◄ Carnets Geol. 19 (11) ►
![]()
Outline:
[Introduction]
[Geological background]
[Material and methods]
[Section of the Leetse Formation at the Zolotoy Creek]
[Systematic palaeontology]
[6. Conclusions] and ... [Bibliographic references]
St. Petersburg State University, Universitetskaya emb.7/9, 199034 St. Petersburg (Russia)
Borissiak Paleontological Institute of the Russian Academy of Sciences, Profsoyuznaya St. 123, 117997, Moscow (Russia)
Published online in final form (pdf) on August 8, 2019
DOI 10.4267/2042/70296
![]()
[Editor: Brian Pratt; technical editor: Bruno Granier]
![]()
New findings of esthonioporate bryozoans of the species Revalotrypa cf. inopinata, representing one of the oldest bryozoans of Baltoscandia, are described from an outcrop located in Leningrad Oblast' (north-western Russia). The colonies of this species are very small and were extracted from limestone-cemented nodules found in glauconitic sandstones of the lower part of the Joa Member (Paroistodus proteus conodont zone, lowermost Floian, Lower Ordovician). Combined X-ray microtomography and the examination of traditional thin sections under a stereomicroscope and a light microscope were used to study the morphology of this species.
Bryozoa;
Ordovician;
Floian;
Baltoscandia;
micro-CT
Fedorov P.V. & Koromyslova A.V. (2019).- New findings of the genus Revalotrypa, the oldest bryozoan genus of Baltoscandia, in north-western Russia.- Carnets Geol., Madrid, vol. 19, no. 11, p. 199-209.
Nouvelles découvertes du genre Revalotrypa, le plus vieux genre bryozoaire de Baltoscandia, au nord-ouest de la Russie.- Les nouvelles découvertes de bryozoaires esthonioporates de l'espèce Revalotrypa cf. inopinata, représentant l'un des plus anciens bryozoaires de Baltoscandie, sont décrites dans un affleurement situé dans l'oblast de Léningrad (nord-ouest de la Russie). Les colonies de cette espèce sont très petites et furent extraites de nodules calcaires cimentés trouvés dans les grès glauconitiques de la partie inférieure du Membre Joa (zone de сonodonte Paroistodus proteus, Floïen basal, Ordovicien inférieur). La microtomographie à rayons X combinée et l'examen de coupes minces traditionnelles au stéréomicroscope et au microscope optique sont utilisés pour étudier la morphologie de cette espèce.
Bryozoaires ;
Ordovicien ;
Floïen ;
Baltoscandie ;
microtomographie
There are not any unequivocal Cambrian bryozoans, including a 'cryptostomate
bryozoan' Pywackia baileyi Landing
in Landing et al., 2010,
described from the Upper Cambrian (Tinũ Formation) of Mexico (Landing
et al., 2010, 2015), which has been
reinterpreted as an octocoral with a 'pennatulacean-like morphology' (Taylor
et al., 2013). Tremadocian to early Floian (Early Ordovician) bryozoans are poorly represented in the
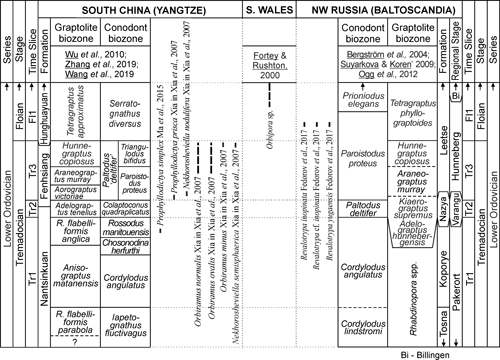
geological record (Fig. 1 ![]() ), while late Floian bryozoans are comparatively common,
especially in north-western Russia (Modzalevskaya, 1953; Pushkin & Popov,
1999; Gorjunova, 2009; Koromyslova,
2011). The oldest known unequivocal bryozoans in the
world are cryptostomate and esthonioporate bryozoans from the Tremadocian of
central China (Hu & Spjeldnæs,
1991; Xia et
al., 2007; Zhang et al., 2009; Ma et al.,
2014b, 2015).
The esthonioporate bryozoan species Revalotrypa
inopinata Fedorov et
al., 2017, and
R. yugaensis Fedorov et
al., 2017, from the lowermost Floian (Lower Ordovician) of the Leningrad
Oblast', Russian Federation, are the oldest known unequivocal bryozoans of
Baltoscandia, and most likely, the oldest of the entire Baltica paleocontinent.
The esthonioporate species Orbipora sp., described by Taylor & Cope
(1987)
from the Ogof Hen Formation (lower Floian) of South Wales, United Kingdom, is
younger than R. yugaensis and R. inopinata, since Fortey & Rushton
(2000)
correlated the Ogof Hen Formation
with the upper part of the T. phyllograptoides graptolite zone (Fig. 1
), while late Floian bryozoans are comparatively common,
especially in north-western Russia (Modzalevskaya, 1953; Pushkin & Popov,
1999; Gorjunova, 2009; Koromyslova,
2011). The oldest known unequivocal bryozoans in the
world are cryptostomate and esthonioporate bryozoans from the Tremadocian of
central China (Hu & Spjeldnæs,
1991; Xia et
al., 2007; Zhang et al., 2009; Ma et al.,
2014b, 2015).
The esthonioporate bryozoan species Revalotrypa
inopinata Fedorov et
al., 2017, and
R. yugaensis Fedorov et
al., 2017, from the lowermost Floian (Lower Ordovician) of the Leningrad
Oblast', Russian Federation, are the oldest known unequivocal bryozoans of
Baltoscandia, and most likely, the oldest of the entire Baltica paleocontinent.
The esthonioporate species Orbipora sp., described by Taylor & Cope
(1987)
from the Ogof Hen Formation (lower Floian) of South Wales, United Kingdom, is
younger than R. yugaensis and R. inopinata, since Fortey & Rushton
(2000)
correlated the Ogof Hen Formation
with the upper part of the T. phyllograptoides graptolite zone (Fig. 1 ![]() ). There are two further candidates for the oldest bryozoan species,
including the cystoporate bryozoan Ceramopora?
unapensis Ross, 1966, from the
uppermost Tremadocian or lowermost Floian deposits of Oklahoma, USA, and
esthonioporate bryozoan Dianulites
borealis Astrova, 1965, from Early Ordovician deposits of Arctic Russia (Astrova,
1965; Ernst et al., 2014).
However, the classification of Ceramopora?
unapensis as a bryozoan was
subsequently questioned by Taylor (1993), whereas the precise age of D.
borealis is uncertain. According to Ernst et
al. (2014), D. borealis is found
in uppermost Tremadocian deposits, whilst Nekhorosheva
(2015) suggested
that D. borealis is not older than the
Floian stage.
). There are two further candidates for the oldest bryozoan species,
including the cystoporate bryozoan Ceramopora?
unapensis Ross, 1966, from the
uppermost Tremadocian or lowermost Floian deposits of Oklahoma, USA, and
esthonioporate bryozoan Dianulites
borealis Astrova, 1965, from Early Ordovician deposits of Arctic Russia (Astrova,
1965; Ernst et al., 2014).
However, the classification of Ceramopora?
unapensis as a bryozoan was
subsequently questioned by Taylor (1993), whereas the precise age of D.
borealis is uncertain. According to Ernst et
al. (2014), D. borealis is found
in uppermost Tremadocian deposits, whilst Nekhorosheva
(2015) suggested
that D. borealis is not older than the
Floian stage.
In this article, findings of esthonioporate bryozoans of the genus Revalotrypa are described from a new location. Specimens of Revalotrypa cf. inopinata were extracted from a thin layer of dense sandstone nodules cemented by limestone, occurring within glauconitic sandstones of the lower part of the Leetse Formation. The nodule-bearing layer has been correlated with the base of the Floian Stage (Lower Ordovician).
The aims of this study are: (1) to describe the new outcrop yielding the oldest bryozoans of Baltoscandia, and (2) to describe the morphological characteristics of new specimens of the esthonioporate bryozoan genus Revalotrypa from Baltoscandia.
|
Figure 1:
Geographic and stratigraphic distribution of
Tremadocian to early Floian bryozoans. |
In north-western Russia, Ordovician sedimentary
rocks comprise an east-west trending elongated plateau, bounded in the north by
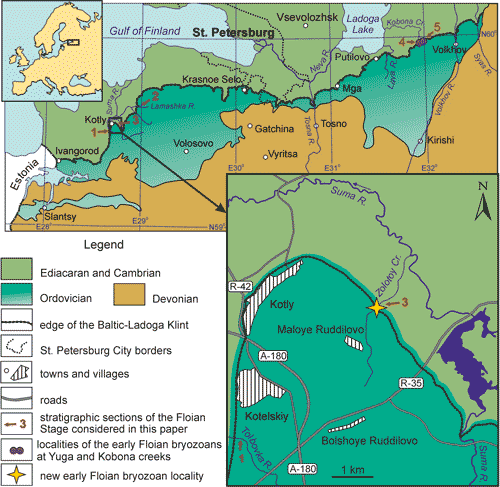
the Baltic-Ladoga Klint (Fig. 2 ![]() ).
Ordovician strata are composed mainly of limestone (Selivanova & Kofman,
1971) with the exception of the lower part, which is represented by
non-carbonate sequences. These include the lower Tremadocian Obolus Sandstone
and the Dictyonema Shale (the Tosna and Koporye formations,
respectively), glauconite-bearing quartz sandstone of the middle Tremadocian
Nazya Formation, and glauconitic sandstone of the upper Tremadocian to lower
Floian Leetse Formation (Popov et
al., 1989). Sedimentary disconformities are described from the base of both
the Nazya Formation and the Leetse Formation. The Leetse Formation is
conformably overlain by a medium-bedded glauconitic limestone of the Päite
Member, informally known as the 'Lower Dikary Member' (upper Floian). The Päite
Member forms part of the Volkhov Formation, which comprises the base of the
Ordovician carbonate sequence.
).
Ordovician strata are composed mainly of limestone (Selivanova & Kofman,
1971) with the exception of the lower part, which is represented by
non-carbonate sequences. These include the lower Tremadocian Obolus Sandstone
and the Dictyonema Shale (the Tosna and Koporye formations,
respectively), glauconite-bearing quartz sandstone of the middle Tremadocian
Nazya Formation, and glauconitic sandstone of the upper Tremadocian to lower
Floian Leetse Formation (Popov et
al., 1989). Sedimentary disconformities are described from the base of both
the Nazya Formation and the Leetse Formation. The Leetse Formation is
conformably overlain by a medium-bedded glauconitic limestone of the Päite
Member, informally known as the 'Lower Dikary Member' (upper Floian). The Päite
Member forms part of the Volkhov Formation, which comprises the base of the
Ordovician carbonate sequence.
Lower Ordovician rocks were deposited in the epeiric Baltoscandian paleobasin, which covered a significant portion of the East European Craton during the Ediacaran - Early Paleozoic (e.g., Nikishin et al., 1996; Sliaupa et al., 2006). Facies equivalents of the Leetse Formation include gray and reddish bioclastic limestones in central Sweden and graptolite shale in the Oslo region of Norway (Pärnaste & Bergström, 2013), however, bryozoans are absent from the coeval Scandinavian deposits (e.g., Lindskog et al., 2018). Furthermore, no bryozoans have been discovered in deposits of the Hunneberg and Billingen regional stages in Estonia and Belarus, which belong to the same facies zone as the Leningrad Oblast' (Männil, 1959; Pushkin, 1987).
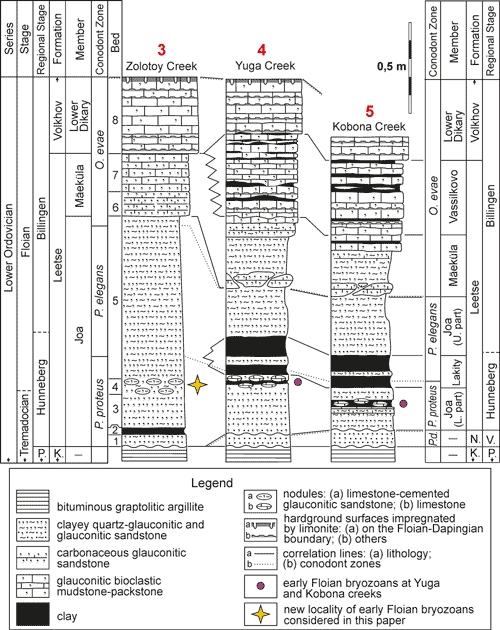
The Leetse Formation is divided into four
members in the Russian part of the Baltic-Ladoga Klint (anonymous, 1987). In
the west, it includes only 2 members: the Joa and the Mäeküla members (Fig. 3 ![]() ). The Joa Member consists of weakly-cemented quartz-glauconitic and
glauconitic sandstone, with layers of quartz or glauconitic-quartz sandstone at
the base, and locally, intercalated layers of clay. The Mäeküla Member
comprises more tightly cemented calcareous glauconitic sandstone and sandy
limestone. In the east, pale gray clay with interlayers of purplish-gray clay of
the Lakity Member wedge into the Joa Member, and the dark-green rocks of the Mäeküla
Member are replaced by variegated limestone with interlayers of clay, comprising
the Vassilkovo Member (see Fedorov et
al., 2017, for details).
). The Joa Member consists of weakly-cemented quartz-glauconitic and
glauconitic sandstone, with layers of quartz or glauconitic-quartz sandstone at
the base, and locally, intercalated layers of clay. The Mäeküla Member
comprises more tightly cemented calcareous glauconitic sandstone and sandy
limestone. In the east, pale gray clay with interlayers of purplish-gray clay of
the Lakity Member wedge into the Joa Member, and the dark-green rocks of the Mäeküla
Member are replaced by variegated limestone with interlayers of clay, comprising
the Vassilkovo Member (see Fedorov et
al., 2017, for details).
In general, the thickness of the Leetse Formation in the vicinity of Saint-Petersburg varies from about 0.5 to 2.7 m, progressively decreasing in a south to south-easterly direction (Fedorov & Ershova, 2012), whilst the age of the basal layers of the Leetse Formation also decreases in the same direction (Ershova, 2008).
The Leetse Formation comprises three North
Atlantic conodont zones (Fig. 3 ![]() ): Paroistodus
proteus, Prioniodus elegans, and Oepikodus
evae (Tolmacheva, 2001; Ershova,
2008). Löfgren (1993)
subdivided the P. proteus conodont
zone into 4 subzones, however, Tolmacheva (2001) and Tolmacheva et
al. (2001) recognized only the upper subzone (Oelandodus
elongates - Acodus deltatus
deltatus) at the base of the most complete sections of north-western Russia.
In the Diabasbrottet stratotype section, the base of this subzone is just below
the base of the Tetragraptus approximatus graptolite
zone, i.e., the base of the Floian stage (Bergström et al.,
2004).
): Paroistodus
proteus, Prioniodus elegans, and Oepikodus
evae (Tolmacheva, 2001; Ershova,
2008). Löfgren (1993)
subdivided the P. proteus conodont
zone into 4 subzones, however, Tolmacheva (2001) and Tolmacheva et
al. (2001) recognized only the upper subzone (Oelandodus
elongates - Acodus deltatus
deltatus) at the base of the most complete sections of north-western Russia.
In the Diabasbrottet stratotype section, the base of this subzone is just below
the base of the Tetragraptus approximatus graptolite
zone, i.e., the base of the Floian stage (Bergström et al.,
2004).
|
Figure 2:
Schematic geological map of Saint-Petersburg
and the Leningrad Oblast' (modified after Sokolova, 1971). Arrows 1-5
mark studied sections of the Leetse Formation; arrows 3, 4, 5 mark localities of the oldest
bryozoans of Baltoscandia. The new occurrence at the Zolotoy Creek is shown by
arrow 3 and marked with an asterisk on the insert map. |
|
Figure 3: Stratigraphic correlation of the Lower Ordovician sections yielding the
oldest bryozoans of Baltoscandia,
see text for details. Abbreviations: P., Pakerort; K., Koporye; N., Nazya; V.,
Varangu; P.d., Paltodus deltifer. |
Based on the findings of Ershova (2008),
we selected the thickest and most stratigraphically complete sections of the
Leetse Formation in the Saint-Petersburg area (Leningrad Oblast') to study for
bryozoan fauna in the lower part of the Joa Member. The thickest sections at the
Lava River and Putilovo Quarry were not considered because their fauna has
already been studied in detail (Tolmacheva et
al., 2001). Three localities were chosen and sampled during this study: the
Tolbovka River, Lamashka River, and Zolotoy Creek, which is the left-hand
tributary of the Suma River (Fig. 2 ![]() ).
Faunal remains comprising a carbonate skeleton were observed in deposits
correlated to the upper part of conodont zone P.
proteus (according to Ershova, 2008) in two of the three localities,
including on the left bank of the Tolbovka River, about 0.8 km south of
Kotelskiy village, and on the right bank of the Lamashka River. The coeval
nodule horizon was discovered in outcrops correlating to below the top of the P. proteus conodont zone on the right bank of the Zolotoy Creek,
0.9 km north-east of the Maloye Ruddilovo village, however fossilized
carbonate skeletal remains were not observed at outcrop (Fig. 3
).
Faunal remains comprising a carbonate skeleton were observed in deposits
correlated to the upper part of conodont zone P.
proteus (according to Ershova, 2008) in two of the three localities,
including on the left bank of the Tolbovka River, about 0.8 km south of
Kotelskiy village, and on the right bank of the Lamashka River. The coeval
nodule horizon was discovered in outcrops correlating to below the top of the P. proteus conodont zone on the right bank of the Zolotoy Creek,
0.9 km north-east of the Maloye Ruddilovo village, however fossilized
carbonate skeletal remains were not observed at outcrop (Fig. 3 ![]() ).
).
Thin intervals of glauconitic sandstone containing fossilized skeletal remains were sampled from the Tolbovka and Lamashka rivers. Approximately 5 kg of weakly-cemented, clayey, carbonate-bearing glauconitic sandstone was extracted from each outcrop. The samples were repeatedly washed with clean water to remove the mud component. After drying, the sandy residue was sieved into >1 mm and <1 mm size fractions. The >1 mm fraction was examined under an ordinary binocular microscope. As fossilized skeletal remains were not observed at outcrop, complete nodules were sampled from the Zolotoy Creek section, along with the host clayey glauconitic sandstone. The total weight of the sample exceeded 15 kg. These samples were prepared in the same way as detailed above, with the surface of the nodules and >1 mm fraction examined under a binocular microscope.
Only small valves of strongly dolomitized articulated brachiopods were found in samples from the Tolbovka River. The samples from Lamashka River contained only decalcified shells of articulated brachiopods, preserved as ferruginous internal/external molds in a weakly-consolidated clayey sand matrix. However, two very small bryozoans were obtained, together with numerous valves of small rhynchonelliform brachiopods and fragments of lingulids, from the host sandstone containing the nodular layer at Zolotoy Creek. A further attempt to obtain additional bryozoan samples from the nodules through repeated freezing and defrosting was unsuccessful.
Since the two bryozoan colonies were very small, we firstly investigated the material using non-destructive methods, i.e., scanning electron microscopy (SEM) and X-ray microtomography (micro-CT). The SEM analyses were performed at the Center for Microscopy and Microanalysis of Research Park of Saint-Petersburg State University, Russian Federation, where a scanning electronic microscope Tabletop TM 3000 (Hitachi) was used. The colonies were then scanned with a Skyscan 1172 (Bruker Corporation) at the Center for X-ray Diffraction Studies in the same institute. No filter was used and the voltage for analyses was 74 kV, while the current was 133 mA. Samples were rotated through 180° during examination and the rotation angle was 0.7°. The exposure was 1010 ms for each sample. The pixel resolution was slightly different for the two bryozoans. Sample 1 (PIN 5075/1013) was scanned with a resolution of 1.17 µm, while the pixel size was 1.17224 µm. Sample 2 (PIN 5075/1014) was scanned with a resolution of 1.24 µm, while the pixel size was 1.24132 µm. For each sample, 770 virtual sections were produced. Virtual three-dimensional (3D) models of the colony surfaces were created from the two-dimensional data. The raw micro-CT data were processed using the CTVox and DataViewer software.
The zooecia of the studied bryozoan specimens were fully filled by micrite, which resulted in a low contrast between the sediment and the calcite colony walls on the micro-CT sections. For this reason, traditional thin sections were made at the A.A. Borissiak Palaeontological Institute of the Russian Academy of Sciences (PIN), Moscow, Russian Federation, to study the internal structure of the specimens. The thin sections were studied and imaged using a Leica M165C stereomicroscope (SM) and a Zeiss Axioplan 2 transmitted light microscope (TLM).
Dimensions are given in the text as the observed range, followed in parentheses by the arithmetic mean ± standard deviation, number of colonies used (N), and total number of measurements made (n). All measurements were made using the micro-CT images.
The material forming the basis of this study is archived in the bryozoan collection of the Laboratory of Higher Invertebrates at the PIN, under number 5075.
The following beds are exposed in order of
decreasing stratigraphic age (i.e., from the base to the top of the succession; Fig. 3 ![]() ):
):
Bed 1 has a thickness of 0.07 m and comprises sandstone, which is dark-green, quartz-glauconitic, fine-grained, muddy, completely bioturbated, and weakly-cemented. Bed 1 overlies the top of the Dictyonema Shale and the contact is represented by a disconformity.
Bed 2 has a thickness of 0.03 m and comprises clay, which is whitish, sandy, glauconite-bearing, and contains rare fragments of graptolite rhabdosomes.
Bed 3 has a thickness of 0.18 m and comprises sandstone, which is dark-green, quartz-glauconitic, clayey, fine-grained, weakly-cemented, and completely bioturbated.
Bed 4 has a thickness of 0.08 m and comprises sandstone, which is dark-green, glauconitic, very fine-grained to fine-grained, clayey, weakly-cemented and completely bioturbated, with traces of deposit feeders infilled by a whitish clay. The sandstone includes disjointed nodules of dense sandstone cemented by limestone, ranging in size from a small coin to a fist. Two very small colonies of the bryozoan species Revalotrypa cf. inopinata Fedorov et. al., 2017 and numerous valves of small rhynchonelliform brachiopods and fragments of lingulids were found on the surface of the nodules and in the host sediments between them.
Bed 5 has a thickness of 0.89 m and comprises sandstone, which is dark-green, glauconitic, very fine-grained, clayey and muddy, weakly-cemented, and completely bioturbated, with numerous traces of deposit feeders infilled with whitish or violet-gray clay.
Bed 6 has a thickness of 0.14 m and comprises sandstone, which is dark-green, glauconitic, very fine-grained, densely cemented with limestone, and completely bioturbated.
Bed 7 has a thickness of 0.21 m and comprises sandy limestone, which is dark green, glauconitic, dolomitized, and completely bioturbated.
Bed 8 has a thickness of 0.42 m and comprises strongly dolomitized limestone, which is glauconitic and light-gray, with yellow bands of limonite impregnation along several hardground surfaces. The thickest band is located under the top of the bed, which is bored by numerous Gastrochaenolites ichno sp.
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Superorder Palaeostomata Ma et al., 2014a
Order Esthonioporata Astrova, 1978
Family Revalotrypidae Gorjunova, 1988
Genus Revalotrypa Bassler, 1952
Revalotrypa cf. inopinata Fedorov et al., 2017
Material: PIN 5075/1013
(Fig. 4 ![]() ), PIN 5075/1014
(Fig. 5
), PIN 5075/1014
(Fig. 5 ![]() ).
).
Occurrence: Joa Member of the Leetse Formation (upper layers of the Paroistodus proteus conodont zone, Hunneberg Regional Stage, lowermost Floian), right bank of the Zolotoy Creek near Maloye Ruddilovo, Leningrad Oblast', Russian Federation.
Description (measurements
based on specimen PIN 5075/1013): Colonies grain-shaped, longitudinally
elongated,
presumably formed by encrusting around a small object (Figs.
4.A, .D, .F ![]() ,
5.A-B, .G
,
5.A-B, .G ![]() ), and 2.5 mm long by 1.0 mm wide.
Secondary overgrowths absent. Autozooecia tubular. Autozooecial apertures
rounded to oval, 200-230 µm (215 ± 14 µm; N 1, n 6) long by 170-190 µm
(180 ± 9 µm; N 1, n 6) wide, with five to six apertures per 2 mm distance.
Autozooecial diaphragms not observed. Neozooecia abundant, separating
autozooecia in one or two rows, with up to ten neozooecia surrounding each
autozooecial aperture (Fig. 4.A-B, .E
), and 2.5 mm long by 1.0 mm wide.
Secondary overgrowths absent. Autozooecia tubular. Autozooecial apertures
rounded to oval, 200-230 µm (215 ± 14 µm; N 1, n 6) long by 170-190 µm
(180 ± 9 µm; N 1, n 6) wide, with five to six apertures per 2 mm distance.
Autozooecial diaphragms not observed. Neozooecia abundant, separating
autozooecia in one or two rows, with up to ten neozooecia surrounding each
autozooecial aperture (Fig. 4.A-B, .E ![]() ). Neozooecial apertures polygonal
and 80-110 µm (93 ± 14 µm; N 1, n 6) in diameter. One colony [PIN
5075/1013] exhibits an area with constriction, presumably consisting of small
autozooecia and neozooecia (Fig. 4.A
). Neozooecial apertures polygonal
and 80-110 µm (93 ± 14 µm; N 1, n 6) in diameter. One colony [PIN
5075/1013] exhibits an area with constriction, presumably consisting of small
autozooecia and neozooecia (Fig. 4.A ![]() ), 350 µm wide.
), 350 µm wide.
Remarks: Two very small poorly preserved colonies of bryozoans were
studied from
the early Floian of Leningrad Oblast', Russian Federation, which presumably belong to
the species Revalotrypa cf. inopinata based on a comparable colony form (Fedorov et
al., 2017: Fig. 5). However, the preservation state of the studied material
does not allow a definitive assignment. 3D models of the colony's surface of
sample PIN 5075/1013 have been produced (Fig. 4.A-B ![]() ), which facilitate a
description of the external morphology of Revalotrypa cf. inopinata without using SEM images,
which are of poor quality (Fig. 4.D-E
), which facilitate a
description of the external morphology of Revalotrypa cf. inopinata without using SEM images,
which are of poor quality (Fig. 4.D-E ![]() ).
However, the surface of the other specimen [PIN 5075/1014] is covered by
glauconite grains (Fig. 5.A-B
).
However, the surface of the other specimen [PIN 5075/1014] is covered by
glauconite grains (Fig. 5.A-B ![]() ), precluding our ability to
study its external morphology. The internal structures of these bryozoans are
unclear, because their zooecia were filled by micrite and diagenetic
recrystallization to microspar has affected their calcite walls. As a result,
both the micro-CT sections (Figs. 4.C
), precluding our ability to
study its external morphology. The internal structures of these bryozoans are
unclear, because their zooecia were filled by micrite and diagenetic
recrystallization to microspar has affected their calcite walls. As a result,
both the micro-CT sections (Figs. 4.C ![]() ,
5.C-D
,
5.C-D ![]() ) and
the thin sections (Figs. 4.F-G
) and
the thin sections (Figs. 4.F-G ![]() ,
5.E-G
,
5.E-G ![]() ) have a low contrast
between the sediment and the calcite walls.
) have a low contrast
between the sediment and the calcite walls.
|
Figure 4:
Revalotrypa cf. inopinata
Fedorov et al., 2017, PIN 5075/1013, early Floian. A-C. Micro-CT
images. A. Surface model, front view of grain-shaped
colony showing apertures of autozooecia (Az) and neozooecia (Nz), and area with constriction (C). B. Surface model, top view. C. Model of longitudinal section showing
several zooecial apertures (Z). D-E. SEM
images. D. General view of colony. E. Surface of colony. F-G. SM images. F. Longitudinal thin section through the central part of the colony
showing almost radially located zooecia. G. Longitudinal thin section, zooecia (outline
marked) filled by microspar. |
|
Figure 5: Revalotrypa cf. inopinata
Fedorov et al., PIN 5075/1014, early Floian. A. SEM image of colony.
B-D. Micro-CT
images. B. Surface model of grain-shaped colony, front view. C. Model of
longitudinal section showing a few zooecial tubes (arrowed). D. Model of
tangential section, top view, showing two autozooecial apertures (Az). E-G. Longitudinal thin sections at different levels near the central part of
the colony showing almost radially located zooecia
(Z). E. TLM image. F. SM image. G. SM image showing the outline of the zooecia. |
Bryozoans Revalotrypa cf. inopinata Fedorov et al., 2017, are reported from a new location in the Leningrad Oblast', increasing the number of outcrops containing the oldest bryozoans in Baltoscandia to three. The oldest bryozoan fauna can be found here only in the upper part of the P. proteus conodont zone, near the base of the most complete sections of glauconitic sandstones, which contain nodular layers enriched with pure limestone that has not been subjected to secondary dolomitization.
Combined X-ray microtomography and the study of traditional thin sections under a stereomicroscope and a light microscope represent the most promising approaches for studying the morphology of these very small bryozoans. 3D modeling of a colony usually replaces 'traditional' thin-section study, because micro-CT allows sections through the whole colony providing information on the internal morphology, but in cases where zooecia are partly filled by microspar, resulting in a low contrast between the sediment and calcite colony walls in the micro-CT image, thin sections can be used.
Steven J. Hageman (Dept. of Geology Appalachian State University, Boone) is thanked for helpful reviews and comments on the manuscript. Financial support of the Russian Foundation for Basic Research (Research Project No. 18-05-00245-A, 18-04-01046-A) is gratefully acknowledged.
anonymous (1987).- Resolutions of the Interdepartmental Conference on the Stratigraphy of the Ordovician and Silurian of the East European Platform, 1987, including Regional Stratigraphic Schemes.- Vsesoyuznyi Nauchno-Issledovatelskiy Geologicheskiy Institut, Leningrad, 114 p. + 4 Regional Stratigraphic Schemes. (in Russian).
Astrova G.G. (1965).- Morphology, history of development, and systematics of Ordovician and Silurian Bryozoa.- Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, Moscow, vol. 106, 432 p. (in Russian).
Astrova G.G. (1978).- Historical development, systematics and phylogeny of Bryozoa. Order Trepostomata.- Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, Moscow, vol. 169, 240 p. (in Russian).
Bassler R.S. (1952).- Taxonomic notes on genera of fossil and recent Bryozoa.- Journal of the Washington Academy of Sciences, Baltimore, vol. 42, p. 381-385.
Borg F. (1926).- Studies on Recent cyclostomatous Bryozoa.- Zoologiska Bidrag från Uppsala, vol. 10, p. 181-507.
Bergström S.M., Löfgren A. & Maletz J. (2004).- The GSSP of the Second (Upper) Stage of the Lower Ordovician Series: Diabasbrottet at Hunneberg, Province of Västergötland, Southwestern Sweden.- Episodes, Seoul, vol. 27, p. 265-272.
Ehrenberg C.G. (1831).- Symbolæ physicæ, seu icones et descriptiones animalium evertebratorum, sepositis insectis, quae ex itineribus per Libyam, Ægyptum, Nubiam, Dongalam, Syriam, Arabiam et Habessiniam publico institutis sumptu Friderici Guilelmi Hemprich et Christiano Godofredi Ehrenberg medicinae et chirurgiae doctorum, studio annis MDCCCXX-MDCCCXXV redierunt.- G. Reimer, Berlin, 128 p.
Ernst A., Bogolepova O.K., Hubmann B., Golubkova E.Y., & Gubanov A.P. (2014).- Dianulites (Trepostomata, Bryozoa) from the Early Ordovician of Severnaya Zemlya, Arctic Russia.- Geological Magazine, Cambridge, vol. 151, no. 2, p. 328-338.
Ershova V.B. (2008, unpublished).- Condensed deposits of the Varangu and Latorp Regional Stages (Lower Ordovician) of the Russian part of the Baltic-Ladoga Klint.- PhD thesis, St. Petersburg State University, 203 p. (in Russian).
Fedorov P.V. & Ershova V.B. (2012).- Significance of detailed lithostratigraphical studies for reconstruction of history and paleogeography of ancient epeiric seas. In: Sitnikov T.A., Platonov M.V., Tugarova M.A., Brusnizyn A.I., Rybalko A.E. & Lalomov A.V. (eds.), Leningrad school of lithology. Materials of All-Russian meeting of lithology dedicated to the 100th anniversary of L.B. Rukhin (25-29 September 2012, St. Petersburg).- St. Petersburg State University, p. 258-260. (in Russian).
Fedorov P.V., Koromyslova A.V. & Martha S.O. (2017).- The oldest bryozoans of Baltoscandia from the lowermost Floian (Ordovician) of north-western Russia: Two new rare, small and simple species of Revalotrypidae.- PalZ, Frankfurt am Main, vol. 91, no. 3, p. 353-373.
Fortey R.A. & Rushton A.W.A. (2000).- South Wales. In: Gregory F.J. (ed.), A revised correlation of Ordovician rocks in the British Isles.- Dorset Press, Dorchester, p. 13-17.
Gorjunova R.V. (1988).- On the systematic position of the genus Revalotrypa (Bryozoa).- Paleontologicheskiy zhurnal, Moscow, vol. 2, p. 31-36 (in Russian).
Gorjunova R.V. (2009).- Evolution of the colonial growth habit in the Ordovician bryozoans of the class Stenolaemata: Feeding adaptations (Leningrad Region, Russia).- Paleontological Journal, Moscow, vol. 43, p. 1390-1405.
Hu Z.-X. & Spjeldæs N. (1991).- Early Ordovician bryozoans from China. In: Bigey F. (ed.), Bryozoaires actuels et fossiles: Bryozoa living and fossil. Papers presented at the 8th International Conference on Bryozoa, Paris, 1989.- Société des Sciences Naturelles de l'Ouest de la France, Nantes, p. 179-185.
Koromyslova A.V. (2011).- Bryozoans of the Latorp and Volkhov horizons (Lower-Middle Ordovician) of the Leningrad Region.- Paleontological Journal, Moscow, vol. 45, p. 887-980.
Landing E., Antcliffe J.B., Brasier M.D. & English A.B. (2015).- Distinguishing Earth's oldest known bryozoan (Pywackia, Late Cambrian) from pennatulacean octocorals (Mesozoic-Recent).- Journal of Paleontology, Cambridge, vol. 89, p. 292-317.
Landing E., English A. & Keppie J.D. (2010).- Cambrian origin of all skeletalized metazoan phyla-Discovery of Earth's oldest bryozoans (Upper Cambrian, southern Mexico).- Geology, vol. 38, p. 547-550.
Lindskog A., Lindskog A.M.L., Johansson J.V., Ahlberg P. & Eriksson M.E. (2018).- The Cambrian-Ordovician succession at Lanna, Sweden: Stratigraphy and depositional environments.- Estonian Journal of Earth Sciences, Tallinn, vol. 67, p. 133-148.
Löfgren A. (1993).- Conodonts from the Lower Ordovician at Hunneberg, south-central Sweden.- Geological Magazine, Cambridge, vol. 120, p. 215-232.
Ma J.Y., Buttler C.J. & Taylor P.D. (2014a).- Cladistic analysis of the "trepostome" suborder Esthonioporina and the systematics of Palaeozoic bryozoans. In: Rosso A., Wyse Jackson P.N. & Porter J. (eds.), Bryozoan Studies 2013: Proceedings of the 16th International Bryozoology Association Conference, Catania, Sicily.- Studi Trentini Di Scienze Naturali, Trento, vol. 94, p. 153-161.
Ma J.-Y., Taylor P.D. & Xia F.-S. (2014b).- New observations on the skeletons of the earliest bryozoans from the Fenhsiang Formation (Tremadocian, Lower Ordovician), Yichang China.- Palaeoworld, Amsterdam, vol. 23, p. 25-30.
Ma J.-Y., Taylor P.D., Xia F.-S. & Zhan R.-B. (2015).- The oldest known bryozoan: Prophyllodictya (Cryptostomata) from the lower Tremadocian (Lower Ordovician) of Liujiachang, southwestern Hubei, central China.- Palaeontology, Oxford, vol. 58, p. 925-934.
Männil R.M. (1959, unpublished).- Questions of stratigraphy and bryozoans of the Ordovician of Estonia.- PhD thesis, Akademiya nauk Estonskoj SSR, Tallinn, 852 p. (in Russian).
Modzalevskaya E.A. (1953).- Ordovician trepostomes of the Baltic Sea Region and their stratigraphic significance.- Trudy Vsesojuznogo neftyanogo nauchno-issledovatelskogo geologorazvedochnogo instituta Novaja serija, Moscow, vol. 78, p. 91-196 (in Russian).
Nekhorosheva L.V. (2015).- To the question of the age of the Kruzhilikhа Formation of the Ordovician Severnaya Zemlya suite in connection with the findings in it of bryozoans Dianulites borealis Astrova (Order Trepostomata).- Regional'naya geologiya i metallogeniya, St. Petersburg, vol. 62, p. 45-48 (in Russian).
Nikishin A.M., Ziegler P.A., Stephenson R.A., Cloetingh, S.A.P.L., Furne A.V., Fokin P.A., Ershov A.V., Bolotov S.N., Korotaev M.V., Alekseev A.S., Gorbachev V.I., Shipilov E.V., Lankreijer A., Bembinova E.Yu. & Shalimov I.V. (1996).- Late Precambrian to Triassic history of the East European Craton: Dynamics of sedimentary basin evolution.- Tectonophysics, Amsterdam, vol. 268, p. 23-63.
Ogg J.G., Ogg G.M., & Gradstein F.M. (2016).- A Concise Geologic Time Scale.- Elsevier, Amsterdam, 234 p.
Pärnaste H. & Bergström J. (2013).- The asaphid trilobite fauna: Its rise and fall in Baltica.- Palæogeography, Palæoclimatology, Palæoecology, Amsterdam, vol. 389, p. 64-77.
Popov L.E., Kazanovitch K.K., Borovko N.G., Sergeeva S.P. & Sobolevskaya R.F. (1989).- The key sections and stratigraphy of the Cambrian-Ordovician phosphate bearing Obolus beds on the northeastern Russian platform.- Trudy mezhvedomstvennogo stratigraphicheskogo komiteta SSSR, St. Petersburg, vol. 18, 222 p. (in Russian).
Pushkin V.I. (1987, unpublished).- Ordovician bryozoans from the East European Platform (composition, distribution, community).- PhD thesis, Palaeontological Institute of the Academy of Sciences of the USSR, Moscow, 423 p. (in Russian).
Pushkin V.I. & Popov L.E. (1999).- Early Ordovician bryozoans from north-western Russia.- Palaeontology, Oxford, vol. 42, p. 171-189.
Ross J.P. (1966).- Early Ordovician ectoproct from Oklahoma.- Oklahoma Geological Notes, Norman, vol. 26, no. 8, p. 218-224.
Selivanova V.A. & Kofman V.S. (1971).- Geology of USSR. Vol. I. Leningrad, Pskov, and Novgorod regions: Geological descriptions.- Nedra, Moscow, 502 p. (in Russian).
Sliaupa S., Fokin P., Lazauskiene J. & Stephenson R.A. (2006).- The Vendian - Early Palaeozoic sedimentary basins of the East European Craton.- Geological Society of London, Memoirs, vol. 32, p. 449-462.
Sokolova T.N. (1971).- Geological map of the Pre-Quaternary deposits of Leningrad, Pskov, and Novgorod regions. Scale: 1:1000000. In: Selivanova V.A. (ed.), Geology of USSR. I. Leningrad, Pskov, and Novgorod regions: Appendix 1.- Nedra, Moscow (in Russian).
Suyarkova A.A. & Koren' T.N. (2009).- Advances in the Ordovician graptolite biostratigraphy of the St. Petersburg area Russia.- Estonian Journal of Earth Sciences, Tallinn, vol. 58, no. 1, p. 3-9.- DOI:10.3176/earth.2009.1.01.
Taylor P.D. (1993).- Bryozoa. In: Benton M.J. (ed.), The fossil record 2.- Chapman & Hall, London, p. 465-489.
Taylor P.D. & Cope J.C.W. (1987).- A trepostome bryozoan from the lower Arenig of south Wales: Implications of the oldest described bryozoan.- Geological Magazine, Cambridge, vol. 124, no. 4, p. 367-371.
Taylor P.D., Berning B. & Wilson M.A. (2013).- Reinterpretation of the Cambrian 'bryozoan' Pywackia as an octocoral.- Journal of Paleontology, Cambridge, vol. 87, no. 6, p. 984-990.
Tolmacheva T.Y. (2001, unpublished).- Conodont biostratigraphy and diversity in the Lower-Middle Ordovician of Eastern Baltoscandia (St. Petersburg region, Russia) and Kazakhstan.- Comprehensive summary of doctoral dissertation, Uppsala University, 40 p.
Tolmacheva T.Y., Koren T.N., Holmer L.E., Popov L.E. & Raevskaya E. (2001).- The Hunneberg Stage (Ordovician) in the area east of St. Petersburg, north-western Russia.- Paläontologische Zeitschrift, Frankfurt am Main, vol. 74, p. 543-561.
Wang Z.- H., Zhen Y.Y., Bergström S.M., Wu R.- C., Zhang Y.- D. & Ma X. (2019).- A new conodont biozone classification of the Ordovician System in South China.- Palaeoworld, Amsterdam, vol. 28, no. 12, p. 173-186.
Wu R., Stouge S., Li Z. & Wang Z. (2010).- Lower and Middle Ordovician conodont diversity of the Yichang Region, Hubei Province, Central China.- Bulletin of Geosciences, Prague, vol. 85, p. 631-644.
Xia F.-S., Zhang S.-G. & Wang Z.-Z. (2007).- The oldest bryozoans: New evidence from the Late Tremadocian (Early Ordovician) of East Yangtze Gorges in China.- Journal of Paleontology, Cambridge, vol. 81, p. 1308-1326.
Zhang S.-G., Xia F.-S., Yan H.-J. & Wang Z.-Z. (2009).- Horizon of the oldest known bryozoans (Ordovician).- Palaeoworld, Amsterdam, vol. 18, p. 67-73.
Zhang Y., Zhan R., Zhen Y., Wang Z., Wenwei Y., Fang X., Ma X. & Zhang J. (2019).- Ordovician integrative stratigraphy and timescale of China.- Science China Earth Sciences, Berlin, vol. 62, p. 61-88.