◄ Carnets Geol. 20 (2) ►
![]()
Outline:
[1. Introduction]
[2. Geological and paleontological framework]
[3. Systematic paleontology]
[4. Discussion]
and ... [Bibliographic references]
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Gruppo Avis Mineralogia e Paleontologia Scandicci, p.za Vittorio Veneto 1, Badia a
Settimo, 50018 Scandicci (Italy);
Studio Tecnico Geologia e Paleontologia, via F.lli Rosselli 4, S.
Casciano Val di Pesa, 50026 Firenze (Italy)
Geotecnica Ricci, via Arenazze 6/8, 66100 Chieti (Italy)
Dipartimento di Scienze della Terra, Universitŕ di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Published online in final form (pdf) on February 22, 2020
DOI 10.4267/2042/70716
![]()
[Editor: Bruno Granier;
language editors: Stephen Carey and Bruno Ferré]
![]()
The extinct family Squalodontidae consists of heterodont, medium-sized odontocetes, featuring a long rostrum that houses large, procumbent incisors and heavily ornamented postcanine teeth carrying accessory denticles, hence their vernacular name, "shark-toothed dolphins". These longirostrine toothed whales are often seen as bridging the anatomical gap between archaic Oligocene odontocetes and their late Miocene to Holocene relatives. Possibly among the major marine predators of their time, the shark-toothed dolphins are important components of several lower Miocene marine-mammal assemblages from the North Atlantic and Mediterranean/Paratethysian realms. In the present work, a partial skull of Squalodontidae is described from the strata of the Bolognano Formation cropping out in the northeastern sector of the Montagna della Majella massif (Abruzzo, central Italy), which has previously yielded a rich lower Miocene marine-vertebrate assemblage, including eleven taxa of elasmobranchs as well as subordinate teleosts and very fragmentary remains of marine reptiles and mammals. The specimen consists of the anterodorsal portion of a rostrum, preserving parts of both premaxillae and left maxilla, and the anteriormost seven upper left teeth. This partial skull is here identified as belonging to the genus Squalodon, whose presence in the Montagna della Majella vertebrate assemblage had already been tentatively proposed on the basis of two fragmentary teeth. The paleontological significance of this find is discussed in the broader framework of the Euromediterranean record of Squalodon.
• Burdigalian;
• lower Miocene;
• Mediterranean Basin;
• Neoceti;
• paleobiogeography;
• paleoecology;
• Squalodontidae;
• toothed whales
Collareta A., Di Cencio A., Ricci R. & Bianucci G. (2020).- The shark-toothed dolphin Squalodon (Cetacea: Odontoceti) from the remarkable Montagna della Majella marine vertebrate assemblage (Bolognano Formation, central Italy).- Carnets Geol., Madrid, vol. 20, no. 2, p. 19-28.
Le dauphin ŕ dents de requin Squalodon (Cetacea: Odontoceti) du remarquable assemblage de vertébrés marins de Montagna della Majella (Formation de Bolognano, Italie centrale).- La famille éteinte des Squalodontidae comprend des odontocčtes de taille moyenne, hétérodontes, présentant un long rostre qui abrite de grandes dents incisives procombantes et de post-canines trčs ornées comportant des denticules accessoires, d'oů leur nom vernaculaire "dauphins ŕ dents de requin". Ces odontocčtes longirostres sont souvent perçus comme des formes intermédiaires comblant le fossé anatomique entre les odontocčtes oligocčnes archaďques et leurs parents du Miocčne tardif jusqu'ŕ l'Holocčne. Probablement parmi les principaux prédateurs marins de leur époque, les dauphins ŕ dents de requin sont d’importants éléments au sein de plusieurs assemblages de mammifčres marins du Miocčne inférieur des domaines nord-atlantique et méditerranéen/para-téthysien. Dans le présent travail, un crâne partiel de Squalodontidae est décrit dans les couches de la Formation de Bolognano affleurant dans le secteur nord-est du massif de Montagna della Majella (Abruzzes, Italie centrale), qui a livré par le passé un riche assemblage de vertébrés marins du Miocčne inférieur, comprenant onze taxons d'élasmobranches, ainsi que des téléostéens moins nombreux et des restes trčs fragmentaires de reptiles et de mammifčres marins. Ce spécimen comprend la partie antéro-dorsale du rostre, comportant des parties des deux prémaxillaires et du maxillaire gauche, et les sept dents supérieures gauches les plus antérieures. Ce crâne partiel est identifié ici comme appartenant au genre Squalodon, dont la présence dans l'assemblage de vertébrés de Montagna della Majella avait déjŕ été proposée provisoirement sur la base de deux dents fragmentaires. La signification paléontologique de cette découverte est discutée dans le contexte plus large du registre euro-méditerranéen du genre Squalodon.
• Burdigalien ;
• Miocčne inférieur ;
• bassin méditerranéen ;
• Neoceti ;
• paléobiogéographie ;
• paléoécologie ;
• Squalodontidae ;
• odontocčtes
The extinct family Squalodontidae consists of heterodont, medium-sized odontocetes, featuring a long rostrum that houses large, procumbent incisors and heavily ornamented postcanine teeth provided with accessory denticles, hence their vernacular name "shark-toothed dolphins" (Marx et al., 2016, and references therein). Known from a rather rich fossil record which ranges from the upper Oligocene (Chattian) to the upper middle Miocene (Serravallian), squalodontids are important components of several lower Miocene marine-mammal assemblages from the North Atlantic and Mediterranean/Paratethysian realms (e.g., Dooley, 1998). Possibly among the major marine predators of their time, squalodontids are often seen as bridging the anatomical gap between the archaic Oligocene odontocetes and their late Miocene to Holocene relatives (Godfrey, 2013; Marx et al., 2016). For a long time, these longirostrine toothed whales have been regarded as members of the crown Odontoceti clade, in the superfamily Platanistoidea, which also includes the South Asian river dolphin Platanista gangetica (e.g., Muizon, 1991, 1994; Fordyce, 1994; Fordyce & Muizon, 2001; Tanaka & Fordyce, 2016); now however, many phylogenetic analyses recover squalodontids as late-branching stem odontocetes (e.g., Geisler & Sanders 2003; Tanaka & Fordyce, 2014; Churchill et al., 2016; Bianucci et al., 2018a). The current lack of consensus upon the phylogenetic position of the squalodontids is highlighted by the observation that several recent papers dealing with the affinities of the heterodont long-snouted toothed whales present both the aforementioned solutions as plausible (e.g., Tanaka & Fordyce, 2015a, 2015b; Lambert et al., 2018; Muizon et al., 2018; Viglino et al., 2018).
The type genus of Squalodontidae, i.e., Squalodon, has been identified from several lower–middle Miocene deposits of the North Atlantic (including the southern North Sea), the Mediterranean (including some Italian sites), and the Paratethys (e.g., Bianucci & Landini, 2002; Dooley, 2003; Symeonidis et al., 2004; Marsili et al., 2007). In the remarkable lower Miocene marine-vertebrate assemblage of the Bolognano Formation (Montagna della Majella massif, central Apennines, central Italy), Squalodon was tentatively identified on the basis of two isolated partial teeth whose preservation state fails to permit a definitive and unambiguous genus-level assignment (Marsili et al., 2007). In the present paper, we report on a partial squalodontid rostrum, exhibiting seven articulated teeth, from the Bolognano strata exposed close to the village of Pretoro, in the northeastern sector of the massif. This fossil specimen is here referred to Squalodon sp. and its paleontological significance is briefly discussed in the broader framework of the Euromediterranean record of this genus.
The
Montagna della Majella, from which the fossil specimen described here comes, is
a wide massif located in the central Apennines (Abruzzo region, central Italy).
The massif (Fig. 1 ![]() ) is largely underlain by a thick package of upper Jurassic to
upper Miocene carbonates developed on the northern margin of the Apulian
platform and forming an east-verging, box-shaped anticline. From the Santonian,
the Mesozoic–Cenozoic succession of the Montagna della Majella testifies to
open-shelf and carbonate-ramp depositional environments (Bally,
1954; Crescenti
et al., 1969; Catenacci,
1974; Accarie, 1988; Vecsei
et al., 1998; Ghisetti & Vezzani,
2002; Scisciani et al., 2002; Rustichelli
et al., 2012, 2013; Brandano
et al., 2013). The
youngest lithostratigraphic unit in the succession is the upper Oligocene to
upper Miocene Bolognano Formation, a sedimentary wedge composed of three stacked
warm-water carbonate-ramp depositional systems (Mutti et al.,
1997; Vecsei & Sanders,
1999; Danese, 1999; Marsili et al.,
2007). The
middle portion of the Bolognano Formation, which has been referred to the lower
Burdigalian SBZ 25 zone of Cahuzac and Poignant
(1997), has
yielded a rich marine-vertebrate assemblage, including eleven taxa of
elasmobranchs (Carcharias acutissima, Carcharias cuspidata, Carcharocles
chubutensis, Cosmopolitodus hastalis, Isurus oxyrinchus, "Isurus"
sp., Hemipristis serra, Galeocerdo aduncus, Negaprion cf.
eurybathrodon, Parotodus benedeni, and Physogaleus contortus),
eight taxa of teleosts (Chilomycterus sp., Diplodus sp. and two
indeterminate sparids, Diodon sp., Labrodon sp., Sphyraena
sp., and Trigonodon jugleri), an indeterminate crocodile, and two taxa of
odontocetes (an indeterminate kentriodontid and a squalodontid-like taxon
tentatively identified as Squalodon). On the whole, this fossil
vertebrate assemblage depicts a warm-water, food-rich paleoenvironment located
close to the coast (Marsili et al., 2007);
moreover, its taxonomic composition (including an elasmobranch taxon, C.
chubutensis, which is only known from lower Miocene deposits; Carrillo-Briceńo
et al., 2019) supports the estimated
Burdigalian age for the middle portion of the Bolognano Formation.
) is largely underlain by a thick package of upper Jurassic to
upper Miocene carbonates developed on the northern margin of the Apulian
platform and forming an east-verging, box-shaped anticline. From the Santonian,
the Mesozoic–Cenozoic succession of the Montagna della Majella testifies to
open-shelf and carbonate-ramp depositional environments (Bally,
1954; Crescenti
et al., 1969; Catenacci,
1974; Accarie, 1988; Vecsei
et al., 1998; Ghisetti & Vezzani,
2002; Scisciani et al., 2002; Rustichelli
et al., 2012, 2013; Brandano
et al., 2013). The
youngest lithostratigraphic unit in the succession is the upper Oligocene to
upper Miocene Bolognano Formation, a sedimentary wedge composed of three stacked
warm-water carbonate-ramp depositional systems (Mutti et al.,
1997; Vecsei & Sanders,
1999; Danese, 1999; Marsili et al.,
2007). The
middle portion of the Bolognano Formation, which has been referred to the lower
Burdigalian SBZ 25 zone of Cahuzac and Poignant
(1997), has
yielded a rich marine-vertebrate assemblage, including eleven taxa of
elasmobranchs (Carcharias acutissima, Carcharias cuspidata, Carcharocles
chubutensis, Cosmopolitodus hastalis, Isurus oxyrinchus, "Isurus"
sp., Hemipristis serra, Galeocerdo aduncus, Negaprion cf.
eurybathrodon, Parotodus benedeni, and Physogaleus contortus),
eight taxa of teleosts (Chilomycterus sp., Diplodus sp. and two
indeterminate sparids, Diodon sp., Labrodon sp., Sphyraena
sp., and Trigonodon jugleri), an indeterminate crocodile, and two taxa of
odontocetes (an indeterminate kentriodontid and a squalodontid-like taxon
tentatively identified as Squalodon). On the whole, this fossil
vertebrate assemblage depicts a warm-water, food-rich paleoenvironment located
close to the coast (Marsili et al., 2007);
moreover, its taxonomic composition (including an elasmobranch taxon, C.
chubutensis, which is only known from lower Miocene deposits; Carrillo-Briceńo
et al., 2019) supports the estimated
Burdigalian age for the middle portion of the Bolognano Formation.
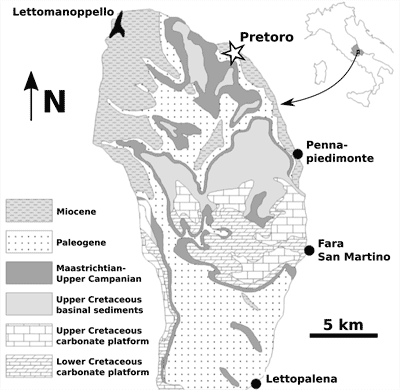
The
cetacean fossil described here comes from the Bolognano strata exposed at a
quarry close to the village of Pretoro (approximate geographic coordinates: 42°13'N,
14°08'E), in the northeastern portion of the Montagna della Majella (Fig. 1 ![]() ). The specimen was collected several decades ago by a local quarryman and its
exact stratigraphic position is unknown. Nevertheless, the fossiliferous lower
Miocene beds of the Bolognano Formation are well exposed in the northeastern
portion of the Montagna della Majella, southeast of Lettomanoppello (Marsili
et al., 2007).
Considering that all other discoveries of marine vertebrates (including
odontocete remains) from this area of the massif come from the middle portion of
the Bolognano Formation, an early Miocene age can be confidently proposed for
the fossil specimen described in the present paper.
). The specimen was collected several decades ago by a local quarryman and its
exact stratigraphic position is unknown. Nevertheless, the fossiliferous lower
Miocene beds of the Bolognano Formation are well exposed in the northeastern
portion of the Montagna della Majella, southeast of Lettomanoppello (Marsili
et al., 2007).
Considering that all other discoveries of marine vertebrates (including
odontocete remains) from this area of the massif come from the middle portion of
the Bolognano Formation, an early Miocene age can be confidently proposed for
the fossil specimen described in the present paper.
|
Figure 1:
Location of the
site where the fossil cetacean specimen MSNUP I-16944 was found (Pretoro, black
star) and schematic regional geological map. Modified after Marsili et
al. (2007). |
CETACEA Brisson, 1762
PELAGICETI Uhen, 2008
NEOCETI Fordyce & Muizon, 2001
ODONTOCETI Flower, 1867
SQUALODONTIDAE Brandt, 1873
Squalodon Grateloup, 1840
Squalodon sp.
Material and repository. An incomplete rostrum, featuring seven articulated teeth, enclosed in a limestone block kept at Museo di Storia Naturale dell'Universitŕ di Pisa (=MSNUP) with accession number MSNUP I-16944.
Occurrence. MSNUP I-16944 was collected at a
quarry close to the village of Pretoro (approximate geographic coordinates:
42°13'N, 14°08'E), in the northeastern portion of the Montagna della Majella
massif (central Apennines, central Italy), where the lower Miocene strata of the
Oligo-Miocene Bolognano Formation are exposed (Fig. 1 ![]() ).
).
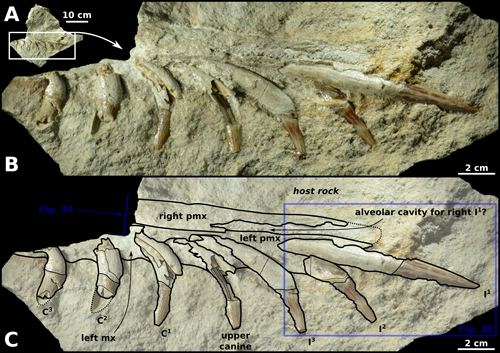
Description, comparisons, and remarks. MSNUP I-16944 consists of the anterodorsal portion of
the rostrum of an odontocete, preserving parts of both premaxillae and left
maxilla, as well as the anteriormost seven upper left teeth. This fragment of
rostrum is about 20 cm long (not including the length of the first incisor) and
is preserved within a hard limestone block that could not be prepared
mechanically without damaging the cetacean fossil; as such, only the right
ventrolateral aspect (Fig.
2 ![]() )
and a transverse section (Fig. 3.A
)
and a transverse section (Fig. 3.A ![]() )
of the rostrum
can be observed and described. In the exposed right ventrolateral view, the
premaxillae and right maxilla appear as strongly abraded. The lateral margins of
the preserved anterior portion of the rostrum are almost straight and roughly
parallel to each other, recalling the condition observed in various early
Neogene groups of heterodont odontocetes, including Squalodontidae. An
anteroposteriorly elongated opening that occurs at the anterior termination of
the left premaxilla is here interpreted as representing the alveolar cavity of
the missing first upper right incisor (Fig.
2
)
of the rostrum
can be observed and described. In the exposed right ventrolateral view, the
premaxillae and right maxilla appear as strongly abraded. The lateral margins of
the preserved anterior portion of the rostrum are almost straight and roughly
parallel to each other, recalling the condition observed in various early
Neogene groups of heterodont odontocetes, including Squalodontidae. An
anteroposteriorly elongated opening that occurs at the anterior termination of
the left premaxilla is here interpreted as representing the alveolar cavity of
the missing first upper right incisor (Fig.
2 ![]() ). At the level of the posterior
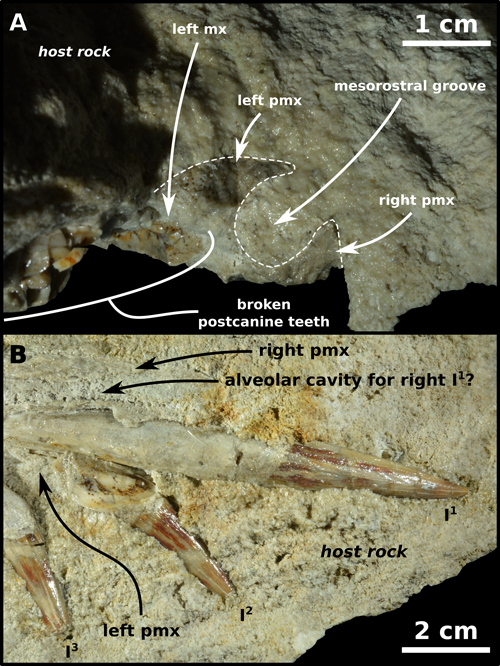
preserved end of the rostrum, the transversely sectioned mesorostral groove is
roughly oval (i.e., slightly higher than wide) and open upwards (Fig.
3.A
). At the level of the posterior
preserved end of the rostrum, the transversely sectioned mesorostral groove is
roughly oval (i.e., slightly higher than wide) and open upwards (Fig.
3.A ![]() ).
).
The
seven anteriormost upper left teeth are preserved in situ, i.e.,
partly embedded in the left premaxilla and maxilla, the labial face of these
teeth being hidden by the hard host rock (Fig. 2 ![]() ). The preserved teeth are
the three incisors (I1 to I3), the canine, and the three
anteriormost postcanines (i.e., the cheek teeth C1 to C3).
All of these teeth are single-rooted and have similar (i.e., roughly
conical) crown morphologies. Mesial/distal carinae (i.e., keels) appear
to be present at least on the crowns of I1, C2, and C3,
providing them with slightly ogival transverse sections. The crown of the
incisors is considerably shorter than the root and strongly procumbent. The
anteriormost incisor (I1) is almost horizontal, and is dorsal to I2;
in turn, I2 and I3 are moderately curved and have a more
ventrolateral orientation (Figs.
2
). The preserved teeth are
the three incisors (I1 to I3), the canine, and the three
anteriormost postcanines (i.e., the cheek teeth C1 to C3).
All of these teeth are single-rooted and have similar (i.e., roughly
conical) crown morphologies. Mesial/distal carinae (i.e., keels) appear
to be present at least on the crowns of I1, C2, and C3,
providing them with slightly ogival transverse sections. The crown of the
incisors is considerably shorter than the root and strongly procumbent. The
anteriormost incisor (I1) is almost horizontal, and is dorsal to I2;
in turn, I2 and I3 are moderately curved and have a more
ventrolateral orientation (Figs.
2 ![]() ,
3.B
,
3.B ![]() ). The roots of I2 and I3
exhibit distinct bulges close to the root-crown passage. The canine is badly
damaged but seems to be more strongly curved than the adjoining incisor, with a
proportionally shorter root. The morphology of
the three preserved anteriormost postcanine teeth recalls that of the canine,
from which they mainly differ by their stronger curvature and shorter crowns
that are more posteriorly oriented. No accessory denticles could be observed
along the crowns of C1, C2, and C3. Enamel
ornamentation is present on the crown of all the preserved teeth but
particularly evident on C2 and C3; it consists of
delicate, slightly undulatory, closely spaced apicobasal ridges (Figs.
2
). The roots of I2 and I3
exhibit distinct bulges close to the root-crown passage. The canine is badly
damaged but seems to be more strongly curved than the adjoining incisor, with a
proportionally shorter root. The morphology of
the three preserved anteriormost postcanine teeth recalls that of the canine,
from which they mainly differ by their stronger curvature and shorter crowns
that are more posteriorly oriented. No accessory denticles could be observed
along the crowns of C1, C2, and C3. Enamel
ornamentation is present on the crown of all the preserved teeth but
particularly evident on C2 and C3; it consists of
delicate, slightly undulatory, closely spaced apicobasal ridges (Figs.
2 ![]() ,
3.B
,
3.B ![]() ).
Dental wear is ubiquitous in form of apical wear facets, here interpreted as
attritional.
).
Dental wear is ubiquitous in form of apical wear facets, here interpreted as
attritional.
|
Figure
2:
MSNUP
I-16944, partial skull of Squalodon sp., enclosed in a limestone block
from the Bolognano Formation of central Italy. A) General view of the
fossil-bearing block. B) Incomplete rostrum in right-ventrolateral view and C)
corresponding line drawing (note that the teeth are displayed in lingual view). |
|
Figure
3:
MSNUP I-16944, partial skull of Squalodon sp.,
enclosed in a limestone block from the Bolognano Formation of central Italy. A)
Detail of the transverse section of the rostrum at the level of the root of the
first upper postcanine (C1) left tooth; note the dorsally open
mesorostral groove (the dashed white line denotes the extent of the bone). B)
Detail of the upper left incisors in lingual view; note the root of I1
situated dorsal to that of I2, the strong protrusion of the teeth,
and the gentle longitudinal fluting of the tooth crowns. |
MSNUP
I-16944 is attributed to Squalodontidae owing to the observation of large,
procumbent incisors, whose crowns display gentle longitudinal fluting of the
dental enamel, with the root of I1 being placed dorsal to that of I2
(e.g., Dooley, 1998; Marx
et al., 2016; Muizon
et al., 2018; Viglino
et al., 2018).
Four genera of Squalodontidae are currently represented by diagnostic material: Squalodon,
Eosqualodon, Phoberodon, and Kelloggia (a possible junior
synonym of Squalodon; Fordyce and Muizon,
2001) (Marx et al., 2016; Muizon
et al., 2018);
recently, however, Phoberodon has been recognized as a stem odontocete or
an early-diverging platanistoid not closely related to the squalodontids (Viglino
et al., 2018).
Whereas the mesorostral groove of MSNUP I-16944 is widely open dorsally at the
level of the root of the first upper postcanine left tooth, the mesorostral
groove of the European form Eosqualodon is almost completely roofed by
the medial portion of the premaxillae from the apex of the rostrum to a much
more posterior level (personal observation on Eosqualodon langewieschei).
Eosqualodon further differs from MSNUP I-16944 in its pattern of dental
ornamentation, which consists of stronger and less closely spaced ridges and
sulci. MSNUP I-16944 also clearly differs from the South American form Phoberodon,
in which the third upper incisor is curved lingually (with the apex pointing
medially) and the upper canine and anteriormost cheek teeth exhibit widely
separated enamel striations (Viglino
et al., 2018). By
contrast, MSNUP I-16944 closely matches the morphology of Squalodon,
which is known from several specimens of the Euromediterranean region (Fig.
4 ![]() ).
).
|
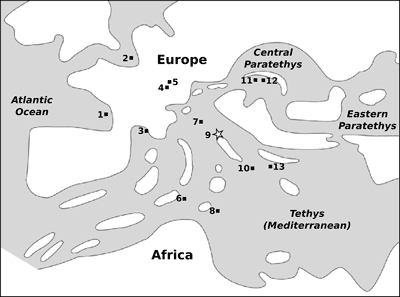
Figure
4:
Occurrences of the genus Squalodon
from lower Miocene deposits of Europe (note that attribution to the early
Miocene of the Apulian remains is tentative). 1- Léognan, Bordeaux,
southwestern France; 2- Antwerp, northern Belgium; 3- Bari, Saint-Paul-Trois-Châteaux, southern France; 4- Switzerland (various
localities); 5- Baltringen, Baden-Württemberg, southwestern Germany; 6-
Ragusa, Sicily, southern Italy; 7- Libano and Belluno, Veneto, northern
Italy; 8- Malta; 9- Montagna della Majella, Abruzzo, central Italy; 10-
Cursi-Melpignano, Apulia, southern Italy; 11- Horné Strháre, Banská
Bystrica, southern Slovakia; 12- Mucín, Banská Bystrica, southern Slovakia;
13- Tsotilion, western Macedonia, Greece. Data after Bianucci & Landini
(2002), Symeonidis et al.
(2004), and the present study. The
paleogeographic reconstruction of the Euromediterranean region in early Miocene
times is from Rögl (1998), modified after Symeonidis et al. (2004). The southwestern German and
Swiss records are interpreted as reflecting temporary marine transgression over
these western Paratethysian sectors. |
Species-level attribution of MSNUP I-16944 is challenged by the fragmentary nature of the available material. For example, the Montagna della Majella specimen cannot be compared with the Italian species Squalodon peregrinus, for which neither the anterior portion of the rostrum nor the teeth are known (Fornasiero & Del Favero, 2014). Moreover, an unambiguous species-level determination of these remains would need a detailed revision of several extinct nominal species currently attributed to the genus Squalodon (e.g., Symeonidis et al., 2004), which is beyond the scope of this paper. However, the following considerations might lend some support to a tentative referral of MSNUP I-16944 to Squalodon bellunensis: 1) no unambiguous dissimilarities in either size or dental morphology distinguish MSNUP I-16944 from specimens of S. bellunensis for which the anteriormost portion of the rostrum and the corresponding upper teeth are preserved, e.g., a specimen kept at Museo di Geologia e Paleontologia dell'Universitŕ di Padova (=MGPUP) under accession number MGPUP 26131 (Table 1); 2) the type material of S. bellunensis comes from a locality that is (and was, in early Miocene times) spatially close to the discovery site of the Montagna della Majella fossil squalodontid; 3) the type horizon of S. bellunensis and the fossiliferous portion of the Bolognano Formation are early Miocene in age, and as such, roughly coeval (in particular, they have been referred to the upper Aquitanian–lower Burdigalian and to the lower Burdigalian, respectively). Nevertheless, given the limited diagnostic value of the preserved skull parts, MSNUP I-16944 is here referred to Squalodon sp.
Table 1: Tooth crown height of the upper incisors of Squalodon sp. (specimen MSNUP I-16944 from the lower Miocene of the Montagna della Majella massif, central Italy) and Squalodon bellunensis (specimen MGPUP 26131 from the lower Miocene of Libano and Belluno, northern Italy). Measurements of MSNUP I-16944 refer to the left tooth row; measurements of MGPUP 26131 refer to the right tooth row. All measurements are expressed in millimeters.
| I3 | I2 | I1 | |
| MSNUP I-16944 | 24 | 33 | 48 |
| MGPUP 26131 | 27 | 30 | 50 |
As sometimes observed in the dental remains of juveniles and young adults of extinct odontocetes whose teeth were progressively worn with age due to abrasion or occlusion during feeding (e.g., the stem beaked whale Messapicetus; Bianucci et al., 2016), apical dental wear is less pronounced in MSNUP I-16944 than in the anteriormost upper teeth of MGPUP 26131 and other full-grown members of Squalodon. This difference in the extent of apical dental wear, coupled with the aforementioned overall similarities in size, might indicate that MSNUP I-16944 is a young adult individual.
The recovery and description of MSNUP I-16944 allows us to confirm the presence of Squalodon in the Montagna della Majella marine vertebrate assemblage, thus supporting previous inferences based on much more fragmentary dental material (Marsili et al., 2007). Moreover, it shows that significant skeletal remains of marine mammals, and not only isolated teeth, might be recovered from the Bolognano Formation. Lending indirect support to this suggestion, Dewaele et al. (2018) have recently proposed that the holotype of the archaic monachine seal Noriphoca gaudini collected in the 19th century from the vicinities of Roccamorice (northwestern sector of the Montagna della Majella massif; Guiscardi, 1870) comes from basal beds of the Bolognano Formation exposed there. These observations encourage the continued search for fossil marine mammals in the Bolognano strata of the Montagna della Majella massif.
In the lower Miocene of the
Mediterranean Basin, Squalodon is a relatively common odontocete genus,
being known from several deposits of France, Italy, Malta and Greece (Fig.
4 ![]() ).
In particular, systematically informative specimens of Squalodon come
from Bari (near Taulignan), Saint-Paul-Trois-Châteaux, southeastern France (Squalodon
bariensis: Gervais, 1853; Muizon,
1988, 1991) and Libano and
Belluno, Veneto,
northern Italy (Squalodon bariensis, Squalodon bellunensis, and Squalodon
peregrinus: Dal Piaz, 1900–1917; Rothausen,
1968; Bianucci et al.,
2018b). In addition to the Venetian
occurrences, in the vicinity of the discovery site of MSNUP I-16944, the genus Squalodon
has been reported from Aquitanian (or Langhian) strata of the Pietra Leccese,
Apulia, southern Italy (as Squalodon sp.: Bianucci et al.,
1994), the lower Miocene of
Ragusa,
Sicily, southern Italy (as Squalodon dalpiazi: Fabiani,
1949), the lower Miocene of Malta (Bianucci
et al., 2012), and the
lower Miocene of Tsotilion, western Macedonia, Greece (as Squalodon cf. bellunensis:
Symeonidis et al., 2004). In combination with the observations from the
literature cited above, our record of a partial skull of Squalodon sp.
from central Italy indicates that the epeiric seas of the central
Mediterranean basin comprised a high-use area for the Squalodon populations
during the first part of the Miocene.
).
In particular, systematically informative specimens of Squalodon come
from Bari (near Taulignan), Saint-Paul-Trois-Châteaux, southeastern France (Squalodon
bariensis: Gervais, 1853; Muizon,
1988, 1991) and Libano and
Belluno, Veneto,
northern Italy (Squalodon bariensis, Squalodon bellunensis, and Squalodon
peregrinus: Dal Piaz, 1900–1917; Rothausen,
1968; Bianucci et al.,
2018b). In addition to the Venetian
occurrences, in the vicinity of the discovery site of MSNUP I-16944, the genus Squalodon
has been reported from Aquitanian (or Langhian) strata of the Pietra Leccese,
Apulia, southern Italy (as Squalodon sp.: Bianucci et al.,
1994), the lower Miocene of
Ragusa,
Sicily, southern Italy (as Squalodon dalpiazi: Fabiani,
1949), the lower Miocene of Malta (Bianucci
et al., 2012), and the
lower Miocene of Tsotilion, western Macedonia, Greece (as Squalodon cf. bellunensis:
Symeonidis et al., 2004). In combination with the observations from the
literature cited above, our record of a partial skull of Squalodon sp.
from central Italy indicates that the epeiric seas of the central
Mediterranean basin comprised a high-use area for the Squalodon populations
during the first part of the Miocene.
The putative closest extant
relative of squalodontids, the South Asian river dolphin, is one of the two (or
three, if the likely extinct baiji, Lipotes vexillifer,
is taken into account) living cetacean species strictly restricted to freshwater
habitats, besides the Amazon river dolphin Inia geoffrensis. The
occurrence of Squalodon sp. reported herein is in a nearshore but truly
marine paleoenvironmental setting, and the same can be said for most of the
other Squalodon localities shown in Figure
4 ![]() . The Libano–Belluno
assemblage, which includes at least three species of Squalodon (Bianucci
et al., 2018b), is found in sandstones that
testify to a delta-front depositional setting (e.g., Cason et
al., 1981; Costa et al.,
1996; Ghibaudo et al., 1996) - an observation that suggests
that Squalodon might have inhabited both shelfal environments and
estuarine (or even riverine) waters. A similar ability to live in both fresh and
marine water is observed in some extant genera of true dolphins (e.g., Sotalia,
the tucuxi and Guiana dolphins, and Orcaella, the Irrawaddy and
Australian snubfin dolphins) and might have characterized other odontocete
lineages in the past (e.g., some platanistids and lipotids; Lambert
et al., 2008; Bianucci
et al., 2013; Boessenecker
& Poust, 2015). Somewhat
akin to these extant odontocetes, squalodontids have been interpreted as nektic
or nektobenthic cetaceans that thrived in fluvial and/or neritic environments,
feeding near the bottom (Landini et al., 2005; Bianucci & Landini,
2002). Interestingly, as reported
above, the genera that are currently assigned to Squalodontidae - Eosqualodon, Squalodon (possibly including the genus Kelloggia)
and, perhaps, Phoberodon - are only known
from the coasts of the Atlantic realm and its northeastern cul-de-sac, the Mediterranean/Paratethysian basins. In spite of their
rich and long fossil record, spanning the late Oligocene and most of the
Miocene, it seems therefore that the squalodontids did not undergo dispersal to
the Indo-Pacific waters, which has instead been recently established for a much
lesser known and more elusive group of heterodont long-snouted odontocetes
such as Inticetidae, an extinct family that is known from a very few discoveries
in the Miocene of Peru, the Atlantic coast of the U.S.A., France and Italy (Peri
et al., 2019, and references therein). If Squalodontidae were indeed
freshwater-tolerant organisms that thrived in river mouths and marginal-marine
settings, a habitat preference for transitional and nearshore environments might
explain the relatively restricted paleobiogeographic distribution of this
otherwise successful and long-ranging family of longirostrine shark-toothed
dolphins.
. The Libano–Belluno
assemblage, which includes at least three species of Squalodon (Bianucci
et al., 2018b), is found in sandstones that
testify to a delta-front depositional setting (e.g., Cason et
al., 1981; Costa et al.,
1996; Ghibaudo et al., 1996) - an observation that suggests
that Squalodon might have inhabited both shelfal environments and
estuarine (or even riverine) waters. A similar ability to live in both fresh and
marine water is observed in some extant genera of true dolphins (e.g., Sotalia,
the tucuxi and Guiana dolphins, and Orcaella, the Irrawaddy and
Australian snubfin dolphins) and might have characterized other odontocete
lineages in the past (e.g., some platanistids and lipotids; Lambert
et al., 2008; Bianucci
et al., 2013; Boessenecker
& Poust, 2015). Somewhat
akin to these extant odontocetes, squalodontids have been interpreted as nektic
or nektobenthic cetaceans that thrived in fluvial and/or neritic environments,
feeding near the bottom (Landini et al., 2005; Bianucci & Landini,
2002). Interestingly, as reported
above, the genera that are currently assigned to Squalodontidae - Eosqualodon, Squalodon (possibly including the genus Kelloggia)
and, perhaps, Phoberodon - are only known
from the coasts of the Atlantic realm and its northeastern cul-de-sac, the Mediterranean/Paratethysian basins. In spite of their
rich and long fossil record, spanning the late Oligocene and most of the
Miocene, it seems therefore that the squalodontids did not undergo dispersal to
the Indo-Pacific waters, which has instead been recently established for a much
lesser known and more elusive group of heterodont long-snouted odontocetes
such as Inticetidae, an extinct family that is known from a very few discoveries
in the Miocene of Peru, the Atlantic coast of the U.S.A., France and Italy (Peri
et al., 2019, and references therein). If Squalodontidae were indeed
freshwater-tolerant organisms that thrived in river mouths and marginal-marine
settings, a habitat preference for transitional and nearshore environments might
explain the relatively restricted paleobiogeographic distribution of this
otherwise successful and long-ranging family of longirostrine shark-toothed
dolphins.
We are grateful to the anonymous quarryman who discovered and collected the partial squalodontid skull MSNUP I-16944. Thanks are also due to Federico Alcini for facilitating the study of the cetacean specimen described here, and to Giorgio Carnevale for fruitful discussions on certain aspects of the Montagna della Majella marine-vertebrate assemblage.
This paper greatly benefited from constructive comments and thoughtful suggestions by Olivier Lambert (Institut Royal des Sciences Naturelles de Belgique) and Christian de Muizon (Muséum National d'Histoire Naturelle) - thank you very much! Thanks are also due to Bruno Granier (Université de Bretagne Occidentale), Stephen Carey (Federation University Australia), and Bruno Ferré (Sotteville-lčs-Rouen) for their most valuable editorial support.
Accarie H. (1988).- Dynamique sédimentaire et structurale au passage plate-forme/bassin. Les facies carbonatés crétacés et tertiaires du Massif de la Maiella (Abruzzes, Italie).- École Normale Supérieure des Mines de Paris, Mémoires des Sciences de la Terre, vol. 5, 162 p.
Bally A.W. (1954).- Geologische Untersuchungen in den SE-Abruzzen.- Ph.D. Thesis, University of Zurich, 289 p.
Bianucci G., Bosio G., Malinverno E., Muizon C. de, Villa I. M., Urbina M. & Lambert O. (2018a).- A new large squalodelphinid (Cetacea, Odontoceti) from Peru sheds light on the Early Miocene platanistoid disparity and ecology.- Royal Society Open Science, vol. 5, no. 4, article #172302, 32 p.
Bianucci G., Collareta A., Bosio G., Landini W., Gariboldi K., Gioncada A., Lambert O., Malinverno E., Muizon C. de, Varas-Malca R., Villa I.M, Coletti G. & Urbina M. (2018b).- Taphonomy and palaeoecology of the lower Miocene marine vertebrate assemblage of Ullujaya (Chilcatay Formation, East Pisco Basin, southern Peru).- Palæogeography, Palæoclimatology, Palæoecology, vol. 511, p. 256-279.
Bianucci G., Collareta A., Post K., Varola A. & Lambert O. (2016).- A new record of Messapicetus from the Pietra leccese (late Miocene, southern Italy): antitropical distribution in a fossil beaked whale (Cetacea, Ziphiidae).- Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy), Milano, vol. 122, p. 63-74.
Bianucci G., Gatt M., Catanzariti R., Sorbi S., Bonavia C.G., Curmi R. & Varola A. (2011).- Systematics, biostratigraphy and evolutionary pattern of the Oligo-Miocene marine mammals from the Maltese Islands.- Geobios, Villeurbanne, vol. 44, p. 549-585.
Bianucci G., Lambert O., Salas-Gismondi R., Tejada J., Pujos F., Urbina M. & Antoine P.-O. (2013).- A Miocene relative of the Ganges river dolphin (Odontoceti, Platanistidae) from the Amazonian Basin.- Journal of Vertebrate Paleontology, vol. 33, p. 741-745.
Bianucci G. & Landini W. (2002).- Change in diversity, ecological significance and biogeographical relationships of the Mediterranean Miocene toothed whale fauna.- Geobios, Villeurbanne, vol. 35, p. 19-28.
Bianucci G., Landini W. & Varola A. (1994).- New remains of Cetacea Odontoceti from the Pietra Leccese (Apulia, Italy).- Bollettino della Societŕ Paleontologica Italiana, Modena, vol. 33, p. 231-242.
Boessenecker R.W. & Poust A.W. (2015).- Freshwater occurrence of the extinct dolphin Parapontoporia (Cetacea: Lipotidae) from the upper Pliocene nonmarine Tulare Formation of California.- Palaeontology, London, vol. 58, p. 489-496.
Brandano M., Scrocca D., Lipparini L., Petracchini L., Tomassetti L., Campagnoni V., Meloni D. & Mascaro G. (2013).- Physical stratigraphy and tectonic settings of Bolognano Formation (Majella): a potential carbonate reservoir.- Journal of Mediterranean Earth Sciences, Rome, Special Issue 2013, p. 151-176.
Brandt J.F. (1873).- Untersuchungen über die fossilen und subfossilen Cetaceen Europa's.- Mémoires de l'Académie impériale des sciences de St. Pétersbourg (7e série), Saint Petersburg, vol. 22, p. 1-361.
Brisson A.D. (1762).- Regnum Animale in Classes IX. Distributum, sive synopsis methodica.- Edito altera auctior, Theodorum Haak, Leiden, 296 p. URL: https://www.biodiversitylibrary.org/item/89810#page/5/mode/1up
Cahuzac B. & Poignant A. (1997).- Essai de biozonation de l'Oligo-Miocčne dans les bassins européens ŕ l'aide des grands foraminifčres néritiques.- Bulletin de la Société géologique de France, Paris, vol. 168, p. 155-169.
Carrillo-Briceńo J.D., Luz Z., Hendy A., Kocsis L., Aguilera O. & Vennemann T. (2019).- Neogene Caribbean elasmobranchs: diversity, paleoecology and paleoenvironmental significance of the Cocinetas Basin assemblage (Guajira Peninsula, Colombia).- Biogeosciences, vol. 6, p. 33-56.
Cason C., Grandesso P., Massari F. & Stefani C. (1981).- Deposi deltizi nella Molassa Cattiano-Burdigaliana del Bellunese.- Memorie di Scienze Geologiche, Padova, vol. 34, p. 325-354.
Catenacci V. (1974).- Note illustrative della Carta Geologica d'Italia. Foglio 147 "Lanciano".- Servizio Geologico d'Italia, Roma.
Churchill M., Martinez-Caceres M., Muizon C. de, Mnieckowski J. & Geisler J.H. (2016).- The origin of high-frequency hearing in whales.- Current Biology, London, vol. 26, p. 2144-2149.
Costa V., Doglioni C., Grandesso P., Masetti D., Pellegrini G.B. & Tracanella E. (1996).- Note illustrative della Carta Geologica d'Italia. Foglio 63 "Belluno".- Servizio Geologico d'Italia, Roma.
Crescenti U., Crostella A., Donzelli G. & Raffi G. (1969).- Stratigrafia della serie calcarea dal Lias al Miocene nella regione marchigiano-abruzzese (II–Litostratigrafia, biostratigrafia, paleogeografia).- Memorie della Societŕ Geologica Italiana, Roma, vol. 8, p. 343-420.
Dal Piaz G. (1900).- Sopra alcuni resti di Squalodon della arenaria miocenica di Belluno.- Palaeontographia italica, Pisa, vol. 6, p. 303-314.
Dal Piaz G. (1901).- Di alcuni resti di Cyrtodelphis sulcatus della arenaria miocenica di Belluno. Parte prima.- Palaeontographia italica, Pisa, vol. 7, p. 287-292.
Dal Piaz G. (1903).- Sugli avanzi di Cyrtodelphis sulcatus della arenaria di Belluno.- Palaeontographia italica, Pisa, vol. 9, p. 187-220.
Dal Piaz G. (1904).- Neosqualodon, nuovo genere della famiglia degli Squalodontidi.-- Mémoire de la Societé Paléontologique Suisse, Basel, vol. 31, p. 1-19.
Dal Piaz G. (1916a).- Gli Odontoceti del Miocene Bellunense. Parte seconda. Squalodon.- Memorie dell'Istituto di Geologia dell'Universitŕ di Padova, vol. 4, p. 1-79.
Dal Piaz G. (1916b).- Gli Odontoceti del Miocene Bellunense. Parte terza. Squalodelphis fabianii.- Memorie dell'Istituto di Geologia dell'Universitŕ di Padova, vol. 5, p. 1-28.
Dal Piaz G. (1916c).- Gli Odontoceti del Miocene Bellunense. Parte quarta. Eoplatanista italica.- Memorie dell'Istituto di Geologia dell'Universitŕ di Padova, vol. 5, p. 1-23.
Dal Piaz G. (1917).- Gli Odontoceti del Miocene Bellunense. Cyrtodelphis, Acrodelphis, Protodelphinus, Ziphiodelphis, Scaldicetus. Conclusioni generali e cosiderazioni filogenetiche. [manuscript dated 1916].- Memorie dell'Istituto di Geologia dell'Universitŕ di Padova, Allegato 4, p. 1-122.
Danese E. (1999).- Upper Miocene carbonate ramp deposits from the southernmost part of Maiella Mountain (Abruzzo, Central Italy).- Facies, Erlangen, vol. 41, p. 41-54.
Dewaele L., Lambert O. & Louwye S. (2018).- A critical revision of the fossil record, stratigraphy and diversity of the Neogene seal genus Monotherium (Carnivora, Phocidae).- Royal Society Open Science, vol. 5, article #171669, 31 p.
Dooley A.C. Jr (1998).- A review of the North America Squalodontidae (Mammalia, Cetacea).- Ph.D. Thesis, Louisiana State University, Baton Rouge, 271 p.
Dooley A.C. Jr (2003).- A review of the eastern north American Squalodontidae (Mammalia: Cetacea).- Jeffersoniana, Martinsville, vol. 11, p. 1-26.
Fabiani R. (1949).- Osservazione sulle forme di Neosqualodon del Miocene della Sicilia.- Rendiconti dell'Accademia Nazionale dei Lincei (serie 8), Roma, vol. 6, p. 428-430.
Flower W.H. (1867).- Description of the skeleton of Inia geoffrensis and of the skull of Pontoporia blainvillii, with remarks on the systematic position of these animals in the order Cetacea.- Transactions of the Zoological Society of London, London, vol. 6, p. 87-116.
Fordyce R.E. (1994).- Waipatia maerewhenua, new genus and new species (Waipatiidae, new family), an archaic late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand.- Proceedings of the San Diego Society of Natural History, San Diego, vol. 29, p. 147-176.
Fordyce R.E. & Muizon C. de (2001).- Evolutionary history of Cetaceans: a review. In: Buffrenil V. de & Mazin J.M. (eds.), Secondary adaptations of tetrapods to life in water.- Dr. Pfeil Verlag, Munich, p. 163-233.
Fornasiero M. & Del Favero L. (2014).- I Cetacei fossili del Museo di Geologia e Paleontologia dell'Universitŕ di Padova.- Museologia Scientifica Memorie, Firenze, vol. 13, p. 62-69
Geisler J.H. & Sanders A.E. (2003).- Morphological evidence for the phylogeny of Cetacea.- Journal of Mammalian Evolution, vol. 10, p. 23-129.
Gervais P. (1853).- Description de quelques espčces de phoques et des cétacés.- Annales de Science naturelle (Zoologie), Paris, vol. 20, p. 291-292.
Ghibaudo G., Grandesso P., Massari F. & Uchman A. (1996).- Use of trace fossils in delineating sequence stratigraphic surfaces (Tertiary, Venetian Basin, northeastern Italy).- Palæogeography, Palæoclimatology, Palæoecology, vol. 120, 261- 279.
Ghisetti F. & Vezzani L. (2002).- Normal faulting, extension and uplift in the outer thrust belt of the central Apennines (Italy): role of the Caramanico fault.- Basin Research, vol. 14, p. 225-236.
Godfrey S.J. (2013).- On the olfactory apparatus in the Miocene odontocete Squalodon sp. (Squalodontidae).- Comptes Rendus Palevol, Paris, vol. 12, p. 519-530.
Grateloup J.P.S. (1840).- Description d'un fragment de machoire fossile d'un genre nouveau de reptile (saurien).- Actes de l'Académie Nationale des Sciences Belles-Letres et Arts de Bordeaux, vol. 2, p. 201-210.
Guiscardi G. (1870).- Sopra un teschio fossile di foca.- Rendiconti dell'Accademia delle Science Fisiche e Matematiche, Napoli, vol. 5, p. 1-8.
Lambert O., Schlögl J. & Kováč M. (2008).- Middle Miocene toothed whale with Platanista-like teeth from the Vienna Basin (Western Carpathians, Slovakia).- Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen, Stuttgart, vol. 250, p. 157-166.
Lambert O., Muizon C. de, Malinverno E., Di Celma C., Urbina M. & Bianucci G. (2018).- A new odontocete (toothed cetacean) from the early Miocene of Peru expands the morphological disparity of extinct heterodont dolphins.- Journal of Systematic Palaeontology, London, vol. 16, p. 981-1016.
Landini W., Bianucci G., Bisconti M., Carnevale G., Sorbini C. & Varola A. (2005).- Il Miocene. I vertebrati marini. In: Bonfiglio L. (ed.), Paleontologia dei Vertebrati in Italia. Evoluzione biologica, significato ambientale e paleogeografia.- Memorie del Museo Civico di Storia Naturale di Verona (Serie II), vol. 6, p. 145-154.
Marsili S., Carnevale G., Danese E., Bianucci G. & Landini W. (2007).- Early Miocene vertebrates from Montagna della Maiella, Italy.- Annales de Paléontologie, Paris, vol. 93, p. 27-66.
Marx F.G., Lambert O. & Uhen M.D. (2016).- Cetacean paleobiology.- John Wiley & Sons, Chichester, 319 p.
Muizon C. de (1988).- Les Relations Phylogénétiques des Delphinida (Cetacea, Mammalia).- Annales de Paleontologie, Paris, vol. 74, p. 159-227.
Muizon C. de (1991).- A new Ziphiidae (Cetacea) from the Early Miocene of Washington State (USA) and phylogenetic analysis of the major groups of odontocetes.- Bulletin du Musée National d'Histoire Naturelle (série 4), Paris, vol. 12, 279-326.
Muizon C. de (1994).- Are the squalodonts related to the platanistoids?- Proceedings of the San Diego Society of Natural History, San Diego, vol. 29, p. 135-146.
Muizon C. de, Lambert O. & Bianucci G. (2018).- River dolphins, evolution. In: Würsig B., Thewissen J.G.M. & Kovacs K. (eds.), Encyclopedia of Marine Mammals, 3rd Edition.- Academic Press, San Diego, p. 829-835.
Mutti M., Bernoulli D. & Stille P. (1997).- Temperate carbonate platform drowning linked to Miocene oceanographic events: Maiella platform margin, Italy.- Terra Nova, vol. 9, p. 122-125.
Peri E., Collareta A., Insacco G. & Bianucci G. (2019).- An Inticetus-like (Cetacea: Odontoceti) postcanine tooth from the Pietra leccese (Miocene, southeastern Italy) and its palaeobiogeographical implications.- Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, Stuttgart, vol. 291, p. 221-228.
Rögl F. (1998).- Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene).- Annalen des Naturhistorischen Museums in Wien (serie A), vol. 99, p. 279-310.
Rothausen K. (1968).- Die systematische Stellung der europäischen Squalodontidae (Odontoceti, Mamm.).- Paläontologische Zeitschrift, Stuttgart, vol. 42, p. 83-104.
Rustichelli A., Tondi E., Agosta F., Cilona A. & Giorgioni M. (2012).- Development and distribution of bed-parallel compaction bands and pressure solution seams in carbonates (Bolognano Formation, Majella Mountain, Italy).- Journal of Structural Geology, vol. 37, p. 181-199.
Rustichelli A., Tondi E., Agosta F., Di Celma C. & Giorgioni M. (2013).- Sedimentologic and diagenetic controls on pore-network characteristics of Oligocene–Miocene ramp carbonates (Majella Mountain, central Italy). Sedimentologic and diagenetic controls on pore-network characteristics.- AAPG Bulletin, vol. 97, p. 487-524.
Scisciani V., Tavarnelli E. & Calamita F. (2002).- The interaction of extensional and contractional deformation in the outer zones of the central Apennines, Italy.- Journal of Structural Geology, vol. 24, p. 1647-1658.
Symeonidis N.K., Kazár E. & Roussiakis S.J. (2004).- Shark-toothed dolphin remains (Mammalia, Cetacea, Squalodontidae) from the early Miocene of Greece.- Annalen des Naturhistorischen Museums in Wien (serie A), vol. 105, p. 307-331.
Tanaka Y. & Fordyce R.E. (2014).- Fossil dolphin Otekaikea marplesi (latest Oligocene, New Zealand) expands the morphological and taxonomic diversity of Oligocene cetaceans.- PLOS ONE, vol. 9, no. 9, article #e107972, 30 p.
Tanaka Y. & Fordyce R.E. (2015a).- A new Oligo-Miocene dolphin from New Zealand: Otekaikea huata expands diversity of the early Platanistoidea.- Palaeontologia Electronica, vol. 18, article #18.2.23A, 71 p.
Tanaka Y. & Fordyce R.E. (2015b).- Historically significant late Oligocene dolphin Microcetus hectori Benham 1935: a new species of Waipatia (Platanistoidea).- Journal of the Royal Society of New Zealand, Wellington, vol. 45, p. 135-150.
Tanaka Y. & Fordyce R.E. (2016).- Papahu-like fossil dolphin from Kaikoura, New Zealand, helps to fill the Early Miocene gap in the history of Odontoceti.- New Zealand Journal of Geology and Geophysics, Wellington, vol. 59, p. 551-567.
Uhen M.D. (2008).- New protocetid whales from Alabama and Mississippi, and a new cetacean clade, Pelagiceti.- Journal of Vertebrate Paleontology, vol. 28, p. 589-593.
Vecsei A. & Sanders D.G.K. (1999).- Facies analysis and sequenze stratigraphy of a Miocene warm-temperate carbonate ramp, Montagna della Maiella Italy.- Sedimentary Geology, vol. 123, p. 103-127.
Vecsei A., Sanders D.G.K., Bernoulli D., Eberli G.P. & Pignatti J.S. (1998).- Cretaceous to Miocene sequence stratigraphy and evolution of the Maiella carbonate platform margin Italy. In: De Graciansky P.C., Jacquin T. & Vail P.R. (eds.), Mesozoic and Cenozoic sequence stratigraphy of European basins.- Society of Economic Paleontologists and Mineralogists Special Publication, Tulsa, vol. 60, p. 53-74.
Viglino M., Buono M.R., Fordyce R.E., Cuitińo J.I. & Fitzgerald E.M. (2018).- Anatomy and phylogeny of the large shark-toothed dolphin Phoberodon arctirostris Cabrera, 1926 (Cetacea: Odontoceti) from the early Miocene of Patagonia (Argentina).- Zoological Journal of the Linnean Society, London, vol. 185, p. 511-542.