◄ Carnets Geol. 16 (26) ►
![]()
Contents
[1. Introduction]
[2. Geological setting and stratigraphic framework]
[3. Systematic paleontology]
[4. Discussion on Paleobiogeography]
[5. Conclusion] and ...
[Bibliographic references]
Department of Geology, Faculty of science, University
of Isfahan, P.O. Box 81746 (Iran)
corresponding
author;
Department of Geology, Faculty of science, University
of Isfahan, P.O. Box 81746 (Iran)
Exploration Directorate of the National Iranian Oil Company, 1st Dead end, Sheikh Bahaee Sq., Seoul Ave., P.O. Box 193956669, Tehran (Iran)
Department of Geology, Faculty of Earth Sciences, Shahid Beheshti University, Tehran (Iran)
CR2P, UMR 7207, Sorbonne
Universités CNRS MNHN UPMC, T. 46-56, E.5, case 104, 4 place
Jussieu, F-75252 Paris cedex 05 (France)
Published online in final form (pdf) on December 31, 2016
[Editor:
Bruno Granier; language editor: Stephen Carey]
![]()
Two species of spatangoid echinoid belonging to Heteraster Orbigny, 1853, are described from Lower Cretaceous deposits (Gadvan and Dariyan formations) at the Kuh-e Gadvan and Banesh sections situated in Zagros Mountains, south of Iran. Heteraster couloni (Agassiz, 1839) is confirmed in the Barremian and Heteraster delgadoi (Loriol, 1884) in the late Aptian-Albian of Iran. The occurrences of Heteraster and other spatangoid echinoids in Iran show a clear distinction between assemblages from the southern and northern margins of the Tethys during the Early Cretaceous.
Spatangoida;
Toxasteridae;
Heteraster;
Dariyan Formation;
Gadvan Formation;
Zagros;
systematics.
Yavari M., Yazdi M., Gahalavand H., Kamyabi Shadan H., Adabi M.H. & Villier L. (2016).- Early Cretaceous Toxasterid Echinoid Heteraster from the high Zagros basin, south of Iran.- Carnets Geol., Madrid, vol. 16, no. 26, p. 615-632.
Oursins toxasteridés du genre Heteraster dans le Crétacé inférieur du haut bassin du Zagros, Sud de l'Iran.- Deux espèces de spatangues attribuées au genre Heteraster Orbigny, 1853, sont décrites à partir de spécimens collectés dans les séries du Crétacé inférieur (formations de Gadvan et de Dariyan) des localités de Kuh-e Gadvan et Banesh, dans les montagnes du Zagros, au sud de l'Iran. La présence d'Heteraster couloni (Agassiz, 1839) est confirmée dans le Barrémien et Heteraster delgadoi (Loriol, 1884) est reconnu pour la première fois dans l'Aptien supérieur/Albien d'Iran. Les occurrences d'Heteraster et d'autres spatangues en Iran permettent de distinguer clairement des associations d'espèces typiques des marges sud et nord de la Téthys au cours du Crétacé inférieur.
Spatangoida ;
Toxasteridae ;
Heteraster ;
Formation Dariyan ;
Formation Gadvan ;
Zagros ;
systématique.
Heteraster Orbigny, 1853, is a widespread Cretaceous genus of toxasterid echinoids reported to occur at the northern and southern margins of Tethys (western and southern Europe, North Africa and the Middle East), the Americas from the USA to Chile, and Japan. Heteraster originated during the Hauterivian, with its earliest occurrence in Portugal, and went extinct during the Early Cenomanian, with its latest occurrences in Europe and the USA. Defined by the alternation of two types of pore pairs in ambulacrum III, Heteraster constitutes a monophyletic group, with roots in the Toxaster lineage (Devriès, 1960; Rey, 1972; Villier et al., 2004). Heteraster reaches its maximum geographical range and its maximum species diversity during the Aptian-Albian. Its evolutionary success is linked to adaptation of the ambulacral structure to enhanced gaseous exchange (Villier et al., 2001).
Gauthier (1902) first reported Heteraster the Zagros region of Iran. Recent investigations resulted in additional records in the Kopeh Dagh Basin, northwest Iran (Taherpour Khalilabad et al., 2011) and in the Kerman area of central Iran (Vaziri & Arab, 2011). This present work provides descriptions and taxonomic assignments of Heteraster species found in sections of the High Zagros (southwest Iran), and includes a discussion of their relationships with other Iranian occurrences.
The fossil
echinoids were collected from the Dariyan Formation at Kuh-e Banesh in the south
of Iran, 80 km north of Shiraz at 52°25'57" longitude and 30°8'59"
latitude, and the Gadvan Formation at Kuh-e Gadvan, 50 km northeast of Shiraz at
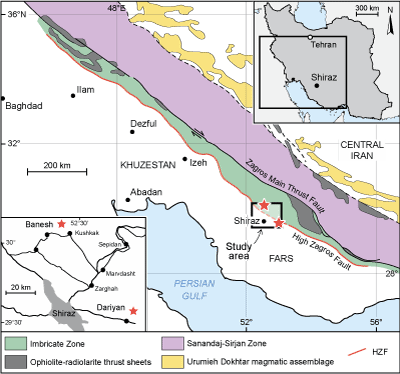
52°56'14.6" longitude and 29°36'53.8" latitude (Fig. 1 ![]() ). Structurally,
the localities belong to the High Zagros (Fig. 1
). Structurally,
the localities belong to the High Zagros (Fig. 1 ![]() ), which is delineated by the
High Zagros fault to the southwest and by the main Zagros thrust fault to the
northeast (Alavi, 2004, 2007). The structural unit is known as the
imbricated belt (Sepehr & Cosgrove,
2004) or the High Zagros
Basin (Berberian, 1995). The High Zagros belongs to the southern Iranian
blocks. During the Cretaceous, the High Zagros formed the northeast margin of
the Arabian plate, along the southern margin of the Neotethys Ocean (Stampfli
& Borel, 2002).
), which is delineated by the
High Zagros fault to the southwest and by the main Zagros thrust fault to the
northeast (Alavi, 2004, 2007). The structural unit is known as the
imbricated belt (Sepehr & Cosgrove,
2004) or the High Zagros
Basin (Berberian, 1995). The High Zagros belongs to the southern Iranian
blocks. During the Cretaceous, the High Zagros formed the northeast margin of
the Arabian plate, along the southern margin of the Neotethys Ocean (Stampfli
& Borel, 2002).
|
Figure 1:
Structure of the Zagros basin (modified after Alavi, 2007; Sepehr
& Cosgrove, 2004), and location of study area. |
In the High
Zagros, Lower Cretaceous deposits include the Barremian Gadvan Formation
overlain by the Aptian to Early Albian Dariyan Formation and the early/middle
Albian Kazhdumi Formation (Buchem et
al., 2010). At Kuh-e Gadvan, the Gadvan Formation is 95 m thick and consists
of marl with intercalated argillaceous limestone
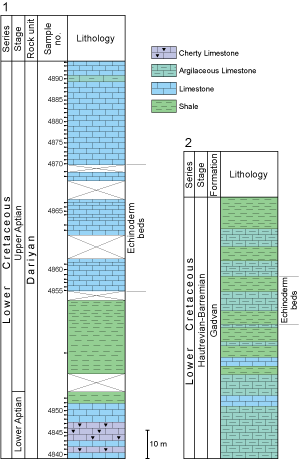
(Fig. 2.B ![]() ). Echinoids were
collected from the middle part of the Gadvan section. The Dariyan Formation is
275 m thick and can be divided into three units, the lower, middle and upper
Dariyan. The lower Dariyan consists of thick- to medium-bedded limestone with
benthic foraminifers and bivalves. The middle Dariyan is made of medium-bedded
limestone with black chert beds, black shale with
radiolarian and ammonite casts, and marl with planktonic foraminifera
(Fig. 2.A
). Echinoids were
collected from the middle part of the Gadvan section. The Dariyan Formation is
275 m thick and can be divided into three units, the lower, middle and upper
Dariyan. The lower Dariyan consists of thick- to medium-bedded limestone with
benthic foraminifers and bivalves. The middle Dariyan is made of medium-bedded
limestone with black chert beds, black shale with
radiolarian and ammonite casts, and marl with planktonic foraminifera
(Fig. 2.A ![]() ).
Carbon isotopes of the middle Dariyan display the characteristic features of
Oceanic Anoxic Event (OAE) 1a (Moosavizadeh et
al., 2014). The upper Dariyan includes two parts. The lower part consists of
buff to grey medium- to thick-bedded limestone with abundant orbitolinids and
echinoids, and local occurrences of iron oxide on bedding surfaces (Kheradpir,
1975)
(Fig. 2
).
Carbon isotopes of the middle Dariyan display the characteristic features of
Oceanic Anoxic Event (OAE) 1a (Moosavizadeh et
al., 2014). The upper Dariyan includes two parts. The lower part consists of
buff to grey medium- to thick-bedded limestone with abundant orbitolinids and
echinoids, and local occurrences of iron oxide on bedding surfaces (Kheradpir,
1975)
(Fig. 2 ![]() ). The upper part comprises thick to massive limestone with
abundant orbitolinids, bivalves and gastropods. The uppermost limestone layer of
the Dariyan Formation is intensively bioturbated which could indicate a
hardground surface (Mojab, 1974). The age of the Dariyan Formation at the
type section was established as early Aptian to middle Albian based on
foraminifera observed in thin sections (James & Wynd,
1965).
). The upper part comprises thick to massive limestone with
abundant orbitolinids, bivalves and gastropods. The uppermost limestone layer of
the Dariyan Formation is intensively bioturbated which could indicate a
hardground surface (Mojab, 1974). The age of the Dariyan Formation at the
type section was established as early Aptian to middle Albian based on
foraminifera observed in thin sections (James & Wynd,
1965).
|
Figure
2: Stratigraphic columns of sections studied
showing echinoid distribution. 1 Kuh-e Banesh. 2 Kuh-e Gadvan. |
Twenty-one specimens of toxasterid echinoids were collected at Kuh-e Banesh from the Dariyan Formation and at Kuh-e Gadvan from the Gadvan Formation. Some specimens were distorted and imperfect so that only 13 specimens could be measured. All specimens were photographed in adoral, aboral, lateral, frontal and posterior views. The arrangement of pore pairs in the ambulacra and the situation of apical disc were observed, photographed and drawn under a stereomicroscope. Fifteen rock samples were taken from echinoderm beds from which 52 thin sections were prepared in order to study the benthic foraminifera. All specimens are deposited in the Exploration Directorate, National Iranian Oil Company.
Description of the new material found in Iran justifies a thorough reappraisal of the identified species. Heteraster is a diverse genus that includes 87 nominal species, of which no comprehensive taxonomic revision has been published to date. Emended diagnoses are proposed for the two reported Iranian taxa following the results of Villier (2001). Without a phylogenetic framework for Heteraster species that would allow recognition of their apomorphic traits, the proposed species diagnoses list all descriptive aspects that could be of use for differentiation from other Heteraster species. The character states listed in the diagnoses refer to the type specimens, and consider the species' morphological variability. Abundant comparative material was assessed in European institutions: Aix-Marseille Université (Marseille, France), Museu Geológico (Lisbon, Portugal), Museum für Naturkunde (Berlin, Germany), Natural History Museum (London, United Kingdom), Muséum National d'Histoire Naturelle (Paris, France), Université de Bourgogne (Dijon, France), Université Claude-Bernard Lyon 1 (Villeurbanne, France), Université de Paul Sabatier (Toulouse, France), Université Pierre & Marie Curie (Paris, France), Université de Rennes 1 (Rennes, France) and Université de Poitiers (Poitiers, France).
The variability of quantitative traits is gauged from linear parameters measured with a Vernier caliper on several specimens per species from the Iranian material:
L: test length; W: test width ; H: test height; Lap: length of apical disc; Wap: width of apical disc; LI: length of petal I or V; LII: length of petal II or IV; LIII: length of ambulacrum III up to ambitus; Lpc: length of periproct; Wpc: width of periproct; Lpr: length of peristome; Wpr: width of peristome; W/L: relative width/elongation of test; H/L: relative height of test; LI/LII: relative length of posterior and anterior petals.
The echinoid classification follows Kroh & Smith (2010).
Class Echinoidea Leske, 1778
Cohort Irregularia Latreille, 1825
Superorder Atelostomata Zittel, 1879
Order Spatangoida Claus, 1876
Family stem group Toxasteridae Lambert, 1920
Genus Heteraster Orbigny, 1853
Type species. Spatangus oblongus Brongniart, 1821, p. 555, by subsequent designation of Loriol (1884: 622).
Synonymy. The genus Heteraster embraces the nominal genera Enallaster Orbigny, 1853, Washitaster Lambert, 1927a, and Paraheteraster Nisiyama, 1968 (Villier, 2001; Markov & Solovjev, 2002; Villier et al., 2004).
Diagnosis. Test low, rectangular, triangular or rounded in profile. Apical disc is ethmophract. The two columns of the anterior paired petals are always differentiated with reduced, small pore pairs in the anterior column and large asymmetrical pore pairs in the posterior column. Two types of pore pairs alternate in ambulacrum III. The pores simply vary in length or differ more markedly in bearing small isopores or anisopores and enlarged, strongly asymmetrical anisopores. The plates of the frontal ambulacrum are not aligned in a regular row, with the smaller pore pairs associated with reduced triangular plates. Plastron plates are elongated, with the two plates varying from asymmetrical to symmetrical. Parafascioles are expressed in various conditions from simple alignments of granules to large bands, or discontinuous narrow bands on the test flanks.
Occurrence. Heteraster ranges from the Hauterivian to the Cenomanian of the Tethyan margins (Middle East, North Africa and Europe), the Americas and Japan.
Remarks. Heteraster is easily distinguished from any other toxasterid echinoids by displaying an alternation of two forms of pore pairs in the unpaired, frontal ambulacrum (isopores or small diplopores, and elongate diplopores), and the differentiation of pore-pair types in the anterior and posterior column of the paired anterior ambulacra. Macraster transiens (Devriès, 1956) is a species initially described as Heteraster due to an alternation of pore-pair types in the frontal ambulacrum. However, symmetry, concavity and elongation of the paired ambulacra, as well as elongation of the labral plate lead to classification of the species with Macraster (Villier et al., 2004). Differentiation between the posterior and anterior columns of paired petals occurs in other toxasterids and represents a plesiomorphic character in the Spatangoida. It is also observed outside of the Spatangoida, in Hemipneustidae and a few other Holasteroida.
Heteraster delgadoi (Loriol, 1884)
(Figs. 3 ![]() - 4
- 4 ![]() - 5
- 5 ![]() -
6
-
6 ![]() - 7
- 7 ![]() - 8
- 8 ![]() ;
Table 1)
;
Table 1)
Synonymy, and abridged citation list.
Enallaster delgadoi Loriol, 1884: 619, Pl. 34, figs. 1-4. Loriol, 1887: 377-378, Pl. 16, figs. 4-5. Loriol, 1888: 87, Pl. 16, figs. 1-4. Cotteau, 1893: 357-358. Loriol, 1897: 165-166. Lambert, 1902: 19-20, Pl. 1, fig. 16. Fourtau, 1912: 51. Fourtau, 1921: 79. Blanckenhorn, 1926: 98, Pl. 8, fig. 25. Devriès, 1955: 316.
Heteraster delgadoi Lambert, 1922: 16. Lambert & Thiéry, 1924: 438. Lambert, 1927b: 46-47. Lambert, 1935: 365. Keller & Vautrin, 1937: 160-161. Devriès, 1960: 73. Mouty et al., 2003: 432.
non Heteraster cf. delgadoi Taherpour Khalilabad et al., 2011: 83-94, Pl. 2, fig. 2; Pl. 4, fig. 2.
Heteraster tissoti Villalba-Curras, 1993: 363-365, Pl. 26, figs. 8-10.
Heteraster cf. greenowi Villalba-Curras, 1993: 340-341.
Type material. The specimen figured by Loriol (1888: Pl. 16, fig. 1) is stored in the collections of the Geological Survey of Portugal, Lisbon. It comes from Albian marl of Varzea, Sintra area, Portugal.
Material. Seven measurable specimens from Kuh-e Banesh section (HDBAp.A-G), Dariyan Formation.
Diagnosis. Test heartshaped, with a marked angular frontal groove. Test profile elevated, ovoid, angular. Posterior truncation of the test subvertical, usually slightly concave. Apical disc eccentric, shifted toward the posterior part when the test increases in size. Unpaired frontal ambulacrum of "Enallaster" type with alternation of one or two pairs of anisopores with one pair of isopores. Small isopores hosted by reduced triangular plates. Inner pore rounded, with a deeply incised neural canal. External pore ovate to rectangular, and transversally transversely elongate. Small isopores with erect partition between the two pores. Anisopores with rounded to ovate internal pore and enlarged slitlike external pore, pores being separated by a wide, narrow partition. Anterior paired petals flexuous near apical disc becoming straight and parallel-sided, open distally. Columns strongly asymmetrical either in shape or number of pore pairs, with about half as many pores in anterior column. Isopores of anterior column rounded or punctiform. Anisopores of posterior column with rounded to ovate internal pore and elongate slit-like external pore. Posterior petals short, ovate to lanceolate and symmetrical, with similar pore pairs in anterior and the posterior columns. Apical system of ethmophract type, with madreporite partially or totally separating posterior genital plates. Periproct ovate, elongate in the horizontal plane and often angular at the base. Peristome large, pentagonal, slightly oriented toward anterior, with straight anterior margin. Parafasciole developed on test flanks. Paired anterior petals bordered by large tubercles with enlarged scrobicular area, often recessed. Labral plate triangular. Sternal plates short.
Description of material.
The test of the Iranian specimens is slightly longer than wide and high (Figs. 3 ![]() - 4
- 4 ![]() ). Test shape is ovate to slightly heart-shape in ambital outline with
shallow frontal sulcus. Posterior face truncated. In lateral view test has a
rounded anterior face, close to vertical posterior face and flat base. Apical
disc is ethmophract with four genital pores
(Fig. 5
). Test shape is ovate to slightly heart-shape in ambital outline with
shallow frontal sulcus. Posterior face truncated. In lateral view test has a
rounded anterior face, close to vertical posterior face and flat base. Apical
disc is ethmophract with four genital pores
(Fig. 5 ![]() )
and is located slightly
posterior of test center.
)
and is located slightly
posterior of test center.
|
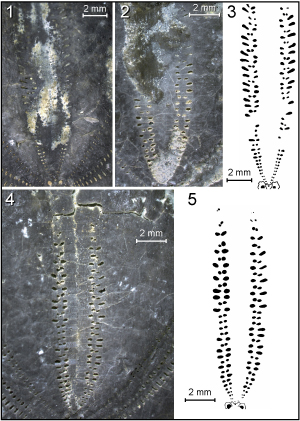
Figure
3: Heteraster
delgadoi. Specimen HDBAp.B: 1 - Aboral view. 2 - Oral view. 3 - Lateral
view. 4 - Anterior view. Specimen HDBAp.A: 5 - Aboral view. 6 - Oral view. 7 -
Lateral view. |
|
Figure
4: Heteraster delgadoi.
Specimen HDBAp.G: 1 - Aboral view. 2 - Oral view. 3 - Anterior view. 4 - Lateral
view. 5 - Posterior view. Specimen HDBAp.C: 6 - Aboral view. 7 - Oral view. 8 -
Anterior view. 9 - Lateral view. |
|
Figure
5:
Heteraster delgadoi.
Specimen HDBAp.F: 1 - Apical disc (photograph). 2 - Camera lucida drawing of
apical disc. |
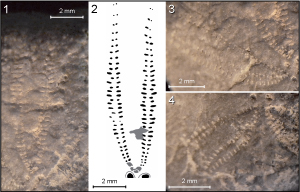
Frontal
ambulacrum is recessed, and situated in shallow anterior sulcus with two
alternating types of pore pairs, namely, wide isopores and anisopores with
welldeveloped slit-like external pore, especially in middle part of ambulacrum (Fig.
6 ![]() ). Alternation of pore pairs is quite regular with one small isopore for
two anisopores (or isopores with elongated pores at juvenile stage).
). Alternation of pore pairs is quite regular with one small isopore for
two anisopores (or isopores with elongated pores at juvenile stage).
Paired
ambulacra petaloid, large and flush with test surface. Anterior paired petals
slightly flexed close to apical disc and open distally (Fig. 7 ![]() ). Posterior
column of anterior petals bears elongate, slit-like, and conjugate anisopores,
outer series of pore pairs being wider than inner ones (Fig.
7.3-4
). Posterior
column of anterior petals bears elongate, slit-like, and conjugate anisopores,
outer series of pore pairs being wider than inner ones (Fig.
7.3-4 ![]() ). Anterior
column of pore pair has small and simple isopores. In some specimens alternating
small and more elongate pore pairs occur along medial and terminal parts of
anterior columns of anterior paired petals (Fig. 7.1-2
). Anterior
column of pore pair has small and simple isopores. In some specimens alternating
small and more elongate pore pairs occur along medial and terminal parts of
anterior columns of anterior paired petals (Fig. 7.1-2 ![]() ). Anterior paired petals
extend about three quarters of distance to ambitus.
). Anterior paired petals
extend about three quarters of distance to ambitus.
Posterior
petals are short (about two thirds length of anterior petal) and straight to
arcuate, and taper distally. pore pairs in both columns of petals are slightly
elongate and conjugate and outer series of pore pairs are slightly more elongate
than inner ones (Fig. 8 ![]() ).
).
|
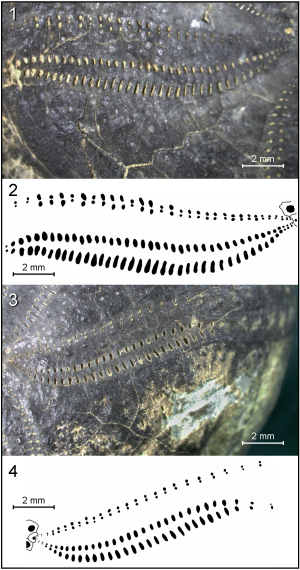
Figure
6: Heteraster
delgadoi. 1 - Frontal ambulacrum of specimen HDBAp.B. 2 - Frontal ambulacrum
of specimen HDBAp.A. 3 - Camera lucida drawing of frontal ambulacrum of specimen
HDBAp.B. 4 - Frontal ambulacrum of specimen HDBAp.F. 5 - Camera lucida drawing
of frontal ambulacrum of specimen HDBAp.F. |
|
Figure
7: Heteraster
delgadoi. Specimen HDBAp.B: 1 - Petal IV of specimen, showing two different
types of pore pairs in anterior and posterior columns, and alternation between
short and elongate pore pairs along medial and terminal parts of the anterior
columns. 2 - Camera lucida drawing of arrangement of pore pairs along petal IV.
Specimen HDBAp.G: 3 - Petal II. 4 - Camera lucida drawing of pore pairs
arrangement along petal II. |
|
Figure
8: Heteraster
delgadoi. Specimen HDBAp.F: Petal V. Pore pairs in posterior and anterior
columns are similar in size and shape. Outer series of pore pairs are slit-like
and longer than inner ones. |
Peristome is located near anterior margin, sub-pentagonal, relatively small, recessed, and without any surrounding rim. Periproct is small and visible in apical face, longitudinally ovate, and located at top of steeply sloping posterior face. Preservation of test surface precludes observation of possible fascioles. Anterior interambulacra develop coarse, enlarged tubercles, with depressed bare base between petals and frontal groove.
Table 1: Quantitative data showing variation in the test shape in Heteraster delgadoi.
| Collection number | L | W | H | W/L | H/L | LI | LII | LIII | LI/ LII | Lpr | Wpr | Lpc | Wpc | Lap | Wap |
| HDBAp.A | - | 29.1 | 17.1 | - | - | 8.6 | 12.6 | 16.6 | 0.68 | - | - | - | - | 1.9 | 2.2 |
| HDBAp.B | - | 31.7 | 17.4 | - | - | 10.6 | 16.4 | 17.8 | 0.64 | - | - | - | - | 2.2 | 2.3 |
| HDBAp.C | 32.2 | - | 17.1 | - | 0.53 | 9.3 | 13.5 | 14.2 | 0.69 | - | - | - | - | 2.3 | 2.4 |
| HDBAp.D | - | 35.8 | 19.2 | - | - | 12.2 | 18.8 | 19.2 | 0.65 | 4.4 | 4.9 | - | - | 2.4 | 2.6 |
| HDBAp.E | - | 27.3 | 17.9 | - | - | 9.4 | 14.9 | 14.1 | 0.64 | 3.6 | 3.8 | - | - | 2.1 | 2.2 |
| HDBAp.F | 27.9 | 25.8 | - | 0.92 | - | 9.3 | 13.6 | 14.8 | 0.68 | - | - | - | - | 1.8 | 1.9 |
| HDBAp.G | 26.5 | 25.3 | 15.1 | 0.95 | 0.57 | 7.2 | 11.8 | 10.5 | 0.61 | - | - | 3.95 | 3.3 | 2.3 | 2.4 |
Remarks. H. delgadoi has strong affinities with H. texanus (Roemer, 1852) and some other American species. However, these later species exhibit a narrower labrum and more elongate sternal plates, and usually do not develop enlarged scrobiculated tubercles in the anterior part of the test as found in H. delgadoi (Villier et al., 2004, fig. 2D). Heteraster delgadoi is similar to the Mediterranean species H. pomeli Ficheur, 1900, H. solignaci Lambert, 1931, H. tissoti Coquand, 1862, and H. zircensis Szörényi, 1955, but they differ in the test shape, the symmetry of the paired petals and the position of the apical disc. H. pomeli has a much more inflated dome-shaped test, a more central apical disc and slightly sinuous paired petals. H. solignaci is a species with a small, low test, and with less petaloid posterior paired petals. H. tissoti has a large, flattened test with the apical disc situated far to the posterior. Among species of the Mediterranean region, H. zircensis is the closest species to H. delgadoi, from which it differs in the more central position of the apical disc, and the wider angle of the posterior petals.
Heteraster delgadoi seems relatively common in the Dariyan Formation of the Kuh-e Banesh section, where they are likely of Aptian age. H. delgadoi is also reported in Aptian strata in Portugal (Rey, 1972) and Spain (Villalba-Curras, 1993). However, the occurrences of H. delgadoi are usually associated in the literature with the Knemiceras beds, Knemiceras being an ammonite genus of Albian age. A Cenomanian age was proposed in early twentieth-century papers for some Knemiceras beds that are nowadays regarded as late Albian (Vraconian). Fourtau (1912, 1921) mentioned H. delgadoi in the Albian of Djebel el Mistan (Egypt) without illustrating specimens, but the description matches the definition of the species, and this occurrence can be considered valid.
Taherpour Khalilabad et al. (2011) report Heteraster cf. delgadoi from the Kopet-Dagh mountains (northern Iran). The ambulacra of the illustrated specimen (Taherpour Khalilabad et al., 2011: Fig. 2a-c) do not match the patterns encountered in H. delgadoi. In this latter paper, almost all illustrated specimens of Toxaster spp. and Heteraster spp. likely belong to Heteraster renngarteni Poretzkaja, 1961.
Occurrences. Aptian to Albian of western and southern margins of the Tethys: Iberian Peninsula (Spain, Portugal), Levant and Middle East (Egypt, Israel, Lebanon, Syria, Iran).
Heteraster couloni (Agassiz, 1839)
Synonymy, and abridged citation list.
Holaster couloni Agassiz, 1839: 22-23, Pl. 4, figs. 9-10.
Toxaster couloni Agassiz & Desor, 1847: 132. Orbigny, 1850: 88. Renevier, 1853: 11. Ogérien & Michalet, 1867: 545.
Heteraster couloni Orbigny, 1855: 179-181, Pl. 848. Pictet, 1857: 195. Desor, 1858: 356. Dujardin & Hupé, 1862: 595. Ooster, 1865: 105. Lory, 1864: 314. Jaccard, 1869: 144. Loriol, 1873: 357-358, Pl. 29, fig. 9. Lambert & Thiéry, 1924: 438. Lambert & Jeannet, 1928: 190. Denizot, 1934: 142. Devriès, 1955: 316. Rey, 1972: 481-482, Fig. 154, Pl. 20, figs. 12-14. Zaghbib-Turki, 1987: 327-331, Fig. 110, Pl. 15, fig. 1. Melikov, 1989: 156.
Enallaster couloni Loriol, 1884: 624. Révil, 1887: 22. Savin, 1903: 33-34. Savin, 1905: 23-24.
Toxaster sabugensis Loriol, 1888: 92, Pl. 17, figs. 1-2. Lambert & Thiéry, 1924: 436. Devriès, 1956: 261.
Heteraster aff. couloni Lees, 1928: 643-644, Pl. 44, fig. 5. Hudson & Chatton, 1959. Rey, 1972: 484-485, Fig. 155, Pl. 22, figs. 1-2. Granier, 2008.
Heteraster sabugensis Devriès, 1960: 78. Rey, 1972: 465-468, Figs. 145-146; Pl. 20, figs. 4-6.
Type material. Casts of the type specimen of Holaster couloni are housed in the Musée de Neuchâtel under the collection number Syn. p. 356 (Lambert & Jeannet, 1928). The types were collected from Barremian limestone at Mormont, Switzerland. Plaster casts of the type specimen are available in several institutions (e.g., MNHN, UCBL).
Material. Six measurable specimens from Kueh-e Gadvan section (HCGAp.1-6), Gadvan Formation, beds numbers 4555-4560.
Diagnosis. Test large, ovate. Posterior side of test rounded, vertical to slightly overhanging. Aboral surface inflated, with interambulacrum 5 forming a slight carina. Anterior groove regularly concave, forming a regularly rounded notch at the ambitus. Test profile ovate to rectangular. Frontal ambulacrum bearing anisopores of varying size. Large anisopores with an ovate internal pore and an elongate, slit-like outer pore. Small anisopores with an internal rounded pore, without clearly differentiated neural canal, and a rectangular elongate external pore, both pores separated by a well-developed interporal partition. The plates bearing the small anisopores are usually short often occluded. Alternating pattern of pore-pair types with two to three pairs of large pores for one pair of small pores. Anterior paired petals large, wide, slightly flexuous with an open sub-petaloid termination. The anterior petal column exhibits anisopores made of a rounded internal pore and an elongate external pore, sometimes alternating with a few isopores. Posterior column has strongly asymmetrical anisopores with a rounded to oval internal pore and a long slit-like external pore. Interporal partition narrow, flat. Posterior paired petals relatively long, forming a wide angle. Ambulacral columns of posterior petals open distally, subpetaloid, asymmetrical, anterior one being narrow, straight, and posterior one being wide and flexuous. Pore pairs expressed as in anterior petals. Apical disc eccentric, displaced posteriorly. Apical disc ethmophract, relatively elongate with ocular plate IV often inserted in contact with madreporite. Periproct rounded to slightly transversally elongate. Peristome sub-pentagonal, transverse, with narrow labrum margin. Sternal plates asymmetrical. Ornament of upper test surface made of small primary tubercles. No fasciole, with the exception of local granule concentrations and alignments on lateral flanks.
Description of material.
Test elongate, cordate, with regularly curved lateral flanks and broad, shallow
anterior groove (Fig. 9 ![]() ). Profile ovate and weakly truncated posteriorly. Apical
disc lies in posterior part of test. Apical disc ethmophract with enlarged
genital 2 (madreporite), three remaining genital plates being of similar size
(Fig. 10
). Profile ovate and weakly truncated posteriorly. Apical
disc lies in posterior part of test. Apical disc ethmophract with enlarged
genital 2 (madreporite), three remaining genital plates being of similar size
(Fig. 10 ![]() ). Ocular plate IV located close to or in contact with genital 2.
Frontal ambulacrum is a wide, shallow sulcus forming a rounded notch at anterior
margin (Fig. 9.1-2
). Ocular plate IV located close to or in contact with genital 2.
Frontal ambulacrum is a wide, shallow sulcus forming a rounded notch at anterior
margin (Fig. 9.1-2 ![]() ). Pore pairs of two types, small isopores and wide,
slit-like anisopores (Fig. 11.1-2
). Pore pairs of two types, small isopores and wide,
slit-like anisopores (Fig. 11.1-2 ![]() ). Anisopores with particularly elongate
external pore in middle part of ambulacrum. Pore-pair arrangement irregular with
one isopore alternating with two or three anisopores. Pore pairs progressively
smaller and less differentiated closer to the apical disc, of elongate isopore
type. All paired ambulacra long, petaloid and flush with test surface (Fig.
9.1, 9.6, 9.9
). Anisopores with particularly elongate
external pore in middle part of ambulacrum. Pore-pair arrangement irregular with
one isopore alternating with two or three anisopores. Pore pairs progressively
smaller and less differentiated closer to the apical disc, of elongate isopore
type. All paired ambulacra long, petaloid and flush with test surface (Fig.
9.1, 9.6, 9.9 ![]() ). All ambulacra have a wide interporiferous area. Anterior paired
petals extend about 3/4 of distance between apex and ambitus; relatively flexed
posteriorly close to apical disc, then straight toward ambitus and open distally
(Fig. 10.3
). All ambulacra have a wide interporiferous area. Anterior paired
petals extend about 3/4 of distance between apex and ambitus; relatively flexed
posteriorly close to apical disc, then straight toward ambitus and open distally
(Fig. 10.3 ![]() ).
Posterior column of anterior petals bears elongate, slit-like and
conjugate anisopores, outer pores wider than inner ones. Anterior column of pore
pair has small, less elongate isopores. Posterior paired petals are short, about
two thirds of length of anterior paired petals. Anterior pore column is
straight, posterior column slightly flexed, especially close to apical disc
(Fig. 10
).
Posterior column of anterior petals bears elongate, slit-like and
conjugate anisopores, outer pores wider than inner ones. Anterior column of pore
pair has small, less elongate isopores. Posterior paired petals are short, about
two thirds of length of anterior paired petals. Anterior pore column is
straight, posterior column slightly flexed, especially close to apical disc
(Fig. 10 ![]() ). In posterior pore column of petals, outer pores slit-like, inner
pores smaller and elongate. Pore pairs of anterior columns of petals are also
anispores, but with narrower interporal partition. Peristome pentagonal, sunken
and lacking rim; located near anterior test margin (Fig. 9.5
). In posterior pore column of petals, outer pores slit-like, inner
pores smaller and elongate. Pore pairs of anterior columns of petals are also
anispores, but with narrower interporal partition. Peristome pentagonal, sunken
and lacking rim; located near anterior test margin (Fig. 9.5 ![]() ). Periproct small,
visible in apical face, transversally ovate, and located at top of weakly
truncated posterior face (Fig. 9.8
). Periproct small,
visible in apical face, transversally ovate, and located at top of weakly
truncated posterior face (Fig. 9.8 ![]() ). No fasciole present. Plastron plating not
seen in specimens to hand (Fig. 9.2, 9.7
). No fasciole present. Plastron plating not
seen in specimens to hand (Fig. 9.2, 9.7 ![]() ). Tuberculation includes fine and
uniformly small primary tubercles.
). Tuberculation includes fine and
uniformly small primary tubercles.
|
Figure
9: Heteraster
couloni. Specimen HCGAp.C: 1 - Aboral view. 2 - Oral view. 3 - Posterior
view. 4 - Lateral view. 5 - Posterior and oral view. 6 - Oral view. 7 - Aboral
view. 8 - Posterior view. 9 - Lateral view. |
|
Figure
10: Heteraster couloni.
Specimen HCGAp.C: 1 Apical disc (photograph). 2 - Camera lucida drawing of
apical disc. |
|
Figure
11: Heteraster
couloni. Specimen HCGAp.C: 1 - Frontal ambulacrum. 2 - Camera lucida of
frontal ambulacrum. 3 - Petal IV, specimen. 4 - petal V. |
Remarks. Several nominal species are rather similar to H. couloni: H. renngarteni Poretzkaja, 1961, H. heckeri Melikov, 1989, H. magnus Poretzkaja, 1961, H. lepidus (Loriol, 1888), H. oblongus (Brongniart, 1821), and H. renevieri (Desor, 1858). H. couloni can be distinguished from the Barremian species H. lepidus by usually larger size, its deeper frontal groove, and the structure of its frontal ambulacra that exhibit smaller and less differentiated anisopores. H. couloni differs from H. oblongus by lesser posteriorward projection of the apical disc, and its higher test profile, less flexuous anterior petals, proportionally longer posterior petals and usually less elongate ambital shape (Loriol, 1873; Rey, 1972). H. sabugensis (Loriol, 1888) is considered a synonym of H. couloni, although specimens usually have a lower test (Villier, 2001). The diagnostic characters of H. sabugensis recognized by Rey (1972) are a slightly truncated posterior margin, an inflated test in its posterior part, and the lack of occluded plates in the frontal ambulacra for the smaller type of pore pairs. All characters cited so far fall within the variability of H. couloni, especially when compared with the oldest populations of the Jura Mountains in eastern France. From the late Hauterivian to the late Barremian of western Europe, there is an obvious trend in the evolution of H. couloni marked by a progressive differentiation of pore pairs in the frontal ambulacrum, a widening of the posterior test, a relative elongation of the lateral interambulacra, and a slight migration of the peristome toward a more central position. H. renevieri is grossly similar to the most inflated variant of H. couloni but is smaller and never develops a notch at the ambitus.
H. renngarteni, H. magnus and H. heckeri form a morphologically homogeneous group of nominal species that should be considered synonymous, H. renngarteni being the senior name (Villier, 2001). Compared to H. couloni, H. renngarteni has a less eccentrically placed apical disc, a lesser development of the anterior petals compared to the posterior ones, and narrower anterior petals.
Maran (1996) describes a fauna from the Barremian of eastern Serbia in which several species are listed (H. couloni, H. lepidus, H. crisminensis (Loriol, 1888), and H. constrictus (Fourtau, 1921)), but it seems that all illustrated specimens belong to H. couloni. Lees (1928: 643-644, Pl. 44, fig. 5) describes four specimens from Oman, assigned to H. aff. couloni. The material is fragmentary which precludes a definitive identification, but an assignment to H. couloni is retained here. Most recent populations of H. couloni found in Europe (especially in southeast France) often correspond to inflated specimens that were incorrectly attributed to H. renevieri by some authors (Démoly, 1928; Savin, 1905).
Table 2: Quantitative data showing variation in test shape in Heteraster couloni.
| Collection number | L | W | H | W/L | H/L | LI | LII | LIII | LI/ LII | Lpr | Wpr | Lpc | Wpc | Lap | Wap |
| HCGAp.a | 26.7 | 23.6 | 10.8 | 0.88 | 0.40 | 6.9 | 10.6 | 11.3 | 0.65 | 3.1 | 3.2 | 2.7 | 3.2 | - | - |
| HCGAp.b | 38.6 | 32.4 | 15.5 | 0.84 | 0.40 | 11.4 | 16.5 | 19.2 | 0.69 | 4.1 | 4.7 | - | - | 3.1 | 2.7 |
| HCGAp.c | 17.1 | 14.9 | 8.2 | 0.87 | 0.48 | 5.3 | 7.9 | 8.2 | 0.67 | 1.8 | 1.8 | - | - | 1.4 | 1.2 |
| HCGAp.d | 19.8 | 17.4 | 9.8 | 0.89 | 0.49 | - | 8.9 | - | - | 2.3 | 2.5 | 2.2 | - | - | - |
| HCGAp.e | 24.1 | 21.8 | 11.6 | 0.9 | 0.48 | 6.1 | 9.3 | 11.2 | 0.66 | 2.6 | 3.0 | - | - | - | - |
| HCGAp.f | 16.8 | 14.6 | 8.6 | 0.87 | 0.51 | - | - | - | - | 2.4 | 2.7 | 1.5 | 1.8 | - | - |
Occurrence. In the studied section, this species is found in the Barremian marly limestone of the Gadvan Formation. H. couloni is reported as common in Europe (Portugal, Spain, France, Switzerland, Serbia), with sparser occurrences along the southern margin of the Tethys (Algeria, Tunisia, Oman, Zagros). Its age ranges from late Hauterivian to late Barremian.
H. couloni is a widespread species in Europe, with sparse occurrences in northern Africa. The new data from Gadvan confirm the presence of H. couloni in Iran during the Barremian, which was only tentatively suggested by former authors. The species is reported from other Middle Eastern countries, including United Arab Emirates (Hudson & Chatton, 1959) and Oman (Granier, 2008). Among the species of Heteraster, H. couloni has one of the longest stratigraphic ranges (Hauterivian to Barremian) and greatest geographical extent (Portugal westward to Iran eastward). It can be understood as a ubiquitous species of the southern Tethys margin during the Barremian.
Heteraster musandamensis Lees, 1928, is a species from the early Aptian of the Middle East, morphologically close to H. oblongus with which it was often confused. It is identified from the Sarmord Formation in northern Iraq as Heteraster oblongus var. musandamensis by Dunnington et al. (1959) and as Heteraster oblongus from the Kazhdumi Formation in Zagros by Jones (2012). A redefinition of the species and a check for further occurrences is appropriate, but Heteraster musandamensis seems to be endemic to the Middle Eastern part of the southern Tethys margin during the Aptian.
H. delgadoi is here recognized unambiguously for the first time in Iran. The species is well known from the Levant area (Egypt, Israel, Lebanon, Syria), and from southern Europe. H. delgadoi was distributed throughout the southern margin of the Tethys during the late Aptian-Albian, with the Iranian material being the easternmost occurrences of the species.
Macraster Roemer, 1888, is a highly diversified genus from the Albian of Iran that shows a biogeographic distribution similar to that of Heteraster. Macraster is reported only from the Zagros area. Two species, Macraster longesulcatus (Cotteau & Gauthier, 1895) and Macraster obtritus Lambert, 1931, are known elsewhere on the southern margin (North Africa) and the western Tethys (Europe) (Kamyabi Shadan et al., 2014).
Several echinoid species are mentioned by Collignon (1981) from the Albian strata of the Kazhdumi Formation. The fauna seems to be highly endemic, with potential relationships with the fauna of Malagasy. However, the toxasterid echinoid taxa of the Zagros area is in need of comprehensive review.
H. renngarteni occurs in the Aptian of the Tirgan Formation in central (Tabas block) and northern Iran (Kopeh-Dagh). The species was described initially from the south Caucasus (Turkmenistan and Azerbaijan) from shallow water carbonate deposits of Barremian age. Based on facies homogeneity and large foraminifer assemblages, Prosorovsky (1990) demonstrated that the south Caucasian area, including Kopeh-Dagh, is biogeographically uniform. After the closure of the Paleothethys, the blocks forming the Central-East-Iranian microcontinent, that includes the Tabas Block, aggregated to Laurasia (and the Caucasus) during the Late Triassic (Mattei et al., 2015). Despite the temporary opening of some deep-marine basins during the Jurassic and the Early Cretaceous, continuous shallow-water shelves were likely to have existed between the Caucasus and central Iran, as witnessed by the development of the Tirgan Formation in both regions. Thus, H. renngarteni may be an endemic species of the northern Tethys margin.
The paleobiogeographical relationship of the Iranian spatangoid fauna during the Early Cretaceous becomes clearer as historical material is revised and new occurrences are reported. A fauna typical of the southern Tethyan margin emerged during the Barremian to Albian interval that includes cosmopolitan species (H. couloni, H. delgadoi, Macraster longesulcatus, Macraster obtritus) and a few endemic species of the Cretaceous margins of the Arabian plate (Heteraster musandamensis). The spatangoid fauna of the northern Tethyan margin seems less diverse, with H. renngarteni the sole species encountered in northern and central Iran during the Barremian. Connections with the south Caspian area (Turkmenistan, Azerbaijan) exist but relationships with other parts of the Tethyan margins remain unclear.
The authors thank Alexandre Lethiers for is help with figure preparation, and the reviewers for their helpful comments on an earlier version of the paper.
Agassiz L. (1839).- Description des échinodermes fossiles de la Suisse.- Nouveaux Mémoires de la Société Helvétique des Sciences naturelles, Genève, vol. 3, 101 p.
Agassiz L. & Desor E. (1847).- Catalogue raisonné des familles, des genres et des espèces de la classe des échinodermes.- Annales des Sciences Naturelles, Paris, vol. 3, 167 p.
Alavi M. (2004).- Regional stratigraphy of the Zagros fold-thrust belt of Iran and its proforeland evolution.- American Journal of Science, New Haven, CT, vol. 304, no. 1, p. 1-20.
Alavi M. (2007).- Structures of the Zagros fold-thrust belt in Iran.- American Journal of Science, New Haven, CT, vol. 307, no. 9, p. 1064-1095.
Berberian M. (1995).- Master "blind" thrust faults hidden under the Zagros folds; active basement tectonics and surface morphotectonics.- Tectonophysics, vol. 241, p. 193-224.
Blanckenhorn M. (1926).- Die Seeigelfauna der Kreide Palästinas.- Palaeontographica - Beitrage zur Naturgeschichte der Vorzeit, Cassel, vol. 67, 113 p.
Brongniart A. (1821).- Sur les caractères zoologiques des formations, avec application de ces caractères à la détermination de quelques terrains de Craie.- Annales des Mines, Paris, (Série 1), vol. 6, tome 4, p. 537-572.
Buchem F.S.P. van, Baghbani D., Bulot L.G., Caron M., Gaumet F., Hosseini S.A., Keyvani F., Schroeder R., Swennen R., Vedrenne V. & Vincent B. (2010).- Barremian-lower Albian sequence-stratigraphy of southwest Iran (Gadvan, Dariyan and Kazhdumi formations) and its comparison with Oman, Qatar and the United Arab Emirates.- GeoArabia Special Publication, Manama, no. 4, p. 503-548.
Claus C.F.W. (1876).- Grundzüge der Zoologie.- N.G. Elwert, Marburg and Leipzig, 3rd edition, 1254 p.
Collignon M. (1981).- Faunes Albo-Cénomaniennes de la Formation des marnes de Kazhdumi, région du Fars-Khuzestan.- Documents du Laboratoire de Géologie de Lyon, Villeurbanne, Hors-Série 6, p. 251-291.
Coquand H. (1862).- Géologie et paléontologie de la région sud de la province de Constantine.- Mémoires de la Société d'Émulation de la Provence, Marseille, 274 p.
Cotteau G. (1893).- Sur quelques espèces d'échinides au Liban.- Comptes rendus du Congrès de l'Association française pour l'Avancement des Sciences, Paris, 22e session, p. 346-360.
Cotteau G.H. & Gauthier V. (1895).- Paléontologie. In: Études géologiques. Mission scientifique en Perse par J. de Morgan.- E. Leroux Éditeur, Paris, tome IIIe, partie II, 107 p.
Démoly M. (1928).- Note de M. Jules Lambert sur les échinides de la Savoie et de l'Isère communiqués par M. Démoly.- Bulletin de la Société d'Histoire naturelle de Savoie, Chambéry, (2e série), tome 21, p. 139-153.
Denizot G. (1934).- Description des massifs de Marseilleveyre et de Puget.- Annales du Muséum d'Histoire naturelle de Marseille, Tome XXVI, 237 p.
Desor E. (1855-58).- Synopsis des échinides fossiles.- Ch. Reynwald, Paris ; Kreidel & Niedner, Wiesbaden, 490 p.
Devriès A. (1955).- Note sur le genre Heteraster.- Bulletin de la Société géologique de France, Paris, (6e série), tome 5e, p. 315-323.
Devriès A. (1956).- Sur une nouvelle espèce d'échinide en Algérie : Enallaster transiens, Pomel.- Publications du Service de la Carte géologique d'Algérie, Alger, vol. 8, p. 251-267.
Devriès A. (1960).- Contribution à l'étude de quelques groupes d'échinides fossiles d'Algérie.- Publications du Service de la Carte géologique d'Algérie, Alger, vol. 3, p. 1-279.
Dujardin F. & Hupé H. (1862).- Suites à Buffon. Histoire naturelle des Zoophytes Échinodermes.- Librairie Encyclopédie de Roret, Paris, 628 p.
Dunnington H.V., Wetzel R. & Morton D.M. (1959).- Iraq, Mesozoic and Palaeozoic.- Lexique stratigraphique international, Paris, Asie, vol. 3(10a), 333 p.; In: Granier B. (ed.), Electronic version.- Lexique stratigraphique international (nouvelle version), Comité Français de Stratigraphie, Paris. URL: http://paleopolis.rediris.es/LEXICON/IRAQ/
Ficheur E. (1900).- Note sur quelques échinides nouveaux de l'Aptien d'Algérie.- Bulletin de la Société géologique de France, Paris, vol. 28, p. 590-596.
Fourtau R. (1912).- Contribution à l'étude des échinides fossiles de la Syrie.- Mémoires présentés et lus à l'Institut égyptien, Le Caire, tome VII, fasc. II, p. 41-68.
Fourtau R. (1921).- Catalogue des invertébrés fossiles de l'Égypte représentés dans les collections du Musée de Géologie au Caire. Terrains crétacés. Troisième partie : Échinodermes (Supplément).- Geological Survey of Egypt, Palaeontological Series, Cairo, vol. 5, p. 1-101.
Gauthier M.V. (1902).- Études géologiques. Partie III. Échinides. Supplément.- Mission scientifique en Perse, Paris, tome III, p. 109-190 (Pls. XVII-XXIV). URL: https://www.rechercheisidore.fr/search/resource/?uri=10670/1.36g29g
Granier B.R.C. (2008).- Holostratigraphy of the Kahmah regional series in Oman, Qatar, and the United Arab Emirates.- Carnets Geol., Madrid, vol. 8, no. A07 (CG2008_A07), p. 1-34.
Hudson R.G.S. & Chatton M. (1959).- The Musandam Limestone (Jurassic to Lower Cretaceous) of Oman, Arabia.- Notes et Mémoires sur le Moyen-Orient, Paris, vol. 3, p. 69-93.
Jaccard A. (1869).- Jura vaudois et neuchâtelois.- Matériaux pour la carte géologique de la Suisse, Berne, vol. 6, 340 p.
James G.A. & Wynd J.G. (1965).- Stratigraphic nomenclature of Iranian Oil Consortium Agreement Area.- American Association of Petroleum Geologists, Bulletin, Tulsa, vol. 49, no. 12, p. 2182-2245.
Jones R.W. (2012).- Applied Palaeontology.- Cambridge University Press, 452 p.
Kamyabi Shadan H., Villier L., Sadeghi A. & Adabi M.H. (2014).- A revision of the Macraster species (Echinodermata, Echinoidea) occurring in the Albian deposits of the Zagros basin, Southwest Iran.- Annales de Paléontologie, Paris, vol. 100, p. 51-62.
Keller A. & Vautrin H. (1937).- Nouvelle contribution à l'étude des échinides de la Syrie et du Liban.- Notes et Mémoires du Haut-Commissariat de la République française en Syrie et au Liban, Beyrouth, vol. 2, p. 137-164.
Kheradpir A. (1975, unpublished).- Stratigraphy of the Khami Group in southwest Iran.- Iranian Oil Operating Companies, Geological and Exploration Division, report no. 1235, 67 p.
Kroh A. & Smith A.B. 2010.- Classification and phylogeny of post-Palaeozoic echinoids.- Journal of Systematic Palaeontology, London, vol. 7, p. 147-212.
Lambert J. (1902).- Description des échinides fossiles de la province de Barcelone. Ière partie.- Mémoires de la Société géologique de France, tome IX, no. 24, p. 1-57 (Pls. IX-XII). URL: http://jubilotheque.upmc.fr/ead.html?id=GM_000924_001
Lambert J. (1920).- Sur quelques genres nouveaux d'échinides.- Mémoires de la Société académique d'Agriculture, des Sciences, Arts et Belles-Lettres du Département de l'Aube, Troyes, tome LXXIV, p. 145-172 (Pls. I-II).
Lambert J. (1922).- Échinides fossiles de la province de Santander.- Trabajos del Museo Nacional de Ciencias Naturales, Madrid, (Serie Geologíca), vol. 28, p. 7-26.
Lambert J. (1927a).- Considérations sur les échinides de la Comanche Série du Texas.- Bulletin de la Société géologique de France, Paris, (4e série), tome XXVI, p. 263-278.
Lambert J. (1927b).- Révision des échinides fossiles de Catalogne.- Memorias del Museo de Ciencias Naturales de Barcelona, (serie geológica), vol. 1, no. 1, p. 1-102 (Pls. I-IV).
Lambert J. (1931).- Étude sur les échinides fossiles du Nord de l'Afrique.- Mémoires de la Société géologique de France, Paris, (nouvelle série), no. 16, p. 5-108.
Lambert J. (1935).- Échinides crétacés d'Espagne.- Boletin de la Sociedad Española de Historia Natural, Madrid, vol. 35, p. 513-526.
Lambert J. & Jeannet A. (1928).- Nouveau catalogue des moules d'échinides fossiles du Musée d'Histoire naturelle de Neuchâtel exécutés sous la direction de L. Agassiz & E. Desor.- Mémoires de la Société Helvétique des Sciences Naturelles, Zürich, vol. 64, no. 2, p. 79-233.
Lambert J. & Thiéry P. (1924).- Essai de nomenclature raisonnée des échinides.- Librairie L. Ferrière, Chaumont, fasc. 6-7, p. 385-512.
Latreille P.A. (1825).- Familles naturelles du règne animal.- Baillière, Paris, 570 p.
Lees G.M. (1928).- The geology and tectonics of the Oman and of parts of south-eastern Arabia.- Quarterly Journal of the Geological Society of London, vol. 84, p. 585-670.
Leske N.G. (1778).- Jacobi Theodori Klein Naturalis dispositio Echinodermatum.- Officiana Gleditschiana, Lipsis, 278 p.
Loriol P. de (1873).- Échinologie helvétique. Description des oursins fossiles de la Suisse. Deuxième partie : Échinides de la période crétacée.- Matériaux pour la Paléontologie Suisse ou Recueil de Monographies sur les Fossiles du Jura et des Alpes, Genève, vol. 6, 398 p.
Loriol P. de (1884).- Notes pour servir à l'étude des échinodermes I.- Recueil Zoologique Suisse, Genève, vol. 1, p. 605-643.
Loriol P. de (1887).- Notes pour servir à l'étude des échinodermes. II.- Recueil Zoologique Suisse, Genève, vol. 4, no. 3, p. 365-407.
Loriol P. de (1888).- Recueil d'études paléontologiques sur la faune crétacique du Portugal. Volume II. Description des échinodermes.- Communication des Travaux géologiques du Portugal, Lisbonne, vol. 7, no. 2, 122 p.
Loriol P. de (1897).- Notes pour servir à l'étude des échinodermes. VI.- Revue Suisse de Zoologie et annales du Muséum d'Histoire naturelle de Genève, vol. 5, p.141-178.
Lory C. (1864).- Description géologique du Dauphiné (Isère, Drôme et Hautes Alpes).- F. Savy, Paris, 748 p.
Maran A.A. (1996, unpublished).- Echinoidea from the Urgonian sediments of eastern Serbia (regular and irregular sea urchins, classification, stratigraphy, paleoecology).- Master's Thesis, Faculty of Mining and Geology, Belgrade, 77 p.
Markov A.V. & Solovjev A.N. (2002).- Echinoids of the genus Washitaster (Spatangoida): Morphology and taxonomic position.- Paleontological Journal, Moscow, vol. 16, no. 4, p. 368-379.
Mattei M., Cifelli F., Muttoni G. & Rashid H. (2015).- Post-Cimmerian (JurassicCenozoic) paleogeography and vertical axis tectonic rotations of central Iran and the Alborz Mountains.- Journal of Asian Earth Sciences, Hong Kong, vol. 102, p. 92-101.
Melikov O.H. (1989).- Morphological peculiarity of the new species of the genus Heteraster d'Orbigny, 1853 (Echinoidea) from the Urgonian facies of Azerbaijan. In: Kalo D.L., Rojnov C.W., Solovjev A.N., Stukalina G.A. & Xints L.M. (eds.), Fossil and recent echinoderm researches.- Academy of Sciences of the Estonian SSR, Tallinn, p. 156-160 (in Russian).
Mojab F. (1974).- Some Cretaceous (Albian) Invertebrates from Banesh, Southern Iran.- Bulletin of the Indian Geologists' Association, Chandigarh, vol. 7, no. 1, p. 13-34.
Moosavizadeh M.A., Mahboubi A., Moussavi-Harami R. & Kavoosi M.A. (2014).- Early Aptian oceanic anoxic event (OAE 1a) in northeastern Arabian plate setting: An example from Dariyan Formation in Zagros fold-trust belt, SE Iran.- Arabian Journal of Geosciences, Riyadh, vol. 7, p. 4745-4756.
Mouty M., Al-Maleh A.K. & Laban H.A. (2003).- Le Crétacé moyen de la chaîne des Palmyrides (Syrie centrale).- Geodiversitas, Paris, vol. 25, no. 3, p. 429-443.
Nisiyama S. (1968).- The echinoid fauna from Japan and adjacent regions part II.- Palaeontological Society of Japan Special Papers, Tokyo, no. 13, p. 1-491 (30 Pls.). URL: http://www.palaeo-soc-japan.jp/download/SP/SP13.pdf
Ogérien F. & Michalet M.E. (1867).- Histoire naturelle du Jura et des départements limitrophes.- Masson, Paris, 570 p.
Ooster W.-A. (1865).- Synopsis des échinodermes fossiles des Alpes Suisses.- Librairie H. Georg, Genève, 131 p.
Orbigny A. d' (1850).- Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- Masson, Paris, vol. 2, 428 p.
Orbigny A. d' (1853-1855).- Échinides crétacés. In: Paléontologie Française. Description zoologique et géologique de tous les animaux mollusques et rayonnés fossiles de France.- Masson, Paris, vol. 6, 596 p.
Pictet F.-J. (1857).- Traité élémentaire de paléontologie, ou, Histoire naturelle des animaux fossiles considérés dans leurs rapports zoologiques et géologiques.- 2e édition, J.B. Baillière et Fils, Paris, 768 p.
Poretzkaja E.S. (1961).- Heteraster du Néocomien du Turkménistan occidental.- Problema neftegazonosti Sredneĭ Azii, Leningrad, vypusk 51, p. 157-175.
Prosorovsky V.A. (1990).- The Urgonian facies of central Asia.- Cretaceous Research, vol. 11, p. 253-260.
Renevier E. (1853).- Note sur le terrain néocomien qui borde le pied du Jura, de Neuchâtel à La Sarraz.- Bulletin de la Société vaudoise de Sciences naturelles, Lausanne, vol. 3, 13 p. URL: https://doi.org/10.3931/e-rara-29747
Révil J. (1887).- Description géologique de la Montagne du Corbelet.- Revue savoisienne, Annecy, 23 p.
Rey J. (1972).- Recherches géologiques sur le Crétacé inférieur de l'Estremadura (Portugal).- Mémoires du Service géologique du Portugal, Lisbonne, vol. 21, 477 p.
Roemer F. (1852).- Die Kreidebildungen von Texas und ihre organischen Einschlüsse.- Adolf Marcus, Bonn, 100 p.
Roemer F. (1888).- Macraster, eine neue Spatangoiden-Gattung aus der Kriede vonTexas.- Neues Jahrbuch für Mineralogie, Geologie und Palöntologie, Stuttgart, Band 5, p. 191195.
Savin L. (1903).- Catalogue raisonné des échinides fossiles du département de la Savoie.- Bulletin de la Société d'Histoire naturelle de Savoie, Chambéry, vol. 2, no. 8, p. 109-315.
Savin L. (1905).- Révision des échinides du département de l'Isère.- Travaux du Laboratoire de Géologie de l'Université de Grenoble, vol. 7, no. 2, p. 152-363.
Sepehr M. & Cosgrove J.W. (2004).- Structural framework of the Zagros Fold-Thrust Belt, Iran.- Marine and Petroleum Geology, vol. 21, p. 829-843.
Stampfli G. & Borel G.D. (2002).- A plate tectonic model for the Paleozoic and Mesozoic constrained by dynamic plate boundaries and restored synthetic oceanic isochrons.- Earth and Planetary Science Letters, vol. 196, p. 17-33.
Szörényi E. (1955).- Échinides crétacés de la Bakony.- Geologica Hungarica, Budapest, (Series Palaeontologica), fasc. 26, 332 p.
Taherpour Khalilabad M., Aryaei A.A., Ashouri A.R. & Ghaderi A. (2011).- Introducing some echinoderms from the Tirgan Formation, Kopeh-Dagh Basin, NE of Iran.- Geopersia, Tehran, vol. 1, no. 1, p. 83-94.
Vaziri M.R. & Arab A.L. (2011).- Morphological variations, patterns of frontal ambulacrum pores and paleoecology of Heteraster renngarteni Poretzkaja (Echinoidea: Spatangoida) from Aptian Sediments of Baghin Area, Kerman, Iran.- Pakistan Journal of Scientific and Industrial Research, Karachi, (Series B: Biological Science), vol. 54, no. 1, p. 41-46.
Villalba-Curras M.P. (1993).- Revisión de los equínoideos del Cretácico inferior y medio español.- Universidad Complutense de Madrid, 568 p.
Villier L. (2001, unpublished).- Évolution du genre Heteraster dans le contexte de la radiation de l'ordre des Spatangoida (Echinoidea, Echinodermata) au Crétacé inférieur.- Thèse de l'Université de Bourgogne, Dijon, 389 p.
Villier L., David B. & Néraudeau D. (2001).- Ontogenetic and morphological evolution of the ambulacral pores in Heteraster (early spatangoids). In: Barker M. (ed.), Echinoderm 2000.- Balkema, Rotterdam, p. 563-567.
Villier L., Néraudeau D., Clavel B., Neumann C. & David B. (2004).- Phylogeny of early Cretaceous spatangoids (Echinoidea: Echinodermata) and taxonomic implications.- Palaeontology, vol. 47, no. 2, p. 265-292.
Zaghbib-Turki D. (1987, unpublished).- Les échinides du Crétacé de Tunisie. Paléontologie générale : Systématique, paléoécologie, paléobiogéographie.- Thèse de la Faculté des Sciences de Tunis, 609 p.
Zittel K. von (1879).- Handbuch der Palaeontologie.- S.R. Oldenbourg, München, 765 p.