◄ Carnets Geol. 19 (19) ►
![]()
Outline:
[1. Introduction]
[2. Geological background and outcrop]
[3. Material and methodsh]
[4. Results]
[5. Discussion and conclusions]
and ...
[Bibliographic references]
Department of Geology, University of Tartu, Ravila 14A, 50411 Tartu (Estonia)
Department of Geology and Natural Resources, Institute of Geosciences, State University of Campinas, 13083-855, Campinas, SP (Brazil)
CENPALEO, UnC, Av. Pres. Nereu Ramos, 1071, Mafra, SC, 89300-000 (Brazil)
Published online in final form (pdf) on December 27, 2019
DOI 10.4267/2042/70636
![]()
[Editor: Bruno Granier]
![]()
Lingulides from Lower Permian of Brazil (24% substrates) showed signs of bioerosion in form of multiple small shallow pits that resemble incomplete Oichnus paraboloides borings. A single lingulide valve showed a centrally located large circular predatory Oichnus simplex boring. Several lingulide shells (c. 21%) show small Arachnostega traces in their interior. The diameter of Arachnostega burrows is relatively constant. These burrows are more similar to juvenile stages of Arachnostega gastrochaena and do not form well-developed meshwork of tunnels. This is the first record of Arachnostega from the interior of lingulide shells and indicates that even very small lingulide shells were suitable substrates for cryptic organisms.
Brachiopods;
borings;
burrows;
Campo Mourão Formation;
Paraná Basin;
Brazil
Vinn O., Zabini C. & Weinschütz L.C. (2019).- Ichnofossils associated with lingulide shells from the Lower Permian of Brazil.- Carnets Geol., Madrid, vol. 19, no. 19, p. 439-444.
Ichnofossiles associés à des coquilles de lingulides du Permien inférieur du Brésil.- Les lingulides du Permien inférieur du Brésil (24% des substrats) portent des traces de bio-érosion sous forme de nombreux petits trous peu profonds qui ressemblent à des perforations incomplètes d'Oichnus paraboloides. Une seule coquille de lingulide montre une grande perforation prédatrice circulaire d'Oichnus simplex localisée en son centre. Plusieurs coquilles de lingulides (environ 21%) présentent des traces d'Arachnostega à l'intérieur. Le diamètre des terriers d'Arachnostega est relativement constant. Ces terriers sont très semblables aux stades juvéniles d'Arachnostega gastrochaena et ne forment pas de réseaux de tunnels bien développés. C'est la première mention d'Arachnostega à l'intérieur de coquilles de lingulides et ce qui prouve que des coquilles de lingulides, même très petites, pouvaient constituer un substrat convenable pour des organismes cryptiques.
Brachiopodes ;
perforations ;
terriers ;
Formation de Campo Mourão ;
Bassin de Paraná ;
Brésil
The Lingulidae Family is part of the Lingulida Order (Holmer & Popov, 2000). The lingulide brachiopods have chitino phosphatic shells (Holmer & Popov, 2000). They have a stratigraphic range from the early Cambrian to Recent (Holmer & Popov, 2000). The data on soft body anatomy of Early Cambrian lingulides from the Chengjiang Fauna (Zhang et al., 2003) include preservation of a pedicle, schizolophous lophophore, as well as a recurved, U-shaped digestive tract with a functional anus showing that major features of soft body anatomy characteristic of Recent lingulates were already present in early Cambrian. Sometimes fossil lingulide shells contain borings.
The earliest predatory borings have been described in Cloudina from the Ediacaran (Bengtson & Zhao, 1992). The Paleozoic record of predation is mostly based on drillings known from the shells of both lingulate and rhynchonellate brachiopods, and a lesser amount from mollusks (Brett & Walker, 2002; Brett, 2003; Harper, 2003; Huntley & Kowalewski, 2007). Vermeij (1977) has shown that predation was an important evolutionary factor since the early Cambrian. It is well known that predation intensities have increased from the Cambrian to the Recent (Vermeij, 1977, 1987; Vermeij et al., 1981). This increase in predation intensities is mirrored by the morphological evolution of the gastropod shell (Vermeij, 1977, 1987; Vermeij et al., 1981).
Often fossilized shells of various invertebrates contain burrows of the other invertebrates. The ichnofossil Arachnostega was first systematically described and named by Bertling (1992) accommodating irregular, branching burrows on the surface of internal moulds of various shelly fossils (e.g., Vinn et al., 2014). Arachnostega presumably belongs to the category fodichnia (Bertling, 1992), but it is equally possible that it in fact represents a dwelling burrow (domichnion) (Fatka et al., 2011). The exact zoological affinities of the tracemakers that produced Arachnostega and their paleoecology are not yet fully understood (Vinn et al., 2018).
The aims of the present note are threefold; firstly, to describe bioerosion of early Permian lingulide shells; secondly, to describe predatory borings in early Permian lingulides and, thirdly, to document the ichnogenus Arachnostega from Lower Permian strata in Brazil for the first time.
Lingulide shells were collected from the Campáleo outcrop,
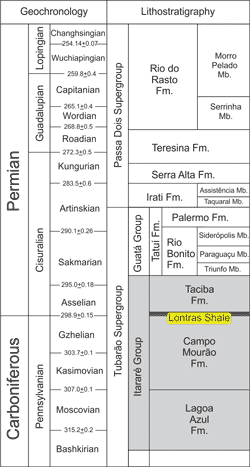
which is located in the city of Mafra in the State of Santa Catarina, Brazil (Fig. 1 ![]() ). The outcrop is positioned in the Lontras Shale
(Fig. 2
). The outcrop is positioned in the Lontras Shale
(Fig. 2 ![]() ), upper part of
Campo Mourão Formation and eastern border of Paraná Basin. The
Campáleo outcrop (UTM 22J 0.618.473, 7.106.243) comprises the first 10 m of the
Lontras Member (Mouro et al., 2017). At the base of outcrop there is a 1-m-thick silty bioturbated argillite
bed (i.e., the Glossifungites suite) and is overlain by a fossiliferous,
1.10-m-thick black shale layer (i.e., the Lontras Shale), followed by a
non-fossiliferous, 7-m-thick shelly rhythmite layer (Mouro et al.,
2017). Lontras Shale can be informally divided into four
levels, including sublevels, distinguished on the bases of rock features, fossil
content, and pyrite concentration (Mouro et al.,
2016, 2017).
Based on the quantity and preservation of the fossils that have been recovered
in the Lontras Shale and are exposed at the Campáleo outcrop, this site has
been identified as an important lower Permian (Cisuralian, 299 myr to 275 myr)
Fossil-Lagerstätte
(Ricetti et al., 2016; Mouro
et al., 2016, 2017).
Lontras Shale is interpreted as a deglaciation phase. The paleoenvironment is
interpreted as a restricted marine area, very close to the coast (Ricetti
et al., 2016; Mouro et
al., 2016, 2017).
), upper part of
Campo Mourão Formation and eastern border of Paraná Basin. The
Campáleo outcrop (UTM 22J 0.618.473, 7.106.243) comprises the first 10 m of the
Lontras Member (Mouro et al., 2017). At the base of outcrop there is a 1-m-thick silty bioturbated argillite
bed (i.e., the Glossifungites suite) and is overlain by a fossiliferous,
1.10-m-thick black shale layer (i.e., the Lontras Shale), followed by a
non-fossiliferous, 7-m-thick shelly rhythmite layer (Mouro et al.,
2017). Lontras Shale can be informally divided into four
levels, including sublevels, distinguished on the bases of rock features, fossil
content, and pyrite concentration (Mouro et al.,
2016, 2017).
Based on the quantity and preservation of the fossils that have been recovered
in the Lontras Shale and are exposed at the Campáleo outcrop, this site has
been identified as an important lower Permian (Cisuralian, 299 myr to 275 myr)
Fossil-Lagerstätte
(Ricetti et al., 2016; Mouro
et al., 2016, 2017).
Lontras Shale is interpreted as a deglaciation phase. The paleoenvironment is
interpreted as a restricted marine area, very close to the coast (Ricetti
et al., 2016; Mouro et
al., 2016, 2017).
Click on thumbnail to enlarge the image. |
Figure 1: Locality map
(modified after Ricetti et al., 2016). |
|
Figure
2:
Stratigraphic position of Lontras Shale in the Lower Permian of Paraná Basin
(modified after Ricetti et
al., 2016). |
A small collection of 58 lingulide shells was searched for associated ichnofossils. Shells with signs of bioerosion, predatory borings and cryptic burrows were selected for photographing. Photographing was carried out using a Zeiss digital camera. Dimensions of ichnofossils were measured from calibrated photos.
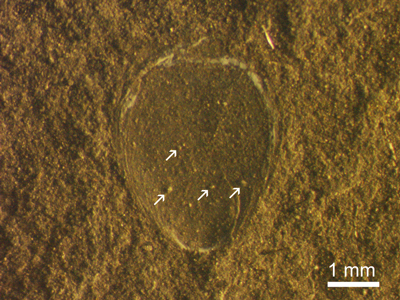
About 24% of 58 lingulides studied
showed signs of bioerosion in form of multiple small shallow pits
(Fig. 3 ![]() ). This
type of pits resemble incomplete Oichnus
paraboloides borings. Their diameters of aperture have a range from 0.03 mm
to 0.17 mm. The diameter of pits in a single lingulid valve is largely variable.
Also, some shells are completely covered with small pits; borings do not show
preference for special shell regions.
). This
type of pits resemble incomplete Oichnus
paraboloides borings. Their diameters of aperture have a range from 0.03 mm
to 0.17 mm. The diameter of pits in a single lingulid valve is largely variable.
Also, some shells are completely covered with small pits; borings do not show
preference for special shell regions.
|
Figure
3: Small rounded pits in the ligulid shell (arrows) indicating the
bioerosion. |
A
single lingulide valve shows a centrally located large circular Oichnus
simplex boring with 0.67 mm wide
(Fig. 4 ![]() ). Same valve also exhibits some
traces of bioerosion.
). Same valve also exhibits some
traces of bioerosion.
|
Figure
4:
Lingulide valve shows a centrally located large circular Oichnus
simplex boring (Oich.). |
Almost
21% of the lingulide shells show small Arachnostega
traces in their interior
(Fig. 5 ![]() ). Arachnostega
traces are visible through somewhat translucent shell material as a colour
contrast. The diameter of burrows is relatively constant and ranges from 0.08 mm
to 0.09 mm. These traces are more similar to juvenile stages of Arachnostega gastrochaena
and do not form well-developed meshwork of tunnels. The Arachnostega burrows are filled with lighter colored sediment than
the dark grey rock matrix.
). Arachnostega
traces are visible through somewhat translucent shell material as a colour
contrast. The diameter of burrows is relatively constant and ranges from 0.08 mm
to 0.09 mm. These traces are more similar to juvenile stages of Arachnostega gastrochaena
and do not form well-developed meshwork of tunnels. The Arachnostega burrows are filled with lighter colored sediment than
the dark grey rock matrix.
|
Figure
5:
Small Arachnostega (Ar.)
traces gleaming from the interior of lingulide shells. |
Bioerosion
Dead or/and living lingulide shells were encrusted by phosphate boring organisms that needed a domicile. Considering the shallow depth of the pits in lingulides, it is likely that the soft body of organisms responsible for pit formation were only partially located within their borings. Thus, the function of these pits may have been rather enabling encrusters stronger contact with substrate than providing a shelter against predators for a true endobiont. It is interesting to notice that bioerosion in phosphatic substrates is rare when compared to calcareous ones (Vinn & Toom, 2016), which partially can be explained by study bias, as most of skeletons of marine organisms are calcareous. The earliest boring in phosphatic substrates are known from the early Cambrian (Vinn & Toom, 2016). The domiciles bored by worm-like organisms into calcareous brachiopod shells are common since the Middle Ordovician (Vinn, 2004, 2005). They have usually been classified as Trypanites or Vermiforichnus (Vinn, 2005). The latter two are much deeper than shallow pits in studied Permian lingulides and were created by worms that could fully retract to their borings. Thus, in calcareous brachiopods borings were usually created by endobiotic organisms, while in studied lingulides they were made by epibiotic organism. The variable diameter of pits in lingulides indicates that they were made by several generations of encrusters and were not a result of a single colonization event by multiple larvae of boring epibionts. Alternatively, different diameters of borings could result from growing organism which over time produce traces of larger diameter. The biological affinities of organisms responsible for creating the shallow pits in lingulides will remain unresolved without soft body preservation, but the architecture of these pits would fit with sponges as the makers. It is difficult to estimate bioerosion intensities in phosphatic lingulides, because of lack of other comparable records, but most of specimens with small pits show moderate bioerosion intensities.
Predation
Most likely large circular Oichnus simplex boring was made by some shell drilling predator, probably by a predatory gastropod, though the boring is slightly too small for common gastropod borings (Zonneveld & Gingras, 2014). The central location of the boring in a lingulide valve is well consistent with the predatory nature of the boring. The boring intensity was relatively low in the studied lingulide community as only one shell with O. simplex was found among 58 lingulide specimens. There are no reports on the boring frequency in Permian lingulides (Huntley & Kowalewski, 2007), but some data on boring frequencies on articulate brachiopods are available for the Carboniferous (see Harper, 2003, and Klompmaker et al., 2019, for review) and Permian (Hoffmeister et al., 2004). Carboniferous articulate brachiopods show boring frequencies ranging from 3.3 to 17.1 % (Harper, 2003), which is much higher than boring frequency of studied Permian lingulides. Therefore, it is very likely that lingulides were not among favorite prey groups in the early Permian of Brazil. In contrast, some Cambrian lingulides exhibit fairly similar boring intensities to the studied lingulides, being <2% of the population (Harper, 2003). The relatively low boring frequency of studied lingulides may reflect also their infaunal life mode, and their moderate bioerosion intensities could result from post mortem bioerosion.
Cryptic organisms
The earliest examples of Arachnostega known to date are of Cambrian age (e.g., Fatka et al., 2011), but the ichnogenus did not achieve global distribution and become common until the Ordovician (e.g., Aceñolaza et al., 2003).This is the first record of Arachnostega from the interior of lingulide shells. Dead lingulide shells offered shelter for cryptic organisms that left behind Arachnostega traces. Arachnostega traces were probably made by small polychaetes that inhabited soft mud filling of the dead lingulide shell. It is interesting to note that even very small lingulide shells were suitable substrates cryptic organisms such as Arachnostega trace makers. However, the Arachnostega trace makers in Permian of Brazil must have been much smaller than their representatives in the Ordovician of Baltica (Vinn et al., 2014) or Miocene of Egypt (Vinn et al., 2018).
Financial support to OV was provided by the Estonian Research Council, project IUT20-34. CZ thanks Fapesp for financial support, project 2017/10956-5. The authors are grateful to the reviewers for their constructive comments on the manuscript and to Mark A. Wilson for linguistic corrections.
Aceñolaza G., Gutiérrez-Marco J.C. & Peralta S. (2003).- Arachnostega gastrochaenae Bertling (traza fósil) en las secuencias volcaniclásticas de la Formación Suri, Sistema de Famatina, Argentina.- Ameghiniana, Buenos Aires, vol. 40, p. 315-322.
Bengtson S. & Zhao Y. (1992).- Predatorial borings in late Precambrian mineralized exoskeletons.- Science, Washington, vol. 257, p. 367-369.
Bertling M. (1992).- Arachnostega n. ichnog. - burrowing traces in internal moulds of boring bivalves (Late Jurassic, Northern Germany).- Paläontologische Zeitschrift, Heidelberg, vol. 66, p. 177-185.
Brett C.E. (2003).- Durophagous predation in Paleozoic marine benthic assemblages. In: Kelley P., Kowalewski M. & Hansen T. (eds.), Predator prey interactions in the fossil record.- Kluwer Academic-Plenum Publishers, Lawrence (KS), p. 401-432.
Brett D.E. & Walker S.E. (2002).- Predators and predation in Paleozoic marine environments.- Paleontological Society Papers, Tulsa, vol. 8, p. 93-118.
Fatka O., Mikulá R., Szabad M., Micka V. & Valent M. (2011).- Arachnostega Bertling, 1992 in the Drumian (Cambrian) sediments of the Teplá-Barrandian region (Czech Republic).- Acta Geologica Polonica, Warsaw, vol. 61, p. 367-381.
Harper E.M. (2003).- Assessing the importance of drilling predation over the Palaeozoic and Mesozoic.- Palæogeography, Palæoclimatology, Palæoecology, vol. 210, p. 185-198.
Hoffmeister A.P., Kowalewski M., Baumiller T.K. & Bambach R.K. (2004).- Drilling predation on Permian brachiopods and bivalves from the Glass Mountains, west Texas.- Acta Palaeontologica Polonica, Warsaw, vol. 49, p. 443-454.
Holmer L.E. & Popov L.E. (2000).- Class Lingulata. In: Kaesler R.L. (ed.), Treatise on Invertebrate Paleontology. Part H. Revised Brachiopoda. Geological Society of America, Boulder, and University of Kansas, Lawrence, Vol. 2, pp. 30-146.
Huntley J.W. & Kowalewski M. (2007).- Strong coupling of predation intensity and diversity in the Phanerozoic fossil record.- Proceedings of the National Academy of Sciences, Washington, vol. 104, p. 15006-15010.
Klompmaker A.A., Kelley P.H., Chattopadhyay D., Clements J.C., Huntley J.W. & Kowalewski, M. (2019).- Predation in the marine fossil record: Studies, data, recognition, environmental factors, and behavior.- Earth-Science Reviews, vol. 194, p. 472-520.
Mouro L.D., Rakociński M., Marynowski L., Pisarzowska A., Musabelliu S., Zatoń M., Carvalho M.A., Fernandes A.C.S. & Waichel B.L. (2017).- Benthic anoxia, intermittent photic zone euxinia and elevated productivity during deposition of the Lower Permian, post-glacial fossiliferous black shales of the Paraná Basin, Brazil.- Global and Planetary Change, vol. 158, p. 155-172.
Mouro L.D., Zatoń M., Fernandes A.C.S. & Waichel B.L. (2016).- Larval cases of caddisfly (Insecta: Trichoptera) affinity in Early Permian marine environments of Gondwana.- Scientific Reports, London, vol. 6, article 19215, 7 p.
Ricetti J.H.Z., Schneider J.W., Iannuzzi R. & Weinschütz L.C. (2016).- Anthracoblattina Mendesi Pinto et Sedor (Blattodea, Phyloblattidae): The most completely preserved South American Palaeozoic cockroach.- Revista Brasileira de Paleontologia, Rio de Janeiro, vol. 19, p. 181-194.
Vermeij G.J. (1977).- The Mesozoic marine revolution: evidence from snails, predators and grazers.- Paleobiology, New York, vol. 3, p. 245-258.
Vermeij G.J. (1987).- Evolution and escalation: An ecological history of life.- Princeton University Press, Princeton (NJ), 527 p.
Vermeij G.J., Schindel D.E. & Zipser E. (1981).- Predation through geological time: evidence from gastropod shell repair.- Science, Washington, vol. 214, p. 1024-1026.
Vinn O. (2004).- The earliest known Trypanites borings in the shells of articulate brachiopods from the Arenig (Ordovician) of Baltica.- Proceedings of the Estonian Academy of Sciences Geology, Tallinn, vol. 53, p. 257-266.
Vinn O. (2005).- The distribution of worm borings in brachiopod shells from the Caradoc Oil Shale of Estonia.- Carnets Geol., Madrid, vol. 5, no. A03 (CG2005_A03), p. 1-11.
Vinn O. & Toom U. (2016).- Borings in phosphatized siltstone pebbles from the Cambrian of Estonia (Baltica).- Geological Magazine, New York, vol. 153, p. 635-642.
Vinn O., El-Sabbagh A.M., El-Hedeny M.M., Rashwan M.A. & Al Farraj S.A. (2018).- A new record and a new ichnospecies of Arachnostega from the Middle Miocene of Egypt.- Proceedings of the Geologists' Association, Oxford, vol. 129, p. 790-796.
Vinn O., Wilson M.A., Zatoń M. & Toom U. (2014).- The trace fossil Arachnostega in the Ordovician of Estonia (Baltica).- Palaeontologia Electronica, Amherst, vol. 17, no. 3, article 17.3.41A, 9 p.
Zhang X.-G., Hou X.-G. & Emig C.C. (2003).- Evidence of lophophore diversity in Early Cambrian Brachiopoda.- Proceedings of the Royal Society (Biological Sciences), London, vol. 270 (Suppl.), p. S65-S68.
Zonneveld J.-P. & Gingras M.K. (2014).- Sedilichnus, Oichnus, Fossichnus and Tremichnus: 'Small round holes in shells' re-visited.- Journal of Paleontology, Tulsa, vol. 88, p. 895-905.