◄ Carnets Geol. 19 (12) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting, material and methods]
[3. Systematic palaeontology]
[4. Concluding remarks]
and ... [Bibliographic references]
Department of Palaeontology and Stratigraphy, Faculty of Earth Science,
University of Silesia in Katowice, Będzińska Str. 60, PL-41-200 Sosnowiec
(Poland)
Published online in final form (pdf) on October 10, 2019
DOI 10.4267/2042/70491
![]()
[Editor: Bruno Granier;
language editor: Simon Mitchell]
![]()
Lower Jurassic (Hettangian-Pliensbachian) shallow-marine ooidal limestones of southern Montenegro contain a large number of isocrinid ossicles. They are assigned to the following taxa: Isocrinus psilonoti (Quenstedt), Isocrinus sp., and Pentacrinites cf. fossilis Blumenbach. The echinoderm assemblage also yields cyrtocrinid ossicles (Cotylederma sp., Cyrtocrinina indet.) and echinoid spines (only spotted in thin sections); however, these elements are rare. Given the fact that the recorded assemblage comes from a single locality, there is a growing need for further research that will require intense sampling to compile and complete the faunal list of crinoids and other echinoderm taxa.
echinoderms;
crinoids;
Lower Jurassic;
Hettangian-Pliensbachian;
Montenegro
Salamon M.A. (2019).- A new prospect in crinoid (Crinoidea, Echinodermata) research: An example from the Lower Jurassic of Montenegro.- Carnets Geol., Madrid, vol. 19, no. 12, p. 211-220.
Une nouvelle perspective dans l'étude des crinoïdes (Crinoidea, Echinodermata) : Un exemple du Jurassique inférieur du Monténégro.- Les calcaires oolithiques du Jurassique inférieur (Hettangien-Pliensbachien) du sud du Monténégro renferment un grand nombre d'éléments squelettiques de crinoïdes isocrinides. Ils sont attribués aux taxons suivants : Isocrinus psilonoti (Quenstedt), Isocrinus sp. et Pentacrinites cf. fossilis Blumenbach. L'échinofaune comporte également des pièces de crinoïdes cyrtocrinides (Cotylederma sp., Cyrtocrinina indet.) et des radioles d'oursin (seulement repérés en lames minces); toutefois, ces éléments sont rares. Compte tenu du fait que l'assemblage répertorié provient d'une seule localité, il importe de poursuivre les recherches, ce qui nécessitera, entre autres, un échantillonnage plus intensif afin d'établir et de compléter la liste des faunes de crinoïdes et autres échinodermes.
échinodermes ;
crinoïdes ;
Jurassique inférieur ;
Hettangien-Pliensbachien ;
Monténégro
To date Early Jurassic crinoids of the former Yugoslavia have remained undescribed. However, as documented in the present study, they represent a common component of the fossil remains that can be found in the Hettangian-Pliensbachian interval. The existing literature, mostly geological or general paleontological papers (yet not directly, partly or exclusively, dedicated to crinoids), only briefly mentions their presence in sedimentary rocks (e.g., ČaĐenović et al., 2008; Črne & Goričan, 2008; Radulović, 2013; Bucković & Sviličić, 2016). The latter authors mentioned or illustrated echinoderms (mostly crinoids in thin sections) from the Lower Jurassic of Croatia, Montenegro, Serbia or Slovenia. Especially abundant are echinoderms in Montenegro, with local mass-occurrences within ooidal limestones (e.g., Fig. 4.f in Črne & Goričan, 2008; this study). Đaković et al. (2017) illustrated isocrinid pluricolumnals and columnals from the Rumija Mountains (Montenegro). The latter authors assigned their material to Chladocrinus basaltiformis (Miller), however, it is difficult to determine that this material really belongs to this taxa (there is no tuberculation present on the lateral surfaces, and all but one of the articular facets has a visible ornamentation). These are also mentioned in the Liassic of Greece (e.g., Karakitsios et al., 2015). Still noteworthy is the only detailed study of Early Jurassic (Pliensbachian) crinoids from Balkans (Bulgaria): Klikushin (1987) identified cyrtocrinids [Cyrtocrinida; Cotylederma manchevi Klikushin], isocrinids [Isocrinida; Chladocrinus basaltiformis (Miller), "Isocrinus" schlumbergeri (Loriol), Seirocrinus laevisutus (Pompecki)], and millericrinids [Millericrinida; Amaltheocrinus amalthei (Quenstedt)]. Significantly the crinoids documented from the Sinemurian-Toarcian interval of the neighbouring countries are more diverse. Delogu & Nicosia (1987) and Nicosia (1991) reported 14 different taxa in NW Turkey. Besides cyrtocrinids, isocrinids and millericrinids, they also noticed comatulids (Comatulida). A similar level of taxonomic diversity among crinoids for the same interval can be found in Italy (15 taxa of cyrtocrinids, isocrinids and millericrinids; Parona, 1892; Manni & Nicosia, 1990, 1999).
Present investigations, as a preliminary study, had been carried out in the surroundings of Tejani; they document a low diversity, isocrinid-dominated assemblage. There are good opportunities to find material in the Lower Jurassic oolitic limestone belt, which stretches for over a few tens of kilometers, south- and southeast-wards close to the Albanian border, and north- and northwest-wards along Lake Skadar.
This preliminary note aims to describe the crinoids, and to compare them with previously recorded forms from adjacent areas.
The
study area is located in the southern part of Montenegro, close to the Albanian
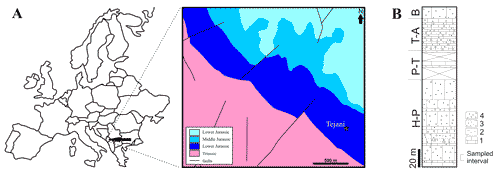
border (Fig. 1 ![]() ). The outcrops around Tejani consist of carbonate sediments of
the Lower and possibly Middle Jurassic (Črne & Goričan,
2008; Fig. 1
). The outcrops around Tejani consist of carbonate sediments of
the Lower and possibly Middle Jurassic (Črne & Goričan,
2008; Fig. 1 ![]() ). The lowermost deposits are thin-bedded limestones
that lack any
macrofaunal remains. Above this are thick-bedded ooidal limestones, ca. 100 m-thick,
containing numerous isocrinids, echinoids (Fig. 2.G
). The lowermost deposits are thin-bedded limestones
that lack any
macrofaunal remains. Above this are thick-bedded ooidal limestones, ca. 100 m-thick,
containing numerous isocrinids, echinoids (Fig. 2.G ![]() ), algae, and foraminifers.
According to Črne & Goričan
(2008), the
foraminiferal assemblage displays index taxa indicative of the
Hettangian-Pliensbachian interval [e.g., Agerina
martana (Farinacci)]. Higher deposits are marls, marly
limestones, and limestones, with scarce microfossils, which are mostly represented by
radiolarians and sponge spicules. They are overlain by oolitic limestones, with
locally abundant crinoids, brachiopods, and foraminifers, indicative of the
middle? Toarcian-Aalenian interval. The topmost part of the Tejani section is
represented by thick-bedded ooidal limestones with algae and foraminifers, most
probably of Bajocian age (Črne & Goričan,
2008).
), algae, and foraminifers.
According to Črne & Goričan
(2008), the
foraminiferal assemblage displays index taxa indicative of the
Hettangian-Pliensbachian interval [e.g., Agerina
martana (Farinacci)]. Higher deposits are marls, marly
limestones, and limestones, with scarce microfossils, which are mostly represented by
radiolarians and sponge spicules. They are overlain by oolitic limestones, with
locally abundant crinoids, brachiopods, and foraminifers, indicative of the
middle? Toarcian-Aalenian interval. The topmost part of the Tejani section is
represented by thick-bedded ooidal limestones with algae and foraminifers, most
probably of Bajocian age (Črne & Goričan,
2008).
Field
investigations in the Tejani area had been conducted in 2017-2018. From the
lower part of the section (Rumija Oolites Fm, Hettangian-Pliensbachian; Fig. 1 ![]() ) 32 small (0.4 kg - 0.7 kg) samples were
collected.
Additionally a ~40 kg bulk sample of weathered material was also collected.
In the Laboratory of Palaeontology and Stratigraphy at the University of Silesia
in Katowice, the weathered sample was washed through with hot water and screened on a
sieve column, using 1.0, 0.5, 0.315 and 0.1 mm mesh sizes. After
drying residues at 220°C, the fossil remains were picked
manually under a binocular microscope.
) 32 small (0.4 kg - 0.7 kg) samples were
collected.
Additionally a ~40 kg bulk sample of weathered material was also collected.
In the Laboratory of Palaeontology and Stratigraphy at the University of Silesia
in Katowice, the weathered sample was washed through with hot water and screened on a
sieve column, using 1.0, 0.5, 0.315 and 0.1 mm mesh sizes. After
drying residues at 220°C, the fossil remains were picked
manually under a binocular microscope.
Fossils showing on rock surface were photographed and, subsequently, treated with Glauber's salt (i.e., frozen and thawed at least 9 times and, after relaxation, washed in the same way as the weathered sample). An additional set of ten thin sections was prepared and analyzed by means of microfacies.
The investigated material is housed at the Faculty of Earth Sciences, University of Silesia in Katowice, Sosnowiec, under catalogue number GIUS 8-3667.
|
Figure 1: A. Map of Europe with area of Montenegro shaded in black, and the geological map of Tejani area (taken from Čađenović et al., 2014; simplified). B. Studied section. H-P - Hettangian-Pliensbachian, P-T - Pliensbachian-Toarcian, T-A - Toarcian-Aalenian, B-Bajocian (taken from Črne & Goričan, 2008, modified). Figure 1 : A. Carte de
l'Europe, le
Monténégro est indiqué en noir, et carte géologique de la région de Tejani
(tirée de Čađenović
et al.,
2014, simplifiée). B. Coupe étudiée. H-P -
Hettangien-Pliensbachien, P-T - Pliensbachien-Toarcien, T-A -
Toarcien-Aalénien, B-Bajocien (modifié d'après Črne & Goričan,
2008). |
Systematics follows the taxonomic views expressed by Hess (2006) and Hess & Messing (2011).
Order Isocrinida Sieverts-Doreck in Moore et al., 1952
Suborder Isocrinina Sieverts-Doreck in Ubaghs, 1953
Family Isocrinidae Gislén, 1924
Subfamily Isocrininae Gislén, 1924
Genus Isocrinus Meyer in Agassiz, 1836
Type species: Isocrinites pendulus Meyer in Agassiz, 1836
Isocrinus psilonoti (Quenstedt, 1858)
(Fig. 3.J-K ![]() )
)
1989 Isocrinus psilonoti (Quenstedt); Simms, Figs. 10-16; Pl. 1, fig. 1; Pl. 7, figs. 1-7, 11-12, 15, 18-21; Pl. 8, figs. 24, 29.
Material. 17 isolated columnals (internodals and nodals).
Description. Columnals are pentalobate to sub-stellate, and stellate. Nodals and internodals are of almost the same size. Columnal diameter ranges from 3.2 to 7.0 mm. Articular facets are flat and covered by drop-like petal floors. Each petal is surrounded by max. 24 culmina. Radial crenulae forming a slit visible as a radial pore on the latus. Latera is straight and smooth. Cirrus scars are small.
Discussion. Głuchowski (1987) collected Isocrinus tuberculatus (Miller) and 'I'. angulatus (Oppel) from the same horizon (Hettangian-Sinemurian) in the Tatra Mountains (southern Poland). This author stated that they differ in morphology of facets and latera. In the case of 'I'. angulatus, the facets are covered by large culmina that are in contact in the radial part and thus cover nearly the whole facet surface. In contrast to I. tuberculatus, crenulation of adjacent petals is not in contact in the radial part. Moreover, the facets of I. tuberculatus are covered by small tubercles. When comparing facets of 'I'. angulatus with those of I. psilonoti found in Tejani, they are identical. A similar opinion was presented by Simms (1989, 2010) who also pointed out that I. psilonoti differs from all pre-Toarcian representatives of Isocrinus in the lack of ornamentation of columnal latera. He also noted that the cirrus scars of I. psilonoti are larger than those of other Lower Jurassic taxa recorded from Europe [I. robustus (Wright) and I. tuberculatus; see also Głuchowski, 1987; Klikushin, 1992].
Isocrinus sp.
(Figs. 2.A-F ![]() ,
3.E-H
,
3.E-H ![]() )
)
Material. Several columnals (nodals and internodals, pluricolumnals), brachials and cirrals. We must bear in mind that some of the ossicles classified here as Isocrinus sp. could belong to I. psilonoti (Quenstedt) as described below.
Description. Columnals are circular, pentagonal, pentalobate to sub-pentalobate. Nodals are higher than internodals. Columnal diameter ranges from 0.6 mm up to 3.9 mm. Small columnals are nearly as high as wide. Articular facets are not visible (or very poorly visible; covered by rock matrix), with smooth latera (very often smeared by rock matrix). Morphology of cirrus scars is not visible. Cirrals are elliptical in outline, with smooth surface. Proximal cirrals are very short; distal cirrals are longer. Cirral articulum is concave, with distinct, sometimes very high perilumen. A few cirrals bearing a sigmoidal perilumen, inclined ca. 20° to short axis. Fulcral ridge of cirral possesses paired pointed projections adorally. Cirral latera is smooth. Brachials have smooth surfaces; however, in some cases small granulae are visible. Proximal brachials are wider than high, meanwhile distal brachials are small and much higher than wide. Almost all isolated brachials are covered by strongly lithified rock matrix. In thin sections some cryptosyzygial brachials are visible.
Discussion.
Some isocrinid taxa were described from the Lower Jurassic strata of Europe.
Most of them possess pentagonal to pentalobate or sub-pentalobate columnals (Loriol,
1889; Parona, 1892; Jäger,
1985; Klikushin, 1987; Simms,
1989; Nicosia, 1991; Hess,
2006). However some
circular columnals from Tejani could also be classified as balanocrinids. Despite
the lack of observation of their articular faces their circular columnal shape
is usually preserved for this isocrinid group (comp. e.g., Simms,
1989,
Pl. 10, figs. 18-20, 25; Hess 2006, Pl. 24, fig. 2; see
Fig. 4.f in Črne
& Goričan, 2008, and Fig. 2.E ![]() here). According to Hess
& Pugin (1983) and Hess
(2012), balanocrinids are absent in the the north
here). According to Hess
& Pugin (1983) and Hess
(2012), balanocrinids are absent in the the north-western Tethysian carbonate platform settings, but commonly occur in
siliciclastic deposits, especially those bordering the open-oceanic Tethys. On
the other hand, these crinoids are a dominant component in the shelf carbonate
areas of central Europe (e.g., Poland; Salamon & Zatoń,
2006; Salamon, 2008; Zatoń et
al., 2008).
It cannot be excluded that some pentagonal columnals classified here as Isocrinus sp. belong to other crinoid groups (e.g., millericrinids). Hess (2006) illustrated pentagonal columnals and described them as millericrinids (Millericrinus? cf. adneticus, Millericrinus? quinquepictus; see Pl. 13, figs. 7, 11-15; Pl. 14, figs. 5-6 in Hess, 2006). The latter author proceeded likewise with unornamented secundibrachials that possessed oblique articular articulation: they were described as Catinocrinus (Pl. 21, fig. 1.a in Hess, 2006). Such an ornamentation can be discerned in some specimens from Tejani.
Suborder Pentacrinitina Gray, 1842
Family Pentacrinitidae Gray, 1842
Genus Pentacrinites Blumenbach, 1804
Type species. Pentacrinites fossilis Blumenbach, 1804
Pentacrinites cf. fossilis Blumenbach, 1804
(Fig. 3.D ![]() )
)
1804 Pentacrinites fossilis; Blumenbach, no. 7, pt. 70, Fig. 70.
Material. 6 isolated cirrals plus a dozen cirrals on slab surfaces and in TS.
Description. Cirrals are compressed and ellipsoidal in outline. They are recrystallized, and their articular facets are not visible.
Discussion. Among the Early Jurassic representatives of genus Pentacrinites, P. fossilis is the commonest one within the Sinemurian-Pliensbachian interval. Complete specimens or columnals are recorded in the Caucasus, England and Germany (Sieverts-Doreck, 1978; Klikushin, 1987; Simms, 1989). Unique, ellipsoidal or rhomboidal in outline, cirrals from Early Jurassic strata were also classified as P. fossilis (e.g., Hess, 2006; Salamon et al., 2008; Hess & Messing, 2011, Fig. 21.i). Additionally Simms (1989) mentioned two representatives of this genus [P. dorecki Simms and P. dichotomus (McCoy)], but underlined that they are seldom found in the fossil record and are only known from the British Isles. Therefore, their occurrence in the Balkans seems unlikely. On the hand, it cannot be formally excluded because Pentacrinites is commonly regarded as a pseudo-planktonic taxon with high dispersal capabilities.
Order Cyrtocrinida Sieverts-Doreck in Moore et al., 1952
Suborder Holopodina Arendt, 1974
Family Cotyledermatidae Wright, 1876
Genus Cotylederma Quenstedt, 1852
Type species. Cotylederma lineati Quenstedt, 1856
Cotylederma sp.
(Fig. 3.I ![]() )
)
Material. 3 isolated and partly preserved primibrachials.
Description. Primibrachials are very thin and wide: ranging from 0.6 mm high and 1.9 mm wide, up to 1.2 mm high and 3.7 mm wide. They are slightly asymmetrical. Articular facets are almost identical. One lateral edge is higher than the other. Aboral ligament fossae is not distinct, and adoral muscle fossae is rather small and moderately deep. Fulcral ridge is separating the aboral side from the adoral one; it is sharp and without crenulae.
Discussion. It is difficult to conclusively state which species of Cotylederma are present at Tejani. Among the many species (for details see Quenstedt, 1856; Loriol, 1889; Klikushin, 1987; Manni & Nicosia, 1990; Jäger, 1991; Nicosia, 1991; Hess, 2006; Hess et al., 2011) almost (or all) present more or less asymmetrical radials and rather thin brachials. Hess et al. (2011) reported that brachials from Upper Jurassic-Lower Cretaceous strata of eastern Poland are practically indistinguishable from the Lower Jurassic brachials described by Jäger (1991) or Nicosia (1991).
Suborder Cyrtocrinina Sieverts-Doreck, 1952
Cyrtocrinina indet.
Material. 4 isolated primibrachials, 32? isolated brachials and 5? isolated columnals plus some remains occurring on the slab surfaces and in TS.
Description. Primibrachials are fused and massive with lateral extension. Proximal facets with pronounced adoral furrow, and distal ones synostosial. One of these latter facets is crescentic and crypto-syzygial. Brachial plates are small and thin, rectangular or more often U-shaped. Feeding groove displaced to only one side. Muscular fossae are rather small and very often indistinct. Some brachials having synostoses on every side, thin, without any pinnular socket. Other brachials are muscular. Columnals are very low and elliptical in outline, their facet being covered by short and relatively thick crenulae. Latera is smooth? Among the material is a certain percentage of small, thin brachials with food groove displaced to one side.
|
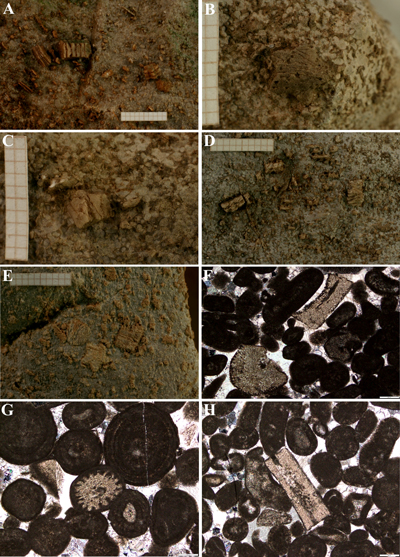
Figure 2: Crinoids (isocrinids) from Tejani section, Montenegro, Lower Jurassic. A-E. Abraded columnals, pluricolumnals, brachials and cirrals visible on slab surfaces. Ruler as a scale. F-H. Ooid grainstone with fragments of echinoid. Thin sections. Scale bar equals 0.2 mm. F. Cryptosyzygial brachial (in the lower part), and undeterminatable brachial, cyrtocrinid? (above). G. Echinoid spine section. H. Cyrtocrinid? columnal section. Figure
2 : Crinoïdes (isocrinides)
de la coupe de Tejani, Monténégro, Jurassique inférieur. A-E. Columnales,
pluri-columnales, brachiales et cirrales érodées visibles à la surface des
blocs. La règle donne l'échelle. F-H. Ooid grainstone avec des fragments
d'échinoïdes. Lames minces. Échelle = 1
mm. F. Brachiale cryptosyzygiale (dans la partie inférieure), et brachiale
indéterminable, cyrtocrinide? (au-dessus). G. Section de radiole d'oursin. H. Section de
columnale de cyrtocrinide ? |
|
Figure 3: Crinoids from Tejani section, Montenegro, Lower Jurassic. Scale bar equals 1 mm. A. Cyrtocrinid columnal, articular face. B. Isocrinid pluricolumnal, latera. C. Cyrtocrinid? brachial, proximal view. D. Pentacrinites cf. fossilis Blumenbach cirral, articular face. E. Isocrinid cirral, articular face. F-G. Isocrinid columnals, articular faces. Arrow indicates an epibiont. H. Juvenile isocrinid, nodal, latera. I. Cotylederma sp., primibrachial, adoral view. J-L. Isocrinus psilonoti (Quenstedt) columnals, articular faces. Arrow indicate an epibiont. Figure 3 : Crinoïdes de la coupe de
Tejani, Monténégro, Jurassique inférieur. Échelle = 1 mm. A. Columnale de
cyrtocrinide, face articulaire. B. Pluri-columnale d'isocrinid, face latérale.
C. Brachiale de cyrtocrinide?, vue proximale. D. Cirrale de Pentacrinites
cf. fossilis Blumenbach, face articulaire. E. Cirrale
d'isocrinide, face articulaire. F-G. Columnales d'isocrinide, faces
articulaires. La flèche montre l'épibionte. H. Nodale d'isocrinide juvénile,
face latérale. I. Primibrachiale de Cotylederma sp., vue adorale.
J-L. Columnales d'Isocrinus psilonoti (Quenstedt). La flèche
montre l'épibionte. |
Almost all new crinoid clades that originated in the fossil record after the end-Permian mass extinction, are also known in the Jurassic period, except for some "microcrinoids" (e.g., Kristan-Tollmann, 1980, 1990) and roveacrinids (e.g., family Somphocrinidae; Hess & Messing, 2011). There are also some doubts concerning the early records of cyrtocrinids. Hess & Messing (2011) claimed that they appear only in the Hettangian but, somewhat earlier, the first of these authors (Hess, 2006) stated that some still undescribed forms resembling cyrtocrinids (eudesicrinids) occur in the Carnian Hanwang Formation of China. Likewise Salamon et al. (2009) described some Rhaetian columnals from the Tatra Mountains (southern Poland), probably representing cyrtocrinids or less likely millericrinids; on the other hand, these columnals do not belong to encrinids that had their last appearance in the middle Carnian.
Simms (1988, 1990) stated that crinoid diversity was very low during the Early Jurassic (Sinemurian) and did not start increasing until mid-Sinemurian. He also stated that crinoid diversity levels comparable with those of the Late Triassic were achieved not earlier than the Middle Jurassic. These two assertions (Simms, 1988, 1990) are not supported by recent data on crinoid diversity dynamics by Gorzelak et al. (2016), which demonstrated that the mean standing diversity of crinoids linearly increased during this time interval. One has to bear in mind that the Triassic/Jurassic mass extinction was significantly less severe than the previous major one (P/T) (Twitchett & Oji, 2005). According to Hess (2006) the species richness of crinoids known from the earliest Jurassic is surprisingly high, and the crinoid diversity in the Early Jurassic is nearly comparable to that of the Late Jurassic (Oxfordian and Kimmeridgian). On the other hand, the Early Jurassic increase in crinoid diversity was largely due to the fast radiation of cyrtocrinids. This latter author described the most diverse crinoid fauna yet recorded from the Lower Jurassic (Pliensbachian): from Arzo (Switzerland), besides 13 cyrtocrinid genera he listed also millericrinids (9 genera), comatulids (2 genera), isocrinids (2 genera), and a large number of undeterminable ossicles. Similar crinoid faunas were also reported from the adjacent countries. In the first study ever published, Parona (1892) recorded nine taxa from the Sinemurian (or Pliensbachian) of northern Italy, dominated by millericrinids and isocrinids, and accompanied by Eudesicrinus mayalis (Deslongchamps & Deslongchamps). Manni & Nicosia (1990, 1999) completed this list with six taxa of cyrtocrinids from the Pliensbachian and Toarcian of central Italy. Similar patterns of diversification of Early Jurassic crinoids are also noted from Bulgaria (Pliensbachian), England (Sinemurian and Pliensbachian), France (Pliensbachian-Toarcian), Germany (Sinemurian and Pliensbachian), Russia (Caucasus, Pliensbachian), Turkey (Sinemurian and Pliensbachian) and the Ukraine (Crimea, Sinemurian and Pliensbachian; Loriol, 1889; Delogu & Nicosia, 1987; Jäger, 1985, 1991, 1993; Klikushin, 1987; Simms, 1989; Nicosia, 1991; Hess & Thuy, 2017; Table 1). Hess & Thuy (2017) recently suggested that the Pliensbachian/Toarcian is a hotspot of comatulid radiation.
Table 1. List
of Early Jurassic crinoid taxa noted world-wide (data after Loriol,
1889; Delogu & Nicosia,
1987; Jäger, 1985,
1991, 1993;
Klikushin, 1987, 1992; Simms,
1989; Nicosia, 1991; Hess
2006; Salamon et al., 2008; Hess
& Thuy, 2017).
Table 1. Liste des taxons de crinoïdes du Jurassique inférieur répertoriés à l'échelle mondiale (données compilées de Loriol, 1889; Delogu & Nicosia, 1987; Jäger, 1985, 1991, 1993; Klikushin, 1987, 1992; Simms, 1989; Nicosia, 1991; Hess, 2006; Salamon et al., 2008; Hess & Thuy, 2017).
| area (in alphabetical order) |
number of crinoid taxa |
number of cyrtocrinid taxa |
number of isocrinid taxa |
number of millericrinid taxa |
number of comatulid taxa |
| Algeria and Morocco | 3 | 0 | 3 | 0 | 0 |
| Bulgaria | 5 | 1 | 3 | 1 | 0 |
| England | 12 | 1 | 10 | 0 | 1 |
| France | 19 | 3 | 5 | 3 | 8 |
| Germany | 18 | 5 | 9 | 4 | 0 |
| Italy | 15 | 7 | 2 | 6 | 0 |
| Poland and Slovakia (Pieniny Klippen Belt) | 6 | 1 | 4 | 0 | 1 |
| Russia (including Siberia), plus Armenia, Azerbaijan, Georgia | 11 | 0 | 6 | 5 | 0 |
| Switzerland | 26 | 13 | 2 | 9 | 2 |
| Ukraine | 8 | 0 | 3 | 5 | 0 |
| Turkey | 14 | 5 | 3 | 5 | 1 |
The present study only documented 5 crinoid taxa, but all the samples
were taken merely from a single horizon at Tejani (Fig. 1 ![]() ). Similarly
low-diversity crinoid assemblages, dominated by isocrinids (5 taxa), are known
from the Lower Jurassic (Sinemurian-Toarcian) deposits of Algeria, Armenia,
Azerbaijan, Georgia, Russian Siberia and Slovakia but, once again in these locations, the samples originated
from isolated horizons, and not from comprehensive formations (Klikushin,
1992; Salamon et al., 2008).
Furthermore the presence of millericrinids at Tejani cannot be excluded (see Systematic
palaeontology section above).
). Similarly
low-diversity crinoid assemblages, dominated by isocrinids (5 taxa), are known
from the Lower Jurassic (Sinemurian-Toarcian) deposits of Algeria, Armenia,
Azerbaijan, Georgia, Russian Siberia and Slovakia but, once again in these locations, the samples originated
from isolated horizons, and not from comprehensive formations (Klikushin,
1992; Salamon et al., 2008).
Furthermore the presence of millericrinids at Tejani cannot be excluded (see Systematic
palaeontology section above).
We should also take into account that the studied deposits document a shallow-water high-energy environment, unfavourable to the appropriate preservation of echinoderm remains necessary for their accurate classification. Crinoids in this location are highly fragmented: pluri-columnals are relatively rare, complete calyces are absent, cirrals are typically broken; articular surfaces of ossicles are strongly abraded. Subsequently these elements might have undergone some transport (details in Gorzelak & Salamon, 2013). Before final burial the articular faces of columnals in some studied fossils were recovered and overgrown by epibionts (foraminifers, annelids); these latter are also displaying very small and rounded holes likely produced by acrothoracican cirripeds, algae, fungi, polychaetids, sipunculans or even sponges. Furthermore, a strong micritization of ooid cortex, dissolution of fragments of echinoderm elements, as well as the lack of preserved ooid cores, which were dissolved and replaced by sparite/microsparite crystals, point to the early diagenetic changes (comp., Richter, 1983). In marine environments, ooids are typically formed in intertidal and shallow-subtidal settings, and in carbonate platforms (between lagoons and open sea; Wilson, 1975). Their occurrence at great depths is due to redeposition by turbidity currents (Flügel, 2004). In the material at hand, however, no evidence of ooid redeposition is noticed (e.g., rounded bioclasts). The symmetrical shapes observed in the studied ooids, their good sorting and the absence of large irregularly shaped forms indicate a high-energy depositional environment within a wave zone (Flügel, 2004). This is consistent with the conclusions of Črne & Goričan (2008) that are based on sedimentary structures observed in Tejani.
The observations reported above let us expect that a systematical sampling of the Lower Jurassic deposits representing different facies cropping out along Lake Skadar (Montenegro) should provide additional materials, hopefully with better preservation, and therefore more suitable for taxonomic study and potential to enlarge the faunal list with additional taxa. These expectations are also supported by the observations of Črne & Goričan (2008, Fig. 2) who reported the presence of crinoids not only in the Hettangian-Pliensbachian interval, but also in younger deposits of this area. Most importantly, however, further documentation of the earliest Jurassic cyrtocrinids in these shallow marine facies would be of particular importance as it may challenge the view that cyrtocrinids originated in deep-sea environments and subsequently migrated to shallow Tethysian shelves from the Sinemurian onward (Hess & Thuy, 2017).
The author is greatly indebted to reviewers for their constructive comments and helpful suggestions. Thanks are due to Dr Wojciech Krawczyński and Dr Krzysztof Szopa (both from the University of Silesia in Katowice, Sosnowiec, Poland) for preparing photographs, and Dr Przemysław Gorzelak for help during field works. The epibiont assignment of Prof. Michał Zatoń (University of Silesia in Katowice, Sosnowiec, Poland) is gratefully acknowledged. Special thanks go to Dr Bruno Ferré (Sotteville-lès-Rouen, France) and Dr Robert Niedźwiedzki (University of Wrocław, Poland) for helpful discussion concerning crinoids in TS. Dr Damjan ČaĐenović (Geological Survey of Montenegro, Podgorica) is acknowledged for providing some geological information and making some valuable comments on the early draft of the manuscript.
Agassiz J.L.R. (1836).- Prodrome d'une Monographie des Radiaires ou Échinodermes.- Mémoires de la Société des Sciences Naturelles de Neuchâtel, vol. 1, p. 168-199.
Arendt Y.A. (1974).- Morskie lilii. Tsirtokrinidy.- Trudy Paleontologicheskego Instituta, Akademia Nauk SSSR, vol. 144, p. 1-251.
Blumenbach I.F. (1804).- 70. Pentacrinites fossilis.- Abbildungen naturhistorischer Gegenstände, Göttingen, Heft 7, 36 p. URL: https://gdz.sub.uni-goettingen.de/id/PPN625161807_0007
Bronn H.G. & Roemer F. (1851-1856).- Lethaea geognostica oder Abbildung und Beschreibung der für die Gebirgs-Formationen bezeichnendsten Versteinerungen. Erster Band.- Schweizerbart, Stuttgart, viii + 412 p., Atlas (124 Pls.).
Bucković D. & Sviličić V. (2016).- Jurassic sequences of Soice and Rovinj localities (Karst Dinarides, Croatia) as examples of long-distance tectonic-controlled deposition on the Adriatic-Dinaric carbonate platform.- International Journal of Earth & Environmental Sciences, Bangalore, vol. 110, p. 1-8.
ČaĐenović D., Kilibarda Z. & Radulović N. (2008).- Late Triassic to Late Jurassic evolution of the Adriatic Carbonate Platform and Budva Basin, Southern Montenegro.- Sedimentary Geology, vol. 2004, p. 1-17.
ČaĐenović D., Radulović N., Milutin J. & Đaković M. (2014).- New geological map of Montenegro, representation of results from sheet Podgorica-3, 1:50000. In: Pangeo Austria 2014.- Berichte des Institutes für Erdwissenschaften Karl-Franzens-Universität Graz, Band 20/1, p. 34.
Črne A. & Goričan . (2008).- The Dinaric Carbonate Platform margin in the Early Jurassic: A comparison between successions in Slovenia and Montenegro.- Italian Journal of Geosciences, Roma, vol. 127, p. 389-405.
Đaković M., Čađenović D., Radulović B., Milić M. & Radulović N. (2017).- Chladocrinus basaltiformis (Miller) and Zeilleria mutabilis (Oppel) from the Pliensbachian oolitic limestone of the Rumija Mt. (Southern Montenegro).- EGU series: Émile Argand Conference, 13th Workshop on Alpine Geological Studies September 7th-18th 2017, Zlatibor Mts. (Serbia), p. 30.
Delogu D. & Nicosia U. (1987).- A new cyrtocrinid from the Lias of Turkey: Capsicocrinus souti n.gen., n.sp.- Geologica Romana, vol. 23, p. 157-162.
Flügel E. (2004).- Microfacies of carbonate rocks.- Springer Verlag, Berlin-Heidelberg-New York, 976 p.
Gislén T. (1924).- Echinoderm Studies.- Zoologiska Bidrag, Uppsala, vol. 9, 330 p.
Głuchowski E. (1987).- Jurassic and early Cretaceous Articulate Crinoidea from the Pieniny Klippen Belt and Tatra Mts, Poland.- Studia Geologica Polonica, Krakowie, vol. XCIV, p. 6-102.
Gorzelak P. & Salamon M.A. (2013).- Experimental tumbling of echinoderms - taphonomic patterns and implications.- Palæogeography, Palæoclimatology, Palæoecology, vol. 386, p. 569-574.
Gorzelak P., Salamon M.A., Trzęsiok D., Lach R. & Baumiller T.K. (2016).- Diversity dynamics of post-Palaeozoic crinoids in quest of the factors affecting crinoid macroevolution.- Lethaia, vol. 49, p. 231-244.
Gray J.E. (1842).- Synopsis of the contents of the British Museum, 44th edition.- British Museum, London, IV+308 p.
Hess H. (2006).- Crinoids (Echinodermata) from the Lower Jurassic (Upper Pliensbachian) of Arzo, southern Switzerland.- Schweizerische Paläontologische Abhandlungen, Basel, vol. 126, p. 1-144.
Hess H. (2012).- Crinoids from the Middle Jurassic (Bajocian-Lower Callovian) of Ardèche, France.- Swiss Journal of Palaeontology, vol. 131, p. 211-253.
Hess H. & Messing C.G. (2011).- Treatise on invertebrate paleontology, part T (revised), volume 3: Echinodermata 2 (Crinoidea).- The University of Kansas, Paleontological Institute, Lawrence, 261 p.
Hess H. & Pugin L. (1983).- Balanocrinus berchteni n.sp., un nouveau crinoïde bajocien des Préalpes médianes frigourgeoises.- Eclogae Geologicae Helvetiae, vol. 76, p. 691-700.
Hess H., Salamon M.A. & Gorzelak P. (2011).- Late Jurassic-Early Cretaceous (Tithonian-Berriasian) cyrtocrinids from south-eastern Poland.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 260, p. 119-128.
Hess H. & Thuy B. (2017).- Extraordinary diversity of feather stars (Echinodermata: Crinoidea: Comatulida) from a Lower Jurassic (PliensbachianToarcian) rock reef of Feuguerolles (Normandy, France).- Swiss Journal of Palaeontology, vol. 136, p. 301-321.
Jäger M. (1985).- Die Crinoiden aus dem Pliensbachium (mittlerer Lias) von Rottorf am Klei und Empelde (Süd-Niedersachsen).- Beiträge die Naturhistorische Gesellschaft Hannover, vol. 128, p. 71-151.
Jäger M. (1991).- Die Crinoiden aus dem Pliensbachium vom Rhein-Main-Donau-Kanal bei Sulzkirchen. Teil 1: Cyrtocriniden mit reduziertem Stiel: Eudescicrinus und Cotylederma.- Archeopteryx, Munich, Band 9, p. 35-46.
Jäger M. (1993).- Die Crinoiden aus dem Pliensbachium vom Rhein-Main-Donau-Kanal bei Sulzkirchen. Teil 2: Gestielte Cyrtocrinina und Millericrinina: Amaltheocrinus, Sroshaecrinus?, Plicatocrinus und Nachtrag zu Teil 1 (Eudesicrinus).- Archeopteryx, Munich, Band 11, p. 73-104.
Karakitsios V., Kvaček Z. & Mantzouka D. (2015).- The first plant megafossil in the Early Jurassic of Greece: Brachyphyllum (Coniferales) from the Lower Posidonia Beds (Toarcian) in the Ionian zone (NW Greece) and its palaeogeographic implications.- Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, Stuttgart, vol. 278, p. 79-94.
Klikushin V.G. (1987).- Crinoids from the Middle Liassic Rosso ammonitico beds.- Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, Stuttgart, vol. 175, p. 235-260.
Klikushin V.G. (1992).- Fossil pentacrinid crinoids and their occurrence in the USSR.- Leningrad Paleontological Laboratory, 358 p.
Kristan-Tollmann H. (1980).- Tulipacrinus tulipa n.g. n.sp., eine Mikrocrinoide aus der alpinen Obertrias.- Annalen des Naturhistorischen Museums in Wien, vol. 83, p. 215-229.
Kristan-Tollmann H. (1990).- Mikrocrinoiden aus der Obertrias der Tethys.- Geologisch- paläontologische Mitteilungen Innsbruck, vol. 17, p. 51-100.
Loriol P. de (1889).- Paléontologie francaise, ou description des fossiles de la France, s. 1, Animaux invertébrés. Terrain jurassique 11, Crinoïdes.- Masson, Paris, 1207 p.
Manni R. & Nicosia U. (1990).- New crinoids from Liassic sediments nearby Tivoli (central Italy): Rearrangment in the systematics of eudesicrinids and cotyledermatids.- Geologica Romana, vol. 26, p. 81-92.
Manni R. & Nicosia U. (1999).- The oldest cyrtocrinid, Nerocrinus petri n. gen., n. sp., and the Nerocrinidae, n. fam.- In: Candia-Carnevali M.D. & Bonasero F. (eds.), Echinoderm research.- Balkema, Rotterdam, p. 327-331.
Moore R.C., Lalicker C.G. & Fischer A.G. (1952).- Invertebrate fossils.- McGraw-Hill, New York-Toronto- London, 766 p.
Nicosia U. (1991).- Mesozoic crinoids from the north-western Turkey.- Geologica Romana, vol. 27, p. 389-436.
Parona C.F. (1892).- Revisione della fauna liasica di Gozzano in Piemonte.- Memorie della Reale Accademia delle Scienze di Torino, vol. 43, 59 p.
Quenstedt P.A. (1852).- Handbuch der Petrefactenkunde.- Laupp, Tübingen, 792 p.
Quenstedt P.A. (1856-1858).- Der Jura.- Laupp, Tübingen, 842 p.
Quenstedt F.A. (1876).- Petrefactenkunde Deutschlands. 1. Abteilung, 4.- Fues's Verlag, Leipzig, 742 p.
Radulović B. (2013).- Lower Jurassic bivalves of eastern Serbia.- Beringeria, Würzburg, vol. 43, p. 3-61.
Richter D.K. (1983).- Calcareous ooids: A synopsis. In: Peryt T. (ed.), Coated grains.- Springer Verlag, Berlin-Heidelberg-New York-Tokyo, p. 71-99.
Salamon M.A. (2008).- Jurassic cyrtocrinids (Cyrtocrinida, Crinoidea) from extra-Carpathian Poland.- Palaeontographica Abteilung A, Stuttgart, vol. 285, p. 77-99.
Salamon M.A. & Gorzelak P. (2010).- Cyrtocrinids (Echinodermata, Crinoidea) from Upper Jurassic tramberk type limestones in southern Poland.- Palaeontology, London, vol. 53, p. 869-885.
Salamon M.A., Gorzelak P. & Schweigert G. (2008).- Early Jurassic (Sinemurian-Toarcian) crinoids (Crinoidea) from Central Europe (Poland, Slovakia, Germany).- Freiberger Forschungshefte C528, vol. 16, p. 93-103.
Salamon M.A., Gorzelak P. & Zatoń M. (2009).- In quest of cyrtocrinid origins: Evidence from Late Triassic ossicles from the Tatra Mountains.- Acta Palaeontologica Polonica, Warsaw, vol. 54, p. 171-174.
Salamon M.A. & Zatoń M. (2006).- Balanocrinus hessi n. sp., a new crinoid (Echinodermata) from the Callovian (Middle Jurassic) of southern Poland.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 240, p. 1-17.
Sieverts-Doreck H. (1952).- Articulata. In: Ubaghs G. (ed.), Classe des Crinoïdes.- Traité de Paléontologie, vol. 3, p. 658-773.
Sieverts-Doreck H. (1978).- Nachweis der Pentacrinites-fossilis-Gruppe im schwäbischen Arietenkalk (Lias alpha 3, unteres Sinemurium).- Stuttgarter Beiträge zur Naturkunde B, vol. 35, 15 p.
Simms M.J. (1988).- Patterns of evolution among Lower Jurassic crinoids.- Historical Biology, vol. 1, p. 17-44.
Simms M.J. (1989).- British Lower Jurassic crinoids.- Monograph of the Palaeontographical Society, London, vol. 142, p. 1-103.
Simms M.J. (1990).- Crinoid diversity and the Triassic/Jurassic boundary.- Cahiers de l'Université Catholique de Lyon, vol. 3, p. 67-77.
Simms M.J. (2010).- Stereom microstructure of columnal latera: A character for assessing phylogenetic relationships in articulate crinoids.- Swiss Journal of Palaeontology, vol. 130, p. 143-154.
Twitchett R.J. & Oji T. (2005).- Early Triassic recovery of echinoderms.- Comptes Rendus Palevol, vol. 4, no. 6, p. 531-542.
Ubaghs G. (1953).- Sous-Classe 4, Articulata J.S. Miller. In: Piveteau- J. (ed.), Traité de Paléontologie 3.- Masson, Paris, p. 756-765.
Wilson J.L. (1975).- Carbonate facies in geologic history.- Springer Verlag, New York, 471 p.
Wright T. (1876).- On the Cotyledermidae.- Geological Magazine, vol. 3, p. 94-95.
Zatoń M., Salamon M. & Kaźmierczak J. (2008).- Cyrtocrinids (Crinoidea) and associated stalked crinoids from the Lower/Middle Oxfordian (Upper Jurassic) shelfal deposits of southern Poland.- Geobios, Villeurbanne, vol. 41, p. 559-569.