◄ Carnets Geol. 18 (11) ►
![]()
Sommaire
[1 - Introduction]
[2 - Localités étudiées]
[3 - Matériel et méthodes]
[4 - Architecture et endosquelette]
[5 - Critčres d'identification des genres et des espčces]
[6 - Biostratigraphie]
[7 - Systématique]
[8 - Conclusions] et ...
[Références bibliographiques]
118 avenue de Flandre, F-59290 Wasquehal (France)
Published online in final form (pdf) on September 4, 2018
DOI 10.4267/2042/68382
[Éditeur scientifique : Bruno Granier ;
"language editor" (extended abstract) : Robert W. Scott]
![]()
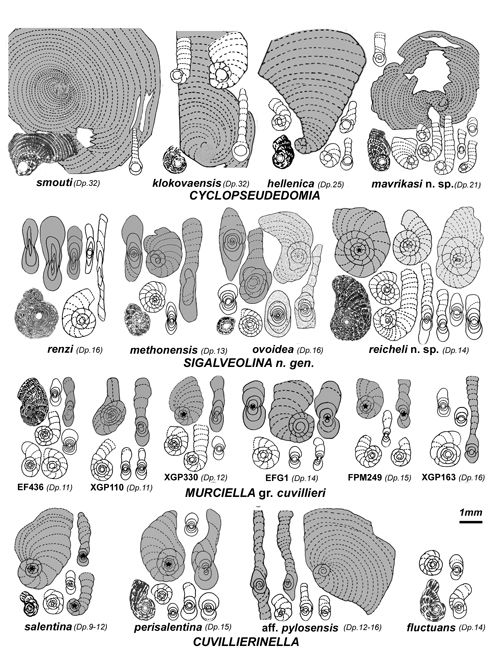
Les trois taxons de la famille des Rhapydioninidae analysés sont issus du genre Cuvillierinella Papetti & Tedeschi,
1965, et sont regroupés au sein de la sous-famille Cuvillierinellinae. Ils sont réexaminés ŕ partir des types et de diverses populations nouvelles qui permettent d'approfondir et renouveler leur connaissance. Leurs tests A sont tous entičrement planispiralés et dotés ŕ des degrés divers de la
"structure hélicoďdale", renommée ici structure confluente. Le genre Murciella est représenté par l'unique espčce M. cuvillieri, encore proche de Cuvillierinella mais entičrement planispiralée ; les six populations étudiées, apparemment toutes de męme âge, réparties de l'Espagne au Dodécančse, témoignent d'une homogénéité d'ensemble qui les isole des autres taxons de la famille ; l'holotype de l'espčce est interprété avec une grande vraisemblance comme représentant la génération microsphérique, ŕ la lumičre de sections centrées de męme morphologie appartenant ŕ des populations nouvellement décrites. Le genre Sigalveolina n. gen., individualisé par la réalisation parfaite de la structure confluente, est représenté par quatre espčces ; il s'agit pour trois d'entre elles du groupe précédemment dénommé
"Murciella gr. renzi" et d'une quatričme, nouvelle, S. reicheli, qui correspondrait au mythique matériel, parfois évoqué sous le nom de
"Cosinella", anciennement découvert par Schlumberger dans les confins italo-slovčnes. Le genre Cyclopseudedomia, caractérisé par son grand proloculus A et ses tests des deux générations presque entičrement évolutes est reconsidéré au moyen de la confrontation de quatre espčces dont trois déjŕ connues et une nouvelle, S. mavrikasi, en provenance des confins italo-slovčnes et d'Italie méridionale. On note en outre la présence ponctuelle d'un organisme rapproché de Pseudonummoloculina kalantarii
qui semble donner, avec d'autres espčces attribuées au męme genre ŕ d'autres époques et en d'autres lieux, une image du groupe de Miliolacea dont sont issues les diverses lignées constituant les Alveolinacea.
Il s'agit ainsi de la derničre partie d'une confrontation générale des taxons méditerranéens fini-crétacés de la famille des Rhapydioninidae, qui fait suite ŕ celles des genres Rhapydionina
- Fanrhapydionina puis Cuvillierinella - Metacuvillierinella menées précédemment par l'auteur. Un tableau général des traits exo- et endosquelettiques mis en śuvre par cette famille et une récapitulation dessinée des morphologies de la plupart des taxons qui la constituent en donnent une vue synthétique. Leur répartition stratigraphique en trois zones du Campanien Maastrichtien est précisée ; on peut douter que ces données soient aisément transposables ŕ des régions qui n'auraient pas été affectées par trois périodes d'émersion qui semblent avoir été ŕ l'origine des disparitions et apparitions successives dont semble témoigner la zonation.
• Campanien-Maastrichtien ;
• Alveolinacea ;
• Rhapydioninidae ;
• domaine méditerranéen ;
• Murciella ;
• Sigalveolina n. gen. ;
• Cyclopseudedomia ;
• nouveau genre ;
• nouvelles espčces.
Fleury J.-J. (2018).- Rhapydioninidés du Campanien-Maastrichtien en région méditerranéenne : Les genres Murciella, Sigalveolina n. gen. et Cyclopseudedomia.- Carnets Geol., Madrid, vol. 18, no. 11, p. 233-280.
Campanian-Maastrichtian Rhapydioninidae in the Mediterranean area: Genera
Murciella, Sigalveolina n.
gen., and Cyclopseudedomia.- This is the third and last part of a review of the Rhapydioninidae,
benthic foraminifera that inhabited almost alone the most central areas of carbonate
platforms of the Mediterranean realm. A global schematic table (Fig. 18 ![]() , in
addition to Figs.
15
, in
addition to Figs.
15
![]() -
16
-
16
![]() -
17
-
17
![]() ) shows their regional
development. The
stratigraphic succession of this family seems to have been dominated by three
periods of emersion (one being documented, the others only inferred) that limit
the time distribution of most of these taxa. Thus it is doubtful that this
distribution can be used with certainty in other areas; for example the mention
of "Cuvillierinella? sp."
(possibly a Metacuvillierinella?) in
the Upper Maastrichtian of Iran is a hint of such a discrepancy (Schlagintweit & Rashidi, 2016).
) shows their regional
development. The
stratigraphic succession of this family seems to have been dominated by three
periods of emersion (one being documented, the others only inferred) that limit
the time distribution of most of these taxa. Thus it is doubtful that this
distribution can be used with certainty in other areas; for example the mention
of "Cuvillierinella? sp."
(possibly a Metacuvillierinella?) in
the Upper Maastrichtian of Iran is a hint of such a discrepancy (Schlagintweit & Rashidi, 2016).
Rhapydionininae
and
Pseudedomiinae subfamilies are summarily
considered here, either because the facts have been established previously, or
because the knowledge of them is still fragmentary. The Rhapydionininae subfamily,
summarily depicted in Figure 18 ![]() , is characterized mainly by its endoskeleton,
lacking the confluent structure, with a wide peripheral zone and a very early
unrolling trend (either cylindrical: Rhapydionina,
with small proloculus, or flabelliform: Fanrhapydionina,
with large proloculus). The milioline origin of this subfamily is indicated by
species of the R. dercourti group,
which are faintly streptospiral in the initial stages of both generations (Fleury, 2014). The Pseudedomiinae subfamily is not the subject of these
studies, being essentially represented by taxa in more Eastern regions. Besides,
nothing definite can be deduced from the minimal knowledge of the group (see
review in Mavrikas
et al., 1994), until the type of the type species is still
poorly known.
, is characterized mainly by its endoskeleton,
lacking the confluent structure, with a wide peripheral zone and a very early
unrolling trend (either cylindrical: Rhapydionina,
with small proloculus, or flabelliform: Fanrhapydionina,
with large proloculus). The milioline origin of this subfamily is indicated by
species of the R. dercourti group,
which are faintly streptospiral in the initial stages of both generations (Fleury, 2014). The Pseudedomiinae subfamily is not the subject of these
studies, being essentially represented by taxa in more Eastern regions. Besides,
nothing definite can be deduced from the minimal knowledge of the group (see
review in Mavrikas
et al., 1994), until the type of the type species is still
poorly known.
Cuvillierinellinae
subfamily (Fleury, 2016) is made up of six genera, including Cuvillierinella from which the others are clearly derived. But the
origin of this genus remains uncertain. However, the presence of Pseudonummoloculina
aff. kalantarii in our samples enables
the consideration of what would be such an ancestor and how it could be named.
We can try to imagine it from a simple principle and a few observations. The
principle is the independence of evolutionary characters, commonly observed
elsewhere in the family but quite obvious in the type population of Cuvillierinella
salentina (see De
Castro, 1988; Fleury, 2016), in which coexist the
well-known type and specimens of overall same appearance, either streptospiral
and devoid of partitions, or planispiral with a highly specialized endoskeleton.
The observations are those that can be performed on species described by various
authors under the name of Pseudonummoloculina.
These organisms change from streptospiral to planispiral stage during their
development. Some have a unique "notched" opening, others a double row
of openings (and complex combinations of these two rather indistinct types) and
some have no endoskeleton while others display rudimentary partitions. Varying
combinations of these characters defy the conventional criteria of
classification, each combination (changing during the ontogenetic development)
being likely to justify a generic or specific status according to the arbitrary
ordinary standards. These organisms are however in all cases isolated in time
and most often deprived of clearly identifiable descendants (with the exception
of P. pecheuxi Fourcade & Fleury, 2001, which is the origin of the
species of Praechubbina genus). They
probably do not constitute a unique group, but represent a step in an iterative
evolution, leading members of a group of Miliolacea to the Alveolinacea, as Reichel
(1936,
1937) and Pęcheux (2002) conceived it and as
Fleury and Fourcade
(1990) illustrated. We are thus led to apply the generic term
Pseudonummoloculina
to indicate a transitional state, that is to say an evolving stage. We are here
confronted with the fundamental difficulty of adapting a nomenclature of
creationistic essence to the perpetual reorganization of living organisms. It is
therefore futile to characterize by terminology the temporary results;
the term Pseudonummoloculina seems
better to express a vague and essentially unstable notion related to the
realities of evolution, ... at least as long as evolving networks of various
known stages are not identified.
The five genera derived from Cuvillierinella
have a wide range of morphology (Fleury, 2016, Fig. 13). Each genus is
characterized by a dominant evolutionary trend, variously associated with secondary independent features.
Metacuvillierinella
and Pseudochubbina genera are treated
superficially in this text and are probably derived from Cuvillierinella or its supposed close ancestor. Metacuvillierinella
is the closest, comparable in its young stage to typical streptospiral tests of C.
salentina, comprising an early well developed wide meshed endoskeleton. It
is distinguished by its adult advolute coiling without final uncoiling and its
very low dimorphism of generations, restricted to the initial coiling. Pseudochubbina
is linked to Cuvillierinella perisalentina,
also hesitating on its coiling mode ("Slow Axial Rotation" in Fleury, 2016), and sharing the "Scattered Secondary Chamberlets"
(Fleury, 2016), which are the only likely criteria for the origin of this genus.
Murciella, Sigalveolina
n. gen. and Cyclopseudedomia, are
reviewed here from the types and various new populations, which allows deepening and
renewing of the knowledge of them. They are all fully planispirally coiled in the A
generation and to varying degrees display an endoskeleton that previously was
called "helicoidal structure", here renamed 'confluent structure'.
Murciella is
very close to Cuvillierinella as
evidenced by the coexistence of specimens having characteristics of both types
in the population of the Cuvillierinella
type species. The existence of populations including both Cuvillierinella and Murciella,
without morphological intermediaries, as well as the existence of populations
containing only one or the other of these taxa emphasizes their independence.
The six various populations assigned to M.
cuvillieri, known from Spain to the Eastern Mediterranean sea, are likely to
belong to the same zone (CsB6a) and were never associated with species of the
overlying zone. These populations constitute a homogeneous set close to that of C.
salentina appearance, with a little wider range of the A proloculus and a
finer endoskeletal mesh. Tests of the B generation in several populations
suggest that the holotype of the type species represents this generation, which
appears to be less influenced by uncoiling than C.
salentina.
Sigalveolina n.
gen. has been long confused with Murciella,
too incompletely known to be distinguished before now. This new genus
characterizes younger levels (zone CsB6b). This study shows that the high degree
of confluent structure that gives its individuality coincides with a
morphological diversity illustrated by four distinct species, including a new
one, Sigalveolina reicheli. This new
species probably corresponds to the mythical material, known as
"Cosinella", formerly discovered by Schlumberger from the
Italo-Slovenian border. A tests of the four species are larger in size than
those of Murciella, at diameters of
proloculus of the same order of magnitude; they are only rarely uncoiled,
with the apparent exception of Sigalveolina reicheli
n. sp., which adopts this trend only at a very late development stage. B tests
also tend to be involute at a later stage, in contrast to the early uncoiling of
previous and next genera.
Cyclopseudedomia is
characterized by a very large A proloculus and an extremely reduced involute
stage resulting in a longer uncoiled uniserial stage. This outstanding
evolutionary trend is modulated in four species, including three already
described and a new one, C. mavrikasi
from the Italo-Slovenian border and Southern Italy. This trend is well marked in
tests of both generations. In A tests, the involute part never exceeds 1.5
whorls around a large proloculus, and the uncoiled part is either cylindrical or
flabelliform. B tests are mainly made of large evolute flabelliform flange,
reaching a discoidal stage in the type species only; these flanges are always
flat and increase in thickness very slightly towards the periphery.
Finally, it must be noted that
data in Figure 18 ![]() , reduced to the A tests, are imprecise
inasmuch as the genera
generally include the features of several species, which are distinguished by
nuances affecting their characters, but also because the time span of the genera
and the included species likely does not correspond to the entire
period of time covered by the zone in which they are recognized. This means that
Figure 18
, reduced to the A tests, are imprecise
inasmuch as the genera
generally include the features of several species, which are distinguished by
nuances affecting their characters, but also because the time span of the genera
and the included species likely does not correspond to the entire
period of time covered by the zone in which they are recognized. This means that
Figure 18 ![]() represents a state of schematic knowledge that can be improved only by
specific careful field studies.
represents a state of schematic knowledge that can be improved only by
specific careful field studies.
• Campanian-Maastrichtian;
• Alveolinacea;
• Rhapydioninidae;
• méditerranean domain;
• Murciella;
• Sigalveolina n. gen.;
• Cyclopseudedomia;
• new genus;
• new species.
Ce travail est le dernier d'une trilogie consacrée aux Rhapydioninidae campano-maastrichtiens du domaine méditerranéen, un groupe qui peuple principalement et presque ŕ lui seul la partie la plus interne des plates-formes carbonatées périadriatiques. Si ces plates-formes apparaissent comme le foyer des genres Cuvillierinella, Cyclopseudedomia, Metacuvillierinella, Murciella, Rhapydionina et Sigalveolina n. gen. qui y prospčrent, quelques-uns de ces taxons ont été accueillis en Espagne et aux abords du "Middle East", ŕ la faveur de l'installation plus ou moins fugace des "facičs d'émersion" qu'ils affectionnent. L'ensemble de ces études ne pourra cependant pas ętre considéré comme exhaustif, dans la mesure oů des genres tels que Pseudedomia Henson, 1948, ou Pseudochubbina De Castro, 1990, qui avoisinent ou parfois accompagnent les peuplements étudiés sont encore trop imprécisément connus pour faire l'objet de synthčses détaillées.
L'ensemble des données réunies ici ne devrait pas conduire ŕ de trop rapides conclusions. On sait que ces plates-formes correspondent actuellement ŕ des paysages souvent désolés qui n'attirent pas la curiosité du géologue et que les épaisses séries de carbonates apparemment monotones qui les constituent, souvent dépourvues de repčres évidents, peuvent décourager le chercheur. Cependant, en considération des riches résultats d'une étude trčs précise d'un de ces domaines, tel que celui du massif du Gavrovo en Grčce (Mavrikas, 1993), on peut supposer que bien des documents nous manquent encore. C'est donc en attendant avec confiance que de futures études ouvrent de nouveaux horizons que le présent dossier peut ętre présenté, certainement provisoire mais exhaustif des connaissances actuelles sur le sujet tel qu'il est limité.
Quoi qu'il en soit, ce travail comme les deux qu'ils l'ont précédé dans le męme esprit (Fleury, 2014, 2016), apporte une dimension nouvelle ŕ la connaissance de ce groupe. Fruit d'une recherche sur le terrain étalée sur plusieurs décennies et d'un travail de laboratoire plus long encore, il établit des comparaisons entre des populations isolées dans l'espace d'espčces le plus souvent connues jusqu'alors de leurs seules localités types et de quelques rares gisements superficiellement exploités. On en vient ainsi ŕ préciser la signification de chacune d'elles et ŕ poser les fondements d'une classification qui tend ŕ s'approcher de l'idéal naturel souhaitable.
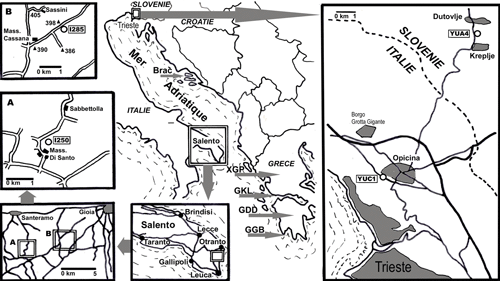
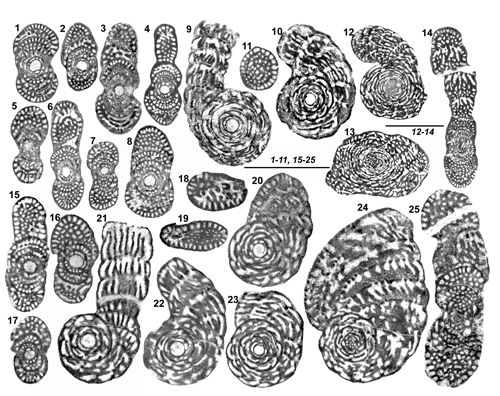
Les localités signalées par la suite sont en petit nombre, choisies en fonction de l'intéręt des populations qui s'y trouvent et, en conséquence, du grand nombre de préparations dont celles-ci ont fait l'objet. Celles de Grčce, comme les échantillons qui en proviennent, sont représentées par un groupe de 3 lettres
(Fig. 1 ![]() ), utilisé dans les précédentes publications de l'auteur, depuis 1980. Ce sont, d'est en ouest et du sud vers le nord, en Grčce : FPM Île d'Astypalia (Dodécančse), GGB Région de Pylos-Méthoni au sud-ouest du Péloponnčse, GDD Massif du Mainalon ŕ l'est du Péloponnčse, GKL Massif du Klokova au sud-ouest de la Grčce continentale, XGP Massif du Gavrovo, au cśur de la Grčce continentale. Les localités d'Italie méridionale et des confins Italo-Slovčnes sont également situés sur la
Figure 1
), utilisé dans les précédentes publications de l'auteur, depuis 1980. Ce sont, d'est en ouest et du sud vers le nord, en Grčce : FPM Île d'Astypalia (Dodécančse), GGB Région de Pylos-Méthoni au sud-ouest du Péloponnčse, GDD Massif du Mainalon ŕ l'est du Péloponnčse, GKL Massif du Klokova au sud-ouest de la Grčce continentale, XGP Massif du Gavrovo, au cśur de la Grčce continentale. Les localités d'Italie méridionale et des confins Italo-Slovčnes sont également situés sur la
Figure 1 ![]() ; celles d'Espagne sont rappelées dans le texte ŕ partir des données d'Éric Fourcade.
; celles d'Espagne sont rappelées dans le texte ŕ partir des données d'Éric Fourcade.
|
Figure 1 : Localités citées dans ce texte. En Grčce, les diverses localités sont représentées par un groupe de 3 lettres, toujours identique pour les męmes localités depuis le travail de Fleury (1980). Sont également décrites dans le texte les localités des confins italo-slovčnes YUA4 et YUC1 (d'aprčs Bignot, 1972) et d'Italie méridionale I250 (A) et I285 (B). Les localités d'Espagne, non situées ici, sont rapportées dans le texte, d'aprčs Fourcade (1966) et Hamaoui & Fourcade (1973). |
Le matériel examiné provient dans tous les cas de calcaires massifs dont les organismes ne peuvent ętre dégagés sans grands dommages. L'étude de sections en lames minces est donc le seul moyen d'accéder ŕ leur connaissance. Ces lames ont été dans un premier temps effectuées au hasard, ce qui est parfois suffisant pour en connaître les principaux caractčres, notamment ceux de l'endosquelette. Mais les particularités de l'enroulement ne sont pas ordinairement déductibles des sections quelconques, spécialement pour les tests de génération B, parfois trčs rares. Des recherches plus précises sont souvent nécessaires ; elles commencent sur le terrain, oů les échantillons doivent ętre soigneusement sélectionnés et oů des éclats de roche révélateurs doivent ętre scrutés et récoltés. On doit également explorer longuement des plaquettes sciées selon des plans variés, progressivement usées jusqu'ŕ l'apparition éventuelle de sections éclairantes. C'est un long et fastidieux travail, qui seul permet de disposer des connaissances indispensables ŕ la compréhension des relations entre les divers taxons examinés.
Le matériel réuni au cours des années d'activité académique de l'auteur est compris dans une collection d'environ 8000 lames minces conservée par le Département des Sciences de la Terre de l'Université des Sciences et Techniques de Lille (USTL), ŕ Villeneuve d'Ascq. L'identité de chaque lame est assurée par un numéro d'échantillon (toujours indiqué dans les publications oů elle a fait l'objet d'une figuration) et un numéro de fabrication inclus dans la préparation. En outre, des numéros de collection sont attribués aux échantillons et portés sur les lames ; ils sont indiqués dans les notes de l'auteur depuis longtemps et permettent une recherche systématique aisée dans l'ensemble de la collection, ŕ partir des diverses publications.
On notera que c'est par erreur que, ŕ la suite de confusions et de malentendus, cette collection a été annoncée (Fleury, 2014, p. 175) comme devant ętre confiée au Museum National d'Histoire Naturelle de Paris ; elle restera la propriété de l'USTL. On confirme ici cependant que les collections de J. Cuvillier, M. Neumann et É. Fourcade seront déposées auprčs du MNHN.
La Figure 2 ![]() présente les principaux traits de l'architecture des Rhapydioninidae, ŕ partir de deux exemples développés dans ce qui suit. Les tests de génération mégalosphérique (A) sont seuls traités sur cette figure mais on notera que mis ŕ part leur parties népioniques (de type miliolin : quinquéloculin ou streptospiralé), les tests microsphériques (B) présentent le męme endosquelette que ceux qui sont illustrés. On peut en effet distinguer, suivant Reichel
(1936), un exosquelette
"qui forme carapace" d'un endosquelette "constitué des dépôts internes", bien que cette distinction toute théorique
ne soit essentiellement utile que pour l'exposé.
présente les principaux traits de l'architecture des Rhapydioninidae, ŕ partir de deux exemples développés dans ce qui suit. Les tests de génération mégalosphérique (A) sont seuls traités sur cette figure mais on notera que mis ŕ part leur parties népioniques (de type miliolin : quinquéloculin ou streptospiralé), les tests microsphériques (B) présentent le męme endosquelette que ceux qui sont illustrés. On peut en effet distinguer, suivant Reichel
(1936), un exosquelette
"qui forme carapace" d'un endosquelette "constitué des dépôts internes", bien que cette distinction toute théorique
ne soit essentiellement utile que pour l'exposé.
Les tests sont limités par une paroi calcaire imperforée, dénommée muraille, dont les inflexions périodiques, nommées septes, limitent les loges successives. Chaque septe est ŕ l'origine une face orale perforée d'ouvertures (Fig. 1.b-c ![]() ), laquelle ne devient ŕ proprement parler septe que lorsqu'elle est recouverte par la loge suivante. C'est ce qui pourrait justifier que certains auteurs aient suggéré de nommer différemment ces perforations passant au cours du temps d'une fonction de communication avec l'extérieur ŕ une communication entre loges successives. Il ne s'ensuit cependant aucun changement morphologique et cette proposition n'est pas retenue, ŕ l'exemple de la bouche humaine qui ne change pas de nom lors du passage de la fonction d'élocution ŕ celle d'absorption de nourriture, en particulier. On nomme ouvertures primaires celles qui, situées ŕ la périphérie du septe, correspondent et donnent naissance aux logettes primaires de la loge suivante et ouvertures secondaires celles qui, situées dans la partie centrale du septe, correspondent et donnent naissance aux logettes secondaires. On retiendra que si les ouvertures secondaires partagent le męme axe et sont en parfaite continuité avec les logettes qui les suivent, les ouvertures primaires présentent un axe oblique sur celui des logettes correspondantes et sont décalées par rapport ŕ cet axe comme en témoignent les bonnes sections convenablement orientées, telles que celles des
Figures 2.4, .11
), laquelle ne devient ŕ proprement parler septe que lorsqu'elle est recouverte par la loge suivante. C'est ce qui pourrait justifier que certains auteurs aient suggéré de nommer différemment ces perforations passant au cours du temps d'une fonction de communication avec l'extérieur ŕ une communication entre loges successives. Il ne s'ensuit cependant aucun changement morphologique et cette proposition n'est pas retenue, ŕ l'exemple de la bouche humaine qui ne change pas de nom lors du passage de la fonction d'élocution ŕ celle d'absorption de nourriture, en particulier. On nomme ouvertures primaires celles qui, situées ŕ la périphérie du septe, correspondent et donnent naissance aux logettes primaires de la loge suivante et ouvertures secondaires celles qui, situées dans la partie centrale du septe, correspondent et donnent naissance aux logettes secondaires. On retiendra que si les ouvertures secondaires partagent le męme axe et sont en parfaite continuité avec les logettes qui les suivent, les ouvertures primaires présentent un axe oblique sur celui des logettes correspondantes et sont décalées par rapport ŕ cet axe comme en témoignent les bonnes sections convenablement orientées, telles que celles des
Figures 2.4, .11 ![]() ,
7.5
,
7.5 ![]() ,
9.11-13
,
9.11-13 ![]() ,
11.13
,
11.13 ![]() ,
14.9-10
,
14.9-10 ![]() . On notera comme caractčre de signification douteuse l'existence chez deux espčces du genre Cyclopseudedomia
(Figs. 11.8, .12
. On notera comme caractčre de signification douteuse l'existence chez deux espčces du genre Cyclopseudedomia
(Figs. 11.8, .12 ![]() ,
13.8
,
13.8 ![]() ) d'une fine costulation externe, inhabituelle chez les Alveolinacea. Bien que ce trait semble propre ŕ ce genre, sa signification demeure douteuse dans la mesure oů il reste trčs discret et donc sensible aux altérations diagénétiques qui peuvent le rendre inobservable chez d'autres taxons.
) d'une fine costulation externe, inhabituelle chez les Alveolinacea. Bien que ce trait semble propre ŕ ce genre, sa signification demeure douteuse dans la mesure oů il reste trčs discret et donc sensible aux altérations diagénétiques qui peuvent le rendre inobservable chez d'autres taxons.
Les tests A comprennent un proloculus (premičre loge) de section ordinairement circulaire communicant avec la loge suivante par le flexostyle (développé sur la surface du proloculus ŕ la maničre du germe de marron, selon l'heureuse expression de Reichel,
1936). Chez certains de ces tests, les loges suivantes s'organisent selon une spire dont le caractčre streptospiralé peut-ętre limité aux premiers tours (Cuvillierinella par exemple) ou persister dans les stades adultes (Pseudochubbina, par exemple) ; chez d'autres, entičrement planispiralés (les espčces qui sont traitées ici), les loges suivant le proloculus constituent un ensemble nautiloďde subsphérique ŕ aplati qui peut constituer la totalité du test. Les tests B présentent une trčs petite loge, rarement observée, suivie par un ensemble de petites loges streptospiralées ou d'allure quinquéloculine. Dans les deux générations, les stades adultes comprennent éventuellement une partie rectiligne unisériée (abréviée PRU par la suite), évolute, soit cylindrique
(Fig. 2.1 ![]() ), soit flabelliforme
(Fig. 2.6, .9
), soit flabelliforme
(Fig. 2.6, .9 ![]() : les 5 derničres loges), dans laquelle chaque loge n'est en contact qu'avec celle qui la précčde et celle qui la suit ; un cas extręme est fourni par Cyclopseudedomia smouti
(Fig. 11.10
: les 5 derničres loges), dans laquelle chaque loge n'est en contact qu'avec celle qui la précčde et celle qui la suit ; un cas extręme est fourni par Cyclopseudedomia smouti
(Fig. 11.10 ![]() ) dont les loges terminales des tests B sont annulaires. Un stade intermédiaire, dit pseudoévolute, correspond au passage de la partie involute ŕ la partie évolute, lorsque le pas de la spire s'accroît rapidement, alors que les loges restent au contact de la partie involute
(Fig. 2.9
) dont les loges terminales des tests B sont annulaires. Un stade intermédiaire, dit pseudoévolute, correspond au passage de la partie involute ŕ la partie évolute, lorsque le pas de la spire s'accroît rapidement, alors que les loges restent au contact de la partie involute
(Fig. 2.9 ![]() : 5 ou 6 loges précédant les 5 derničres) ; ce stade est plus ou moins bref mais peut persister longuement chez certains tests adultes (Cuvillierinella, tests B, par exemple : voir Fleury, 2016, Fig. 14). Pour mémoire, on évoquera encore l'enroulement advolute, cas particulier de l'enroulement involute dans lequel les tours successifs ne se recouvrent que partiellement (Metacuvillierinella in Fleury, 2016, Figs. 10-11).
: 5 ou 6 loges précédant les 5 derničres) ; ce stade est plus ou moins bref mais peut persister longuement chez certains tests adultes (Cuvillierinella, tests B, par exemple : voir Fleury, 2016, Fig. 14). Pour mémoire, on évoquera encore l'enroulement advolute, cas particulier de l'enroulement involute dans lequel les tours successifs ne se recouvrent que partiellement (Metacuvillierinella in Fleury, 2016, Figs. 10-11).
Structure coaxiale (Fig. 2.12-14 ![]() ). On nomme ainsi la disposition ordinaire, commune ŕ tous les taxons de la famille, des logettes qui divisent les loges. Ces volumes tubulaires ménagés au sein des
"dépôts internes" divisent le protoplasme de chaque loge en colonnes isolées, parallčles entre elles (coaxiales) ŕ l'origine, qui débouchent dans un espace indivis ménagé ŕ proximité du septe, l'espace préseptal. Cet ensemble constitue
l'unique élément fondamental caractérisant les Alveolinacea en général et les Rhapydioninidae en particulier.
). On nomme ainsi la disposition ordinaire, commune ŕ tous les taxons de la famille, des logettes qui divisent les loges. Ces volumes tubulaires ménagés au sein des
"dépôts internes" divisent le protoplasme de chaque loge en colonnes isolées, parallčles entre elles (coaxiales) ŕ l'origine, qui débouchent dans un espace indivis ménagé ŕ proximité du septe, l'espace préseptal. Cet ensemble constitue
l'unique élément fondamental caractérisant les Alveolinacea en général et les Rhapydioninidae en particulier.
Les logettes s'organisent en deux ensembles. Les unes, formant une seule couche adjacente ŕ la paroi, d'axes toujours parallčles ŕ l'axe de la loge sont dites logettes primaires (les premičres apparues et parfois demeurant les seules divisions des loges) ; elles sont limitées par les cloisonnettes qui joignent les septes successifs
(Figs. 5.14 ![]() , 11.14
, 11.14 ![]() ,
12.6
,
12.6 ![]() ), dont la section est triangulaire ŕ la marge de l'espace préseptal (pour les plus lisibles exemples, voir
Figs. 9.1, .6
), dont la section est triangulaire ŕ la marge de l'espace préseptal (pour les plus lisibles exemples, voir
Figs. 9.1, .6 ![]() ,
10.14
,
10.14 ![]() pour Sigalveolina n. gen. ;
Figs. 11.12, .15
pour Sigalveolina n. gen. ;
Figs. 11.12, .15 ![]() ,
12.16
,
12.16 ![]() ,
13.14
,
13.14 ![]() ,
14.26
,
14.26 ![]() pour Cyclopseudedomia et Fleury, 2014, Fig.
1H pour Rhapydionina); ces logettes caractérisent une zone marginale. Les autres, dites logettes secondaires (apparaissant en second, parfois absentes, et accessoires, ŕ l'origine, mais vouées dans bien des cas ŕ la structure confluente décrite plus loin) sont dispersées dans une masse plus ou moins compacte dénommée endosquelette central. L'espace préseptal męme est occupé par des piliers préseptaux apparaissant lorsque l'endosquelette central est bien développé (les piliers sont donc une structure annexe de celui-ci) et partiellement divisé dans sa partie marginale par les cloisonnettes qui s'y prolongent. On rappellera que l'expression
"piliers résiduels", parfois utilisée, est issue d'une théorie depuis longtemps abandonnée selon laquelle les Rhapydioninidés seraient issus de Soritidés dont les piliers interseptaux auraient en partie fusionné pour constituer l'endosquelette central.
pour Cyclopseudedomia et Fleury, 2014, Fig.
1H pour Rhapydionina); ces logettes caractérisent une zone marginale. Les autres, dites logettes secondaires (apparaissant en second, parfois absentes, et accessoires, ŕ l'origine, mais vouées dans bien des cas ŕ la structure confluente décrite plus loin) sont dispersées dans une masse plus ou moins compacte dénommée endosquelette central. L'espace préseptal męme est occupé par des piliers préseptaux apparaissant lorsque l'endosquelette central est bien développé (les piliers sont donc une structure annexe de celui-ci) et partiellement divisé dans sa partie marginale par les cloisonnettes qui s'y prolongent. On rappellera que l'expression
"piliers résiduels", parfois utilisée, est issue d'une théorie depuis longtemps abandonnée selon laquelle les Rhapydioninidés seraient issus de Soritidés dont les piliers interseptaux auraient en partie fusionné pour constituer l'endosquelette central.
Les termes de "planchers" et "couche basale", parfois utilisés pour décrire l'aspect de l'endosquelette central nécessitent d'ętre reconsidérés. On évoquera également les aspects variés de la disposition des logettes secondaires au sein de l'endosquelette.
Les planchers ("lames parallčles ŕ la surface ; ils délimitent les couches de logettes" selon Reichel,
1936) correspondent ŕ des structures que bien des auteurs, parmi lesquels celui du présent travail, ont cru discerner chez de nombreux Alveolinidae ŕ tests fusiformes et chez certains Rhapydioninidae (particuličrement dans les sections de type équatorial, voir par exemple
Figs. 2.4, .15 ![]() ,
5.11-12
,
5.11-12 ![]() ,
7.5
,
7.5 ![]() ,
9.11-13
,
9.11-13 ![]() , ou encore in Fleury, 2016, Fig. 6.2). Pourtant, dans le cas général, et en sections axiales, bien des irrégularités de détail se présentent et de nombreux exemples remarquables semblent de nature ŕ discréditer cette conception, comme en témoignent les
Fig. 2.16-24
, ou encore in Fleury, 2016, Fig. 6.2). Pourtant, dans le cas général, et en sections axiales, bien des irrégularités de détail se présentent et de nombreux exemples remarquables semblent de nature ŕ discréditer cette conception, comme en témoignent les
Fig. 2.16-24 ![]() . Ainsi, des genres aussi divers que Sellialveolina Colalongo,
1963
(Fig. 2.16
. Ainsi, des genres aussi divers que Sellialveolina Colalongo,
1963
(Fig. 2.16 ![]() ), Chubbina Robinson,
1968
(Fig. 2.17-18
), Chubbina Robinson,
1968
(Fig. 2.17-18 ![]() ), Murciella
(Fig. 6.6, 8
), Murciella
(Fig. 6.6, 8 ![]() ) et Cyclopseudedomia
(Figs. 11.15
) et Cyclopseudedomia
(Figs. 11.15 ![]() ,
13.15
,
13.15 ![]() ,
14.20
,
14.20 ![]() ) montrent-ils, soit dans les derničres loges de la partie involute, soit dans celles des parties évolutes, une organisation anarchique des logettes qui ne peut ętre décrite en termes de planchers. Il en est de męme chez le genre Rhapydionina Stache,
1913, dont la partie terminale unisériée présente des configurations trčs diverses, permettant d'en distinguer plusieurs types :
) montrent-ils, soit dans les derničres loges de la partie involute, soit dans celles des parties évolutes, une organisation anarchique des logettes qui ne peut ętre décrite en termes de planchers. Il en est de męme chez le genre Rhapydionina Stache,
1913, dont la partie terminale unisériée présente des configurations trčs diverses, permettant d'en distinguer plusieurs types :
les logettes secondaires sont peu nombreuses, irréguličrement dispersées et totalement indépendantes des primaires, en fonction de la coalescence des cloisonnettes ŕ des distances variables de la muraille
(Fig. 2.19-21, .24 ![]() );
);
les logettes secondaires sont disposées en quinconce par rapport aux primaires, leur limite formant une ligne réguličrement indentée
(Fig. 2.22 ![]() ) ;
) ;
les logettes secondaires, noyées dans la masse centrale, se trouvent ŕ des distances trčs variables de la muraille, apparaissant parfois comme de simples subdivisions distales des primaires
(Fig. 2.23 ![]() ).
).
Ces observations donnent entičrement raison ŕ Hamaoui & Fourcade (1973), qui niaient l'existence de planchers chez les Rhapydioninidae. Elles semblent permettre de soutenir l'hypothčse d'une élaboration de l'endosquelette central des Rhapydioninidae ŕ partir de la coalescence des cloisonnettes, ŕ l'endroit oů, géométriquement, elles se rencontrent.
La couche basale correspond ŕ une notion introduite également par Reichel (1936, "dépôt plus ou moins épais qui s'étend sur la surface du tour précédent et constitue le revętement interne des loges"). Il existe effectivement toujours chez les Alveolinacea un dépôt qui répond ŕ cette définition mais il n'est généralement pas évoqué, tant il est banal et peu épais; il peut ętre qualifié de "couche basale au sens large". On lui a attribué une signification particuličre dans les cas oů ce dépôt est épais et donc trčs remarquable, dans deux types de configurations bien différents.
Le plus souvent (parfois sous le nom de "flosculinisation") ce terme caractérise l'épaississement apparaissant chez certaines Alveolina et autres genres cénozoďques : il est parfois également réparti dans toute la loge, structuré de stries qui témoigneraient de son accroissement progressif (voir Hottinger, 1960) et parfois parcouru de canaux irréguliers ("logettes supplémentaires") ; ces canaux ne débouchent pas dans l'espace préseptal (limité ŕ une partie du septe), oů n'accčdent que les logettes primaires.
Par extension, le męme terme a été utilisé pour décrire un épaississement qui se développe exclusivement vers les pôles des tests B du genre Subalveolina Reichel, 1936, parcouru de canaux plus ou moins bien organisés débouchant dans l'espace préseptal, lequel a le męme développement que le septe. Ces canaux sont plutôt irréguliers et dispersés chez S. dordonica Reichel, 1936 (Pl. IV.2 de cet auteur; voir aussi Vicedo et al., 2009, Fig. 5.3), mais "plus densément répartis (…) comme chez Praealveolina" chez S. perebaskini Reichel, 1953.
Les épaississements observés chez Alveolina et chez Subalveolina présentent donc entre eux une analogie de position mais ne sont pas homologues. La notion de couche basale au sens restreint qui s'applique ŕ Alveolina ne semble donc pas plus adaptée ŕ caractériser l'épaississement de Subalveolina que l'endosquelette central des Rhapydioninidae.
Par ailleurs, Reichel (1953) ne voit qu'une différence de détail entre l'irrégularité des logettes secondaires de S. dordonica ŕ la régularité de celles de S. perebaskini ou de Praealveolina, qui ne lui semblent donc pas de nature différente. Ces deux aspects ont leurs équivalents chez les Rhapydioninidae. D'une part, le dispositif SSC (Scattered secondary chamberlets in Fleury, 2016), dans lequel la distance entre logettes est plus grande que leur diamčtre, connu chez Pseudochubbina, Cuvillierinella perisalentina Fleury, 2016 et que l'on retrouve chez Rhapydionina
(Fig. 2.19-23 ![]() ) et Cyclopseudedomia
(Figs. 2.8
) et Cyclopseudedomia
(Figs. 2.8 ![]() ,
11.8, .11, .15
,
11.8, .11, .15 ![]() ,
12.11, .16
,
12.11, .16 ![]() ,
13.15
,
13.15 ![]() ) correspond ŕ celui de S. dordonica. D'autre part, le dispositif plus ordinaire, dans lequel les logettes sont séparées par une paroi mince, comme chez Sellialveolina
(Fig. 2.16
) correspond ŕ celui de S. dordonica. D'autre part, le dispositif plus ordinaire, dans lequel les logettes sont séparées par une paroi mince, comme chez Sellialveolina
(Fig. 2.16 ![]() ), Chubbina
(Fig. 2.17-18
), Chubbina
(Fig. 2.17-18 ![]() ), Murciella
(Figs. 5.4
), Murciella
(Figs. 5.4 ![]() ,
6.8
,
6.8 ![]() ,
7.7
,
7.7 ![]() ) et Sigalveolina
(Figs. 8.3, .19
) et Sigalveolina
(Figs. 8.3, .19 ![]() ,
10.13-14
,
10.13-14 ![]() ) correspond ŕ celui de S. perebaskini. Il n'y a donc pas lieu de distinguer fondamentalement ces deux dispositifs, dont on ne vient d'évoquer que les cas les plus évidents, entre lesquels on peut trouver des exemples moins tranchés. Mais il reste que le dispositif SSC peut servir ŕ caractériser certains taxons, dont les plus notables sont Pseudochubbina et Cyclopseudedomia.
) correspond ŕ celui de S. perebaskini. Il n'y a donc pas lieu de distinguer fondamentalement ces deux dispositifs, dont on ne vient d'évoquer que les cas les plus évidents, entre lesquels on peut trouver des exemples moins tranchés. Mais il reste que le dispositif SSC peut servir ŕ caractériser certains taxons, dont les plus notables sont Pseudochubbina et Cyclopseudedomia.
En conclusion, ni la théorie des planchers, ni celle d'une équivalence de l'endosquelette central des Rhapydioninidae avec la couche basale prise dans la signification qu'elle présente chez Alveolina ne semblent devoir s'appliquer ŕ cette famille. L'endosquelette central des Rhapydioninidae serait issu du développement des cloisonnettes et serait l'homologue de ce qui a été nommé par extension (et ŕ défaut d'autres exemples connus ŕ l'époque) "couche basale" chez Subalveolina.
|
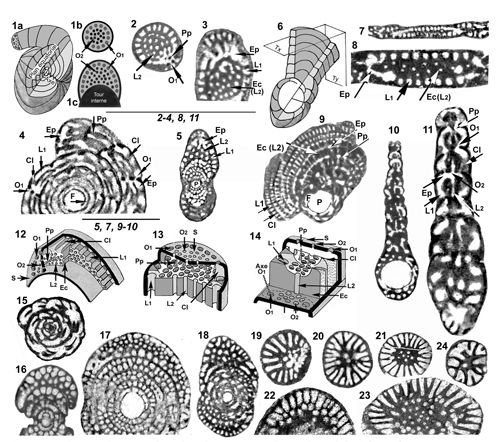
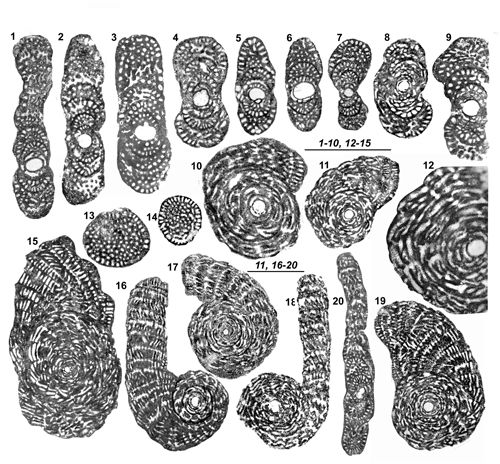
Figure
2 :
1-14 :
Architecture du test et endosquelette des genres Murciella et Cyclopseudedomia (Tests
A). 1-5 : Genre Murciella
ŕ l'exemple de M. cuvillieri.
1a : Aspect extérieur d'un test théorique en partie disséqué montrant
le mode d'enroulement et situant les éléments de description ; 1b et 1c :
Aspect de la face aperturale d'une loge de la partie déroulée finale et
d'une loge de la partie involute (noter que les ouvertures primaires comme les
logettes primaires n'existent pas au contact du tour interne). 2 :
Section transverse d'une partie rectiligne unisériée (dite
PRU par la suite) cylindrique. 3 : Partie de section axiale de deux
loges successives (voir l'entičre section Fig. 6.4 |
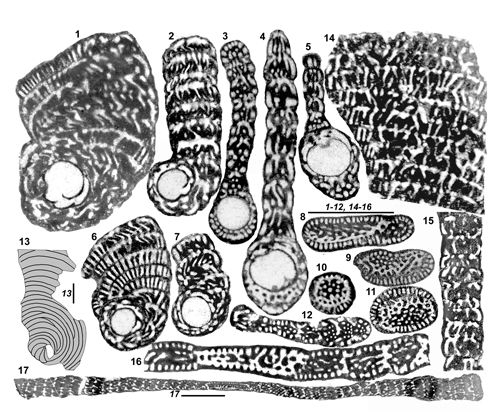
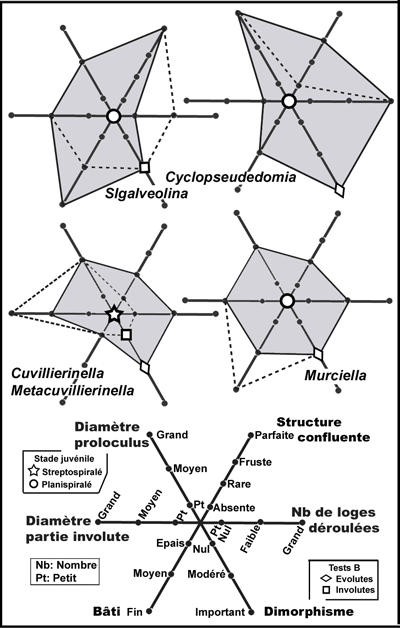
Structure confluente (Fig. 3 ![]() ). Une importante variation de la structure endosquelettique coaxiale décrite ci-dessus a été mise en évidence il y a presque quatre décennies (Fleury,
1979a) sous le nom de
"structure hélicoďdale". Il s'agissait d'un trait jamais observé auparavant chez les Alveolinacea, consistant en une apparente désorganisation des logettes secondaires, perdant le parallélisme habituel de leur axe avec celui des logettes primaires. C'est une espčce en particulier, ("Murciella" renzi, ici attribuée ŕ Sigalveolina n. gen.) qui devait fournir la clé de compréhension de ce caractčre : les logettes de fin diamčtre comprises dans un test trčs aplati créaient des conditions favorables ŕ l'observation en section plane de leur organisation en deux jeux sensiblement orthogonaux orientés ŕ 45° de l'axe des logettes primaires
(Fig. 3.5
). Une importante variation de la structure endosquelettique coaxiale décrite ci-dessus a été mise en évidence il y a presque quatre décennies (Fleury,
1979a) sous le nom de
"structure hélicoďdale". Il s'agissait d'un trait jamais observé auparavant chez les Alveolinacea, consistant en une apparente désorganisation des logettes secondaires, perdant le parallélisme habituel de leur axe avec celui des logettes primaires. C'est une espčce en particulier, ("Murciella" renzi, ici attribuée ŕ Sigalveolina n. gen.) qui devait fournir la clé de compréhension de ce caractčre : les logettes de fin diamčtre comprises dans un test trčs aplati créaient des conditions favorables ŕ l'observation en section plane de leur organisation en deux jeux sensiblement orthogonaux orientés ŕ 45° de l'axe des logettes primaires
(Fig. 3.5 ![]() ). Le plus remarquable de ce dispositif était l'interpénétration de ces logettes ŕ leurs croisements
(Fig. 3.3-14
). Le plus remarquable de ce dispositif était l'interpénétration de ces logettes ŕ leurs croisements
(Fig. 3.3-14 ![]() ), créant ainsi de nouvelles communications protoplasmiques au sein de la loge, concurrentes de celles qui sont ordinairement limitées ŕ l'espace préseptal. Comme certaines sections semblaient montrer que ces logettes s'organisaient en couches adjacentes bien délimitées
(Fig. 3.6
), créant ainsi de nouvelles communications protoplasmiques au sein de la loge, concurrentes de celles qui sont ordinairement limitées ŕ l'espace préseptal. Comme certaines sections semblaient montrer que ces logettes s'organisaient en couches adjacentes bien délimitées
(Fig. 3.6 ![]() ), il était possible d'envisager qu'au sein d'une loge de type cylindrique, les logettes secondaires décrivent deux hélices contrariées de męme axe que la loge, ce qui semblait propre ŕ supporter le nom choisi alors. Mais il est vrai que les loges cylindriques sont rares chez Sigalveolina et il
apparaît probable que l'organisation ne soit pas aussi réguličre que supposée, si bien que fonder un nom sur une image presque virtuelle pouvait sembler peu avisé. C'est ainsi que certains auteurs, bien que n'ayant rien apporté ŕ sa connaissance, ont cru devoir abandonner
le terme de "structure hélicoďdale", jugé probablement trop théorique. On conviendra qu'il vaut mieux se fonder sur le fait fondamental, qui réside dans la communication établie entre les logettes secondaires d'une męme loge, dont résulte une révolution dans la gestion du protoplasme de la loge. On songera dans ces conditions ŕ l'avantage que constituent ces confluences pour les symbiotes, dont on sait qu'ils présentent des rendements optimaux dans d'étroits domaines de luminosité (Lee & Anderson,
1991, par exemple) et qui sont donc dans ce dispositif susceptibles d'adapter constamment leur position en fonction de leurs préférences. Il s'agirait d'un facteur décisif du développement du groupe, qui concerne ŕ des degrés divers les trois genres examinés ci-dessous.
), il était possible d'envisager qu'au sein d'une loge de type cylindrique, les logettes secondaires décrivent deux hélices contrariées de męme axe que la loge, ce qui semblait propre ŕ supporter le nom choisi alors. Mais il est vrai que les loges cylindriques sont rares chez Sigalveolina et il
apparaît probable que l'organisation ne soit pas aussi réguličre que supposée, si bien que fonder un nom sur une image presque virtuelle pouvait sembler peu avisé. C'est ainsi que certains auteurs, bien que n'ayant rien apporté ŕ sa connaissance, ont cru devoir abandonner
le terme de "structure hélicoďdale", jugé probablement trop théorique. On conviendra qu'il vaut mieux se fonder sur le fait fondamental, qui réside dans la communication établie entre les logettes secondaires d'une męme loge, dont résulte une révolution dans la gestion du protoplasme de la loge. On songera dans ces conditions ŕ l'avantage que constituent ces confluences pour les symbiotes, dont on sait qu'ils présentent des rendements optimaux dans d'étroits domaines de luminosité (Lee & Anderson,
1991, par exemple) et qui sont donc dans ce dispositif susceptibles d'adapter constamment leur position en fonction de leurs préférences. Il s'agirait d'un facteur décisif du développement du groupe, qui concerne ŕ des degrés divers les trois genres examinés ci-dessous.
|
Figure 3
: La structure
confluente. 1 : Figure schématique
idéale représentant une portion de loge cylindrique dans laquelle les logettes
primaires (L1) conservent un axe parallčle ŕ celui de la loge, alors que les
logettes secondaires (L2) décrivent deux hélices contrariées obliques sur
l'axe de la loge, ce qui semblait justifier l'ancienne appellation "structure
hélicoďdale" ; l'interpénétration des deux ensembles de
logettes secondaires n'est pas représentée. 2 : Figure schématique représentative de la męme structure
dans une loge aplatie, dans laquelle les logettes secondaires sont organisées
selon des plans ; elles peuvent sembler obliques sur les parois de la loge,
en fonction de l'orientation du plan de section ; l'interpénétration
des logettes secondaires des couches adjacentes n'est pas représentée (Fleury &
Fourcade, 1987). 3-4 :
Modčle matériel d'une partie de loge cylindrique (3) et section (4) de ce
modčle par un plan dont la trace est figurée sur le côté de 3 ;
l'interpénétration des logettes secondaires des différentes couches donne
naissance ŕ des nouvelles communications intraloculaires, dénommées "stolons"
(St). 5-14 : Exemples concrets
de la structure confluente. 5 : Dessin de section de 2 loges de Sigalveolina
renzi (in Fleury, 1979a). 6 : Partie de section de S.
aff. renzi, montrant clairement la
disposition des logettes secondaires en couches parallčles ŕ la paroi de la
loge (GKL414). 7 : section d'un grand test de Cyclopseudedomia ? n. sp. (voir
Fig. 14.29-37 |
L'expérience montre que, au sein d'un groupe particulier tel que celui qui est étudié ici, il est vain de chercher ŕ classifier les caractčres morphologiques en catégories spécifiques ou génériques définies a priori, ou du moins ŕ partir d'une expérience trčs limitée. Nous montrerons que les éléments morphologiques n'ont pas de signification intrinsčque : tel caractčre susceptible de distinguer des espčces pourra se retrouver dans divers genres et inversement tel caractčre permettant de distinguer des genres se retrouvera dans diverses espčces. Une telle complexité est liée au fait que les divers caractčres évoluent indépendamment les uns des autres. Il apparaît plutôt que ce sont les associations de caractčres qui permettent d'entrevoir les tendances évolutives dans leur complexité et de les distinguer dans une perspective d'ensemble. Mais ce mode opératoire nécessite une bonne compréhension globale du groupe considéré et n'est pas ŕ la portée d'une étude superficielle rassemblant des éléments divers hâtivement réunis.
Les critčres qui peuvent servir ŕ l'identification des espčces et des genres sont les suivants :
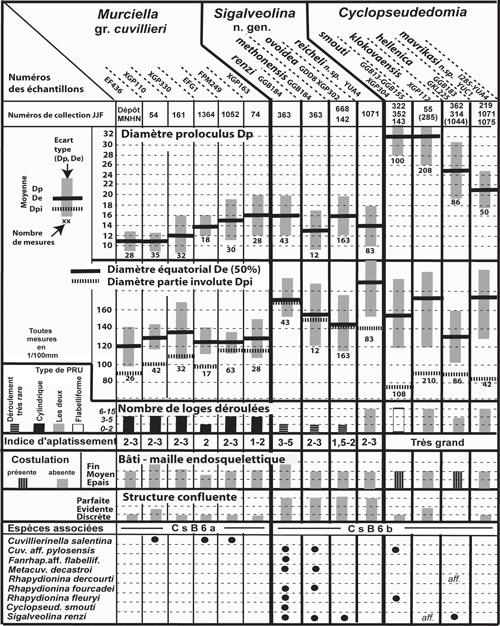
Le diamčtre du proloculus permet en principe de caractériser statistiquement chaque population et nécessite d'ętre toujours exprimé.
Dans ce travail, comme dans les précédents de l'auteur, c'est toujours le
diamčtre interne qui est mesuré (sans qu'il soit tenu compte de l'épaisseur
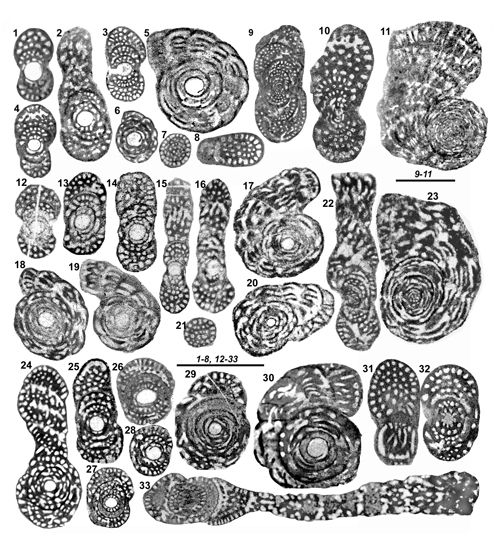
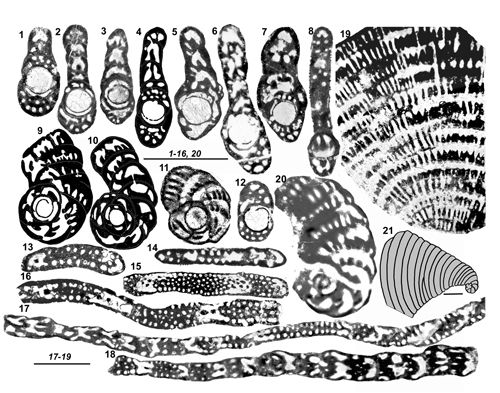
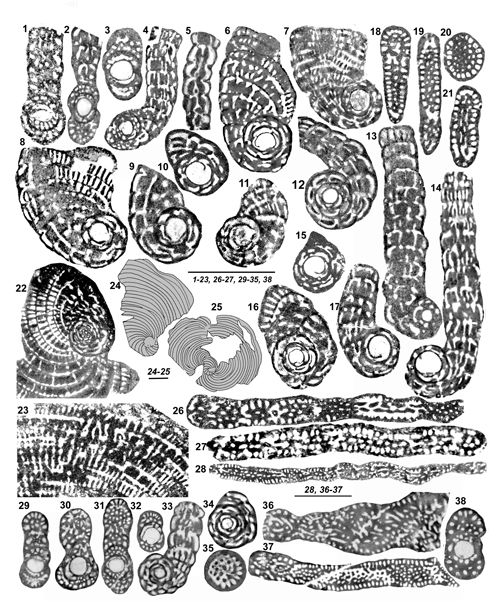
de la paroi). On ne peut cependant pas accorder ŕ ce critčre une importance primordiale chez des organismes répandus sur de vastes territoires et pendant une période de temps relativement importante, sauf lorsqu'il semble indiquer une tendance générale. Par exemple on notera le cas de Murciella gr. cuvillieri
(Figs. 15 ![]() - 16
- 16 ![]() ) représentée par des populations de morphologie relativement homogčnes dont les variations de taille du proloculus sont sans rapport avec les relations de proximité ou d'éloignement des diverses localités dont ils proviennent. On citera encore les variations autour du type signalées dans la description de Sigalveolina renzi (Fig.
8
) représentée par des populations de morphologie relativement homogčnes dont les variations de taille du proloculus sont sans rapport avec les relations de proximité ou d'éloignement des diverses localités dont ils proviennent. On citera encore les variations autour du type signalées dans la description de Sigalveolina renzi (Fig.
8 ![]() ), relativement homogčnes par l'ensemble de leurs caractčres, dont les tailles de proloculus sont trčs diverses. A l'inverse, on rappellera que dans le cas de Rhapydionina gr. liburnica, il a paru utile (Fleury, 2014) de distinguer les populations de Grčce de celle du type d'Istrie, tant la différence de taille du proloculus entre les deux régions apparaissait systématique. Un cas non élucidé se présente chez le genre Sigalveolina : l'examen des tests A de S. ovoidea et S. reicheli n. sp. semble montrer que les grands proloculus correspondent ŕ de (relatifs) petits tests alors que, inversement, les plus petits proloculus correspondent aux plus grands tests (voir
Fig. 16
), relativement homogčnes par l'ensemble de leurs caractčres, dont les tailles de proloculus sont trčs diverses. A l'inverse, on rappellera que dans le cas de Rhapydionina gr. liburnica, il a paru utile (Fleury, 2014) de distinguer les populations de Grčce de celle du type d'Istrie, tant la différence de taille du proloculus entre les deux régions apparaissait systématique. Un cas non élucidé se présente chez le genre Sigalveolina : l'examen des tests A de S. ovoidea et S. reicheli n. sp. semble montrer que les grands proloculus correspondent ŕ de (relatifs) petits tests alors que, inversement, les plus petits proloculus correspondent aux plus grands tests (voir
Fig. 16 ![]() ) ; le matériel réuni est trop peu abondant pour l'établissement de données statistiques, mais la question de l'existence de deux générations A se trouve posée. On rappellera ŕ cette occasion que De Castro
(1990, p. 103) a révélé chez Pseudochubbina globularis (Smout,
1963) une relation de la taille du proloculus avec les variations de divers caractčres, n'affectant cependant pas la taille des tests.
) ; le matériel réuni est trop peu abondant pour l'établissement de données statistiques, mais la question de l'existence de deux générations A se trouve posée. On rappellera ŕ cette occasion que De Castro
(1990, p. 103) a révélé chez Pseudochubbina globularis (Smout,
1963) une relation de la taille du proloculus avec les variations de divers caractčres, n'affectant cependant pas la taille des tests.
Le mode d'enroulement initial des tests A constitue un critčre classique d'utilisation commode qui, comme tous les caractčres évolutifs doit ętre considéré dans la perspective d'ensemble. Si certains taxons semblent liés ŕ l'un ou l'autre des types streptospiralé ou planispiralé, il en est d'autres qui les empruntent concurremment ou successivement. Ainsi, la population du type de Cuvillierinella, espčce en principe tenue pour typiquement streptospiralée montre des spécimens planispiralés ; en outre les espčces C. perisalentina et C. fluctuans Fleury, 2016 présentent des individus qui se partagent entre l'un ou l'autre des deux types d'enroulement initial. Il s'agit donc d'un critčre qui doit ętre pris en considération mais doit ętre envisagé en dehors de tout esprit de systčme.
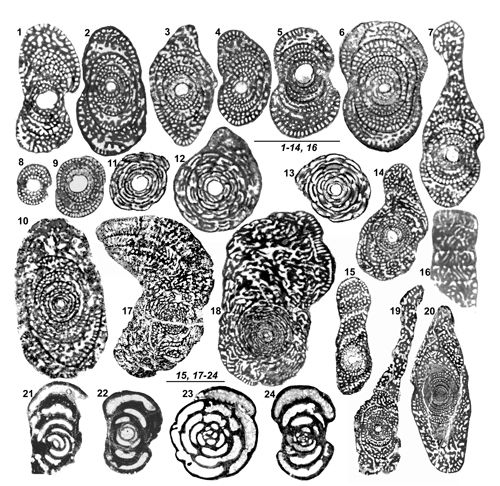
L'importance relative du stade involute chez les tests A est en principe d'usage limité. Il distingue cependant (avec la taille du proloculus) chez le genre Rhapydionina certaines espčces de la zone CsB6b de celles de la zone suivante (Fleury, 2014, Fig. 20). Sa réduction extręme ŕ moins de deux tours enveloppant le proloculus est une caractéristique, parmi d'autres, des genres Cyclopseudedomia et Fanrhapydionina, liée dans les deux cas ŕ des proloculus de trčs grande taille. Au contraire, sa grande importance par rapport ŕ un stade déroulé absent, trčs court ou trčs tardif constitue l'une des caractéristiques du nouveau genre Sigalveolina.
Le mode d'enroulement et de déroulement des tests adultes doit évidemment ętre toujours caractérisé. Dans les genres concernés par cette étude, il est ordinairement planispiralé dčs le stade juvénile des tests A. On observe cependant, trop exceptionnel pour ętre pris en compte autrement que comme la récurrence d'un caractčre ancestral, deux cas (S. renzi,
Fig. 8.11 ![]() ,
et une citation chez S. methonensis) oů l'axe d'enroulement effectue une rotation réguličre
("Slow axial rotation", in Fleury, 2016) et c'est l'une des caractéristiques des genres Pseudochubbina et Metacuvillierinella. Le déroulement final existe chez presque toutes les espčces examinées ici ; il est parfois minoritaire et ne pas constituer la rčgle ; il peut comprendre des loges subsphériques ou aplaties, constituant une PRU cylindrique ou flabelliforme, évolute ou pseudoévolute, ces divers types étant susceptibles de coexister dans la męme génération d'une męme espčce d'une męme localité (S. reicheli n. sp., C. mavrikasi n. sp.,
Figs. 15
,
et une citation chez S. methonensis) oů l'axe d'enroulement effectue une rotation réguličre
("Slow axial rotation", in Fleury, 2016) et c'est l'une des caractéristiques des genres Pseudochubbina et Metacuvillierinella. Le déroulement final existe chez presque toutes les espčces examinées ici ; il est parfois minoritaire et ne pas constituer la rčgle ; il peut comprendre des loges subsphériques ou aplaties, constituant une PRU cylindrique ou flabelliforme, évolute ou pseudoévolute, ces divers types étant susceptibles de coexister dans la męme génération d'une męme espčce d'une męme localité (S. reicheli n. sp., C. mavrikasi n. sp.,
Figs. 15 ![]() - 16
- 16 ![]() ).
).
La forme générale des tests est rarement significative. Ils sont ordinairement de type nautiloďde, de presque sphérique ŕ aplati, ceci męme au sein d'une unique population (M. cuvillieri,
Fig. 5.1-5 ![]() par exemple). Ce critčre pourrait cependant ętre le seul, si l'on y tient, qui puisse ętre qualifié de constituer un
"caractčre spécifique", mais son usage resterait trčs limité si l'on prend en compte l'aspect trčs homogčne de la plupart des tests A des espčces de petite taille. L'indice d'aplatissement des tests
(Fig. 15
par exemple). Ce critčre pourrait cependant ętre le seul, si l'on y tient, qui puisse ętre qualifié de constituer un
"caractčre spécifique", mais son usage resterait trčs limité si l'on prend en compte l'aspect trčs homogčne de la plupart des tests A des espčces de petite taille. L'indice d'aplatissement des tests
(Fig. 15 ![]() ) correspond au rapport du diamčtre équatorial (les tests déroulés étant exclus) ŕ
"l'épaisseur du test", plus grande dimension mesurée parallčlement ŕ l'axe d'enroulement. Cette donnée est peu précise puisque
"l'épaisseur" dépend de l'exacte orientation du plan de section et que les sections axiales passent rarement par le plus grand diamčtre équatorial ; elle donne cependant une idée de la morphologie du test et s'avčre utile ŕ caractériser les cas les plus extręmes, c'est-ŕ-dire les tests les plus plats (S. renzi) et les plus občses (S. ovoidea). Elle n'a pas de signification pour le genre Cyclopseudedomia oů l'indice serait toujours trčs élevé, variant dans de trčs larges proportions en fonction de l'orientation de la section ; on notera cependant que les sections équatoriales pourraient ętre utilisées puisque "l'épaisseur" du test correspond grosso-modo au diamčtre du proloculus.
) correspond au rapport du diamčtre équatorial (les tests déroulés étant exclus) ŕ
"l'épaisseur du test", plus grande dimension mesurée parallčlement ŕ l'axe d'enroulement. Cette donnée est peu précise puisque
"l'épaisseur" dépend de l'exacte orientation du plan de section et que les sections axiales passent rarement par le plus grand diamčtre équatorial ; elle donne cependant une idée de la morphologie du test et s'avčre utile ŕ caractériser les cas les plus extręmes, c'est-ŕ-dire les tests les plus plats (S. renzi) et les plus občses (S. ovoidea). Elle n'a pas de signification pour le genre Cyclopseudedomia oů l'indice serait toujours trčs élevé, variant dans de trčs larges proportions en fonction de l'orientation de la section ; on notera cependant que les sections équatoriales pourraient ętre utilisées puisque "l'épaisseur" du test correspond grosso-modo au diamčtre du proloculus.
L'endosquelette, commun aux tests des deux générations est indépendant des caractčres précédents. Le moment de l'apparition de cet endosquelette chez les tests A n'est pas significatif, puisque s'il est trčs précoce chez les formes planispiralées étudiées ici, il peut l'ętre tout autant chez certaines formes streptospiralées telles que Cuvillierinella perisalentina ou C. fluctuans ou męme certaines formes avancées de C. salentina (voir Fleury, 2016, Figs.
5.25, 6.3 et 7). La particularité des genres ici présentés réside en grande partie dans l'expression de la structure confluente dont le degré de perfection est pris en compte, parmi d'autres critčres, pour distinguer les genres et les espčces. Cette structure est rare mais n'est pas inconnue chez Cuvillierinella (voir Fleury, 2016,
Fig. 6.11) qui serait ŕ l'origine de tous les taxons concernés par cette étude, et l'on peut juger par les exemples de la
Figure 3 ![]() qu'elle est répandue parmi des taxons trčs divers par leur localisation comme par leur âge.
qu'elle est répandue parmi des taxons trčs divers par leur localisation comme par leur âge.
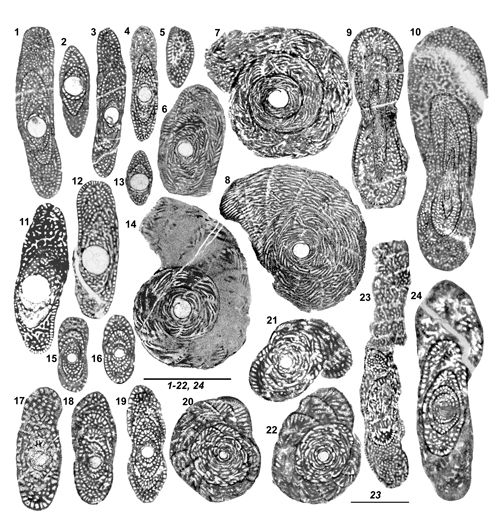
La forme comme le diamčtre absolu et relatif des logettes en section perpendiculaire ŕ leur axe peut ętre utile ŕ remarquer, mais on notera que leur aspect dépend de l'orientation du plan de coupe : une logette cylindrique peut donner une section d'aspect ovale allongé si elle est coupée obliquement par rapport ŕ son axe. Par ailleurs, les variations au sein d'un groupe homogčne selon d'autres critčres incitent ŕ la circonspection (par exemple, pour la taille des logettes secondaires
Fig. 5.1-2 ![]() , pour l'ensemble des logettes Fig. 7.1, .3
, pour l'ensemble des logettes Fig. 7.1, .3 ![]() ), mais il a semblé que des variations systématiques de ces tailles, associées ŕ d'autres critčres, permettaient de distinguer certaines espčces, par exemple S. methonensis de S. renzi.
), mais il a semblé que des variations systématiques de ces tailles, associées ŕ d'autres critčres, permettaient de distinguer certaines espčces, par exemple S. methonensis de S. renzi.
La famille des Rhapydioninidae présente un dimorphisme de générations des plus importants. Celui-ci est marqué dans les stades initiaux, comme chez bien d'autres groupes, mais également dans les stades adultes oů le déroulement, déjŕ parfois notable dans les tests A, prend des proportions inégalées chez certains taxons. Cette génération est donc ŕ la fois "conservatrice" dans ses stades népioniques (elle rappelle un caractčre ancestral) et "novatrice" dans ses stades adultes (elle exacerbe un caractčre de la fin du développement de la génération A). Il s'agit donc d'un critčre essentiel pour discerner les tendances évolutives, clé de la compréhension de la structure du groupe. Sans doute, les tests de cette génération sont rares et les sections significatives trčs difficiles ŕ obtenir. Leur recherche constitue cependant une condition sine qua non pour tenter d'approcher la compréhension du groupe.
En somme, aucun de ces critčres n'est absolu en lui-męme. Ils sont indépendants les uns des autres comme le montre la variété des combinaisons observées
(Fig. 16 ![]() ). Ce sont leurs associations diverses qui permettent de distinguer des lignes évolutives, avec en particulier l'appoint significatif des tests B. S'il est pratique et confortable de juger des éléments de classification ŕ partir de critčres préétablis sur des données trčs fragmentaires, l'expérience montre qu'on ne peut rien attendre de ces méthodes. Il est vrai que ces considérations ne simplifient pas la tâche de celui qui tente de rendre compte des possibles relations naturelles entre les taxons : les guides de conduite, les rčgles couramment admises s'effaçant, plus les données s'accumulent et plus la synthčse est complexe.
). Ce sont leurs associations diverses qui permettent de distinguer des lignes évolutives, avec en particulier l'appoint significatif des tests B. S'il est pratique et confortable de juger des éléments de classification ŕ partir de critčres préétablis sur des données trčs fragmentaires, l'expérience montre qu'on ne peut rien attendre de ces méthodes. Il est vrai que ces considérations ne simplifient pas la tâche de celui qui tente de rendre compte des possibles relations naturelles entre les taxons : les guides de conduite, les rčgles couramment admises s'effaçant, plus les données s'accumulent et plus la synthčse est complexe.
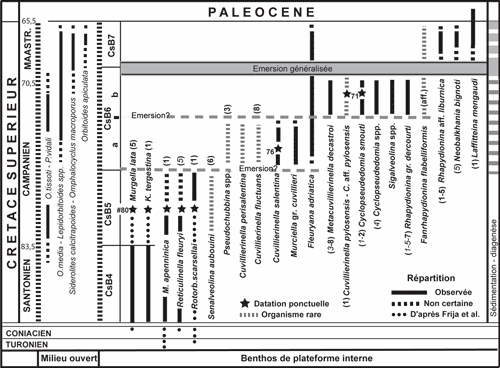
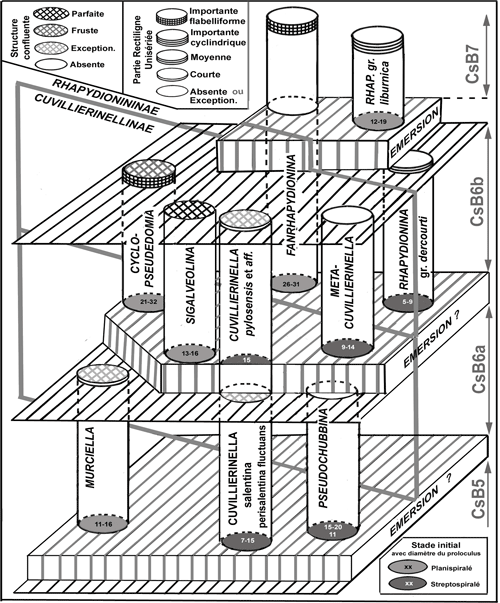
La
Figure 4 ![]() propose une biozonation applicable aux facičs de plate-forme interne pour la période Campanien-Maastrichtien. Issue de l'étude des carbonates de la plate-forme de Gavrovo-Tripolitza en Grčce, elle souffre de la quasi-absence de coupes continues, ŕ l'exception de celle du massif du Klokova (Fleury, 1974, 1980, et en particulier 2016, Fig. 1), oů cependant les organismes les plus typiques de la zone CsB6a ne sont pas observés.
propose une biozonation applicable aux facičs de plate-forme interne pour la période Campanien-Maastrichtien. Issue de l'étude des carbonates de la plate-forme de Gavrovo-Tripolitza en Grčce, elle souffre de la quasi-absence de coupes continues, ŕ l'exception de celle du massif du Klokova (Fleury, 1974, 1980, et en particulier 2016, Fig. 1), oů cependant les organismes les plus typiques de la zone CsB6a ne sont pas observés.
Ainsi, les zones CsB6a et CsB6b bien représentées n'ont jamais été observées en superposition directe sur une męme coupe et leur distinction ne résulte que d'une parfaite ségrégation en deux groupes des organismes qui y participent. Elle réunit donc des observations éparses et tient compte essentiellement des associations constatées par l'auteur dans de nombreuses localités et par Mavrikas (1993) dans le massif du Gavrovo. On notera comme une confirmation les données d'une coupe décrite par Chiocchini & Mancinelli (2001) dans le Latium italien oů sont observées successivement, de bas en haut "Raadshoovenia" salentina (CsB6a), Murciella sp. (CsB6 a ou b), puis Rhapydionina liburnica (CsB7) ; il en est de męme des observations de Reina & Luperto-Sinni (1993) dans la péninsule Salento. Ces résultats sont appliqués par Heba (2008), Heba & Prichonnet (2009) et Le Goff et al. (2015) en Albanie et sont proches de ceux de Cvetko et al. (2001) et Velić (2007) sur le littoral croate (Île de Brač, en particulier).
Les amendements apparus au cours de la derničre décennie ne sont pas négligeables, surtout liés ŕ l'émergence des techniques fondées sur l'étude des isotopes du strontium (SIS : Strontiun isotope stratigraphy), appliquées aux carbonates de diverses plates-formes du domaine périadriatique. Les modifications qu'elles proposent ŕ nos données s'appliquent ŕ trois périodes.
Les zones CsB4 et 5, supposées situées de part et d'autre de la limite Santonien - Campanien et distinguées par la présence puis l'absence de Murgella lata Luperto-Sinni, 1966, et Keramosphaerina tergestina (Stache, 1889), sont mises en cause par les données synthétisées par Frija et al. (2015). Selon ces auteurs, M. lata est présente depuis le Coniacien moyen jusqu'ŕ la limite Campanien inférieur-Campanien moyen (environ 80,5 MA) et K. tergestina (presque jamais contemporaine de la précédente) serait limitée ŕ une courte période située vers la fin du Campanien inférieur. Ce dernier organisme devrait ainsi se trouver dans les męmes couches que celles qui renferment Moncharmontia apenninica (De Castro, 1966), Rotorbinella scarsellai Torre, 1967, Reticulinella fleuryi Cvetko et al., 1997, Calveziconus lecalvezae Caus & Cornella, 1981 (organismes figurés par Fleury, 1970 et/ou 1980, sous des noms parfois erronés), ou encore Senalveolina aubouini Fleury, 1984, en principe propres ŕ la zone CsB5. C'est dire que, si ces données se confirment, la distinction des zones CsB4 et CsB5 est sans fondement et que, de maničre plus générale, toute zonation de la période comprise entre le Turonien et le Campanien supérieur est illusoire : la plus grande partie des organismes identifiés dans cet intervalle apparaîtrait au cours du Turonien - Coniacien et persisteraient au moins jusqu'ŕ l'extręme base du Campanien supérieur. C'est donc que les variantes observées dans les apparentes répartitions stratigraphiques des organismes connus seraient simplement liées aux hasards de l'échantillonnage (peu vraisemblables pour M. lata et K. tergestina, bien discernables sur le terrain) ou ŕ de subtiles modifications du milieu. Il conviendra cependant d'assurer que ces męmes modifications n'influent pas sur les données de la SIS.
La zone CsB6a est caractérisée par des organismes qui sont manifestement influencés par les conditions du milieu ; ils apparaissent synchroniquement avec les "facičs d'émersion" qui vont se développer et s'intensifier jusqu'ŕ la fin du Crétacé. L'âge proche de 76 MA donné par Schlüter et al. (2008) pour la localité type de Cuvillierinella salentina situe la zone au cours du Campanien supérieur.
La zone CsB6b est également située au cours du Campanien supérieur (et Maastrichtien inférieur ?), en raison de l'âge proche de 71 MA attribué par Vicedo et al. (2011) ŕ la localité type de Cyclopseudedomia smouti.
Les facičs d'émersion qui viennent d'ętre évoqués conduisent ŕ rappeler la dépendance essentielle des Rhapydioninidae du Campanien-Maastrichtien ŕ la nature des milieux de vie dans le domaine de la plate-forme interne ŕ circulation restreinte auxquels ils sont clairement inféodés. Ils en constituent d'ailleurs ordinairement les seuls peuplements, avec quelques Miliolidés, Cuneolina, "Discorbis" et gastéropodes. S'il est possible de trouver dans les descriptions qui suivent des références ŕ des tests dissous et/ou recristallisés, des silts vadoses ou męme des amas de cristaux d'anhydrite et des restes de charophytes dans les échantillons fossilifčres, on devra aussi considérer les alternances de carbonates dans lesquels ils sont inclus, qui témoignent fréquemment de dépôts inter- ŕ supratidaux ayant subi parfois des diagenčses en milieux émergés. Bernier et Fleury (1980) en ont donné une image d'ensemble, précisée en divers affleurements par Fleury (1980).
On discutera en conclusion de l'influence éventuelle des phénomčnes tectoniques et eustatiques sur cette zonation, dont les organismes vivant dans de trčs faibles tranches d'eau sont les plus fragiles victimes.
|
Figure 4 : Essai de zonation biostratigraphique des facičs de plate-forme interne pour la période campano-maastrichtienne. Les données éparses et discontinues sont issues des travaux de Fleury (1980, 2014, 2016) et Mavrikas (1993) en Grčce. Ces données sont ici complétées par des datations ponctuelles relatives ŕ la zone CsB5 (Frija et al., 2015), la zone CsB6a (Schlüter et al., 2008) et la zone CsB6b (Vicedo et al., 2011). Les incertitudes sont soulignées par la discontinuité de la ligne qui sépare les colonnes des zones et des étages. Vers la droite, une colonne (Sédimentation diagenčse) résume schématiquement l'évolution des milieux sur la plateforme de Gavrovo-Tripolitza en Grčce (facičs subtidaux en blanc, facičs d'émersion en gris) ; un trait marquant de cette évolution est constitué par l'émersion généralisée proche de la limite Campanien-Maastrichtien (Landrein, 2001). Des phénomčnes de męme nature, traduits par des lignes discontinues, sont supposés entre les zones CsB6b/CsB6a et CsB6/CsB5 en raison des discontinuités des peuplements et diverses indications rapportées dans le texte. Les publications de l'auteur dans lesquelles apparaissent des figurations de taxons provenant de la coupe du Klokova sont indiquées par un chiffre, entre parenthčses, selon le code suivant: 1= 1970; 2= 1974; 3= 1977; 4= 1979a; 5= 1980; 6= 1984; 7= 2014; 8= 2016; ce chiffre est en italique maigre lorsque l'organisme est présenté en légende de la publication citée sous un nom erroné. |
Ordre Foraminifera Eichwald, 1830
Sous-ordre Miliolina Delage & Hérouard, 1896
Super-famille Alveolinacea Ehrenberg, 1839
Famille Rhapydioninidae Keijzer, 1945
Sous-famille Cuvillierinellinae Fleury, 2016
Rappelons que, par différence avec celle des Rhapydionininae, cette sous-famille distingue des genres dotés d'une zone marginale étroite, dont la plupart comporte la structure confluente ; une image schématique d'ensemble en est donnée in Fleury, 2016, Fig. 13.
Le genre Cuvillierinella Papetti & Tedeschi,
1965, est vraisemblablement ŕ l'origine des taxons examinés ci-dessous. Il est maintenant bien connu par sa population type d'Italie comme par plusieurs populations de Grčce, d'Espagne et de Méditerranée orientale (Fleury, 2016) et mérite qu'on en rappelle les caractčres principaux. Les stades juvéniles des deux générations en sont de type miliolin ŕ streptospiralé (exceptionnellement planispiralé chez de rares individus); les tests A nautiloďdes ŕ proloculus sphérique et flexostyle se distinguent par un éventuel court stade terminal cylindrique des tests B de męme morphologie mais susceptibles de développer une grande partie terminale évolute ŕ pseudoévolute (jamais discoďde ŕ ce qu'on en connaît). L'endosquelette est constitué par deux jeux de logettes d'apparition parfois tardive et de diamčtre relativement grand, soit différenciées par leur taille, soit formant un réseau unidimensionnel dense, soit encore dispersées dans un endosquelette central massif (SSC); la structure confluente y est exceptionnelle; l'espace préseptal relativement profond est meublé par des piliers préseptaux rares chez le jeune, disposés en une ou plusieurs rangées dans les derničres loges des tests B. Trois espčces lui sont rapportées de la męme zone CsB6a, mais une quatričme, associée ŕ C. smouti en un seul gisement, atteint la zone CsB6b. Le genre est donc typiquement ambivalent (voir Fleury, 2016 et ici
Fig. 16 ![]() ), męlant les caractčres hérités des Miliolacea ŕ ceux qui sont développés par son plus proche compagnon (Murciella), son proche cousinage vraisemblable (Pseudochubbina) et ses successeurs (Cyclopseudedomia, Metacuvillierinella, Sigalveolina n. gen.).
), męlant les caractčres hérités des Miliolacea ŕ ceux qui sont développés par son plus proche compagnon (Murciella), son proche cousinage vraisemblable (Pseudochubbina) et ses successeurs (Cyclopseudedomia, Metacuvillierinella, Sigalveolina n. gen.).
Genre Murciella Fourcade, 1966
Espčce-type : Murciella cuvillieri Fourcade, 1966.
Diagnose. Genre de la sous-famille Cuvillierinellinae, dimorphique : tests A entičrement planispiralés involutes, parfois terminés par une courte PRU évolute cylindrique ŕ rarement flabelliforme, tests B streptospiralés chez le jeune terminés par une PRU évolute flabelliforme de taille relativement modérée. Endosquelette simple de type coaxial dominant, passant exceptionnellement au type confluent. Diffčre de Cuvillierinella son contemporain par le mode d'enroulement des parties juvéniles de ses tests A et son bâti plus fręle. On ne reconnaît qu'une seule espčce qui peuple la zone CsB6a depuis l'Espagne jusqu'ŕ la Méditerranée orientale, représentée ici par six populations.
Murciella cuvillieri Fourcade, 1966
Diagnose. Espčce type du genre Murciella caractérisée par ses tests A de taille modeste, polymorphes, présentant un déroulement terminal cylindrique ŕ exceptionnellement flabelliforme. Tests B ŕ partie terminale flabelliforme pseudoévolute ŕ évolute de taille médiocre. Bâti et endosquelette de taille moyenne, structure coaxiale, exceptionnellement hélicoďdale. Distincte des espčces du genre Cuvillierinella par ses tests A strictement planispiralés et son endosquelette ŕ maille plus fine. Distincte des espčces du genre Sigalveolina n. gen. par la quasi absence de la structure confluente.
Cette espčce, créée dans un contexte alors peu exploré, a connu une histoire complexe. Elle fut un temps rapportée au genre Raadshoovenia Van den Bold, 1946, lorsque, en raison d'une quasi-similitude entre l'espčce type de ce genre et celle de Cuvillierinella (voir De Castro, 1971), ce genre américain fut soupçonné d'avoir des représentants crétacés européens et que M. cuvillieri, de morphologie proche, puisse lui ętre assimilée. Fleury & Fourcade (1990), s'appuyant sur l'âge cénozoďque du taxon américain établi par Pęcheux (1984) et les caractéristiques propres ŕ Murciella, proposaient d'infirmer cette double hypothčse. Par ailleurs, De Castro (1988) devait révéler que certains tests de morphologie proche de Murciella se trouvaient parmi les populations de la localité type de Cuvillierinella salentina. Pour Fleury (2016), réétudiant ce riche matériel aimablement confié par P. De Castro, les formes de type Murciella de cette population, trčs minoritaires et jamais exactement comparables ŕ M. cuvillieri, peuvent ętre considérées comme des variants intraspécifiques et ne justifient pas que les deux taxons soient confondus. En outre, l'existence exprimée dans les lignes qui suivent de plusieurs populations de M. cuvillieri (EF436, XGP163 et XGP330) dépourvues de tout représentant susceptible d'ętre rapporté ŕ Cuvillierinella, ou encore témoignant de la coexistence des deux types, sans le moindre intermédiaire morphologique (EFG1, XGP110 et FPM249), constitue le signe d'une complčte indépendance des deux taxons.
Les 6 populations décrites sont réparties sur un grand territoire qui va de l'Espagne ŕ la méditerranée orientale, en passant par la Grčce ; elles sont choisies parmi les plus riches et les plus distinctes les unes des autres qui aient été rencontrées. Si elles constituent un ensemble homogčne par rapport ŕ Cuvillierinella, elles montrent cependant entre elles des différences de détail qui méritent d'ętre signalées par la terminologie. Si chacune d'elles peut ętre nommée M. cuvillieri, leur ensemble peut ętre caractérisé par la dénomination M. gr. cuvillieri.
Murciella cuvillieri
Population EF436 (Population type)
Comportant une abondante illustration, la publication d'origine est suffisante pour caractériser l'espčce. L'intéręt que l'on pourra trouver aux nouvelles figures proposées réside dans des grandissements homogčnes, permettant une comparaison visuelle directe aux autres populations décrites ci-aprčs et aux figurations de C. salentina (in Fleury, 2016, Fig. 6). Une grande partie des sections du matériel type a été observée, dessinée et photographiée par l'auteur, avant son dépôt auprčs du Museum National d'Histoire Naturelle, Paris, sous les numéros Lma2000/131 ŕ 150, ŕ l'origine. Selon Marie-Béatrice Forel, Maître de Conférences au MNHN qui a recherché le matériel, celui-ci est finalement conservé dans les collections de géologie sous les nº MNHN-GG-GG2000-131 ŕ 150 et accessible en référence par le lien suivant : https://science.mnhn.fr/institution/mnhn/collection/gg/item/list?full_text=Murciella+cuvillieri. De męme le matériel type des diverses espčces du genre Praechubbina décrites par Fleury & Fourcade (2001), déposé ŕ la męme occasion, sous les nº Lma2000/59 ŕ 130, est conservé sous les nº MNHN-GG-GG2000-59 ŕ 130 et accessible en référence par le lien suivant : https://science.mnhn.fr/institution/mnhn/collection/gg/item/list?full_text=Praechubbina+breviclaustra.
Localisation, conditions de gisement. Dans la localité type, (Sierra de los Agueros, Province de Murcie, Espagne) d'aprčs Fourcade (1966), M. cuvillieri est accompagnée de quelques organismes (oogones de charophytes, Ostracodes, "Discorbis", Nummoloculines et Milioles), indiquant un milieu isolé du marin franc, en l'absence de tout représentant du genre Cuvillierinella. La population se trouve dans des calcaires compris entre des couches ŕ Lacazina elongata Munier-Chalmas, attribuables au Santonien et des couches ŕ Orbitoides media Archiac, Omphalocyclus macroporus Lmk et Siderolites cf. calcitrapoides du Maastrichtien. Ces informations ne sont pas précisées par Vicedo et al. (2009) qui ont figuré quelques "topotypes".
|
Figure 5
: Murciella cuvillieri (EF436),
population type. 1-14 : Tests A. 1-7 :
Sections axiales centrées décrivant les divers types morphologiques connus
(voir aussi Fig. 2.5 |
Morphologie, enroulement.
Tests A. Les diverses sections permettent de déduire l'aspect nautiloďde des tests
(Figs. 2.5 ![]() ,
5.1-5
,
5.1-5 ![]() ), plus ou moins ovoďdes ou aplatis, toujours franchement biombiliqués. Le déroulement final, qui n'est pas de rčgle, est parfois cylindrique
(Fig. 5.10
), plus ou moins ovoďdes ou aplatis, toujours franchement biombiliqués. Le déroulement final, qui n'est pas de rčgle, est parfois cylindrique
(Fig. 5.10 ![]() ), exceptionnellement flabelliforme
(Fig. 5.13
), exceptionnellement flabelliforme
(Fig. 5.13 ![]() ). Les sections transverses de loges isolées, rectangulaires ou subcirculaires
(Fig. 5.15-16
). Les sections transverses de loges isolées, rectangulaires ou subcirculaires
(Fig. 5.15-16 ![]() ), peuvent donc appartenir ŕ l'une ou l'autre des générations. Le proloculus sphérique d'un diamčtre moyen de 0,11 mm est suivi d'un flexostyle étroit et court, puis d'une spire serrée dont la hauteur s'accroit faiblement jusqu'ŕ la fin du troisičme ou quatričme tour ; le tour suivant montre un accroissement rapide de sa hauteur, témoignant d'une tendance au déroulement. Les tests déroulés ne sont pas fréquents (environ 10% des sections). Le diamčtre équatorial des tests A certains, partie déroulée comprise, est située entre 0,55 et 1,72 mm ; ce diamčtre ne dépasse pas 1,4 mm pour les tests non déroulés. Pour les 15 sections dépassant 1 mm (tests considérés comme adultes), la moyenne s'établit ŕ 1,2 mm.
), peuvent donc appartenir ŕ l'une ou l'autre des générations. Le proloculus sphérique d'un diamčtre moyen de 0,11 mm est suivi d'un flexostyle étroit et court, puis d'une spire serrée dont la hauteur s'accroit faiblement jusqu'ŕ la fin du troisičme ou quatričme tour ; le tour suivant montre un accroissement rapide de sa hauteur, témoignant d'une tendance au déroulement. Les tests déroulés ne sont pas fréquents (environ 10% des sections). Le diamčtre équatorial des tests A certains, partie déroulée comprise, est située entre 0,55 et 1,72 mm ; ce diamčtre ne dépasse pas 1,4 mm pour les tests non déroulés. Pour les 15 sections dépassant 1 mm (tests considérés comme adultes), la moyenne s'établit ŕ 1,2 mm.
Tests B. L'holotype (Fig. 5.17 ![]() ) a jusqu'ŕ présent constitué une énigme. La section non centrée est en effet différente de toutes les autres connues dans sa population par sa partie terminale flabelliforme et la sinuosité de l'arrangement des logettes secondaires (structure confluente) qui s'y observe. Une section centrée d'une autre population
(Fig. 6.24
) a jusqu'ŕ présent constitué une énigme. La section non centrée est en effet différente de toutes les autres connues dans sa population par sa partie terminale flabelliforme et la sinuosité de l'arrangement des logettes secondaires (structure confluente) qui s'y observe. Une section centrée d'une autre population
(Fig. 6.24 ![]() ) comportant un peloton miliolin initial, et par ailleurs trčs comparable par son aspect d'ensemble, son espace préseptal profond et ses logettes secondaires sinueuses permet maintenant de l'interpréter indubitablement. Une section axiale subcentrée compatible avec la précédente
(Fig. 5.18
) comportant un peloton miliolin initial, et par ailleurs trčs comparable par son aspect d'ensemble, son espace préseptal profond et ses logettes secondaires sinueuses permet maintenant de l'interpréter indubitablement. Une section axiale subcentrée compatible avec la précédente
(Fig. 5.18 ![]() ) complčte l'image de cette génération. Il convient de rappeler que Fourcade (1966, Pl.
2, fig. 10) a figuré une section centrée non déroulée d'un type plus proche des tests A.
) complčte l'image de cette génération. Il convient de rappeler que Fourcade (1966, Pl.
2, fig. 10) a figuré une section centrée non déroulée d'un type plus proche des tests A.
Endosquelette. Le cloisonnement apparaît trčs précocement, dčs le premier tour dans les tests bien conservés
(Figs. 2.5 ![]() , 5.1, .3-6
, 5.1, .3-6 ![]() ). Les logettes secondaires présentent ordinairement un diamčtre un peu plus faible que celui des primaires. La multiplication des niveaux de logettes secondaires apparaît dčs le deuxičme ou troisičme tour selon le caractčre plus ou moins serré de la spire. Si des
"planchers" peuvent sembler parfois séparer ces niveaux de logettes en section équatoriale, il n'en est plus de męme dans les loges adultes oů la disposition prend un aspect irrégulier bien discernable dans les sections axiales. Cette irrégularité est en partie liée ŕ la structure confluente, responsable de l'attribution erronée au genre Murciella de diverses espčces comportant la męme structure
("Murciella gr. renzi" in Fleury, 1979a), ŕ un degré plus accentué : l'orientation ŕ 45°
des logettes de couches adjacentes chez Sigalveolina n'est jamais observable chez Murciella. C'est en considérant les tendances évolutives plutôt que des critčres artificiellement préétablis que sera justifiée la création du nouveau genre Sigalveolina n. gen. pour caractériser l'originalité des espčces qui lui sont attribuées.
). Les logettes secondaires présentent ordinairement un diamčtre un peu plus faible que celui des primaires. La multiplication des niveaux de logettes secondaires apparaît dčs le deuxičme ou troisičme tour selon le caractčre plus ou moins serré de la spire. Si des
"planchers" peuvent sembler parfois séparer ces niveaux de logettes en section équatoriale, il n'en est plus de męme dans les loges adultes oů la disposition prend un aspect irrégulier bien discernable dans les sections axiales. Cette irrégularité est en partie liée ŕ la structure confluente, responsable de l'attribution erronée au genre Murciella de diverses espčces comportant la męme structure
("Murciella gr. renzi" in Fleury, 1979a), ŕ un degré plus accentué : l'orientation ŕ 45°
des logettes de couches adjacentes chez Sigalveolina n'est jamais observable chez Murciella. C'est en considérant les tendances évolutives plutôt que des critčres artificiellement préétablis que sera justifiée la création du nouveau genre Sigalveolina n. gen. pour caractériser l'originalité des espčces qui lui sont attribuées.
Au total, cette population, riche et variée pour les tests A et recélant un exemple trčs vraisemblable de test B est tout ŕ fait propre ŕ caractériser une espčce et un genre de statut irrévocable. Les autres populations d'origines variées qui s'y rattachent et seront décrites ci-dessous permettront de nuancer son individualité mais n'ajouteront rien de fondamental ŕ sa signification.
Murciella cuvillieri
Population XGP110
(Fig. 6.1-14 ![]() )
)
Localisation, conditions de gisement. Deux échantillons, trčs proches l'un de l'autre (XGP110-111) ont été étudiés dans un lot de 42 lames. Ils renferment une męme association proviennent du massif connu des géologues sous le nom ancien de Massif du Gavrovo (Ori Valtou), représentatif de la plate-forme de Gavrovo-Tripolitza en Grčce. L'affleurement est décrit par Mavrikas (1993, p. 77, facičs de type III). Il s'agit de calcaires dont l'examen est rendu difficile par une stratification peu nette et les nombreuses diaclases qui l'affectent ; ils sont principalement caractérisés par l'abondance des tests de petits Rhapydioninidés qui s'y trouvent. Dans le détail, la population de ces échantillons est comprise dans des biomicrites wackestone ŕ débris de rudistes et de gastéropodes, "Discorbis", trčs rares Orbitoides, Cuneolina sp., et Fleuryana adriatica De Castro et al., 1994. Une riche population de Cuvillierinella salentina (environ 80 sections, dont 40 centrées), figurée par Fleury (2016, Fig. 6.1-12) accompagne les Murciella décrites ici, sans qu'existent de formes de morphologie intermédiaire susceptibles de provoquer la confusion.
Morphologie, enroulement.
Tests A. Cette génération est représentée par une cinquantaine de sections centrées, montrant toutes un enroulement strictement planispiralé. Les sections axiales témoignent de tests nautiloďdes, plus ou moins aplatis, biombiliqués, dont les tours de spire présentent en section le męme aspect ogival ou arrondi que la population type. Le déroulement final de type cylindrique (jusqu'ŕ 4 loges) est observé ŕ la faveur de plusieurs sections équatoriales et le déroulement de type flabelliforme ne peut ętre exclu bien que les sections décentrées qui en témoignent puissent appartenir ŕ des tests B. Le proloculus de section circulaire, d'un diamčtre moyen de 0,11 mm est suivi d'un flexostyle étroit et de loges participant ŕ une spire serrée dont la hauteur s'accroit réguličrement jusque vers la fin du quatričme tour, oů elle devient trčs forte, marquant la tendance au déroulement. Le diamčtre équatorial, partie déroulée comprise est situé entre 0,62 et 1,74 mm ; les 22 plus grands mesures excčdent toutes 1,12 mm, pour une moyenne de 1,30 mm.
Tests B. Ces tests sont plus grands que les tests A (jusqu'ŕ 2 et presque 3 mm de diamčtre). Deux sections subcentrées sont connues
(Fig. 6.12-13 ![]() ), qui témoignent d'un déroulement final de type indéterminé ; une section axiale décentrée
(Fig. 6.14
), qui témoignent d'un déroulement final de type indéterminé ; une section axiale décentrée
(Fig. 6.14 ![]() ) indiquerait plutôt un type flabelliforme.
) indiquerait plutôt un type flabelliforme.
Endosquelette. Il apparaît dčs le premier tour. Les logettes primaires sont ordinairement de section transverse ovale ŕ presque rectangulaire et d'un diamčtre plus fort que les secondaires, de section circulaire ; ces derničres sont plus fines et plus serrées que chez toutes les autres populations observées. En conséquence, la structure confluente est discernable dans toutes les sections équatoriales des tests des deux générations.
Au total, comparable par ses dimensions, cette population ne se distingue de celle du type que par la finesse de son bâti et de sa maille endosquelettique, qui permettent d'observer la structure confluente.
|
Figure 6
: Murciella cuvillieri. 1-14 : Population XGP110.
1-8: Sections axiales centrées de tests A (voir une partie de 4, renseignée
sur Fig. 2.3 |
Murciella cuvillieri
Population XGP330
(Fig. 6.15-25 ![]() )
)
Localisation, conditions de gisement. L'échantillon provient du męme gisement que XGP110. La population est représentée par plus de 100 sections, dont 32 centrées, réparties en 10 lames minces. Le calcaire est de type packstone ŕ débris de rudistes, Cuneolina sp. et Milioles. On n'y observe pas de Cuvillierinella.
Morphologie, enroulement.
Les tests A constituent la majorité du peuplement. Ils sont nautiloďdes, trčs proches d'aspect de ceux du type de l'espčce. Le proloculus de section circulaire est d'un diamčtre moyen de 0,12 mm, le flexostyle étroit est d'une longueur égale ŕ la moitié de sa circonférence. Le pas de la spire ne s'accroit fortement qu'aprčs 3 ŕ 4 tours, donnant naissance ŕ une éventuelle PRU, flabelliforme pseudoévolute ou cylindrique de quelques loges (jusqu'ŕ 5) de section ovale ou circulaire d'un diamčtre atteignant 0,56 mm. Le diamčtre équatorial des tests A certains est compris entre 0,40 et 2 mm ; il est de 1,36 mm pour les 16 plus grands tests.
Les tests B sont connus par 2 sections centrées. L'une, équatoriale
(Fig. 6.24 ![]() ), d'un diamčtre de 2 mm, montre des loges initiales organisées selon le mode quinquéloculin et un stade final flabelliforme pseudoévolute. Sa ressemblance avec l'holotype de l'espčce permet d'interpréter surement celui-ci comme représentant cette génération. Une section axiale
(Fig. 6.25
), d'un diamčtre de 2 mm, montre des loges initiales organisées selon le mode quinquéloculin et un stade final flabelliforme pseudoévolute. Sa ressemblance avec l'holotype de l'espčce permet d'interpréter surement celui-ci comme représentant cette génération. Une section axiale
(Fig. 6.25 ![]() ) est compatible avec la section équatoriale.
) est compatible avec la section équatoriale.
Endosquelette. Il apparaît dčs le premier tour de spire. Le bâti est comparable ŕ celui de la population type ; les logettes secondaires de section circulaire sont selon les cas de męme diamčtre ou plus petites que les primaires, de section ovale ŕ rectangulaire. L'espace préseptal est étroit et les piliers préseptaux bien formés. Les sinuosités liées ŕ la structure confluente sont discernables dans toutes les sections équatoriales, au moins dans les derničres loges des tests des deux générations.
Au total, cette population présente de bonnes ressemblances avec la population type, avec des tests ŕ peine plus grands et une structure confluente mieux exprimée.
Murciella cuvillieri
Population EFG1
(Fig. 7.1-11 ![]() )
)
Localisation, conditions de gisement. Cette population provient d'Espagne (Pantano de Camarillos, Province de Murcie). Un lot de 18 sections centrées comprises dans 14 lames a été examiné, provenant d'un don ŕ l'auteur d'Éric Fourcade, qui le considérait comme un "quasi-topotype". Des Cuvillierinella salentina participent ŕ l'association ; Hamaoui et Fourcade (1973) en ont donné quelques illustrations (op. cit., Pl. 2, figs. 1, 3 et 9 ; Pl. 5, figs. 9-10) et elles sont décrites et figurées par Fleury (2016, Fig. 6.19-31).
Morphologie, enroulement.
Les tests A sont nautiloďdes globuleux, jamais aplatis. Le diamčtre du proloculus de section circulaire s'établit en moyenne ŕ 0,14 mm. La spire de 2,5 ŕ 3 tours (exceptionnellement 4) est d'un diamčtre compris entre 0,58 et 1,45 mm (moyenne 0,99) ; la moyenne pour les 8 plus grandes mesures est de 1,23 mm. Des sections circulaires de PRU sont observées, mais les indices de déroulement des derničres loges sont rares.
Les tests B sont représentés par deux sections axiales subcentrées
(Fig. 7.9-10 ![]() ) de tests involutes biombiliqués, dépourvus d'indices de déroulement. Une section équatoriale, peu distincte dans sa partie centrale montre cependant un déroulement flabelliforme.
) de tests involutes biombiliqués, dépourvus d'indices de déroulement. Une section équatoriale, peu distincte dans sa partie centrale montre cependant un déroulement flabelliforme.
Endosquelette. Il apparaît dčs le premier tour. Le bâti semble un peu plus massif que celui du type ; les logettes présentent de faibles différenciations au sein d'un męme test mais des diamčtres différents d'un test ŕ l'autre
(Fig. 7.1, .3 ![]() ). La structure confluente ne peut ętre soupçonnée que dans la section de la
Figure 7.11
). La structure confluente ne peut ętre soupçonnée que dans la section de la
Figure 7.11 ![]() .
.
Au total, cette population reste comparable ŕ celle du type mais se distingue par un proloculus A de taille un peu plus grande et probablement une moindre tendance au déroulement final.
|
Figure 7 : Murciella cuvillieri. 1-11 : Population EFG1. 1-4 : Sections axiales centrées de tests A. Noter la différence de taille des logettes entre les sections 1 et 3, ainsi que la précocité de l'apparition du cloisonnement dans l'ensemble des tests. 5-6 : Sections équatoriales montrant le flexostyle (noter en 5 l'absence de communication directe entre logettes primaires de loges successives). 7-8 : Sections transverses de PRU cylindriques. 9-11 : Sections axiales et équatoriale subcentrées de tests B vraisemblables (10, non observée, est reproduite d'aprčs Hamaoui et Fourcade, 1973, Pl. 5.2, indication de grandissement respectée). 12-23 : Population FPM246. 12-16 : Sections axiales centrées de tests A. Noter le cloisonnement précoce et l'homogénéité de la taille de l'ensemble des logettes. 17-20: Sections équatoriales centrées de tests A montrant une tendance au déroulement final. 21 : Section transverse de PRU cylindrique. 22-23: Sections axiale presque centrée et équatoriale centrée de tests B témoignant d'une tendance modérée au déroulement final. 24-33 : Population XGP163. 24. Section oblique centrée de test A montrant une PRU probablement flabelliforme. 25-28: Sections axiales de tests A ; noter le flexostyle en 28 et le cloisonnement précoce généralisé. 29-30 : Sections équatoriales centrées de tests A ; les derničres loges de 30 témoignent de la structure confluente. 31-32 : Sections décentrées de tests A probables montrant l'arrangement des logettes, toutes sensiblement de męme diamčtre. 33 : section fortement décentrée d'un test comportant une grande PRU flabelliforme, interprétable comme de génération B. Échelles graphiques : 1 mm. |
Murciella cuvillieri
Population FPM249
(Fig. 7.12-23 ![]() )
)
Localisation, conditions de gisement. Cette population provient de l'Île grecque d'Astypalia (Dodécančse). Récoltée par Phédon Marnelis, elle fut transmise par Michel Bonneau, sans indications précises sur le gisement. Il s'agit de deux trčs riches échantillons (FPM 249 et 250) constitués par un calcaire packstone ŕ débris de rudistes et de gastéropodes, Cuneolina sp. et Milioles. Des Cuvillierinella salentina (décrites par Fleury, 2016, p. 212) accompagnent les Murciella, fréquemment recristallisée en leur centre, déjŕ illustrées par Fleury (1979a, Pl. 5, figs. 16, 19, 25-26) sous le nom de Murciella aff. cuvillieri.
Morphologie, enroulement.
Les tests A, connus par environ 200 sections, également en partie recristallisés, appartiennent ŕ une gamme comprise entre des formes globuleuses profondément creusées par l'ombilic et des formes plates présentant une tendance évolute marquée, le proloculus affleurant sur les deux faces. Le proloculus de section circulaire est d'un diamčtre de 0,15 en moyenne ; le flexostyle est indiscernable. Le pas de la spire s'accroît réguličrement jusqu'ŕ l'amorce du déroulement final. Le déroulement des derničres loges est fréquent, formant une PRU cylindrique ou flabelliforme. Le diamčtre équatorial est en moyenne de 1,24 mm pour les plus grands tests.
Les tests B sont rares, de plus grande taille que les autres. Deux bonnes sections
(Fig. 7.22-23 ![]() ), presque centrées sont typiques de cette génération et confirment l'interprétation donnée de l'holotype de l'espčce.
), presque centrées sont typiques de cette génération et confirment l'interprétation donnée de l'holotype de l'espčce.
Endosquelette. Il apparaît dčs le premier tour, au moins dans les rares sections peu recristallisées en leur centre
(Fig. 7.16 ![]() ). Le bâti est comparable ŕ celui de la population type, bien que logettes primaires et secondaires soient plus ordinairement de męme diamčtre et que la forme rectangulaire des primaires ne soit pas toujours discernable. La structure confluente peut-ętre subodorée dans beaucoup de sections mais n'est clairement exprimée que dans les derničres loges de quelques tests
(Fig. 7.17-20, 22-23
). Le bâti est comparable ŕ celui de la population type, bien que logettes primaires et secondaires soient plus ordinairement de męme diamčtre et que la forme rectangulaire des primaires ne soit pas toujours discernable. La structure confluente peut-ętre subodorée dans beaucoup de sections mais n'est clairement exprimée que dans les derničres loges de quelques tests
(Fig. 7.17-20, 22-23 ![]() ).
).
Au total, cette population se distingue essentiellement de celle du type par son proloculus de plus grande taille et une tendance un peu plus forte au déroulement final.
Murciella cuvillieri
Population XGP163
(Fig. 7.24-33 ![]() )
)
Localisation, conditions de gisement. La population provient du męme gisement que celui des échantillons XGP110 et 330. Elle se trouve dans un lot de 9 lames ; il s'agit d'un calcaire packstone ŕ débris de rudistes et rares Milioles. Les Murciella n'y sont accompagnées par aucun représentant du genre Cuvillierinella.
Morphologie, enroulement.
Les tests A sont connus par une cinquantaine de sections, dont une trentaine centrées. Ils sont globuleux, creusés d'un ombilic peu profond. Le proloculus de section circulaire ŕ ovale est d'un diamčtre moyen de 0,16 mm. Le flexostyle étroit et apparemment court est suivi d'une spire de 3 ŕ 4 tours dont le pas croît réguličrement jusqu'ŕ l'amorce de la courte partie finale déroulée dont témoigne une dizaine de sections. Les plus grands tests ont un diamčtre moyen de 1,29 mm. Un seul grand test présentant 4 loges déroulées atteint un diamčtre proche de 2 mm
(Fig. 7.24 ![]() ).
).
Les tests B ne sont vraisemblablement représentés que par une seule section décentrée constitué d'un petit stade involute contrastant avec un grand stade final flabelliforme
(Fig. 7.33 ![]() ).
).
Endosquelette. Le bâti est d'épaisseur moyenne. Les logettes, toutes de section subcirculaire, sont sensiblement équidimensionnelles. La structure confluente n'est jamais bien caractérisée, bien que quelques indices en existent
(Fig. 7.30 ![]() ).
).
Au total, cette population dont les diamčtres de tests entrent dans la norme des précédentes ne semble se distinguer que par la relative grande taille de son proloculus. De telles tailles se trouvent chez des espčces du genre Sigalveolina n. gen, qui marquent leur différence par des tests un peu plus grands bien qu'ŕ peine déroulés et surtout l'acquisition d'une structure confluente beaucoup mieux réalisée. La génération B, trčs imparfaitement connue, constituerait le seul indice d'une certaine individualité de cette population.
En résumé, les populations rapportées ŕ M. cuvillieri participent ŕ un ensemble cohérent, bien que chacune d'entre elles présente une ou plusieurs particularités. La taille du proloculus A se révčle variable, mais dans une gamme homogčne sans discontinuité ; les tests A peuvent apparaître plus ou moins globuleux ou aplatis dans une population que dans les autres mais il existe également des variations dans chacune des populations ; le bâti de l'une des populations est plus fin que celui des autres, mais des variations existent au sein męme de plusieurs populations. Les tests B sont apparemment homogčnes, caractérisés par un faible déroulement flabelliforme, ŕ une exception prčs. Finalement, c'est
"l'air de famille" qui l'emporte : ces populations, malgré leurs différences de détail, forment un ensemble bien différencié de
tous les autres, comme il apparaît sur les Figures 15 ![]() -
16
-
16 ![]() .
.
Par rapport ŕ C. salentina, en ce qui concerne la génération A, l'abandon de l'enroulement streptospiralé juvénile, l'apparition toujours précoce du cloisonnement et la maille endosquelettique plus fine constituent les seules différences notables. Le déroulement plus modeste des tests B est sensible mais n'est pas nécessairement distinctif. En somme, Murciella constitue un avatar de Cuvillierinella, plus "avancé" dans ses stades juvéniles, mais peut-ętre plus "conservateur" dans ses stades adultes de génération B.
Toutes ces populations sont en principe du męme âge, c'est-ŕ-dire participent ŕ la zone CsB6a. Cette attribution est peu douteuse lorsqu'elles sont associées ŕ des C. salentina, mais cet âge n'est admissible pour les populations constituées d'une seule de ces deux espčces qu'en raison de l'absence des taxons propres ŕ la zone CsB6b. Inversement, on doit prendre en considération qu'aucune des nombreuses localités oů l'association des taxons de la zone CsB6b a été observée n'a livré des M. gr cuvillieri.
Sur quelques mentions de Murciella gr. cuvillieri dans la région méditerranéenne.
Une brčve revue des mentions de la littérature permettra de compléter le tableau précédent, bien que les comparaisons précises ne soient généralement pas possibles, en l'absence d'étude exhaustive et de données statistiques.
Christodoulou (1967) a figuré une population provenant de l'île d'Astypalia. Les tests déroulés y sont nombreux, mais le grandissement trop faible (x8,5) des photographies ne permet pas d'identification. Il s'agit d'une population clairement différente de celle de męme provenance décrite ci-dessus (FPM249) ; il ne peut ętre exclu qu'elle soit ŕ rapprocher de Sigalveolina reicheli n. gen., n. sp.
Decrouez & Radoičić (1977) ont figuré 3 sections attribuées ŕ "Raadshoovenia" cuvillieri, comprises dans des couches témoignant de tendances émersives du Sénonien de Serbie occidentale. Leur morphologie comme la claire indication de la structure confluente permettent d'attribuer ces sections ŕ Sigalveolina ovoidea. Il s'agirait de la seule mention de cette espčce en dehors des plates-formes périadriatiques.
Luperto Sinni & Ricchetti (1978) décrivent une succession connue par forage en Italie du sud, oů "Raadshoovenia" salentina et Murciella cuvillieri apparaissent ensemble, au-dessus d'une passée ŕ Orbitoides tissoti et O. media. Les tests de ces 2 espčces, en partie écrasées, sont figurés dans un état de conservation trčs médiocre mais paraissent correctement identifiées.
Fleury (1979a) a publié les photos de 2 groupes de sections rapportées ŕ M. gr. cuvillieri, appartenant ŕ des populations peu fournies et jamais retrouvées en d'autres gisements. L'une
("Murciella n. sp. 1", Pl. 2.21-24) provenant du Péloponnčse central (GDD,
Fig. 1 ![]() ) présente un bâti épais, un enroulement juvénile mixte et un déroulement marqué constituant un cas particulier du type Cuvillierinella. L'autre
("Murciella n. sp. 2", Pl. 2.16-19) a été figurée de nouveau sous le nom de Cuvillierinella sp. 2 in Fleury, 2016 (Fig. 8.10-13) ; son intéręt particulier consiste en ce qu'elle accompagne Pseudochubbina philippsoni (Fleury,
1977) sur la coupe du Klokova et se trouve ętre le plus ancien Rhapydioninidae qui s'y présente.
) présente un bâti épais, un enroulement juvénile mixte et un déroulement marqué constituant un cas particulier du type Cuvillierinella. L'autre
("Murciella n. sp. 2", Pl. 2.16-19) a été figurée de nouveau sous le nom de Cuvillierinella sp. 2 in Fleury, 2016 (Fig. 8.10-13) ; son intéręt particulier consiste en ce qu'elle accompagne Pseudochubbina philippsoni (Fleury,
1977) sur la coupe du Klokova et se trouve ętre le plus ancien Rhapydioninidae qui s'y présente.
Gušić & Jelaska (1990) ont publié (op. cit., Pl. 19, fig. 1), une bonne section déroulée de Murciella cuvillieri typique provenant de l'île Dalmate de Brać (Croatie).
Parente (1994, Pl. 1.4) a figuré, en provenance de la pointe méridionale de la péninsule Salento (Italie du sud), une section décentrée ŕ importante PRU attribuée ŕ M. cuvillieri dont la maille endosquelettique large pourrait appartenir ŕ une Cuvillierinella ; une section typique de C. salentina est par ailleurs figurée dans le męme travail (Pl. 1.3).
Velić (2007) a figuré quelques bonnes sections de M. cuvillieri (op. cit., Fig. 25.5-10), non douteuses bien qu'en partie écrasées, provenant de Makarska, sur le littoral croate, proche de Brać.
Vicedo et al. (2009) ont donné des figurations d'une population présentée comme topotypique, uniquement constituée de tests A trčs compatibles avec la population type. Son originalité est de révéler des espaces préseptaux équipés de trčs nombreux piliers ; la structure confluente y est trčs discrčte. Les tests B ne sont pas observés et les dimensions de proloculus ne sont pas données.
En conclusion, le genre Murciella constitue une entité relativement homogčne, bien représentée dans l'ensemble du domaine méditerranéen. Son association ŕ C. salentina en quelques localités confirme que ces deux taxons caractérisent une męme zone (CsB6a), ŕ laquelle les espčces propres ŕ la zone suivante (CsB6b) ne participent pas.
Genre Sigalveolina n. gen.
Espčce-type : "Murciella" renzi Fleury, 1979a.
Diagnose. Genre de la sous-famille Cuvillierinellinae, dimorphique (ou éventuellement trimorphique) : tests A entičrement planispiralés involutes suivi d'une PRU flabelliforme ŕ cylindrique exceptionnelle ou éventuellement tardive ; tests B presque entičrement involutes de grande taille. Endosquelette de type confluent précoce et clairement exprimé. Diffčre de Murciella, son prédécesseur dans le temps, par des tests A plus grands ŕ taille de proloculus égal, ses tests des 2 générations non déroulés ou trčs tardivement déroulés et sa structure confluente achevée. Diffčre de tous les autres genres par la perfection de sa structure confluente, et particuličrement de son contemporain Cyclopseudedomia par sa faible capacité de déroulement. Quatre espčces sont distinguées, apparemment toutes de męme âge (zone CsB6b), limitées aux alentours de la mer Adriatique.
Origine du nom. Préfixe en hommage ŕ Jacques Sigal, auteur, parmi bien d'autres travaux portant principalement sur les foraminifčres planctoniques, d'un ouvrage longtemps compulsé de synthčse sur les foraminifčres (1952), d'une proposition hétérodoxe de rčgles de nomenclature propres aux foraminifčres (1966) et d'un travail de mémoire sur les micropaléontologues français de la premičre moitié du vingtičme sičcle (1989). Le suffixe rappelle l'appartenance du nouveau genre ŕ la super-famille des Alveolinacea.
Sigalveolina renzi (Fleury, 1979a)
Diagnose. Espčce type du genre Sigalveolina caractérisée essentiellement par ses tests A dotés d'un indice d'aplatissement élevé, non ombiliqués et d'un enroulement lâche associé ŕ une trčs faible tendance au déroulement terminal. Tests B totalement involutes en forme de double massue en section axiale. Bâti fręle et endosquelette ŕ maille équidimensionnelle fine dont les logettes secondaires participent ŕ une structure confluente parfaite ; espace préseptal trčs étroit meublé de nombreux piliers. Distincte de M. gr. cuvillieri par son endosquelette, le grand diamčtre et la faible capacité de déroulement de ses tests A ainsi que par ses tests B involutes ; distincte des autres espčces rapportées au męme genre par ses tests A plats (Indice d'aplatissement de 3 ŕ 5) et par la finesse de sa maille endosquelettique.
Localisation, conditions de gisement. La population type est située par Fleury (1979a) et plus précisément 1980 (Fig. 36, gisement FB), dans l'île de Skhiza, ŕ la pointe sud-ouest du Péloponnčse. Sur une centaine au total, les lames minces de l'échantillon GGB184 ayant fourni le matériel type sont déposées auprčs de l'Université de Lyon, sous les nº FSL147 801 ŕ 831. Il s'agit d'un calcaire biomicrite wackestone-packstone, renfermant également de rares Milioles, Cuneolina sp., Rhapydionina fourcadei Fleury, 2014, Metacuvillierinella decastroi Fleury, 2016 (il s'agit de la localité type de ces deux espčces) et Fleuryana adriatica. L'espčce est connue en une dizaine d'autres localités de Grčce ; elle est signalée en Croatie, trčs douteuse dans la figuration de Gušić et Jelaska (1990, Pl. XIX : il s'agirait plutôt de M. cuvillieri) et non figurée, par Velić (2007).
Morphologie des tests, enroulement.
Tests A. Ces tests sont toujours largement majoritaires. Il s'agit de tests plats, involutes, pratiquement dépourvus d'ombilic, ŕ périphérie aigüe dans les premiers tours, plus arrondie, presque carrée dans les derniers. Le proloculus de section circulaire est d'un diamčtre moyen de 0,16 mm chez la riche population type; le flexostyle est étroit et court. L'enroulement strictement planispiralé est lâche, le pas de la spire s'accroît réguličrement sans que rien ne laisse supposer une tendance au déroulement final. La taille moyenne des tests avoisine 1,70 mm (50% des tests, les plus grands).
Tests B. Aucune section exactement centrée n'est connue. Les seules sections interprétables comme appartenant ŕ cette génération montrent des tests plus grands que les tests A (jusqu'ŕ 3 mm de diamčtre) dont la périphérie des premiers tours est aigüe et celle des derniers, trčs enveloppants, arrondie.
Endosquelette. La principale caractéristique de cette espčce est constituée par la finesse de sa maille endosquelettique, jamais approchée au sein des Rhapydioninidae, sinon par quelques représentants du genre Pseudedomia. Logettes primaires et secondaires sont d'une męme finesse et ne se distinguent que par leur orientation. Les primaires restent parallčles ŕ l'enroulement de la spire, alors que les secondaires, adoptent trčs précocement les obliquités et les interpénétrations caractéristiques de la structure confluente. La confusion est extręme dans les derničres loges des 2 générations, avec l'agrandissement du diamčtre des logettes secondaires. L'espace préseptal est trčs étroit, encombré de nombreux piliers, devenant presque indiscernable dans les sections axiales.
Au total, cette espčce est trčs originale, tant pour sa morphologie d'ensemble que pour la finesse de sa maille endosquelettique et l'absence de déroulement dans les 2 générations.
Variations autour du type : plusieurs populations plus ou moins proches du type par leur endosquelette ont été observées.
La population XGP304 (Fig. 8.13-14
![]() ), provenant du massif du Gavrovo fut récoltée dans un ensemble bréchique d'âge vraisemblablement paléocčne (Mavrikas,
1993, coupe Pirgi 1, p. 83) ; l'échantillon correspond ŕ un calcaire packstone-grainstone ŕ fenestrae, cimentation diagénétique et sédiments internes, ŕ débris de rudistes, Cuneolina sp., Milioles, Rhapydionina dercourti Fleury,
2014, R. fleuryi Vicedo & Caus,
2011 (voir R.
"bulbiformis" in Fleury, 2014, Fig. 10 pour l'ensemble des caractéristiques de cette espčce) et Cyclopseudedomia smouti A et B (voir
Fig. 11.2-3
), provenant du massif du Gavrovo fut récoltée dans un ensemble bréchique d'âge vraisemblablement paléocčne (Mavrikas,
1993, coupe Pirgi 1, p. 83) ; l'échantillon correspond ŕ un calcaire packstone-grainstone ŕ fenestrae, cimentation diagénétique et sédiments internes, ŕ débris de rudistes, Cuneolina sp., Milioles, Rhapydionina dercourti Fleury,
2014, R. fleuryi Vicedo & Caus,
2011 (voir R.
"bulbiformis" in Fleury, 2014, Fig. 10 pour l'ensemble des caractéristiques de cette espčce) et Cyclopseudedomia smouti A et B (voir
Fig. 11.2-3 ![]() ). Un lot de 30 lames minces ne montre que 7 sections centrées, dont le proloculus mesure en moyenne 0,21 (±0,03 mm) pour un diamčtre équatorial de 1,90 mm, marquant des différences avec le type, soulignées par un début de déroulement
(Fig. 8.14
). Un lot de 30 lames minces ne montre que 7 sections centrées, dont le proloculus mesure en moyenne 0,21 (±0,03 mm) pour un diamčtre équatorial de 1,90 mm, marquant des différences avec le type, soulignées par un début de déroulement
(Fig. 8.14 ![]() ).
).
La population I250 (Fig. 8.11-12
![]() ) provient de la région italienne située au sud-ouest de Gioia del Colle, aux abords d'un puits proche du bord de la route conduisant de Laterza ŕ Santeramo, ŕ mi-distance des lieux dits Mass. Di Santo et Sabbettoli
(Fig. 1
) provient de la région italienne située au sud-ouest de Gioia del Colle, aux abords d'un puits proche du bord de la route conduisant de Laterza ŕ Santeramo, ŕ mi-distance des lieux dits Mass. Di Santo et Sabbettoli
(Fig. 1 ![]() , localité A). Il s'agit de calcaires packstone-grainstone renfermant des débris de rudistes et gastéropodes, Cuneolina sp., des Milioles, Fanrhapydionina aff. flabelliformis in Fleury, 2014 et Metacuvillierinella decastroi. Un lot de 33 lames minces fournit 19 sections centrées ; le diamčtre du proloculus est nettement plus grand que celui du type (0,23±0,03 mm), alors que le diamčtre équatorial des tests en reste peu différent (1,80±0,20 mm). C'est la seule population de cette espčce dans laquelle une rotation de l'axe d'enroulement
("Slow axial rotation" in Fleury, 2016, p. 201) ait été observée
(Fig. 8.11
, localité A). Il s'agit de calcaires packstone-grainstone renfermant des débris de rudistes et gastéropodes, Cuneolina sp., des Milioles, Fanrhapydionina aff. flabelliformis in Fleury, 2014 et Metacuvillierinella decastroi. Un lot de 33 lames minces fournit 19 sections centrées ; le diamčtre du proloculus est nettement plus grand que celui du type (0,23±0,03 mm), alors que le diamčtre équatorial des tests en reste peu différent (1,80±0,20 mm). C'est la seule population de cette espčce dans laquelle une rotation de l'axe d'enroulement
("Slow axial rotation" in Fleury, 2016, p. 201) ait été observée
(Fig. 8.11 ![]() ).
).
La population GKL414 (Fleury, 1979a, Pl.
1, figs. 6, 9, 13-14, 16 ; ici Fig. 3.6 ![]() ), connue par seulement 8 sections centrées dans un lot de 80 lames a été attribuée par erreur ŕ l'espčce lors de sa création. Son endosquelette présente les męmes caractéristiques, son proloculus est du męme ordre de grandeur (0,14±0,03 mm) mais une nette tendance au déroulement relativement précoce ne semble pas compatible avec le type.
), connue par seulement 8 sections centrées dans un lot de 80 lames a été attribuée par erreur ŕ l'espčce lors de sa création. Son endosquelette présente les męmes caractéristiques, son proloculus est du męme ordre de grandeur (0,14±0,03 mm) mais une nette tendance au déroulement relativement précoce ne semble pas compatible avec le type.
Une unique grande section plate, dotée d'un endosquelette ŕ maille fine, de tours internes trčs aigus évoquant M. renzi et un enroulement de type pseudoévolute ou advolute trčs original été figurée sous le nom de
"Murciella n. sp. aff. renzi" (in Fleury, 1979a, Pl.
3, fig. 12 ; section reproduite ici sur la Fig. 16 ![]() , flanquée d'un point d'interrogation, ŕ droite du groupe de dessins représentant S. renzi) ; cette section provient d'une population (GGB155) dans laquelle l'espčce, trčs mal représentée, accompagne C. smouti, Rhapydionina dercourti et Metacuvillierinella decastroi. Cette section, avec celles qui sont évoquées dans les remarques précédentes, ouvre d'intéressantes perspectives sur les variations susceptibles d'affecter cette espčce et ses proches.
, flanquée d'un point d'interrogation, ŕ droite du groupe de dessins représentant S. renzi) ; cette section provient d'une population (GGB155) dans laquelle l'espčce, trčs mal représentée, accompagne C. smouti, Rhapydionina dercourti et Metacuvillierinella decastroi. Cette section, avec celles qui sont évoquées dans les remarques précédentes, ouvre d'intéressantes perspectives sur les variations susceptibles d'affecter cette espčce et ses proches.
Les quatre cas particuliers décrits ci-dessus ne sont pas assez bien documentés pour donner lieu ŕ de longues considérations mais montrent que la connaissance de formes diverses apparemment proches de S. renzi est loin d'ętre achevée et se trouve hors de portée du présent travail. De telles variations évoquent un complexe dont on doute qu'il puisse ętre démęlé aisément dans les affleurements de Grčce.
|
Figure 8
: Sigalveolina renzi et S.
methonensis. 1-14 : Sigalveolina
renzi et populations affines. 1-4, 11-13 :
Sections axiales centrées
de tests A; noter la périphérie aigüe des tours internes et la précocité du
cloisonnement ; la section 11 montre une rotation réguličre de l'axe
d'enroulement. 5 : Section
transverse d'une derničre loge (ou d'une PRU) montrant de nombreux piliers
préseptaux. 6 : Section oblique
centrée. 7-8 et 14: Sections équatoriales
centrées de tests A ; noter l'étroitesse de l'espace préseptal,
occupé par de nombreux piliers et la finesse du bâti, mettant en évidence la
structure confluente dčs les tours internes (7 : holotype). 9-10:
Sections axiales subcentrées de tests attribués ŕ la génération B ;
noter la périphérie aigüe des tours internes. (1-10 : GGB184, population
type ; 11-12 : I250 ; 13-14 : XGP304). 15-23 :
Sigalveolina
methonensis. 15-19 : Sections
axiales centrées de tests A ; notez la petite taille des proloculus, le bâti
plus épais et les logettes de diamčtre plus grand que chez S.
renzi. 20-22 : Sections équatoriales centrées de tests A (22 :
holotype). 23-24 : Sections axiales de tests B vraisemblables ; noter
la périphérie arrondie des tours internes. Toutes : GGB184, population
type. Échelles graphiques : 1 mm. |
Sigalveolina methonensis (Fleury, 1979a)
(Fig. 8.15-23 ![]() )
)
Diagnose. Espčce du genre Sigalveolina ŕ proloculus A de petite taille, enroulement trčs serré et capacité de déroulement nulle. Tests B vraisemblables de grande taille, involutes avec partie terminale déroulée. Bâti et endosquelette relativement épais n'entravant pas le développement d'une structure confluente précoce et parfaite. Distincte de M. gr. cuvillieri par son diamčtre équatorial un peu plus grand, l'absence de déroulement chez les tests A et par sa structure confluente bien développée ; se distingue des autres espčces du genre Sigalveolina n. gen. par le proloculus de faible taille et la spire serrée de ses tests A mais ressemble aux tests de męme génération non déroulés de S. reicheli.
Cette espčce n'a pas été retrouvée depuis sa création ce qui, joint au fait qu'elle accompagne S. renzi dans son gisement type, pourrait la rendre suspecte de n'ętre qu'un variant extręme de sa compagne ; on note cependant que rien n'indique sa présence dans les autres populations de S. renzi. Son matériel type est déposé auprčs de l'Université de Lyon sous les nº FSL147803 ŕ 806, 810 ŕ 817, 819-820 et 823.
Localisation, conditions de gisement. Son gisement type (et unique) est le męme que celui du type de S. renzi.
Morphologie des tests, enroulement.
Tests A. Le proloculus de section circulaire (0,13 mm de diamčtre) est plus petit que chez S. renzi et dans la moyenne des M. gr. cuvillieri ; le flexostyle est ŕ peine discernable. Les tests sont relativement plats, faiblement biombiliqués dans les sections parfaitement axiales. Ils sont planispiralés ŕ l'ordinaire, mais une section montre (Fleury, 1979a, Pl. 2.10) un aspect déjŕ observé chez S. renzi
(Fig. 8.2 ![]() ) évoquant un type d'enroulement propre ŕ d'autres taxons de la sous-famille
("Slow axial rotation", Fleury, 2016, p. 201). Le diamčtre équatorial est sensiblement plus petit que celui de S. renzi et se situe au niveau des plus grandes Murciella gr. cuvillieri (1,35 mm en moyenne). Aucune section ne témoigne d'un déroulement.
) évoquant un type d'enroulement propre ŕ d'autres taxons de la sous-famille
("Slow axial rotation", Fleury, 2016, p. 201). Le diamčtre équatorial est sensiblement plus petit que celui de S. renzi et se situe au niveau des plus grandes Murciella gr. cuvillieri (1,35 mm en moyenne). Aucune section ne témoigne d'un déroulement.
Tests B. Comme chez S. renzi, on ne connaît pas de sections centrées. Une grande section, subcentrée
(Fig. 8.24 ![]() ) est distincte de celles qui sont attribuées ŕ S. renzi par les tours internes de contour arrondi. Une autre
(Fig. 8.23
) est distincte de celles qui sont attribuées ŕ S. renzi par les tours internes de contour arrondi. Une autre
(Fig. 8.23 ![]() ) montre une partie terminale de type évolute ou pseudoévolute trčs tardive ; elle est fortement décentrée, mais ce qu'on y distingue, des tours ŕ contour arrondi et des logettes de diamčtre important, la rend compatible avec la précédente.
) montre une partie terminale de type évolute ou pseudoévolute trčs tardive ; elle est fortement décentrée, mais ce qu'on y distingue, des tours ŕ contour arrondi et des logettes de diamčtre important, la rend compatible avec la précédente.
Endosquelette. La maille endosquelettique est plus large que celle de M. renzi, en particulier en raison du diamčtre plus important des logettes primaires. La structure confluente apparaît trčs précocement dans toutes les sections équatoriales. Comme chez S. renzi, la structure devient plus lâche et plus confuse dans les derničres loges des tests B.
Au total, malgré les doutes issus de sa coexistence avec S. renzi, l'espčce en paraît indépendante en raison de la taille du proloculus A et sa spire serrée. Si ses mensurations semblent la rapprocher de M. gr. cuvillieri, sa structure confluente bien exprimée et son refus du déroulement l'en éloignent clairement. Des petits tests de S. reicheli n. sp. pourraient ressembler ŕ cette espčce mais, dans l'ensemble, ils sont de plus grande taille, présentent une spire plus lâche et surtout sont plus sensibles au déroulement.
Sigalveolina ovoidea (Fleury, 1979a)
(Figs. 3.10 ![]() ,
9.1-20
,
9.1-20 ![]() )
)
Diagnose. Espčce du genre Sigalveolina caractérisée par son faible indice d'aplatissement et sa faible capacité de déroulement chez les tests A (dont certains, ŕ petit proloculus, restent totalement involutes malgré leur grande taille). Bâti et endosquelette relativement épais, structure confluente parfaitement réalisée. Tests B présentant un déroulement tardif de type flabelliforme. Distincte de M. gr. cuvillieri par son endosquelette et des autres espčces du genre Sigalveolina par le volume ovoďde de ses tests A.
Cette espčce, alors rapportée au genre Murciella, fut décrite ŕ partir d'une seule population provenant du Péloponnčse central (Fleury, 1979a, Pl. 5, figs. 1-15). Son matériel type est déposé auprčs de l'Université de Lyon sous les nº FSL147846 ŕ 853. L'espčce était principalement caractérisée par son enroulement strictement involute et son volume ovoďde. On y distinguait 2 types de tests :
des tests ŕ gros embryon, d'un diamčtre équatorial de moins de 1 mm pour 2 ŕ 4 tours de spire, pour un proloculus de 0,18 mm en moyenne ;
des tests ŕ petits embryon de diamčtre équatorial compris entre 1 et 2 mm pour un nombre double de tours de spire et un proloculus de 0,11 mm en moyenne.
Localisation, conditions de gisement. Outre le matériel type, on dispose de nouvelles populations (XGP301, 302 et 471), en provenance du massif du Gavrovo, découvertes par Mavrikas
(1993, p. 81) dans un ensemble bréchique ayant par ailleurs livré plusieurs populations de S. renzi et C. klokovaensis. L'espčce se trouve dans 2 types de facičs. Ses représentants sont peu nombreux et dispersés dans une biomicrite
wackestone (XGP301 et 471) ŕ oogones de charophytes, gastéropodes (dissouts et emplis de silts vadoses), ostracodes,
"Discorbis", Cuneolina sp., quinquéloculines banales rares, Rhapydionina gr. dercourti et Pseudonummoloculina aff. kalantarii Schlagintweit & Rashidi 2016
(Fig. 9.22-25 ![]() ), espčce observée exclusivement en compagnie de S. ovoidea. On les trouve par ailleurs trčs abondants dans une biomicrite packstone (XGP302) ŕ rares oogones de charophytes, débris de gastéropodes et rudistes, rares Cuneolina sp., quinquéloculines banales et F. adriatica.
), espčce observée exclusivement en compagnie de S. ovoidea. On les trouve par ailleurs trčs abondants dans une biomicrite packstone (XGP302) ŕ rares oogones de charophytes, débris de gastéropodes et rudistes, rares Cuneolina sp., quinquéloculines banales et F. adriatica.
Morphologie des tests, enroulement.
Tests A. Les tests sont de volume général ovoďde, ŕ périphérie plus ou moins arrondie ou anguleuse, apparaissant profondément biombiliqués en section strictement axiale. La plupart d'entre eux sont involutes, mais au moins deux grands types doivent ętre distingués :
Les tests correspondant aux plus grands proloculus sont dans l'ensemble strictement involutes, mais quelques rares spécimens montrent une tendance au déroulement final
(Fig. 9.7, .14-15 ![]() ). Aucune section circulaire n'est connue dans les riches populations examinées, ce qui conduit ŕ supposer que les PRU (observées uniquement dans des tests coupés axialement), sont toujours de type flabelliforme, probablement pseudoévolute. Le proloculus de ces tests est d'un diamčtre moyen de 0,16 mm pour des diamčtres équatoriaux de l'ordre de 1,50 mm pour 50% de l'effectif choisi parmi les plus grands, ce qui correspond sensiblement aux données de la population type.
). Aucune section circulaire n'est connue dans les riches populations examinées, ce qui conduit ŕ supposer que les PRU (observées uniquement dans des tests coupés axialement), sont toujours de type flabelliforme, probablement pseudoévolute. Le proloculus de ces tests est d'un diamčtre moyen de 0,16 mm pour des diamčtres équatoriaux de l'ordre de 1,50 mm pour 50% de l'effectif choisi parmi les plus grands, ce qui correspond sensiblement aux données de la population type.
Les tests correspondant aux plus petits proloculus, plus rares, se rangent en deux groupes. Les uns, trčs rares, restent totalement involutes et atteignent de grandes tailles en multipliant les tours de spire
(Fig. 9.6, .10 ![]() ). Les autres, plus abondants, mais rarement coupés en leur centre, présentent de grands tests apparemment entičrement planispiralés, développant de grands PRU flabelliformes de type pseudoévolute
(Fig. 9.17-19
). Les autres, plus abondants, mais rarement coupés en leur centre, présentent de grands tests apparemment entičrement planispiralés, développant de grands PRU flabelliformes de type pseudoévolute
(Fig. 9.17-19 ![]() ) ; ils pourraient représenter une éventuelle seconde génération A. Ils ne sont pas pris en compte dans les statistiques.
) ; ils pourraient représenter une éventuelle seconde génération A. Ils ne sont pas pris en compte dans les statistiques.
Tests B. Une seule section faiblement décentrée (Fig.
9.20 ![]() ) est susceptible de représenter cette génération. Elle ressemble ŕ celles qui ont été attribuées ŕ S. renzi, en ce qu'elle est strictement involute, de grande taille, mais les tours internes présentent une périphérie arrondie rappelant les tests A.
) est susceptible de représenter cette génération. Elle ressemble ŕ celles qui ont été attribuées ŕ S. renzi, en ce qu'elle est strictement involute, de grande taille, mais les tours internes présentent une périphérie arrondie rappelant les tests A.
Endosquelette. Le bâti est relativement épais, comparable ŕ celui de S. methonensis ou de certaines M. gr. cuvillieri. Ordinairement, le diamčtre des primaires est supérieur ŕ celui des secondaires, mais il existe des exemples dans lesquels les deux ensembles partagent le męme diamčtre
(Fig. 9.1 ![]() ). Ce diamčtre augmente dans les derničres loges des grands tests. La structure confluente est souvent masquée dans les premiers tours de spire en raison de l'enroulement trčs serré, en particulier dans les sections équatoriales. Cette structure est cependant trčs clairement exprimée dans les derničres loges de tous les grands tests.
). Ce diamčtre augmente dans les derničres loges des grands tests. La structure confluente est souvent masquée dans les premiers tours de spire en raison de l'enroulement trčs serré, en particulier dans les sections équatoriales. Cette structure est cependant trčs clairement exprimée dans les derničres loges de tous les grands tests.
Au total, cette espčce est bien distincte des précédentes par sa morphologie d'ensemble et en particulier son indice d'aplatissement, bien que la plupart des données statistiques ne soient pas trčs éloignées de celles de S. methonensis (Fig. 15 ![]() ). Sa plus grande particularité est constituée par la coexistence de 2 sortes de tests de type A qui pourraient ętre interprétée comme la conséquence d'un trimorphisme, pour autant que la génération B soit correctement identifiée.
). Sa plus grande particularité est constituée par la coexistence de 2 sortes de tests de type A qui pourraient ętre interprétée comme la conséquence d'un trimorphisme, pour autant que la génération B soit correctement identifiée.
Nota : Sur la présence de Pseudonummoloculina aff. kalantarii Schlagintweit & Rashidi,
2016
(Fig. 9.21-24 ![]() ), dans deux échantillons renfermant S. ovoidea du massif du Gavrovo. On observe 16 sections sur un lot de 30 lames de l'échantillon XGP301 et une quinzaine sur un lot de 18 lames de l'échantillon XGP471 (récolté en éboulis). Il s'agit de tests massifs de type miliolin ŕ paroi épaisse, ovoďdes, biombiliqués, ŕ tours de spire serrés, comme chez la plupart des Alveolinidae. Les ouvertures semblent disposées en deux rangées
(Fig. 9.24
), dans deux échantillons renfermant S. ovoidea du massif du Gavrovo. On observe 16 sections sur un lot de 30 lames de l'échantillon XGP301 et une quinzaine sur un lot de 18 lames de l'échantillon XGP471 (récolté en éboulis). Il s'agit de tests massifs de type miliolin ŕ paroi épaisse, ovoďdes, biombiliqués, ŕ tours de spire serrés, comme chez la plupart des Alveolinidae. Les ouvertures semblent disposées en deux rangées
(Fig. 9.24 ![]() , au sud-ouest du centre de la section). Des protubérances visibles dans les sections axiales de plusieurs loges, triangulaires en section, indiquent l'existence d'un cloisonnement rudimentaire ; celui-ci est fréquent dans la partie des loges proche du tour précédent ; la section
Figure 9.24
, au sud-ouest du centre de la section). Des protubérances visibles dans les sections axiales de plusieurs loges, triangulaires en section, indiquent l'existence d'un cloisonnement rudimentaire ; celui-ci est fréquent dans la partie des loges proche du tour précédent ; la section
Figure 9.24 ![]() (dernier tour, ŕ l'ouest du centre) montre cependant que ce cloisonnement peut exister au
"toit" de la loge (partie proche du tour suivant). Un dimorphisme de génération est clairement exprimé par des tests A ŕ proloculus sphérique suivi de loges streptospiralées et des tests B ŕ partie juvénile de type quinquéloculin passant ŕ un stade final planispiralé. L'intéręt de cet organisme est souligné dans la conclusion.
(dernier tour, ŕ l'ouest du centre) montre cependant que ce cloisonnement peut exister au
"toit" de la loge (partie proche du tour suivant). Un dimorphisme de génération est clairement exprimé par des tests A ŕ proloculus sphérique suivi de loges streptospiralées et des tests B ŕ partie juvénile de type quinquéloculin passant ŕ un stade final planispiralé. L'intéręt de cet organisme est souligné dans la conclusion.
|
Figure 9
: 1-20 : Sigalveolina
ovoidea. 1-10, 14-15 :
Sections axiales centrées de tests A illustrant la variété des morphologies ;
noter le cloisonnement précoce dans tous les cas ; le flexostyle est
discernable en 1, 3 et 9 (voir aussi 11) ; la structure confluente n'est
discernable que par la confusion apparente de l'arrangement des logettes (voir
une section des tours internes, Fig. 3.10 |
Sigalveolina reicheli n. sp.
(Fig. 10.1-20 ![]() )
)
Diagnose. Espčce du genre Sigalveolina ŕ proloculus de taille médiocre associé ŕ une tardive capacité ŕ développer une grande PRU cylindrique ou flabelliforme chez les tests A. Structure confluente bien réalisée pour un bâti et un endosquelette relativement épais. Tests B mal connus, apparemment non déroulés, comme ceux des autres espčces du męme genre. Distincte de M. gr. cuvillieri par sa structure confluente bien caractérisée pour des proloculus de męme taille et des autres espčces du męme genre par la grande différence de taille entre la partie involute et celle des tests déroulés ; ses plus petits tests miment ceux de S. methonensis, indiquant ainsi une vraisemblable étroite parenté de ces deux espčces.
Origine du nom. En hommage ŕ Manfred Reichel, "pčre spirituel" des artisans de l'étude des Alveolinacea, dont l'ouvrage de 1964 demeure un modčle inégalé de précise concision. On lui doit quelques mots sur les "Cosinella" de Ch. Schlumberger dont il observa une partie du matériel et chez lequel il reconnut un "dernier tour de Peneroplis" (Reichel, 1937, p. 136).
Matériel type. Plus d'une centaine de sections observées dans 75 lames minces numérotées YUA1. L'holotype
(Fig. 10.19 ![]() ) est compris dans une lame portant le numéro de fabrication 9545, déposé auprčs de l'Université des Sciences et Techniques de Lille sous le numéro 3150. Il correspond ŕ la section équatoriale d'un test de génération A ŕ proloculus de taille moyenne (0,16 mm de diamčtre) de section circulaire et grande PRU flabelliforme. Le diamčtre maximum du test est de 2,65 mm. Il est accompagné des sections provenant du męme échantillon apparaissant sur la
Figure 10.1-4, .6-8, .10-11, .13-19
) est compris dans une lame portant le numéro de fabrication 9545, déposé auprčs de l'Université des Sciences et Techniques de Lille sous le numéro 3150. Il correspond ŕ la section équatoriale d'un test de génération A ŕ proloculus de taille moyenne (0,16 mm de diamčtre) de section circulaire et grande PRU flabelliforme. Le diamčtre maximum du test est de 2,65 mm. Il est accompagné des sections provenant du męme échantillon apparaissant sur la
Figure 10.1-4, .6-8, .10-11, .13-19 ![]() , comprises dans les lames portant les numéros USTL3130 ŕ 53 (comprenant également des sections de Cyclopseudedomia mavrikasi n. sp.). Le reste du matériel sera déposé avec l'ensemble de la collection de l'auteur, dans les męmes lieux.
, comprises dans les lames portant les numéros USTL3130 ŕ 53 (comprenant également des sections de Cyclopseudedomia mavrikasi n. sp.). Le reste du matériel sera déposé avec l'ensemble de la collection de l'auteur, dans les męmes lieux.
Bignot (1972) a figuré sous le nom de "Raadshoovenia cuvillieri" plusieurs sections (op. cit., Pl. 15, figs. 1-7) de son matériel GB994, qui participent clairement ŕ l'espčce qui est décrite ici. En outre, Hamaoui & Fourcade (1973, Pl. 3, figs. 2-4 ; Pl. 5, fig. 3 ; Pl. 11, fig. 1) ont figuré sous le nom de "Murciella cuvillieri" le mythique matériel istriote de Ch. Schlumberger souvent cité, bien que ni décrit ni figuré, dont le Museum National d'Histoire Naturelle de Paris conserve une partie du matériel sous le nom de "Cosinella". Par ailleurs, Vicedo et al. (2009, Figs. 9.1-10 et 19) ont figuré des sections d'un organisme de męme provenance et de męme apparence que le matériel décrit ici, sous le nom de "Murciella renzi" ; il ne peut s'agir de cette espčce qui présente un endosquelette de maille beaucoup plus fine et ne possčde de stade déroulé qu'exceptionnel et trčs bref.
Localisation, conditions de gisement. L'échantillon présentant la plus riche population (YUA4) provient d'un gisement situé en Slovénie, ŕ environ 10 km au nord/nord-est de Trieste
(Fig. 1 ![]() ), précisément au bord de la route joignant Dutovlje ŕ Kreplje, au lieu oů Bignot (1972, p. 62) situe son échantillon GB994. Les calcaires surmontent des affleurements ŕ K. tergestina et les couches environnant l'échantillon renferment des traces de charbon, ainsi que des charophytes. Les listes de fossiles données sous forme synthétique par Bignot ne permettent pas de déduire la position de l'échantillon par rapport ŕ Rhapydionina liburnica (Stache,
1889) citée aux environs. On observe, dans une riche biomicrite wakestone-packstone, des Cuneolina sp., de trčs nombreuses
"Bigenerina et/ou Bolivinopsis" (figurées in Bignot, 1972, Pl. 15,
figs. 8-9 ; Pl. 16, fig. 9), des Milioles, de rares F. adriatica et Cyclopseudedomia mavrikasi n. sp., décrite plus loin. Les sections de la nouvelle espčce n'y sont pas rares (plusieurs par lame) mais souvent difficiles ŕ interpréter en raison de leur conservation défectueuse.
), précisément au bord de la route joignant Dutovlje ŕ Kreplje, au lieu oů Bignot (1972, p. 62) situe son échantillon GB994. Les calcaires surmontent des affleurements ŕ K. tergestina et les couches environnant l'échantillon renferment des traces de charbon, ainsi que des charophytes. Les listes de fossiles données sous forme synthétique par Bignot ne permettent pas de déduire la position de l'échantillon par rapport ŕ Rhapydionina liburnica (Stache,
1889) citée aux environs. On observe, dans une riche biomicrite wakestone-packstone, des Cuneolina sp., de trčs nombreuses
"Bigenerina et/ou Bolivinopsis" (figurées in Bignot, 1972, Pl. 15,
figs. 8-9 ; Pl. 16, fig. 9), des Milioles, de rares F. adriatica et Cyclopseudedomia mavrikasi n. sp., décrite plus loin. Les sections de la nouvelle espčce n'y sont pas rares (plusieurs par lame) mais souvent difficiles ŕ interpréter en raison de leur conservation défectueuse.
Morphologie des tests, enroulement.
Tests A. Sur l'ensemble de la population, le diamčtre moyen du proloculus, de section souvent ovale, est de 0,145 mm ; les diamčtres équatoriaux des plus grands tests (50% des mesures) s'établissent ŕ 1,90 mm, alors que les parties involutes des tests sont limitées ŕ 1,14 mm, ce qui traduit la forte tendance au déroulement tardif. Rappelons que ce męme diamčtre est celui de la moyenne des autres espčces du genre, qui n'est pas atteint par les moyennes des tailles des populations de M. gr. cuvillieri.
Bien qu'une limite tranchée ne puisse ętre définie, on peut tenter de distinguer 2 types de tests :
Les uns, ŕ proloculus de grande taille, ont un aspect nautiloďde évoquant la diversité des morphologies de Murciella gr. cuvillieri ; certains marquent cependant une certaine tendance au déroulement, probablement pseudoévolute.
Les autres, peu nombreux, ŕ proloculus petit (Fig.
10.15, .17, .20 ![]() ) ou moyen
(Fig. 10.16, .18-19
) ou moyen
(Fig. 10.16, .18-19 ![]() ) produisent une PRU, soit de type flabelliforme pseudoévolute en général, soit cylindrique ; ce dernier cas est peu fréquent si l'on en juge par l'extręme rareté des sections circulaires ou ovales observées dans les lames (3 au total).
) produisent une PRU, soit de type flabelliforme pseudoévolute en général, soit cylindrique ; ce dernier cas est peu fréquent si l'on en juge par l'extręme rareté des sections circulaires ou ovales observées dans les lames (3 au total).
Tests B. Une seule section est connue (voir Fig.
16 ![]() , en haut ŕ gauche du groupe de dessins, indication du grandissement d'origine respectée), figurée par Hamaoui & Fourcade (1973, Pl. 2,
figs. 2-3) en provenance du matériel de Schlumberger ; il s'agit d'un test non déroulé. On ne sait pas ŕ quel type de test adulte appartiendrait la section de partie juvénile publiée par Vicedo et al. (2009, Fig. 9).
, en haut ŕ gauche du groupe de dessins, indication du grandissement d'origine respectée), figurée par Hamaoui & Fourcade (1973, Pl. 2,
figs. 2-3) en provenance du matériel de Schlumberger ; il s'agit d'un test non déroulé. On ne sait pas ŕ quel type de test adulte appartiendrait la section de partie juvénile publiée par Vicedo et al. (2009, Fig. 9).
Bâti, endosquelette. Le bâti est comparable ŕ celui des M. gr. cuvillieri les plus massives, comme des S. methonensis ou ovoidea, moins fręle que chez S. renzi. Les logettes secondaires apparaissent d'un diamčtre inférieur ŕ celui des primaires et donnent souvent l'impression d'une organisation désordonnée dans les tests coupés axialement ou dans les sections transverses de PRU. Leur organisation selon le mode de la structure confluente est déjŕ discernable dans les tours internes et devient évidente dans les sections équatoriales des derničres loges des grands tests. L'espace préseptal est étroit et occupé par de nombreux piliers.
Au total, l'espčce ne se démarque pas clairement de M. gr. cuvillieri par la morphologie de ses petits tests, ni par la taille de son proloculus mais se distingue clairement par sa structure confluente bien exprimée. De męme, un rapprochement peut-ętre fait avec S. methonensis dont elle partage la taille du proloculus, l'enroulement serré des premiers tours et le profil en section axiale. Sa forte et tardive tendance au déroulement l'en distingue cependant. L'existence de deux types de tests A supposés rappelle le cas de S. ovoidea.
En conclusion, le genre Sigalveolina n. gen., s'il a été longtemps confondu avec le précédent, trouve son unité dans la structure confluente bien réalisée, partagée par quatre espčces distinctes par leur variété morphologique; celles-ci ne sont par ailleurs qu'exceptionnellement déroulées dans leurs générations A, sauf S. reicheli n. sp. qui ne cčde ŕ cette tendance qu'ŕ un stade de développement trčs tardif. Les tests B marquent également une tendance ŕ rester longuement involute qui contraste avec le déroulement précoce du genre précédent. Ces espčces caractérisent, en conjonction avec Cyclopseudedomia, la zone CsB6b.
|
Figure 10
: Sigalveolina reicheli
n. sp. 1-9 : Sections axiales centrées de tests A ŕ grand
proloculus ; le flexostyle est visible en 3, 4, 5 et 8 ; le
cloisonnement est précoce dans tous les cas, les piliers préseptaux sont
particuličrement visibles en 2 ; 1 témoigne d'une tendance au déroulement
final. 10-12 : Sections équatoriales
centrées ŕ grand proloculus ; la structure confluente est ŕ peine
discernable sur 10 en raison de l'état médiocre de la conservation du test
mais plus certaine sur 11 et 12. 13-14 :
Sections de PRU cylindriques de tests A vraisemblables. 15-19 : Sections équatoriales de tests ŕ proloculus petit (15
et 17) ŕ moyen (16, 18-19) et PRU cylindriques (16-18) ou flabelliformes (15 et
19) ; la structure confluente est clairement exprimée dans tous les cas, dčs
les premier tours de spire (19 : holotype). 20 : Section axiale centrée d'un test ŕ petit proloculus
(probablement double) et PRU flabelliforme. Échelles graphiques :
1 mm. |
Genre Cyclopseudedomia Fleury, 1974
Espčce-type : Cyclopseudedomia smouti Fleury, 1974.
Diagnose. Genre de la sous-famille Cuvillierinellinae, dimorphique : tests A ŕ grand proloculus, trčs précocement déroulés ŕ loges évolutes constituant une PRU flabelliforme ou cylindrique; tests B essentiellement évolutes ŕ loges formant de grands tests plats en forme d'éventails ou męme discoďdes. Endosquelette de type coaxial passant ŕ une structure confluente éventuelle, toujours fruste. Diffčre de tous les autres genres par le déroulement de ses deux générations. Quatre espčces sont distinguées, vraisemblablement toutes de męme âge (zone CsB6b), limitées ŕ la région périadriatique.
Ce genre a été créé ŕ partir d'une seule espčce différente de tous les taxons connus ŕ l'époque, mais qui semblait partager avec certains représentants alors admis du genre Pseudedomia une grande capacité de déroulement ; sa génération B, représentée par des tests discoďdes imposait un préfixe caractérisant cette particularité. On sait maintenant que ses tests ŕ loges annulaires sont uniques dans la famille comme dans la sous-famille. Il s'agissait du type extręme d'une tendance évolutive maintenant reconnue chez 3 autres espčces.
Cyclopseudedomia smouti Fleury, 1974
(Figs. 2.6-11 ![]() ,
3.14
,
3.14 ![]() , 11.1-18
, 11.1-18 ![]() )
)
L'espčce type du genre est sans doute le plus remarquable taxon de la famille. Aucun autre ne l'approche par l'originalité de ses deux générations : ses tests B, en particulier, sont seuls ŕ produire des loges annulaires et les tests A n'en sont pas éloignés. En outre, c'est ŕ peine si deux autres espčces européennes (Fanrhapydionina flabelliformis et Cyclopseudedomia klokovaensis) partagent les grandes dimensions de son proloculus A. Le matériel type décrit par Fleury (1974) était constitué par les tests des deux générations clairement identifiées et largement illustrées. On notera que Vicedo et al.
(2011), revisitant la localité type de ce taxon, ont jugé utile d'en donner de nombreuses photos et une nouvelle description trčs fidčles au travail de 1974 ; ces auteurs n'ont cependant pas observé les détails qui permettent de construire un modčle structural tel que celui de la
Figure 11.18 ![]() .
.
Diagnose. Espčce type du genre Cyclopseudedomia, dimorphique et costulée : tests A comportant un petit nombre de loges involutes enveloppant un proloculus de grande taille, suivies de loges pseudoévolutes ŕ évolutes formant une PRU flabelliforme semi-circulaire ; tests B constitués essentiellement de loges annulaires formant un test discoďde sensiblement plat, de grand diamčtre. Endosquelette central comprenant des logettes secondaires irréguličres par leur disposition et leurs orientations passant ŕ un type confluent fruste. Distincte des autres espčces du genre par le mode de déroulement propre ŕ ses tests des deux générations.
Localisation, conditions de gisement. Lors de sa création, l'espčce était connue de trois gisements, deux de la région de Pylos (GGB) et un du massif du Klokova (GKL). Son matériel type est déposé auprčs de l'Université de Lyon sous les nº FSL115656 ŕ 666. De nouvelles localités ont été découvertes depuis, prčs de Pylos (GGB520-521) et dans le massif du Gavrovo (XGP166 et 304). Dans tous ces échantillons, les tests des deux générations sont associés, accompagnés de Milioles, Sigalveolina renzi (et/ou formes affines) et Rhapydionina gr. dercourti. Les sédiments sont des biomicrites wackestone renfermant des tests de mollusques (rudistes et gastéropodes) souvent dissous par les eaux météoriques et remplis de silts vadoses. La présence de rares oogones de charophytes signale la proximité de domaines émergés, annoncés par les conditions de la diagenčse.
Les diverses populations connues présentent des mensurations trčs homogčnes, qui ne nécessitent pas de descriptions individuelles. On ne s'étonnera pas que la plupart des figurations de ce travail soient issues de la population du type, qui avait fait l'objet de nombreuses préparations pour régler le mystčre des sections insolites des grands tests B du nouveau taxon associés ŕ ceux de la męme génération de Cuvillierinella aff. pylosensis, (attribuée ŕ C. salentina en 1974) alors tout ŕ fait inconnus chez ce genre ; c'est le seul exemple connu de cette association.
Morphologie, enroulement.
Les tests A (Figs. 2.7-11 ![]() , 11.1-9
, 11.1-9 ![]() ,
16
,
16 ![]() ) comportent un proloculus de grand diamčtre (0,32 mm en moyenne), ouvrant sur un flexostyle volumineux
(Figs. 2.9
) comportent un proloculus de grand diamčtre (0,32 mm en moyenne), ouvrant sur un flexostyle volumineux
(Figs. 2.9 ![]() ,
11.1
,
11.1 ![]() ) mais court, suivi d'un seul tour planispiralé involute nautiloďde presque sphérique de 6 ou 7 loges, puis d'une partie flabelliforme plate (jamais cylindrique) comprenant jusqu'ŕ plus de 10 loges pseudoévolutes puis évolutes
(Fig. 2.9
) mais court, suivi d'un seul tour planispiralé involute nautiloďde presque sphérique de 6 ou 7 loges, puis d'une partie flabelliforme plate (jamais cylindrique) comprenant jusqu'ŕ plus de 10 loges pseudoévolutes puis évolutes
(Fig. 2.9 ![]() )) dont les derničres peuvent ętre semi-circulaires.
)) dont les derničres peuvent ętre semi-circulaires.
Les tests B (Figs. 3.14 ![]() , 11.10-17
, 11.10-17 ![]() ,
16
,
16 ![]() ) comprennent, autour d'une partie initiale peu précisément décryptée (voir Fleury, 1974, Fig. 3), des loges embrassantes disposées ŕ la ressemblance des
"palmiers" des pâtissiers, devenant rapidement annulaires (Fleury, 1974, Pl.
52, fig. 1); l'épaisseur du test, trčs faible au centre, croît réguličrement et faiblement vers la périphérie
(Fig. 11.17
) comprennent, autour d'une partie initiale peu précisément décryptée (voir Fleury, 1974, Fig. 3), des loges embrassantes disposées ŕ la ressemblance des
"palmiers" des pâtissiers, devenant rapidement annulaires (Fleury, 1974, Pl.
52, fig. 1); l'épaisseur du test, trčs faible au centre, croît réguličrement et faiblement vers la périphérie
(Fig. 11.17 ![]() ), au contraire des tests de męme génération des Cuvillierinella associées (Fleury, 1974, Pl.
52, fig. 6 ; 2016, Fig. 14), d'épaisseur constante et organisés autour d'une forte nodosité centrale.
), au contraire des tests de męme génération des Cuvillierinella associées (Fleury, 1974, Pl.
52, fig. 6 ; 2016, Fig. 14), d'épaisseur constante et organisés autour d'une forte nodosité centrale.
Endosquelette. Il apparaît dčs le premier demi-tour de spire, c'est-ŕ-dire dčs les toutes premičres loges
(Figs. 2.10 ![]() ,
11.3-4
,
11.3-4 ![]() ). Les logettes primaires présentent un diamčtre ordinairement supérieur ŕ celui des secondaires
(Figs. 2.8
). Les logettes primaires présentent un diamčtre ordinairement supérieur ŕ celui des secondaires
(Figs. 2.8 ![]() ,
11.1, .8, .15
,
11.1, .8, .15 ![]() ), les unes et les autres étant de section circulaire lorsqu'elles sont coupées perpendiculairement ŕ leur axe ; mais les primaires peuvent apparaître oblongues
(Fig. 11.8
), les unes et les autres étant de section circulaire lorsqu'elles sont coupées perpendiculairement ŕ leur axe ; mais les primaires peuvent apparaître oblongues
(Fig. 11.8 ![]() ) ou presque rectangulaires lorsqu'elles sont coupées obliquement. Les primaires restent perpendiculaires aux septes
("parallčles au sens de l'enroulement" selon l'expression de Reichel,
1936) et sont disposées selon le mode
"continu", alignées d'une loge ŕ la suivante et réguličrement espacées, en général. Les secondaires apparaissent dans la plupart des sections comme noyées sans ordre dans la masse de l'endosquelette central, c'est le type de disposition nommé
"SSC" en l'absence de toute structure assimilable ŕ des "planchers"; les sections équatoriales des loges adultes montrent qu'elles ont tendance ŕ prendre des orientations obliques sur celle des primaires et qu'elles peuvent devenir coalescentes
(Figs. 2.14
) ou presque rectangulaires lorsqu'elles sont coupées obliquement. Les primaires restent perpendiculaires aux septes
("parallčles au sens de l'enroulement" selon l'expression de Reichel,
1936) et sont disposées selon le mode
"continu", alignées d'une loge ŕ la suivante et réguličrement espacées, en général. Les secondaires apparaissent dans la plupart des sections comme noyées sans ordre dans la masse de l'endosquelette central, c'est le type de disposition nommé
"SSC" en l'absence de toute structure assimilable ŕ des "planchers"; les sections équatoriales des loges adultes montrent qu'elles ont tendance ŕ prendre des orientations obliques sur celle des primaires et qu'elles peuvent devenir coalescentes
(Figs. 2.14 ![]() ,
11.6
,
11.6 ![]() en particulier). Il s'agit donc d'une structure confluente, qui peut ętre qualifiée de fruste puisqu'elle semble irréguličre et surtout que les 2 familles de logettes qui y participent ne sont pas orthogonales. L'espace préseptal est relativement étroit, occupant environ un cinquičme de la distance entre 2 septes consécutifs. Les cloisonnettes qui limitent les logettes primaires s'y terminent en pointe de section triangulaire
(Fig. 11.12, .15
en particulier). Il s'agit donc d'une structure confluente, qui peut ętre qualifiée de fruste puisqu'elle semble irréguličre et surtout que les 2 familles de logettes qui y participent ne sont pas orthogonales. L'espace préseptal est relativement étroit, occupant environ un cinquičme de la distance entre 2 septes consécutifs. Les cloisonnettes qui limitent les logettes primaires s'y terminent en pointe de section triangulaire
(Fig. 11.12, .15 ![]() ). Les piliers préseptaux forment une masse confuse ŕ leur enracinement dans l'endosquelette central puis s'individualisent ŕ proximité du septe
(Fig. 11.12
). Les piliers préseptaux forment une masse confuse ŕ leur enracinement dans l'endosquelette central puis s'individualisent ŕ proximité du septe
(Fig. 11.12 ![]() ). Les ouvertures primaires sont toujours disposées en dehors de l'axe des logettes correspondantes et présentent un axe oblique sur celui des logettes qui leur correspondent
(Figs. 2.11
). Les ouvertures primaires sont toujours disposées en dehors de l'axe des logettes correspondantes et présentent un axe oblique sur celui des logettes qui leur correspondent
(Figs. 2.11 ![]() ,
11.13
,
11.13 ![]() en particulier) : il en résulte que les logettes primaires de loges successives ne communiquent jamais directement entre elles
(Fig. 11.13-14
en particulier) : il en résulte que les logettes primaires de loges successives ne communiquent jamais directement entre elles
(Fig. 11.13-14 ![]() ), mais toujours par l'intermédiaire de l'espace préseptal.
), mais toujours par l'intermédiaire de l'espace préseptal.
Au total, cette espčce est la plus remarquable de toutes celles qui constituent la famille des Alveolinidae. Elle constitue, par son déroulement, un cas extręme auquel ses trois congénčres semblent se mesurer, ce qui impose le mode de présentation de leurs diagnoses.
|
Figure 11
: Cyclopseudedomia smouti, type du genre (voir aussi Fig. 2.6-11 |
Cyclopseudedomia klokovaensis (Fleury, 1979a)
(Figs. 3.13 ![]() ,
12.1-17
,
12.1-17 ![]() )
)
C'est au genre Murciella que cette espčce fut attribuée lors de sa création puisque la structure confluente n'était alors connue que chez le type de ce genre et quelques nouvelles espčces vraisemblablement apparentées (alors dénommées "Murciella" gr. renzi). Par la suite, ce caractčre étant reconnu dans divers contextes, il devint évident qu'il n'avait aucune signification générique, mais qu'il s'agissait d'un stade évolutif partagé par plusieurs groupes (Fleury & Fourcade, 1987). Dans ces conditions, pour les męmes auteurs (1990), sa forte tendance au déroulement rangeait clairement cette espčce aux côtés de C. smouti.
Diagnose. Espčce du genre Cyclopseudedomia, dimorphique, apparemment non costulée, proche du type par la taille de son proloculus A, le nombre de loges qui l'entourent et les tests B plats de grande taille; elle en diffčre par des tests A comportant une PRU flabelliforme ou cylindrique et des tests B dépourvus de loges annulaires. Structure confluente évidente bien qu'imparfaite.
Localisation, conditions de gisement. L'espčce était connue ŕ l'origine d'un seul échantillon du massif du Klokova (GKL76) ; son matériel type est déposé auprčs de l'Université de Lyon, sous les nº FSL147832 ŕ 845. Elle a été retrouvée en abondance par G. Mavrikas dans le massif du Gavrovo en 4 populations trčs homogčnes et comparables ŕ celle du type, bien que l'holotype apparaisse finalement assez marginal. Les deux plus riches populations nouvelles proviennent de calcaires bréchiques décrits par Mavrikas,
(1993, p. 81, coupe de Pirgi). Des calcaires de type packstone ŕ débris de rudistes et Milioles (XGP112-113) renferment les tests A en abondance; des biomicrites
wackestone ŕ petits débris (XGP115) ont livré de nombreux tests B.
Le peuplement est dans les deux cas presque monospécifique mais de rares débris d'Orbitoides, quelques sections de Cuneolina sp., Accordiella sp., et Sigalveolina aff. renzi ont été observées. L'espčce a été citée dans le massif du Mainalon en Péloponnčse (GDD,
Fig. 1 ![]() ), par Zambetakis-Lekkas
(1988).
), par Zambetakis-Lekkas
(1988).
Morphologie, enroulement.
Tests A. Le proloculus, de section ordinairement subcirculaire mais parfois d'allure bosselée, est d'un diamčtre moyen de 0,32 mm, avec un maximum de 0,52 mm (ce qui constitue un record absolu pour ce groupe). Le flexostyle, bien observé en plusieurs sections est court et volumineux
(Fig. 12.1-2 ![]() ) ; il est suivi de 1 ou 2 loges dépourvues de logettes secondaires, lesquelles apparaissent avant la fin du premier tour de spire
(Fig. 12.3
) ; il est suivi de 1 ou 2 loges dépourvues de logettes secondaires, lesquelles apparaissent avant la fin du premier tour de spire
(Fig. 12.3 ![]() ), accompagnées des piliers préseptaux
(Fig. 12.4
), accompagnées des piliers préseptaux
(Fig. 12.4 ![]() ). La partie spiralée involute correspondant ŕ environ 1 ŕ 1,5 tour de spire est suivie d'une PRU cylindrique ou flabelliforme
(Fig. 12.1-2
). La partie spiralée involute correspondant ŕ environ 1 ŕ 1,5 tour de spire est suivie d'une PRU cylindrique ou flabelliforme
(Fig. 12.1-2 ![]() ), de section circulaire ŕ oblongue ou rectangulaire ou en haltčre (épaissie aux 2 extrémités,
Fig. 12.12
), de section circulaire ŕ oblongue ou rectangulaire ou en haltčre (épaissie aux 2 extrémités,
Fig. 12.12 ![]() ) ; on y compte jusqu'ŕ 12 loges.
) ; on y compte jusqu'ŕ 12 loges.
Tests B. Les grands tests se trouvent ŕ l'état de débris dans les échantillons riches en tests A, mais abondent et sont entiers dans ceux oů les A sont rares. La partie juvénile n'a pas été observée en raison de sa faible épaisseur. Une section
(Fig. 12.13 ![]() ) montre pour les stades suivants une spire comparable ŕ celle des tests A puis des loges trčs embrassantes qui ne semblent pas susceptibles d'atteindre l'état annulaire. L'épaisseur de ces tests avoisine 0,2 mm au centre et 0,7 mm ŕ la périphérie ; le plus grand diamčtre observé dépasse 5 mm.
) montre pour les stades suivants une spire comparable ŕ celle des tests A puis des loges trčs embrassantes qui ne semblent pas susceptibles d'atteindre l'état annulaire. L'épaisseur de ces tests avoisine 0,2 mm au centre et 0,7 mm ŕ la périphérie ; le plus grand diamčtre observé dépasse 5 mm.
Endosquelette. Les premičres logettes secondaires apparaissent dčs le premier demi-tour de spire ; elles constituent alors une seule rangée et peuvent sembler limitées par un
"plancher", auquel correspond une rangée de piliers préseptaux réguličrement espacés
(Fig. 12.3-4 ![]() ). Le diamčtre des 2 ensembles de logettes n'est pas alors trčs différent et il ne l'est pas non plus dans certaines loges du stade déroulé
(Fig. 12.8
). Le diamčtre des 2 ensembles de logettes n'est pas alors trčs différent et il ne l'est pas non plus dans certaines loges du stade déroulé
(Fig. 12.8 ![]() , vers la droite et
Fig. 12.9
, vers la droite et
Fig. 12.9 ![]() vers le haut), si bien que l'on peut supposer que l'aspect oblong ŕ rectangulaire des logettes primaires dans de nombreuses sections soit lié ŕ une obliquité du plan de section sur leur axe ; le plus clair exemple est donné par la section de la
Figure 12.9
vers le haut), si bien que l'on peut supposer que l'aspect oblong ŕ rectangulaire des logettes primaires dans de nombreuses sections soit lié ŕ une obliquité du plan de section sur leur axe ; le plus clair exemple est donné par la section de la
Figure 12.9 ![]() qui montre des sections de logettes circulaires vers le haut, alors qu'elles sont trčs allongées dans la partie inférieure de la figure. Quoi qu'il en soit, il est peu de sections qui ne témoignent pas de la structure confluente. Elle n'est pas réellement discernable dans le premiers tour de spire, d'aspect souvent confus en section équatoriale, mais elle est clairement exprimée dans les parties évolutes de toutes les sections par les obliquités des 2 ensembles de logettes secondaires par rapport aux primaires
(Fig. 12.1-7
qui montre des sections de logettes circulaires vers le haut, alors qu'elles sont trčs allongées dans la partie inférieure de la figure. Quoi qu'il en soit, il est peu de sections qui ne témoignent pas de la structure confluente. Elle n'est pas réellement discernable dans le premiers tour de spire, d'aspect souvent confus en section équatoriale, mais elle est clairement exprimée dans les parties évolutes de toutes les sections par les obliquités des 2 ensembles de logettes secondaires par rapport aux primaires
(Fig. 12.1-7 ![]() ). Elle est également discernable dans les sections transverses de PRU de tests A par certaines orientations affectant les alignements des logettes secondaires et des piliers, rectilignes et obliques sur la muraille
(Fig. 12.8-9, .11
). Elle est également discernable dans les sections transverses de PRU de tests A par certaines orientations affectant les alignements des logettes secondaires et des piliers, rectilignes et obliques sur la muraille
(Fig. 12.8-9, .11 ![]() ). Les sections de tests B sont également sans ambiguďté, les sections équatoriales en particulier montrant les 2 ensembles de logettes se partageant deux directions franchement obliques (bien que non exactement orthogonales) sur l'axe des logettes primaires (discernables ŕ la marge gauche de la section
Fig. 12.14
). Les sections de tests B sont également sans ambiguďté, les sections équatoriales en particulier montrant les 2 ensembles de logettes se partageant deux directions franchement obliques (bien que non exactement orthogonales) sur l'axe des logettes primaires (discernables ŕ la marge gauche de la section
Fig. 12.14 ![]() ). Les diverses sections de type axial
"Ty" (Fig. 12.4, .15
). Les diverses sections de type axial
"Ty" (Fig. 12.4, .15 ![]() ) montrent bien la position des ouvertures primaires en dehors de l'axe des logettes correspondantes : comme chez C. smouti, ces logettes de 2 loges successives ne communiquent jamais directement d'une loge ŕ la suivante. L'espace préseptal est ordinairement étroit, mais l'exemple de la
Figure 12.2
) montrent bien la position des ouvertures primaires en dehors de l'axe des logettes correspondantes : comme chez C. smouti, ces logettes de 2 loges successives ne communiquent jamais directement d'une loge ŕ la suivante. L'espace préseptal est ordinairement étroit, mais l'exemple de la
Figure 12.2 ![]() montre qu'il peut occuper exceptionnellement jusqu'ŕ 2/5 de la distance entre les septes consécutifs ; la
Figure 12.16
montre qu'il peut occuper exceptionnellement jusqu'ŕ 2/5 de la distance entre les septes consécutifs ; la
Figure 12.16 ![]() montre un espace préseptal coupé selon l'orientation
"Tx", tout ŕ fait comparable ŕ celui de C. smouti (Fig.
11.12
montre un espace préseptal coupé selon l'orientation
"Tx", tout ŕ fait comparable ŕ celui de C. smouti (Fig.
11.12 ![]() ).
).
Au total, cette espčce est proche de C. smouti par sa forte et précoce tendance au déroulement évolute dans les deux générations. Elle s'en distingue par un déroulement plus modeste et une structure confluente mieux réalisée. Elle apparaît globalement plus "avancée" que C. hellenica et C. mavrikasi n. sp.
|
Figure 12
: Cyclopseudedomia klokovaensis. 1-2, 6-7 : Sections équatoriales
centrées de tests A représentatives des divers types de PRU ; toutes témoignent
clairement de la structure confluente ; noter en 6 les logettes primaires
ne communiquant jamais directement d'une loge ŕ la suivante. 3-5 :
Sections axiales centrées (tests A) ; noter la précocité du
cloisonnement (en 3) et des piliers préseptaux (en 4). 8-12 :
Sections transverses de PRU de type circulaire, ovale et rectangulaire (tests A
probables). 13 : Dessin schématique
d'une section de grand test flabelliforme (B probable).
14 : Portion de section peu oblique sur le plan d'étalement d'un
grand test B (voir aussi Fig. 3.13 |
Cyclopseudedomia hellenica Fleury, 1979b
(Fig. 13.1-21 ![]() )
)
Cette espčce constitue un intéressant intermédiaire entre l'ancętre de type Cuvillierinella et les espčces plus accomplies du genre Cyclopseudedomia.
Diagnose. Espčce du genre Cyclopseudedomia, dimorphique, partageant la costulation et la tendance évolutive du type, mais différente par plusieurs caractčres: taille du proloculus plus petite, nombre de loges déroulées plus faible, PRU flabelliforme ou cylindrique chez les tests A; loges peu embrassantes, n'atteignant vraisemblablement pas le stade semi-circulaire chez les tests B. Structure confluente apparemment absente.
Localisation, gisements. L'espčce n'a pas été retrouvée depuis sa création. Elle était fondée sur l'étude de quatre populations du Péloponnčse et du massif du Klokova ; son matériel d'origine est déposé auprčs de l'Université de Lyon, sous les nº FSL 147828 et 147854 ŕ 873. Les 2 plus riches de ces populations (GGB183 et GKL225) sont revisitées ici ŕ la lumičre des connaissances acquises sur les autres espčces du genre. Le calcaire wackestone renfermant la population type (GGB183) renferme de rares algues dasycladales, des Cuneolina sp., des Milioles et Rhapydionina aff. dercourti (in Fleury, 2014, Fig. 13). Le calcaire wackestone GKL225 réunit Cuneolina sp., Fleuryana adriatica, Sigalveolina renzi et Rhapydionina sp. ; des amas de pseudomorphoses d'anhydrite soulignent la proximité de milieux émergés.
Morphologie, enroulement (voir Fleury, 1979b, Figs. 1 et 3).
Tests A. Le proloculus sphérique de 0,25 mm de diamčtre en moyenne est suivi d'un flexostyle long d'un tiers de tour
(Fig. 13.9 ![]() ) et de section semi-circulaire
(Fig. 13.4
) et de section semi-circulaire
(Fig. 13.4 ![]() ). Les toutes premičres loges formant le premier demi-tour ne sont pas divisées
(Fig. 13.1-5, .9
). Les toutes premičres loges formant le premier demi-tour ne sont pas divisées
(Fig. 13.1-5, .9 ![]() ) mais les loges directement superposées au flexostyle le sont. Les loges suivantes participent ŕ une spire dont le pas s'accroît réguličrement sur environ 2 tours, avant que ne se constitue un court stade déroulé comprenant 2 ŕ 3 loges. Ce stade est ordinairement aplati
(Fig. 13.6, .8, .13-14
) mais les loges directement superposées au flexostyle le sont. Les loges suivantes participent ŕ une spire dont le pas s'accroît réguličrement sur environ 2 tours, avant que ne se constitue un court stade déroulé comprenant 2 ŕ 3 loges. Ce stade est ordinairement aplati
(Fig. 13.6, .8, .13-14 ![]() ), mais une section équatoriale dotée d'un petit proloculus (0,18 mm) et d'un stade juvénile streptospiralé est doté d'une partie terminale constituée de 3 loges cylindriques
(Fig. 13.10
), mais une section équatoriale dotée d'un petit proloculus (0,18 mm) et d'un stade juvénile streptospiralé est doté d'une partie terminale constituée de 3 loges cylindriques
(Fig. 13.10 ![]() ). L'ensemble du test est d'un diamčtre moyen de 1,07 mm avec un maximum observé de 1,88. De rares sections témoignent d'une fine costulation externe
(Fig. 13.8
). L'ensemble du test est d'un diamčtre moyen de 1,07 mm avec un maximum observé de 1,88. De rares sections témoignent d'une fine costulation externe
(Fig. 13.8 ![]() ).
).
Tests B. Les plus grands tests observables sont connus par des débris, dont aucun ne montre le stade initial. Une trčs médiocre section, décentrée
(Fig. 13.20 ![]() ), présentant une partie spiralée d'au moins 2 tours et plusieurs loges déroulées est supposée représenter la partie jeune de cette génération. Les plus grandes sections observées correspondent ŕ des tests flabellés mesurant jusqu'ŕ 9,5 mm, le rayon d'un test atteignant 7,5 mm ; rien n'indique cependant l'existence de loges annulaires. Admettant que les échecs ŕ trouver un test complet soient dus ŕ une grande fragilité de la limite entre les parties involute et évolute, une reconstruction d'ensemble a été tentée sur la
Figure 13.21
), présentant une partie spiralée d'au moins 2 tours et plusieurs loges déroulées est supposée représenter la partie jeune de cette génération. Les plus grandes sections observées correspondent ŕ des tests flabellés mesurant jusqu'ŕ 9,5 mm, le rayon d'un test atteignant 7,5 mm ; rien n'indique cependant l'existence de loges annulaires. Admettant que les échecs ŕ trouver un test complet soient dus ŕ une grande fragilité de la limite entre les parties involute et évolute, une reconstruction d'ensemble a été tentée sur la
Figure 13.21 ![]() .
.
Endosquelette. A la suite du flexostyle des tests A, 1 ou 2 loges trčs plates, étalées largement sur le proloculus, ne sont pas subdivisées
(Fig. 13.1-4, .9 ![]() ). L'endosquelette apparaît dčs le premier tour de spire et la loge superposée au flexostyle présente déjŕ parfois des logettes secondaires
(Fig. 13.4
). L'endosquelette apparaît dčs le premier tour de spire et la loge superposée au flexostyle présente déjŕ parfois des logettes secondaires
(Fig. 13.4 ![]() ) qui continuent d'exister, en petit nombre dans les 2 tours involutes mais deviennent exceptionnelles dans la partie déroulée aplatie des tests A, oů l'endosquelette central est réduit ŕ une simple lame sagittale
(Fig. 13.13-14
) qui continuent d'exister, en petit nombre dans les 2 tours involutes mais deviennent exceptionnelles dans la partie déroulée aplatie des tests A, oů l'endosquelette central est réduit ŕ une simple lame sagittale
(Fig. 13.13-14 ![]() ) ; les piliers préseptaux sont rarement observés de maničre certaine
(Fig. 13.10, .13
) ; les piliers préseptaux sont rarement observés de maničre certaine
(Fig. 13.10, .13 ![]() ). L'endosquelette des tests B est également réduit ŕ sa plus simple expression dans les loges les moins épaisses, proches de la partie initiale
(Fig. 13.17
). L'endosquelette des tests B est également réduit ŕ sa plus simple expression dans les loges les moins épaisses, proches de la partie initiale
(Fig. 13.17 ![]() ). Les logettes secondaires, apparemment désordonnées, réapparaissent dans les loges plus épaisses, vers la périphérie des tests ; elles peuvent devenir nombreuses et accompagnées de piliers préseptaux formant une ou plusieurs rangées
(Fig. 13.15-18
). Les logettes secondaires, apparemment désordonnées, réapparaissent dans les loges plus épaisses, vers la périphérie des tests ; elles peuvent devenir nombreuses et accompagnées de piliers préseptaux formant une ou plusieurs rangées
(Fig. 13.15-18 ![]() ). Rien ne semble indiquer une organisation confluente des logettes secondaires, mais il est vrai que de bonnes sections équatoriales dans des parties épaisses du test, oů les logettes secondaires trouvent la place pour s'organiser, nous manquent. L'une des particularités de l'espčce réside dans la profondeur importante de l'espace préseptal oů, comme chez les autres espčces du genre, les cloisonnettes se terminent en pointe triangulaire
(Fig. 13.14
). Rien ne semble indiquer une organisation confluente des logettes secondaires, mais il est vrai que de bonnes sections équatoriales dans des parties épaisses du test, oů les logettes secondaires trouvent la place pour s'organiser, nous manquent. L'une des particularités de l'espčce réside dans la profondeur importante de l'espace préseptal oů, comme chez les autres espčces du genre, les cloisonnettes se terminent en pointe triangulaire
(Fig. 13.14 ![]() ).
).
Au total, cette espčce dont un test A évoque encore Cuvillierinella
(Fig. 13.10 ![]() ), se rattache indubitablement au genre Cyclopseudedomia. L'interprétation de l'absence de structure confluente est liée aux états divers dans lesquels on trouve cette structure chez les autres espčces partageant les męmes caractéristiques : on la voit
apparaître et se perfectionner chez les autres espčces, passant de rudimentaire chez C. mavrikasi n. sp.
(Fig. 14.22
), se rattache indubitablement au genre Cyclopseudedomia. L'interprétation de l'absence de structure confluente est liée aux états divers dans lesquels on trouve cette structure chez les autres espčces partageant les męmes caractéristiques : on la voit
apparaître et se perfectionner chez les autres espčces, passant de rudimentaire chez C. mavrikasi n. sp.
(Fig. 14.22 ![]() ), ŕ fruste chez C. smouti
(Fig. 11.14
), ŕ fruste chez C. smouti
(Fig. 11.14 ![]() ), et plus aboutie chez C. klokovaensis (Fig.
12.14
), et plus aboutie chez C. klokovaensis (Fig.
12.14 ![]() ), ce qui permet d'admettre qu'elle existe ŕ l'état latent chez C. hellenica, comme elle existe exceptionnellement chez
Cuvillierinella. On notera d'ailleurs que le développement de ce caractčre est grosso-modo corrélé ŕ la taille du proloculus et celle des tests A et B (voir
Figs. 15
), ce qui permet d'admettre qu'elle existe ŕ l'état latent chez C. hellenica, comme elle existe exceptionnellement chez
Cuvillierinella. On notera d'ailleurs que le développement de ce caractčre est grosso-modo corrélé ŕ la taille du proloculus et celle des tests A et B (voir
Figs. 15 ![]() - 16
- 16 ![]() ).
).
|
Figure 13
: Cyclopseudedomia hellenica. 1-7 : Sections axiales centrées de tests A ; noter le
flexostyle en 4 et le cloisonnement toujours trčs précoce. 8 : Section décentrée de type Ty (voir
Fig. 2.6 |
Cyclopseudedomia mavrikasi n. sp.
(Fig. 14.1-28 ![]() ).
).
Cette nouvelle espčce s'impose par ses particularités et ses gisements éloignés les uns des autres, depuis l'Istrie jusqu'au sud de l'Italie. Ce sont probablement des représentants de cette espčce qui ont été attribués par Vicedo et al. (2009) ŕ Cyclopseudedomia hellenica, ŕ partir d'un matériel incomplet.
Diagnose. Espčce du genre Cyclopseudedomia, dimorphique, non costulée, partageant la tendance évolutive du type mais différente par plusieurs caractčres: proloculus plus petit, nombre de loges involutes plus grand, déroulement flabelliforme ou cylindrique pour les tests A; loges n'atteignant pas le stade annulaire chez les tests B. Structure confluente trčs fruste.
Origine du nom. L'espčce est dédiée ŕ Georges Mavrikas, dont le travail de thčse sur le massif du Gavrovo constitue un modčle d'utilisation de la micropaléontologie aux reconstructions paléogéographiques et structurales. L'auteur lui doit maintes découvertes dont ce travail est émaillé.
Matériel type. Le matériel type est constitué par un lot d'environ 30 sections comprises dans 10 lames minces portant le numéro YUC1. Les sections figurées de l'échantillon YUC1 et I285 sont déposées auprčs de l'USTL sous les
nº 3154 ŕ 58 et 3159 ŕ 61, respectivement. Des sections de cette espčce accompagnant Sigalveolina reicheli n. sp. dans l'échantillon YUA4 sont comprises dans le lot 3130 ŕ 53. L'holotype est une section équatoriale centrée
(Fig. 14.6 ![]() ) comprenant un proloculus de section ovale (0,18x0,24 mm) suivi d'un court flexostyle, d'une loge apparemment non subdivisée, puis de 2 tours de spire comprenant environ 18 loges formant une PRU flabelliforme, dont seules les 2 derničres sont détachées du tour précédent. Les loges du premier tour spiralé montrent les ouvertures primaires situées hors de l'axe des logettes correspondantes ; les derničres loges de la partie flabellée montrent des logettes secondaires obliques sur l'orientation des primaires et des piliers préseptaux. Cette section est comprise dans une lame portant le numéro d'échantillon (YUC1) un numéro de fabrication (556) et le numéro de collection (Coll.1075), déposée sous le nº 3158 auprčs de l'Université des Sciences et techniques de Lille.
) comprenant un proloculus de section ovale (0,18x0,24 mm) suivi d'un court flexostyle, d'une loge apparemment non subdivisée, puis de 2 tours de spire comprenant environ 18 loges formant une PRU flabelliforme, dont seules les 2 derničres sont détachées du tour précédent. Les loges du premier tour spiralé montrent les ouvertures primaires situées hors de l'axe des logettes correspondantes ; les derničres loges de la partie flabellée montrent des logettes secondaires obliques sur l'orientation des primaires et des piliers préseptaux. Cette section est comprise dans une lame portant le numéro d'échantillon (YUC1) un numéro de fabrication (556) et le numéro de collection (Coll.1075), déposée sous le nº 3158 auprčs de l'Université des Sciences et techniques de Lille.
Localisation, gisements. L'espčce est reconnue en 3 gisements
(Fig. 1 ![]() ). La population type de l'espčce (YUC1) provient d'Italie, région de Trieste, prčs d'Opicina, oů Bignot (1972, p. 68) situe son échantillon GB558. Il s'agit d'un calcaire packstone ŕ trčs abondants
"Bolivinopsis sp. et/ou Bigenerina sp." (in Bignot, 1972) et Cuneolina sp. ; on observe plusieurs sections par lame dans un lot de 10 lames minces. Une autre riche population (YUA4) provient de Slovénie, au bord de la route joignant Dutovlje ŕ Kreplje, oů Bignot (1972, p. 62) situe son échantillon GB994. Il s'agit d'un calcaire
wackestone oů abondent les Bolivinopsis et/ou Bigenerina ; de rares Milioles accompagnent les Sigalveolina reicheli n. sp. décrites précédemment ; chaque lame d'un lot de 76 unités renferme plusieurs sections des 2 espčces. Le troisičme gisement est situé en Italie méridionale, dans la péninsule Salentina, non loin de Bari, prčs de Gioia del Colle. Il s'agit d'un ensemble de dalles calcaires situé au bord de la route conduisant de Gioia ŕ Matera, ŕ environ 5km du passage de l'autoroute A14 ; l'échantillon I285 correspond ŕ un grainstone renfermant Cuneolina sp. et de rares Milioles. Les tests médiocrement conservés sont au nombre de plusieurs par lame mince dans un lot de 6. Ces divers échantillons témoignent de milieux marins subtidaux, apparemment non soumis ŕ l'émersion.
). La population type de l'espčce (YUC1) provient d'Italie, région de Trieste, prčs d'Opicina, oů Bignot (1972, p. 68) situe son échantillon GB558. Il s'agit d'un calcaire packstone ŕ trčs abondants
"Bolivinopsis sp. et/ou Bigenerina sp." (in Bignot, 1972) et Cuneolina sp. ; on observe plusieurs sections par lame dans un lot de 10 lames minces. Une autre riche population (YUA4) provient de Slovénie, au bord de la route joignant Dutovlje ŕ Kreplje, oů Bignot (1972, p. 62) situe son échantillon GB994. Il s'agit d'un calcaire
wackestone oů abondent les Bolivinopsis et/ou Bigenerina ; de rares Milioles accompagnent les Sigalveolina reicheli n. sp. décrites précédemment ; chaque lame d'un lot de 76 unités renferme plusieurs sections des 2 espčces. Le troisičme gisement est situé en Italie méridionale, dans la péninsule Salentina, non loin de Bari, prčs de Gioia del Colle. Il s'agit d'un ensemble de dalles calcaires situé au bord de la route conduisant de Gioia ŕ Matera, ŕ environ 5km du passage de l'autoroute A14 ; l'échantillon I285 correspond ŕ un grainstone renfermant Cuneolina sp. et de rares Milioles. Les tests médiocrement conservés sont au nombre de plusieurs par lame mince dans un lot de 6. Ces divers échantillons témoignent de milieux marins subtidaux, apparemment non soumis ŕ l'émersion.
Morphologie, enroulement.
Tests A. La moyenne des diamčtres de proloculus s'établit ŕ 0,21 mm pour des tests d'un diamčtre de 1,75 mm dans l'ensemble des 3 populations. L'holotype comprend la plupart des caractčres déductible de l'ensemble des sections équatoriales des 3 populations étudiées. Outre les PRU flabelliformes ordinaires, bien que rares dans la population type, les trois populations en renferment de cylindriques
(Fig. 14.4-5, .12-14, .17 ![]() ) ; l'existence de ces deux types est confirmée par les sections transverses de PRU
(Fig. 14.18-21
) ; l'existence de ces deux types est confirmée par les sections transverses de PRU
(Fig. 14.18-21 ![]() ).
).
Tests B. Ces tests sont bien représentés dans la population type, en sections équatoriales plus ou moins centrées. L'un d'eux montre le peloton initial caractéristique
(Fig. 14.22 ![]() ). Dans leur ensemble, ils comportent une partie juvénile constituée d'un petit nombre de tours, suivie de nombreuses loges trčs embrassantes, qui ne semblent pas atteindre le stade annulaire. Les tests apparaissent d'épaisseur trčs constante, sans épaississement ŕ la périphérie ; ils semblent entičrement évolute, y compris dans leur partie initiale, comme C. smouti. Il est vraisemblable que le test plat figuré par Bignot
(1971, Pl.
4, fig. 3), dont
"la partie enroulée est visible" (et ne peut donc pas ętre une Rhapydionina B, voir Fleury, 2014) soit attribuable ŕ cette espčce : l'échantillon dont il provient (GB1813) est en effet signalé par Bignot (1972, p. 216) comme caractérisé par la présence de
"Raadshoovenia cuvillieri", nom donné alors ŕ Sigalveolina reicheli n. sp.
). Dans leur ensemble, ils comportent une partie juvénile constituée d'un petit nombre de tours, suivie de nombreuses loges trčs embrassantes, qui ne semblent pas atteindre le stade annulaire. Les tests apparaissent d'épaisseur trčs constante, sans épaississement ŕ la périphérie ; ils semblent entičrement évolute, y compris dans leur partie initiale, comme C. smouti. Il est vraisemblable que le test plat figuré par Bignot
(1971, Pl.
4, fig. 3), dont
"la partie enroulée est visible" (et ne peut donc pas ętre une Rhapydionina B, voir Fleury, 2014) soit attribuable ŕ cette espčce : l'échantillon dont il provient (GB1813) est en effet signalé par Bignot (1972, p. 216) comme caractérisé par la présence de
"Raadshoovenia cuvillieri", nom donné alors ŕ Sigalveolina reicheli n. sp.
Endosquelette. Le plus grand nombre des tests comporte un endosquelette de type coaxial, constitué de logettes toutes parallčles entre elles. Les piliers préseptaux sont parfois difficiles ŕ distinguer dans les sections de PRU flabelliformes, oů on peut les confondre avec les cloisonnettes ; ils sont aisément lisibles dans les sections de PRU cylindriques
(Fig. 14.13-14, .17 ![]() ) ; ils apparaissent clairement dans les sections transverses de type
"Tx", en particulier Figure 14.26
) ; ils apparaissent clairement dans les sections transverses de type
"Tx", en particulier Figure 14.26 ![]() , comme dans les sections du męme type de C. smouti
(Fig. 11.12, .15
, comme dans les sections du męme type de C. smouti
(Fig. 11.12, .15 ![]() ) et C. klokovaensis (Fig.
12.16
) et C. klokovaensis (Fig.
12.16 ![]() ). Les logettes secondaires visibles dans les sections transverses de PRU, ordinairement d'un diamčtre inférieur ŕ celui des primaires, apparaissent disposées sans ordre au sein de l'endosquelette central. En outre, certaines sections de tests A
(Fig. 14.4, .6, .14
). Les logettes secondaires visibles dans les sections transverses de PRU, ordinairement d'un diamčtre inférieur ŕ celui des primaires, apparaissent disposées sans ordre au sein de l'endosquelette central. En outre, certaines sections de tests A
(Fig. 14.4, .6, .14 ![]() ) montrent des obliquités dans l'orientation des logettes secondaires, parfois concourantes, confirmées par des sections équatoriales de tests B
(Fig. 14.23
) montrent des obliquités dans l'orientation des logettes secondaires, parfois concourantes, confirmées par des sections équatoriales de tests B
(Fig. 14.23 ![]() ) : une structure confluente trčs imparfaite se trouve ici réalisée.
) : une structure confluente trčs imparfaite se trouve ici réalisée.
Au total, la nouvelle espčce partage les tendances évolutives communes ŕ celles du genre Cyclopseudedomia Elle est relativement proche de C. hellenica mais s'en distingue principalement par le déroulement plus important de ses tests des 2 générations et l'amorce de la structure confluente, moins bien réalisée que chez C. smouti et C. klokovaensis.
En conclusion, le genre Cyclopseudedomia est caractérisé par de trčs grands proloculus A et une extręme réduction de la partie involute au profit de la PRU, tendance évolutive fondamentale modulée par les deux générations de quatre espčces ; une structure confluente fruste s'y développe chez trois espčces et une fine costulation est reconnue chez deux d'entre elles. Ces espčces, associées ordinairement ŕ celles du genre Sigalveolina n. gen., caractérisent la zone CsB6b.
Il reste cependant ŕ examiner bričvement un organisme qui s'intčgre mal dans le cadre qui vient d'ętre mis en place.
|
Figure 14
: 1-28: Cyclopseudedomia mavrikasi
n.
sp. 1-21 :
Tests A. 1-4 : Sections axiales centrées ; noter le cloisonnement
précoce généralisé. 5 :
Section longitudinale d'une PRU ; noter la position des ouvertures
primaires situées en dehors de l'axe des logettes correspondantes (voir aussi
8-10 et 12). 6-17 : Sections équatoriales
centrées montrant la variété des modes de déroulement (PRU cylindrique ŕ
flabelliforme) ; 6 (holotype) et 14 comportent des indices de structure
confluente (logettes secondaires obliques sur l'axe des loges) ; 13 est
subcentrée. 18-21 : Sections
transverses de PRU circulaires ŕ rectangulaires ; noter les logettes
secondaires désordonnées. 22-28 :
Tests B. 22-23 : Sections équatoriales
de grands tests flabelliformes. 22 comporte un peloton caractéristique; 23
montre des logettes secondaires irréguličres, confluentes dans quelques cas,
constituant un dispositif comparable ŕ celui de C.
smouti. 24-25 : Croquis schématiques de 2 sections de grands tests
flabelliformes. 26 : Section
perpendiculaire au rayon d'un grand test montrant la terminaison triangulaire
des cloisonnettes dans l'espace préseptal et les piliers organisés en deux
rangées. 27-28: Portions de sections
perpendiculaires au plan d'étalement de grands tests flabelliformes (B
probables). 1-5, 8-10, 12-13, 15, 18-21, 23, 26-28: YUA4 ; 6, 16, 22,
24-25: YUC1 ; 7, 11, 14, 17: I285. 29-37 :
Cyclopseudedomia ? n. sp. 29-32:
Sections axiales centrées de tests A. Noter le flexostyle en 29 et le
cloisonnement précoce généralisé.
33-34 : Sections équatoriales de tests A ; les derničres loges
de la partie déroulée de 33 témoignent d'une structure confluente fruste. 35 :
Section transverse de PRU montrant de nombreux piliers préseptaux. 36-37 :
Sections trčs décentrées de grands tests plats, montrant les logettes
secondaires d'orientations variées et les nombreux piliers (voir aussi Fig.
3.7 |
Cyclopseudedomia ? n. sp.
Localisation, gisement. Cet organisme provient d'un échantillon (XGP288) récolté par G. Mavrikas aux environs de Miléa, dans la partie méridionale du massif du Gavrovo. Il s'agit d'un calcaire biomicritique wackestone ŕ débris de gastéropodes, "Discorbis" et Milioles, en l'absence de tout marqueur permettant de le dater.
Morphologie, enroulement. On observe deux types de tests, si différents l'un de l'autre que leur appartenance ŕ un męme taxon (espčce ou genre) est purement hypothétique.
Les petits tests (environ 1 mm), de génération A, sont de petite taille caractérisés par un proloculus de grande taille (0,20±0,05 mm pour 35 mesures), relativement aux espčces des genres Murciella et Sigalveolina n. gen. examinées précédemment, pour des diamčtres de tests non déroulés de 0,75±0,10 mm (50% des 34 mesures, les plus grandes) plus proches des valeurs notées chez les espčces de Cyclopseudedomia que de celles des 2 autres genres. La maille endosquelettique fine se pręte ŕ l'expression de la structure confluente dans la plupart des sections, en particulier dans les PRU, cylindriques dans leur ensemble
(Fig. 14.33 ![]() ).
).
Les grands tests (de génération B ?), indubitablement plats et évolutes sont connus par trois sections incomplčtes trčs décentrées de dimension proche de 5 mm. La structure confluente y est trčs clairement observable
(Figs. 3.7 ![]() ,
14.36-37
,
14.36-37 ![]() ). Il est possible que ces tests appartiennent ŕ la męme espčce que les tests A précédents, mais aucune certitude ne peut en ętre acquise, dans la mesure oů d'autres rares petits tests de type A, dotés d'un proloculus de taille comparable et d'un endosquelette plus massif
(Fig. 14.38
). Il est possible que ces tests appartiennent ŕ la męme espčce que les tests A précédents, mais aucune certitude ne peut en ętre acquise, dans la mesure oů d'autres rares petits tests de type A, dotés d'un proloculus de taille comparable et d'un endosquelette plus massif
(Fig. 14.38 ![]() ) participent ŕ l'association.
) participent ŕ l'association.
Au total, on trouve ici des caractčres męlant la tendance évolutive de Sigalveolina (développement d'une structure confluente bien exprimée) ŕ celle de Cyclopseudedomia (grande taille du proloculus et tendance marquée au déroulement évolute), qui souligne encore une fois l'indépendance des caractčres évolutifs. La singularité de l'association de caractčres habituellement disjoins ne permet pas de conclure mais ouvre d'évidentes perspectives ŕ des recherches futures.
On ne saurait terminer cet examen sans faire allusion ŕ Twaraina seiglici Robinson, 1993, que, comme le faisait remarquer son auteur, l'on aurait probablement classée parmi les espčces du genre Cyclopseudedomia si elle n'était d'âge éocčne moyen et connue de la région Caraďbe. On retient comme traits de ressemblance remarquable la grande taille du proloculus et la tendance au déroulement évolute des tests A, comme une probable structure confluente fruste (Robinson, 1993, Pl. 4.5), mais l'absence de tests B connus limite l'analyse d'une éventuelle parfaite convergence.
Un renouvellement significatif de la connaissance de la famille Rhapydioninidae résulte des trois parties de l'examen mené par l'auteur. Dans l'ensemble, la représentation schématique des relations entre les divers éléments qui la constituent, selon Fleury & Fourcade (1990, Figs. 2 et 4) n'est gučre modifiée, si ce n'est par l'adjonction de la sous-famille Cuvillierinellinae Fleury, 2016, détachée de celle des Rhapydionininae. On rappellera que l'image d'un tronc commun cylindrique dont émergent des rameaux correspondant aux diverses familles et sous-famille, si elle est originale dans sa forme, représente ce qui pouvait ętre déduit des observations de Reichel (1936, 1937, 1964) sur les Alveolinidae, confortées par celles de maints auteurs et en particulier chez les Rhapydioninidae. Chaque entité prend sa source chez quelque Miliolacea et suit sa propre voie indépendamment de toutes les autres, dans le carcan strict imposé par la norme familiale : l'organisation cellulaire fondamentale dirigée par la dualité logettes-espace préseptal n'entrave pas une liberté adaptative permettant toutes les fantaisies morphologiques, y compris une altération essentielle du fonctionnement cellulaire marquée par la structure confluente. Six sous-familles illustrent les diverses modalités de cette diversification.
8.1 : Sur la structure de la famille Rhapydioninidae : Sous-familles et genres.
Les sous-familles ont été distingués par Fleury & Fourcade (1990) ; seule la sous-famille Cuvillierinellinae (Fleury, 2016) a été créée depuis cette étude.
Les sous-familles américaines (Chubbininae et Neomurciellininae) sont peu concernées par ces études, bien qu'elles n'apparaissent pas fondamentalement différentes de celles du domaine euro-asiatique ; on rappellera que des vues simplifiées de ces deux entités sont illustrées par Fleury & Fourcade (1990, Figs. 8-9, dont la premičre est reprise par Vicedo, 2008, Fig. 89), puis remodelées par Fleury (2016, Fig. 4).
La sous-famille Sellialveolininae, restreinte ŕ la période médio-crétacée, apparaît indépendante des autres; elle est envisagée dans son ensemble par Fleury et Fourcade (1990, Fig. 5), et reprise par Vicedo (2008, Fig. 87).
La sous-famille Pseudedomiinae réunit des taxons encore mal connus, peuplant essentiellement des régions plus orientales que la Méditerranée, au Campanien-Maastrichtien (les mentions de périodes antérieures étant liées ŕ des erreurs d'interprétation longuement récidivées). Elle souffre d'un défaut originel, celui d'une espčce type (Pseudedomia multistriata Henson, 1948) représentée par une unique section décentrée et incomplčte d'un test B vraisemblable. Smout
(1963) a éclairé autant que possible cette section et proposé d'attribuer ŕ la męme espčce des petits tests lenticulaires de génération A ; bien qu'attrayante, cette hypothčse impliquant un dimorphisme de générations exceptionnel demande confirmation. Un ensemble de figurations est proposé par Fleury & Fourcade,
1990, Fig. 7
("Pseudedomia" globularis et "Chubbina ?" philippsoni ayant été depuis rapportées au genre Pseudochubbina) ; un état des connaissances a été établi par Mavrikas et al.,
1994, ŕ l'occasion de la découverte d'un représentant du genre en Grčce, comportant en particulier la structure confluente (voir
Fig. 3.9 ![]() ).
).
Les sous-familles Rhapydionininae et Cuvillierinellinae, alors confondues, ont fait l'objet d'une premičre synthčse, par Fleury & Fourcade (1990, Fig. 6), reprise par Vicedo (2008, Fig. 88). Les divers taxons de la premičre ont été étudiés par Fleury (2014), ceux de la seconde envisagés dans leur ensemble et illustrés par Fleury (2016, Fig. 13).
La sous-famille Rhapydionininae est caractérisée essentiellement par son endosquelette qui ménage une zone périphérique singuličrement large associée ŕ une trčs précoce tendance au déroulement, soit cylindrique (Rhapydionina, ŕ petit proloculus), soit flabelliforme (Fanrhapydionina, ŕ grand proloculus). La structure confluente y est inconnue. L'origine milioline de cet ensemble est indiquée par les espčces du groupe de R. dercourti Fleury, 2014, discrčtement streptospiralées dans les stades initiaux.
La sous-famille Cuvillierinellinae a fait l'objet principal des études récentes de l'auteur. Les genres Cuvillierinella, Murciella, Sigalveolina n. gen. et Cyclopseudedomia sont représentés par les sections les plus significatives de leurs espčces, dessinées sur la
Figure 16 ![]() .
.
Le genre Cuvillierinella est maintenant bien connu, par son espčce type qui, dans sa localité type a fourni ŕ De Castro (1988) un matériel trčs riche, confié ŕ Fleury (2016) ; on y observe que les morphotypes streptospiralés habituels coexistent avec des exemplaires pratiquement dépourvus d'endosquelette et d'autres encore, pratiquement planispiralés. L'ensemble témoigne en lui-męme des stades qui ont pu conduire d'une forme milioline d'origine ŕ des types planispiralés ŕ endosquelette typique de la famille, et dimorphisme de générations bien marqué. C'est clairement ŕ partir de ces variants et de quelques formes proches (C. fluctuans et C. perisalentina in Fleury, 2016) que se développent les divers rameaux représentés par les genres décrits ici.
Le genre Metacuvillierinella Fleury, 2016, est probablement de męme origine que Cuvillierinella, par son mode d'enroulement juvénile et par le détail de son organisation endosquelettique. Il emprunte cependant une voie évolutive originale, traduite par son enroulement adulte de type advolute et, paradoxalement, l'absence de toute tendance au déroulement final dans les deux générations, qui ne se distinguent que par leur stade juvénile.
Le genre Pseudochubbina De Castro, 1990, reste en partie énigmatique, bien qu'il soit représenté par quatre espčces euro-asiatiques; la principale difficulté de son interprétation résulte des doutes qui demeurent sur les caractéristiques des tests de génération B. Pour le reste, son enroulement streptospiralé persistant et ses logettes secondaires dispersées dans une masse compacte (SSC) semblent attester d'une proche parenté avec Cuvillierinella perisalentina Fleury, 2016. C'est en tout état de cause, ŕ l'heure actuelle, la seule hypothčse qui soit susceptible de rendre compte de l'origine de ce genre.
Les trois genres qui font l'objet de cette étude sont confrontés sur les diverses figures résumant les analyses précédentes. La
Figure 15 ![]() réunit les données morphologiques mesurables et comparables réunies précédemment et rappelle les associations auxquelles participent les diverses espčces. La
Figure 16
réunit les données morphologiques mesurables et comparables réunies précédemment et rappelle les associations auxquelles participent les diverses espčces. La
Figure 16 ![]() regroupe un choix de sections jugées caractéristiques permettant une comparaison visuelle immédiates entre les diverses espčces ; celles du genre Cuvillierinella y sont adjointes. La
Figure 17
regroupe un choix de sections jugées caractéristiques permettant une comparaison visuelle immédiates entre les diverses espčces ; celles du genre Cuvillierinella y sont adjointes. La
Figure 17 ![]() , plus schématique, permet une lecture immédiate des critčres qui sont retenus pour la distinction des divers genres et comporte l'indication de cas particuliers.
, plus schématique, permet une lecture immédiate des critčres qui sont retenus pour la distinction des divers genres et comporte l'indication de cas particuliers.
Le genre Murciella, tel qu'il est compris ici, se rattache clairement ŕ Cuvillierinella, comme indiqué par d'exceptionnels morphotypes se trouvant dans la population type de l'espčce type (De Castro,
1988 ; Fleury, 2016) et ne s'en distingue fondamentalement que par sa partie juvénile entičrement planispiralée. La
Figure 17 ![]() résume graphiquement les différences entre les deux genres ; la variante qui s'y trouve marquée correspond ŕ la population XGP110
(Fig. 6
résume graphiquement les différences entre les deux genres ; la variante qui s'y trouve marquée correspond ŕ la population XGP110
(Fig. 6 ![]() ) dont le bâti est particuličrement fin. Ces 2 genres partagent, au moins en partie, une męme répartition stratigraphique (zone CsB6a) et se trouvent dans tout le domaine méditerranéen.
) dont le bâti est particuličrement fin. Ces 2 genres partagent, au moins en partie, une męme répartition stratigraphique (zone CsB6a) et se trouvent dans tout le domaine méditerranéen.
Le nouveau genre Sigalveolina, correspondant aux espčces précédemment dénommées
"Murciella gr. renzi", se distingue principalement par sa structure confluente, associé ŕ un ensemble de caractčres schématisés sur la
Figure 17 ![]() ; les deux variantes notées sur cette figure sont constituées par le bâti relativement épais de S. methonensis (Fig.
8
; les deux variantes notées sur cette figure sont constituées par le bâti relativement épais de S. methonensis (Fig.
8 ![]() ) et les loges (tardivement) déroulées de S. reicheli (Fig.
10
) et les loges (tardivement) déroulées de S. reicheli (Fig.
10 ![]() ). Les diverses espčces, en compagnie de celles du genre suivant caractérisent la zone CsB6b.
). Les diverses espčces, en compagnie de celles du genre suivant caractérisent la zone CsB6b.
Le genre Cyclopseudedomia développe particuličrement une forte tendance au déroulement commun aux deux générations ; une structure confluente imparfaite s'y associe, sauf dans un cas particulier (C. hellenica, Fig.
13 ![]() ), indiqué sur la
Figure 17
), indiqué sur la
Figure 17 ![]() . On rappelle que męme en l'absence de sections parfaitement centrées, les tests B ont une épaisseur trčs faible en leur centre, alors que les Cuvillierinella parfois associées présentent en leur centre un test subglobuleux de męme morphologie que la génération A : ils ne devraient pas ętre confondus.
. On rappelle que męme en l'absence de sections parfaitement centrées, les tests B ont une épaisseur trčs faible en leur centre, alors que les Cuvillierinella parfois associées présentent en leur centre un test subglobuleux de męme morphologie que la génération A : ils ne devraient pas ętre confondus.
|
Figure 15 : Tableau récapitulatif des caractéristiques essentielles des taxons examinés (tests A). Chaque population est représentée par son numéro d'échantillon et par son numéro dans la collection de l'auteur (sauf pour EF436, déposé auprčs du Museum National d'Histoire Naturelle, Paris). Les moyennes et écarts-types sont accompagnés du nombre de sections observées. Le diamčtre équatorial des tests (De) correspond ŕ 50% des mesures (les plus grandes) afin de tenter d'éliminer les tests immatures ; on donne en outre le diamčtre de la partie enroulée involute (Dpi), permettant de visualiser directement l'importance du déroulement. L'indice d'aplatissement correspond au rapport du diamčtre équatorial ŕ l'épaisseur maximale du test mesurée parallčlement ŕ l'axe. Les données relatives au nombre de loges déroulées confirment les données précédentes et précisent le type de PRU (cylindrique ou flabelliforme). Les caractčres du bâti et de la structure confluence, peu aisées ŕ quantifier, font l'objet d'appréciations subjectives ; on signale les tests pourvus d'une costulation. |
|
Figure 16 : Architecture comparée de divers taxons de la sous famille Cuvillierinellinae. Chaque espčce ou population est représentée par les sections jugées les plus caractéristiques parmi celles figurées précédemment. Les holotypes sont représentés par leurs photos. Les sections de tests A apparaissent en blanc et la moyenne du diamčtre du proloculus (Dp., en centičmes de mm) est donnée ŕ la suite du nom de l'espčce ou de la population ; les sections correspondant éventuellement ŕ une deuxičme génération A sont faiblement grisées. Les tests B sont représentés en grisé sombre et comportent une étoile lorsque la section est centrée. |
|
Figure 17 : Diagrammes comparatifs des principales caractéristiques des genres étudiés (tests A). Les genres Cuvillierinella et Metacuvillierinella (représentés par des figures emboitées, le second en pointillés), sont résumés d'aprčs Fleury (2016). Les diagrammes des trois autres genres comportent des lignes pointillées rendant compte de données exceptionnelles, explicitées dans le texte. |
On a signalé précédemment l'association exclusive de cet organisme ŕ S. ovoidea, dans deux populations. L'aspect d'ensemble de ces tests est celui d'espčces crétacées rapportées dans le domaine euroasiatique au genre Pseudonummoloculina, telles que P. aurigerica Calvez, 1988 (espčce type du genre), de l'Albien de France méridionale, une forme indéterminée figurée par De Castro (1987, Fig. 3), du Cénomanien d'Italie, une autre espčce indéterminée illustrée par Hottinger et al. (1989, Pl. 22, figs. 1-5) et P. kalantarii Schlagintweit & Rashidi, 2016, du Maastrichtien d'Iran. Chez ces trois taxons, les auteurs décrivent comme principal trait l'existence d'une ouverture unique allongée bordée de crénulations ("fente en croissant, ŕ bord crénelé", "single aperture with weavy margins", "apertures with numerous short notches … protruding from the chamber roof and floor"). Ce n'est pas le cas de l'espčce mexicaine P. pecheuxi Fourcade & Fleury, 2001 (ŕ l'origine vraisemblable des diverses espčces du genre Praechubbina des męmes auteurs), oů l'on discerne plutôt 2 niveaux d'ouvertures, comme chez la forme grecque ici décrite. Par ailleurs, aucun des descripteurs des sections citées ne fait allusion ŕ un systčme de cloisonnement, dont l'existence semble cependant indiquée par la section de la Fig. 15b de Schlagintweit et Rashidi (au sud-sud-ouest du proloculus). C'est ce qui justifie la référence au taxon iranien qui a été choisie pour évoquer l'organisme grec.
On évoquera finalement un cas particulier, unique mais bien connu et illustré (Hottinger et al., 1989), celui de Hellenalveolina tappanae, d'un type pseudonummoloculinin affirmé par son enroulement streptospiralé, et une ouverture en fente crénelée, doté en outre d'un systčme de cloisonnement aussi bien développé que chez de typiques Alveolinidae, tels que Ovalveolina, par exemple. Il s'agirait d'un exemple remarquable d'aboutissement d'un des cas transitionnels qui viennent d'ętre évoqués.
Pour notre systčme de classification, dont la clé réside dans des définitions formelles et des critčres distinctifs parfois choisis a priori (souvent en raison des informations incomplčtes inhérentes ŕ notre discipline), il n'existe en principe aucune analogie entre l'organisme grec et l'iranien. On considčrera cependant que ce systčme fixiste est issu d'un temps oů les organismes étaient réputés indépendants les uns des autres, ce qui n'est plus admissible alors que l'accumulation des connaissances nous montre qu'il n'en est rien et que l'on peut souvent entrevoir le détail de leurs relations. C'est-ŕ-dire que, sauf pour les caractčres structuraux fondamentaux (ceux qui permettent la distinction entre Alveolinacea et Soritacea, par exemple), tous les détails d'organisation (tels que ceux de l'ouverture), doivent ętre considérés comme évolutifs et susceptibles de passer d'un type ŕ un autre en continuité sans qu'il soit nécessaire d'y voir plus qu'un ajustement progressif ŕ des contraintes diverses, internes ou environnementales. C'est la raison pour laquelle on ne juge pas ici nécessaire de fonder des distinctions d'ordre générique sur d'éventuelles différences dans la morphologie des ouvertures. De la męme maničre, l'acquisition d'un cloisonnement se situe dans la logique de ce que, ŕ la suite de Reichel, on s'attend ŕ trouver chez les entités susceptibles d'avoir conduit des Milolacea ŕ l'organisation propre aux Alveolinacea. Tel qu'il est ainsi compris, le genre Pseudonummoloculina témoignerait des modifications diverses, indépendantes les uns des autres, produites ŕ différentes époques, au sein d'un groupe de Miliolacea pour donner naissance aux divers rameaux qui, les uns aprčs les autres, depuis le milieu du Crétacé jusqu'ŕ l'Actuel, adoptent le mode d'organisation des Alveolinacea. Il n'y aurait, dans ces conditions, aucune raison de subdiviser ce genre, dont "l'air de famille" est bien marqué, qui représenterait simplement un état transitoire, itératif, entre deux modes d'organisation plus stables. Dans ces conditions, ce serait le statut spécifique qui serait le mieux adapté ŕ distinguer les différentes réalisations correspondant ŕ ce stade évolutif. L'organisme qui nous occupe ici serait l'un des témoins de ce type éphémčre, lui-męme apparemment sans avenir en raison de son âge, mais participant ŕ l'image plus ou moins fidčle de ce que furent ses prédécesseurs, plus heureux dans leur descendance. Ils auraient été trop fugaces pour que nous ayons pu accéder encore ŕ leur connaissance, mais nous pouvons au moins imaginer ŕ quoi ils pourraient ressembler.
8.3 Sur la répartition des Rhapydioninidae : L'influence des variations du milieu.
Les Rhapydioninidae, organismes qui peuplent presque seuls les régions les plus isolées des plates-formes carbonatées du domaine méditerranéen, vivent dans une tranche d'eau toujours peu épaisse et sont donc soumis ŕ toutes les vicissitudes de la vie de la plate-forme, aussi bien d'origine tectonique qu'eustatique.
Les premičres, tectoniques, ont été bien documentées par Mavrikas (1993) dans le massif du Gavrovo, vers la fin du Crétacé. Cet auteur distingue en particulier deux phases d'âge "santonien supérieur ŕ campanien basal" et "campanien inférieur sommital ŕ campanien supérieur" et une troisičme, plus hypothétique, "fini crétacée" ; ces phénomčnes ne sont pas précisément calibrés par rapport aux zones CsB et ne peuvent donc pas ętre pris en compte pour juger de leur influence sur les limites de ces zones. Ils soulignent cependant une grande instabilité, qui semble ętre assez générale dans l'ensemble des plates-formes adriatico-égéennes, d'aprčs l'examen des données bibliographiques.
Par ailleurs, des périodes d'émersion, interprétées comme d'origine eustatique apparaissent probables. L'une d'elles est documentée sur le terrain en Grčce (Landrein, 2001 ; Landrein et al., 2001), entre les zones CsB6b et CsB7. Une autre, non encore observée sur le terrain, semble probable entre les zones CsB6a et CsB6b, en raison du changement drastique qui s'opčre dans les peuplements de Rhapydioninidae. Il est possible męme que la zone CsB6a corresponde ŕ une lacune de sédimentation ou ŕ une ablation dans certaines régions : c'est ainsi que l'on peut tenter de rendre compte de l'absence des témoins de cette zone dans le massif de Pylos-Méthoni, oů pourtant les carbonates campano-maastrichtiens affleurent largement (Fleury et al., 1978). Une autre encore, envisagée par Heba et al. (2009) en Albanie, est possible entre les zones CsB5 et CsB6 ; celle-ci serait la cause de la disparition (au moins locale) des organismes présents sur la plate-forme depuis le Turonien-Coniacien, au profit des Rhapydioninidae. On notera encore que Steuber & Schlüter (2012) dénombrent au cours du Campanien plusieurs discontinuités dans la distribution stratigraphique des rudistes, qui seraient corrélées ŕ des périodes d'abaissement du niveau marin.
La Figure 18 ![]() donne un tableau d'ensemble de l'évolution régionale des Rhapydioninidae, tenant compte des éventualités précédemment exprimées et admettant que chaque zone soit strictement limitée par les conséquences de phénomčnes d'origine eustatique. Les données qui y sont consignées, réduites aux tests A, restent fragiles dans la mesure oů les genres réunissent en général les traits de plusieurs espčces distinguées par des nuances affectant ces caractčres ; la représentation est d'autant plus approximative que les genres (et a fortiori les espčces qu'ils englobent) n'ont pas nécessairement une durée de vie correspondant ŕ l'entičre période de temps de la zone dans laquelle ils sont reconnus. De telles incertitudes rendent aléatoire les possibilités d'utilisation de la zonation proposée ici dans des régions qui pourraient n'avoir pas connu les męmes vicissitudes avec la męme acuité. On notera comme un indice de cette difficulté les sections de
"Cuvillierinella ? sp." (ou éventuellement Metacuvillierinella ?) figurées par Schlagintweit
et Rashidi (2016), en provenance de couches bien postérieures ŕ l'apparition de Omphalocyclus macroporus et Loftusia sp. dans le Zagros Iranien. La
Figure 18
donne un tableau d'ensemble de l'évolution régionale des Rhapydioninidae, tenant compte des éventualités précédemment exprimées et admettant que chaque zone soit strictement limitée par les conséquences de phénomčnes d'origine eustatique. Les données qui y sont consignées, réduites aux tests A, restent fragiles dans la mesure oů les genres réunissent en général les traits de plusieurs espčces distinguées par des nuances affectant ces caractčres ; la représentation est d'autant plus approximative que les genres (et a fortiori les espčces qu'ils englobent) n'ont pas nécessairement une durée de vie correspondant ŕ l'entičre période de temps de la zone dans laquelle ils sont reconnus. De telles incertitudes rendent aléatoire les possibilités d'utilisation de la zonation proposée ici dans des régions qui pourraient n'avoir pas connu les męmes vicissitudes avec la męme acuité. On notera comme un indice de cette difficulté les sections de
"Cuvillierinella ? sp." (ou éventuellement Metacuvillierinella ?) figurées par Schlagintweit
et Rashidi (2016), en provenance de couches bien postérieures ŕ l'apparition de Omphalocyclus macroporus et Loftusia sp. dans le Zagros Iranien. La
Figure 18 ![]() exprime donc un état schématique des connaissances et des hypothčses, dont on peut attendre de substantielles améliorations, mais seulement au prix de précises études de terrain.
exprime donc un état schématique des connaissances et des hypothčses, dont on peut attendre de substantielles améliorations, mais seulement au prix de précises études de terrain.
|
Figure 18 : Principales caractéristiques (tests A) des genres européens de la famille des Rhapydioninidae dans leur cadre stratigraphique. Chaque cylindre représente un espace évolutif défini par les caractčres initiaux, l'importance relative des parties enroulées-déroulées et l'endosquelette. Les épisodes d'émersion sont commentés dans le texte. |
L'auteur doit beaucoup ŕ de nombreux collčgues et amis. D'une part ceux qui lui ont fait confiance et aidé ŕ obtenir les moyens destinés ŕ la poursuite d'un but lointain et apparemment utopique, qui consistait ŕ tenter la synthčse d'un groupe peu répandu, dispersé et d'accčs incommode ; il s'agit de Jacques Paquet, Jean Dercourt et Hervé Chamley, tour ŕ tour directeurs de l'UFR des Sciences de la Terre de l'Université de Lille 1 et/ou de diverses équipes associées au CNRS. D'autre part, ceux qui, sur le terrain, ou plus centrés sur la micropaléontologie ou la sédimentologie, tels Paul Bernier, Michel Bonneau, Irfan Cibaj, Piero De Castro, Patrick De Wever, Danielle Decrouez, Éric Fourcade, Claude Guernet, Grigor Heba, Alexandra et Spyros Lekkas, Jean-Paul Loreau, François Michaud, Madeleine Neumann, Jacques Sigal et François Thiébault, ont été d'un grand soutien matériel et/ou psychologique au cours des années. Marie-Béatrice Forel a eu l'amabilité et le courage de rechercher et trouver la piste du matériel type de Murciella cuvillieri mal aiguillé lors de son dépôt. En outre, les Rédacteurs en chef et Secrétaires de rédaction des revues ayant accepté d'accueillir des notes volumineuses peu rémunératrices en termes d'audience ont également puissamment participé ŕ la réalisation d'un projet porté pendant presque un demi-sičcle. Michel Septfontaine a usé avec modération de son pouvoir de rapporteur et a su exprimer clairement sa compréhension des idées et principes diffus dans ce texte. À tous et toutes : merci.
Bernier P. & Fleury J.-J. (1980).- La plate-forme carbonatée de Gavrovo-Tripolitza (Grčce). Évolution des conditions de sédimentation au cours du Mésozoďque.- Géologie méditerranéenne, Marseille, t. VII, nº 3, p. 247-259.
Bignot G. (1971).- Contribution ŕ l'étude des espčces liburniennes des genres Rhapydionina Stache, 1913 et Rhipidionina Stache, 1913.- Revue de Micropaléontologie, Paris, vol. 13, nº 4, p. 222-236.
Bignot G. (1972).- Recherches stratigraphiques sur les calcaires du Crétacé supérieur et de l'Éocčne d'Istrie et des régions voisines.- Thčse, Université Paris VI, Travaux du Laboratoire de Micropaléontologie, nº 2, 354 p.
Calvez H. (1988).- Pseudonummoloculina aurigerica n. gen., n. sp. et Dobrogelina ? angulata n. sp. Deux foraminifčres nouveaux de l'Albien calcaire des Pyrénées franco-espagnoles.- Revue de Paléobiologie, Genčve, vol. spécial 2 (Benthos '86), p. 391-399.
Caus E. & Cornella A. (1981).- Calveziconus lecalvezae n.gen. n.sp., Orbitolinidé campanien de la bordure méridionale des Pyrénées.- Cahiers de Micropaléontologie, Paris, vol. 4, p. 27-34.
Chiocchini M. & Mancinelli A. (2001).- Sivasella monolateralis Sirel & Gunduz, 1978 (Foraminiferida) in the Maastrichtian of Latium (Italy).- Revue de Micropaléontologie, Paris, vol. 44, nº 4, p. 267-277.
Christodoulou G. (1967).- Contribution to the geology of the Astypalaea island.- Annales de l'Université Aristote, Thessalonique, vol. 10, p. 171-180.
Colalongo M.L. (1963).- Sellialveolina viallii n. gen., n. sp. di Alveolinid cenomaniano dell'Appenninino meridional.- Giornale di Geologia (Serie 2a), Bologna, vol. XXX (1962), p. 361-371.
Cvetko B., Gušić I. & Schroeder R. (1997).- Reticulinella fleuryi (Foraminiferida) from the Upper Cretaceous (Upper Santonian - Middle Campanian) of the island of Brač, Croatia.- Revue de Micropaléontologie, Paris, vol. 40, p. 131-139.
Cvetko B., Gušić I., Jelaska V. & Buković D. (2001).- Stratigraphy and microfacies of the Upper Cretaceous Pučišća Formation, Island of Brač, Croatia.- Cretaceous Research, vol. 22, p. 591-613.
De Castro P. (1966).- Sulla presenza di un nuovo genere di Endothyridae nel Cretacico superiore della Campania.- Bollettino della Societŕ dei Naturalisti in Napoli, vol. 75, p. 1-33.
De Castro P. (1971).- Osservazioni su Raadshoovenia Van Den Bold e i suoi raporti col nuovo genere Scandonea (Foraminiferida, Miliolacea).- Bollettino della Societŕ dei Naturalisti in Napoli, vol. 80, p. 161-236.
De Castro P. (1987).- On some foraminifera and algae in Apennine Upper Cretaceous and Paleocene.- Memorie della Societŕ Geologica Italiana, Roma, vol. 40, p. 109-124
De Castro P. (1988).- Les Alvéolines du Crétacé d'Italie.- Revue de Paléobiologie, Genčve, vol. spécial 2 (Benthos '86), p. 401-416.
De Castro P. (1990).- Osservazioni paleontologiche sul cretacico della localitŕ-tipi di Raadshoovenia salentina e su Pseudochubbina n. gen.- Quaderni dell'Accademia pontaniana, Napoli, 10, p. 1-116.
De Castro P., Drobne K. & Gušić I. (1994).- Fleuryana adriatica n. gen., n. sp. (Foraminiferida) from the upermost Maastrichtian of the Brač Island (Croatia) and some other localities on the Adriatic Carbonate Platform.- Razprave IV. razreda SAZU, Ljubljana, vol. 35, nº 8, p. 129-149.
Decrouez D. & Radoičić R. (1977).- Nummoloculina irregularis n. sp., nouveau foraminifčre du Sénonien (Santonien) de la Serbie occidental (Dinarides yougoslaves).- Notes du Laboratoire de Paléontologie de l'Université de Genčve, nº 7, p. 1-5.
Fleury J.-J. (1970).- Le Sénonien et l'Éocčne ŕ microorganismes benthoniques du Klokova (Zone du Gavrovo, Akarnanie, Grčce continentale).- Revue de Micropaléontologie, Paris, vol. 13, nº 1, p. 30-44.
Fleury J.-J. (1974).- Contribution ŕ la connaissance des Rhapydionininae (Foraminifčres, Alveolinidae) crétacés.- Geobios, Villeurbanne, vol. 7, nº 4, p. 307-322.
Fleury J.-J. (1977).- Deux Rhapydionininae (Foraminifčres, Alveolinidae) d'affinités américaines dans le Crétacé supérieur de Grčce (Zone de Gavrovo-Tripolitza).- Revue de Micropaléontologie, Paris, vol. 20, nº 2, p. 77-90.
Fleury J.-J. (1979a).- Le genre Murciella (Foraminifčre, Rhapydionininae), dans le Crétacé supérieur de Grčce (zone de Gavrovo-Tripolitza).- Geobios, Villeurbanne, vol. 12, nº 2, p. 149-185.
Fleury J.-J. (1979b).- Ŕ propos d'une nouvelle espčce du Crétacé terminal de Grčce. Place du genre Cyclopseudedomia parmi les Rhapydionininae (Foraminifčres, Alveolinidae).- Revue de Micropaléontologie, Paris, vol. 22, nº 1, p. 19-28.
Fleury J.-J. (1980).- Les zones de Gavrovo-Tripolitza et du Pinde-Olonos (Grčce continentale et Péloponnčse du Nord). Evolution d'une plate-forme et d'un bassin dans leur cadre alpin.- Société géologique du Nord, Lille, Publication 4, 651 p.
Fleury J.-J. (1984).- Senalveolina aubouini, n. gen., n. sp., Alveolinidae nouveau du Sénonien de Grčce.- Revue de Micropaléontologie, Paris, vol. 27, nº 3, p. 171-188.
Fleury J.-J. (2014).- Données nouvelles sur Rhapydionina Stache, 1913 et Fanrhapydionina n. gen., un groupe de Rhapydioninidae (Alveolinacea, Foraminifera) foisonnant en région périadriatique au Campanien-Maastrichtien.- Geodiversitas, Paris, vol. 36, nº 2, p. 173-208.
Fleury J.-J. (2016).- Cuvillierinella salentina (Foraminifera, Rhapydioninidae) and its kinship in the Western Mediterranean during the Campanian-Maastrichtian.- Revue de Micropaléontologie, Paris, vol. 59, p. 200-224.
Fleury J.-J., Thiébault F. & Tsoflias P. (1978).- Stratigraphie et structure du Massif de Pylos (Zone de Gavrovo-Tripolitza, Péloponnčse sud-occidental, Grčce).- Annales de la Société géologique du Nord, Lille, vol. 98, p. 223-232.
Fleury J.-J. & Fourcade É. (1987).- Un nouvel Alveolinidae du Paléocčne-Éocčne inférieur du Mexique. Ses affinités avec les genres Murciella et Yaberinella.- Revue de Micropaléontologie, Paris, vol. 30, nº 2, p. 91-110.
Fleury J.-J. & Fourcade É. (1990).- La super-famille Alveolinacea (foraminifčres) : Systématique et essai d'interprétation phylogénétique.- Revue de Micropaléontologie, Paris, vol. 33, nº 3-4, p. 241-268.
Fourcade É. (1966).- Murciella cuvillieri n. gen., n. sp., nouveau foraminifčre du sud-est de l'Espagne.- Revue de Micropaléontologie, Paris, vol. 9, nº 3, p. 147-155.
Fourcade É. & Fleury J.-J. (2001).- Origine, évolution et systématique de Praechubbina n. gen., foraminifčre Alveolinacea de Crétacé supérieur du Guatemala et du Mexique.- Revue de Micropaléontologie, Paris, vol. 44, nº 2, p. 125-157.
Frija G., Parente M., Di Lucia M. & Mutti M. (2015).- Carbon and strontium isotope stratigraphy of the Upper Cretaceous (Cenomanian-Campanian) shallow-water carbonates of southern Italy: Chronostratigraphic calibration of larger foraminifera stratigraphy.- Cretaceous Research, vol. 53, p. 110-139.
Gušić I. & Jelaska V. (1990).- Upper Cretaceous stratigraphy of the Island of Brač.- Jugoslaneska akademija znanosti I umjetnosti, Institut za geološka Istraživanja, OOUR za Geologiju, Zagreb, 160 p.
Hamaoui M. & Fourcade E. (1973).- Révision des Rhapydionininae (Alveolinidae, Foraminifčres).- Bulletin du Centre de Recherche SNPA, Pau, vol. 7, nº 2, p. 361-435.
Heba G. (2008).- Évolution de la plate-forme carbonatée de Kruja en Albanie, du Crétacé ŕ l'Éocčne.- Thčse, Université du Québec, Montréal et Chicoutimi, 196 p.
Heba G. & Prichonnet G. (2009).- L'instabilité fini-Crétacé-Éocčne de la marge passive d'Apulie en Albanie (plate-forme carbonatée de Kruja) et ses impacts sur la sédimentation.- Bulletin de la Société géologique de France, Paris, vol. 180, nº 5, p. 431-448.
Heba G., Prichonnet G. & El Albani A. (2009).- Meteoric diagenesis of Upper Cretaceous and Paleocene-Eocene shallow-water carbonates in the Kruja platform (Albania): geochemical evidence.- Geologica carpathica, Bratislava, vol. 60, nº 2, p. 165-179.
Henson F.R.S. (1948).- Larger imperforate Foraminifera of south-western Asia.- British Museum of Natural History, London, 127 p.
Hottinger L. (1960).- Recherche sur les Alvéolines paléocčnes et éocčnes.- Mémoires de la Société paléontologique suisse, Basel, vol. 75-76, 232 p.
Hottinger L., Drobne K. & Caus E. (1989).- Late Cretaceous, large, complex miliolids (Foraminifera) endemic in the Pyrenean faunal province.- Facies, Erlangen, vol. 21, nº 1, p. 99-134.
Landrein P. (2001).- Fonctionnement sédimentaire et diagenčse d'une plate-forme carbonatée isolée du Crétacé supérieur: la zone de Gavrovo-Tripolitza (Grčce).- Thčse Doctorat, Université de Bourgogne, Dijon 154 p.
Landrein P., Loreau J.-P. & Fleury J.-J. (2001).- Émersion généralisée intra-maastrichtienne de la plate-forme de Gavrovo-Tripolitza (Grčce), effets sur les populations de foraminifčres Rhapydionininae.- Bulletin de la Société géologique de France, Paris, vol. 172, nº 1, p. 85-98.
Lee J.J. & Anderson O.R. (1991).- Symbiosis in foraminifera. In: Lee J.J. & Anderson O.R. (eds.), Biology of foraminifera.- Academic press, London, p. 157-220.
Le Goff J., Cerepi A., Ghysels G., Swennen R., Loisy C., Heba G., El Desouky H. & Muska K. (2015).- Meter-scale cycles as a proxy for the evolution of the Apulian Carbonate Platform during the late Cretaceous (Llogora Pass, Albania).- Facies, Erlangen, vol. 61, p. 21-47.
Luperto Sinni E. (1966).- Nuovo genere di foraminifero del Senoniano delle Murge.- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 4, nº 2, p. 263-268.
Luperto Sinni E. & Ricchetti G. (1978).- Studio micropaleontologico-stratigraphico di una successone carbonatica del Cretaceo superior rilevat nel sottosuolo delle Murge sud-orientali.- Rivista Italiana Paleontologica, Milano, vol. 84, nº 3, p. 561-666.
Mavrikas G. (1993).- Évolution crétacée-éocčne d'une plate-forme carbonatée des Héllénides externes. La plate-forme des Ori Valtou ("massif du Gavrovo"), zone de Gavrovo-Tripolitza (Grčce continentale).- Société géologique du Nord, Lille, Publication 20, 239 p.
Mavrikas G., Fleury J.-J. & Fourcade É. (1994).- Implications paléobiogéographiques de la présence de Pseudedomia (Foraminifčre) dans le Maastrichtien méditerranéen.- Comptes Rendus de l'Académie des Sciences (Série II), Paris, vol. 318, p. 849-855.
Papetti L. & Tedeschi D. (1965).- Nuovo genere di foraminifero del Santoniano superior.- Geologica Romana, vol. 4, p. 119-128.
Parente M. (1994).- A revised stratigraphy of the Upper Cretaceous to Oligocene units from southeastern Salento (Apulia, southern Italy).- Bollettino della Societŕ Paleontologica Italiana, Milano, vol. 33, nº 2, p. 155-170.
Pęcheux J.-F. (1984).- Le Sénonien supérieur-Tertiaire du Chiapas (SE Mexique) et ses macroforaminifčres.- Thčse 3e Cycle, Université de Nice, 154 p.
Pęcheux J.-F. (2002).- Des Miliolacea aux Alveolinacea (grands Foraminifčres): la faune du Crétacé supérieur et du Tertiaire inférieur du Chiapas (S.E. Mexique).- Geobios, Villeurbanne, vol. 35, p. 469-488.
Reichel M. (1936).- Étude sur les Alvéolines. Mémoires de la Société paléontologique suisse, Basel, vol. 57, nº 4, p. 1-93.
Reichel M. (1937).- Étude sur les Alvéolines. Mémoires de la Société paléontologique suisse, Basel, vol. 59, nº 3, p. 95-147.
Reichel M. (1953).- Les caractčres embryonnaires de Subalveolina.- Eclogae geologicae Helvetiae, Basel, vol. 40, nº 2, p. 256-262.
Reichel M. (1964).- Alveolinidae. In: Loeblich A.R. & Tappan H. (eds.), Sarcodina chiefly "Thecamoebians" and Foraminifera. In: Moore R.C. (ed.), Treatise in Invertebrate Paleontology, Part C, Protista 2.- The Geological Society of America, Tulsa, and The University of Kansas Press, Lawrence, p. C503--510.
Reina A. & Luperto-Sinni E. (1993).- Considerazioni chronostratigrafiche e paleogeografiche sui calcari a Rhapydionina liburnica affioranti nei dintorni di Veglie (Salento, Puglia).- Bollettino della Societa Geologica Italiana, Roma, vol. 112, p. 273-279.
Robinson E. (1968).- Chubbina, a new Cretaceous Alveolinid genus from Jamaica and Mexico.- Palaeontology, London, vol. 11, nº 4, p. 526-534.
Robinson E. (1993).- Some imperforate Larger Foraminifera from the Paleogene of Jamaica ant the Nicaragua Rise.- Journal of Foraminiferal Research, Lawrence, vol. 23, nº 1, p. 47-65.
Schlagintweit F. & Rashidi K. (2016).- Some new and poorly known benthic foraminifera from Late Maastrichtian shallow-water carbonates of the Zagros zone, SW Iran.- Acta palaeontologica romaniae, vol. 12, nº 1, p. 53-70.
Schlüter M., Steuber T. & Parente M. (2008).- Chronostratigraphy of Campanian-Maastrichtian platform carbonates and rudist associations of Salento (Apulia, Italy).- Cretaceous Research, vol. 29, p. 100-114.
Sigal J. (1952).- Ordre des foraminifčres. In : Piveteau J. (éd.), Traité de paléontologie. vol. 1.- Masson, Paris, p. 133-178 & 192-301.
Sigal J. (1966).- Le concept taxinomique de spectre. Exemples d'application chez les Foraminifčres. Propositions de rčgle de nomenclature.- Société géologique de France, Paris, Mémoire hors-série 3, 126 p.
Sigal J. (1989).- Les recherches sur les foraminifčres fossiles en France des environs de 1930 ŕ l'immédiat aprčs-guerre.- Société géologique du Nord, Lille, Publication 16, 161 p.
Smout A.H. (1963).- The genus Pseudedomia and its phyletic relationships, with remarks on Orbitolites and other complex foraminifera. In: Konigswald E.V. (ed.), Evolutionary trends in Foraminifera.- Elsevier, Amsterdam, p. 224-281.
Stache G. (1889).- Die Liburnische Stufe und deren Grenzhorizonte.- Abhandlungen des Geologische Reichsanstalt, Wien, Band 13, 170 p.
Steuber T. & Schlüter M. (2012).- Strontium isotope stratigraphy of Upper Cretaceous rudist bivalves : biozones, evolutionary patterns and sea-level change calibrated to numerical ages.- Earth Science Reviews, vol. 114, p. 42-60.
Torre M. (1967).- Alcuni foraminiferi de Cretacico superior della penisola Sorrentina.- Bollettino della Societŕ dei Naturalisti in Napoli, vol. 75, p. 409-429.
Van den Bold W.A. (1946).- Contribution to the study of Ostracoda with special reference to the Tertiary and Cretaceous microfauna of the Carribean region.- Thčse, Rijks-Univ. Utrecht, De Bussy, Amsterdam, 167 p.
Velić I. (2007).- Stratigraphy and paleobiogeography of Mesozoic benthic Foraminifera of the Karst Dinarides (SE Europe).- Geologica Croatica, Zagreb, vol. 60, nº 1, p. 1-113.
Vicedo V. (2008).- Morfoestructura de los generos cretacicos de los Rhapydioninidae (Foraminifera).- Tesis Doctoral, Universitat Autonoma de Barcelona, Bellaterra, 168 p.
Vicedo V., Aguilar-Lekkas M., Caus. E. & Hottinger L. (2009).- Fusiform and laterally compressed alveolinaceans (Foraminiferida) from both side of the Late Cretaceous Atlantic.- Neues Jarbuch für geologie und Paleontologie, Frankfurt am Main, vol. 253, nº 2-3, p. 229-247.
Vicedo V., Frijia G., Parente M. & Caus E. (2011).- The late Cretaceous genera Cuvillierinella, Cyclopseudedomia, and Rhapydionina (Rhapydioninidae, Foraminiferida) in shallow-water carbonates of Pylos (Peloponnese, Greece).- Journal of Foraminiferal Research, Lawrence, vol. 41, nº 2, p. 167-181.
Zambetakis-Lekkas A. (1988).- Biostratigraphie de la série crétacée de la zone de Tripolitza dans le massif du Mainalon (Péloponnčse central, Grčce).- Revue de Paléobiologie, Genčve, vol. spécial 2 (Benthos '86), p. 477-482.