◄ Carnets Geol. 16 (27) ►
![]()
Contents
[1. Introduction]
[2. Geological background and material]
[3. Results]
[4. Discussion]
[5. Conclusions and suggestions for further studies]
and ...
[Bibliographic references]
Institute of Geological Sciences, Jagiellonian University, Oleandry 2a, 30-063 Kraków (Poland)
Department of Geology, Paleontology and Fossil Fuels, Sofia University "St. Kliment Ohridski", 15 Tzar Osvoboditel Bd., 1000 Sofia (Bulgaria)
Department of Geology, Paleontology and Fossil Fuels, Sofia University "St. Kliment Ohridski", 15 Tzar Osvoboditel Bd., 1000 Sofia (Bulgaria)
Institute of Geological Sciences, Jagiellonian University, Oleandry 2a, 30-063 Kraków (Poland)
Published online in final form (pdf) on December 31, 2016
[Editor:
Robert W. Scott; technical editor: Bruno Granier]
![]()
Euendolithic microorganisms (boring endoliths) syn-vivo associated with modern corals are commonly reported, but their fossil record is extremely rare. This paper reports the new finding recognized in the colonial scleractinian coral Clausastrea saltensis from the Upper Barremian of Bulgaria. Large microborings (up to 50 µm, most ca. 15-25 µm in diameter) filled with calcite cement are distributed medially along coral septa of some corallites. Borings were produced by microeuendoliths growing from the skeleton interior outward during the life of the coral host. They are compared to traces produced by the recent oligophotic filamentous chlorophyte Ostreobium, which is known to be the most common skeleton-dwelling alga in modern living corals and regarded as neutral or beneficial to the coral. In terms of general morphology, diameter and distribution pattern, the borings are similar to those recently recognized in the Early Cretaceous microsolenid coral.
microborings;
euendoliths;
symbiosis;
corals;
Barremian;
Bulgaria.
Kołodziej B., Idakieva V., Ivanov M. & Salamon K. (2016).- New record of endolithic algae syn-vivo associated with an Early Cretaceous coral.- Carnets Geol., Madrid, vol. 16, no. 27, p. 633-640.
Découverte nouvelle d'algues endolithes affectant un corail d'âge Crétacé inférieur de son vivant.- Des microorganismes euendolithiques (endolithes perforants) sont souvent signalés affectant des coraux modernes de leur vivant, alors qu'a contrario ils sont extrêmement rares dans le registre fossile. Cette publication fait état de leur présence documentée récemment dans un corail scléractiniaire colonial, Clausastrea saltensis, du Barrémien supérieur de Bulgarie. Des perforations relativement larges (jusqu'à 50 µm de diamètre, dans la plupart des cas de l'ordre de 15 à 25 µm) colmatées par un ciment calcitique sont réparties selon le plan médian des cloisons coralliennes de quelques corallites. Ces perforations sont dues à des euendolithes microscopiques qui se développaient du vivant de leur hôte corallien depuis l'intérieur de son squelette vers l'extérieur. Elles sont comparées aux traces laissées par le chlorophyte filamenteux oligophotique Ostreobium, une algue actuelle vivant au coeur du squelette de coraux modernes, en fait la plus fréquemment observée, et dont l'impact sur ces derniers est considéré comme neutre, voire comme bénéfique. En raison de leur forme générale, de leur diamètre et de leur modèle de répartition, ces perforations sont similaires à celles récemment identifiées dans le corail microsolénidé d'âge Crétacé inférieur.
microperforations ;
euendolithes ;
symbiose ;
coraux ;
Barrémien ;
Bulgarie.
Eukaryotic organisms may be animal or plant holobionts that are associated with microbial communities. The term coral holobiont refers to a dynamic, complex association between a coral host and internal and external microbiota: dinoflagellates (zooxanthellae), other algae, bacteria, archaea, viruses, fungi, and protozoans living in the coral tissue, mucus microlayer and skeleton (Rohwer et al., 2002). Recently, substantial progress has been made in studies of coral-associated microorganisms because interactions between them are crucial for coral health and disease, hence for the functioning of coral reefs (e.g., Rosenberg & Loya, 2004; Thompson et al., 2015). Zlatarski (2010) drew attention to the concept of coral holobionts in a paper addressed to researchers of fossil corals.
Geochemical and biochemical criteria, and fine-scale growth banding of the skeleton hold potential to identify symbiotic zooxanthellae of fossil corals (Frankowiak et al., 2016, and references therein), but to reveal these features requires exceptionally well preserved skeletons. Among coral-associated microorganisms, euendolithic algae, cyanobacteria and fungi are endoliths that actively penetrate hard substrates (Golubic et al., 1981). These endoliths leave specific boring traces with fossilization potential even in poorly preserved skeletons. However, the fossil record of such coral-endolith associations are extremely rare, even though traces of boring microorganisms that colonized skeletons of live corals are as old as the Ordovician (Elias & Lee, 1993). The best preserved microendoliths are those described by Kołodziej et al. (2012a) from the Lower Cretaceous (Aptian) microsolenid corals from Romania. These authors also discussed other scarce fossil records. This paper reports large microborings produced by endoliths associated with a coral from the Barremian of Bulgaria.
During the Barremian-Early Aptian (Early Cretaceous) several carbonate platforms existed on the northern Tethyan margin in Bulgaria (Minkovska et al., 2002). Abundant and diversified corals (about 120 species) were described from marls and limestones of the Lovech Urgonian Group in the Central Fore-Balkan. Level-bottom assemblages and coral bioconstructions (mostly matrix-supported reefs) represent clear- and turbid-water environments (e.g., Zlatarski, 1967; Idakieva & Ivanov, 2002; Idakieva, 2008; Kołodziej et al., 2012b).
The studied specimen comes
from the Upper Barremian coral-bearing
marls of the Smochan
Formation (Idakieva
& Ivanov, 2002).
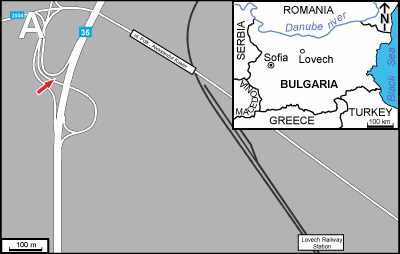
Marls crop out along the road in the northern part of Lovech (ca. 140 km ENE of
Sofia), ca. 1 km NW from the Lovech Railway Station (Fig. 1 ![]() ). Ten transverse and longitudinal uncovered thin sections of standard size
(25 mm x 35 mm) were prepared from a colony of Clausastrea
saltensis Alloiteau,
1960, in which
microendoliths were recognized.
). Ten transverse and longitudinal uncovered thin sections of standard size
(25 mm x 35 mm) were prepared from a colony of Clausastrea
saltensis Alloiteau,
1960, in which
microendoliths were recognized.
Thin sections were analyzed using binocular microscope Nikon SMZ 1000, petrographic microscope Zeiss Axiolab and scanning electron microscope Hitachi S-4700. Additionally, thin sections were studied using cold cathodoluminescence microscopy, apparatus Cambridge Image Technology Ltd, 8200 Mk3. All observations were carried out at the Institute of Geological Sciences, Jagiellonian University. Microboring diameters were measured using the Palaeontological Database System PaleoTax, module PaleoTax/Measure (http://www.paleotax.de/measure). Measurements were statistically analysed with the PAST software (Hammer et al., 2001). The specimen and thin sections are housed in the Institute of Geological Sciences, Jagiellonian University, Kraków.
|
|
|
Figure 1:
A) Location of sampling site. B)
The outcrop in Lovech showing coral-bearing marls of
the Smochan Formation. |
The
microeuendoliths, further referred to as microendoliths or microborings, were
recognized in septa of the thamnasterioid (corallites
without intervening wall) scleractinian
coral, Clausastrea
saltensis Alloiteau,
1960, of the Family Montlivaltiidae Dietrich, 1926,
Suborder
Astreoina Alloiteau, 1952. They occur in three of the ten thin sections made from
different parts of the colony. Microendoliths are common in septa in transverse
thin section Lov 1c (Figs. 2 ![]() & 4
& 4 ![]() ). Longitudinal sections of borings are rare (Lov 1b;
Fig. 3
). Longitudinal sections of borings are rare (Lov 1b;
Fig. 3 ![]() ). Microborings are absent in
dissepiments, that is tabular or vesicular skeletal elements connecting
neighbouring septa.
). Microborings are absent in
dissepiments, that is tabular or vesicular skeletal elements connecting
neighbouring septa.
The aragonitic coral
skeleton was neomorphically
transformed to calcite and recrystallized (Fig. 2.A ![]() ). Septa are not micritized. Interseptal space
is filled with blocky calcite cement, only locally with fine-grained sediment (Fig.
2.E
). Septa are not micritized. Interseptal space
is filled with blocky calcite cement, only locally with fine-grained sediment (Fig.
2.E ![]() ). Calcite cement is lighter (more transparent)
than neomorphic calcite (pseudospar) forming the coral skeleton. Observations
under petrographic (Figs. 2
). Calcite cement is lighter (more transparent)
than neomorphic calcite (pseudospar) forming the coral skeleton. Observations
under petrographic (Figs. 2 ![]() - 3
- 3 ![]() ) and scanning electron microscope (Fig. 4
) and scanning electron microscope (Fig. 4 ![]() ) revealed that calcite crystals
filling microboring tunnels are coarser than neomorphic calcite of the coral
skeleton, which is reflected by different solubilities of both calcite types (Fig.
4.B
) revealed that calcite crystals
filling microboring tunnels are coarser than neomorphic calcite of the coral
skeleton, which is reflected by different solubilities of both calcite types (Fig.
4.B ![]() ). Only locally microborings are filled with micrite (Figs.
2.B-C
). Only locally microborings are filled with micrite (Figs.
2.B-C ![]() & 3.D
& 3.D ![]() ) and rarely with pyrite crystals (Fig.
4.B
) and rarely with pyrite crystals (Fig.
4.B ![]() ). Studies under
crossed nicols and
under cathodoluminescent microscope revealed neither new microborings nor
enhanced their morphology.
). Studies under
crossed nicols and
under cathodoluminescent microscope revealed neither new microborings nor
enhanced their morphology.
In transverse sections, microborings are distributed medially, along the
septa (Figs. 2.B-F ![]() & 4
& 4 ![]() ),
both in their outer and inner parts, which are close to the corallite centre (Fig.
2.E
),
both in their outer and inner parts, which are close to the corallite centre (Fig.
2.E ![]() ). Locally microborings are clearly clustered (Fig.
2.D
). Locally microborings are clearly clustered (Fig.
2.D ![]() ). A few microendoliths are cut
in the longitudinal direction evidencing that they are filamentous (Fig. 3
). A few microendoliths are cut
in the longitudinal direction evidencing that they are filamentous (Fig. 3 ![]() ). The presence of lateral branches is uncertain both in transverse (Fig.
2.F
). The presence of lateral branches is uncertain both in transverse (Fig.
2.F ![]() ) and longitudinal (Fig.
3.C-D
) and longitudinal (Fig.
3.C-D ![]() )
sections. Two filaments are segmented (Fig. 3.B-C
)
sections. Two filaments are segmented (Fig. 3.B-C ![]() ).
).
|
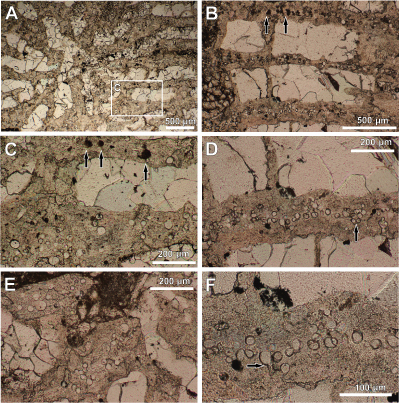
Figure
2: Transverse sections through Clausastrea
saltensis. A) General morphology of the corallite; Close-up of boxed area is
shown in image C. B-F) Septa with microendoliths. Microborings are filled mostly
with calcite sparite cement, rarely by micrite (arrow in B and C). In image D
microborings are clearly clustered (arrow); in E microendoliths occur in the
inner part (at the corallite centre) of coral septa; arrow in F shows two filaments squeezed in the same borehole, possibly above the
branch point. |
|
Figure
3: A-D) Longitudinal sections through Clausastrea
saltensis showing septa with microendoliths. A) Arrows indicate long algal
filament. B) Longitudinal (left white arrow) and transverse/oblique sections
(right white arrow). Black arrow shows segmented
filament; C) Segmented
filament (lower arrow) and the cross wall in the filament (upper arrow)
suggesting the onset of a narrower lateral branch. D) Two filaments filled with micrite
interpreted as possible lateral branching (arrow) or alternatively as closely
located filaments. |
|
|
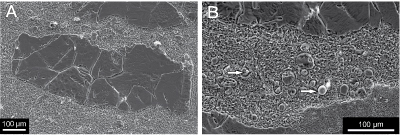
Figure
4: SEM
images of microendoliths in coral septum in the transverse section. Image B is
enlargement of the septum partially visible in the lower part of the image A.
Note differences in crystal sizes of neomorphic
calcite forming the coral skeleton and calcite cement filling microboring
tunnels. Arrows indicate pyrite crystals in microborings. |
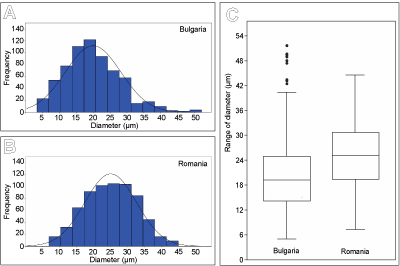
662 measurements of microboring diameters in transverse section were made
using the program PaleoTax/Measure. Microborings vary in diameter ranging from 5
to 52 µm, most of them are between ca. 15-25 µm. The histogram
showing the frequency distribution of diameters is
slightly right-skewed (Fig. 5.A & 5.C ![]() ). For comparison, 662 measurements of microborings in
the Lower Aptian microsolenid coral (the best preserved sample R31; Kołodziej
et al., 2012a) from Romania range from
7 to 45 µm in diameter, mostly between ca. 18-32 µm. The histogram is
normally distributed with a
flat top (Fig. 5.B-C
). For comparison, 662 measurements of microborings in
the Lower Aptian microsolenid coral (the best preserved sample R31; Kołodziej
et al., 2012a) from Romania range from
7 to 45 µm in diameter, mostly between ca. 18-32 µm. The histogram is
normally distributed with a
flat top (Fig. 5.B-C ![]() ).
).
|
Figure
5:
Frequency of microboring diameters. A-B) Histograms
showing frequency of the minimum and maximum diameters of microborings in Clausastrea
saltensis from
Bulgaria (A) and in the microsolenid coral from the Lower Aptian of Romania (B) described
by Kołodziej et al. (2012a). C) Boxplot depicting median and range of microboring
diameters in Bulgarian and Romanian corals. |
Distribution
patterns of microendoliths differ in live and dead corals. Dead
and denuded parts of the skeleton are colonized at the surface and bored inward.
Skeletons of live corals are bored from the inside upward and outward by
microorganisms living in close proximity to and below the coral tissue and keeping
pace with the rate of coral growth (Le Campion-Alsumard et
al., 1995). Filamentous morphology, diameter and
distribution patterns of microendoliths described here are very similar to those
revealed recently in
skeletons of microsolenid corals from the Lower Aptian of Romania (Kołodziej
et al., 2012a). The presence of
filament branching is uncertain because of very rare and poorly preserved
longitudinal sections of microendoliths in C.
saltensis. For example, possible branching shown in Figure
3.D ![]() may be
alternatively interpreted as a result of close position of two unbranched
filaments. Microborings
are distributed medially along coral septa
evidencing they are not post-mortem, but instead
microorganisms lived in the skeleton of a growing coral. It is worth underlining
that septa are not micritized in Bulgarian and Romanian corals with
microendoliths. Thus there is no evidence of post-mortem activity of boring
microorganisms, although micritization is a common phenomenon in many fossil
corals, including those from the Lower Cretaceous of Bulgaria (e.g., Kołodziej
et al., 2012b, Figs. 10C, 14E, 15D,
22A). Diameters of microendoliths
described here are similar to those recognized in Romanian corals. Histograms
of the frequency of microendolith diameters are
normally distributed and slightly right-skewed in the case of microendoliths in the Bulgarian coral. That
indicates the presence of one dominant morphotype (species?).
may be
alternatively interpreted as a result of close position of two unbranched
filaments. Microborings
are distributed medially along coral septa
evidencing they are not post-mortem, but instead
microorganisms lived in the skeleton of a growing coral. It is worth underlining
that septa are not micritized in Bulgarian and Romanian corals with
microendoliths. Thus there is no evidence of post-mortem activity of boring
microorganisms, although micritization is a common phenomenon in many fossil
corals, including those from the Lower Cretaceous of Bulgaria (e.g., Kołodziej
et al., 2012b, Figs. 10C, 14E, 15D,
22A). Diameters of microendoliths
described here are similar to those recognized in Romanian corals. Histograms
of the frequency of microendolith diameters are
normally distributed and slightly right-skewed in the case of microendoliths in the Bulgarian coral. That
indicates the presence of one dominant morphotype (species?).
The spectacularly well-preserved, iron-stained microendoliths from Romania allowed their more detailed analysis and comparison with the modern euendolithic siphonalean chlorophyte Ostreobium (Kołodziej et al., 2012a). It is worth noting that algal filaments in Romanian and Bulgarian corals have large diameters. Filaments of recent Ostreobium usually have smaller diameters. For example, Lukas (1974) described from living corals filaments of Ostreobium quekettii Bornet & Flahault, 1889, with diameters of 1-25 μm (typically 2-5 μm). In Ostreobium constrictum Lukas, 1974, two types of filaments occurred: cylindrical with diameter of 3-20 μm, and inflated ones with diameter of 13-60 μm. Ostreobium quekettii is a modern producer of the ichnospecies Ichnoreticulina elegans (Radtke, 1991). Its galleries have diameters of ca. 3 µm (see Tribollet, 2008), which are much less than microendoliths described here and than most of those described by Kołodziej et al. (2012a).
Although microendoliths are ubiquitous in the modern living corals, they are poorly taxonomically diversified and include algae (chlorophytes, rhodophytes), cyanobacteria and fungi (Le Campion-Alsumard et al., 1995; Golubic et al., 2005; Tribollet, 2008). Ostreobium quekettii is the most common autotrophic euendolith and was recognized in over 85% of coral species across a wide geographic and bathymetric range (Lukas, 1974; Le Campion-Alsumard et al., 1995; Tribollet, 2008; Gutner-Hoch & Fine, 2011).
Autotrophic and heterotrophic microorganisms that actively bore into rocks (mostly carbonates) might gain advantages in nutrient acquisition, finding a niche with limited competition, protection from some physical environmental stressors normally experienced by free-living algae, exploitation of organic matter, protection from predatory grazing, and desiccation. A skeleton of a living coral is an extreme habitat, yet some photosynthetic algae and cyanobacteria show extraordinary ability to cope with low-light conditions (Schlichter et al., 1997; Shashar et al., 1997; Golubic et al., 2005; Tribollet, 2008).
Microborers that colonize dead carbonate substrates are important primary producers and greatly contribute to bioerosion and sedimentation (Tribollet et al., 2011). The role of the euendolithic communities within skeletons of living corals for their health and diseases remains poorly understood (for review: Tribollet, 2008). Although, rapid blooms of algal endoliths may cause over 25% of skeletal carbonate to be dissolved and removed (Le Campion-Alsumard et al., 1995), most of the studies on endolithic algae from reef corals suggest a neutral or positive effect on coral health, while fungi are potentially pathogenic. Microendoliths can translocate fixed carbon to the coral tissue (e.g., Schlichter et al., 1997), and may be important for the survival of some bleached corals providing an alternative source of energy (Fine & Loya, 2002).
Although its exact character is difficult to determine, syn-vivo coral-microendolith associations described here can be termed as facultative ectosymbiotic relationships in a broad (and original) sense of the term symbiosis. The broad meaning of this term is accepted by many biologists (e.g., Paracer & Ahmadjian, 2000) and, especially by palaeontologists (e.g., Darrell & Taylor, 1993).
Diagenetic bias (early or late diagenesis) seems to be the main control of the poor fossil record of this ecological group, especially of these microendoliths that left very thin microborings. Aragonitic skeletons of scleractinians are commonly neomorphically transformed to calcite, recrystallized, or dissolved (moldic porosity is commonly later filled with calcite cement). However, as previously observed by Kołodziej et al. (2012a) in the Lower Cretaceous corals of Romania, microendoliths may be well preserved even in coral skeletons strongly affected by diagenesis. In Romanian corals the filaments show diverse states of preservation, which indicate that responsible taphonomic processes (mostly impregnation by iron oxides) operated on a small scale.
Alternatively, but less possible, the patchy distribution of microendoliths may be interpreted to be an increase in growth and diameter of microendoliths, in some parts of the colony, for instance during coral bleaching. That might favour preservation of large filaments.
This report provides rare evidence of fossil microendolith-coral association. Although they are not as well preserved as the Early Cretaceous microendoliths described by Kołodziej et al. (2012a), the new record confirms that some coral-associated microendoliths may be large enough to be examined optically with standard petrographic microscope. However, careful observations of corals are required, because microendoliths might be preserved only locally.
The study of fossil microendoliths that inhabited growing corals is still in its infancy. This type of symbiotic association was certainly much more common, but it is extremely sparse fossil record is likely due to taphonomic loss. Moreover, microborings may be overlooked during taxonomic studies, or may be confused with relics of coral skeletal microstructure. They may co-occur with post-mortem borings, which makes their identification more difficult. In particular, further research should be oriented towards revealing thin microendoliths (ca. 5 μm and smaller). This requires well preserved coral specimens and SEM observations of natural or resin casts (Kołodziej & Radtke, 1999; K. Salamon, work in progress) using casting-embedding, which is commonly used for modern and fossil material where microborings remained empty (Golubic et al., 1970). Corals associated with ferruginous sediments, like those from the Cenomanian of Saxony (Löser, 2014), are potentially perspective material for study of symbiotic microendoliths that may be preserved as natural casts (K. Salamon, work in progress). However, as shown by this report, large microendoliths may be readily visible under a petrographic microscope, even though they are not impregnated by other minerals.
Research focused on this aspect of coral palaeoecology will certainly reveal new findings. Such discoveries will be potentially significant to the understanding of palaeoecology of corals, microendoliths and their evolutionary relationships. Although much information will remain unrevealed due to taphonomic biases, new fossil records and rapidly improving understanding of modern coral-endolithic associations may provide valuable insight into the palaeoecology of ancient reefs. Potentially valuable research material includes corals showing skeletal bandings, which may be related to increased algal or fungal growth associated with environmental stress (e.g., Le Campion-Alsumard et al., 1995; Golubic et al., 2005; Carilli et al., 2010). Distribution of fine microborings in some Palaeozoic corals are compared by Risk et al. (1987) and Elias and Lee (1993) to algal banding in modern corals.
Lastly, due to the presence of early calcite-filled borings great care must be taken when even well preserved aragonitic corals are used as trace element archives applied to palaeoenvironmental studies. Proper sampling techniques are required to avoid calcite cement-filled microborings produced by symbiotic microendoliths in recent corals (Nothdurft & Webb, 2009).
The studies were conducted in the frame of the research grant financed by the Polish Ministry of Science and Higher Education (No. 2P04D 028 29). Dr. Vassil Zlatarski (Bristol, Rhode Island, US) participated actively during our field studies. Prof. Elizabeth Chacón (Linares, Nuevo Leon, Mexico) and Prof. Stejpko Golubic (Boston, US) are acknowledged for corrections and constructive critical comments that improved the manuscript. We are grateful to Prof. Robert W. Scott (Tulsa, US) for linguistic corrections.
Carilli J.E., Godfrey J., Norris R.D., Sandin S.A. & Smith J.E. (2010).- Periodic endolithic algal blooms in Montastraea faveolata corals may represent periods of low-level stress.- Bulletin of Marine Science, Miami, vol. 86, no. 3, p. 709-718.
Darrell J.G. & Taylor P.D. (1993).- Macrosymbiosis in corals: A review of fossil and potentially fossilizable examples.- Courier Forschunginstitut Senckenberg, Frankfurt am Main, vol. 164, p. 185-198.
Elias R.J. & Lee D.-J. (1993).- Microborings and growth in Late Ordovician halysitids and other corals.- Journal of Paleontology, Lawrence, vol. 67, no. 6, p. 922-934.
Fine M. & Loya Y. (2002).- Endolithic algae - An alternative source of photoassimilates during coral bleaching.- Proceedings of the Royal Society of London, vol. 269, no. 1497, p. 1205-1210.
Frankowiak K., Kret S., Mazur M., Meibom A., Kitahara M.V. & Stolarski J. (2016).- Fine-scale skeletal banding can distinguish symbiotic from asymbiotic species among modern and fossil scleractinian corals.- PLoS ONE, San Francisco, vol. 11, no. 1, e0147066.
Golubic S., Brent G. & Le Campion-Alsumard T. (1970).- Scanning electron microscopy of endolithic algae and fungi using a multipurpose casting-embedding technique.- Lethaia, Oslo, vol.3, no. 2, p. 203-209.
Golubic S., Friedmann I. & Schneider J. (1981).- The lithobiontic ecological niche, with special reference to microorganisms.- Journal of Sedimentary Petrology, Tulsa, vol. 51, no. 2, p. 475-478.
Golubic S., Radtke G. & Le Campion-Alsumard T. (2005).- Endolithic fungi in marine ecosystems.- Trends in Microbiology, London, vol. 13, no. 5, p. 229-235.
Gutner-Hoch E. & Fine M. (2011).- Genotypic diversity and distribution of Ostreobium quekettii within scleractinian corals.- Coral Reefs, New York, vol. 30, no. 3, p. 643-650.
Hammer Ø., Harper D.A.T. & Ryan P.D., 2001. Past: Paleontological statistics software package for education and data analysis.- Palaeontologia Electronica, vol. 4, no. 1, art. 4, 9 p. URL: http://folk.uio.no/ohammer/past/
Idakieva V. (2008).- Taxonomy of scleractinian corals from the Barremian-Lower Aptian of Central North Bulgaria (Lovech Urgonian Group).- Annuaire de l'Université de Sofia "St. Kliment Ohridski", Faculté de Géologie et Géographie, vol. 101, p. 5-32.
Idakieva V. & Ivanov M. (2002).- Coral buildups in the Smochan Formation (Urgonian, Lower Cretaceous) around Lovech (Central North Bulgaria).- Annuaire de l'Universite de Sofia "St. Kliment Ohridski", Faculte de Géologie et Géographie, Sofia, vol. 95, p. 31-45. [In Bulgarian, English abstract].
Kołodziej B., Golubic S., Bucur I.I., Radtke G. & Tribollet A. (2012a).- Early Cretaceous record of microboring organisms in skeletons of growing corals.- Lethaia, Oslo, vol. 45, no. 1, p. 34-45.
Kołodziej B., Ivanov M. & Idakieva V. (2012b).- Prolific development of pachythecaliines in Late Barremian, Bulgaria: Coral taxonomy and sedimentary environment.- Annales Societatis Geologorum Poloniae, Kraków, vol. 82, no. 4, p. 291-330.
Kołodziej B. & Radtke G. (1999).- Relation of coral microstructure and microendoliths attack (Paleocene). In: 8th International Symposium on Fossil Cnidaria and Porifera, Sendai.- Abstracts, p. 101.
Le Campion-Alsumard T., Golubic S. & Hutchins P. (1995).- Microbial endoliths in the skeletons of live and dead corals: Porites lobata (Moorea, French Polynesia).- Marine Ecology Progress Series, Oldendorf (Luhe), vol. 117, no. 1-3, p. 149-157.
Löser H. (2014).- Korallen.- Geologica Saxonica, Dresden, vol. 60, no. 1, p. 17-56.
Lukas K.J. (1974).- Two species of the chlorophyte genus Ostreobium from skeletons of Atlantic and Caribbean reef corals.- Journal of Phycology, Moss Landing, vol. 10, no. 3, p. 331-335.
Minkovska V., Peybernès B., Nikolov T. & Ivanov M. (2002).- Paleogeographic reconstruction of a segment of the North-Tethyan margin in Bulgaria from Barremian to Albian.- Eclogae Geologicae Helvetiae, Basel, vol. 95, no. 1, p. 183-195.
Nothdurft L.D. & Webb G.E. (2009).- Earliest diagenesis in scleractinian coral skeletons: Implications for palaeoclimate-sensitive geochemical archives.- Facies, Erlangen, vol. 55, no. 2, p. 1612-4820.
Paracer S. & Ahmadjian V. (2000).- Symbiosis. An introduction to biological associations.- Oxford University Press, New York, 291 p.
Risk M.J., Pagani S.E. & Elias R.J. (1987).- Another internal clock: Preliminary estimates of growth rates based on cycles of algal boring activity.- Palaios, Lawrence, vol. 2, no. 4, p. 323-331.
Rohwer F., Seguritan V., Azam F. & Knowlton N. (2002).- Diversity and distribution of coral-associated bacteria.- Marine Ecology Progress Series, Oldendorf (Luhe), vol. 243, p. 1-10.
Rosenberg E. & Loya Y. (eds., 2004).- Coral health and disease.- Springer-Verlag, Berlin Heidelberg. 488 p.
Schlichter D., Kampmann H. & Conrady S. (1997).- Trophic potential and photoecology of endolithic algae living within coral skeletons.- Marine Ecology, Berlin, vol. 18, no. 4, p. 299-317.
Shashar N., Banaszak A.T., Lesser M.P. & Amrami D. (1997).- Coral endolithic algae: Life in a protected environment.- Pacific Science, Honolulu, vol. 51, no. 2, p. 167-173.
Thompson J.R., Rivera H.E., Closek C.J. & Medina M. (2015).- Microbes in the coral holobiont: Partners through evolution, development, and ecological interactions.- Frontiers in Cellular and Infection Microbiology, Lausanne, vol. 4, Article 176, no. 4, p. 1-20.
Tribollet A. (2008).- The boring microflora in modern coral reef ecosystems: A review of its roles. In: Wisshak M. & Tapanila L. (eds), Current developments in bioerosion.- Springer-Verlag, Berlin, p. 67-94.
Tribollet A., Radtke G. & Golubic S. (2011).- Bioerosion. In: Reitner J. & Thiel V. (eds), Encyclopedia of geobiology.- Springer, Dordrecht, p. 117-134.
Zlatarski V. (1967).- Note sur le genre Clausastrea d'Orbigny (Ordre Madreporaria).- Bulletin of the Geolological Insitute, Sofia, (Series Paleontology), vol. 16, p. 23-34.
Zlatarski V. (2010).- Palaeobiological perspectives on variability and taxonomy of scleractinian corals.- Palaeoworld, Nanjing, vol. 19, no. 3-4, p. 333-339.