◄ Carnets Geol. 18 (2) ►
![]()
Outline:
[1. Whale barnacles ...]
[2. Fossil whale barnacles from Sicily ...]
[3. Geological and biostratigraphic framework]
[4. Systematic palaeontology]
[5. Palaeobiogeographical and palaeoecological implications] and ...
[Bibliographic references]
Corresponding author
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Museo Civico di Storia Naturale di Comiso, Via degli Studi 9, 97013 Comiso (Italy)
Museo Civico di Storia Naturale di Comiso, Via degli Studi 9, 97013 Comiso (Italy)
Istituto di Geoscienze e Georisorse, IGG-CNR, via G. Moruzzi 1, 56124 Pisa (Italy)
Zeeuwsch Genootschap der Wetenschappen, P.O. Box 378, 4330 AJ Middelburg (The Netherlands);
Koninklijk Belgisch Instituut voor Natuurwetenschappen, Operationele Directie Aarde en Geschiedenis van het Leven, Vautierstraat 29, 1000 Brussel (Belgium)
Vicolo Monteleone 4, 90133 Palermo (Italy)
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Published online in final form (pdf) on March 19, 2018
DOI 10.4267/2042/65747
[Editor: Bruno Granier;
language editor: Stephen Carey]
![]()
We report on three shells of whale barnacle (Cirripedia: Coronulidae) collected from Pleistocene shallow-marine deposits exposed at Cinisi (northwestern Sicily, southern Italy). These specimens are identified as belonging to the extinct species Coronula bifida Bronn, 1831. Calcareous nannoplankton analysis of the sediment hosting the coronulid remains places the time of deposition between 1.93 and 1.71 Ma (i.e., at the Gelasian-Calabrian transition), an interval during which another deposit rich in whale barnacles exposed in southeastern Apulia (southern Italy) formed. Since Coronula Lamarck, 1802, is currently found inhabiting the skin of humpback whales [Cetacea: Balaenopteridae: Megaptera novaeangliae (Borowski, 1781)], and considering that the detachment of extant coronulids from their hosts' skin has been mainly observed in occurrence of cetacean breeding/calving areas, the material here studied supports the existence of a baleen whale migration route between the central Mediterranean Sea (the putative reproductive ground) and the North Atlantic (the putative feeding ground) around 1.8 Ma, when several portions of present-day southern Italy were still submerged. The early Pleistocene utilization of the epeiric seas of southern Italy as breeding/calving areas by migrating mysticetes appears to be linked to the severe climatic degradation that has been recognized at the Gelasian-Calabrian transition and that is marked in the fossil record of the Mediterranean Basin by the appearance of "northern guests" such as Arctica islandica (Linnaeus, 1767) (Bivalvia: Veneroida). The subsequent abandonment of the Mediterranean Sea by most species of mysticetes is likely to have resulted from the progressive emergence of shallow-water coastal environments that occurred in Calabrian and Middle Pleistocene times.
• Cirripedia;
• Coronulidae;
• Mysticeti;
• Balaenopteridae;
• habitat loss;
• Mediterranean Basin;
• cetacean migrations;
• phoresis;
• palaeobiogeography
Collareta A., Insacco G., Reitano A., Catanzariti R., Bosselaers M., Montes M. & Bianucci G. (2018).- Fossil whale barnacles from the lower Pleistocene of Sicily shed light on the coeval Mediterranean cetacean fauna.- Carnets Geol., Madrid, vol. 18, no. 2, p. 9-22.
Des balanes de baleine fossiles du Pléistocčne inférieur de Sicile renseignent sur la faune de cétacés contemporaine en mer Méditerranée.- Nous signalons trois coquilles de balanes de baleine (Cirripedia: Coronulidae) provenant des dépôts marins peu profonds du Pléistocčne de Cinisi (nord-ouest de la Sicile, Italie méridionale). Ces spécimens sont identifiés comme appartenant ŕ l'espčce disparue Coronula bifida Bronn, 1831. L'analyse du nannoplancton calcaire du sédiment renfermant les restes de coronulidés date la période de dépôt entre 1,93 et 1,71 Ma (c'est-ŕ-dire au passage Gélasien-Calabrien), période ŕ laquelle est également connu un autre dépôt riche en balanes de baleine, situé au sud-est de l'Apulie (Italie méridionale). Bien que Coronula Lamarck, 1802, soit actuellement trouvée en position fixée sur la peau de baleines ŕ bosse [Cetacea : Balaenopteridae : Megaptera novaeangliae (Borowski, 1781)], et sachant que le décollement des coronulidés actuels de la peau de leur hôte a principalement été observé sur les sites de reproduction et de vęlage des cétacés, le matériel présenté plaide en faveur de l'existence d'une route migratoire active pour les baleines ŕ fanons entre la mer Méditerranée centrale (le site de reproduction présumé) et l'Atlantique nord (l'aire d'alimentation présumée) autour de 1,8 Ma, alors que plusieurs parties de l'Italie méridionale actuelle étaient encore sous les eaux. La fréquentation au Pléistocčne inférieur des mers épicontinentales d'Italie méridionale comme aires de reproduction et de vęlage par les mysticčtes migrateurs semble ętre liée ŕ la dégradation climatique dramatique qui a eu lieu au passage Gélasien-Calabrien et qui se marque dans le registre fossile du bassin méditerranéen par l'apparition d' "espčces nordiques" telles qu'Arctica islandica (Linnaeus, 1767) (Bivalvia : Veneroida). Le délaissement consécutif de la mer Méditerranée par la plupart des espčces de mysticčtes est probablement la conséquence de l'émersion progressive des environnements côtiers peu profonds qui s'est produite au Calabrien et au Pléistocčne moyen.
• Cirripedia ;
• Coronulidae ;
• Mysticeti ;
• Balaenopteridae ;
• perte d'habitat ;
• bassin méditerranéen ;
• migration des
cétacés ;
• phorésie ;
• paléobiogéographie
Whale barnacles (Crustacea: Cirripedia: Coronulidae) are epizoic organisms that live exclusively on the skin of cetaceans (e.g., Fertl, 2002). While also present on a few species of toothed whales (Cetacea: Odontoceti), these crown-shaped, symbiotic cirripedes are typically found in association with baleen-bearing whales (Cetacea: Mysticeti). The whale barnacles that currently inhabit the skin of mysticetes show a high degree of host specificity; for example, Coronula diadema (Linnaeus, 1767), which likely represents the most widespread coronulid species, attaches preferentially to the humpback whale Megaptera novaeangliae (Borowski, 1781), a member of the family Balaenopteridae, although it has occasionally been reported from other cetacean taxa (including two other balaenopterid species). Other species of whale barnacles exhibit an even greater host specificity; for example, Cetopirus complanatus (Mörch, 1853) is considered exclusive to the skin of the right whales (genus Eubalaena Gray, 1864, currently comprising three species belonging to the family Balaenidae), and Cryptolepas rhachianecti Dall, 1872, is regarded as characteristic of the gray whales (Eschricthius robustus (Lilljeborg, 1861), the sole living member of the family Eschrichtiidae).
The fossil record of the family Coronulidae is rather scanty. Whale barnacles appear to have originated from an ancient dispersal of members of the family Chelonibiidae (a cirripedian lineage of epizoic phoronts of turtles, sirenians, crabs, and other marine vertebrates and invertebrates) on mysticete cetaceans (Collareta et al., 2016a). Although all the fossil occurrences of whale barnacles published so far are reasonably referable to the Pliocene or Quaternary (Collareta et al., 2016a, and previous references therein), some unpublished coronulid specimens from Taiwan may be older (i.e., late Miocene; John Buckeridge, personal communication, 2018). Based on the remarkable host preferences of extant whale barnacle species, and considering that the detachment of coronulid shells from their host's skin has been observed along migration routes and especially in cetacean breeding/calving grounds (Bianucci et al., 2006b, and references therein), the fossil remains of whale barnacles have recently been interpreted as markers of ancient mysticete distributional and migration patterns (Bianucci et al., 2006a, 2006b; Álvarez-Fernández et al., 2014; Bosselaers & Collareta, 2016; Collareta, 2016; Collareta et al., 2016b, 2017, in press; Bosselaers et al., 2017). From this perspective, whale barnacle remains cease to be exclusively regarded as body fossils, assuming instead an additional role – i.e., providing indirect evidence for the passage of their cetacean hosts – that makes them conceptually similar to trace fossils.
The present paper aims at (1) reporting on new fossil remains of coronulid barnacles from lower Pleistocene deposits exposed at Cinisi (northwestern Sicily, southern Italy, central Mediterranean) and (2) outlining their palaeobiogeographical and palaeoecological implications for the coeval cetacean fauna. In doing so, a brief account of the history of scientific research on fossil whale barnacles in Sicily is provided here for the first time.
The still fragmentary Mediterranean fossil record of Coronulidae consists of findings from a few upper Pliocene to Recent sites of Italy (e.g., De Alessandri, 1895, 1906; Menesini, 1968; Bossio et al., 1993; Dominici et al., 2011; Collareta, 2016; Collareta et al., 2016b, in press), Cyprus (Simonelli, 1893; De Alessandri, 1906), and Spain (Álvarez-Fernández et al., 2014; Bosselaers et al., 2017). Based on this record, mainly consisting of upper Pliocene to mid-lower Pleistocene occurrences of the extinct species Coronula bifida Bronn, 1831, and few upper lower Pleistocene (Calabrian) occurrences of the extant species Coronula diadema, Bianucci et al. (2006a, 2006b) proposed that, unlike today, balaenopteroid whales may have used the Mediterranean as a breeding ground during the latest Neogene and part of the Quaternary.
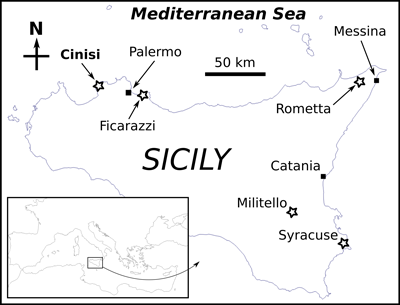
In Sicily (southern Italy, central Mediterranean), fossil whale barnacles
have a long but largely neglected history of discovery and study. To our
knowledge, Aradas (1854) was the first scientist to report on fossil
whale barnacles from Sicily (Fig. 1.a ![]() ). Aradas (1854) described a complete
shell (which he attributed to Coronula diadema) from 'Tertiary' beds
exposed at Militello (Catania Province, Fig.
2
). Aradas (1854) described a complete
shell (which he attributed to Coronula diadema) from 'Tertiary' beds
exposed at Militello (Catania Province, Fig.
2 ![]() ),
and mentioned also a similar
specimen previously found in calcareous deposits near Syracuse (Fig.
2
),
and mentioned also a similar
specimen previously found in calcareous deposits near Syracuse (Fig.
2 ![]() ).
).
Seguenza (1873) reattributed the coronulid specimen
from Militello described by Aradas (1854) to Coronula bifida (a
species that is also known from a few other Italian localities) and tentatively
proposed a Zanclean (i.e., early Pliocene) age for that record. Seguenza
(1873) also cited the presence of shells of C. bifida at Syracuse
(Fig.
2 ![]() ) and reported on three wall plates belonging to the same species found at
Rometta (Messina Province, Fig.
2
) and reported on three wall plates belonging to the same species found at
Rometta (Messina Province, Fig.
2 ![]() ) in 'upper Zanclean marls', without
providing any further indication. Marls and marly clays are common lithologies
in the Piacenzian (i.e., upper Pliocene) to middle Pleistocene
shallow-marine 'Rometta Succession' exposed in northeastern Sicily and
investigated by Seguenza in the 1870s (Di Stefano et al.,
2007). Therefore, the fossil remains of C. bifida from Rometta described
by Seguenza
(1873) are most likely not older than the late Pliocene.
) in 'upper Zanclean marls', without
providing any further indication. Marls and marly clays are common lithologies
in the Piacenzian (i.e., upper Pliocene) to middle Pleistocene
shallow-marine 'Rometta Succession' exposed in northeastern Sicily and
investigated by Seguenza in the 1870s (Di Stefano et al.,
2007). Therefore, the fossil remains of C. bifida from Rometta described
by Seguenza
(1873) are most likely not older than the late Pliocene.
In 1895, De Gregorio created the new species (and subgenus) Coronula
(Flabelcorona) ficarazzensis De Gregorio, 1895, based on a complete
coronulid shell from the "Post-Pliocčne" of Ficarazzi (Palermo
Province, Fig.
2 ![]() ). On the basis of the very approximate and generic description
and figures (Fig. 1.b
). On the basis of the very approximate and generic description
and figures (Fig. 1.b ![]() ) provided by De Gregorio (1895), this taxon was
tentatively synonymized with Coronula balaenaris Darwin,
1854 (= Cetopirus
complanatus) by De Alessandri (1906), whereas Pilsbry
(1916)
considered the "perfectly useless subgenus Flabelcorona" as possibly
indistinguishable from Coronula reginae Darwin, 1854
(a probable
junior synonym of C. diadema: see Dominici et al.,
2011,
and Bosselaers & Collareta,
2016, for further comment).
The current location of the holotype of Coronula (Flabelcorona)
ficarazzensis is not known (Carolina D'Arpa, 2015,
personal communication); as such, its systematic assignment remains an open question.
Nevertheless, some characters illustrated by De Gregorio
(1895) – e.g.,
the dome-like shape of the shell, the narrow and transversely striated true
radius, the moderately thick compound radius, and the presence of rather
frequently bifurcating external ribs flaring downwards in a fan-like triangular
fashion (Fig. 1.b
) provided by De Gregorio (1895), this taxon was
tentatively synonymized with Coronula balaenaris Darwin,
1854 (= Cetopirus
complanatus) by De Alessandri (1906), whereas Pilsbry
(1916)
considered the "perfectly useless subgenus Flabelcorona" as possibly
indistinguishable from Coronula reginae Darwin, 1854
(a probable
junior synonym of C. diadema: see Dominici et al.,
2011,
and Bosselaers & Collareta,
2016, for further comment).
The current location of the holotype of Coronula (Flabelcorona)
ficarazzensis is not known (Carolina D'Arpa, 2015,
personal communication); as such, its systematic assignment remains an open question.
Nevertheless, some characters illustrated by De Gregorio
(1895) – e.g.,
the dome-like shape of the shell, the narrow and transversely striated true
radius, the moderately thick compound radius, and the presence of rather
frequently bifurcating external ribs flaring downwards in a fan-like triangular
fashion (Fig. 1.b ![]() ) – could suggest a relationship between Coronula
(Flabelcorona) ficarazzensis and Cetopirus fragilis Collareta et
al., 2016b, whose holotype and only known specimen comes from
lower Pleistocene deposits exposed at Otranto (southeastern Apulia, southern Italy:
see Discussion below for further details).
) – could suggest a relationship between Coronula
(Flabelcorona) ficarazzensis and Cetopirus fragilis Collareta et
al., 2016b, whose holotype and only known specimen comes from
lower Pleistocene deposits exposed at Otranto (southeastern Apulia, southern Italy:
see Discussion below for further details).
Finally, Cipolla (1978) reported on shell elements of Coronula
bifida from lower Pleistocene sediments exposed along a coastal cliff west
of the city of Cinisi (Palermo Province, Fig.
2 ![]() ), without providing any
illustration or description of these remains. Unfortunately, the work of Cipolla
(1978) was published in a local magazine of natural sciences (Il Naturalista
Siciliano), which is not widely distributed outside Sicily; as a result, the
presence of fossil whale barnacles in the lower Pleistocene of Cinisi was almost
completely overlooked by the scientific community. Here we report on
the discovery of three new fossil specimens of Coronula bifida from the
coastal cliff studied by Cipolla (1978) and discuss their
palaeobiological meaning.
), without providing any
illustration or description of these remains. Unfortunately, the work of Cipolla
(1978) was published in a local magazine of natural sciences (Il Naturalista
Siciliano), which is not widely distributed outside Sicily; as a result, the
presence of fossil whale barnacles in the lower Pleistocene of Cinisi was almost
completely overlooked by the scientific community. Here we report on
the discovery of three new fossil specimens of Coronula bifida from the
coastal cliff studied by Cipolla (1978) and discuss their
palaeobiological meaning.
|
Figure 1:
Early works on the fossil record of whale barnacles from Sicily. a.
Frontispiece of the pioneering work by Aradas (1854). b. Original
illustration of the holotype of Coronula (Flabelcorona) ficarazzensis De
Gregorio, 1895. A description of the features depicted in Fig. 1b is
provided in the main text of the paper by De Gregorio
(1895). |
|
Figure 2:
Location of the finds of fossil whale barnacles (stars) from Sicily
(see text for details and sources of data). The toponym of the study site is
highlighted in bold. |
The
sedimentary succession studied by Cipolla (1978) is about 12
m thick and crops
out just west of Cinisi, along a coastal cliff running between two
localities known as "La Ciucca" and "Torre Molinazzo" (indicative
geographic coordinates: N 38°09'50", E 13°05'07"). This succession
belongs to the "Calcareniti e sabbie di Castellammare lithofacies", a lower Pleistocene sedimentary unit lying unconformably on
pre-Quaternary rocks (Catalano
et al., 2004). This unit consists mostly of calcarenites, yellowish
carbonate-rich sands, and minor conglomeratic and clayey beds, deposited in an inner-shelf environment (Catalano
et al., 2004). The portion of this unit exposed at the study site
(Fig. 3.a ![]() ) hosts a macrofossil assemblage that includes bivalves, gastropods,
echinoids, cirripedes, bryozoans, brachiopods, ostracods, and fragmentary
remains of crabs and fish (Ruggieri, 1976; Cipolla,
1978). Indeterminate cetacean remains including worn fragmentary postcrania are also
present (personal observation by A.C., G.I., and G.B.; Fig.
3.b
) hosts a macrofossil assemblage that includes bivalves, gastropods,
echinoids, cirripedes, bryozoans, brachiopods, ostracods, and fragmentary
remains of crabs and fish (Ruggieri, 1976; Cipolla,
1978). Indeterminate cetacean remains including worn fragmentary postcrania are also
present (personal observation by A.C., G.I., and G.B.; Fig.
3.b ![]() ). A
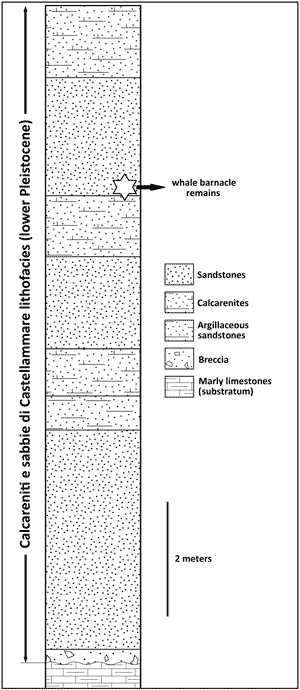
12-m-thick stratigraphic section (Fig. 4
). A
12-m-thick stratigraphic section (Fig. 4 ![]() ) was measured by Cipolla
(1978). The whale barnacle specimens described herein were reported by M.M. and
collected by G.I. during fieldwork on behalf of Museo di Storia Naturale di
Comiso (Comiso, Ragusa Province, Sicily), and come from the base of an interval
of yellowish, bioturbated, fossil-rich, locally indurated sand located in the
upper portion of the Cinisi section (Fig. 4
) was measured by Cipolla
(1978). The whale barnacle specimens described herein were reported by M.M. and
collected by G.I. during fieldwork on behalf of Museo di Storia Naturale di
Comiso (Comiso, Ragusa Province, Sicily), and come from the base of an interval
of yellowish, bioturbated, fossil-rich, locally indurated sand located in the
upper portion of the Cinisi section (Fig. 4 ![]() ). This interval has also produced
shelly remains referred by Cipolla (1978) to the veneroid bivalve Arctica
islandica (Linnaeus, 1767).
). This interval has also produced
shelly remains referred by Cipolla (1978) to the veneroid bivalve Arctica
islandica (Linnaeus, 1767).
Samples of the sandy sediment body hosting the coronulid specimens were collected for biostratigraphic purposes. Calcareous nannofossil analyses were carried out by one of us (R.C.) using a polarized light microscope on a smear-slide prepared from the sample, following standard techniques (Bown & Young, 1998). The calcareous nannofossil assemblages of the samples studied do not differ significantly from each other. The co-occurrence of abundant Calcidiscus macintyrei (Bukry & Bramlette, 1969) Loeblich & Tappan, 1978, and rare, small (<3.5 μm), unidentified Gephyrocapsa Kamptner, 1943, coupled with the absence of Discoaster brouweri Tan, 1927, emend. Bramlette & Riedel, 1954, allowed us to attribute the assemblage detected in the analyzed samples to the lower Pleistocene Mediterranean Neogene Nannoplankton (= MNN) zone 19a of Rio et al. (1990), corresponding to the Calcareous Nannofossil Plio-Pleistocene (= CNPL) zone 7 of the more recent Neogene and Quaternary biozonation of Backman et al. (2012). The bio-events that bound this zone have recently been calibrated by Backman et al. (2012) to 1.93 Ma and 1.71 Ma; consequently, the analyzed samples should be regarded as latest Gelasian to earliest Calabrian (i.e., early Santernian, following the 'Italian Marine Stages' regional scheme of Gibbard & Cohen, 2008). Our biostratigraphic results partially contradict those of Ruggieri (1976, 1987) and Cipolla (1978), who referred to the deposits exposed at Cinisi as middle Calabrian (i.e., Emilian substage of the 'Italian Marine Stages' regional scheme) while evoking the persistence or reappearance of some 'Pliocene' (i.e., pre-Emilian) taxa.
|
Figure
3:
a. Exposure of the "Calcareniti e sabbie di Castellammare
lithofacies" west of the city of Cinisi. b. An indeterminate cetacean
vertebra cropping out from a sandy horizon at the study site. |
|
Figure
4:
Schematic stratigraphic
section of the sedimentary succession exposed at the study site (indicative
geographic coordinates: N 38°09'50", E 13°05'07"). Redrawn and
modified after Cipolla (1978). |
Class MAXILLOPODA Dahl, 1956
Subclass CIRRIPEDIA Burmeister, 1834
Order SESSILIA Lamarck, 1818
Suborder BALANOMORPHA Pilsbry, 1916
Family CORONULIDAE Leach, 1817
Genus Coronula Lamarck, 1802
Diagnosis (after Buckeridge, 1983). Body within shell wall composed of six equal-sized compartments; opercular valves present; parietes with similar structure throughout, without internal midribs; parietal ribs radiate accordion-like to end as T-shaped flanges forming exterior of wall; radii less than half thickness of parietes, orifice of body chamber larger than basal opening; sheath smooth, extending entire length of inner wall; radiating ribs on either side of sutures unbranched or asymmetrically branched; opposed sides of terminal flanges crenulate.
Type species. Lepas diadema Linnaeus, 1767. Early Pleistocene to Recent, cosmopolitan.
Distribution. Upper Pliocene (Piacenzian) to Recent (Collareta et al., 2016a), cosmopolitan (Buckeridge, 1983). Currently known as an exclusive symbiont of cetaceans, displaying a high host specificity for the balaenopterid baleen whale species Megaptera novaeangliae (Borowski, 1781).
Coronula bifida Bronn, 1831
(Fig. 5 ![]() )
)
1831 Coronula bifida Bronn, p. 126.
1854 Coronula barbara Darwin, p. 421, Pl. 15, fig. 6.
1951 Coronula dormitor Pilsbry & Olsson, p. 202.
1968 Coronula bifida bifida Menesini, p. 387, Pls. 1-3, figs. 1a-2b; Pl. 4, figs. 1, 3, 5.
1968 Coronula bifida barbara Menesini, p. 395, Pl. 3, figs. 3a-5c; Pl. 4, figs. 2, 4, 6.
Diagnosis. Shell globose, with longitudinal convex parietal ribs that often bifurcate, and with strong transverse ridges; radii moderately thick, displaying a roughly constant width (or slightly widening downwards) in the upper two thirds of the shell, and then becoming progressively narrower downwards in their lowermost third; body chamber relatively shallow, sub-cylindrical; sheath less than half of the total height of the shell.
Distribution. Upper Pliocene (Piacenzian) to mid-lower Pleistocene. Known as fossil from marine deposits of Belgium (as "C. barbata", error pro C. barbara: Marquet et al., 2009), California (U.S.A.) (as C. barbara: Zullo, 1969), Ecuador (as C. dormitor: Pilsbry & Olsson, 1951), England (as C. barbara: Darwin, 1854), and Italy (both as C. barbara and C. bifida: Bianucci et al., 2006b, and previous references therein; Collareta, 2016; Collareta et al., 2016b, in press).
Material
and repository. Three
rather large specimens kept at Museo Civico di Storia Naturale di Comiso (=
MSNC), Comiso, Ragusa Province, Sicily, under accession numbers MSNC 4454 (Fig.
5.a-c ![]() ), MSNC 4555
(Fig. 5.d-f
), MSNC 4555
(Fig. 5.d-f ![]() ), and MSNC 4556
(Fig. 5.g-h
), and MSNC 4556
(Fig. 5.g-h ![]() ). MSNC 4554 and
MSNC 4555 are two complete and articulated shells, whereas MSNC 4556 is an
incomplete shell consisting of two articulated compartments (rostrum and right
latus).
). MSNC 4554 and
MSNC 4555 are two complete and articulated shells, whereas MSNC 4556 is an
incomplete shell consisting of two articulated compartments (rostrum and right
latus).
Occurrence. Lower Pleistocene shallow-marine deposits outcropping at Cinisi (Palermo Province, northwestern Sicily, southern Italy; indicative geographic coordinates: N 38°09'50", E 13°05'07") and belonging to the "Calcareniti e sabbie di Castellammare lithofacies" as defined by Catalano et al. (2004). The sediment embedding the whale barnacles is attributed to the calcareous nannoplankton zone MNN 19a of Rio et al. (1990), that is, to the interval 1.93 - 1.71 Ma (according to Backman et al., 2012).
Remarks.
The
following set of characters helps in distinguishing the extinct species Coronula
bifida from other coronulid taxa (including the extant species Coronula
diadema): (1) prominent transverse growth ridges on the external surface of
the parietal ribs (e.g., Fig. 5.f ![]() ); (2) frequently bifurcated parietal
ribs (e.g., Fig. 5.b-c
); (2) frequently bifurcated parietal
ribs (e.g., Fig. 5.b-c ![]() ); (3) external radii displaying a roughly constant
width (or slightly expanding in width downwards) in the upper two thirds of the
shell, and then becoming progressively narrower downwards in their lowermost
third (e.g., Fig. 5.c, e-f
); (3) external radii displaying a roughly constant
width (or slightly expanding in width downwards) in the upper two thirds of the
shell, and then becoming progressively narrower downwards in their lowermost
third (e.g., Fig. 5.c, e-f ![]() ); (4) basal margin of the sheath which does
not project freely (e.g., Fig. 5.h
); (4) basal margin of the sheath which does
not project freely (e.g., Fig. 5.h ![]() );
and (5) relatively shallow body chamber,
shorter than half of the total height of the shell (e.g., Fig.
5.h
);
and (5) relatively shallow body chamber,
shorter than half of the total height of the shell (e.g., Fig.
5.h ![]() ) (see
also Menesini, 1968). All these characters can be observed in the
fossil coronulid specimens from Cinisi, thus allowing an unambiguous
species-level determination. Smoothed surfaces (e.g., Fig.
5.a
) (see
also Menesini, 1968). All these characters can be observed in the
fossil coronulid specimens from Cinisi, thus allowing an unambiguous
species-level determination. Smoothed surfaces (e.g., Fig.
5.a ![]() ) and fractures (e.g.,
Fig. 5.d, g
) and fractures (e.g.,
Fig. 5.d, g ![]() ) are mostly observed in the
upper half of the three specimens, suggesting that this part of the
shells was exposed above the skin of the cetacean, as also indicated by the
shallow body chamber of MSNC 4554. By contrast, in C. diadema most of the
shell is exposed outside the skin of the host, so that specimens belonging to
this extant species frequently display evidences of abrasion and wearing in the
upper two thirds (or more) of the shell (Bianucci et al.,
2006a; Dominici
et al., 2011). Marks due to encrustation by non-epizoic barnacles (cf. Anellusichnus
isp. Santos et al., 2005) and other invertebrates (e.g.,
bryozoans) are observable on all specimens (e.g., Fig.
5.c
) are mostly observed in the
upper half of the three specimens, suggesting that this part of the
shells was exposed above the skin of the cetacean, as also indicated by the
shallow body chamber of MSNC 4554. By contrast, in C. diadema most of the
shell is exposed outside the skin of the host, so that specimens belonging to
this extant species frequently display evidences of abrasion and wearing in the
upper two thirds (or more) of the shell (Bianucci et al.,
2006a; Dominici
et al., 2011). Marks due to encrustation by non-epizoic barnacles (cf. Anellusichnus
isp. Santos et al., 2005) and other invertebrates (e.g.,
bryozoans) are observable on all specimens (e.g., Fig.
5.c ![]() ), similar to
features reported by Bianucci et al. (2006a) for fossil shells of C.
diadema from the lower Pleistocene (probably Calabrian; Collareta,
2016) of Ecuador.
), similar to
features reported by Bianucci et al. (2006a) for fossil shells of C.
diadema from the lower Pleistocene (probably Calabrian; Collareta,
2016) of Ecuador.
Based on the occurrence of a single, well-preserved, large (i.e., displaying a diameter of 60 - 70 mm) shell of Coronula diadema from the upper lower Pleistocene of Riparbella (Tuscany, central Italy), Dominici et al. (2011) proposed that C. diadema is a direct descendant of C. bifida and that the evolution of the former included a pronounced increase in size of the adult shell. Interestingly, one of the specimens of C. bifida described herein (MSNC 4554) exhibits a maximum diameter of 73 mm, that is, seemingly slightly larger than that of the specimen of C. diadema described by Dominici et al. (2011). Moreover, with regard to 56 well-preserved shells of Coronula diadema from the lower Pleistocene Canoa and Tablazo Formations of Ecuador, Bianucci et al. (2006b) noted that their average size is distinctly greater than that observed for extant individuals of Coronula diadema from the skin of humpback whales taken off Madagascar (Angot et al., 1951). Bianucci et al. (2006b) argued that the fragile construction apparent in juveniles of Coronula could render them less likely to fossilize, at least as complete and articulated shells, thus significantly biasing the size parameters of fossil whale barnacle assemblages. Therefore, we contend that the evolutionary trend towards an increase in size proposed by Dominici et al. (2011) for Coronula still needs to be confirmed.
|
Figure
5:
MSNC 4554, MSNC 4555, and MSNC 4556, three fossil
shells of Coronula bifida Bronn, 1831, from lower Pleistocene
deposits exposed near Cinisi (Palermo
Province, western Sicily, southern Italy). a-c. Specimen MSNC 4554. d-f.
Specimen MSNC 4555. g-h. Specimen MSNC 4556. Panels a and d: subapical
view; panel b: basal view; panels c, e, f, and g: lateral view; panel h:
internal
view. |
During the Pliocene, mysticetes were abundant and diverse along the coasts of Italy, being represented by several skeletons referred to species of at least three families (Balaenidae, Balaenopteridae, and Eschrichtiidae) (e.g., Bisconti, 2002, 2003, 2007, 2008, 2009; Bianucci et al., 2009). In turn, the balaenopterid fin whale Balaenoptera physalus (Linnaeus, 1758) is the sole baleen whale species known to inhabit the modern waters of the Mediterranean Sea regularly (e.g., Cagnolaro et al., 1993) and is recognized as forming a genetically distinct sub-population displaying largely resident (i.e., non-migratory) habits (Panigada & Notarbartolo di Sciara, 2012, and references therein). The transition from the Pliocene to the extant baleen whale fauna of the Mediterranean Sea is still obscure, as the Pleistocene record of mysticetes (and of cetaceans generally) is very poor in this basin (e.g., Collareta et al., 2016b, and references therein), although a few recent discoveries from Italy (e.g., Margiotta & Varola, 2007; Bianucci et al., 2012; Tsai et al., 2017) are helping to close this gap. Nevertheless, Pleistocene remains of whale barnacles still play a pivotal role in illustrating the dynamics of the cetacean fauna of the Mediterranean Sea during the Quaternary as reliable indicators of ancient baleen whale populations.
As reported above, the sediment hosting the coronulid specimens from Cinisi was referred to zone MNN 19a of Rio et al. (1990) by means of calcareous nannoplankton biostratigraphy. Interestingly, the same lower Pleistocene biozone was identified in fossiliferous deposits of southeastern Sicily featuring remains of a pygmy right whale (cf. Caperea sp. Gray, 1864) (Tsai et al., 2017), as well as in inner-neritic deposits of southeastern Apulia (southern Italy) containing fossil whale barnacles (attributed to Coronula bifida and Cetopirus fragilis) (Collareta, 2016; Collareta et al., 2016b, in press). The latter occurrence was recently interpreted as indirect evidence of an ancient breeding/calving ground shared by humpback whales (or other balaenopterids closely related to Megaptera Gray, 1846) and right whales (or other balaenids closely related to Eubalaena) (Collareta et al., 2016b). Our new finds from Cinisi confirm that, during the early Pleistocene, whales colonized by Coronula bifida (most likely belonging to the family Balaenopteridae, in light of the host preferences of the Recent species Coronula diadema) frequented the shallow coastal waters of the central Mediterranean Sea (presumably for breeding and calving purposes, considering that the detachment of whale barnacles from living whales has been primarily observed in zones of reproduction). Furthermore, since the fossil whale barnacles from Otranto and Cinisi come from sedimentary units that were deposited during the same, relatively short period of time (i.e., between 1.93 and 1.71 Ma), they seemingly depict the same phase of dispersal of migrating North Atlantic cetaceans within the Mediterranean Basin, and in particular in the epeiric seas located off the coasts of modern southern Italy. From this perspective, baleen whales inhabiting the high-latitude areas of the northeastern Atlantic could have chosen to use the central region of the Mediterranean Basin as a winter breeding/calving ground during the Gelasian-Calabrian transition, possibly as a response to Northern Hemisphere climate change (see also Clementz et al. (2014), Boessenecker & Fordyce (2015), Marx & Fordyce (2015), and Tsai (2017) for an updated discussion about the emergence of migratory habits in mysticetes). Indeed, recent research indicates that the waters of the central Mediterranean Sea saw a strong increase in thermal seasonality, coupled with a strong decrease of average winter temperature, around 1.80 Ma (Crippa et al., 2016). This severe climatic degradation is famously marked by the appearance of cold-water North Atlantic invertebrate taxa ("northern guests") such as the ocean quahog bivalve Arctica islandica (a species that has been reported as occurring alongside Coronula bifida in the deposits exposed at the study site) in the Mediterranean fossil record around 2.0-1.8 Ma (e.g., Gibbard & Head, 2010; Crippa & Raineri, 2015, and references therein). Such a drastic basin-wide cooling episode reflected global climate degradation that implies, in the Atlantic Ocean, both a significant shift of the polar/subpolar water fronts towards the Equator and a remarkable lowering of sea surface temperatures at low latitudes, thus ultimately allowing a cool-temperate Southern Hemisphere form such as Caperea to cross the normally impassable tropics and enter the Mediterranean Basin (Tsai et al., 2017).
Since several currently emerged portions of southern Italy were still submerged at the time of the Gelasian-Calabrian transition (Santangelo et al., 2012), lagoons punctuated by islands and other sheltered shallow neritic habitats potentially suitable as breeding/calving grounds for mysticete cetaceans should have been probably well-represented in the central part of the Mediterranean Basin, at the foot of the rising southern Apennine chain (a palaeoenvironmental scenario somewhat reminiscent of today's inshore reproduction sites of Eschrichtius robustus and Megaptera novaeangliae along the coasts of Baja California; e.g., Scammon, 1968; Gendron & Urban, 1993; Hindell, 2009). Due to a long-lasting pattern of widespread emergence of coastal epeiric sea regions, most of these key environments suffered a progressive reduction during the late early (Calabrian) and Middle Pleistocene (Santangelo et al., 2012), thus possibly leading to the abandonment of the central Mediterranean nursery areas by most large species of migrating mysticetes in favor of breeding and calving grounds located outside the Mediterranean Basin (e.g., in the Gulf of Biscay or off the coasts of northwest Africa). The persistence of large mysticetes in the central Mediterranean at the end of the early Pleistocene is nevertheless witnessed by a partial balaenopterid skeleton, provisionally attributed to the genus Balaenoptera Lacépčde, 1804, from Calabrian deposits exposed at San Giuliano (Matera Province, Basilicata, Italy) (Bianucci et al., 2012). As reported above, the youngest fossil occurrence of Coronula from the Mediterranean Basin is represented so far by a single shell collected from inner-shelf deposits exposed near Riparbella (Tuscany, central Italy) in mudstones tentatively referred by Dominici et al. (2011) to the mid-upper Calabrian zone MNN 19e of Rio et al. (1990), corresponding to zone CNPL 9 of Backman et al. (2012), regarded as ranging from 1.25 to 1.06 Ma.
In conclusion, investigations focusing on the fossil record of whale barnacles (Coronula and allied forms) highlight a rather complex and previously unexpected Quaternary history for the Mediterranean mysticetes. New additions to the still fragmentary Italian Pleistocene record of fossil mysticetes will hopefully further elucidate the biogeographical patterns and dynamics of the Mediterranean baleen whale fauna during the Quaternary.
Note added in proofs: During the proofreading of the present paper, another contribution (Collareta et al., in press) dealing with the Mediterranean record of Coronula was accepted for publication in an international journal. In this forthcoming paper, the hypothesis of a baleen whale migratory route active in early Pleistocene times between the central Mediterranean Sea and the northeastern Atlantic is further investigated in the light of oxygen-isotope analyses of a fossil whale barnacle shell from Apulia.
We
are grateful to Daniele Raimondi (Universitŕ degli Studi di Palermo) who
took part in the discovery of one of the whale barnacle specimens described in
the present paper. Walter Landini (Universitŕ di Pisa) and Enrico Di
Stefano (Universitŕ degli Studi di Palermo) contributed greatly to the
initiation of the present research. Carolina D'Arpa (Universitŕ degli Studi di
Palermo) and Sebastiano Italo Di Geronimo (Universitŕ degli Studi di
Catania) are thanked for their help in reconstructing
the history of scientific research on fossil whale barnacles from Sicily. John
W.M. Jagt (Natuurhistorisch Museum Maastricht) kindly shared with us an
electronic version of the work by De Gregorio (1895), an excerpt of which
is reported in Fig. 1.a ![]() of the present paper.
of the present paper.
We are deeply indebted to John Buckeridge (RMIT University) and Olivier Lambert (Institut Royal des Sciences Naturelles de Belgique), whose constructive criticisms and thoughtful remarks greatly improved an early version of the paper. Thanks are also due to Bruno Granier (Université de Bretagne Occidentale) and to Stephen Carey (Federation University Australia) for their most valuable editorial support.
Álvarez-Fernández E., Carriol R.-P., Jordá J.F., Aura J.E., Avezuela B., Badal E., Carrión Y., García-Guinea J., Maestro A., Morales J.V., Perez G., Perez-Ripoll M., Rodrigo M.J., Scarff J.E., Villalba M.P. & Wood R. (2014).- Occurrence of whale barnacles in Nerja Cave (Málaga, southern Spain): Indirect evidence of whale consumption by humans in the Upper Magdalenian.- Quaternary International, Oxford, vol. 337, p. 163-169.
Angot M. (1951).- Rapport scientifique sur les expéditions baleiničres autour de Madagascar.- Mémoires de l'Institut de Recherche Scientifique de Madagascar, Séries A, Antananarivo, vol. 6, p. 439-486.
Aradas A. (1854).- Monografia del genere Coronula e descrizione di alcune altre nuove specie di conchiglie siciliane.- Atti dell'Accademia Gioenia di Scienze Naturali, Catania, vol. 9, p. 57-72.
Backman J., Raffi I., Rio D., Fornaciari E. & Pälike H. (2012).- Biozonation and biochronology of Miocene through Pleistocene calcareous nannofossils from low and middle latitudes.- Newsletters on Stratigraphy, Leiden, vol. 45, p. 221-244.
Bianucci G., Di Celma C., Landini W. & Buckeridge J. (2006a).- Palaeoecology and taphonomy of an extraordinary whale barnacle accumulation from the Plio-Pleistocene of Ecuador.- Palæogeography, Palæoclimatology, Palæoecology, vol. 242, p. 326-342.
Bianucci G., Landini W. & Buckeridge J. (2006b).- Whale barnacles and Neogene cetacean routes.- New Zealand Journal of Geology and Geophysics, Wellington, vol. 49, p. 115-120.
Bianucci G., Landini W., Varola A., Bianco S. & De Siena A. (2012).- From sea to lake: The giant baleen whale of San Giuliano (Matera, Italy): An evidence of Pleistocene body size increase acceleration. In: Rosso A. & Sanfilippo R. (eds.), Giornate di Paleontologia, XII Edizione, Volume dei Riassunti.- Catania, p. 35.
Bianucci G., Sorbi S., Vaiani S.C. & Landini W. (2009).- Pliocene marine mammals from Italy: A systematic and stratigraphic overview. In: Fanti F. & Spalletta C. (eds.), International conference on Vertebrate palaeobiogeography and continental bridges across Tethys, Mesogea, and Mediterranean Sea, Abstract Book.- Bologna, p. 912.
Bisconti M. (2002).- Morphology and phylogenetic relationships of a new eschrichtiid genus (Cetacea: Mysticeti) from the Early Pliocene of northern Italy.- Zoological Journal of the Linnean Society, London, vol. 153, p. 161-186.
Bisconti M. (2003).- Evolutionary history of Balaenidae.- Cranium, Leiden, vol. 20, p. 9-50.
Bisconti M. (2007).- A new basal balaenopterid whale from the Pliocene of northern Italy.- Palaeontology, London, vol. 50, p. 1103-1122.
Bisconti M. (2008).- An early Late Pliocene right whale (genus Eubalaena) from Tuscany (central Italy).- Bollettino della Societa Paleontologica Italiana, Modena, vol. 41, p. 83-92.
Bisconti M. (2009).- Taxonomy and evolution of the Italian Pliocene Mysticeti (Mammalia, Cetacea): A state of the art.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 48, p. 147-156.
Boessenecker R.W. & Fordyce R.E. (2015).- Anatomy, feeding ecology, and ontogeny of a transitional baleen whale: A new genus and species of Eomysticetidae (Mammalia: Cetacea) from the Oligocene of New Zealand.- PeerJ, San Diego (CA), vol. 3, article #e1129.
Borowski G.H. (1781).- Gemmeinuzige Naturgeschichte des Theirreichs, Band 2.- Gottlieb August Lange, Berlin, 196 p. URL: https://www.biodiversitylibrary.org/item/89998#page/7/mode/1up
Bosselaers M. & Collareta A. (2016).- The whale barnacle Cryptolepas rhachianecti (Cirripedia: Coronulidae), a phoront of the grey whale Eschrichtius robustus (Cetacea: Eschrichtiidae), from a sandy beach in The Netherlands.- Zootaxa, vol. 4154, p. 332-338.
Bosselaers M., Nieulande M. van & Collareta A. (2017).- A new record of Cetopirus complanatus (Cirripedia: Coronulidae), an epibiont of right whales (Cetacea: Balaenidae: Eubalaena spp.), from a beach deposit of Mediterranean Spain.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. CXXIV, p. 43-48.
Bossio A., Landini W., Mazzei R., Ragaini L., Salvatorini G. & Varola A. (1993).- I fossili pliocenici di Sant'Andrea. In: Guida alle escursioni, XII Convegno della Societŕ Paleontologica Italiana.- Lecce, p. 69-77.
Bown P.R. & Young J.R. (1998).- Techniques. In: Bown P.R. (ed.), Calcareous nannofossil biostratigraphy.- Kluwer Academic Publishers, Dordrecht.
Bramlette M.N. & Riedel W.R. (1954).- Stratigraphic value of discoasters and some other microfossils related to Recent coccolithophores.- Journal of Paleontology, New York, vol. 28, p. 385-403.
Bronn H.G. (1831).- Italiens Tertiär-Gebilde und deren organische Einschlüsse: vier Abhandlungen.- Groos, Heidelberg, 176 p.
Buckeridge J.S. (1983).- Fossil barnacles (Cirripedia: Thoracica) of New Zealand and Australia.- New Zealand Geological Survey Paleontological Bulletin, Wellington, vol. 50, p. 1-151.
Bukry D. & Bramlette M.N. (1969).- Some new and stratigraphically useful calcareous nannofossils of the Cenozoic.- Tulane Studies in Geology and Paleontology, New Orleans, vol. 7, p. 131-142.
Burmeister H.I. (1834).- Beiträge zur Naturgeschichte der Rankenfüsser (Cirripedia).- G. Reimer, Berlin, 60 p.
Cagnolaro L., Notarbartolo di Sciara G. & Podestŕ M. (1993).- Profilo della cetofauna dei mari italiani.- Supplemento alle Ricerche di Biologia della Selvaggina, Bologna, vol. 21, p. 101-114.
Catalano R., Agate M., Basilone L., Di Maggio C., Mancuso M. & Sulli A. (2004).- Foglio 593. Castellammare del Golfo.- Note Illustrative della Carta Geologica d'Italia, Roma, 239 p.
Cipolla P. (1978).- Macrofauna del Pleistocene inferiore di Cinisi (Sicilia).- Il Naturalista Siciliano, Serie IV, Palermo, vol. II, p. 55-67.
Clementz M.T., Fordyce R.E., Peek S.L. & Fox D.L. (2014).- Ancient marine isoscapes and isotopic evidence of bulk-feeding by Oligocene cetaceans.- Palæogeography, Palæoclimatology, Palæoecology, vol. 400, p. 28-40.
Collareta A. (2016).- Fossil turtle and whale barnacles (Crustacea: Cirripedia: Coronuloidea) kept at the Museo di Storia Naturale dell'Universitŕ di Pisa: An annotated catalogue.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 123, p. 41-45.
Collareta A., Marean C.W., Jerardino A. & Bosselaers M. (2017).- Cetopirus complanatus (Cirripedia: Coronulidae) from the Middle Pleistocene human settlement of Pinnacle Point 13B (Mossel Bay, South Africa).- Zootaxa, Auckland, vol. 4237, p. 393-400.
Collareta A., Bosselaers M. & Bianucci G. (2016a).- Jumping from turtles to whales: A Pliocene fossil record depicts an ancient dispersal of Chelonibia on mysticetes.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), Milan, vol. 122, p. 35-44.
Collareta A., Margiotta S., Varola A., Catanzariti R., Bosselaers M. & Bianucci G. (2016b).- A new whale barnacle from the early Pleistocene of Italy suggests an ancient right whale breeding ground in the Mediterranean.- Comptes Rendus Palevol, Paris, vol. 15, p. 473-481.
Collareta A., Regattieri E., Zanchetta G., Lambert O., Catanzariti R., Bosselaers M., Covelo P., Varola A. & Bianucci G. (in press).- New insights on ancient cetacean movement patterns from oxygen-isotope analyses of a Mediterranean Pleistocene whale barnacle.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart. DOI: 10.1127/njgpa/2018/0729
Crippa G., Angiolini L., Bottini C., Erba E., Felletti F., Frigerio C., Hennissen J.A.I., Leng M.J., Petrizzo M.R., Raffi I., Raineri G. & Stephenson M.H. (2016).- Seasonality fluctuations recorded in fossil bivalves during the early Pleistocene: Implications for climate change.- Palæogeography, Palæoclimatology, Palæoecology, vol. 446, p. 234-251.
Crippa G. & Raineri G. (2015).- The genera Glycymeris, Aequipecten and Arctica, and associated mollusk fauna of the Lower Pleistocene Arda River section (Northern Italy).- Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), Milan, vol. 121, p. 61-101.
Dahl E. (1956).- Some crustacean relationships.- In: Wingstrand K.G. (ed.), Bertil Hanström: Zoological papers in honor of his sixty-fifth birthday.- Zoological Institute, Lund, p. 138-147.
Dall W.H. (1872).- On the parasites of the cetaceans of the N.W. coast of America, with descriptions of new forms.- Proceedings of the California Academy of Sciences, San Francisco, vol. 4, p. 299-301.
Darwin C. (1854).- A Monograph on the subclass Cirripedia with figures of all the species. The Balanidae, The Verrucidae, etc.- Ray Society, London, 684 p.
De Alessandri G. (1895).- Contribuzione allo studio dei Cirripedi fossili d'Italia.- Bollettino della Societŕ Geologica Italiana, Roma, vol. 13, p. 234-314.
De Alessandri G. (1906).- Studi monografici sui cirripedi fossili d'Italia.- Palaeontographia Italica, Pisa, vol. 12, p. 207-324.
De Gregorio A. (1895).- Note sur un astéride et un cirripčde du Postpliocčne de Sicile des genres Astrogonium et Coronula.- Annali di Geologia e Paleontologia di Palermo, Palermo, vol. 17, p. 1-7.
Di Stefano A., Longhitano S. & Smedile A. (2007).- Sedimentation and tectonics in a steep shallow-marine depositional system: Stratigraphic arrangement of the Pliocene-Pleistocene Rometta Succession (NE Sicily, Italy).- Geologica Carpathica, Bratislava, vol. 58, p. 71-87.
Dominici S., Bartalini M., Benvenuti M. & Balestra B. (2011).- Large kings with small crowns: A Mediterranean Pleistocene whale barnacle.- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 50, p. 95-101.
Fertl D. (2002).- Barnacles. In: Perrin W.F., Wursig B. & Thewissen J.G.M. (eds.), Encyclopedia of marine Mammals.- Academic Press, San Diego, p. 75-78.
Gendron D. & Urban J. (1993).- Evidence of feeding by humpback whales (Megaptera novaeangliae) in the Baja California breeding ground, Mexico.- Marine Mammal Science, Oxford, vol. 9, p. 76-81.
Gibbard P. & Cohen K.M. (2008).- Global chronostratigraphical correlation table for the last 2.7 million years.- Episodes, Beijing, vol. 31, p. 243-247.
Gibbard P.L. & Head J.J. (2010).- The newly-ratified definition of the Quaternary System/Period and redefinition of the Pleistocene Series/Epoch, and comparison of proposals advanced prior to formal ratification.- Episodes, Beijing, vol. 33, p. 152-158.
Gray J.E. (1846).- On the British Cetacea.- Annals and Magazine of Natural History, Series 1, London, vol. 17, p. 82-8.
Gray J.E. (1864).- On the Cetacea which have been observed in the seas surrounding the British Islands.- Proceedings of the Zoological Society of London, London, vol. 1864, p. 195-248.
Hindell M.A. (2009).- Breeding sites. In: Perrin W.F., Wursig B. & Thewissen J.G.M. (eds.), Encyclopedia of marine Mammals (Second edition).- Academic Press, San Diego, p. 156-158.
Kamptner E. (1943).- Zur Revision der Coccolithineen-Spezies Pontosphaera huxleyi Lohm.- Anzeiger der Akademie der Wissenschaften in Wien, Mathematische-Naturwissenchaftliche Klasse, Vienna, vol. 80, p. 73-49.
Lacépčde B.G.É. (1804).- Histoire naturelle des Cétacées. Chez Plassan Imprimeur-Libraire, Paris, 329 p.
Lamarck J.B. de (1802).- Mémoire sur la Tubicinelle.- Annales du Muséum National d'Histoire Naturelle, Paris, vol. 1, p. 461-464.
Lamarck J.B. de (1818).- Histoire naturelle des animaux sans vertčbres, 5.- Verdičre, Paris, 612 p.
Leach W.E. (1817).- Distribution systématique de la classe des Cirripčdes.- Journal de Physique, de Chimie et d'Histoire Naturelle, Paris, t. LXXXV, p. 67-69.
Lilljeborg W. (1861).- Hvalben funna ijorden pĺ Gräsön I Roslagen i Sverige.- Forhandlinger ved de skandinaviske Naturforskeres, Kopenhagen, vol. 8 (1860), p. 599-616.
Linnaeus C. (1758).- Systema Naturae sive Regna Tria Naturae, secundum Classes, Ordines, Genera, Species, cum characteribus, differentiis, synonymis, locis, Tomus I. Editio decima, reformata.- Stockholm, 824 p.
Linnaeus C. (1767).- Systema Naturae sive Regna Tria Naturae, secundum Classes, Ordines, Genera, Species, cum characteribus, differentiis, synonymis, locis, Tomus I. Editio duodecima, Reformata. L. Salvius, Stockholm, 1327 p.
Loeblich A.R. & Tappan H. (1978).- The coccolithophorid genus Calcidiscus Kamptner and its synonyms.- Journal of Paleontology, New York, vol. 52, p. 1390-1392.
Margiotta S. & Varola A. (2007).- Il paleosito di Cutrofiano (Salento): Proposta per l'istituzione di un parco-museo.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 112, p. 1-8.
Marquet R., Collins J., Jagt J. & Bakel B. van (2009).- The invertebrate fauna of the Pliocene in Belgium. Part I. Annellida, Anthozoa and Crustacea.- Palaeofocus, Mortsel, vol. 2, p. 41-61.
Marx F.G. & Fordyce R.E. (2015).- Baleen boom and bust: A synthesis of mysticete phylogeny, diversity and disparity.- Royal Society Open Sciences, vol. 2, article #140434.
Menesini E. (1968).- Osservazioni su Coronula bifida Bronn.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 75, p. 387-398.
Mörch O.A.L. (1853).- Cephalophora. In Catalogus Conchyliorum.- Typis Ludovici Kleini, Copenhagen, p. 65-68.
Panigada S. & Notarbartolo di Sciara G. (2012).- Balaenoptera physalus (Mediterranean subpopulation). In: The IUCN red list of threatened species 2012, e.T16208224A17549588. URL: https://doi.org/10.2305/IUCN.UK.2012.RLTS.T16208224A17549588.en
Pilsbry H.A. (1916).- The sessile barnacles (Cirripedia) contained in the collections of the U.S. National Museum; including a monograph of the American species.- Bulletin of the United States National Museum, Washington D.C., vol. 93, p. 1-366.
Pilsbry H.A. & Olsson A.A. (1951).- Tertiary and Cretaceous Cirripedia from northwestern South America.- Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 103, p. 197-210.
Rio D., Raffi I. & Villa G. (1990).- Pliocene-Pleistocene calcareous nannofossil distribution patterns in the Western Mediterranean.- Proceedings of the Ocean Drilling Program, Scientific Results, Washington D.C., 107, 513-533.
Ruggieri G. (1976).- La Ostracofauna pleistocenica della Falesia di Cinisi (Sicilia).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 15, p. 85-106.
Ruggieri G. (1987).- La ricomparsa nel Pleistocene marino del Mediterraneo di molluschi pliocenici.- Atti della Societŕ Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 94, p. 1-10.
Santangelo N., Di Donato V., Lebreton V., Romano P. & Ermolli E.R. (2012).- Palaeolandscapes of Southern Apennines during the late Early and the Middle Pleistocene.- Quaternary International, vol. 267, p. 20-29.
Santos A., Mayoral E. & Muńiz F. (2005).- Bioerosion scars of acorn barnacles from the southwestern Iberian Peninsula, upper Neogene.- Rivista Italiana di Paleontologia e Stratigrafia (Research In Paleontology and Stratigraphy), Milan, vol. 111, p. 181-189.
Scammon C.M. (1968).- The marine mammals of the North-Western coast of North America described and illustrated: Together with an account of the American whale fishery (re-issued volume).- Dover Publications, New York, 319 p.
Seguenza G. (1873).- Ricerche paleontologiche intorno ai Cirripedi terziari della provincia di Messina. Con appendice intorno ai Cirripedi viventi nel Mediterraneo e sui fossili terziari dell'Italia Meridonale. Parte I.- Atti dell'Accademia Pontaniana, Napoli, vol. 10, p. 267-311.
Simonelli V. (1893).- Fossili terziari e postpliocenici dell'Isola di Cipro.- Memorie della Reale Accademia delle Scienze dell'Istituto di Bologna, Serie V, Bologna, vol. 3, p. 353-362.
Tan S.H. (1927).- Discoasteridae incertae sedis.- Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen-Amsterdam, Amsterdam, vol. 30, p. 411-419.
Tsai C.-H. (2017).- A Miocene breeding ground of an extinct baleen whale (Cetacea: Mysticeti).- PeerJ, San Diego (CA), vol. 5, article #e3711.
Tsai C.-H., Collareta A., Fitzgerald E.M.G., Marx F.G., Kohno N., Bosselaers M., Insacco G., Reitano A., Catanzariti R., Oishi M. & Bianucci G. (2017).- Northern pygmy right whales highlight Quaternary marine mammal interchange.- Current Biology, London, vol. 27, p. R1058-R1059.
Zullo V.A. (1969).- Thoracic Cirripedia of the San Diego Formation, San Diego County, California.- Contributions in Science, Natural History Museum of Los Angeles County, Los Angeles, vol. 159, p. 1-25.