◄ Carnets Geol. 20 (13) ►
![]()
Outline:
[Introduction]
[Geological setting]
[The Cretaceous of Aksai Chin]
[Material and methods]
[Paleontological note]
[Age and stratigraphic considerations]
[Comparison of the Aksai Chin fauna with other Neotethys faunas]
[Conclusions]
and ...
[Bibliographic references]
State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008 (China)
* Corresponding author
ISPRA - Geological Survey of Italy, via V. Brancati 48, 00144 Rome (Italy)
Strata GeoResearch Srl Spin Off INGEO Dept. of Engineering and Geology, University D'Annunzio Chieti-Pescara Via Dei Vestini, 31 66100 Chieti (Italy)
University of Trieste, Dept. of Mathematics and Geosciences, via Weiss 2, 34128 Trieste (Italy)
Published online in final form (pdf) on September 21, 2020
DOI 10.2110/carnets.2020.2013
![]()
[Editor:
Robert W. Scott; technical editor: Bruno Granier]
![]()
The rudist fauna collected in western Tibet in the Aksai Chin area by the De Filippi expedition in 1914 has been reprised and redescribed. This fauna is composed of Radiolites cf. lusitanicus, Radiolites sp., Gorjanovicia cf. endrissi, ? Sauvagesia sp., Sphaerulites sp., Durania sp., and Gyropleura sp. The rudist-bearing beds belong to the Tielongtan Group of the Tianshuihai terrane. The Turonian-? Coniacian Xiloqzung Formation (Fm.) bear the older rudists (Radiolites cf. lusitanicus, Radiolites sp., Sphaerulites sp., Durania sp.), whereas younger ages have been determined through microfossil analysis which, compared with the western Neotethys records, suggests an early-mid Campanian age. This allowed to ascribe the younger rudists of the collection (Gorjanovicia cf. endrissi, ? Sauvagesia sp., Gyropleura sp., Radiolites sp.) to the Dongloqzung Formation. Our data confirm that rudist-bearing facies in the Tielongtan Group extend at least up to the middle Campanian. The Aksai Chin rudist assemblage should belong to the Southwestern Asian assemblage of the Eastern Mediterranean Subprovince.
ò Hippuritida;
ò Tethysian biostratigraphy;
ò Upper Cretaceous of Tibet;
ò paleobiogeography;
ò museum collection
Sha J., Fabbi S., Cestari R. & Consorti L. (2020).- Stratigraphic and taxonomic considerations on the Late Cretaceous rudist fauna of Aksai Chin (Western Tibet, China) from the De Filippi Collection.- Carnets Geol., Madrid, vol. 20, no. 13, p. 249-272.
ConsidÚrations stratigraphiques et taxonomiques sur la faune de rudistes du CrÚtacÚ supÚrieur d'Aksai Chin (Tibet occidental, Chine) appartenant Ó la Collection De Filippi.- La faune de rudistes rÚcoltÚe dans la rÚgion d'Aksai Chin dans l'ouest du Tibet par l'expÚdition De Filippi en 1914 est rÚexaminÚe et redÚcrite. Cette faune est composÚe de Radiolites cf. lusitanicus, Radiolites sp., Gorjanovicia cf. endrissi, ? Sauvagesia sp., Sphaerulites sp., Durania sp. et Gyropleura sp. Les couches Ó rudistes appartiennent au Groupe Tielongtan du terrane de Tianshuihai. La Formation Xiloqzung du Turonien - ? Coniacien recÞle les rudistes les plus anciens (Radiolites cf. lusitanicus, Radiolites sp., Sphaerulites sp., Durania sp.), tandis que l'analyse du contenu microfossilifÞre par comparaison avec celui de la NÚotÚthys occidentale suggÞre un Ôge Campanien infÚrieur Ó moyen pour les rudistes les plus jeunes. Ceci permet d'attribuer les rudistes les plus jeunes appartenant Ó la collection (Gorjanovicia cf. endrissi, ? Sauvagesia sp., Gyropleura sp. et Radiolites sp.) Ó la Formation de Dongloqzung. Nos donnÚes confirment que les faciÞs Ó rudistes du Groupe Tielongtan sont prÚsents au moins jusqu'au Campanien moyen. L'association de rudistes de l'Aksai Chin devrait Ûtre rattachÚe Ó l'association du sud-ouest asiatique au sein de la Sous-Province mÚditerranÚenne orientale.
ò Hippuritida ;
ò biostratigraphie de la TÚthys ;
ò CrÚtacÚ supÚrieur du Tibet ;
ò palÚobiogÚographie ;
ò collection de musÚe
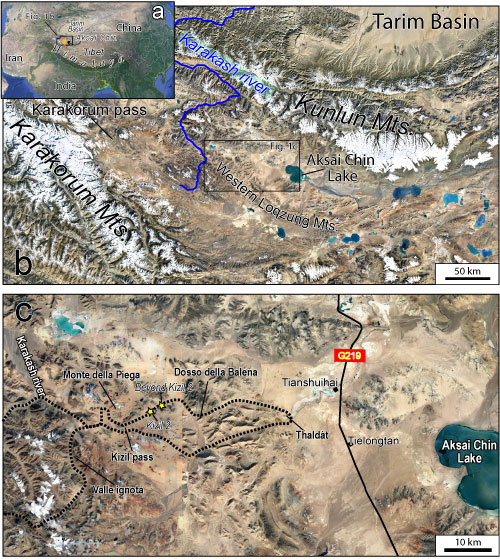
The Aksai Chin is a remote region at the western termination of the Tibetan Plateau (China), extending at its westernmost point to the Karakorum Pass (Fig.
1.a ![]() ). It is essentially a wide system of plateaus, partly occupied by salt lakes (e.g., Aksai Chin Lake), the average elevation of which is greater than 4800 m. Northward, it is bordered by and separated from the Tarim Basin by the Kunlun Range, a mountain chain that exceeds 7000 m
asl. The Upper Karakash River, which is one of the main tributaries of the Tarim River, flows in the western part of the Aksai Chin, along a deeply incised valley. In the central sector of the region a high mountain ridge exceeding 6400 m
asl, is known as Western Loqzung Mountains/Luokezongshan (Fig.
1.b
). It is essentially a wide system of plateaus, partly occupied by salt lakes (e.g., Aksai Chin Lake), the average elevation of which is greater than 4800 m. Northward, it is bordered by and separated from the Tarim Basin by the Kunlun Range, a mountain chain that exceeds 7000 m
asl. The Upper Karakash River, which is one of the main tributaries of the Tarim River, flows in the western part of the Aksai Chin, along a deeply incised valley. In the central sector of the region a high mountain ridge exceeding 6400 m
asl, is known as Western Loqzung Mountains/Luokezongshan (Fig.
1.b ![]() ).
).
Aksai Chin is almost uninhabited and very difficult for researchers to investigate because the average altitude is more than 4800 m
asl, with a frigid climate and reduced oxygen levels, which result in a wide desert land. Furthermore, travelling is extremely difficult across the region with a single, wide, long road (G219) (Fig.
1.c ![]() )
built at the end of 20th century.
)
built at the end of 20th century.
Because of such issues, the geology of Aksai Chin was sporadically investigated through time and, with few exceptions, most of the significant expeditions in the area were made in the first half of 20th century (Hedin,
1909; De Terra, 1932; Norin,
1946). After such pioneering explorations this remote area was almost forgotten by geologists until the
'80s of the 20th century, when it was investigated by the "Comprehensive Scientific Expedition to the Tibetan Plateau" in the years 1987-1990 (Chinese Academy of Sciences,
1998, 2000). Also it was only marginally the object of studies of the huge Sino-English
"geotraverse of Tibet" (Chang et al.,
1986, 1989; Sino-UK Geological Expedition Team,
1990), and subsequently of the Sino-French expedition, which followed the route of the G219 Highway (Matte et al.,
1996; Gaetani, 2011). Regarding the area east of the Karakorum Pass, and in particular between the Upper Karakash Valley and the ThaldÓt Basin (i.e., Loqzung
Mts./Luokezongshan subregion - Chinese Academy of Sciences, 2000), the first scientific data known to date are those collected by the Italian De Filippi Expedition made in 1913-1914 (De Filippi,
1915; Dainelli, 1934; reviewed by Gaetani, 2011). This expedition crossed the Western Loqzung Mts., through the Kizil Pass and reached the ThaldÓt Basin and the Aksai Chin Lake (Fig.
1.c ![]() ). Later data was collected by the "Comprehensive Scientific Expedition to the Tibetan Plateau" (Chinese Academy of Sciences,
1998, 2000). During the
De Filippi expedition, which crossed the Himalayan Range from Kashmir to Baltistan and Xinjiang and lasted for about two years, almost one month was spent in the Aksai Chin Region (Dainelli, 1934). The ~1000 rock and fossil samples collected during the expedition are stored at the Natural History Museum of Florence University (Italy), and represent still today one of the most complete geological archives of the area (Gaetani, 2011).
). Later data was collected by the "Comprehensive Scientific Expedition to the Tibetan Plateau" (Chinese Academy of Sciences,
1998, 2000). During the
De Filippi expedition, which crossed the Himalayan Range from Kashmir to Baltistan and Xinjiang and lasted for about two years, almost one month was spent in the Aksai Chin Region (Dainelli, 1934). The ~1000 rock and fossil samples collected during the expedition are stored at the Natural History Museum of Florence University (Italy), and represent still today one of the most complete geological archives of the area (Gaetani, 2011).
The first goal of this paper is to re-evaluate in modern terms the taxonomy of this remarkable rudist collection from the De Filippi expedition in order to refine the Upper Cretaceous stratigraphy. The second goal is to place this area into a wider paleobiogeographic framework by comparing it with more recently published research (Wen, 1998; Wen et al., 1998; Chinese Academy of Sciences, 2000).
|
Figure 1:
(a) and (b) Location and
geographical setting of the study area (basemap retrieved from Google Earth).
(c) close-up of the study area, the dashed line is the route of the De Filippi
expedition through the W-Loqzung Mts., yellow stars indicate sampling localities. |
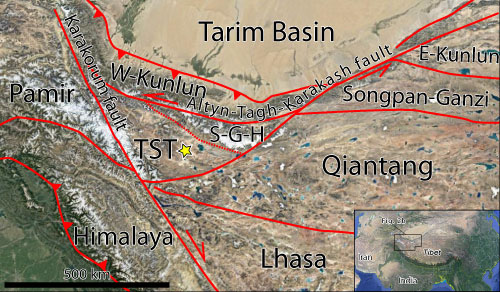
The geological setting of the study area is complex because of the superimposition of multiple orogenic phases (e.g., Molnar & Tapponnier,
1975; Sengor et al.,
1988; DeCelles et al.,
2002; Yin & Harrison,
2000; Mattern & Schneider,
2000). The subsequent accretion of crustal blocks at the southern margin of the Siberian Craton or paleo-Asia (Fig. 2 ![]() )
persisted from Paleozoic to Cenozoic times (Dewey et al.,
1988; Windley, 1988; Chang et al.,
1989; Gaetani, 1997; Zanchi & Gaetani,
2011; Angiolini et al., 2013; Faisal et al.,
2014). The northernmost of these crustal blocks is represented by the western Kunlun Range, a Paleozoic orogen rejuvenated during the early Cenozoic collisional stage of the Himalaya-Karakorum-Tibet orogenic system (Tapponnier & Molnar,
1977; Chang et al., 1986,
1989; Yongun & Hsu,
1994; Matte et al.,
1996; Chinese Academy of Sciences, 2000; Xiao et al.,
2005; Zhang et al., 2019). The southern boundary of the western Kunlun range is represented by the regional scale Altyn Tagh û Karakash strike-slip fault or Kunlun-Maqe Fault
(Fig. 2
)
persisted from Paleozoic to Cenozoic times (Dewey et al.,
1988; Windley, 1988; Chang et al.,
1989; Gaetani, 1997; Zanchi & Gaetani,
2011; Angiolini et al., 2013; Faisal et al.,
2014). The northernmost of these crustal blocks is represented by the western Kunlun Range, a Paleozoic orogen rejuvenated during the early Cenozoic collisional stage of the Himalaya-Karakorum-Tibet orogenic system (Tapponnier & Molnar,
1977; Chang et al., 1986,
1989; Yongun & Hsu,
1994; Matte et al.,
1996; Chinese Academy of Sciences, 2000; Xiao et al.,
2005; Zhang et al., 2019). The southern boundary of the western Kunlun range is represented by the regional scale Altyn Tagh û Karakash strike-slip fault or Kunlun-Maqe Fault
(Fig. 2 ![]() ; Matte et al.,
1996; Sha & F³rsich,
1999; Xiao et al., 2005; Sha et al.,
2004; Zanchi & Gaetani, 2011).
; Matte et al.,
1996; Sha & F³rsich,
1999; Xiao et al., 2005; Sha et al.,
2004; Zanchi & Gaetani, 2011).
A complex system of terranes south of the Kunlun block, drifted northward after the fragmentation of the northern margin of Gondwana (Sengor et al.,
1988). It was accreted during the late Middle-Late Permian to the Eocene
(Fig. 2 ![]() ), and includes the Songpan-Ganzi-Hoxhil terrane, the Qiantang composite terrane, the Tianshuihai terrane, the Lhasa terrane, and the Himalaya terrane (Molnar & Tapponnier,
1975; Huang & Chen,
1987; Dewey et al., 1988; Windley,
1988; Chang et al., 1989; Gaetani et al.,
1990; Matte et al.,
1996; Sha & F³rsich,
1999; Sha,
1998; Yin & Harrison,
2000; Sha et al., 2004; Zanchi & Gaetani,
2011; Sha, 2018).
), and includes the Songpan-Ganzi-Hoxhil terrane, the Qiantang composite terrane, the Tianshuihai terrane, the Lhasa terrane, and the Himalaya terrane (Molnar & Tapponnier,
1975; Huang & Chen,
1987; Dewey et al., 1988; Windley,
1988; Chang et al., 1989; Gaetani et al.,
1990; Matte et al.,
1996; Sha & F³rsich,
1999; Sha,
1998; Yin & Harrison,
2000; Sha et al., 2004; Zanchi & Gaetani,
2011; Sha, 2018).
|
Figure
2:
Tectonic framework of the
study area. The yellow star indicates the study area. TST=Tianshuihai terrane,
S-G-H=Songpan-Ganzi-Hoxhil terrane; Modified after Yin & Harrison,
2000 (basemap retrieved from Google Earth). |
Most of the Aksai Chin Region belongs to the Tianshuihai terrane (Pan et al.,
1992; Matte et al.,
1996; Chinese Academy of Sciences, 2000; Wittlinger et al.,
2004; Xiao et al., 2005) or
Kara-Kunlun and Uygur terranes (Mattern et al., 1996; Mattern & Schneider,
2000). The Tianshuihai terrane is a small crustal block interposed between the larger
Kunlun, Pamir, Songpan-Ganzi-Hoxhil, and Qiantang terranes (Fig. 2 ![]() ). It was interpreted as an accretionary wedge produced by the northward subduction of the Paleotethys oceanic crust beneath the Kunlun block (Mattern & Schneider,
2000; Xiao et al., 2005). The general stratigraphy of the Tianshuihai terrane
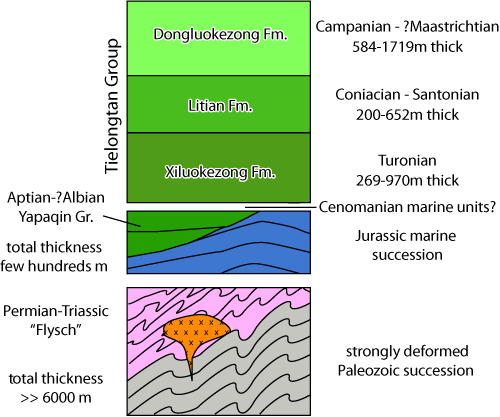
(Fig.
3
). It was interpreted as an accretionary wedge produced by the northward subduction of the Paleotethys oceanic crust beneath the Kunlun block (Mattern & Schneider,
2000; Xiao et al., 2005). The general stratigraphy of the Tianshuihai terrane
(Fig.
3 ![]() ) is characterized by a thick (>> 6000 m)
? Carboniferous to Permian succession consisting of fusulinid limestones (marbles), green schists and brachiopod-bearing marbles that overly poorly known early Paleozoic rocks (Matte et al.,
1996). These are overlain by
Permo-Triassic dark grey slates and barely metamorphic flyschoid sandstones (Chang et al.,
1989; Matte et al.,
1996; Mattern et al., 1996). The stratigraphic succession was strongly deformed during the Triassic -
? Early Jurassic (Cimmerian) orogeny (Mattern & Schneider,
2000; Sengor et al., 1988). Arc magmatism affected the Tianshuihai accretionary wedge, with batholiths intruding the sedimentary succession trending parallel to the orogen (Mattern & Schneider,
2000). The 215 and 171 Ma ages of the plutonic rocks (mostly
granitoid) were determined using various radiometric methods (40Ar/39Ar,
K/Ar, U/Pb) (Matte et al., 1996; Mattern & Schneider,
2000). This magmatic activity ceased in the Middle Jurassic (Mattern & Schneider,
2000).
) is characterized by a thick (>> 6000 m)
? Carboniferous to Permian succession consisting of fusulinid limestones (marbles), green schists and brachiopod-bearing marbles that overly poorly known early Paleozoic rocks (Matte et al.,
1996). These are overlain by
Permo-Triassic dark grey slates and barely metamorphic flyschoid sandstones (Chang et al.,
1989; Matte et al.,
1996; Mattern et al., 1996). The stratigraphic succession was strongly deformed during the Triassic -
? Early Jurassic (Cimmerian) orogeny (Mattern & Schneider,
2000; Sengor et al., 1988). Arc magmatism affected the Tianshuihai accretionary wedge, with batholiths intruding the sedimentary succession trending parallel to the orogen (Mattern & Schneider,
2000). The 215 and 171 Ma ages of the plutonic rocks (mostly
granitoid) were determined using various radiometric methods (40Ar/39Ar,
K/Ar, U/Pb) (Matte et al., 1996; Mattern & Schneider,
2000). This magmatic activity ceased in the Middle Jurassic (Mattern & Schneider,
2000).
Unconformably covering the Paleozoic-Triassic deformed rocks is a marine sedimentary succession consisting of a few
hundred (?) meters of Jurassic black shales, sandstones and limestones (Fig.
3 ![]() ). This is, in turn, covered by a thick Cretaceous stratigraphic succession made of conglomerates, sandstones and limestones, containing macrofossils such as ammonites, echinoids, bivalves (including rudists), corals among others (Parona,
1928; Stefanini, 1928; Dainelli,
1933-1934; Wen et al.,
1998, 2000a, 2000b). The Jurassic-Cretaceous sedimentary successions of the Aksai Chin are poorly investigated, but since the end of the last century, a general stratigraphic subdivision was established by Wen et al.
(1998, 2000a, 2000b) based on paleontology and superposition. In this paper we contribute to the knowledge of the Cretaceous stratigraphy of the area through the analysis of rudists collected in the surroundings of the Kizil Pass by the Italian scientists Giotto Dainelli and Olinto Marinelli in 1914. These fossils are from the Tielongtan Group (see below), which crops out, strongly deformed, in the Western Loqzung Mts. (Dainelli,
1933-1934; Matte et al.,
1996; Wen et al.,
2000a, 2000b).
). This is, in turn, covered by a thick Cretaceous stratigraphic succession made of conglomerates, sandstones and limestones, containing macrofossils such as ammonites, echinoids, bivalves (including rudists), corals among others (Parona,
1928; Stefanini, 1928; Dainelli,
1933-1934; Wen et al.,
1998, 2000a, 2000b). The Jurassic-Cretaceous sedimentary successions of the Aksai Chin are poorly investigated, but since the end of the last century, a general stratigraphic subdivision was established by Wen et al.
(1998, 2000a, 2000b) based on paleontology and superposition. In this paper we contribute to the knowledge of the Cretaceous stratigraphy of the area through the analysis of rudists collected in the surroundings of the Kizil Pass by the Italian scientists Giotto Dainelli and Olinto Marinelli in 1914. These fossils are from the Tielongtan Group (see below), which crops out, strongly deformed, in the Western Loqzung Mts. (Dainelli,
1933-1934; Matte et al.,
1996; Wen et al.,
2000a, 2000b).
|
Figure
3:
Sketch of the general
stratigraphy of the W-Loqzung Mts. (modified
after Matte et al., 1996; Wen
et al., 2000a, 2000b). |
Stratigraphy
A discontinuous stratigraphic succession crops out in the Aksai Chin, characterizing the Loqzung Mts./Luokezongshan stratigraphic subregion (Wen et al.,
1998, 2000a, 2000b), previously studied by pioneers such as Hedin
(1909), Parona (1917;
1928), Stefanini (1917), DouvillÚ
(1926) and Dainelli
(1933-1934). The Lower Cretaceous includes both continental and marine units: the
Aptian-? Albian Yapaqin Group
(Fig.
3 ![]() ) is made of interbedded marine (limestone) and non-marine facies (Sun & Zhang,
1983; Wen et al.,
2000a); this unit likely corresponds to the possibly Albian crinoid red limestone and underlying conglomerates, described by Parona (1928) and Dainelli
(1933-1934), and to the Albian limestones inferred by Matte et al.
(1996). The occurrence of Barremian limestone with Choffatella sp. and orbitolinids described by DouvillÚ
(1916, 1926) in samples collected by Hedin
(1909) is considered not reliable.
) is made of interbedded marine (limestone) and non-marine facies (Sun & Zhang,
1983; Wen et al.,
2000a); this unit likely corresponds to the possibly Albian crinoid red limestone and underlying conglomerates, described by Parona (1928) and Dainelli
(1933-1934), and to the Albian limestones inferred by Matte et al.
(1996). The occurrence of Barremian limestone with Choffatella sp. and orbitolinids described by DouvillÚ
(1916, 1926) in samples collected by Hedin
(1909) is considered not reliable.
The Lower Cretaceous is disconformably covered by marls and limestones belonging to the Tielongtan Group
(Fig.
3 ![]() ), which marks a regional transgression considered it to be Cenomanian (Sun & Zhang,
1983; Xi et al., 2019) or Turonian (Wen et al.,
2000a, 2000b). Elsewhere it directly overlies the Upper Jurassic (e.g., Sun & Zhang,
1983). The Tielongtan Gp. is a very thick (up to >3300 m) stratigraphic unit. It is subdivided into the Xiloqzung/Xiluokezong (269-970 m thick), Litian (200-652 m thick) and Donglokzung/Dongluokezong (584-1719 m thick) formations. Its age was suggested as being late Turonian to Campanian (even to early Maastrichtian) based on rudists and gastropods (Wen et al.,
1998, 2000a, 2000b). Recently, some authors inferred the range of Tielongtan Group to be early Cenomanian to late Maastrichtian (e.g., Xi et al.,
2019), although they do not provide any good evidence.
), which marks a regional transgression considered it to be Cenomanian (Sun & Zhang,
1983; Xi et al., 2019) or Turonian (Wen et al.,
2000a, 2000b). Elsewhere it directly overlies the Upper Jurassic (e.g., Sun & Zhang,
1983). The Tielongtan Gp. is a very thick (up to >3300 m) stratigraphic unit. It is subdivided into the Xiloqzung/Xiluokezong (269-970 m thick), Litian (200-652 m thick) and Donglokzung/Dongluokezong (584-1719 m thick) formations. Its age was suggested as being late Turonian to Campanian (even to early Maastrichtian) based on rudists and gastropods (Wen et al.,
1998, 2000a, 2000b). Recently, some authors inferred the range of Tielongtan Group to be early Cenomanian to late Maastrichtian (e.g., Xi et al.,
2019), although they do not provide any good evidence.
Palaeontology
The fossil assemblages that are the object of this study, led Parona
(1917,
1928) to subdivide the Upper Cretaceous succession in two parts:
1. A Cenomanian -
? "Senonian" limestone and marly limestone with abundant macrofossils such as ammonoids in the lowermost part, with rudist bivalves, echinoids and miliolids (Parona,
1917,
1928; Dainelli, 1933-1934);
2. An
? "upper Senonian" limestone with bivalves (including rudists), bryozoans, brachiopods, and possibly orbitoids (Parona,
1928; Dainelli, 1933-1934). This subdivision was obviously only tentative, and they did not reconstruct a reliable stratigraphy, but the stratigraphic succession seems roughly to correspond to the Tielongtan Gp., which is composed of interbedded brown calcareous sandstones, marls, red conglomerates, gray massive bioclastic limestones, and brown sandstones, also bearing bioclastic limestones in the upper part (Yang et al.,
2014). This group yields, in fact, abundant marine bivalves, in particular, rudists, which are abundant along the eastern slopes of Western Loqzung Mts. (Wen et al.,
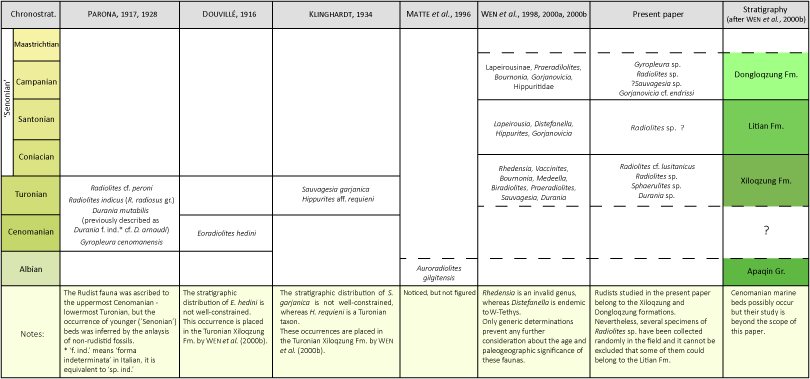
2000a, 2000b). According to Wen et al.
(2000b) the Xiloqzung/Xiluokezong Formation yields a rich rudist assemblage with Rhedensia, Vaccinites, Bournonia, Medeella, Biradiolites, Praeradiolites, Sauvagesia, and Durania. The Litian Formation yields Lapeirousia, Distefanella, Hippurites, and Gorjanovicia. The Dongloqzung/Dongluokezong Formation yields Lapeirousinae, Praeradiolites, Bournonia, Gorjanovicia, and Hippuritidae
(Table 1 ![]() ). Some of these determinations should be reviewed and are not considered here.
). Some of these determinations should be reviewed and are not considered here.
Table 1: Stratigraphic chart of rudist taxa of the Aksai Chin region cited in the literature and notes.
|
This is a study of rudist-bearing samples stored in the Natural History Museum of the University of Florence
(Table 2), from the Aksai Chin area
(Fig. 1 ![]() ). Sampling was carried out by unconventional methods, and it is very difficult to refer each specimen to a precise locality. Parona
(1917,
1928) and Dainelli (1934) erroneously said that the samples were from the "Lingzi-Thang area", which is actually the Western Loqzung Mts. The actual collection area is marked by well-developed rudist banks, with specimens collected in the surroundings of Kizil Pass, "along the valley descending from the pass toward NE in the left riverside" and "beyond Kizil II camp site" (Parona,
1917). In addition, we have tried our best to refer each specimen to a formation, following the stratigraphic nomenclature established by Wen et al.
(1998, 2000a, 2000b).
). Sampling was carried out by unconventional methods, and it is very difficult to refer each specimen to a precise locality. Parona
(1917,
1928) and Dainelli (1934) erroneously said that the samples were from the "Lingzi-Thang area", which is actually the Western Loqzung Mts. The actual collection area is marked by well-developed rudist banks, with specimens collected in the surroundings of Kizil Pass, "along the valley descending from the pass toward NE in the left riverside" and "beyond Kizil II camp site" (Parona,
1917). In addition, we have tried our best to refer each specimen to a formation, following the stratigraphic nomenclature established by Wen et al.
(1998, 2000a, 2000b).
Unfortunately, most of the nearly 100 specimens of the collection are broken isolated fragments or fragmental in whole rock. The list of specimens in the collection, with register numbers (Table 2), includes several specimens that are grouped under the same register number.
Ten samples have been cut to check the shell structure and for general microfacies analysis, resulting in 20 thin sections. Due to limitations of the preservation of fossil material and that only a few specimens were sectioned, only gross descriptions of the main features of the rudists have been perfomed. Shell shape and structure description follows Dechaseaux et al. (1969), Pons & Vicens (2008), Pons et al. (2012), and Skelton (2013).
Abbreviations: AB=Anterior Band; PB=Posterior Band; L= Ligament ridge; CA= Cardinal apparatus; IGF = Register number.
Table 2: Aksai Chin Rudist collection stored at the Natural History Museum of Florence University (Italy). A catalogue number can include one or more specimens.
| IGF | Description | N. | Order | Family | Genus | Species | Locality | Nation | Epoch | Age | Year | Collectors |
| 103252 | Fragments with rudists and matrix | 7 | Hippuritoida | Monopleuridae | - | Gyropleura cenomanensis | Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103253 | Fragments with rudists and matrix | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103254 | Fragments with rudists and matrix | 11 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103255 | Fragments with rudists and matrix | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103256 | Fragments with rudists and matrix | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103257 | Fragments with rudists and matrix | 8 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103258 | Single specimen | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites cf. peroni | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103259 | Broken specimens | 7 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | Aksai Chin | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103260 | Fragments | 2 | Hippuritoida | Radiolitidae | Radiolites | Radiolites cf. peroni | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103261 | Fragments | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites cf. peroni | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103262 | Fragments | 2 | Hippuritoida | Radiolitidae | Radiolites | Radiolites cf. peroni | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103263 | Fragments | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites cf. peroni | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103264 | Fragments | 4 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103265 | Fragments | 7 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103266 | Fragments | 3 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103267 | Fragments | 15 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | Aksai Chin | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103268 | Single specimen | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites indicus | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103269 | Fragments | 15 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103270 | Two specimens | 2 | Hippuritoida | Radiolitidae | - | Durania mutabilis | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103942 | Fragments with rudists and matrix | 1 | Hippuritoida | Monopleuridae | Gyropleura | Gyropleura cenomanensis | Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103943 | Fragments with rudists and matrix | 1 | Hippuritoida | Monopleuridae | Gyropleura | Gyropleura cenomanensis | Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103944 | Fragments with rudists and matrix | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103945 | Fragments with rudists and matrix | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103946 | Fragments with rudists and matrix | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
| 103947 | Fragments with rudists and matrix | 1 | Hippuritoida | Radiolitidae | Radiolites | Radiolites sp. | beyond Kizil 2 | CHINA | Upper Cretaceous | Turonian | 1914 | Dainelli & Marinelli |
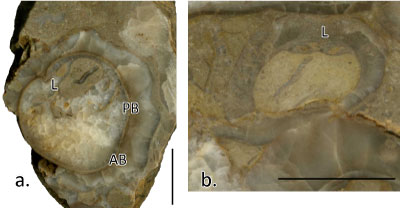
The rudist bivalves (Order HIPPURITIDA Newell, 1965) of the De Filippi fossil collection were originally determined by Parona (1917, 1928), who recognized three (or possibly more) species of radiolitids and one of monopleurids: Radiolites indicus (Stoliczka) (Radiolites radiosus Orb. group); Radiolites cf. peroni (Choffat); Radiolites sp.; Durania f. ind. (cf. D. arnaudi (Choff.)), then identified as Durania mutabilis and Gyropleura cenomanensis (Orb.).
Family Radiolitidae Orbigny, 1847
Original determination (Parona, 1928): Radiolites cf. peroni (Choffat)
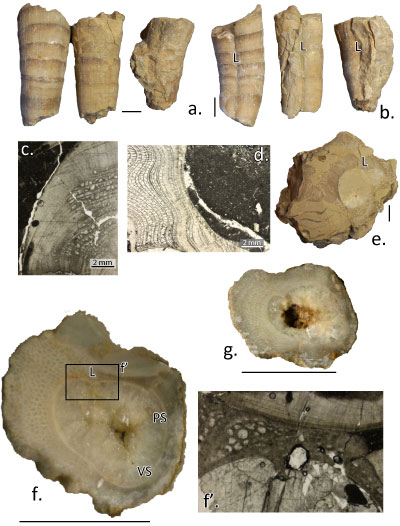
Radiolites cf. lusitanicus (Bayle, 1857)
(Fig. 4.a-g ![]() )
)
Material: 5 quite well-preserved right valves and 7 fragments (IGF 103258, 103260, 103261, 103262, 103263).
Locality: beyond Kizil 2 (Fig. 1 ![]() )
)
Inferred formation: Xiloqzung Fm.
Age: ? Turonian-? Coniacian
Description: Elongate shell, subcylindrical, up to >90 mm high showing slightly inclined growth lamellae sets and dominant normal cellular shell structure
(Fig. 4.a, g, g' ![]() ). In transverse sections the cardinal apparatus is well developed
(Fig. 4.f
). In transverse sections the cardinal apparatus is well developed
(Fig. 4.f ![]() ). On the external side, well impressed growth cycles are present with robust non-protruding costae
(Fig. 4.a-b, d
). On the external side, well impressed growth cycles are present with robust non-protruding costae
(Fig. 4.a-b, d ![]() ). Growth cycles are well impressed also interiorly, in the inner shell of elongated specimens, from which the outer shell layer has spalled away
(Fig. 4.e
). Growth cycles are well impressed also interiorly, in the inner shell of elongated specimens, from which the outer shell layer has spalled away
(Fig. 4.e ![]() ). The radial bands are on indented folds producing smooth sinuses
(Fig. 4.a-b, g
). The radial bands are on indented folds producing smooth sinuses
(Fig. 4.a-b, g ![]() ). The ligament ridge is well marked and projected towards the ventral side
(Fig. 4.e-g
). The ligament ridge is well marked and projected towards the ventral side
(Fig. 4.e-g ![]() ).
).
Discussion: This fossil material was described as Radiolites cf. peroni by Parona (1917, 1928). Radiolites peroni was considered as a distinct species but only differs from Radiolites lusitanicus by the number of folds in the interband, which is variable and not a distinctive character (Steuber, 1999). For this reason and the common occurrence of both species in the same stratigraphic interval (Pejovic, 1957; Berthou, 1973; Steuber, 1999) led Steuber (1999) to consider R. peroni a junior synonym of R. lusitanicus. More fossil material close to the type locality in Portugal has been recently described, with a shell structure marked by well differentiated lamellar sets interiorly inclined with regular, well developed, small costae, externally barely visible radial bands and normal cells passing locally to very elongate cells from the inner towards the external part of the outer shell layer (Troya-GarcÝa, 2016).
Due to the poorly preserved morphostructural features of the studied material, a definite attribution of this fossil material to this species is not possible due to: (a) the notable presence of compact structure together with the normal cellular structure of growth lamellae sets
(Fig. 4, inset g' ![]() ), (b) the ligament ridge that is more projected toward the inner cavity in our material with respect to the material described by Troya-GarcÝa (2016), and (c) the unclear organization of radial bands. Despite this, we have no sufficient data to refute the original determination by Parona, thus we stand on its attribution, updated to modern taxonomy (e.g., Steuber,
1999).
), (b) the ligament ridge that is more projected toward the inner cavity in our material with respect to the material described by Troya-GarcÝa (2016), and (c) the unclear organization of radial bands. Despite this, we have no sufficient data to refute the original determination by Parona, thus we stand on its attribution, updated to modern taxonomy (e.g., Steuber,
1999).
R. lusitanicus has a broad geographical distribution, being reported from both the southern and northern margins of Neotethys, and from Portugal to the Mediterranean area, Middle-East, Afghanistan and western Tibet (e.g., Choffat, 1886; DouvillÚ, 1910; Parona, 1917, 1928; Steuber, 1999; Steuber & L÷ser, 2000; El Hedeny, 2007). Originally described as a Turonian species, its stratigraphic range is between the late Cenomanian and the Coniacian (Steuber, 1999; El Hedeny, 2007; Zakhera, 2011).
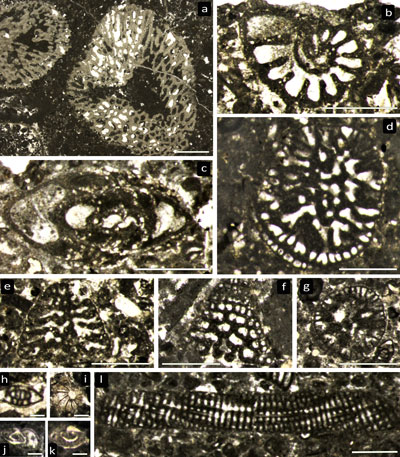
|
Figure
4:
Radiolites aff. lusitanicus (ex cf. peroni
in Parona, 1928). (a)
IGF103260 (figured as R. cf. peroni by Parona,
1928, Pl.
16, fig. 2), two joined specimens in lateral view. (b, c) IGF 103261, lateral
dorsal view and longitudinal section (showing slightly inclined growth lamellae
sets) of right valve, respectively. (d) IGF103260, lateral dorsal view of right
valve. (e) IGF 103263, dorsal view of right valve, only a thin layer of inner
shell preserved, with well impressed growth cycles and invaginated ligament
ridge. (f) IGF 103261, transverse section of right valve. (g, g') IGF 103258,
transverse section of two right valves with detail of the outer shell layer with
normal cellular (left) and compact (right) structure in inset f'. Scale bars = 1 cm. |
Original determination (Parona, 1928): Radiolites sp.
Several specimens, mainly fragments, in the collection were described by Parona (1928) as Radiolites sp. (Table 2). Most of the material shows no diagnostic characters, and its determination is thus left as Radiolites sp. sensu Parona (1928). But some distinction is made between two groups of specimens that show different characters. Note that register numbers can be repeated because different taxa can be found in the same polished slab.
Radiolites sp. 1
(Fig. 5.a-e ![]() )
)
Material: Several poorly preserved right valves and fragments (IGF 103253, 103254, 103255, 103256, 103257, 103259, 103264, 103265, 103266, 103267, 103269, 103944, 103946, 103947).
Locality: Kizil 2; beyond Kizil 2.
Inferred formation: Xiloqzung/Litian/Dongloqzung Fm.
Age: ? Turonian to ? Campanian
Description: Only right valves, commonly in the form of inner shell fragments, from which the outer shell layer has spalled away, have been studied. Shape is conical to low cylindrical, with well-developed growth cycles
(Fig. 5.a-b ![]() ). Shell is marked by growth lamellar sets with both compact and normal cellular structure
(Fig. 5.c-d
). Shell is marked by growth lamellar sets with both compact and normal cellular structure
(Fig. 5.c-d ![]() ), with this last more developed on the dorsal side.
), with this last more developed on the dorsal side.
Discussion: This material differs from other specimens by its elongate cylindrical shape and the dominant compact structure
(Fig. 5.e ![]() ), which contrasts to the cellular structure of Radiolites sp. 2.
), which contrasts to the cellular structure of Radiolites sp. 2.
Radiolites sp. 2
(Fig. 5.f-g ![]() )
)
Material: Few poorly preserved right valves and fragments (IGF 103253, 103254, 103254B, 103267).
Locality: Kizil 2; beyond Kizil 2.
Inferred formation: Litian/Dongloqzung Fm.
Age: ? Campanian
Description: Only right valves have been studied; shell form is conical to low cylindrical, structure of growth lamellar sets is normal cellular with large cells more developed in the thicker dorsal side with short ligament ridge
(Fig. 5, inset f' ![]() ),
whereas it is compact in the thinner antero-ventral side, where the radial structures are marked by slightly developed folds (as in 103267,
Fig. 5.f-g
),
whereas it is compact in the thinner antero-ventral side, where the radial structures are marked by slightly developed folds (as in 103267,
Fig. 5.f-g ![]() ).
).
Discussion: This material differs from the other group by its cylindro-conical shape, the dominant normal cellular structure
(Fig. 5.f-g ![]() ) and radial band organization.
) and radial band organization.
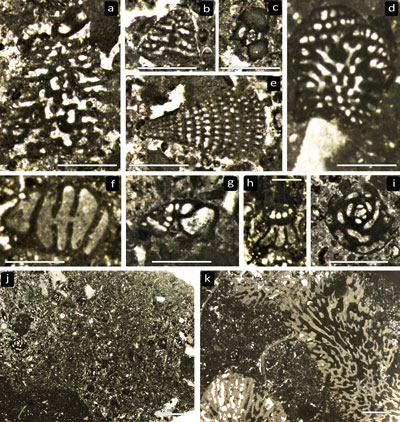
|
Figure
5:
Radiolites sp. (a,
b) IGF103265, ventral and dorsal views, respectively of elongate specimens, only
a very thin layer of the inner shell is preserved, with well impressed growth
cycles and invaginated ligament ridge. (c, d) IGF103267 and IGF 103269
respectively, thin sections of the outer shell showing growth lamellar sets with
both compact and normal cellular structures. (e) IGF103267, transverse section
of the right valve showing dominant compact structure. (f, g) IGF103267,
transverse sections of right valves showing normal cellular with well-developed
cells in the dorsal side and short ligament ridge (inset f'). Scale bars = 1
cm. |
Besides these two groups, other material originally described as Radiolites sp. has been tentatively assigned to different taxa.
Gorjanovicia cf. endrissi (Boehm, 1927)
(Fig. 6.a ![]() )
)
Material: IGF 103945, 103947
Locality: beyond Kizil 2
Inferred formation: Dongloqzung Fm.
Age: Campanian
Description: Two transverse sections of right valves are characterized by a thick compact outer shell layer; radial bands are marked by slightly developed folds with a slight external central indentation of the AB, which is separated from the PB by a narrow interband; a thin triangular ligamentary infolding is also present
(Fig. 6.a ![]() ).
).
Discussion: The radial bands organization, the thick compact outer shell and the ligamentary infolding suggest a similarity of our specimens to Gorjanovicia endrissi as described by Fenerci-Masse et al. (2011). The lack of any well-preserved material only allows an uncertain attribution to Gorjanovicia cf. endrissi.
? Sauvagesia sp.
(Fig. 6.b ![]() )
)
Material: IGF 103947
Locality: beyond Kizil 2
Inferred formation: Dongloqzung Fm.
Age: Campanian
Description: Another taxon previously assigned to Radiolites sp. is a single specimen cut in a polished section from a rock slab (IGF103947). The transverse section of the lower (right) valve, juvenile stage, shows a sub-rounded external perimeter and thick outer shell layer composed of growth lamellar sets with normal cellular structure. Ligament ridge is present and delimiting an undeveloped cardinal apparatus, at the edge of the dorsal side
(Fig. 6.b ![]() ).
).
Discussion: Although determined by a well-preserved single specimen, the polished slab reveals a shape and structure that could be possibly assigned to Sauvagesia sp. by the characteristic small cells of the cellular shell structure.
|
Figure
6:
(a) IGF103267, Transverse section of the right valve of Gorjanovicia
cf. endrissi. (b) IGF 103947, transverse section of the right valve,
juvenile stage, of ? Sauvagesia sp. Scale
bars = 1 cm. |
Original determination (Parona, 1917): Radiolites indicus (Stoliczka)
Sphaerulites sp.
(Fig. 7.a-d ![]() )
)
Material: 1 right valve (IGF 103268)
Locality: beyond Kizil 2
Inferred formation: Xiloqzung Fm.
Age: ? Turonian-? Coniacian (for its occurrence in association with R. cf. lusitanicus).
Description: This single specimen of the collection consists of a large right valve (up to 200 mm in diameter) partly deformed and broken on the ventral side
(Fig. 7.a ![]() ). The shell is cylindro-conical, at least 35 cm high, the right (lower) valve is very thick in the antero-dorsal side (up to 80 mm) with a wide radial, down-and-outward fold separating two badly preserved radial structures
(Fig. 7.a
). The shell is cylindro-conical, at least 35 cm high, the right (lower) valve is very thick in the antero-dorsal side (up to 80 mm) with a wide radial, down-and-outward fold separating two badly preserved radial structures
(Fig. 7.a ![]() ). Shell is composed of growth lamellar sets with normal cellular structure and cells commonly are elongate in the outer part, where lamellar sets are downfolded
(Fig. 7.a, c-d
). Shell is composed of growth lamellar sets with normal cellular structure and cells commonly are elongate in the outer part, where lamellar sets are downfolded
(Fig. 7.a, c-d ![]() ) giving rise to a foliaceous aspect. Growth cones cannot be observed in the outer surface as it is eroded and badly preserved. Ligament ridge is robust at the base
(Fig. 7.b
) giving rise to a foliaceous aspect. Growth cones cannot be observed in the outer surface as it is eroded and badly preserved. Ligament ridge is robust at the base
(Fig. 7.b ![]() ), presumably with a long pedicle. Left valve is absent.
), presumably with a long pedicle. Left valve is absent.
Discussion: The specimen was assigned to Radiolites indicus by Parona (1917), but the few preserved features suggest a different assignment to the genus Sphaerulites. The shell structure is marked by growth lamellar sets with normal cellular structure composed of elongate cells. The radial bands, although not clearly visible, are separated by a typical main fold in between them. The cellular structure differs slightly from the radial, essentially discontinuous ridges of the cellular structure typical of Sphaerulites (e.g., Troya-GarcÝa, 2016). The poorly preserved features and the scarcity of material do not allow a specific assignment, but we consider invalid the generic emendation by Parona, due to the analyzed characters, which led us to assign this specimen to Sphaerulites, rather than to Radiolites.
Sphaerulites indicus is a species not commonly reported in the scientific literature. It was established by Stolizcka (1871) from specimens in Upper Cretaceous brownish marly limestone at Odium (NW from Ottakkovil) south-east India. DouvillÚ (1910) suggested that S. indicus was similar to R. peroni, but he did not emend the taxon, and he noted that the figures by Stolizcka and the quality of the material was poor. S. indicus was then described as Radiolites indicus by Parona (1917) and listed in K³hn (1932) and Sanchez (1981). Our unique specimen and the sectioning limitations do not allow a better taxonomic redescription that could result in a definite specific determination. Only few species are currently assigned to genus Sphaerulites, and S. indicus is not included (as in Troya-GarcÝa, 2016). Due to the scarce reports and poor quality of the material, S. indicus would be possibly an invalid taxon. In addition, the hypothetical geographical distribution of the species should oddly be limited only to southeastern India and, eventually, the Aksai Chin area. These issues led us to consider, that if the species is valid, its actual occurrence in the Himalayan area is very dubious, and thus we assign our specimen to Sphaerulites sp.
|
Figure
7:
Sphaerulites sp. (a)
IGF103268, conical shaped specimen seen from above showing thick ligament ridge;
in the ventral side the presumably position of the radial bands is separated by
an upward fold. (b) IGF103268, internal view of growth cycles and robust
ligament ridge. (c) IGF103268, growth lamellar sets with normal cellular
structure characterized by elongated cells. (d) IGF103268, side view of
sub-horizontal and downfolded lamellae sets projecting laterally in
grain-supported sediment. |
Original determination (Parona, 1928): Durania mutabilis (Stol.)
Durania sp.
(Fig. 8.a-b ![]() )
)
Material: 1 almost complete right valve, 1 broken and weathered right valve, 1 fragment of right valve (IGF 103256, 103270).
Locality: beyond Kizil 2
Inferred formation: Xiloqzung
Age: ? Turonian-? Coniacian (for its occurrence in association with R. cf. lusitanicus)
Description. Although the material is not well preserved, the few specimens have an elongate cylindrical shape
(Fig. 8.b ![]() ) and well-developed growth laminar sets with normal cellular structure
(Fig. 8.a
) and well-developed growth laminar sets with normal cellular structure
(Fig. 8.a ![]() ). The radial bands have growth laminar sets that are concave in radial section and they are marked externally by a dozen thin costellae
(Fig. 8.b
). The radial bands have growth laminar sets that are concave in radial section and they are marked externally by a dozen thin costellae
(Fig. 8.b ![]() ) separated by a protruding main interband fold. Well-developed folds form external costae; no ligament ridge has been detected.
) separated by a protruding main interband fold. Well-developed folds form external costae; no ligament ridge has been detected.
Discussion. The studied material was first described as Durania f. ind. (cf. D. arnaudi (Choff.)) by Parona (1917) and then as Durania mutabilis (Stol.) by Parona (1928), who erroneously placed Radiolites mutabilis Stoliczka in "Durania mutabilis". However later R. mutabilis was reassigned to Bournonia mutabilis by DouvillÚ (1910), which is reported in the Maastrichtian Ottakkovil Fm. of southern India (Mohanti & Pons, 1999; Dihman et al., 2019). Except for the papers by Parona, "D. mutabilis" was no longer cited in the palaeontological literature (with the only exception of a paper by Morris & Skelton (1995), who considered D. mutabilis as a senior synonym of D. gaensis), and is considered the result of an erroneous taxonomic attribution by Parona. The described specimens can be, therefore, determined only at the generic rank, as Durania sp.
Family Monopleuridae (Munier-Chalmas, 1873)
Gyropleura sp. (Fig. 8.c-g ![]() )
)
Material: 4 rock samples with several specimens embedded in limestone matrix, some thin section has been performed (IGF 103252, 103942, 103943)
Locality: Kizil 2
Inferred formation: Dongloqzung Fm.
Age: Campanian
Description: Small (< 20 mm) spirogyrate valves (Fig.
8.c, e ![]() ), difficult to study in terms of shell structures and morphostructural analysis. A thin section performed on the whole rock has shown shell fragments made of plano-parallel growth laminar sets with compact calcite in the outer shell layer and the inner layer made of neomorphic calcite, in some case showing the middle layer, aragonitic in origin
(Fig. 8.f-g
), difficult to study in terms of shell structures and morphostructural analysis. A thin section performed on the whole rock has shown shell fragments made of plano-parallel growth laminar sets with compact calcite in the outer shell layer and the inner layer made of neomorphic calcite, in some case showing the middle layer, aragonitic in origin
(Fig. 8.f-g ![]() ).
).
Discussion: Gyropleura is a monopleurid rudist represented by few samples in the studied collection. The specimens described as Gyropleura cenomanensis by Parona
(1917) are difficult to determine at specific rank, being small and embedded in the rock
(Fig. 8.d-e ![]() ) and not showing any distinctive character. For this reason, nothing can be added in a paleontological point of view. It is worth noting that when morphostructural analyses can be performed only with difficulty, specific attribution is easily erroneous. Similar spirogyrate rudists occur beginning in the Aptian and up to the end of the Cretaceous (Steuber et al.,
2016). The studied specimens are associated with a Campanian microfossil assemblage and are considered as Gyropleura sp.
) and not showing any distinctive character. For this reason, nothing can be added in a paleontological point of view. It is worth noting that when morphostructural analyses can be performed only with difficulty, specific attribution is easily erroneous. Similar spirogyrate rudists occur beginning in the Aptian and up to the end of the Cretaceous (Steuber et al.,
2016). The studied specimens are associated with a Campanian microfossil assemblage and are considered as Gyropleura sp.
|
Figure
8:
(a, b) Durania sp., IGF103270, (a) transverse
section of right valve showing well developed folds and radial bands; (b)
ventral view of a right valve of Durania sp. showing ribs corresponding
to the radial bands (figured by Parona, 1917,
p. 135). IGF103270. (c-g) Gyropleura
sp., (c) IGF 103943, ventral view of a single spirogyrate valve; (d) IGF 103942,
fragments of made of compact calcite structure, embedded in grain-supported
limestone; (e) IGF 103252, bouquet of several spirogyrate valves of Gyropleura
sp.; (f) IGF103942, thin section of a shell fragment: outer shell layer (OL) is
made of compact calcite lamellar sets, a very thin middle shell layer (ML)
separate it from the inner shell layer (IL) made of white neomorphic calcite
substituting the original aragonite. Scale bars 1 cm. (g.) IGF103942, same shell
fragment shown in (f) with crossed polars, showing calcite crystals oriented
perpendicular to the growth lines, typical of rudists. |
Rudists
Comparing the original sedimentological description of the succession, which yielded the studied material (as said, collected more than one century ago in an inaccessible area, Dainelli, 1933-1934), with the recently assessed stratigraphic setting of the area (Wen et al., 2000a, 2000b), we infer that the rudist beds represented in the collection belong to the Tielongtan Group.
The lower part of the Tielongtan Gp. (Xiloqzung Formation) is Turonian-Coniacian p.p. according to Wen et al.
(2000a, 2000b). The older rudists determined at generic rank by Wen et al.
(1998, 2000a, 2000b) in the Xiloqzung Fm. were considered as Turonian, by the occurrence of Vaccinites, Bournonia and other taxa
(Table 1 ![]() ), which ruled out the hypothesis of a Cenomanian age.
), which ruled out the hypothesis of a Cenomanian age.
Cenomanian and Turonian beds bearing Eoradiolites hedini, Sauvagesia garianica and Hippurites aff. requieni were noted by DouvillÚ
(1916) and Klinghardt
(1934), who studied samples collected by S. Hedin and H. De Terra
(Table 1 ![]() ), respectively. Nevertheless, the stratigraphic range of E. hedini and Sauvagesia garianica being not well constrained, and H. requeni definitely a Turonian taxon, these occurrences have to be considered as most likely Turonian.
), respectively. Nevertheless, the stratigraphic range of E. hedini and Sauvagesia garianica being not well constrained, and H. requeni definitely a Turonian taxon, these occurrences have to be considered as most likely Turonian.
As we have already discussed, age determinations by Parona (1917, 1928), who ascribed the studied rudist fauna to the upper Cenomanian-Turonian, are not reliable. The age of the older rudist assemblage of our material (R. cf. lusitanicus, Durania sp., Sphaerulites sp., Radiolites sp. 1) is not well-constrained as well, but the occurrence of Radiolites cf. lusitanicus, which is a Cenomanian-Coniacian taxon (Steuber, 1999), is significant, whereas other taxa only determined at generic rank do not provide any additional information, being Durania and Radiolites Albian-Maastrichtian genera, and Sphaerulites a Cenomanian-Campanian genus (Steuber et al., 2016).
The Turonian-Coniacian p.p. age of the Xiloqzung Fm. is fully consistent with the stratigraphic distribution of Radiolites lusitanicus and is thus inferred for the rudist-bearing beds of the succession characterized by its occurrence, which should belong to the Xiloqzung Fm.
Ages younger than the Coniacian were inferred in the literature and confirmed by analyzing the studied material. The rudist assemblages of the Litian Formation, composed of Lapeirousia, Hippurites, etc. determined by Wen et al. (1998, 2000a), were considered as being approximately Santonian but the lack of specific determinations does not allow a greater precision of stratigraphic attribution. Our material provided no additional data regarding this unit.
The upper part of the Tielongtan Gp. is represented by the the Dongloqzung Fm.; the occurrence of Lapeirousinae, which did not appear until Santonian, and Praeradiolites, Bournonia,
etc. (Table 1 ![]() ), which appeared slightly earlier but extended into Maastrichtian, led Wen et al.
(1998, 2000a, 2000b) to infer a mainly Campanian or possibly Maastrichtian age.
), which appeared slightly earlier but extended into Maastrichtian, led Wen et al.
(1998, 2000a, 2000b) to infer a mainly Campanian or possibly Maastrichtian age.
Some of the studied rudists have been referred to this unit (Gorjanovicia cf. endrissi, ? Sauvagesia sp., Radiolites sp., Gyropleura sp.) with the only exception of G. cf. endrissi, which is essentially a Campanian taxon (Fenerci-Masse et al., 2011), other taxa do not provide any additional information, but their age has been constrained based on micropaleontological analysis (see below).
Benthic Foraminifera
Along with the study of rudist assemblages, micropaleontological analyses performed through thin sections on the rock matrix embedding the rudist specimens revealed the occurrence of some stratigraphically relevant foraminifers, which provide important information on the temporal position of the samples.
The carbonate matrix embedding Radiolites sp. is represented by a bioclastic packstone-wackestone and subordinate floatstone
(Figs. 9.a ![]() ,
10.k
,
10.k ![]() ), with Dicyclina schlumbergeri Munier-Chalmas (Fig.
9.l
), with Dicyclina schlumbergeri Munier-Chalmas (Fig.
9.l ![]() ), possible Murgeina apula (Luperto Sinni)
(Fig. 9.h-i
), possible Murgeina apula (Luperto Sinni)
(Fig. 9.h-i ![]() ), possible Navarella Ciry & Rat
(Fig. 9.c
), possible Navarella Ciry & Rat
(Fig. 9.c ![]() ), a possible Nezzazatinella sp.
(Fig. 9.b
), a possible Nezzazatinella sp.
(Fig. 9.b ![]() ), and a probable specimen structurally close to a macrospheric
? Keramosphaerina Stache (Fig.
10.i
), and a probable specimen structurally close to a macrospheric
? Keramosphaerina Stache (Fig.
10.i ![]() ). We have observed three features supporting the identification of Navarella
(Fig. 9.c
). We have observed three features supporting the identification of Navarella
(Fig. 9.c ![]() ): 1) the microgranular shell texture; 2) the accumulation of coarse carbonate grains over the outer shell surface, and 3) the streptospiral arrangement typical of Navarella. The geometry of chambers over the central area of the cross section suggests a degree of involution rather than a perfectly symmetrical disposition of lumens that, in a planispiral geometry, should appear on both sides. Regarding
? Keramosphaerina sp. (Fig. 10.i
): 1) the microgranular shell texture; 2) the accumulation of coarse carbonate grains over the outer shell surface, and 3) the streptospiral arrangement typical of Navarella. The geometry of chambers over the central area of the cross section suggests a degree of involution rather than a perfectly symmetrical disposition of lumens that, in a planispiral geometry, should appear on both sides. Regarding
? Keramosphaerina sp. (Fig. 10.i ![]() ), another possible identification may be a microspheric Hellenalveolina (Hottinger et al.,
1989). That species, however, has more distinctly streptospiral chambers than our specimen. Furthermore, the proloculus size is substantially smaller than that of a megalospheric Hellenalveolina. Moreover, Hellenalveolina seems geographically restricted into the Upper Cretaceous of Europe. Having a look at the initial chambers of Keramosphaerina in Devoto
(1964) we have found a close match. Anyway, due to the scarcity of material at our disposal we prefer keeping this identification under open nomenclature. The identification of Murgeina apula (see Chiocchini et al.,
2012) is supported by the thickly calcified umbos on both sides of the test. Nezzazatinella sp. can be linked to the Santonian taxon figured by Chiocchini et al.
(2012) as N. picardi, but a more specific taxonomic designation is hampered by the lack of diagnostic sections.
), another possible identification may be a microspheric Hellenalveolina (Hottinger et al.,
1989). That species, however, has more distinctly streptospiral chambers than our specimen. Furthermore, the proloculus size is substantially smaller than that of a megalospheric Hellenalveolina. Moreover, Hellenalveolina seems geographically restricted into the Upper Cretaceous of Europe. Having a look at the initial chambers of Keramosphaerina in Devoto
(1964) we have found a close match. Anyway, due to the scarcity of material at our disposal we prefer keeping this identification under open nomenclature. The identification of Murgeina apula (see Chiocchini et al.,
2012) is supported by the thickly calcified umbos on both sides of the test. Nezzazatinella sp. can be linked to the Santonian taxon figured by Chiocchini et al.
(2012) as N. picardi, but a more specific taxonomic designation is hampered by the lack of diagnostic sections.
This western Tibetan foraminiferal assemblage is also known in the western Neotethys where it ranges from upper Santonian to at least middle Campanian (Radoičić et al., 2010; Frijia et al., 2015; Consorti et al., 2017); this age is attributed to the Litian and Donglozung formations.
The packstone matrix embedding Gyropleura sp. contains, along with other abundant undetermined microfossils, the stromatoporoid Sarmentofascis zamparelliae Schlagintweit, Frijia & Parente
(Figs. 9.a ![]() ,
10.k
,
10.k ![]() ), Calveziconus lecalvezae Caus
(Figs. 9.d, f
), Calveziconus lecalvezae Caus
(Figs. 9.d, f ![]() ,
10.a, d
,
10.a, d ![]() ), a small and high-conical agglutinated foraminifer closely resembling the Orbitolinidae gen. ind. in Luperto Sinni & Ricchetti
(1978:
Pl. 43, figs. 3, 7), Cuneolina sp., (Fig. 10.e
), a small and high-conical agglutinated foraminifer closely resembling the Orbitolinidae gen. ind. in Luperto Sinni & Ricchetti
(1978:
Pl. 43, figs. 3, 7), Cuneolina sp., (Fig. 10.e ![]() ) and Cibicidoides sp.
(Fig. 9.j-k
) and Cibicidoides sp.
(Fig. 9.j-k ![]() ). In particular, the identification of Calveziconus lecalvezae is supported by its subepidermal network formed by two orders of beams and rafters, and by radial undulated septula. Orbitolinidae gen.
ind. (Figs. 9.e, g
). In particular, the identification of Calveziconus lecalvezae is supported by its subepidermal network formed by two orders of beams and rafters, and by radial undulated septula. Orbitolinidae gen.
ind. (Figs. 9.e, g ![]() ,
10.b
,
10.b ![]() ) lacks the second order of beams. Their occurrence in such levels is justified by Luperto Sinni & Ricchetti
(1978) as well, who found some strictly comparable forms in the Upper Cretaceous carbonates of southern Italy, and in co-assemblage with Calveziconus. Their identification should be accompanied by a close taxonomical analysis, likely resulting in the description of a new taxon, which falls beyond the scope of the present contribution. This foraminiferal assemblage, if correlated with the European records, suggests an age ranging between the lower and the middle Campanian. This age of the European assemblage is firmly constrained by the Strontium Isotope Stratigraphy studies made through the levels bearing Calveziconus lecalvezae (Steuber et al.,
2005;
Frijia et al., 2015; Villalonga et al.,
2019) and Sarmentofascis zamparelliae (Schlagintweit et al.,
2016). Cibicidoides sp. is commonly recorded in the Maastrichtian (Consorti & Rashidi,
2019) and the lower Paleocene (see Gavelinellidae indet. in Serra-Kiel et al.,
2020), but its stratigraphic range would widen into older or younger levels, thus limiting its biostratigraphic potential. Besides giving critical and more detailed stratigraphic information on the Tibetan rudists, this assemblage allows us to definitely consider that the rudist-bearing facies of the Tielongtan Gp. extends up to, at least, the middle Campanian.
) lacks the second order of beams. Their occurrence in such levels is justified by Luperto Sinni & Ricchetti
(1978) as well, who found some strictly comparable forms in the Upper Cretaceous carbonates of southern Italy, and in co-assemblage with Calveziconus. Their identification should be accompanied by a close taxonomical analysis, likely resulting in the description of a new taxon, which falls beyond the scope of the present contribution. This foraminiferal assemblage, if correlated with the European records, suggests an age ranging between the lower and the middle Campanian. This age of the European assemblage is firmly constrained by the Strontium Isotope Stratigraphy studies made through the levels bearing Calveziconus lecalvezae (Steuber et al.,
2005;
Frijia et al., 2015; Villalonga et al.,
2019) and Sarmentofascis zamparelliae (Schlagintweit et al.,
2016). Cibicidoides sp. is commonly recorded in the Maastrichtian (Consorti & Rashidi,
2019) and the lower Paleocene (see Gavelinellidae indet. in Serra-Kiel et al.,
2020), but its stratigraphic range would widen into older or younger levels, thus limiting its biostratigraphic potential. Besides giving critical and more detailed stratigraphic information on the Tibetan rudists, this assemblage allows us to definitely consider that the rudist-bearing facies of the Tielongtan Gp. extends up to, at least, the middle Campanian.
|
Figure
9:
Chronostratigraphically important benthic Foraminifera
and stromatoporoids recovered from the matrix of the rudist specimens. (a) IGF
103944, Sarmentofascis zamparelliae Schlagintweit et al..
Scale bar 2 mm. (b) IGF 103269, Nezzazatinella sp. Scale bar 0.5 mm. (c) IGF 103264, Navarella sp. Ciry & Rat.
Scale bar 0.5 mm. (d, f) IGF 103942 and 103946, Calveziconus lecalvezae Caus,
basal and longitudinal sections respectively. Scale bar 0.5 mm. (e, g) IGF 103264 and 103946,
Orbitolinidae gen. ind. Scale bar 0.5 mm. (h, i) IGF 103269, Murgeina apula
(Luperto Sinni). Scale bar 0.1 mm. (j, k) IGF 103942, Cibicidoides
sp. Scale bar 0.1 mm. (l) IGF 103942, Dicyclina schlumbergeri Munier-Chalmas.
Scale bar 0.5 mm. |
|
Figure
10:
Benthic Foraminifera and facies of the matrix of rudist
specimens. (a, d) IGF 103269; 103943, Calveziconus lecalvezae Caus.
Scale bar 0.5 mm. (b) IGF 103269, Orbitolinidae gen. ind. Scale bar 0.5
mm. (c) IGF 103264, Thick-shelled undetermined miliolid. Scale bar 0.5 mm. (e) IGF 103269, Cuneolina sp.
Scale bar 0.5 mm. (f, g) IGF 103942; 103946, Nezzazatinella sp. Scale bar
0.5 mm. (h) IGF 103942, Nezzazatinella sp. scale bar 0.25 mm. (i) IGF103254, megalospheric ? Keramosphaerina sp.
Scale bar 1 mm. (j) IGF 103254, Packstone facies rich in miliolid foraminifers.
Scale bar 2 mm. (k) IGF 103944, floatstone with Sarmetofascis zamparelliae.
Scale bar 2 mm. |
Rudist faunas from the Himalaya-Karakorum-Tibet range and the Tarim basin are known from the late Aptian and are well-developed into the Maastrichtian (Lan & Wei, 1995; Wen et al., 1998; Scott et al., 2010; Rao et al., 2015, 2017, 2020; Sha & Cestari, 2016; Rao, 2019).
Scott et al. (2010) reported 36 genera and 44 species occurring in the region in the Albian-Maastrichtian interval. Most of the taxa were considered endemic and only 10 species were considered as being cosmopolitan.
In particular, the uppermost Lower Cretaceous Langshan Formation and the Upper Cretaceous Tielongtan Group of Lhasa, Qiantang and Tianshuiai terranes and the Tarim basin, yielded rich rudist faunas (Wen, 1998; Wen et al., 1998; Scott et al., 2010). The genera determined from the Langshan Fm. include Monopleura, Auroradiolites, Eoradiolites, Requienia, Magallanesia, Sellaea, and Toucasia (Scott et al., 2010; Rao et al., 2015, 2017). Differently, the Upper Cretaceous Tielongtan Group yielded a fauna dominated by warm, shallow-water Tethysian rudists, such as Hippurites, Vaccinites, Praeradiolites, Biradiolites, Durania, Sauvagesia, Distefanella, and Lapeirousia, among others. They are more abundant and diverse than rudists of the Himalaya, Shiqunhe-Xainza (Southeastern Tethys) and southwestern margin of the Tarim Basin (Northeastern Tethys) (Wen, 1999; Scott et al., 2010). In the Kunlun terrane and Tarim Basin, Late Cretaceous rudists are Biradiolites, Gyropleura, Lapeirousella, Osculigera, and Sauvagesia (Lan & Wei, 1995; Wen, 1999; Rao, 2019).
The rudist fauna of the Aksai Chin in the Tielongtan Group is not rich and has low variability compared with published data. In addition, it is similar in terms of age with the Western Tarim fauna (Yigeziya and Wuyitake formations) described by Lan & Wei (1995) such as the Campanian age of Gyropleura beds.
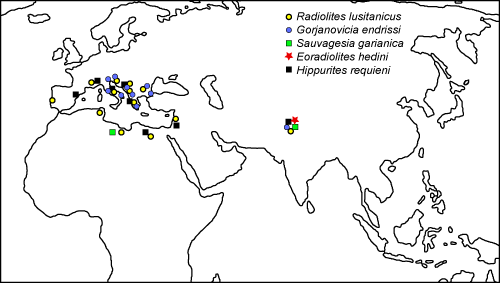
The taxa described in this paper should be considered cosmopolitan; in particular Radiolites lusitanicus and Gorjanovicia endrissi are known from most of the
Tethysian realm
(Fig. 11 ![]() ).
).
Radiolites indicus (sensu Parona), as said, is considered an invalid taxon and is useless for paleogeographic considerations. Gyropleura sp., Praeradiolites sp., Sphaerulites sp., Sauvagesia sp., Durania sp., and Radiolites sp. provide no further information on their distribution pattern, not being determined at specific rank. Along with the described rudists, also the Turonian Sauvagesia garianica and Hippurites aff. requieni described by Klinghardt (1934) are considered as cosmopolitan taxa, the first having been found in the Aksai Chin, Lybia and Tunisia (Parona, 1921; Busson, 1967), and the second being a common taxon of both the northern and southern margins of Neotethys (e.g., Cestari & Sartorio, 1995; Steuber & L÷ser, 2000).
In contrast, the Albian - Cenomanian rudists Auroradiolites gilgitensis (= Eoradiolites gilgitensis) and Eoradiolites hedini (= Praeradiolites hedini) described by Matte et al.
(1996) and DouvillÚ (1916) are considered as endemic taxa (Rao et al.,
2015), being only known from the northeastern margin of the Neotethys
(Fig. 11 ![]() ).
).
No further paleobiogeographic consideration can be done regarding the rudist assemblages described by Wen et al. (1998, 2000a, 2000b), being only generic determinations of cosmopolitan genera, except for Rhedensia, which is not a valid genus according to Steuber et al. (2016) and Distefanella, which is a radiolitid with an extremely elongate shell whose occurrence is limited to the central Mediterranean Tethys (Cestari et al., 2018).
The Aksai Chin rudist assemblage should belong to the Southwestern Asian assemblage of the Eastern Mediterranean subprovince, as defined by Kauffman (1973) and Scott et al. (2010).
Concerning benthic foraminifers and Demospongiae, the specimens identified in the studied microfacies show strong affinities with that known in the shallow-water deposits of central-western Neotethys including the central and southern Apennines (Italy: Frijia et al., 2015; Schlagintweit et al., 2016; Consorti et al., 2017), Dinarides (Croatia: Steuber et al., 2005), Apulian platform (Italy: Luperto Sinni & Ricchetti, 1978) and southeastern Pyrenees (northeast Spain: Villalonga et al., 2019, among others). It is thus affirmed that during the Campanian a biogeographic connection, represented by foraminifers and Demospongiae, was established between the Tibetan and Mediterranean areas. The confidence in the age assignment is provided by a direct correlation with the European fauna, which stratigraphic age is calibrated by means of Strontium Isotope (e.g., Steuber et al., 2005; Frijia et al., 2015; Villalonga et al., 2019). However, we cannot exclude the presence of a biogeographic effect, likely due to faunal dispersal, which may slightly affect such age-correlation. Like other shallow-water taxa, large Upper Cretaceous agglutinating and porcelaneous foraminifera were possibly subject to provincialism and periodic populations diversification (e.g., Hottinger et al., 1989; Goldbeck & Langer, 2009). This is the reason why Navarella from Tibet would not exactly match with the European forms. Even in Europe, the specific diversity (or the occurrence of different varieties) may be higher than the expected, as Maync (1954) demonstrates. In the Tibetan samples, in fact, there are certain morphotypes such as Nezzazatinella sp. and ? Keramosphaerina sp., or the Orbitolinidae gen. ind., that cannot be assigned to any species so far known. These are likely new taxa, but their detailed systematic placing is hampered by the lack of additional material.
|
Figure
11:
Geographical distribution of the rudist species from the
Western Loqzung Mountains discussed in this paper. |
The rudist fauna of the De Filippi collection from western Tibet has been reprised and redescribed. The previous taxonomic attribution of this fauna has been mostly considered inaccurate, with the only exception of R. cf. lusitanicus, which has been renamed after the recent emendation of the taxon R. peroni. Regarding the other specimens no definitely specific attributions have been possible due to the poor quality of the material. The fauna is mostly composed of radiolitids, R. cf. lusitanicus, two more species of Radiolites, Gorjanovicia cf. endrissi, a possible ? Sauvagesia sp., a Durania and a specimen, which was previously described as Radiolites indicus but is most likely a Sphaerulites. The only non-radiolitid taxon which was determined in the studied rudist assemblage is the monopleurid Gyropleura sp. (previously erroneously determined as G. cenomanensis).
The rudists have been collected in beds belonging to the Tielongtan Group. The oldest rudists belong to the Xiloqzung Formation and their age is Turonian-? Coniacian. Younger ages have been determined through the analysis of the carbonate matrix embedding the rudists, which, compared with the western Neotethys records suggest a lower-middle Campanian age, allowing to ascribe such specimens to the Litian Formation and Dongloqzung Formation. Our data confirm that rudist-bearing facies in the Tielongtan Gp. extends at least up to the middle Campanian.
The described rudist fauna shows some similarity with the Western Tarim fauna and is composed of cosmopolitan taxa Radiolites lusitanicus and Gorjanovicia cf. endrissi, which are in fact known from most of the Tethysian realm.
Benthic Foraminifers and Demospongiae in the Dongloqzung Formation and associated with the younger rudists of the studied fauna (Gorjanovicia cf. endrissi, ? Sauvagesia sp., Gyropleura sp., Radiolites sp.) suggest that during the Campanian a biogeographic connection existed between the Tibetan and Mediterranean areas. However, it cannot be excluded that an effect due to faunal dispersal may slightly affect the age-correlation and the evolution of possibly endemic taxa. The Aksai Chin rudist assemblage should belong to the Southwestern Asian assemblage of the Eastern Mediterranean Subprovince.
We are grateful to Stefano Dominici of the Natural History Museum of Florence University (Italy) for his kindness and for giving us the permissions to study the material of the De Filippi Collection. We also are grateful to the careful reviews by the rudist specialists J.M. Pons, R.W. Scott and P.W. Skelton, which greatly improved the quality of the original manuscript. This work is a contribution to the StrataGeoResearch Project "Tethyan Carbonate Platforms". This work was supported by the National Natural Science Foundation of China (41730317), Chinese Academy of Geological Sciences (DD20190009).
Angiolini L., Zanchi A., Zanchetta S., Nicora A. & Vezzoli G. (2013).- The Cimmerian geopuzzle: New data from South Pamir.- Terra Nova, vol. 25, no. 5, p. 352-360.
Bayle E. (1857).- Nouvelles observations sur quelques espÞces de rudistes.- Bulletin de la SociÚtÚ GÚologique de France, vol. 2, no. 14, p. 647-719.
Berthou P.-Y. (1973).- Le CÚnomanien de l'EstrÚmadure portugaise.û Servicos geol¾gicos de Portugal, Mem¾ria (N. S.), vol. 23, 169 p.
Boehm J. (1927).- Beitrag zur Kenntnis der Senon fauna der Bithynischen Halbinsel.- Paleontographica, vol. 69, p. 187û 222.
Busson G. (1967).- MÚsozo´que saharien-1 Partie - L'ExtrÛme-Sud Tunisien.- Centre Recherche. Zones Arides (SÚries gÚologiques), CNRS, vol. 8, 194 p.
Cestari R., Pons J.M., Vicens E. & Di Michele P. (2018).- Distefanella Parona, 1901 a radiolitid rudist from Late Cretaceous platform margin in central-eastern Mediterranean Tethys.- Cretaceous Research, vol. 90, p. 265-282.
Cestari R. & Sartorio D. (1995).- Rudists and facies of the periadriatic domain.- Agip, San Donato Milanese, Italy, 207 p.
Chang C., Chen N., Coward M.P., Wanming D., Dewey J.F., Gansser A., Harris N.B.W, Chengwei J., Kidd W.S.F., Leeder M.R., Huan L., Jinlu L., Chengjie L., Houjun M., Molnar P., Pan Y., Pan Y., Pearce J.A., Shackleton R.M., Smith A.B., Sun Y., Ward M., Watts D.R., Xu J., Xu R., Yin J. & Zhang Y. (1986).- Preliminary conclusions of the Royal Society/Academia Sinica 1985 Geotraverse of Tibet.- Nature, vol. 323, p. 501û507.
Chang C.F., Pan Y.S. & Sun Y.Y. (1989).- The tectonic evolution of Qinghai-Tibet Plateau: A review. In: Sengor A.M.C. (Ed.), Tectonic evolution of the Tethyan region.- Springer, Dordrecht, p. 415-476.
Chinese Academy of Sciences (1998).- Paleontology of the Karakorum-Kunlun Mountains.- Science Press, Beijing, 365 p. [in Chinese with English abstract].
Chinese Academy of Sciences (2000).- Geological evolution of the Karakorum-Kunlun Mountains.- Science Press, Beijing. 525 p. [in Chinese].
Chiocchini M., Pampaloni M.L. & Pichezzi R.M. (2012).- Microfacies and microfossils of the Mesozoic carbonate successions of Latium and Abruzzi (Central Italy).- Memorie per Servire alla Descrizione della Carta Geologica D'Italia, vol. 17, 269 p.
Choffat P. (1886).- Recueil d'Útudes palÚontologiques sur la faune crÚtacique du Portugal. EspÞces nouvelles ou peu connues.- Comunicaþoes dos Serviþos geologicos de Portugal, 40 p.
Consorti L., Frijia G. & Caus E. (2017).- Rotaloidean foraminifera from the Upper Cretaceous carbonates of Central and Southern Italy and their chronostratigraphic age.- Cretaceous Research, vol. 70, p. 226-243.
Consorti L. & Rashidi K. (2019).- Remarks on Fissoelphidium operculiferum Smout, 1955 (larger Foraminifera, Maastrichtian of Zagros) and comments on the associated rotaloidean and other lamellar perforate Foraminifera.- Cretaceous Research, vol. 94, p. 59-71.
Dainelli G. (1933-1934).- La serie dei Terreni, in Relazioni Scientifiche della Spedizione Italiana De Filippi nell'Himalaia, Caracorum e Turchestan Cinese, serie II û Resultati geologici e geografici, vol. 2.- Zanichelli, Bologna, 458 p. + 542 p.
Dainelli G. (1934).- La esplorazione della regione fra l'HimÓlaja occidentale e il Caracor¨m, in Relazioni Scientifiche della Spedizione Italiana De Filippi nell'Himalaia, Caracorum e Turchestan Cinese, serie II û Resultati geologici e geografici, vol. 1.- Zanichelli, Bologna, 434 p.
De Filippi F. (1915).- Spedizione scientifica italiana in India e in Asia Centrale negli anni 1913-1914, tenuta dal capo della spedizione nell'anfiteatro Augusteo il 15 Aprile 1915.- Bollettino della SocietÓ Geografica Italiana, vol. LII, p. 645-685.
De Terra H. (1932).- Wissenschaftliche Ergebnisse der Dr. Trinkler'schen Zentralasien-expedition, vol. 2.- Reimer/Vohsen, Berlin, 121 p.
DeCelles P.G., Robinson D.M. & Zandt G. (2002).- Implications of shortening in the Himalayan fold-thrust belt for uplift of the Tibetan Plateau.- Tectonics, vol. 21, no. 6 (12), p. 1-25.
Deschaseaux C., Coogan A. & Cox L. (1969).- Systematic descriptions; HippuritoidaûHippuritacea. In: Cox L.R., Newell N.D., Boyd D.W., Branson C.C., Casey R., Chavan A., Coogan A.H., Dechaseaux C., Fleming C.A., Haas F., Hert-Lein L.G., Kauffman E.G., Keen A.M., Larocque A., Mcalester A.L., Moore R.C., Nuttall C.P., Perkins B.F., Puri H.S., Smith L.A., Soot Ryen T., Stenzel H.B., Trueman E.R., Turner R.D. & Weir J. (Eds.), Treatise on Invertebrate Paleontology, Part N, 3, Mollusca 6, Bivalvia.- Geological Society of America and University of Kansas Press, N749ûN817.
Devoto G. (1964).- Il passaggio Cretacico-Paleocene nei Monti Lepini e il problema relativo a Keramosphaera tergestina.- Geologica Romana, vol. 3, p. 49-64.
Dewey J.F., Shackleton R. M., Chang C. & Sun Y. (1988).- The tectonic development of the Tibetan plateau.- Philosophical Transactions of the Royal Society of London A, vol. 327, p. 379û413.
Dihman H., Prasad G.V. & Goswami A. (2019).- Parataxonomy and Palaeobiogeographic significance of dinosaur eggshell fragments from the Upper Cretaceous strata of the Cauvery Basin, South India.- Historical Biology, vol. 31, no. 10, p. 1310-1322.
DouvillÚ H. (1910).- Études sur les Rudistes. Rudistes de Sicile, d'AlgÚrie, d'Égypte, du Liban et de la Perse.- MÚmoires de la SociÚtÚ gÚologique de France (PalÚontologie), vol. 41, 95 p.
DouvillÚ H. (1916).- Les calcaires Ó Orbitolines et Ó Radiolites du Thibet. In: Hedin S. (Ed.), Southern Tibet û Discoveries in former times compared with my own researches in 1906û1908, Vol. 5.- Lithographic Institute of the General Staff of the Swedish Army, Stockholm, p. 145û147.
DouvillÚ H. (1926).- Fossiles recueillis par Hedin dans le Kashmir en 1906 et les Pamirs en 1914 ; leur description.- Records of the Geological Survey of India, vol. 58, p. 349û357.
El-Hedeny M.M. (2007).- New taxonomic and biostratigraphic data on the ? Upper CenomanianûTuronian Radiolitidae (Bivalvia: Hippuritoidea) of Abu Roash, Western Desert, Egypt.- Neues Jahrbuch f³r Geologie und Palõontologie-Abhandlungen, vol. 244, no. 1, p. 79-98.
Faisal S., Larson K. P., Cottle J. M. & Lamming J. (2014).- Building the Hindu Kush: Monazite records of terrane accretion, plutonism and the evolution of the HimalayaûKarakoramûTibet orogen.- Terra Nova, vol. 26, no. 5, p. 395-401.
Fenerci-Masse M., Skelton P.W. & Masse J.P. (2011).- The rudist bivalve genus Gorjanovicia (Radiolitidae, Hippuritoidea) a revision of species based on quantitative analysis of morphological characters.- Palaeontology, vol. 54, p. 1û23.
Frijia G., Parente M., Di Lucia M. & Mutti M. (2015).- Carbon and strontium isotope stratigraphy of the Upper Cretaceous (Cenomanian-Campanian) shallow-water carbonates of southern Italy: Chronostratigraphic calibration of larger foraminifera biostratigraphy.- Cretaceous Research, vol. 53, p. 110-139.
Gaetani M. (1997).- The Karakorum Block in Central Asia, from Ordovician to Cretaceous.- Sedimentary Geology, vol. 109, p. 339-359.
Gaetani M. (2011).- Fossili della collezione De Filippi, raccolti da Giotto Dainelli e Olinto Marinelli. In: Monechi S. & Rook L. (Eds.), Il Museo di Storia Naturale dell'UniversitÓ di Firenze, volume III, Le Collezioni Geologiche e Paleontologiche.- Firenze University Press, p. 100-103.
Gaetani M., Pognante U. & Gosso G. (1990).- A geological transect from Kun Lun to Karakorum (Sinkiang, China). The western termination of the Tibetan Plateau. Preliminary note.- Terra Nova, vol. 2, p. 23-30.
Goldbeck E.J. & Langer M. (2009).- Biogeographic provinces and patterns of diversity in selected Upper Cretaceous (Santonian-Maastrichtian) larger foraminifera.- SEPM Special Publication, vol. 93, p. 187û232.
Hedin S. (1909).- Trans-Himalayan Discoveries and Adventures in Tibet, Vol. 1.- Macmillan, London, 436 p.
Hottinger L., Drobne K. & Caus E. (1989).- Late Cretaceous, larger, complex miliolids (Foraminifera) endemic in the Pyrenean Faunal Province.- Facies, vol. 21, p. 99û134.
Huang J. & Chen B. (1987).- The evolution of the Tethys in China and adjacent region.- Geological Publishing House, Beijing, 109 p.
Kauffman E.G. (1973).- Cretaceous Bivalvia. In: Hallam A. (Ed.), Atlas of palaeobiogeography.- Elsevier Scientific Publishing Company, Amsterdam, p. 353û383.
Klinghardt F. (1934).- Rudisten aus West-Tibet (Von der Forschungsreise Trinkler-De Terra 1928/29.).- Zeitschrift der Deutschen Geologischen Gesellschaft, vol. 86, p. 110-114.
K³hn O. (1932).- Rudistae from Eastern Persia.- Records of the Geological Survey of India, vol. 46, p. 151û179.
Lan X. & Wei J. (1995).- Late Cretaceousûearly Tertiary marine bivalve fauna from the western Tarim Basin.- Chinese Science House, Beijing, 212 p.
Luperto Sinni E. & Ricchetti G. (1978).- Studio micropaleontologico-stratigrafico di una successione carbonatica del Cretaceo superiore rilevata nel sottosuolo delle Murge sud orientali.- Rivista Italiana di Paleontologia e Stratigrafia, vol. 84, p. 561-666.
Matte P., Tapponnier P., Arnaud N., Bourjot L., Avouac J.P., Vidal P., Liu Q., Pan Y.S. & Wang Y. (1996).- Tectonics of Western Tibet, between the Tarim and the Indus.- Earth and Planetary Science Letters, vol. 142, p. 311-330.
Mattern F. & Schneider W. (2000).- Suturing of the Proto- and Paleo-Tethys oceans in the West Kunlun (Xijiang, China).- Journal of Asian Earth Sciences, vol. 18, p. 637-650.
Mattern F., Schneider W., Li Y. & Li X. (1996).- A traverse through the West Kunlun (Xinjiang, China): tentative geodynamic implications for the Paleozoic and Mesozoic.- Geologische Rundschau, vol. 85, p. 705-722.
Maync W. (1954).- The genus Navarella Ciry and Rat, 1951, in the Maastrichtian of Switzerland.- Contributions from the Cushman Foundation for Foraminiferal Research, vol. v, part 3, p. 138-144.
Mohanti M. & Pons J.M. (1999).- The rudist Bournonia mutabilis (Stoliczka, 1871) redescription based on new material from the Ottakkovil Formation (Maastrichtian), Ariyalur Region, South India.- Geobios, vol. 32, no. 6, p. 853-859.
Molnar P. & Tapponnier P. (1975).- Cenozoic tectonics of Asia: Effects of a continental collision.- Science, vol. 189, no. 4201, p. 419-426.
Morris N.J. & Skelton P.W. (1995).- Late Campanian - Maastrichtian rudists from the United Arab Emirates - Oman border region.- Bulletin of the British Museum (Natural History) (Geology Series), vol. 51, p. 277-305.
Munier-Chalmas H. (1873).- Prodr¶me d'une classification des rudistes.- Journal de Conchyliologie, vol. 13, p. 71-75.
Newell N.D. (1965).- Classification of the Bivalvia.- American Museum Novitates, no. 2206, p. 1û25.
Norin E. (1946).- Geological Exploration in Western Tibet: Report on Sino-Swedish Expedition 29.- Sweden Aktiebolaget Thule, Stockholm, 214 p.
Orbigny A. d' (1847).- Sur les brachiopodes ou palliobranches.- Comptes Rendus Hebdomadaires des SÚances de l'AcadÚmie des Sciences, vol. 25, p. 266û269.
Pan Y., Zheng D. & Zhang Q. (1992).- Introduction to integrated scientific investigation on Karakorum and Kunlun Mountains.- China Meterological Press, Beijing, p. 1û92.
Parona C.F. (1917).- Faune cretaciche del Caracorum e degli Altipiani tibetani.- Rendiconti della Reale Accademia dei Lincei, Classe di Scienze, matematiche e naturali (serie 5), vol. 26, no. 2, p. 53-57.
Parona C.F. (1921).- Fauna del neocretacico della Tripolitania.- Memorie per servire alla Descrizione della Carta geologica d'Italia, vol. 8, p. 1û21.
Parona C.F. (1928).- Faune Cretaciche del Caracorum. In: Fossa Mancini E., Parona C.F. & Stefanini G. (Eds.), Fossili del Secondario e del Terziario. Relazioni Scientifiche della Spedizione Italiana De Filippi nell'Himalaia, Caracorum e Turchestan Cinese, serie II (Resultati geologici e geografici), Vol. 6.- Zanichelli, Bologna, 344 p.
Pejovic D. (1957).- Geologie und Tektonik der weiteren Umgebung von Pocuta (Westserbien) mit besonderer Ber³cksichtung der Biostratigraphie der oberkretazischen Bildungen.- Posebna Izdana, Geoloski Institut "Jovan Zujovic", vol. 8, 147 p.
Pons J.M. & Vicens E. (2008).- The structure of the outer shell layer in radiolitid rudists, a morphoconstructional approach.- Lethaia, vol. 41, no. 3, p. 219-234.
Pons J.M., Vicens E., Chikhi-Aouimeur F. & Abdallah H. (2012).- Taxonomical and biostratigraphical significance of the North African radiolitid rudist bivalve Praeradiolites biskraensis (Coquand, 1880).- Palaeontology, vol. 55, no. 3, p. 677-695.
Radoičić R., Radulović V., Rabrenović D. & Radulović B. (2010).- The age of the brachiopod limestones from Guča, western Serbia.- Annales gÚologiques de la PÚninsule Balkanique, vol. 71, p. 73û93.
Rao X. (2019).- Biradiolites from the Yigeziya Formation of the Southwestern Tarim Basin.- Open Journal of Geology, vol. 9, no. 10, p. 562-565.
Rao X., Skelton P.W., Sano S.I., Li C., Pan Y., Luo H., Cai H., Peng B. & Sha, J. (2017).- Evolution and palaeogeographical dispersion of the radiolitid rudist genus Auroradiolites (Bivalvia, Hippuritida) with descriptions of new material from Tibet and archived specimens from Afghanistan.- Papers in Palaeontology, vol. 3, p. 297-315.
Rao X., Skelton P.W., Sano S.I., Zhang Y., Zhang Y., Pan Y., Cai H., Peng B., Zhang T. & Ma Z. (2020).- Shajia, a new genus of polyconitid rudist from the Langshan formation of the Lhasa block, Tibet, and its palaeogeographical implications.- Cretaceous Research, vol. 105, p. 104-151.
Rao X., Skelton P.W., Sha J., Cai H. & Iba Y. (2015).- Mid-Cretaceous rudists (Bivalvia: Hippuritida) from the Langshan Formation, Lhasa block, Tibet.- Papers in Palaeontology, vol. 1, no. 4, p. 401-424.
Sanchez V. (1981).- Hippuritidae y Radiolitidae (Bivalvia). Catalogo de especies.- Universidad Autonoma de Barcelona, Publicaciones Geol., vol. 15, 228 p.
Schlagintweit F., Frijia G. & Parente M. (2016).- Sarmentofascis zamparelliae n. sp., a new demosponge from the lower Campanian of southern Italy.- Cretaceous Research, vol. 57, p. 157-164.
Scott R.W., Wan X., Sha J. & Wen S.X. (2010).- Rudists of Tibet and the Tarim Basin, China: significance to Requieniidae phylogeny.- Journal of Paleontology, vol. 84, no. 3, p. 444-465.
Sengor A.M.C., Altiner D., Cin A., Ustaomer T. & Hsu K.J. (1988).- Origin and assembly of the Tethyside orogenic collage at the expense of Gondwana Land.- Geological Society London Special Publications, vol. 37, p. 119-181.
Serra-Kiel J., Vicedo V., Baceta J.I., Bernaola G. & Robador A. (2020).- Paleocene Larger Foraminifera from the Pyrenean Basin with a recalibration of the Paleocene Shallow Benthic Zone.- Geologica Acta, vol. 18.8, 69 + III p.
Sha J.G. (1998).- Characteristics of stratigraphy and palaeontology of Hohxil, Qinghai: Geographic significance.- Acta Palaeontologica Sinica, vol. 37, no. 1, p. 85-96 [in Chinese with English abstract].
Sha J.G. (2018).- Fossils witnessed the uplift of Tibetan Plateau. In: Rong J.Y., Yuan X.L., Zhan R.B. & Deng T. (Eds.), Biological evolution and environment.- Heifei, University of Sciences & Technology of China Press, p. 298-329 [in Chinese].
Sha J.G. & Cestari R. (2016).- Late Aptian-Albian Yasin-type rudist assemblage in the Himalayas: Palaeobiogeographic implications.- Cretaceous Research, vol. 65, p. 34-47.
Sha J.G. & F³rsich F.T. (1999).- Palaeotethys Ocean closed before Capitanian times in Hohxil area (W China): New data on the temporal extend of the Palaeotethys.- Neues Jahrbuch f³r Geologie und Palõontologie Monatshefthe, vol. 1999, no. 7, p. 440-448.
Sha J.G., Johnson A.L. & F³rsich F.T. (2004).- From deep-sea to high mountain ranges: Palaeogeographic and biotic changes in Hohxil, the source area of the Yangtze River (Tibet Plateau) since the Late Palaeozoic.- Neues Jahrbuch fur Geologie und Palaontologie Abhandlungen, vol. 233, no. 2, p. 169-195.
Sino-UK Geological Expedition Team (1990).- Geological evolution in the Tibetan Plateau.- Science Press, Beijing, 415 p.
Skelton P.W. (2013).- Rudist classification for the revised Bivalvia volumes of the 'Treatise on Invertebrate Paleontology'.- Caribbean Journal of Earth Science, vol. 45, p. 9-33.
Stefanini G. (1917).- Echinidi mesozoici del Caracorum raccolti dalla "Spedizione italiana nell'Asia Centrale (1913-1914).- Rendiconti della Reale Accademia dei Lincei, Classe di Scienze, matematiche e naturali (serie 5), vol. 26, p. 49-50.
Stefanini G. (1928).- Echinidi mesozoici del Caracor¨m. In: Fossa Mancini E., Parona C.F. & Stefanini G. (Eds.), Fossili del Secondario e del Terziario. Relazioni Scientifiche della Spedizione Italiana De Filippi nell'Himalaia, Caracorum e Turchestan Cinese, serie II (Resultati geologici e geografici), Vol. 6.- Zanichelli, Bologna, 344 p.
Steuber T. (1999).- Cretaceous rudists of Boeotia, central Greece.- Special Papers in Palaeontology, vol. 61, 229 p.
Steuber T., Korbar T., Jelaska V. & Gusić I. (2005).- Strontium isotope stratigraphy of Upper Cretaceous platform carbonates of the island of Brac (Adriatic Sea, Croatia): implications for global correlation of platform evolution and biostratigraphy.- Cretaceous Research, vol. 26, p. 741-756.
Steuber T. & L÷ser H. (2000).- Species richness and abundance patterns of Tethyan Cretaceous rudist bivalves (Mollusca: Hippuritacea) in the central-eastern Mediterranean and Middle East, analysed from a palaeontological database.- Palµogeography, Palµeoclimatology, Palµoecology, vol. 162, p. 75-104.
Steuber T., Scott R.W., Mitchell S.F. & Skelton P.W. (2016).- Treatise on Invertebrate Paleontology, Part N, Revised, Volume 1, Chapter 26C: Stratigraphy and diversity dynamics of Jurassic-Cretaceous Hippuritida (rudist bivalves).- Treatise Online, no. 81, p. 1-17.
Stolizcka F. (1871).- Cretaceous fauna of southern India.- Palaeontologia indica, vol. 3, 537 p.
Sun D.L. & Zhang B.G. (1983).- Aspects of the marine Cretaceous of China.- Cretaceous Research, vol. 4, p. 145-158.
Tapponnier P. & Molnar P. (1977).- Active faulting and tectonics of China.- Journal of Geophysical Research, vol. 82, p. 2905û2930.
Troya-GarcÝa L. (2016).- Rudistas (Hippuritida, Bivalvia) del Cenomaniense-Coniaciense (Cretßcico superior) del Pirineo meridional-central paleontologÝa y bioestratigrafÝa.- PhD Thesis, Universitat Aut‗noma de Barcelona, 519 p.
Villalonga R., Boix C., Frijia G., Parente M., Bernaus J.M. & Caus E. (2019).- Larger foraminifera and strontium isotope stratigraphy of middle Campanian shallow-water lagoonal facies of the Pyrenean Basin (NE Spain).- Facies, vol. 65, no. 2, article 20, 23 p.
Wen S.X. (1998).- Palaeontological aspects in Karakorum and Kunlum region. In: Comprehensive Scientific Expedition to the Tibetan Plateau, Chinese Academy of Sciences, Palaeontology of the Karakorum-Kunlun Mountains.- Science Press, Beijing, p. 15-26 [in Chinese with English abstract].
Wen S.X. (1999).- Cretaceous bivalve biogeography in Qinghai-Xizang Plateau.- Acta Palaeontologica Sinica, vol. 38, p. 1-30.
Wen S.X., Sha J.G., Zhang B.G. & Cai H.W. (2000b).- Marine Cretaceous. In: Nanjing Institute of Geology and Palaeontology Academia Sinica (Ed.), Stratigraphical Studies in China (1979-1999).- China University of Science and Technology Press, Hefei, p. 329-346 [in Chinese].
Wen S.X., Sun D.L., Yin J.X., Chen T.E. & Luo H. (1998).- Stratigraphic outline In Karakorum and Kunlum region. In: Comprehensive Scientific Expedition to the Tibetan Plateau, Chinese Academy of Sciences, Palaeontology of the Karakorum-Kunlun Mountains.- Science Press, Beijing, p. 5-13 [in Chinese with English abstract].
Wen S.X., Sun D.L., Yin J.X., Chen T.E. & Luo H. (2000a).- Stratigraphy. In: Comprehensive Scientific Expedition to the Tibetan Plateau, Chinese Academy of Sciences, Geological evolution of the Karakorum-Kunlun Mountains.- Science Press, Beijing, p. 6-92 [in Chinese].
Windley B.F. (1988).- Tectonic framework of the Himalaya, Karakoram and Tibet, and problems of their evolution.- Philosophical Transactions of the Royal Society of London A, vol. 326, p. 3-16.
Wittlinger G., Vergne J., Tapponnier P., Farra V., Poupinet G., Jiang M., Su H., Herquel G. & Paul A. (2004).- Teleseismic imaging of subducting lithosphere and Moho offsets beneath western Tibet.- Earth and Planetary Science Letters, vol. 221, p. 117-130.
Xi D., Wan X., Li G. & Li G. (2019).- Cretaceous integrative stratigraphy and timescale of China.- Science China Earth Sciences, vol. 62, p. 256-286.
Xiao W.J., Windley B.F., Liu D.Y., Jian P., Liu C.Z., Yuan C. & Sun M. (2005).- Accretionary tectonics of the West Kunlun orogen, China: a Paleozoic-Early Mesozoic, long-lived active continental margin with implications for the growth of southern Eurasia.- The Journal of Geology, vol. 113, p. 687-705.
Yang W., Xie Y., Fu S., Ren Y., Zhou J., Wang X., Zhang J., Yin D. & Zhao S. (2014).- The Tianshuihai leadûzinc deposit, Xinjiang province, NW China: A successful case of multi-scale geochemical mapping.- Journal of Geochemical Exploration, vol. 139, p. 136-143.
Yin A. & Harrison T.M. (2000).- Geologic evolution of the Himalayan-Tibetan orogen.- Annual Review of Earth and Planetary Sciences, vol. 28, p. 211-280.
Youngun Y. & Hs³ K. (1994).- Origin of the Kunlun Mountains by arc-arc and arc-continent collisions.- Island Arc, vol. 3, p. 75-89.
Zakhera M.S. (2011).- Cenomanian-Turonian rudists from western Sinai, Egypt: Systematic paleontology and paleoecology.- Geobios, vol. 44, no. 4, p. 409-433.
Zanchi A. & Gaetani M. (2011).- The geology of the Karakoram range, Pakistan: the new 1:100,000 geological map of Central-Western Karakoram.- Italian Journal of Geosciences, vol. 130, p. 161û262.
Zhang C.L., Zou H.B., Ye X.T. & Chen X.Y. (2019).- Tectonic evolution of the West Kunlun Orogenic Belt along the northern margin of the Tibetan Plateau: Implications for the assembly of the Tarim terrane to Gondwana.- Geoscience Frontiers, vol. 10, no. 3, p. 973-988.