentric ribbing of variable prominence and spacing. Both valves moderately convex. Maximum width at mid-length of shell. Shell thickened in both valves between anterior adductors. External features, such as shape, size, and dimensional ratios of valves, have no taxonomic value (Emig,
1982, 1983, 2002; Biernat & Emig, 1993). Consequently, they cannot be used to define either genus or species but may be provided as descriptive information additional to diagnosis.
); shell surface bears only subconcentric ribbing of variable prominence and spacing. Both valves moderately convex. Maximum width at mid-length of shell. Shell thickened in both valves between anterior adductors. External features, such as shape, size, and dimensional ratios of valves, have no taxonomic value (Emig,
1982, 1983, 2002; Biernat & Emig, 1993). Consequently, they cannot be used to define either genus or species but may be provided as descriptive information additional to diagnosis.
Ventral valve and body area. Length: 12.6-22.0 mm, mean: 17.3 mm; width: 6.5-11.0 mm, mean: 8.5 mm; width/length ratio: 0.44-0.55, mean: 0.50 (n=7) (Tables 1
- 2; Figs. 2 ![]() - 3
- 3 ![]() ). A pair of narrow, subparallel grooves arises a short distance posterior to anterior adductors to merge, and terminates at level of posterior adductor. Umbonal region pointed to rostrated and continuous with posterolateral margins; large propareas separated by broad pedicle groove. Mantle canals are strongly incurved, like dorsal canals. Perimial line, surrounding muscles (Figs. 3
). A pair of narrow, subparallel grooves arises a short distance posterior to anterior adductors to merge, and terminates at level of posterior adductor. Umbonal region pointed to rostrated and continuous with posterolateral margins; large propareas separated by broad pedicle groove. Mantle canals are strongly incurved, like dorsal canals. Perimial line, surrounding muscles (Figs. 3 ![]() & 7
& 7 ![]() ), is distinct. Internal median oblique muscle scar narrows near its middle. Scar of heart-like shaped posterior adductor commonly covers junction of pair of grooves (body
nerves) (Fig. 3
), is distinct. Internal median oblique muscle scar narrows near its middle. Scar of heart-like shaped posterior adductor commonly covers junction of pair of grooves (body
nerves) (Fig. 3 ![]() ).
).
|
Click on thumbnail to enlarge the image. |
Figure 2: "Lingula" krausei: a. ventral valve (external view); b. dorsal body side; c. ventral body side; d. dorsal valve (external view); e. ventral body side, showing nerve impressions; f. ventral body side. Scales are 0.5 cm. See also Fig. 3 |
Dorsal valve and body area. Length: 16.2-21.8 mm, mean: 19.8 mm; width: 9.3-11.1 mm, mean: 10.4 mm; width/length ratio: 0.51-0.57, mean: 0.53 (n=3) (Tables 1
- 2; Figs. 2 ![]() - 3
- 3 ![]() ). A central narrow ridge extends 2-3 mm posteriorly from anterior oblique muscle scars. Umbonal region is rounded, with very weak median beak. Mantle canals are strongly incurved on both ventral and dorsal side. Perimial line is well-marked.
). A central narrow ridge extends 2-3 mm posteriorly from anterior oblique muscle scars. Umbonal region is rounded, with very weak median beak. Mantle canals are strongly incurved on both ventral and dorsal side. Perimial line is well-marked.
|
Click on thumbnail to enlarge the image. |
Figure 3 : Muscles disposition and variability, and main anterior mantle canals of "Lingula" krausei, Lingularia similis and L. smirnovae. For muscle arrangement, see Fig. 4 |
|
Click on thumbnail to enlarge the image. |
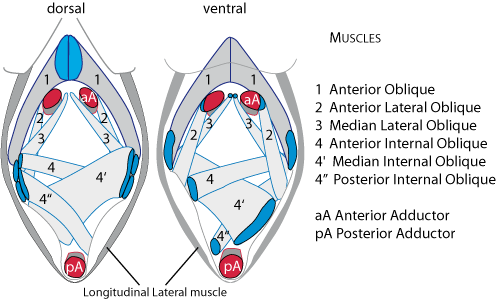
Figure 4: Muscle arrangement on dorsal and ventral faces (Lingula anatina is used as living example allowing body dissection). Note: internal face of a fossil valve acts as a mould because the body is located between two valves. In living specimens valves must be removed to see canal mantles and body muscles. No muscle scars can be seen on such valves. The position is as in life oriented anterior-posterior and left-right. Failure to observe these protocols may introduce confusion and misinterpretation. |
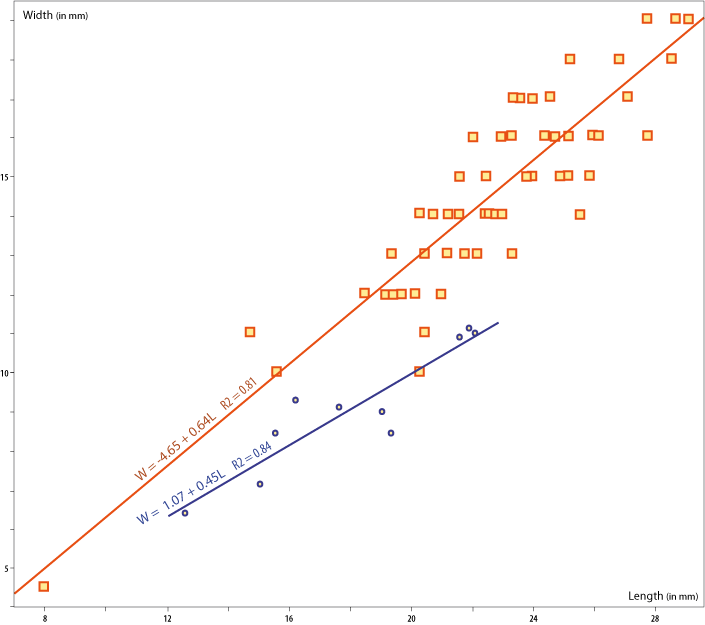
In specimens of L. krausei mean W/L ratio is 0.51 compared to 0.65 in specimens of L. similis, while the other ratios, H/L and H/W, are similar (Table 1; L= shell length, W=width, H=height). Regression curve is W=1.07+0.45L R2=0.84 for krausei and W=-4.65+0.64L R2=0.81 for similis (Fig. 5 ![]() ). Muscular disposition is similar in both species, as well as lophophoral cavity extension (Table 2). However, the extension of tips of mantle canals is similar to that of extant Glottidia (Table 2: three specimens measured), where tips reach closer to anterior margin than in Jurassic specimens.
). Muscular disposition is similar in both species, as well as lophophoral cavity extension (Table 2). However, the extension of tips of mantle canals is similar to that of extant Glottidia (Table 2: three specimens measured), where tips reach closer to anterior margin than in Jurassic specimens.
|
Click on thumbnail to enlarge the image. |
Figure 5: Frequency distribution of shell Width to Length ratios (W/L) and regression plots for "Lingula" krausei, in blue, and Lingularia similis in red. Data from Table 1. L. krausei has been recorded from Cenomanian of Cape Rozewie (Poland) and L. similis from Middle Jurassic of Spitsbergen (Biernat & Emig, 1993). Variation within a species of shell growth may be related to ecological conditions (Emig,
1997). |
Not all characters described herein, e.g., shell outine (see Emig, 2002), can be considered as taxonomically significant, but as helpful ensuring more reliable identification. Variation in taxonomic characters cannot be used to describe a new species, nor the stratigraphic range to identify a fossil taxon. For example, Peng & Shi
(2008) in commenting on Emig's
(2003) diagnosis of the
Family Lingulidae stated that "the pseudointerarea is not always well developed as has been suggested by Williams
(1977) and Archbold
(1981)" (our emphasis). The term "well-developed" is vague and may be related to the size of an individual. Small specimens are immature ones and so have a small pseudo-interarea as noted by Peng & Shi
(2008), Holmer & Bengtson
(2009), and Williams
(1977), the latter observing that a pseudointerarea is absent in Apsilingula parkesensis Williams,
1977. All the specimens studied by these authors are millimetric in length (Table 1; Figs. 6 ![]() - 7
- 7 ![]() ). Such individuals have a broadly rounded anterior margin that becomes nearly straight to straight in mature specimens.
). Such individuals have a broadly rounded anterior margin that becomes nearly straight to straight in mature specimens.
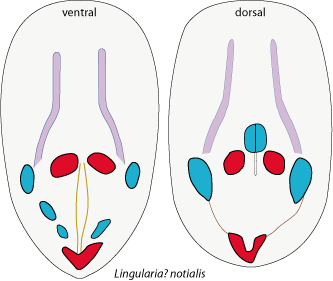
Biernat & Emig (1993) reported that "photographs of internal moulds of "Lingula" krausei from the Cretaceous, sent by L. Holmer, illustrate a great deal of similarity between this latter species and Lingularia similis, particularly in the disposition of mantle canals and of the ventral muscles." So it is surprising that Holmer & Bengtson (2009), when describing the new species Lingularia ? notialis from Brazil did not compare it to krausei. We have done so and here assign it to krausei. Holmer & Bengtson (2009) overlooked some muscular scars and the posterior adductor scar as complete. The additional material has led to a better grasp of the variation in the taxon's characters and, hence, a better description.
|
Click on thumbnail to enlarge the image. |
Figure 6: Muscle disposition established from original figures of Holmer & Bengtson (2009), modified. Original length is respectively 6.2 and 5.8 mm. |
In the case of "Lingula" bagualensis Wilckens, 1905, from southern Patagonia which is morphologically similar to Lingularia? notialis (Fig. 6 ![]() ), Holmer & Bengtson
(1996) initially referred to Lingularia but later conditionally placed it in Credolingula Smirnova in Smirnova & Ushatinskaya, 2001. Nevertheless, Holmer & Bengtson
(2009) pointed out that this species is morphologically similar to species of Lingularia. Their data on taxonomic characters seem to be restricted to a few valves for L. notialis and a dorsal and ventral valve for Credolingula bagualensis. In the diagnosis provided by Holmer and Popov in volume 6, p. 2533, of the Treatise (Selden,
2007), Credolingula is characterized by "ventral valve interior with asymmetrical paired umbonal muscle". Consequently, this genus does not belong to the
Family Lingulidae which has an unpaired posterior adductor muscle (Emig,
2003). But Peng & Shi
(2008) described a trapezoidal and horseshoe-shaped posterior adductor scar. Credolingula is incompletely described; before it is used, all taxonomic characters and a diagnosis must be soundly established, and all possible related species redescribed on the basis of well preserved internal sides of each valve (about ten of each is a minimum) (Fig. 7
), Holmer & Bengtson
(1996) initially referred to Lingularia but later conditionally placed it in Credolingula Smirnova in Smirnova & Ushatinskaya, 2001. Nevertheless, Holmer & Bengtson
(2009) pointed out that this species is morphologically similar to species of Lingularia. Their data on taxonomic characters seem to be restricted to a few valves for L. notialis and a dorsal and ventral valve for Credolingula bagualensis. In the diagnosis provided by Holmer and Popov in volume 6, p. 2533, of the Treatise (Selden,
2007), Credolingula is characterized by "ventral valve interior with asymmetrical paired umbonal muscle". Consequently, this genus does not belong to the
Family Lingulidae which has an unpaired posterior adductor muscle (Emig,
2003). But Peng & Shi
(2008) described a trapezoidal and horseshoe-shaped posterior adductor scar. Credolingula is incompletely described; before it is used, all taxonomic characters and a diagnosis must be soundly established, and all possible related species redescribed on the basis of well preserved internal sides of each valve (about ten of each is a minimum) (Fig. 7 ![]() ).
).
|
Click on thumbnail to enlarge the image. |
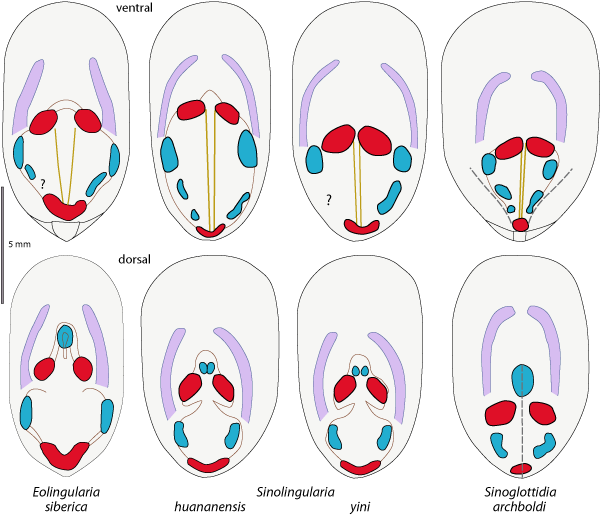
Figure 7: Disposition of muscles and anterior mantle canals in the type species of Eolingularia
n. gen. compared to the species of Sinolingularia and Sinoglottidia, as figured by Peng & Shi
(2008). |
Holmer & Bengtson (2009) alluded in several places to shell structure, especially in comparing Lingula and Lingularia, with reference to Cusack et al. (1999). Mistaken observations made by Cusack et al. (1999) include that several lingulides species belong to Lingularia and others to Glottidia, and that the genus Glottidia is known only from the Eocene and Lingula probably from the Oligocene. Consequently, their data need further investigation based on valid identification of the adult forms.
According to Baliński (1997), more paleontological data are needed to trace the process of gradual morphological modification of the embryonic lingulid shell.
Posenato (2016) is the latest of several authors to observe that the study and application of internal taxonomic characters suggests a more complicated phylogenetic history than previously thought. This point has indeed been emphasized by Emig since 1982. Discovery of new phylogenetic characters in living species is much more efficient than in fossil remains. The latter are very often described on the basis of incomplete characters as well as non-taxonomic ones or on stratigraphic data. It is important to bear in mind that when not all taxonomic characters are preserved the identification cannot be established or a new taxon described. Sometimes several tens or hundreds of specimens must be sampled to yield a sound identification.
Class Lingulata Gorjansky et Popov, 1985
Order Lingulida Waagen, 1885
Family Lingulidae Menke, 1828
Diagnosis (Emig, 2003).- Shell elongate oval to subrectangular, gently and subequally biconvex, subequivalved; lateral margins generally subparallel. Ventral valve with wide triangular pedicle groove and lateral vestigial propareas. Dorsal valve with posterior margin rounded, with a more or less developed median beak; anterior adductor and anterior oblique muscle scars closely spaced. Asymmetrical muscle system with internal oblique muscles; unpaired posterior adductor muscle; pedicle nerve curving around posterior adductor muscle. Mantle canal system bifurcate; vascula lateralia of both valves converging anteriorly to become subparallel; vascula media absent. Long flexible pedicle; lophophore spirolophous, with apices of spires directed medially.
Posenato (2016) suggests emendation of the diagnosis of the Family Lingulidae by adding "asymmetrical paired umbonal or unpaired muscles". Thus one of the main characters of the family is eliminated without explanation nor any reference to the recent diagnosis proposed and discussed by Emig (2003). An asymmetrical muscle system and a single umbonal muscle can be considered as apomorphies at least among the Linguloidea and a symetrical muscle system and a pair of umbonal muscles as plesiomorphies, as shown in the other families of this superfamily. Consistent with Emig's (2003) diagnosis, only five genera are included, i.e., Dignomia, Eolingularia, Lingularia, Lingula and Glottidia. It should be noted that in cladistics a taxon can only have one state of a character. Consequently, the genus Trentingula, described by Posenato (2016) with paired umbonal muscles, cannot be included in the Lingulidae. Nevertheless, it is likely that T. lorigae has an unpaired umbonal muscle (Posenato, 2016, Pl. 1). However no information regarding the umbonal muscle is provided on the other new species, except for the case of unpaired muscles of Pl. 2, fig. 6 (Posenato, 2016). For the most part the umbonal muscles are covered by the valve which has to be removed to identify the type of muscle, and cannot be identified as paired or unpaired.
The subfamilies proposed by several authors (see Peng & Shi, 2008) are not retained here because the diagnoses have characters, which apply exclusively to distinguish fossil taxa. Furthermore, the genera covered by these diagnoses are not identified and the generic diagnoses are modified according to the "new" proposed characters. In some cases taphonomic artefacts, including internal calcification, so-called muscle scars, and folds described as septa, have been used to create new fossil species.
Specimens from the Triassic of Moya (Province of Cuenca, SE Iberian Ranges, Spain; Márquez-Aliaga et al.,
2007) are similar to those previously described from Jalance (Province of Valencia, SE Iberian Ranges, Spain) and Henarejos (Province of Cuenca) by Prats et al.
(1987) and Márquez-Aliaga et al.
(1999), respectively. Here they are assigned to Lingularia. Many valves are deformed, mainly along the longitudinal median line (Fig. 8 ![]() ). Such features should not be interpreted as primary, but as taphonomic. Probable rapid dehydratation and heating induced the mass mortality of the lingulides and deformation of their valves. In Moya about two hundred specimens of Lingularia sp. (Fig. 8
). Such features should not be interpreted as primary, but as taphonomic. Probable rapid dehydratation and heating induced the mass mortality of the lingulides and deformation of their valves. In Moya about two hundred specimens of Lingularia sp. (Fig. 8 ![]() ; Tables 1
- 2) have been carefully studied, but it has not been possible to identify the species. Thus, tens of specimens may sometimes be needed to validate a character and its variability.
; Tables 1
- 2) have been carefully studied, but it has not been possible to identify the species. Thus, tens of specimens may sometimes be needed to validate a character and its variability.
|
Click on thumbnail to enlarge the image. |
Figure 8: Triassic Lingularia sp. from Moya (SE Iberian Ranges, Spain):
A. Showing valve deformation (arrows) related to taphonomic processes (see text).
B. Entire specimen fossilized with pedicle and body parts (ventral side). Remnants of ventral valve are whitish (from Márquez-Aliaga et al.,
2007). |
Genus Lingularia Biernat et Emig, 1993
Diagnosis.- Shell elongate-oval in outline, lateral margins subparallel, anterior margin broadly rounded. Valves weakly to strongly convex. Dorsal valve with posterior margin rounded, narrow median beak sometimes present, and a narrow internal central ridge extending over about 1 mm posterior to anterior oblique muscle scars. Ventral valve with triangular umbo formed by small to large internal propareas, continuous with posterolateral margins and separated by a deep pedicle groove; pair of narrow subparallel, V-shaped, grooves extends internally from the anterior adductor pair to posterior adductor where grooves join. Posterior adductor scar heart-like in outline. Main ventral canals shorter than dorsal canals, S-shaped on ventral and dorsal side.
Lingularia similis Biernat et Emig, 1993
Now regarded as a synonym of Lingula krausei Dames, 1874 - see ICZN (1999): article 67.1.
Lingularia smirnovae Biernat et Emig, 1993.
Lingularia michailovae Smirnova, 2001 in Smirnova & Ushatinskaya (2001) – no diagnosis provided by Smirnova.
Lingularia salymica Smirnova, 2015 in Smirnova et al. (2015) – no diagnosis provided by Smirnova.
Genus Eolingularia Bitner et Emig, n. gen.
Diagnosis.- Shell elongate-oval in outline, lateral margins subparallel, anterior margin broadly rounded. Valves weakly to strongly convex. Dorsal valve with posterior margin rounded, narrow median beak sometimes present, and a narrow internal central ridge extending over about 1 mm posterior to anterior oblique muscle scars. Ventral valve with triangular umbo formed by small to large internal propareas, continuous with posterolateral margins and separated by a deep pedicle groove; pair of narrow subparallel, V-shaped, grooves extends internally from the anterior adductor pair to posterior adductor where grooves join. Posterior adductor scar heart-like in outline. Main ventral and dorsal canals curved and convergent anteriorly (Fig. 7 ![]() ).
).
Etymology of the name: from eo ("dawn" in Greek) and lingularia.
Eolingularia ranges from Carboniferous to Triassic in Russia, China, and Spain.
According to the original diagnosis of Sinolingularia by Peng & Shi (2008), only one character distinguishes this genus from Lingularia: "Both ventral and dorsal mantle canals (vascula lateralia) subparallel and convergent anteriorly" (Peng & Shi, 2008), which is a taxonomic character in Eolingularia. However, these authors stated that "the new genus closely resembles Lingularia Biernat et Emig, 1993 but differs by its clearly separated dorsal antero-lateral muscle scars, V-shaped pedicle nerve grooves in the ventral valve that extend posteriorly without merging, and its rudimentary pseudo-interarea". In our opinion, none of these latter characters allows the genera to be distinguished because of their variability (see Biernat & Emig, 1993; Emig, 2002, 2003).
Similarly, from the original diagnosis of Sinoglottidia Peng et Shi,
2008, it seems clear that a single character distinguishes this genus from Lingularia is: "Dorsal valve with a median septum extending from posterior adductor scar to antero-lateral muscle scars; … Both ventral and dorsal mantle canals (vascula lateralia) subparallel and convergent anteriorly". It should be noted that the so-called median septum coud be a post-mortem folding due to taphonomic processes (see Fig. 8 ![]() ). No view of this septum from the internal side of dorsal valves is shown. Mantle canals are similar as in Eolingularia.
). No view of this septum from the internal side of dorsal valves is shown. Mantle canals are similar as in Eolingularia.
Another argument against the erection of this genus is the small size of the specimens which were probably immature with incompletely developed characters. The difficulty in identifying small (<10 mm) immature individuals has been emphasized repeatedly in living forms by Emig (1982). Moreover, Emig (2002) pointed out that, as in extant lingulides, the importance of a small median ridge or septum generally increases with the growth of the shell. The anterior oblique muscles are anchored on.
Lingularia siberica Biernat et Emig, 1993
Diagnosis.- Numerous fine radial striae and commonly concentric microlines between thicker growth lines generally visible on the shell. Umbonal region rounded on dorsal valve, pointed on ventral valve; small propareas separated by a deep striated pedicle groove, not continuous with the internal valve face. Body muscles characterized by internal anterior oblique located near anterior oblique; internal posterior oblique not observed. Mantle canals almost curved, convergent, on both sides (Fig. 7 ![]() ).
).
Finally, we consider that the status of the Sinolingularia huananensis Peng et Shi,
2008, and Sinoglottidia archboldi Peng et Shi,
2008, remains contentious until their taxonomic characters have been clearly defined and their variability established. For example, Peng & Shi
(2008) distinguish huananensis from yini as follows: "Antero-lateral muscle scars in dorsal valve lacrymiform, separated anteriorly and connected posteriorly, and antero-lateral muscle scars in dorsal valve lacrymiform, separated anteriorly and connected posteriorly"; but, in both species the antero-lateral muscle is "closely located near central muscle scars". Anatomically, on the dorsal side the antero-lateral muscle is named in this paper, the anterior oblique muscle following Emig's nomenclature
(1982) (Fig. 4 ![]() ); the oblique muscles act in scissor-like movements of the valves, an action that requires strong anchorage on the valves (Emig,
1997). Dorsally the anterior oblique muscles are anchored on a small and short median ridge which is lacking or not readily visible in immature individuals – it may be present in
Fig. 6A of Peng & Shi
(2008). The central muscle is the anterior adductor – which, with the posterior adductor acts to close the shell. Further, the positions of the muscles are dependent on the age of the individual, as they shift with the growth of the shell throughout life. Consequently muscle scars show greater or lesser variability in their disposition and shape (Fig. 7
); the oblique muscles act in scissor-like movements of the valves, an action that requires strong anchorage on the valves (Emig,
1997). Dorsally the anterior oblique muscles are anchored on a small and short median ridge which is lacking or not readily visible in immature individuals – it may be present in
Fig. 6A of Peng & Shi
(2008). The central muscle is the anterior adductor – which, with the posterior adductor acts to close the shell. Further, the positions of the muscles are dependent on the age of the individual, as they shift with the growth of the shell throughout life. Consequently muscle scars show greater or lesser variability in their disposition and shape (Fig. 7 ![]() ).
).
Additional comments on several lingulide species cited here follow.
Lingula subspatulata Hall et Meek, 1855
Hall & Meek (1855) remarked about their new species: "The only specimen we have is imperfect, and the shell is preserved only on the margins. It has nearly the proportions of Lingula Rouliniana, (d'Orbigny, Pal. Française, Terrains Crétacés, Brach. p. 10, p. 490, Fig. 1,) but differs in having its greatest width above the middle, while in the European species the greatest width is below the middle. Our species is also more abruptly rounded or sub-truncate at the base. The surface markings are similar." The persistence of this scientific folly may be related to the conservatism of the palaeontological community. A partial explanation is contained in the answer of an American specialist on Mesozoic brachiopods, who in 2003 wrote to one of us (CCE) by email: "[There are people] people who study faunal lists and databases instead of anatomy and taxonomy, and thus perpetuate older nomenclature. As the ecology and distribution of lingulides does not seem to have changed dramatically since their origin, the name "Lingula" has a tremendous amount of inertia" (Emig, 2008).
About this species recorded in the Cenomanian of Texas, Stephenson (1952) pointed out: "In the absence of the soft parts, and the views of the interior, the simplicity and similarity of the shells of many of the members of the group are such as to render difficult their separation into clearly defined species", and further: "Many specimens have suffered more or less mechanical compression". Scott (1970) pointed out that, for specimens ranging from Cenomanian to Maastrichtian, "the absence of radial sculpture might provide a basis for placing these specimens in a new species". Confusion is induced by description of new species on the basis of characters that are neither taxonomically nor phylogenetically valid. Furthermore, a fossil is far from being equivalent to a living individual. On the contrary, living individuals are not described in the same way as fossil ones which are identified solely on shell characters.
Lingula mytiloides Sowerby, 1813
Jack & Etheridge (1892) recorded this species from the Permo-Carboniferous, of Queensland, Australia.
Lingula ovalis Sowerby, 1855 and L. subovalis Davison, 1852
Moore (1870) identified the Cretaceous species, L. subovalis in the same assemblage as the English Jurassic Lingula ovalis Sowerby, and even though he seems to have regarded L. subovalis as a synonym (op. cit., p. 236), the occurrence of L. ovalis is used as evidence for the Jurassic age of the fauna (op. cit., p. 239) – see also Tate (1889).
From the Lower Cretaceous of Queensland, Australia, Jack & Etheridge (1892) stated that the "names probably refer to the same species", following Moore's opinion. Day (1967) referred his Cretaceous lingulide specimens to Lingula cf. subovalis Davidson, and regarded L. ovalis Sowerby as a synonym.
Morton (1982) described Lingula sturti, synonymizing the Australian specimens referred earlier to L. ovalis and L. subovalis. Unfortunately, his description is incomplete and does not allow the identifification the specimens as belonging to a new species.
Miscellaneous Lingulids
Several authors listed Mesozoic Lingula, including Rollier (1916), Terquem (1850) and Davidson (1876). In each case, the description is insufficient to allow identification to species level.
The species described as "Glottidia" rostrata by Thomson and Owen (1979) from the Lower Cretaceous of Alexander Island (Antarctica) does not show the characters of Glottidia (see Emig & Bitner, 2005).
Two species have been recorded by Thomson (1971) in the Lower Cretaceous of Alexander Island, Antarctic Peninsula, L. beanii Philips and L. cf. subovalis Davidson. Both species seem to be cosmopolitan. Such an occurrence can be determined only on the basis of very well defined taxonomic characters which allow confident differentiation of the two forms. This is not the case, so that only one species is likely to be present.
Thanks are due to Prof. J. Dzik (Institute of Paleobiology, Warszawa) for making the material available for study. The photographs were taken by Ms. G. Dziewińska (Institute of Paleobiology, Warszawa) to whom we are grateful. Thanks are due to Prof. F. Álvarez and an anonymous referee for comments, and to Prof. S. Carey for English improvement.
Archbold N.W. (1981).- Lingula (Lingulidae, Brachiopoda) from the Late Artinskian (Permian), Carnarvon Basin, Western Australia.- Proceedings of the Royal Society of Victoria, Melbourne, vol. 92, p. 169-180.
Baliński A. (1997).- Evolution of the embryonic development in lingulid brachiopods.- Acta Palaeontologia Polonica, Warszawa, vol. 42, no. 1, p. 45-56.
Biernat G. & Emig C.C. (1993).- Anatomical distinctions of the Mesozoic lingulide brachiopods.- Acta Palaeontologica Polonica, Warszawa, vol. 38, p. 1-20.
Cusack M., Williams A. & Buckman J.O. (1999).- Chemico-structural evolution of linguloid brachiopod shells.- Palaeontology, vol. 42, no. 5, p. 799-840.
Dames W. (1873).- Notiz über ein Diluvial-Geschiebe cenomanen Alters von Bromberg.- Zeitschrift der Deutschen geologischen Gesellschaft, Berlin, 25, p. 66-70.
Dames W. (1874).- Über Diluvialgeschiebe cenomanen Alter.- Zeitschrift der Deutschen geologischen Gesellschaft, Berlin, vol. 26, no. 4, p. 761-774 (Pl. XXI).
Davidson T. (1852).- A monograph of British Cretaceous Brachiopoda.- Monographs of the Palaeontological Society of London, vol. 1, no. 2, p. 1-54 (5 Pls.).
Davidson T. (1876).- A monograph of the British fossil Brachiopoda. Supplement to the Jurassic and Triassic species.- Palaeontological Society Monographs, London, vol. 4, no. 2, p. 73-185 (Pls. IX-XIII).
Day R.W. (1967).- Marine Lower Cretaceous fossils from the Minmi Member, Blythesdale Formation Roma-Wallumbilla area.- Publications of the geological Survey of Queensland, Brisbane, vol. 335, p. 1-30 (6 Pls.).
Emig C.C. (1982).- Taxonomie du genre Lingula (Brachiopodes, Inarticulés).- Bulletin du Muséum national d'Histoire naturelle de Paris, (4e Série), vol. 4 (Sect. A), no. 3/4, p. 337-367.
Emig C.C. (1983).- Taxonomie du genre Glottidia (Brachiopodes, Inarticulés).- Bulletin du Muséum national d'Histoire naturelle de Paris, (4e Série), vol. 5 (Sect. A), no. 2, p. 469-489.
Emig C.C. (1986).- Conditions de fossilisation du genre Lingula (Brachiopoda) et implications paléoécologiques.- Palæogeography, Palæoclimatology, Palæoecology, vol. 53, p. 245-253.
Emig C.C. (1997).- Ecology of the inarticulated brachiopods. In: Kaesler R.L. (ed.), Treatise on Invertebrate Paleontology. Part H. Revised Brachiopoda.- Geological Society of America, Boulder, and University of Kansas, Lawrence, Vol. 1, p. 473-495.
Emig C.C. (2002).- Tools for linguloid taxonomy: The genus Obolus (Brachiopoda) as an example.- Carnets Geol., Madrid, vol. 1-2, no. A01 (CG2002_A01), p. 1-9.
Emig C.C. (2003).- Proof that Lingula (Brachiopoda) is not a living-fossil, and emended diagnoses of the Family Lingulidae.- Carnets Geol., Madrid, vol. 3, no. L01 (CG2003_L01), p. 1-8.
Emig C.C. (2008).- On the history of the names Lingula, anatina, and on the confusion of the forms assigned them among the Brachiopoda.- Carnets Geol., Madrid, vol. 8, no. A08 (CG2008_A08), p. 1-13.
Emig C.C. & Bitner M.A. (2005).- Glottidia (Brachiopoda: Lingulidae) from the Eocene La Meseta Formation, Seymour Island, Antarctica.- Palæontology, vol. 48, no. 2, p. 423-431.
Hall J. & Meek F.B. (1855).- Descriptions of new species of fossils, from the Cretaceous formations of Nebraska, with observations upon Baculites ovatus and B. compressus, and the progressive development of the septa in Baculites, Ammonites, and Scaphites.- Memoirs of the American Academy of Arts and Sciences, Boston, (2nd series), vol. 5, p. 379-411 (8 Pls.).
Holmer L.E. & Bengtson P. (1996).- Implications of the rare occurrences of brachiopods in the Upper Cretaceous of Sergipe, Brazil. In: Dias-Brito D., Rohn R. & Perinotto J.A. (eds.), Boletim do 4º Simpósio sobre o Cretáceo do Brasil.- UNESP, Rio Claro, p. 67-69.
Holmer L.E. & Bengtson P. (2009).- The first occurrence of a lingulid brachiopod from the Cretaceous of Sergipe, Brazil, with a restudy of "Lingula" bagualensis Wilckens, 1905 from Southern Patagonia.- Paläontogische Zeitschrift, Stuttgart, vol. 83, p. 255-266.
ICZN - International Commission of Zoological Nomenclature (1999).- International code of zoological nomenclature.- Fourth Edition, adopted by the International Union of Biological Sciences, The International Trust for Zoological Nomenclature / The Natural History Museum, London, xxix + 306 p.
Jack R.L. & Etheridge R. (1892).- The geology and palæontology of Queensland and New Guinea.- Beal, Brisbane, 768 p.
Kharitonov V.M., Ivanov A.V. & Seltser V.B. (2003).- Stratigraphy of the Turonian deposits of the Lower Volga.- Volga and Pricaspian Region Resources, Saratov, vol. 36, p. 48-60.
Kiesow J. (1880).- Die geologischen Verhältnisse der Umgebung Danzigs. In: Danzig in naturwissenschaftlicher und medizinischer Beziehung.- Versammlung Deutscher Naturforscher und Ärzte, Danzig, vol. 53, p. 31-39.
Kiesow J. (1881).- Über Cenomanversteinerungen aus dem Diluvium der Umgegend Danzigs.- Schriften der naturforschenden Gesellschaft in Danzig, (N. Folge), Band 5, Heft 1/2, p. 404-417 (1 Pl.).
Leonov G.P. (1974).- Chapter 16: "Zones" as units of regional stratigraphic (geostratigraphic) categories and proper zones (as examplified by stratigraphic subdivision of the Upper Cretaceous deposits of the Volga River region). In: Osnovy stratigrafii (Fundamentals of Stratigraphy).- Izd-vo Moskovskogo universiteta, Moscow, tom 2, p. 146-189.
Márquez-Aliaga A., Emig C.C. & Brito J.M. (1999).- Triassic lingulide brachiopods from the Iberian Range (Spain).- Geobios, Villeurbanne, vol. 32, no. 6, p. 815-821.
Márquez-Aliaga A., Emig C.C. & López-Gómez J. (2007).- Triassic Lingularia (Brachiopoda) from Moya (SE Iberian Ranges, Spain).- XXIII Jornadas de Paleontología, Caravaca de la Cruz, Murcia, Resumen, p. 121-122.
Moore C. (1870).- Australian Mesozoic geology and palaeontology.- Quarterly Journal of the Geological Society of London, vol. 26, p. 226-261.
Morton J.G.G. (1982).- Brachiopoda from the Early Cretaceous of the Southern Eromanga Basin, N.S.W.- Transactions of the Royal Society of South Australia, vol. 106, no. 1/2, p. 35-38.
Nötling F. (1881).- Herr Noetling. In: 3. Protokoll der Juni-Sitzung.- Zeitschrift der Deutschen geologischen Gesellschaft, Berlin, vol. 23, no. 2, p. 352-355.
Nötling F. (1885).- Die Fauna der baltischen Cenoman-Geschiebe.- Paläontologische Abhandlungen, Berlin, vol. 2, no. 4, 199-247, Pls. XXVI-XXIII [IV. Brachiopoda: p. 11-14].
Peng Y. & Shi G.R. (2008).- New Early Triassic Lingulidae (Brachiopoda) genera and species from South China.- Alcheringa, Adelaide, vol. 32, no. 2, p. 149-170.
Posenato R. (2016).- Systematics of lingulide brachiopods from the end-Permian mass extinction interval.- Rivista Italiana di Paleontologia e Stratigrafia, Milano, vol. 122, no. 2, p. 85-108.
Prats J.L., Emig C.C. & Márquez Aliaga A. (1987).- Las Lingula del Triasico: un aspecto tafonómico.- III Jornadas de Paleontología, Leioa 1987, p. 228.
Rollier L. (1916).- Synopsis des Spirobranches (Brachiopodes) jurassiques celto-souabes. 1e Partie (Lingulidés-Spiriféridés).- Abhandlungen der Schweizerische Paläontologische Gesellschaft, Genève, vol. 41, no. 1, 69 p.
Saltykov V.F. & Pervushov E.M. (2007).- The Upper Cretaceous local stratigraphic units from the Tersa Depression (Volgograd Region). In: Cretaceous system of Russia and near foreign countries: Stratigraphical and paleontological problems.- Saratov University, p. 205-221.
Scott R.W. (1970).- Paleoecology and paleontology of the Lower Cretaceous Kiowa Formation, Kansas.- Paleontological Contributions, Lawrence, vol. 52 (Cretaceous 1), 94 p.
Selden P.A. (2007).- Treatise on Invertebrate Paleontology. Part H, Brachiopoda (Revised). Vol. 6: Supplement.- Geological Society of America, Boulder, and University of Kansas, Lawrence, 906 p.
Smirnova T.N. & Ushatinskaya G.T. (2001).- New lingulids (Brachiopoda) from the Lower Cretaceous of European Russia, with notes on the microstructure of their shells.- Paleontologicheskii Zhurnal, Moscow, vol. 4, p. 51-59 [in Russian].
Smirnova T.N., Ushatinskaya G.T., Zhegallo E.A. & Panchenko I.V. (2015).- Lingularia Biernat et Emig, 1993 from the Upper Jurassic of western Siberia: Larval and embryonic shells and shell microstructure.- Paleontological Journal, Moscow, vol. 49, no. 2, p. 125-133.
Stephenson L.W. (1952).- Larger invertebrate fossils of the Woodbine Formation (Cenomanian) of Texas, with decapod crustaceans from the Woodbine Formation of Texas.- U.S. Geological Survey Professional Papers, Washington, vol. 242, 226 p. (59 Pls.).
Tate R. (1889).- On the age of the Mesozoic rocks of the Lake Eyre Basin.- Report First Meeting of the Australasian Association for the Advancement of Science, Sydney, 1888, section C, p. 228-230.
Terquem O. (1850).- Observations sur quelques espèces de Lingules.- Bulletin de la Société géologique de France, Paris, (2e série), vol. 8, no. 2, p. 10-13.
Thomson M.R.A. (1971).- Inarticulate Brachiopoda from the Lower Cretaceous of south-eastern Alexander Island.- British Antarctic Survey Bulletin, Cambridge, vol. 25, p. 85-94.
Thomson M.R.A. & Owen E.F. (1979).- Lower Cretaceous Brachiopoda from south-eastern Alexander Island.- British Antarctic Survey Bulletin, Cambridge, vol. 48, p. 15-36.
Williams A.J. (1977).- Insight into lingulid evolution from the Late Devonian.- Alcheringa, Adelaide, vol. 1, p. 401-406.