◄ Carnets Geol. 20 (14) ►
![]()
Outline:
[Introduction]
[Historical background and taxonomical problems]
[Material studied]
[Systematic micropaleontology]
and ...
[Bibliographic references]
Lerchenauerstr. 167, 80935 Munich (Germany)
Babeş-Bolyai University,
Department of Geology and Center for Integrated Geological Studies, Str. M. Kogălniceanu
1, 400084 Cluj-Napoca (Romania)
World Foraminifera Database, 80 rue Jean Parot, 42100
Saint-Étienne (France)
Published online in final form (pdf) on September 21, 2020
DOI 10.2110/carnets.2020.2014
![]()
[Editor: Bruno Granier;
language editor: Simon Mitchell]
![]()
Orbitolinopsis flandrini Moullade, 1960, was described from Lower Cretaceous Urgonian limestones of southeastern France without the designation of a holotype. It represents a valid taxon since the selection of a holotype is only obligatory for taxa established after 1999 (ICZN, 4th edition, article 72.3). Today the original description is considered to be a mixture of several taxa belonging to Orbitolinopsis Henson, 1948/Cribellopsis Arnaud-Vanneau, 1980 (transverse sections with cupules/septules in the central zone) and Drevennia Arnaud-Vanneau, 1980 (axial sections with an axial "columella-like" column). The short and in any case insufficient original description of O. flandrini, however, stressed the presence of a columellar-like central test as the main characteristic feature. In fact, the majority of the (sub)axial sections showing an axial column (lacking in Orbitolinopsis) are considered to belong to Drevennia (Family Pfenderinidae). Therefore, the new combination Drevennia flandrini (Moullade, 1960) is proposed and a lectotype herein designated from the original illustrations. Drevennia ecougensis, the type-species of the genus, is found to be a subjective junior synonym of the latter. New finds from Serbia and Spain extend the stratigraphic range of D. flandrini, from the upper Berriasian to the lowermost upper Aptian. The first occurrences of Drevennia, Dobrogelina Neagu, 1979, Pfenderina Henson, 1948, and Moulladella Bucur & Schlagintweit, 2018, in the upper Berriasian give evidence for an adaptive radiation of the Pfenderinidae during the time reported and from other, mostly complex larger benthic foraminifera (e.g., Orbitolinidae).
• Foraminifera;
• Orbitolinidae;
• Pfenderinidae;
• taxonomy;
• ICZN;
• Lower
Cretaceous
Schlagintweit F., Bucur I.I. & Le Coze F. (2020).- Lectotype designation for Orbitolinopsis flandrini Moullade, 1960 (Foraminifera): The missing piece of a taxonomic puzzle.- Carnets Geol., Madrid, vol. 20, no. 14, p. 273-282.
Sélection du lectotype d'Orbitolinopsis flandrini Moullade, 1960 (Foraminifère) : La pièce manquante d'un puzzle taxinomique.- Orbitolinopsis flandrini Moullade, 1960, a été décrit dans les calcaires urgoniens (Crétacé inférieur) du sud-est de la France. La validité de ce taxon n'est pas en question, la désignation d'un holotype n'étant obligatoire que pour les taxons décrits après 1989 (CINZ, 4e édition, article 72.3). Aujourd'hui, la description originale de cette espèce est considérée comme fondée sur un mélange de taxons rapportés aux genres Orbitolinopsis Henson, 1948, Cribellopsis Arnaud-Vanneau, 1980 (sections transversales à cupules/septules dans la partie centrale) et Drevennia Arnaud-Vanneau, 1980 (sections axiales avec une colonne axiale "semblable à une columelle"). Toutefois cette description originale, courte et donc insuffisante, soulignait la présence d'une partie centrale du test semblable à une "columelle" comme étant la caractéristique distinctive de l'espèce. De fait, la majorité des sections (sub-) axiales montrant une colonne axiale (absente chez les Orbitolinopsis) sont considérées comme appartenant au genre Drevennia (Famille des Pfenderinidae). Par conséquent, la nouvelle combinaison Drevennia flandrini (Moullade, 1960) est proposée et un lectotype est sélectionné à partir des illustrations originales. Drevennia ecougensis, l'espèce-type du genre, est mise en synonymie avec D. flandrini, cette dernière espèce ayant priorité sur D. ecougensis (synonyme subjectif plus ancien). De nouvelles découvertes en Serbie permettent d'élargir la répartition stratigraphique de D. flandrini du Berriasien supérieur à l'Aptien inférieur. Les premières apparitions de Drevennia, Dobrogelina Neagu, 1979, Pfenderina Henson, 1948, et Moulladella Bucur & Schlagintweit, 2018, au Berriasien supérieur apportent la preuve d'une radiation adaptative des Pfenderinidae à cette époque, phénomène déjà noté pour d'autres types de foraminifères benthiques principalement chez les grands foraminifères complexes (e.g., Orbitolinidae).
• Foraminifères ;
• Orbitolinidae ;
• Pfenderinidae ;
• taxinomie ;
• CINZ ;
• Crétacé inférieur
Urgonian (upper Hauterivian-lower Aptian) shallow-water carbonates of southeastern France display a high diversity of Orbitolinidae. Clavel (2014: Fig. 5) reported 38 species belonging to the subfamilies Orbitolininae (6), Dictyoconinae (21), and Praedictyorbitolininae (11). The genera with the most species are Cribellopsis Arnaud-Vanneau, 1980, Orbitolinopsis Henson, 1948, and Paracoskinolina Moullade, 1965. Depending on the importance of structural elements for generic or specific rank among different workers (e.g., Moullade, 1963; Schroeder, 1964), many of these taxa were, and are still insufficiently understood. One example refers to Orbitolinopsis flandrini Moullade, 1960, initially described from the Barremian of south-eastern France. It has not been included in the compilation of Clavel et al. (2014), because its assignment to Orbitolinopsis has been questioned by several authors since its original description. Based on a literature study and new finds from upper Berriasian shallow-water carbonates of Serbia, the problems related to understanding this taxon are presented herein.
In 1960, Moullade described Orbitolinopsis flandrini n.sp. from the Barremian of the Drôme department in southeastern France. Based on the test morphology, Moullade (1960: p. 190) distinguished a cylindro-conical Form A (op.cit.: Pl. 2, figs. 8-20) and conical Form B (op.cit.: Pl. 2, figs. 21-28). In his rather short description, a reduced central zone similar to a columella (solid reticulum) was stressed as characteristic for the species. Neither a holotype nor a type-locality was indicated. This procedure was in line with the requirements of the International Code of Zoological Nomenclature (ICZN) at that time. It was only with the 4th edition published in the year 2000 that the nomination of a type (holotype or syntype) to be designated in the original publication (established after 1999) became obligatory (article 72.3). This means that O. flandrini was validly published and does not represent a nomen nudum. Two years later, Bassoullet and Moullade (1962) differentiated the subspecies Orbitolinopsis flandrini flandrini for the Form A (mean ratio height/diameter = 2), and Orbitolinopsis flandrini inflatus for the Form B (mean ratio height/diameter 1.4) of Moullade (1960). In the monograph of Moullade (1966: p. 42), both subspecies were again mentioned but for O. flandrini flandrini, some of the original illustrations (Pl. 2, figs. 8, 14-17 in Moullade, 1960) were omitted. The specimen illustrated in figure 17 of Plate 2 of Moullade (1960) was assigned to O. flandrini elongatus Dieni et al., 1963, which was subsequently revised by Arnaud-Vanneau (1980) as Cribellopsis elongata.
One of the first workers commenting on O.
flandrini was Hofker (1966). According to him (op.
cit.: p. 900), O.
flandrini Form B might represent juvenile specimens of Orbitolinopsis kiliani
Henson, 1948. Concerning Form A (= O.
flandrini flandrini), Hofker stated that the "structure is not
quite clear", but the horizontal sections (Moullade,
1960:
Pl. 2, figs.
18-20) "certainly show features of Orbitolinopsis". Arnaud-Vanneau
(1980: p. 700) treated O.
flandrini as an orbitolinid species and attributed it to an undetermined genus
("genres indéterminés d'Orbitolinidae"). In the monograph of the
Urgonian microfauna of southeastern France, Arnaud-Vanneau
(1980) also
established the new genus Drevennia
with the type-species D. ecougensis.
It is worth mentioning that any resemblances with O. flandrini have not been envisaged or discussed by Arnaud-Vanneau
(1980). The possible identity of both taxa was first suggested by Becker
(1999, p. 415), who noted also the lack of a holotype for O. flandrini. As already remarked above, this is not an argument for
the invalidity of the species. According to Becker (1999), the specimens
illustrated by Moullade (1960) in figures 8-9, 11 and 14-16
of Plate 2 are
seemingly morphologically identical to D. ecougensis (Fig.
1.E-F ![]() ). On the other hand, those specimens
illustrated in figures 12-13 and 17-20 of Plate 2 (op.cit.) were interpreted as incertae
sedis. This conclusion is herein accepted. Becker also correctly noted the
lack of the typical central structure of Orbitolinopsis
with cupules (see Arnaud-Vanneau, 1980) in O. ? flandrini raising
doubts about its generic status. Furthermore, Becker (1999) concluded that an exact
definition of O. ? flandrini
is only possible by re-analyzing the original material of Moullade
(1960)
and the designation of a lectotype. Independently from the correct conclusions
of Becker (1999), the specimens illustrated by that author as O. ? flandrini, do in our
opinion not belong to the form described by Moullade (1960). The small
forms are spirally coiled throughout, without showing uniserial adult chambers
and an axial column, and are considered to belong to another genus (neither Drevennia
nor Orbitolinopsis). Finally, Clavel et al.
(2010: p. 6) followed the opinion of Arnaud-Vanneau
(1980) that the assignment of the species flandrini
to the genus Orbitolinopsis was
very doubtful ("très douteuse"). Concerning O.
inflatus (former Form B of Moullade,
1960), Clavel et
al. (2010) concluded that the specimens (here: transverse sections) belong
to different species and genera, namely Cribellopsis
sp. for the transverse sections illustrated in figures 26-28 of Plate 2 of Moullade
(1960) due to the presence of radial partitions in the marginal zone. The axial/subaxial
sections of Moullade (1960: Pl. 2, figs. 22-24) instead were interpreted
as belonging to Orbitolinopsis cuvillieri
due to the presence of cupules.
). On the other hand, those specimens
illustrated in figures 12-13 and 17-20 of Plate 2 (op.cit.) were interpreted as incertae
sedis. This conclusion is herein accepted. Becker also correctly noted the
lack of the typical central structure of Orbitolinopsis
with cupules (see Arnaud-Vanneau, 1980) in O. ? flandrini raising
doubts about its generic status. Furthermore, Becker (1999) concluded that an exact
definition of O. ? flandrini
is only possible by re-analyzing the original material of Moullade
(1960)
and the designation of a lectotype. Independently from the correct conclusions
of Becker (1999), the specimens illustrated by that author as O. ? flandrini, do in our
opinion not belong to the form described by Moullade (1960). The small
forms are spirally coiled throughout, without showing uniserial adult chambers
and an axial column, and are considered to belong to another genus (neither Drevennia
nor Orbitolinopsis). Finally, Clavel et al.
(2010: p. 6) followed the opinion of Arnaud-Vanneau
(1980) that the assignment of the species flandrini
to the genus Orbitolinopsis was
very doubtful ("très douteuse"). Concerning O.
inflatus (former Form B of Moullade,
1960), Clavel et
al. (2010) concluded that the specimens (here: transverse sections) belong
to different species and genera, namely Cribellopsis
sp. for the transverse sections illustrated in figures 26-28 of Plate 2 of Moullade
(1960) due to the presence of radial partitions in the marginal zone. The axial/subaxial
sections of Moullade (1960: Pl. 2, figs. 22-24) instead were interpreted
as belonging to Orbitolinopsis cuvillieri
due to the presence of cupules.
|
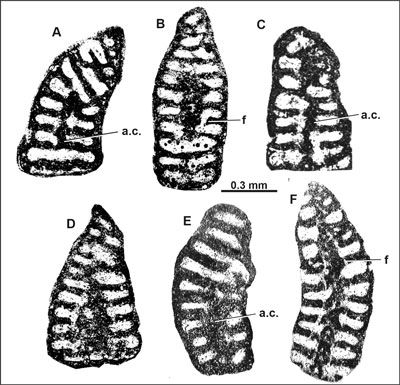
Figure 1:
Drevennia flandrini (Moullade) comb. nov. (A-D:
from Moullade,
1960: Pl. 2, fig. 16 = lectotype
designated herein, figs. 8, 14-15 = paralectotypes, Barremian of southeastern
France; E-F: Drevennia ecougensis from Arnaud-Vanneau,
1980: Pl. 84, figs. 10-11. Abbreviations: a.c. = axial column, f = foramen. |
In summary, in accordance with the observations of previous workers, we conclude that the initial description of O. flandrini Moullade represents a mixture of different species/genera. The axial sections displaying seemingly identical test morphologies, dimensions (see Dimensions in the chapter Systematic description), internal structure, namely the "columellaire central zone" (excluding its belonging to Orbitolinopsis), are considered identical to the form described by Arnaud-Vanneau (1980) twenty years later as Drevennia ecougensis (see Becker, 1999). The transverse sections are considered as belonging to Cribellopsis and Orbitolinopsis (see Hofker, 1966; Clavel et al., 2010) (Table 1).
Table 1: Historical summary.
| Moullade, 1960 | Bassoullet & Moullade, 1962 | Moullade, 1966 | Arnaud-Vanneau, 1980 | Becker, 1999 | Clavel et al., 2010 | This work |
| O.
flandrini
n.sp. Forme A: Pl. 2, figs. 8-20 |
O. flandrini flandrini n.subsp. | O.
flandrini flandrini: Pl. 2, figs. 9-13, 18-20 (non figs. 8, 14-16) of 1960 |
O. ? flandrini: Pl. 2, figs. 9-10, 12, doubtful: Pl. 2, figs. 18-20 not flandrini: Pl. 2, figs. 8, 11, 13-17 |
different taxa: Pl. 2, figs. 8-9, 11, 14-16 might be Drevennia; not determinable: Pl. 2, figs. 12-13, 17-20 |
sharing opinion of Arnaud-Vanneau, 1980, as O. flandrini a doubtful orbitolinid | Drevennia
flandrini nov. comb.: Pl.
2, figs. 8-9, 11, 14-16 of
1960 Lectotype: Pl. 2, fig. 16 in Moullade, 1960 |
| O.
flandrini elongatus
Dieni et al.: Pl. 2, fig. 17 of 1960 |
||||||
| O. flandrini n.sp.
Forme B: Pl. 2, figs. 21-28 |
O. flandrini inflatus n.subsp. | O. flandrini inflata | O. ? inflata: Pl. 2, figs. 21-24 of 1960 |
not mentioned expressis verbis | Cribellopsis sp.: Pl. 2, figs. 25-28 |
sharing opinion of Clavel et al., 2010 |
| O. kiliani: Pl. 2, figs. 22-24 |
taxon inquierendum |
We recently studied a Lower
Cretaceous section from eastern Serbia where specimens of "O." flandrini
have been found (Bucur et al., 2019,
2020). The samples studied belong to a
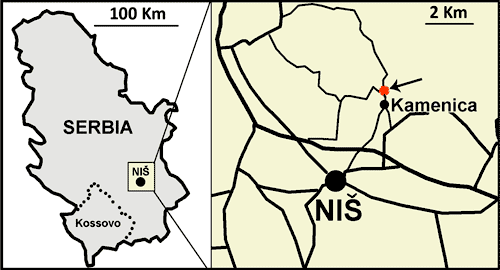
carbonate succession outcroping north-north-east of the city of Niš, in the vicinity
of the village of Kamenica (Fig. 2 ![]() ). The coordinates of the section base are N 43°23'03.7"
and E 21°56'41.6". The limestones from this area belong to the Kurilovo fold
structure (or anticline) (Krstić et al., 1978,
1980) that is part of the Gornjak-Stuva Planina Unit,
the westernmost part of the Carpatho-Balkanids of eastern Serbia. North of the
Danube River it continues with the Sasca Unit belonging to the Getic domain of
the South Carpathians (Săndulescu, 1975, as Sasca-Gornjak Unit). On
the geological map 1:100,000, sheet Aleksinac (Krstić et al.,
1978, 1980), the Lower Cretaceous carbonate deposits of the
Kamenica area are assigned to the Valanginian-Hauterivian and Barremian-Aptian.
New investigations have provided a Berriasian-pro parte Valanginian age for the
Cretaceous succession of the Kamenica section (Bucur et
al., 2019, 2020). It is the type-locality of the giant sized Bispiraloconulus
serbicus (Schlagintweit et al., 2019, for further information). The specimens of "Orbitolinopsis" flandrini
have been observed in the part of the section containing a diverse
assemblage of orbitolinids and other larger benthic foraminifera (e.g.,
spirocyclinids, pfenderinids, coscinoconids). The late Berriasian age is based
on the associated microfaunistic-microfloristic assemblage (see Associated
microfossils in following chapter), and rare calpionellids, such as Calpionellopsis
oblonga Colom, Lorenziella
plicata Remane or Lorenziella
hungarica Knauer & Nagy. Among the numerous Dasycladales,
we just want to mention the occurrence of Falsolikanella
campanensis (Azéma & Jaffrezo) (see also Radoičić,
1978), that represents an upper Berriasian marker taxon according to Granier
(2019). The orbitolinids will be the subject of a separate investigation. We
just name a few: examples: Cribellopsis
neoelongata (Cherchi & Schroeder), Urgonina alpillensis (Foury), Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy,
or O. debelmasi Moullade & Thieuloy.
The present study is based on 20 specimens (15 illustrated herein) of "Orbitolinopsis"
flandrini observed in 10 thin
sections. These are housed in the Geology Department of the Babeş-Bolyai
University, Cluj-Napoca (Ioan I. Bucur collection). One specimen
(Fig. 4.O
). The coordinates of the section base are N 43°23'03.7"
and E 21°56'41.6". The limestones from this area belong to the Kurilovo fold
structure (or anticline) (Krstić et al., 1978,
1980) that is part of the Gornjak-Stuva Planina Unit,
the westernmost part of the Carpatho-Balkanids of eastern Serbia. North of the
Danube River it continues with the Sasca Unit belonging to the Getic domain of
the South Carpathians (Săndulescu, 1975, as Sasca-Gornjak Unit). On
the geological map 1:100,000, sheet Aleksinac (Krstić et al.,
1978, 1980), the Lower Cretaceous carbonate deposits of the
Kamenica area are assigned to the Valanginian-Hauterivian and Barremian-Aptian.
New investigations have provided a Berriasian-pro parte Valanginian age for the
Cretaceous succession of the Kamenica section (Bucur et
al., 2019, 2020). It is the type-locality of the giant sized Bispiraloconulus
serbicus (Schlagintweit et al., 2019, for further information). The specimens of "Orbitolinopsis" flandrini
have been observed in the part of the section containing a diverse
assemblage of orbitolinids and other larger benthic foraminifera (e.g.,
spirocyclinids, pfenderinids, coscinoconids). The late Berriasian age is based
on the associated microfaunistic-microfloristic assemblage (see Associated
microfossils in following chapter), and rare calpionellids, such as Calpionellopsis
oblonga Colom, Lorenziella
plicata Remane or Lorenziella
hungarica Knauer & Nagy. Among the numerous Dasycladales,
we just want to mention the occurrence of Falsolikanella
campanensis (Azéma & Jaffrezo) (see also Radoičić,
1978), that represents an upper Berriasian marker taxon according to Granier
(2019). The orbitolinids will be the subject of a separate investigation. We
just name a few: examples: Cribellopsis
neoelongata (Cherchi & Schroeder), Urgonina alpillensis (Foury), Orbitolinopsis buccifer Arnaud-Vanneau & Thieuloy,
or O. debelmasi Moullade & Thieuloy.
The present study is based on 20 specimens (15 illustrated herein) of "Orbitolinopsis"
flandrini observed in 10 thin
sections. These are housed in the Geology Department of the Babeş-Bolyai
University, Cluj-Napoca (Ioan I. Bucur collection). One specimen
(Fig. 4.O ![]() )
is
from the lower upper Aptian (Gargasian) of the Reocin Formation, Cantabria,
northern Spain. For details on the geological setting the reader is referred to Schlagintweit
et al. (2016).
)
is
from the lower upper Aptian (Gargasian) of the Reocin Formation, Cantabria,
northern Spain. For details on the geological setting the reader is referred to Schlagintweit
et al. (2016).
|
Figure
2:
Location of the Kamenica section near the city of Niš
in eastern Serbia. |
The high-rank classification (Phylum-Class) follows Pawlowski et al. (2013). For the low-rank classification see Kaminski (2014). For a glossary of terms, see Hottinger (2006).
Phylum FORAMINIFERA Orbigny, 1826
Class Globothalamea Pawlowski et al., 2013
Order Loftusiida Kaminski & Mikhalevich in Kaminski, 2004
Suborder Orbitolinina Kaminski, 2004
Superfamily Pfenderinoidea Smout & Sugden, 1962, nom. correct. Kaminski, 2014 (pro Pfenderinacea)
Family Pfenderinidae Smout & Sugden, 1962
Subfamily Pfenderininae Smout & Sugden, 1962, nom. transl. Loeblich & Tappan, 1964
Remarks: Among the four genera comprising the Pfenderininae (see Septfontaine, 1988; Kaminski, 2014), Pfenderella Redmond, 1964, is reported from the Middle Jurassic, while the other three have their first appearances in the (upper) Berriasian: Dobrogelina Neagu, 1979, Pfenderina Henson, 1948 (see Granier, 2019), and Drevennia Arnaud-Vanneau, 1980 (this work). The paleopfenderinid Moulladella Bucur & Schlagintweit, 2019, also has its first appearance in the upper Berriasian documenting a radiation-diversification of this group, and other complex larger benthic foraminifera (e.g., Orbitolinidae) during this time.
Genus Drevennia Arnaud-Vanneau, 1980, emend. herein
Type species: Drevennia ecougensis Arnaud-Vanneau, 1980 = D. flandrini (Moullade, 1960), comb. nov.
Lectotype designation:
From the original illustrations of Orbitolinopsis
flandrini by Moullade (1960), the specimen illustrated in
figure 16 of Plate 2 is selected herein as lectotype. It represents an oblique axial
section of a bent test showing the solid column in the central test part of
some chambers. The lectotype is illustrated herein in Figure
1.A ![]() .
From the
further specimens illustrated by Moullade (1960), only figures
8-9 and 14-15 of Plate 2 belong to the same species as the lectotype
herein designated and are designated as paralectotypes (ICZN, Recommendation 74
F). The collection of Moullade's specimens is
deposited at the Muséum d'Histoire Naturelle de Nice and still awaits
curation (Granier, 2020, pers. comm.).
.
From the
further specimens illustrated by Moullade (1960), only figures
8-9 and 14-15 of Plate 2 belong to the same species as the lectotype
herein designated and are designated as paralectotypes (ICZN, Recommendation 74
F). The collection of Moullade's specimens is
deposited at the Muséum d'Histoire Naturelle de Nice and still awaits
curation (Granier, 2020, pers. comm.).
Diagnosis: Test elongate, cylindrical to cylindro-conical, may be slightly bent at the transition from the trochospiral to uniserial stages. Initial part trochospirally coiled about a central columella. Later uncoiled, with uniserial chambers broader than high and of almost constant height throughout ontogeny. The central part of the uniserial chambers is occupied by an axial column formed by stacked inverted cones protruding slightly into the basal part of the subsequent chamber. Few multiple foramina surround the axial column, and are straight to obliquely arranged. The central column may be transjected by a central canal. The thick wall is homogeneous microgranular to finely agglutinated.
Remarks: In the
original description, D. ecougensis
was poorly illustrated with three illustrations/specimens: one isolated specimen
(Arnaud-Vanneau, 1980: Pl. 55, fig. 8), and two from thin-sections
(op.cit.: Pl. 84, figs. 10-11, axial and axial-oblique sections). The latter two are re-illustrated
herein Fig.
1.E-F ![]() . Transverse sections were not provided. Additionally,
at least one specimen of Pseudolituonella
sp. (op.cit.: Pl. 84, fig. 9) is herein interpreted as a section of D.
ecougensis evidently not passing through the median test plane and therefore
not cutting the axial central column throughout. It is partly sectioned in the
latest chamber, showing also the surrounding cribrate foramina. The dimensions (chamber
height, thickness of wall, septa) also appear to be equal. The genus diagnosis
includes the presence of a more or less opaque columellar mass in the central
part of the chambers ("masse columellaire plus ou moins opaque ... dans la
partie central des loges", Arnaud-Vanneau, 1980: p. 569). Let us
remember that the term columella is typically reported from trochospirally
coiled forms as a "solid, trochospiral structure formed by the basal walls of
spiral chambers coalescing around the coiling axis" (Hottinger,
2006:
p. 11). Consequently, the term columella can only be applied to the early stage
of Drevennia with trochospirally
coiled chambers. In the uniserial stage this central (or axial) element occurs
inside the chambers and is comparable to the structure reported from upper
Cretaceous (lower Turonian) Axicolumella
Hercogová, 1988. This genus has uniserial chambers with
"an axial
column composed of segments, which are components of individual chambers" (Hercogová,
1988: p. 173), and "these segments have the shape of truncated cones standing
on the base with the smaller diameter" (remark: = widening in growth
direction). A comparable structure with constrictions is also present in Drevennia
and is visible in the holotype shown in figure 11 of Plate 84 of Arnaud-Vanneau
(1980) (Fig. 1.F
. Transverse sections were not provided. Additionally,
at least one specimen of Pseudolituonella
sp. (op.cit.: Pl. 84, fig. 9) is herein interpreted as a section of D.
ecougensis evidently not passing through the median test plane and therefore
not cutting the axial central column throughout. It is partly sectioned in the
latest chamber, showing also the surrounding cribrate foramina. The dimensions (chamber
height, thickness of wall, septa) also appear to be equal. The genus diagnosis
includes the presence of a more or less opaque columellar mass in the central
part of the chambers ("masse columellaire plus ou moins opaque ... dans la
partie central des loges", Arnaud-Vanneau, 1980: p. 569). Let us
remember that the term columella is typically reported from trochospirally
coiled forms as a "solid, trochospiral structure formed by the basal walls of
spiral chambers coalescing around the coiling axis" (Hottinger,
2006:
p. 11). Consequently, the term columella can only be applied to the early stage
of Drevennia with trochospirally
coiled chambers. In the uniserial stage this central (or axial) element occurs
inside the chambers and is comparable to the structure reported from upper
Cretaceous (lower Turonian) Axicolumella
Hercogová, 1988. This genus has uniserial chambers with
"an axial
column composed of segments, which are components of individual chambers" (Hercogová,
1988: p. 173), and "these segments have the shape of truncated cones standing
on the base with the smaller diameter" (remark: = widening in growth
direction). A comparable structure with constrictions is also present in Drevennia
and is visible in the holotype shown in figure 11 of Plate 84 of Arnaud-Vanneau
(1980) (Fig. 1.F ![]() ). Axicolumella belongs
to the family Thomasinellidae (see Kaminski, 2014) displaying a
distinctly agglutinated test with bifurcations. We state also some similarities
to the Maastrichtian-Paleocene trochospirally coiled Pachycolumella Septfontaine et al.,
2019. It possesses a conspicuous central columella appearing
as a helicoidal succession of inverted half-cones.
). Axicolumella belongs
to the family Thomasinellidae (see Kaminski, 2014) displaying a
distinctly agglutinated test with bifurcations. We state also some similarities
to the Maastrichtian-Paleocene trochospirally coiled Pachycolumella Septfontaine et al.,
2019. It possesses a conspicuous central columella appearing
as a helicoidal succession of inverted half-cones.
Other species: Drevennia is a monospecific genus.
Occurrences: Spain: Barremian and lower Aptian of Sierra de Montsec, Province Lerida-Huesca (Bassoullet and Moullade, 1962; Schroeder et al., 1982). Upper Barremian of Organyà Basin, Pre-Pyrenees, NE Spain (Bernaus, 2000). Upper Aptian Reocin Formation of northern Spain, previously unrecorded (Schlagintweit et al., 2016: Cuchia quarry section). France: Barremian and Lower Aptian? of southeastern France (Moullade, 1960; Arnaud-Vanneau, 1980).
|
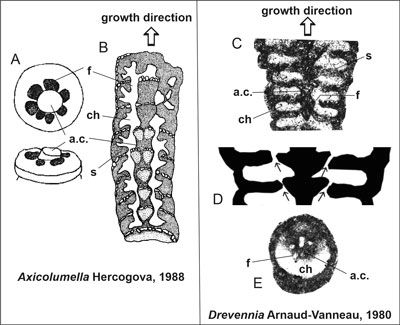
Figure
3:
Comparison of Axicolumella
Hercogová (lower Turonian) (A-B)
and Drevennia Arnaud-Vanneau
(Barremian) (C-E). A
Axicolumella cylindrica (Perner),
broken specimen showing open chamber with central axial column and attaching
cribrate foramina. B ibidem; schematic drawing of longitudinal section showing internal
structure (from Hercogová, 1988, Fig. 11, pars, and Fig. 12, pars,
modified). C Drevennia
flandrini (Arnaud-Vanneau) comb. nov., excerpt from
Fig. 4.E |
Drevennia flandrini (Moullade, 1960), comb. nov.
pars 1960 Orbitolinopsis flandrini n.sp. Forme A - Moullade, p. 190, Pl. 2, figs. 8-9, 14-16.
1980 Drevennia ecougensis n.sp. - Arnaud-Vanneau, p. 569, Pl. 55, fig. 8; Pl. 84, figs. 10-11.
1980 Pseudolituonella sp. - Arnaud-Vanneau, Pl. 84, fig. 9.
1982 Drevennia cf. ecougensis Arnaud-Vanneau - Schroeder et al., Pl. 3, fig. 6.
non 1999 Orbitolinopsis ? flandrini Moullade - Becker, p. 415, Pl. 16, figs. 9-13.
2005 Pseudolituonella gavonensis Foury - Polavder and Radulović, Pl. 1, fig. 7.
Description: Test free, elongate, high conical to
cylindroconical with acute apex, sometimes bent (Figs. 1.F ![]() ,
4.M
,
4.M ![]() ). The initial
part shows a high trochospiral coiling (~4 to ~6 whorls; Fig. 4.M
). The initial
part shows a high trochospiral coiling (~4 to ~6 whorls; Fig. 4.M ![]() ) about a
central columella (Fig. 4.A-C
) about a
central columella (Fig. 4.A-C ![]() ) that may display a central canal
(Fig. 4.K
) that may display a central canal
(Fig. 4.K ![]() ).
Details on the embryonic chambers are not known. The transition to the following
uniserial stage may be marked by a change in growth direction (Figs.
1.A, 1.F
).
Details on the embryonic chambers are not known. The transition to the following
uniserial stage may be marked by a change in growth direction (Figs.
1.A, 1.F ![]() ,
4.M
,
4.M ![]() ). The main test part consists of up to 10 discoidal chambers distinctly
broader than high, almost rectangular in axial sections. Transverse sections of this part
of the test are circular.
The
central part of the chambers is occupied by an axial column (calcitic mass).
Both, chambers and axial column slightly increase in width during ontogeny.
Depending on the plane of the section, the column may appear massive (Figs.
1.D
). The main test part consists of up to 10 discoidal chambers distinctly
broader than high, almost rectangular in axial sections. Transverse sections of this part
of the test are circular.
The
central part of the chambers is occupied by an axial column (calcitic mass).
Both, chambers and axial column slightly increase in width during ontogeny.
Depending on the plane of the section, the column may appear massive (Figs.
1.D ![]() ,
4.E, 4.G
,
4.E, 4.G ![]() ), display an indistinct central hollow (Fig. 4.D, 4.L
), display an indistinct central hollow (Fig. 4.D, 4.L ![]() ), or is pierced by an
undulating (spirally coiled?) canal (Figs. 1.F
), or is pierced by an
undulating (spirally coiled?) canal (Figs. 1.F ![]() ,
4.H, 4.J
,
4.H, 4.J ![]() ).
The column consists of stacked cones (cone-in-cone structure) widening in the growth
direction and slightly extending into the base of the succeeding chamber (Figs.
1.F
).
The column consists of stacked cones (cone-in-cone structure) widening in the growth
direction and slightly extending into the base of the succeeding chamber (Figs.
1.F ![]() ,
3.C-D
,
3.C-D ![]() ,
4.D-E
,
4.D-E ![]() ).
Multiple foramina are arranged straight or with an oblique axis closely around the
axial column (Fig. 3.D
).
Multiple foramina are arranged straight or with an oblique axis closely around the
axial column (Fig. 3.D ![]() ). Wall thick, microgranular to
finely agglutinating, appearing homogeneous.
). Wall thick, microgranular to
finely agglutinating, appearing homogeneous.
Dimensions:
test diameter: 0.4-0.6 mm (Arnaud-Vanneau, 1980: up to 0.42 mm)
test height: 0.7-1.1 mm (Moullade, 1960: always > 1.0 mm; Arnaud-Vanneau, 1980: up to 1.125 mm)
thickness septa: 0.030-0.045 mm
chamber height (adult): 0.11-0.14 mm
diameter foramina: 0.020-0.025 mm
wall thickness: 0.04-0.05 mm
number of chambers last 0.5 mm: 5 (rarely 6) (Moullade, 1960: 4-6*; Arnaud-Vanneau, 1980: 5-6* = measured from illustrations)
|
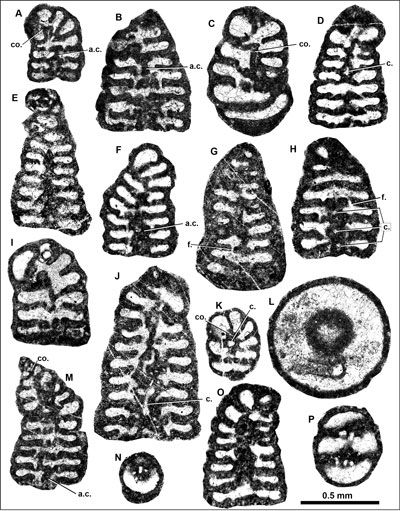
Figure
4:
Drevennia
flandrini (Moullade) comb. nov. from the upper Berriasian of the
Kamenica section, eastern Serbia (A-N,
P), and upper Aptian of the Cuchia quarry section, northern Spain (O). A,
C, I Oblique axial sections of juvenile specimens. B
Oblique section of the uncoiled adult part. C,
K Oblique sections, including trochospirally coiled part. D, F, G-H Oblique axial sections only partly transjecting the
trochospirally coiled part. E Axial
section (detail shown in Fig. 3.C |
Stratigraphy: The specimens illustrated herein from eastern Serbia are of late Berriasian age. The Barremian age of the sections (platform-basin transitions) from southeastern France studied by Moullade (1960, 1966) and containing D. flandrini is based on ammonites. Bassoullet and Moullade (1962) mention the species from the "Barremian" of Spain associated with orbitolinids that Schroeder et al. (1982: p. 916) recognized as O. praesimplex. According to Schroeder (1972), the latter species indicates an early Aptian age. The Reocin Formation of Cantabria, N-Spain is late Aptian in age (Schlagintweit et al., 2016, for details). Summarizing, the stratigraphic range of D. flandrini can be indicated as late Berriasian-earliest late Aptian (Gargasian).
Microfacies and palaeoenvironment of the Serbian material: The samples with D. flandrini are orbitolinid-bearing packstones/grainstones/wackestones with common remains of bivalves, gastropods, echinoids, and rare corals. The palaeoenvironment corresponds to an inner platform facies, which also includes oolithic shoals (see also Schlagintweit et al., 2019). The samples are characterized by a highly diverse assemblage of small and large-sized agglutinated and calcareous benthic foraminifers. Calcareous algae (dasycladalean and rarely also gymnocodiacean algae) may be present (Bucur et al., 2020, for details).
We kindly acknowledge the helpful comments and discussion with reviewers Michel Septfontaine and Danielle Decrouez. Thanks to Simon Mitchell who helped with the English text.
Arnaud-Vanneau A. (1980).- L'Urgonien du Vercors septentrional et de la Chartreuse.- Géologie Alpine, Mémoire, Grenoble, vol. 11 (3 volumes), 874 p.
Bassoullet J.-P. & Moullade M. (1962).- Les Orbitolinidae du Crétacé inférieur de la Sierra du Montsech, Province de Lerida (Espagne).- Revue de Micropaléontologie, Paris, vol. 5, no. 2, p. 104-114.
Becker E. (1999).- Orbitoliniden-Biostratigraphie der Unterkreide (Hauterive – Barrême) in den spanischen Pyrenäen (Profil Organyà, Prov. Lérida).- Revue de Paléobiologie, Genève, vol. 18, p. 359-489.
Bernaus J.M. (2000).- L'Urgonien du bassin d'Organyà (NE de l'Espagne).- Géologie Alpine, Mémoire, Grenoble, vol. 33, 138 p.
Bucur I.I. & Schlagintweit F. (2018).- Moulladella jourdanensis (Foury & Moullade, 1966) n. gen., n. comb.: Valanginian-early late Barremian larger benthic foraminifera from the northern Neotethyan margin.- Acta Palaeontologica Romaniae, Cluj Napoca, vol. 14, no. 2, p. 45-59.
Bucur I.I., Sudar M.N., Schlagintweit F., Polavder S., Săsăran E., Jovanović D. & Pleş G. (2019).- The lowermost Cretaceous from the western part of Carpatho-Balkanides (Eastern Serbia): The Kamenica section.- Abstracts, 12th Romanian Symposium of Paleontology, Cluj-Napoca (September 19-21, 2019), p. 16-17.
Bucur I.I., Sudar M., Schlagintweit F., Pleş G., Săsăran E., Jovanović D., Polavder S. & Radoičić R. (2020).- Lowermost Cretaceous limestones from the Kučaj zone (Carpatho-Balkanides, Eastern Serbia): New data on their age assignment.- Cretaceous Research, vol. 116, article 104575, 20 p.
Clavel B., Busnardo R., Charollais J., Conrad M. & Granier B. (2010).- Répartition biostratigraphique des orbitolinidés dans la biozonation à ammonites (plate-forme urgonienne du Sud-Est de la France. Partie 1 : Hauterivien supérieur-Barrémien basal.- Carnets Geol., Madrid, vol. 10, no. A06 (CG2010_A06), 56 p.
Clavel B., Charollais J., Busnardo R., Granier B., Conrad M.A., Desjacques P. & Metzger J. (2014).- La plate-forme carbonatée urgonienne (Hauterivien supérieur - Aptien inférieur) dans le Sud-Est de la France et en Suisse : Synthèse.- Archives des Sciences, Genève, vol. 67, no. 1, p. 1-97.
Dieni L., Massari F. & Moullade M. (1963).- Sur quelques Orbitolinidae des calcaires à facies "urgonien" du Crétacé inférieur des environs d'Orosei (Sardaigne).- Bolletino della Società Paleontologica Italiana, Milano, vol. 2, no. 2, p. 3-8.
Granier B. (2019).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition.- Cretaceous Research, vol. 93, p. 245-274.
Henson F.R.S. (1948).- New Trochamminidæ and Verneuilinidæ from the Middle East.- Annals and Magazine of Natural History (Series 11), London, vol. XIV [1947], p. 605-630.
Hercogová J. (1988).- Acruliammina, Bdelloidina and Axicolumella n.gen. (Foraminifera) from the Cretaceous transgressive sediments of the Bohemian Massif.- Sbornik geologických věd (Paleontologie), Praha, vol. 29, p. 145-189.
Hofker J. (1966).- Note on Orbitolinopsis kiliani (Prever).- Eclogae Geologicae Helvetiae, Basel, vol. 59, no. 2, p. 887-902.
Hottinger L. (2006).- Illustrated glossary of terms used in foraminiferal research.- Carnets Geol., Madrid, vol. 6, no. M02 (CG2006_M02), 126 p.
International Commission on Zoological Nomenclature (ICZN; Ride W.D.L, Cogger H.G., Dupuis C., Minelli A., Thompson F.C. & Tubbs P.K., Eds.) (1999).- International Code of Zoological Nomenclature. Fourth Edition.- International Trust for Zoological Nomenclature, London.
Kaminski M.A. (2004).- The Year 2000 Classification of the Agglutinated Foraminifera. In: Bubik M. & Kaminski M.A. (Eds.), Proceedings of the Sixth International Workshop on Agglutinated Foraminifera.- Grzybowski Foundation Special Publication, Krakow, vol. 8, p. 237-255.
Kaminski M.A. (2014).- The year 2010 classification of the agglutinated foraminifera.- Micropaleontology, New York, vol. 60, p. 89-108.
Krstić B., Rakić B., Veselinović M., Dolić D., Rakić M., Anđelković J. & Banković V. (1978).- Geological map of RSF Yougoslavia, 1:10000, sheet K 34-20 (Aleksinac).- Geological Institute, Beograd.
Krstić B., Veselinović M., Divljan M. & Rakić M. (1980).- Explanation to the map of RSF Yougoslavia, 1:10000, sheet K 34-20 (Aleksinac).- Geological Institute, Beograd [in Serbian].
Loeblich A.R. Jr & Tappan H. (1964).- Treatise on Invertebrate Paleontology, Part C: Protista 2, Sarcodina, chiefly "Thecamoebians" and Foraminiferida.- Geological Society of America, Boulder, and University of Kansas Press, Lawrence, xxxi + 900 p.
Loeblich A.R. Jr & Tappan H. (1987).- Foraminiferal genera and their classification.- Van Nostrand Reinhold, New York, 2 volumes, 970 p.
Moullade M. (1960).- Sur quelques Foraminifères du Crétacé inférieur des Baronnies (Drôme).- Revue de Micropaléontologie, Paris, vol. 2, p. 131-142.
Moullade M. (1963).- État actuel des connaissances sur les Orbitolinidae (Foraminifères) du Crétacé inférieur mésogéen.- Mémoires du Bureau de Recherches Géologiques et Minières, Orléans, no. 34, p. 513-523.
Moullade M. (1965).- Contribution au problème de la classification des Orbitolinidae (Foraminiferida, Lituolacea).- Comptes rendus hebdomadaires des Séances de l'Académie des Sciences, Paris, vol. 260, p. 4031-4034.
Moullade M. (1966).- Étude stratigraphique et micropaléontologique du Crétacé inférieur de la "fosse vocontienne".- Thèse, Université de Lyon, 369 p.
Neagu T. (1979).- Données nouvelles concernant les représentants de la Famille des Pfenderinidae de l'Éocrétacé de la Dobrogea méridionale (Roumanie).- Revista Española de Micropaleontología, Madrid, vol. 11, no. 3, p. 479-504.
Orbigny A. d' (1826).- Tableau méthodique de la classe des Céphalopodes.- Annales des Sciences Naturelles, Paris, no. 7, p. 245-314.
Pawlowski J., Holzmann M. & Tyszka J. (2013).- New supraordinal classification of Foraminifera: Molecules meet morphology.- Marine Micropaleontology, vol. 100, p. 1-10.
Polavder S. & Radulović B. (2005).- Microfossils and brachiopods from the Lower Barremian at Prekonozi, eastern Serbia.- Annales Géologiques de la Péninsule Balkanique, Beograd, vol. 66 (2004-2005), p. 55-61.
Radoičić R. (1978).- On distribution of Dasycladales in the Lower Cretaceous of Kurilovo, East Serbia.- Bulletin of the Natural History Museum in Belgrade, (Series A), t. 33, p. 183-197 [in Serbian, with English summary].
Redmond C.D. (1964).- The foraminiferal Family Pfenderinidae in the Jurassic of Saudi Arabia.- Micropaleontology, New York, vol. 10, no. 2, p. 251-263.
Săndulescu M. (1975).- Essai de synthèse structurale des Carpathes.- Bulletin de la Société géologique de France (7e série), Paris, vol. XXVII, no. 3, p. 299-358.
Schlagintweit F., Bucur I.I. & Sudar M. (2019).- Bispiraloconulus serbiacus n. gen., n.sp., a giant arborescent benthic foraminifera from the Berriasian of Serbia.- Cretaceous Research, vol. 93, p. 98-106.
Schlagintweit F., Rosales I. & Najarro M. (2016).- Glomospirella cantabrica n.sp., and other benthic foraminifera from Lower Cretaceous Urgonian-type carbonates of Cantabria, Spain.- Geologica Acta, vol. 14, no. 2, p. 113-138.
Schroeder R. (1964).- Kritische Bemerkungen zu den Orbitoliniden-Untersuchungen von M. Moullade.- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Stuttgart, Heft 7, p. 429-439.
Schroeder R. (1972).- Zwei neue Orbitolinidae (Foram.) der spanischen Kreide.- Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Heft 2, p. 108-119.
Schroeder R., Altenbach A., Brinnel P., Cherchi A. & Wallrabe H.-J. (1982).- El Barremiense marino de la Sierra de Montsec (Prov. Lerida-Huesca).- Cuadernos Geología Ibérica, Madrid, no. 8, p. 915-933.
Septfontaine M. (1988).- Vers une classification évolutive des Lituolidés (foraminifères) jurassiques en milieu de plate-forme carbonatée.- Revue de Paléobiologie, Genève, vol. spec. 2 (Benthos '86), p. 229-256.
Septfontaine M., Schlagintweit F. & Rashidi K. (2019).- Pachycolumella nov. gen., shallow-water benthic imperforate foraminifera and its species from the Maastrichtian and Paleocene of Iran.- Micropaleontology, New York, vol. 65, no. 2, p. 145-160.
Smout A.H. & Sugden KW. (1962).- New information on the foraminiferal genus Pfenderina.- Palaeontology, London, vol. 4, part 4, p. 581-591.