◄ Carnets Geol. 20 (18) ►
![]()
Outline:
[1. Introduction]
[2. Stratigraphy]
[3. Systematic palaeontology]
[4. Significance for the evolution of American hippuritids]
[Bibliographic references] and ...
[Appendix]
Department of Geography and Geology, The University of
the West Indies, Mona, Kingston 7 (Jamaica)
Published online in final form (pdf) on November 11, 2020
DOI 10.2110/carnets.2020.2018
![]()
[Editor:
Robert W. Scott; technical editor: Bruno Granier]
![]()
Exceptionally well-preserved (silicified) hippuritid rudists occur in the El Rayo Formation (lower Maastrichtian) of south-western Puerto Rico. Three species belonging to three different genera are present: Caribbea muellerreidi (Vermunt), Laluzia peruviana (Gerth) and Parastroma guitarti (Palmer). Acid digestion of the limestones has resulted in a collection with numerous three-dimensional left and right valves many with the preservation of the minute details of the pore system. The morphological features of each species are described, and many features are illustrated for the first time. The new material, coupled with descriptions from other studies, demonstrates that six genera of endemic hippuritids evolved in two separate radiations in the New World: an older radiation of forms that had pallial canals in their left valves (Barrettia, Whitfieldiella and Parastroma) and a younger radiation of forms lacking pallial canals in their left valves (Laluzia, Caribbea and Praebarrettia). The exquisite preservation also reveals that in these endemic New World hippuritids the sockets for the teeth consisted of slots into which ribs on the teeth fitted; this contrasts with Old World hippuritids that have true sockets formed from upfolds of the tabulae for the teeth. The distinctive morphology of the tooth sockets is here used to define a monophyletic subfamily for which the name Barrettiinae Chubb is available.
• exceptional preservation (silicification);
• rudist bivalves;
• Laluzia;
• Caribbea;
• Parastroma;
• Hippuritidae;
• Puerto Rico;
• Cretaceous
Mitchell S.F. (2020).- Exceptionally well-preserved silicified hippuritid rudist bivalves from the lower Maastrichtian of Puerto Rico.- Carnets Geol., Madrid, vol. 20, no. 18, p. 333-366.
Les rudistes (Bivalvia) silicifi�s et exceptionnellement bien conserv�s du Maastrichtien inf�rieur de Porto Rico.- Des rudistes hippuritid�s exceptionnellement bien conserv�s (silicifi�s) sont observ�s dans la Formation d'El Rayo (Maastrichtien inf�rieur) du sud-ouest de Porto Rico. Trois esp�ces appartenant � trois genres diff�rents sont repr�sent�es : Caribbea muellerreidi (Vermunt), Laluzia peruviana (Gerth) et Parastroma guitarti (Palmer). La dissolution par de l'acide des matrices calcaires a fourni une collection de nombreuses valves senestres et dextres en trois dimensions, dont de nombreuses pr�sentant les menus d�tails du syst�me de pores. Les caract�ristiques morphologiques de chaque esp�ce sont d�crites, et nombre de ces traits sont illustr�s pour la premi�re fois. Ce nouveau mat�riel, associ� aux descriptions pr�sentes dans d'autres �tudes, montre que six genres d'hippuritid�s end�miques ont �volu� en deux radiations distinctes dans le Nouveau Monde : une radiation plus ancienne avec des formes qui comportent des canaux pall�aux dans la valve senestre (Barrettia, Whitfieldiella et Parastroma) et une radiation plus r�cente de formes d�pourvues de canaux pall�aux dans cette m�me valve senestre (Laluzia, Caribbea and Praebarrettia). La d�licate conservation montre �galement que, chez ces hippuritid�s end�miques du Nouveau Monde, les alv�oles pour les dents consistaient en des fentes dans lesquelles les c�tes des dents s'ajustaient ; cela contraste avec les hippuritid�s de l'Ancien Monde qui pr�sentent de v�ritables alv�oles constitu�es par les replis des tabulae pour les dents. La morphologie unique de ces alv�oles dentaires est utilis�e ici pour d�finir une sous-famille monophyl�tique pour laquelle le nom de Barrettiinae Chubb est disponible.
• conservation exceptionnelle
(silicification) ;
� bivalves rudistes ;
� Laluzia ;
� Caribbea ;
� Parastroma ;
� Hippuritidae ;
� Porto Rico ;
� Cr�tac�
Multiple-folded hippuritid rudists, exemplified by Barrettia Woodward, 1862, are amongst the most distinctive fossils of the tropical Upper Cretaceous of the New World, and many different forms have now been described. Yet, the lack of complete morphological descriptions, particularly the details of the left valve (LV) construction and its pore system have led to difficulties in understanding the relationships between the various forms as well as their relations with other hippuritids of the Americas and the Old World. Although the pore systems are now known (to a certain degree - although most have suffered significantly from some amount of weathering) in most forms (e.g., Palmer, 1933; Dommelen, 1971; Mitchell, 2010; Munujos et al., 2016, 2020), other details of the construction of the left valve have only been reconstructed using cut sections, which, because of their complexity, are often difficult to interpret. Norman Sohl's studies of New World Cretaceous gastropods (e.g., Sohl & Kollmann, 1985; Sohl, 1992, 1998) led him to collect vast quantities of material from Puerto Rico, much of which resides in the United States National Museum collection at the Smithsonian Institution. Amongst this material is a large collection of silicified specimens from the El Rayo Formation of Puerto Rico. In 2009, I was invited to the Smithsonian to study the rudist material and was able to carry out a detailed investigation of this material. This unique collection includes exceptionally preserved specimens of rudist bivalves with the shell partially or completely silicified. It presents an unrivalled opportunity to understand the morphology of various rudists, particularly the hippuritids which are represented by three species belonging to three different genera. This paper presents the results of that study.
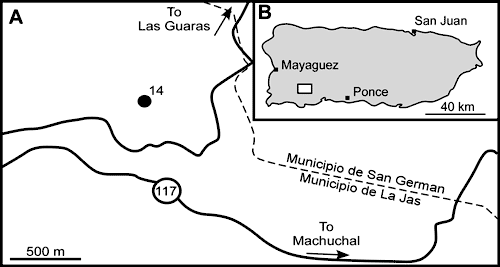
The collection site ('Locality 14') of the rudist (and gastropod)
material from the El Rayo Formation in the Sabana Grande Quadrangle was
described by Sohl and Kollmann
(1985), Sohl (1992) and in
the posthumous publication by Sohl (1998, p. 24). These descriptions are
similar. The locality (Fig. 1
![]() ) is described as being situated on a 125-m-high
hill (1985 and 1998 papers) or 147-m-high hill
(1992 paper) east of Quebrada
Jicara, 4.75 km S-SW of Sabana Grande, Barrio Lajas Arriba, Municipio de Lajas (approximately
Puerto Rico metre grid 23,900 N; 93,400 to 93,600 E), and has the USGS Mesozoic
locality numbers 28664, 28748, 29075, 29088 and 29361. The material was
collected by N.F. Sohl, E.G. Kauffman and W.O. Ross in 1963
and 1964 (Sohl, 1992, p. 433).
) is described as being situated on a 125-m-high
hill (1985 and 1998 papers) or 147-m-high hill
(1992 paper) east of Quebrada
Jicara, 4.75 km S-SW of Sabana Grande, Barrio Lajas Arriba, Municipio de Lajas (approximately
Puerto Rico metre grid 23,900 N; 93,400 to 93,600 E), and has the USGS Mesozoic
locality numbers 28664, 28748, 29075, 29088 and 29361. The material was
collected by N.F. Sohl, E.G. Kauffman and W.O. Ross in 1963
and 1964 (Sohl, 1992, p. 433).
|
Figure 1:
Location map from which the silicified rudists were obtained. Locality 14
from Sohl (1998, p. 24). A, detailed locality. B, location in Puerto Rico
(white
square). |
The succession exposed on the hill was divided into three parts by Sohl (1998): the lower part consisted of massive, grey limestone with ostreid bivalves and other molluscs; the poorly exposed middle part of the hill consisted of inter-bedded brownish shale and limestone, and it was blocks from here that yielded the silicified material described here; and the top of the hill was capped by light-grey limestones containing the rudists Parastroma sp. and Titanosarcolites giganteus (Whitfield).
The silicified residue was separated from the limestone by acidization (Sohl, 1998, p. 25). Since material from here was described in Kauffman and Sohl (1974, Fig. 7F-H) and Sohl and Kollmann (1985), the acid digestion must have been done before those dates (presumably in the late 1960s). It is assumed that this was done using dilute hydrochloric acid, but no further details of the procedure have been recorded in the literature (Kauffman & Sohl, 1974; Sohl & Kollmann, 1985; Sohl, 1992, 1998).
The silicified fauna is highly diverse, and Sohl (1998) recognized nearly 80 genera of gastropods and bivalves including the rudists Antillocaprina, Plagioptychus, Parastroma, Orbignya, Durania, and various other smaller radiolitids. The Durania in this list would now be referred to Macgillavryia, and Orbignya to Caribbea. Sohl (1998) assigned a Maastrichtian age to this assemblage. The presence of Caribbea and Laluzia in association with Parastroma guitarti (Palmer), Coralliochama cf. gboehmi B�se and a single specimen of Oligosarcolites monotubularis (Mitchell & Gunter) (Mitchell, 2013, Fig. 12.1-2; Skelton, 2013, Fig. 14a-b) would suggest assignment to the early Maastrichtian (e.g., Mitchell, 2013; Pons et al., 2013).
The hippuritid rudists described here, Caribbea muellerriedi (Vermunt), Laluzia peruviana Gerth and Parastroma guitarti (Palmer), have very complex shells and numerous terms have been introduced to describe their morphology (e.g., Woodward, 1862; Whitfield, 1897; Dommelen, 1971) together with standard terms (e.g., Moore, 1969). The Appendix provides a list and brief description of these morphological terms. For higher level classification, see Skelton (2013). Abbreviations used in the descriptions are listed in Table 1.
Table 1: Abbreviations used in description of species (see Appendix for details).
AM - anterior myophore
ATS - anterior tooth socket
AT - anterior tooth
CT - central tooth
LV - left valve
P0 - ligamental infold
P1 - first pillar/infold
P2 - second pillar/infold
PM - posterior myophore
PTS - posterior tooth socket
PT - posterior tooth
RV - right valve
Specimens reside at the following repositories: Naturalis, Leiden, The Netherlands (NL); Smithsonian National Museum of Natural History, Washington - D.C. (SNMNH); Museum of Paleontology, Institute of Geology, Universidad Nacional Aut�noma de M�xico (UNAM), Ciudad Universitaria, M�xico (IGM); The Natural History Museum London (BMNH); The University of the West Indies Geology Museum (UWIGM). Specimens in the collection at Washington also have an NS number, which was assigned to specimens as they were being studied/photographed; these numbers are retained here for cross-referencing purposes. Species synonymy lists use marginal annotations (e.g., 'v.', '.', '*') as used by Mathews (1973).
Order HIPPURITIDA Newell, 1965
Suborder HIPPURITIDINA Newell, 1965
Superfamily RADIOLITOIDEA Orbigny, 1847
Family HIPPURITIDAE Gray, 1848
Discussion. The family has been variously split into subfamilies based on the presence/absence of pores (e.g., Grubić, 1979), presence/absence of multiple infolds (e.g., Chubb, 1971) and differences in right valve outer shell structure (e.g., Grubić, 2004). This study of the silicified material from Puerto Rico and investigation of other New World and Old World specimens indicates that three subfamilies can be recognized within the Hippuritidae: the Hippuritinae Mac Gillavry, 1937, the Barrettiinae Chubb, 1971, and the Torreitinae Grubić, 1979.
Subfamily BARRETTIINAE Chubb, 1971
Diagnosis. Hippuritidae in which the sockets for each tooth are formed by two pairs of slots on the inner shell wall.
Discussion. This division of the Hippuritidae into subfamilies is based on major morphological features - differences in the sockets for the teeth and differences in the construction of the left valve - which are ascribed to taxonomic differentiation at a level above that of genus but below that of family. Chubb (1971) based his subfamily Barrettiinae on the presence of multiple infolds (rays) and included within it all the American multiple-fold hippuritids, but excluded the Old World multiple-fold Pironaea Meneghini, which he distinguished based on the radial distance between the two principle pillars (P1 and P2), which was small in Pironaea and large in the Barrettiinae. However, Mitchell (2010) demonstrated that this distance was too highly variable in three New World species to even distinguish species let alone genera or subfamilies, and the radial distance is an unsound criterion upon which to erect a subfamily. Grubić (2004) resurrected the subfamily Barrettiinae Chubb and gave a revised diagnosis based on the structure of the outer shell layer of the right valve. Yet, both Mitchell (2010) and Pons et al. (2010) have subsequently demonstrated that the structure described by Grubić (2004) does not exist and is an erroneous interpretation of previous descriptions.
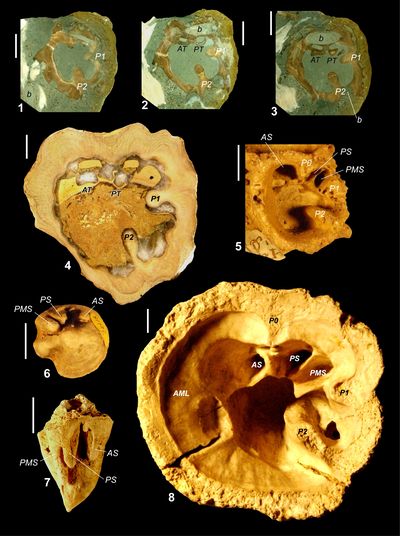
Here, the subfamily Barrettiinae is defined on differences in the socket
construction in the right valve of Caribbean and Central American Barrettiinae
as compared with Old World Hippuritinae (Fig. 2 ![]() ). In the Barrettiinae, the
socket for each tooth is formed by a pair of slots, one pair each for the
posterior and anterior tooth (Fig. 2.1-4
). In the Barrettiinae, the
socket for each tooth is formed by a pair of slots, one pair each for the
posterior and anterior tooth (Fig. 2.1-4 ![]() ). When preserved in three-dimensions,
it is obvious that the teeth are open to the body cavity in the right valve, and
furthermore, the slots are formed from the inner wall of the right valve and do
not involve the tabulae. In some ways this mimics the slots developed for the
teeth in many genera in the family Radiolitidae. In contrast, in the
Hippuritinae Mac Gillavry, 1937, the sockets for the teeth in the right
valve are formed by a lamina that encloses the region where the teeth would fit
(Douvill�, 1891, Pl. 1, fig. 6;
Dechaseaux, 1952, Pl. 2;
Fig. 2.5-8
). When preserved in three-dimensions,
it is obvious that the teeth are open to the body cavity in the right valve, and
furthermore, the slots are formed from the inner wall of the right valve and do
not involve the tabulae. In some ways this mimics the slots developed for the
teeth in many genera in the family Radiolitidae. In contrast, in the
Hippuritinae Mac Gillavry, 1937, the sockets for the teeth in the right
valve are formed by a lamina that encloses the region where the teeth would fit
(Douvill�, 1891, Pl. 1, fig. 6;
Dechaseaux, 1952, Pl. 2;
Fig. 2.5-8 ![]() ); the laminae represent an upfolding of the tabulae that fill the body
cavity. The form of the sockets is clear in three-dimensional forms of both the
Barrettiinae and the Hippuritinae but cannot always be resolved in transverse
sections cut close to the commissure of the right valve. Sections cut close to
the commissure of the right valve typically miss the lamina forming the socket
in the Hippuritinae and do not allow the form of the socket to be determined. In
such cases more adapical transverse sections need to be cut in order to
distinguish between members of the two subfamilies.
); the laminae represent an upfolding of the tabulae that fill the body
cavity. The form of the sockets is clear in three-dimensional forms of both the
Barrettiinae and the Hippuritinae but cannot always be resolved in transverse
sections cut close to the commissure of the right valve. Sections cut close to
the commissure of the right valve typically miss the lamina forming the socket
in the Hippuritinae and do not allow the form of the socket to be determined. In
such cases more adapical transverse sections need to be cut in order to
distinguish between members of the two subfamilies.
|
Figure
2:
Form
of sockets for teeth in Old Word (Hippurites) hippuritids (formed by cavities) and New
World (Carribea) hippuritids (formed by paired slots). 2.1, Caribbea
muellerriedi (Vermunt, 1937),
UWIGM.2010.01.0001 (same specimen as Mitchell,
2010, Fig. 3A, Guinea Corn Formation (lower upper Maastrichian), Central Inlier,
Jamaica. 2.4, Caribbea
maldonensis (Chubb),
no number (UWIGM collection), Maldon Formation (upper upper Maastrichtian),
Maldon Inlier, Jamaica. 2.5, Hippurites sp.
no number (BMNH collection), Turonian, I'iltarium, Uchaux, Vancluse, France. 2.6-7,
Hippurites
lapeiroussii (Goldfuss, 1840), interior mould of the body cavity preserving the form of the sockets for
teeth and posterior myopohre, BMNH. 81722, Maastricht Chalk (Md zone,
Maastrichtian), ENCI Quarry, St. Pietersburg, The Netherlands. 2.8, Hippurites
radiosus Des Moulins. BMNH.PI RU 21, Campanian, Barbezieux, Charente, SW France. P0, P1, P2,
pillars; AT, PT, anterior and posterior teeth in slots; ATS, PTS, anterior and
posterior sockets for the teeth, PM, PMS, posterior myophore and socket,
respectively. |
Grubić (1979) erected the subfamily Torreitinae to distinguish Torreites from the Hippuritinae. The subfamily is accepted here and has sockets formed in the same way as the Hippuritinae but lacks a pore system in the left valve. Table 2 shows a revision of the Hippuritidae as recognized here.
Table 2: Classification of the Hippuritidae with a revision of New World forms.
|
Hippuritinae Mac Gillavry |
|
Hippurites Lamarck, 1801 Pironaea Meneghini, 1868 Pseudovaccinites S�nesse, 1946 Tetracionites Astre, 1931 Vaccinites Fischer, 1887 Yvaniella Milovanović, 1938 |
|
Barrettiinae Chubb |
Caribbea Clade |
Caribbea Grubić, 2004 Laluzia G�tz & Mitchell, 2009 Praebarrettia Trechmann, 1924 |
|
Barrettia Clade |
Barrettia Woodward, 1862 Parastroma Douvill�, 1926 Whitfieldiella Mitchell, 2010 |
|
|
Torreitinae Grubić |
|
Praetorreites Philip & Platel, 1994 Torreites Palmer, 1933 Polytorreites Mitchell & Skelton, 2013 |
Caribbea Clade
Diagnosis. Barrettiinae that lack pallial canals in the inner layer of the left valve.
Remarks. This clade contains three genera: Caribbea Grubić, 2004, Laluzia G�tz & Mitchell, 2009, and Praebarrettia Trechmann, 1924.
Genus Caribbea Grubić, 2004
Type species. Orbignya muellerriedi Vermunt, 1937, from the lower Maastrichtian of the Province of Pinar del Rio, Cuba.
Emended Diagnosis. A hippuritid with two primary infolds (P1 and P2) and an incipient ligamental ridge. The PT and AT have ridges that fit into channeled grooves (sockets) on the CT and inner shell layer of the RV. The PM fits into an embayment in the inner shell layer and is not separated from the body cavity by a lamina. The AM is attached directly to the inner shell layer. The outer shell layer of the RV is composed of funnel plates that are separated by irregular dendritic radial muri or pustules. Pallial canals are absent from the inner shell layer of the LV. The radial canals of the LV are overlain by a pore system characterized by thin-walled irregular to polygonal canals that are subdivided to form porules by merging denticles. The pores are irregularly arranged in the central part of the LV, but form radial lines over numerous narrow radial canals in the limbal zone.
Discussion. The generic placement of the simple hippuritids of the Maastrichtian of the Americas has long been debated. Most authors have used the pillar arrangements (prominent P1 and P2, incipient P0) to place the American forms grouped around muellerriedi within Hippurites Lamarck, 1801 (=Orbignya Fischer, 1887). Mac Gillavry (1937, p. 111) also described muellerriedi under the generic name Hippurites, but suggested it might belong to Hippuritella Douvill�. Mitchell (2010) illustrated the pores in the LV of a specimen of muellerriedi from Jamaica and placed the species in Hippuritella. Grubić (2004) erected the genus Caribbea based on the structure of the outer shell layer of the RV.
The RVs of Hippurites species from the Old World have well-developed P1 and P2, but incipient development of P0; furthermore, their pore systems are characterized by round, linear or vermicular pores that are clearly distinct from the American muellerriedi. The RV of Hippuritella is superficially similar to that of Hippurites, but instead of simple (round, linear or vermicular) pores, the pore system consists of polygonal pores, and a placement of the American muellerriedi in Hippuritella based on the pore system would appear attractive (Mitchell, 2010).
Grubić's (2004) character of vertical canals in the outer shell layer of the RV is now known to be incorrect. This is due to a misunderstanding of the sections and descriptions of the shell wall given by Chubb (1956) for 'Parastroma' maldonensis and by Mac Gillavry (1937) for 'Hippurites' muellerriedi. Because Mac Gillavry (1937) stated that the shell structure of muellerriedi might be similar to some Old World forms (e.g., 'Hippurites' cornucopiae Defrance figured by Douvill�, 1910, Pl. 41, fig. 3, and Hippurites lapeirousei Goldfuss of Parona, 1900, Pl. 1, fig. 2), the structure of the outer shell layer of the RV would appear to be an unsuitable criterion for erecting a new genus.
However, the construction of the sockets for the teeth and PM in muellerriedi, now apparent from the Puerto Rican material described here, contrasts markedly with both Hippurites and Hippuritella. In the latter two genera (together with Vaccinites Fischer), the sockets in the RV for the teeth and PM are vertical cavities developed within the inner shell layer. Transverse sections through the RVs of Hippurites, Hippuritella and Vaccinites, depending on orientation relative to, and distance from the commissure, show the teeth and PM of the LV partially or completely surrounded by the inner shell layer of the RV. In specimens preserved in three-dimensions, such as Hippurites radiosus Des Moulins from the Maastrichtian of France (Dechaseaux, 1952, Pl. 2) or Vaccinites petrocoriensis (Douvill�) from the middle-upper Turonian of Gourd-del-Arche (Douvill�, 1891, Pl. 1, fig. 6), the sockets for the teeth and PM are clearly formed from the tabulate infilling of the adapical part of the body cavity of the RV. In contrast, each socket in C. muellerriedi is represented by an embayment in the dorsal inner shell wall with slots (defined by two thin laminae) developed from the shell wall and the CT; on the ventral side, each tooth is open to the body cavity for its entire length, whereas on the dorsal side each tooth is flanked by an embayment of the inner shell layer. Thus, the quite different construction of the sockets of the RVs of New World Caribbea (slots on the inner layer of the RV) and Old World Vaccinites, Hippurites and Hippuritella (cavities in the tabulate infilling of the adapical body cavity of the RV) justifies the placement of muellerriedi in a separate genus.
The external surface of the RV of specimens of Caribbea from the New World is variable with three contrasting morphotypes. The first morphotype is exhibited by C. muellerreidi and C. maldonensis (Chubb) from the Maldon Inlier (Jamaica: Gunter, 2002); it is rounded other than for three sulci corresponding to the principal folds and a few more sulci between as described below. The second morphotype is shown by 'H. perkinsi' Myers from Cardenas (Mexico) and the Jerusalem Mountain Inlier (Jamaica) and most C. maldonensis from the Central Inlier (Jamaica); these have angular costae developed between the sulci. The third morphotype seen in specimens from the Jerusalem Mountain Inlier (Jamaica) is characterized by fine longitudinal riblets (similar to those described below in Laluzia peruviana). Given the change in outer shell microstructure from C. muellerriedi to C. maldonensis and the corresponding existence of two of the three morphotypes in both species, it is clear that the morphotypes are phenotypic and have little taxonomic value. For descriptive purposes these might be referred to as (in roman text) the 'muellerriedi' morphotype, the 'perkinsi' morphotype and the 'peruviana' morphotype, but without implying any taxonomic significance. As such, only a limited number of species of Caribbea need to be recognized in the Maastrichtian of the Caribbean: C. muellerriedi from the lower to lower upper Maastrichtian of Mexico, Cuba, Jamaica and Puerto Rico, C. maldonensis from the mid upper Maastrichtian of Jamaica, and an undescribed giant form from the uppermost Maastrichtian of Jamaica.
Caribbea muellerriedi Vermunt, 1937
(Figs. 3.1-6 ![]() ,
4.1-4
,
4.1-4 ![]() ,
5.1-6
,
5.1-6 ![]() ,
4.1-4
,
4.1-4 ![]() ,
5.1-6
,
5.1-6 ![]() ,
6.1-6
,
6.1-6 ![]() , 7
, 7 ![]() )
)
v. 1924 Hippurites (Orbignya) sp.; Trechmann, p. 396, Pl. 23, fig. 5.
. 1930 Hippurites (Hippuritella Douvill�; Orbignya Toucas) cf. incisus (Douvill�); M�llerried, p. 165, Figs. 1-2.
*. 1937 Orbignya m�llerriedi Vermunt, p. 261, Fig. 3a-d; Pl. 36, figs. 1-3.
v. 1937 Hippurites muellerriedi (Vermunt); Mac Gillavry, p. 110, Fig. 2 (on p. 123); Pl. 5, fig. 6.
. 1937 Hippurites mullerriedi (Vermunt); Mac Gillavry, p. 111, Pl. 5, fig. 6.
v. 1956 Hippurites (Orbignya) mullerriedi (Vermunt); Chubb, p. 19, Pl. 4, figs. 4-8.
v. 1956 Hippurites (Orbignya) ceibarum; Chubb, p. 19, Pl. 4, figs. 9-10.
v. 1968 Hippurites muellerriedi (Vermunt); Myers, p. 42, Pl. 3, figs. 4-6.
v. 1968 Hippurites perkinsi, n.sp.; Myers, p. 43, Pl. 4, figs. 5-6.
v. 1971 Orbignya mullerriedi Vermunt; Chubb, p. 204, Pl. 49, figs. 1-2.
v. 1971 Orbignya ceibarum (Chubb); Chubb, p. 204, Fig. 6; Pl. 49, fig. 3.
v. 1974 Orbignya Kauffman and Sohl, p. 413, Fig. 7F-H.
. 1975 Hippurites muellerriedi (Vermunt); Lupu, p. 240, Fig. 20.
v. 2004 Caribbea muellerriedi (Vermunt); Grubić, p. 147, Pl. 2, fig. 1.
v. 2004 Caribbea ceibarum (Chubb); Grubić, p. 147, Pl. 2, fig. 2.
v. 2004 Caribbea sladici Grubić, p. 148, Pl. 2, fig. 4.
v. 2006 Hippurites perkinsi Myers; Schafhauser, p. 62, Pl. 16, figs. 8-9.
v. 2010 Hippuritella muellerriedi (Vermunt); Mitchell, Fig. 3A.
. 2013 Caribbea muellerriedi (Vermunt); Pons et al., p. 738-742, Figs. 16.1-16.11, 17.1-17.7, 18.1-18.3.
. 2018 Caribbea muellerriedi (Vermunt); Skelton, Fig. 7.6 (= Fig. 3.4
herein).
Holotype. The holotype (Min.-Geol. Inst., Univ. Utrecht, W. C. 1933, 6, now in Naturalis, Leiden, The Netherlands) from the lower Maastrichtian, Locality L818, province of Pinar del Rio, Western Cuba was figured by Vermunt (1937, Pl. 36, figs. 1-2). A new photograph of it is illustrated in Figure 7 and it shows no apparent differences from the material included in this species from Jamaica, Puerto Rico or Mexico.
Material. Many specimens in the USNM. Material specifically referred to and placed in the type collection: NS31 (USNM 547509) (Locality 1202, USNM 187687), NS32 (USNM 547510) (Locality 1202, USNM 187687), NS35 (USNM 547511) (Locality 1202), NS39 (USNM 547512) (Locality 1445-3), NS7-10 (USNM 547513-547516) (Locality 1202?), NS70 (USNM 547517) (Locality 1202? draw 131, pores), NS33 (USNM 547548) (Locality 1202, USNM 187687), and NS71 (USNM 547518) (draw 131, Locality 1202, tabulae). All from the El Rayo Formation, Puerto Rico.
Extensive comparable material is available from Cuba (Naturalis, Leiden, The Netherlands), Jamaica (UWI Geology Museum) and Mexico (Texas Memorial Museum, The University of Texas at Austin). All this material agrees very well with the material from Puerto Rico described here.
Diagnosis. A small to medium sized species of Caribbea with a wall structure defined by radial muri on funnel plates of the outer shell layer throughout growth.
Description. The RV is conical gradually increasing in diameter. Its surface is
marked by regular furrows (Fig. 3.1-2, 3.5 ![]() ), the three deepest corresponding to
the three pillars (P0, P1 and P2). Between P0 and P1 there is one (rarely two)
further weak furrow(s), and between P1 and P2 another weak furrow; weak furrows
also occur around the remainder of the shell's circumference, but are
generally not as well developed as the ones between the pillars. The surface
between the furrows is regularly convex and smooth.
), the three deepest corresponding to
the three pillars (P0, P1 and P2). Between P0 and P1 there is one (rarely two)
further weak furrow(s), and between P1 and P2 another weak furrow; weak furrows
also occur around the remainder of the shell's circumference, but are
generally not as well developed as the ones between the pillars. The surface
between the furrows is regularly convex and smooth.
The inner layer of the RV is thin (Fig. 3.2 ![]() ); the outer layer of the RV
is composed of funnel plates with irregular dendritic radial
muri (Fig.
3.2, 3.5
); the outer layer of the RV
is composed of funnel plates with irregular dendritic radial
muri (Fig.
3.2, 3.5 ![]() ). The funnel plates are directed outwards and upwards at an angle of about
60� to 70� to the horizontal. P1 and P2 are strong infolds of the outer shell
layer into the body cavity and have a microstructure identical to the remainder
of the RV outer shell wall. P0 is a very weak (incipient) infold into the body
chamber to which the central tooth is attached (Figs. 3.6
). The funnel plates are directed outwards and upwards at an angle of about
60� to 70� to the horizontal. P1 and P2 are strong infolds of the outer shell
layer into the body cavity and have a microstructure identical to the remainder
of the RV outer shell wall. P0 is a very weak (incipient) infold into the body
chamber to which the central tooth is attached (Figs. 3.6 ![]() ,
4.4
,
4.4 ![]() ,
5.2
,
5.2 ![]() ); the central
tooth is an inward projection of the inner shell layer and has two laminae each
on the posterior and anterior sides that create slots for the PT and AT to fit
into; single strong vertical laminae extend on the ventral-side of the wall of
the body cavity where the teeth would fit together with very weak ribs on the
dorsal side to form the sockets for the PT and AT (Fig. 5.2
); the central
tooth is an inward projection of the inner shell layer and has two laminae each
on the posterior and anterior sides that create slots for the PT and AT to fit
into; single strong vertical laminae extend on the ventral-side of the wall of
the body cavity where the teeth would fit together with very weak ribs on the
dorsal side to form the sockets for the PT and AT (Fig. 5.2 ![]() ). The interior of
the RV contains only incipient (weak to very weak) secondary folds into the body
cavity. There is no socket for the PM, which simply fits between the lamina for
the PT and P1 (Figs. 3.5
). The interior of
the RV contains only incipient (weak to very weak) secondary folds into the body
cavity. There is no socket for the PM, which simply fits between the lamina for
the PT and P1 (Figs. 3.5 ![]() ,
4.4
,
4.4 ![]() ,
5.2
,
5.2 ![]() ). No socket is present for the AM, which must
attach directly onto the interior wall of the RV. The tabulae in the body cavity
are plate-like and widely spaced (Fig. 3.3-4
). No socket is present for the AM, which must
attach directly onto the interior wall of the RV. The tabulae in the body cavity
are plate-like and widely spaced (Fig. 3.3-4 ![]() ). The last tabula is inserted
adapically of the point to which the teeth and PM penetrate the RV, so that
transverse sections of a RV never show the teeth and tabulae at the same level (Fig.
3.3
). The last tabula is inserted
adapically of the point to which the teeth and PM penetrate the RV, so that
transverse sections of a RV never show the teeth and tabulae at the same level (Fig.
3.3 ![]() ).
).
The LV is weakly concave, flat or very gently convex (Figs.
4.1, 4.4 ![]() ,
5.1, 5.4
,
5.1, 5.4 ![]() , 6.1-2
, 6.1-2 ![]() ). The AT, PT and PM are arranged in a straight line
(Figs. 4.2-3
). The AT, PT and PM are arranged in a straight line
(Figs. 4.2-3 ![]() ,
6.3-6
,
6.3-6 ![]() ). The AT is broadest and longest and is generally two-pronged
(Figs. 4.2
). The AT is broadest and longest and is generally two-pronged
(Figs. 4.2 ![]() ,
6.3-4
,
6.3-4 ![]() ) and has a rectangular transverse cross-section. The PT is about as
wide as broad (Fig. 6.3-6
) and has a rectangular transverse cross-section. The PT is about as
wide as broad (Fig. 6.3-6 ![]() ); in transverse cross-sections it is strongly curved
with its posterior-ventral aspect highly convex and its anterior-dorsal margin
straight to gently concave. The PM is a narrow to relatively narrow radially
orientated blade (Fig. 6.3-4
); in transverse cross-sections it is strongly curved
with its posterior-ventral aspect highly convex and its anterior-dorsal margin
straight to gently concave. The PM is a narrow to relatively narrow radially
orientated blade (Fig. 6.3-4 ![]() ). The AM extends anteriorly from the AT and gently
slopes towards the ventral aspect adapically; it forms a curved blade that
extends outwards and abapically at an angle of about 45� (Figs.
4.3
). The AM extends anteriorly from the AT and gently
slopes towards the ventral aspect adapically; it forms a curved blade that
extends outwards and abapically at an angle of about 45� (Figs.
4.3 ![]() ,
5.3, 5.6
,
5.3, 5.6 ![]() ,
6.5
,
6.5 ![]() ). The teeth and myophores are connected to a circular
myocardinal yoke,
which lacks openings (Figs. 5.3, 5.6
). The teeth and myophores are connected to a circular
myocardinal yoke,
which lacks openings (Figs. 5.3, 5.6 ![]() ,
6.3-6
,
6.3-6 ![]() ). The PM is constructed on a strong
buttress that extends radially and is supported by two or three very weak radial
buttresses (Fig. 6.4
). The PM is constructed on a strong
buttress that extends radially and is supported by two or three very weak radial
buttresses (Fig. 6.4 ![]() ). A strong buttress also extends between the openings for
P1 and P2 and bifurcates towards the margin of the valve (Figs.
5.2-3
). A strong buttress also extends between the openings for
P1 and P2 and bifurcates towards the margin of the valve (Figs.
5.2-3 ![]() ,
6.3
,
6.3 ![]() ); it
may be supported by a few radiating incipient buttresses. A weak buttress
extends along the ventral side of the opening (O2) above P2 (Fig.
6.2, 6.5
); it
may be supported by a few radiating incipient buttresses. A weak buttress
extends along the ventral side of the opening (O2) above P2 (Fig.
6.2, 6.5 ![]() ). The
myocardinal yoke below the AM is supported by incipient to weak radially
directed buttresses, which may number 7, 8 or 9 (Fig. 6.3,
6.5-6
). The
myocardinal yoke below the AM is supported by incipient to weak radially
directed buttresses, which may number 7, 8 or 9 (Fig. 6.3,
6.5-6 ![]() ). The LV
contains a series of radial canals that extend to the margin of the valve and
are visible when the pore system is not preserved (Figs. 4.1
). The LV
contains a series of radial canals that extend to the margin of the valve and
are visible when the pore system is not preserved (Figs. 4.1 ![]() ,
5.4-5
,
5.4-5 ![]() ). The canals
have maximum widths of 2 mm and the walls between adjacent canals are 0.3 mm
thick. New canals are intercalated between earlier formed canals during growth (Fig.
4.1
). The canals
have maximum widths of 2 mm and the walls between adjacent canals are 0.3 mm
thick. New canals are intercalated between earlier formed canals during growth (Fig.
4.1 ![]() ). The canals open onto the apertural surface of the RV
(Fig. 4.2-3
). The canals open onto the apertural surface of the RV
(Fig. 4.2-3 ![]() ).
In large individuals the limbal zone is different: the large canals continue
as thin canals and many additional thin limbal canals are inserted. The limbal
canals have widths of 0.3 to 0.4 mm and are separated by walls with
thicknesses of 0.3 to 0.4 mm (Figs. 4.1
).
In large individuals the limbal zone is different: the large canals continue
as thin canals and many additional thin limbal canals are inserted. The limbal
canals have widths of 0.3 to 0.4 mm and are separated by walls with
thicknesses of 0.3 to 0.4 mm (Figs. 4.1 ![]() ,
6.1-2
,
6.1-2 ![]() ). The larger radial canals open
onto the adapical portion of the apertural surface of the outer layer of the RV,
the limbal canals open onto the abapical portion of the apertural surface of the
outer layer of the RV (Fig. 6.5-6
). The larger radial canals open
onto the adapical portion of the apertural surface of the outer layer of the RV,
the limbal canals open onto the abapical portion of the apertural surface of the
outer layer of the RV (Fig. 6.5-6 ![]() ). The arrangement of apertural openings of
the radial and limbal canals around the margins of the shell agrees with the
pattern previously described for various Old World hippuritids (e.g.,
Skelton, 1976, Fig. 2; Schumann,
2010). The radial canals are
overlain by the pore system which is uniform across the central region of the LV
(that is no pustules are developed above the inner terminations of newly
intercalated radial canals as in many Old World hippuritids). The pores in the
inner zone covering the normal radial canals are polygonal with diameters of 0.3
to 0.5 mm and with three or four pores situated concentrically over each radial
canal (Fig. 6.2
). The arrangement of apertural openings of
the radial and limbal canals around the margins of the shell agrees with the
pattern previously described for various Old World hippuritids (e.g.,
Skelton, 1976, Fig. 2; Schumann,
2010). The radial canals are
overlain by the pore system which is uniform across the central region of the LV
(that is no pustules are developed above the inner terminations of newly
intercalated radial canals as in many Old World hippuritids). The pores in the
inner zone covering the normal radial canals are polygonal with diameters of 0.3
to 0.5 mm and with three or four pores situated concentrically over each radial
canal (Fig. 6.2 ![]() ). The walls between the pores are about 0.1 mm thick. Each pore
has three or four denticles that in well-preserved regions of well-preserved
specimens merge to form a fine reticulate covering consisting of three or four
porules in each pore (Figs. 4.1
). The walls between the pores are about 0.1 mm thick. Each pore
has three or four denticles that in well-preserved regions of well-preserved
specimens merge to form a fine reticulate covering consisting of three or four
porules in each pore (Figs. 4.1 ![]() ,
6.2
,
6.2 ![]() ). It is likely that originally all pores
were subdivided to form porules, but the finer denticles have been damaged or
were not silicified. The limbal canals are overlain by pores which are radially
elongate (Figs. 4.1
). It is likely that originally all pores
were subdivided to form porules, but the finer denticles have been damaged or
were not silicified. The limbal canals are overlain by pores which are radially
elongate (Figs. 4.1 ![]() ,
6.1-2
,
6.1-2 ![]() ). These radially elongate pores also have fine
denticles that would have subdivided them into two porules, which are arranged
radially (Figs. 4.1
). These radially elongate pores also have fine
denticles that would have subdivided them into two porules, which are arranged
radially (Figs. 4.1 ![]() ,
6.1
,
6.1 ![]() ). The
'oscules' for P1 and P2 are surrounded by a
single line of vertical canals with a reticulate covering; 'oscules' may
even have been completely covered by a reticulate cover as some specimens
preserve part of this (Fig. 6.2
). The
'oscules' for P1 and P2 are surrounded by a
single line of vertical canals with a reticulate covering; 'oscules' may
even have been completely covered by a reticulate cover as some specimens
preserve part of this (Fig. 6.2 ![]() ).
).
|
|
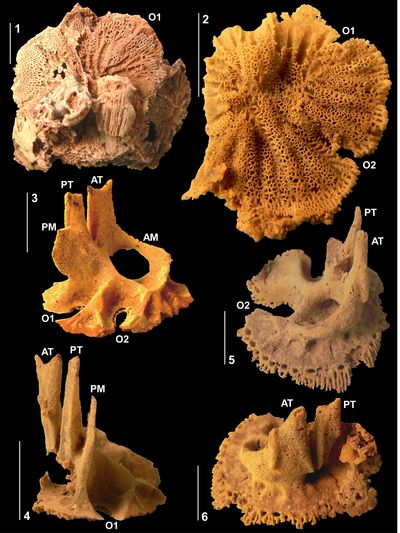
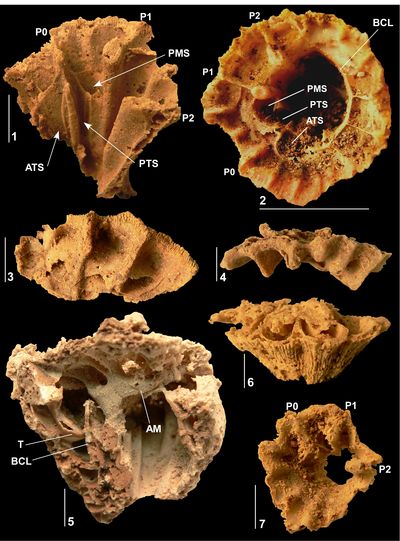
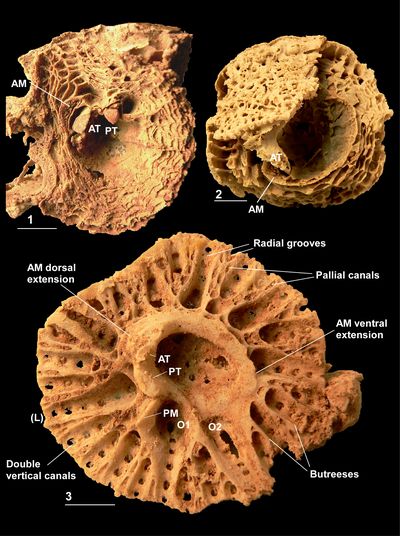
Figure
3:
Caribbea
muellerriedi (Vermunt, 1937), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 3.1-2,
RV (NS32; USNM 547510) attached to
conjoined specimen of Antillocaprina
suboccidentalis Chubb: 3.1,
showing grooves on exterior surface of RV; 3.2,
adapical view showing pillars, slots for teeth, and outer shell layer wall
structure. 3.3-4 (NS71:
USNM 547518), bouquet
of two conjoined RVs, 3.3,
adapical view showing single tabula closing off the body cavity, note that the
sockets for the teeth are not formed by folding of the tabula; 3.4,
flank with outer shell layer broken away showing the tabulae in body cavity. 3.5-6
(NS33: USNM
547548), bouquet of
four RVs: 3.5,
flank showing grooves on exterior surface with rounded costae between; 3.6,
adapical view of interior of RV showing outer shell layer structure, thin inner
shell layer, and myocardinal arrangement; note that sockets for teeth are formed
by slots on the central tooth and that the 'socket' for the PM is open to
the body cavity. P1 and P2, first and second pillars; ATS, anterior socket; PTS,
posterior socket; PMS, posterior myophore socket; ISL, inner shell layer; OSL,
outer shell layer. Scale bar = 10 mm. |
|
|
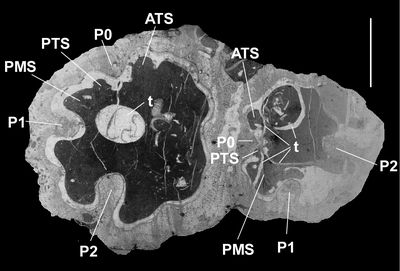
Figure 4:
Caribbea
muellerriedi (Vermunt, 1937), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 4.1, 4.4 (NS33: USNM 547548),
bouquet of four RVs (same as Fig. 3.5-6 |
|
|
Figure
5:
Caribbea
muellerriedi (Vermunt, 1937), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 5.1 (NS10: USNM 547516),
LV, abapical view, showing radial canals and pores. 5.2 (NS50: USNM
547542), RV of a
specimen from a bouquet showing sockets for anterior and posterior teeth and
posterior myophore. 5.3-4 (NS35: USNM 547511),
LV: 5.3,
oblique view showing teeth and myophores; 5.4,
abapical view showing radial canals and partially preserved pores. 5.5-6 (NS31:
USNM
547509), LV, 5.5,
abapical view showing radial canals (no pores preserved); 5.6,
oblique view showing teeth and myophores. P1, first pillar; O1 and O2, oscules
for first and second pillars; AT/ATS, anterior tooth/socket; PT/PTS, posterior
tooth/socket; PM/PMS, posterior myophore/myophore socket; CT, central tooth; RF,
radial canals; B, buttress. Scale bar = 10 mm. |
|
|
Figure 6:
Caribbea
muellerriedi (Vermunt, 1937), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 6.1 (NS39: USNM 547512),
LV, abapical view showing radial canals and well preserved pores and porules. 6.2
(NS70: USNM
547517), LV,
abapical view showing radial canals and well preserved pores and porules. 6.3
(NS9: USNM
547515), LV,
oblique view, showing myocardinal arrangement; note lack of canals penetrating
myocardinal yoke. 6.4 (NS35: USNM 547511),
LV, oblique view showing myocardinal elements. 6.5-6 (NS8A: USNM 547514),
LV showing myocardinal arrangement, oblique views; 6.5,
note lack of canals penetrating myocardinal yoke, 6.6,
note ridges on teeth that fit into the grooved sockets. O1 and O2, oscules for
first and second pillars, AT, anterior tooth; PT, posterior tooth; Am, anterior
myophore; PM, posterior myophore. Scale bar = 10 mm. |
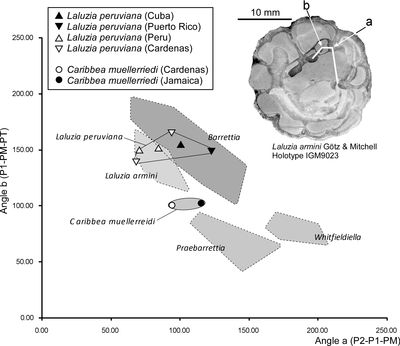
The relationships between the angles P1-PM-PT and P2-P1-PM (Fig. 8 ![]() )
separate different genera
of multiple fold hippuritids of the Caribbean region (Mitchell,
2010). This plot requires
articulated specimens so that the position of the PM can be determined (there is
no enclosed socket in Caribbea) and it
can be determined in specimens from Jamaica and Mexico, but not from Puerto Rico.
Caribbea plots in an intermediate
position between Laluzia and Praebarrettia.
)
separate different genera
of multiple fold hippuritids of the Caribbean region (Mitchell,
2010). This plot requires
articulated specimens so that the position of the PM can be determined (there is
no enclosed socket in Caribbea) and it
can be determined in specimens from Jamaica and Mexico, but not from Puerto Rico.
Caribbea plots in an intermediate
position between Laluzia and Praebarrettia.
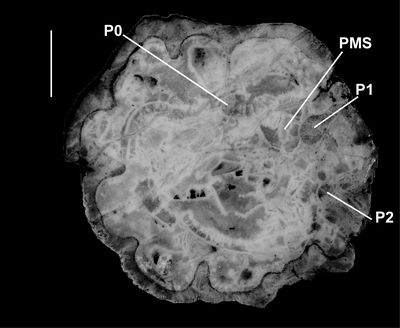
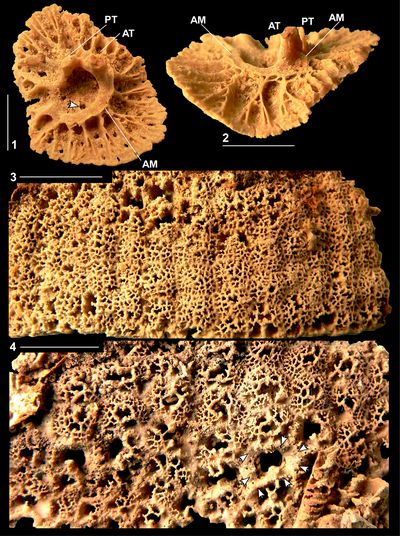
Discussion. The holotype of Orbignya
muellerreidi was figured by Vermunt (1937, Pl. 6, figs. 1-2) and
shows orimentary folds and slots on the CT for the reception of the teeth of the
LV (Fig. 7 ![]() ). The adjoining individual shows a round cavity, however this is not
the cavity for the PM, but an oblique cut through a tabula filling the lower
part of the body cavity. A socket is not present between the ligamentary
infold and P1 for the reception of the PM and this material is fully consistent
with the material described from Puerto Rico in this paper (as well as material
from Jamaica and Mexico). Furthermore, specimens of RVs examined by SFM in 2009
in the collections of the Museo Nacional de Historia Natural, La Habana, Cuba,
are also identical to the Puerto Rican material.
). The adjoining individual shows a round cavity, however this is not
the cavity for the PM, but an oblique cut through a tabula filling the lower
part of the body cavity. A socket is not present between the ligamentary
infold and P1 for the reception of the PM and this material is fully consistent
with the material described from Puerto Rico in this paper (as well as material
from Jamaica and Mexico). Furthermore, specimens of RVs examined by SFM in 2009
in the collections of the Museo Nacional de Historia Natural, La Habana, Cuba,
are also identical to the Puerto Rican material.
|
|
Figure
7:
Caribbea
muellerriedi (Vermunt, 1937), new
photograph of holotype, thin section (somewhat oxidized), province of Pinar del
Rio (lower Maastrichtian), Cuba. Note that sockets for teeth are not enclosed
and that the posterior myophore would have fitted into an embayment in the inner
shell wall, an embayment that is not separated from the body cavity by a laminar.
Scale bar = 10 mm. |
|
|
Figure
8:
Scatter
Plots of myocardinal arrangements of selected hippuritid rudists. Laluzia
peruviana plots
in the field overlapping with Laluzia
armini and Barrettia and not in the
field of Praebarrettia. Caribbea with
its simple infolds plots midway between the fields for Laluzia and Praebarrettia,
and could be the ancestor of either or both of these genera. Fields for Whitfieldiella,
Praebarrettia, Barrettia and Laluzia armini are
taken from G�tz and Mitchell (2009).
Holotype of Laluzia
armini (reproduced
from G�tz and Mitchell,
2009, Fig. 6) shown for interpretation of angles a (P2-P1-PM) and b (P1-PM-PT). |
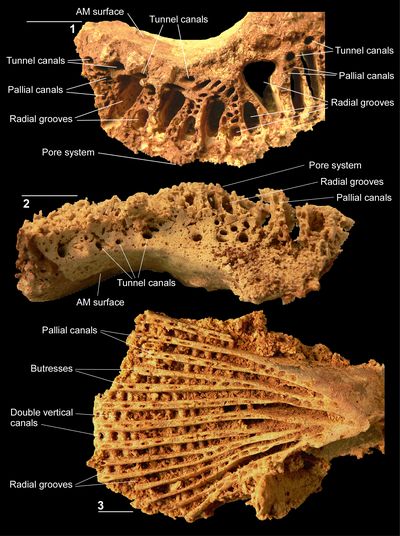
The pore system of C. muellerriedi has also been illustrated in a specimen from the Guinea Corn Formation of Jamaica (Mitchell, 2010). This specimen shows a pore system very similar to that of the Puerto Rican material described here. The inner zone of the LV is characterized by irregular sub-polygonal pores with five or six pores arranged concentrically above each radial canal; the limbal zone has radially arranged lines of radially orientated pores. Both the central zone pores and the limbal zone pores have denticles, which divide central pores into three or four porules, and limbal pores into two porules. The only difference between the pore systems of the Puerto Rican and Jamaican specimens is in the number of pores developed concentrically above the radial canals of the inner zone of the LV; 3 or 4 in the former, and 5 or 6 in the latter. The Jamaican specimen is also larger than the Puerto Rican specimens. The increased number of pores concentrically above each radial canal may be due to phylogenetic size increase passing from the early Maastrichtian (Puerto Rican material) to the mid Maastrichtian (Jamaican material). Caribbea maldonensis is still larger, but the pore system is unknown. Given the rarity with which pores are preserved in New World specimens, it is inadvisable to use this to define species, and in any case only a single specimen from Jamaica has the pore system preserved.
Five further species assigned to Orbignya, Hippurites or Caribbea have been erected in the Central American region. Chubb (1956) erected Hippurites (Orbignya) ceibarum for small forms from Jamaica. They occur with normal-sized specimens of muellerriedi, and other than being smaller, do not appear to differ in any significant way from that species. Hippurites perkinsi was erected by Myers (1968) for material from the Cardenas Formation which occurred together with specimens of C. muellerriedi. Myers' specimens differ from muellerriedi in having more prominent costae on the external surface of their RVs. Numerous additional specimens of H. perkinsi collected by Myers are preserved in the Texas Memorial Museum, Austin, Texas, and were examined by SFM in 2009. Similar forms also occur in the Jerusalem Mountain Limestone in Jamaica. As discussed earlier, these forms are interpreted as phenotypic variants and H. perkinsi is placed in synonymy with C. muellerriedi here. Grubić (2004) erected Caribbea sladici for a specimen from Jamaica that had been illustrated by Trechmann (1924), based on its lack of orimentary folds. In populations of muellerriedi, the presence and strength of orimentary folds is variable and it is doubtful if forms that lack orimentary folds are anything other than morphological variants of C. muellerriedi.
In contrast, I point out that C. maldonensis is distinct, having a different outer shell structure (pustules rather than muri as described by Chubb, 1956, 1971) and a different range (mid Maastrichtian, as opposed to early to early mid Maastrichtian for C. muellerriedi) in the Jamaica (Mitchell, 1999; Steuber et al., 2002; Gunter & Mitchell, 2005). Given that juvenile growth stages of C. maldonensis have an identical RV outer shell layer microstructure to adult forms of C. muellerriedi, it seems likely that the former is the direct descendent of the latter and that differences in outer layer shell microstructure do not warrant the erection of a separate genus for maldonensis, a move that would artificially split a single evolving lineage.
Olsson (1934) erected Orbignya pacifica for a species from the Monte Grande Formation (upper Campanian - lowermost Maastrichtian?) of Peru. This form has multiple infolds and appears to be unrelated to Caribbea, it may represent Laluzia peruviana (Gerth).
Geographic and Stratigraphic Range. Caribbea muellerriedi is widely distributed in the Caribbean and Central American region. In Cuba it is reported from the Habana Formation of the province of Pinar del Rio (Vermunt, 1937; Mac Gillavry, 1937) which I attribute to the early Maastrichtian based on its association with Laluzia. In Jamaica the species is widely distributed in the Jerusalem Mountain Limestone (lowermost upper Maastrichtian), the Titanosarcolites limestone of the Marchmont Inlier (Chubb, 1971) and the lower part (A to Upper C Beds) of the Guinea Corn Formation (Mitchell, 1999, 2010). In Puerto Rico it occurs in the El Rayo Formation which is attributed to the lower Maastrichtian (Mitchell et al., 2011). In Mexico, it is recorded from the Cardenas Formation (Myers, 1968; Pons et al., 2013) which has been dated by ammonites and Sr-isotope values to the lower to lower upper Maastrichtian (Ifrim et al., 2005; Schafhauser et al., 2007). Caribbea muellerriedi is therefore a widespread early to early mid Maastrichtian species.
Genus Laluzia G�tz & Mitchell, 2009
=Gloria Grubić, 2004, type species Gloria vermunti Grubić, 2004 (=Pironaea cf. peruviana Vermunt, 1937), from the Habana Formation of the Pinar del Rio Province, Cuba; non Gloria Barrande, 1881, which is a mid-Palaeozoic cryptodont bivalve.
Type species. Laluzia armini G�tz & Mitchell, 2009, from the lower Maastrichtian Cardenas Formation of San Luis Potosi, Mexico.
Diagnosis. A hippuritid with three primary infolds (P0, P1 and P2) and multiple secondary infolds. The PT and AT have ridges that fit into channeled grooves (sockets) on the CT and inner shell layer/PM lamella of the RV. The PM fits into an embayment in the inner shell layer that is separated from the body cavity by a lamella; the anterior myophore is attached directly to the inner shell layer. The outer shell layer of the RV is composed of funnel plates that are separated by irregular dendritic radial muri. Pallial canals are absent from the inner shell layer of the LV. Vertical canals penetrate the outer zone of the LV above the rays, and radial canals occur in the inner zone. The pore system consists of irregularly distributed, thick-walled irregular pores.
Discussion. The genus Laluzia was erected for a small hippuritid that lacks pallial canals in the inner layer of its LV and had a myocardinal arrangement different from that of Praebarrettia, the only other American multiple-ray hippuritid that lacks pallial canals in its LV. At that time, the affinity of the various forms referred to Pironaea (that is, P. peruviana Gerth and P. cf. P. peruviana Vermunt) from the Americas was unknown. With the new material from Puerto Rico described here which includes details of the pore system, and which I regard as undoubtedly congeneric with Laluzia, differences can be pointed out between Laluzia and Caribbea. In Laluzia, the teeth fit into slots developed from the body cavity lining just as in Caribbea. But Caribbea and Laluzia differ in many other respects: in Laluzia the PM fits into a socket whereas in Caribbea it fits into an embayment; Laluzia and Caribbea have differences in PM locations, LV buttress formation, pore systems and secondary infolds. So, whereas the 'sockets' in the RV of Laluzia and Caribbea are similar and may indicate a common ancestry, other details indicate that the two genera have already significantly diverged.
Pons et al. (2013, 2019) considered that Laluzia was a junior synonym of Praebarrettia Trechmann. However, the myocardinal arrangements of Laluzia and Praebarrettia are different, with Laluzia resembling that of Barrettia and Praebarrettia resembling that of Whitfieldiella (Dommelen, 1971; G�tz & Mitchell, 2009; Mitchell, 2010). Furthermore, the outer shell layer of Laluzia consists of funnel plates with radial muri; in contrast the outer shell layer of the RV of Praebarrettia consists of funnel plates separated by pustules. Although Caribbea shows an evolutionary development of pustules from radial muri, the differences in the myocardinal arrangements are sufficient to justify separation at the generic level. The pore systems of the two genera are also different: in Laluzia, pores are randomly distributed across the reticulum; whereas in Praebarrettia, they are highly ordered occurring on sieve plates directly above the transverse canals (Dommelen, 1971). The stratigraphic distribution of the two genera is also different (Laluzia in the early Maastrichtian and Praebarrettia in the late Maastrichtian). It is possible that Laluzia evolved into Praebarrettia, but this would have required changes in both the position of the PM and the pore system. It seems advisable at the present time to maintain two separate genera.
Laluzia peruviana (Gerth, 1928)
(Figs. 9.1-7 ![]() ,
10.1-6
,
10.1-6 ![]() ,
11.1-6
,
11.1-6 ![]() ,
12
,
12 ![]() )
)
. 1928 Pironaea peruviana: Gerth, p. 235, Figs. 2-3.
v. 1937 Pironaea sp. cf. P. peruviana Gerth; Vermunt, p. 266, Fig. 2i-j.
v. 2004 Gloria vermunti spec. nov.; Grubić, p. 150, Pl. 2, fig. 5.
. 2004 Gloria peruviana (Gerth); Grubić, p. 150, Pl. 2, fig. 7.
. 2004 Praebarrettia sparcilirata (Whitfield, 1897); Philip and Jaillard, p. 45, Pl. 3, figs. 1-2.
. 2013 Praebarrettia sparcilirata (Whitfield) sensu lato, larger morphotype; Pons et al., p. 742-748, Figs. 19.1-10, 21.1-6, 23.1-6.
Type. The material described as Pironaea peruviana by Gerth (1928) is missing from the collections held by Naturalis in Leiden, The Netherlands. However, Gerth's illustrations are sufficient to suggest affinity and a neotype should be selected from Peru.
Material. NS6 (USNM 547519) (locality 1202-3), NS11 (USNM 547520) (locality 1202-3), NS12 (USNM 547521) (locality 1202-3), NS34 (USNM 547522) (locality 1202, NSNM 187687), NS36 (USNM 547523) (locality 1202-10), NS40 (USNM 547549) (locality 1202), NS41 (USNM 547524) (locality 1202-33), NS45 (USNM 547525) (locality 1202-5), NS48 (USNM 547526) (locality 1202-7), NS54 (USNM 547527) (locality 1202), NS48 (USNM 547528) (locality 1202-7), NS49 (USNM 547529) (locality 1202 45), NS51 (USNM 547530) (locality 1202), NS52 (USNM 547531) (locality 1202), NS53 (USNM 547532) (locality 1202), NS66 (USNM 547533) (locality 1202-5 or 17?), and NS73 (USNM 547534) (draw 118, locality 1202). All from the El Rayo Formation, Puerto Rico.
Diagnosis. A large species of Laluzia with broad, short secondary infolds.
Description. RV small to medium sized, conical to cylindrical. External surface with
many rounded costae and in some specimens vertically striated (Fig.
9.6 ![]() ), no growth
lines. The larger furrows correspond to infolds of the outer shell layer. The
outer shell layer is composed of funnel plates that are orientated outwards and
upwards at an angle of about 50�. The funnel plates are ornamented with radial
muri (Fig. 9.1, 9.3
), no growth
lines. The larger furrows correspond to infolds of the outer shell layer. The
outer shell layer is composed of funnel plates that are orientated outwards and
upwards at an angle of about 50�. The funnel plates are ornamented with radial
muri (Fig. 9.1, 9.3 ![]() ). The outer shell layer is folded into the inner shell
layer as a series of rays that may number up to 15 (Fig. 9.2,
9.7
). The outer shell layer is folded into the inner shell
layer as a series of rays that may number up to 15 (Fig. 9.2,
9.7 ![]() ). The body
chamber (body cavity lining) is formed by a continuous lamina that represents
the inner shell layer, and this lamina rises above the apertural surface of the
inner layer as a pronounced wall (Fig. 9.2
). The body
chamber (body cavity lining) is formed by a continuous lamina that represents
the inner shell layer, and this lamina rises above the apertural surface of the
inner layer as a pronounced wall (Fig. 9.2 ![]() ). The inner shell layer is filled
with large partitions formed by vertical radial plates extending from the rays
and horizontal tabulae (Fig. 9.2
). The inner shell layer is filled
with large partitions formed by vertical radial plates extending from the rays
and horizontal tabulae (Fig. 9.2 ![]() ). The central tooth has two laminae each on the
anterior and posterior sides that form slots for the PT and AT to fit into; two
thin laminae project from the interior wall both posterior (that is on the
lamella separating the PM cavity from the body cavity) and anterior of the
central tooth to create two further slots for the AT and PT to fit into (Fig.
9.1-2
). The central tooth has two laminae each on the
anterior and posterior sides that form slots for the PT and AT to fit into; two
thin laminae project from the interior wall both posterior (that is on the
lamella separating the PM cavity from the body cavity) and anterior of the
central tooth to create two further slots for the AT and PT to fit into (Fig.
9.1-2 ![]() ). The PM cavity is separated from the body cavity by a thin lamella
(Fig. 9.1-2
). The PM cavity is separated from the body cavity by a thin lamella
(Fig. 9.1-2 ![]() ).
).
During ontogeny, P1 and P2 develop first, then incipient infoldings develop, but the ligamental ray does not develop until later in ontogeny. This suggests that the ligamental infold is an acquired character and not an ancestral character in Laluzia (a similar situation occurs in the upper Maastrichtian in an undescribed large species of Caribbea).
The LV is flat to gently domed and does not contain pallial canals
(Fig. 9.5 ![]() ). The myocardinal yoke is continuous and elliptical
(Figs. 10.1-2, 10.4-6
). The myocardinal yoke is continuous and elliptical
(Figs. 10.1-2, 10.4-6 ![]() ,
11.1, 11.5-6
,
11.1, 11.5-6 ![]() ). The PT is about half the size of the AT
(Fig. 10.4-6
). The PT is about half the size of the AT
(Fig. 10.4-6 ![]() ). The teeth
are concentrically blade-like. The PM is blade like and situated on a buttress
extending posterior-dorsally from the PT (Figs. 10.4, 10.6
). The teeth
are concentrically blade-like. The PM is blade like and situated on a buttress
extending posterior-dorsally from the PT (Figs. 10.4, 10.6 ![]() ,
11.1, 11.3, 11.6
,
11.1, 11.3, 11.6 ![]() ).
The AM is a surface that is tilted outwards extending around a third of the yoke,
it extends around the anterior side of the AT (Figs. 10.1-2,
10.4-6
).
The AM is a surface that is tilted outwards extending around a third of the yoke,
it extends around the anterior side of the AT (Figs. 10.1-2,
10.4-6 ![]() ,
11.1, 11.6
,
11.1, 11.6 ![]() ). The yoke is supported by very prominent buttresses, which are much more
prominent that those in Caribbea. The
buttresses are single, or bifurcate once. The yoke is penetrated by openings on
the adapical side of the AM (Fig. 10.2, 10.5
). The yoke is supported by very prominent buttresses, which are much more
prominent that those in Caribbea. The
buttresses are single, or bifurcate once. The yoke is penetrated by openings on
the adapical side of the AM (Fig. 10.2, 10.5 ![]() ). Between each pair of buttresses,
a very irregular line of double pores penetrates the LV. The terminal pore is
broadly rounded, and the other pores are irregular and alternate with each other
(Fig. 10.1
). Between each pair of buttresses,
a very irregular line of double pores penetrates the LV. The terminal pore is
broadly rounded, and the other pores are irregular and alternate with each other
(Fig. 10.1 ![]() ). The adapical side of the left valve bears radial canals in the
umbonal (inner) part of the shell (Fig. 10.3
). The adapical side of the left valve bears radial canals in the
umbonal (inner) part of the shell (Fig. 10.3 ![]() ). The radial canals are overlain by
a fine reticulate mesh that has denticles (Figs. 10.3
). The radial canals are overlain by
a fine reticulate mesh that has denticles (Figs. 10.3 ![]() ,
11.2, 11.4-5
,
11.2, 11.4-5 ![]() ). The radial
canals pass down to the round canal at the end of the irregular pore system (Fig.
10.3
). The radial
canals pass down to the round canal at the end of the irregular pore system (Fig.
10.3 ![]() ). Where the canals penetrate the LV, lateral canals are developed.
Lateral openings extend concentrically from the radial canals into the
inter-radial areas. The inter-radial areas are covered by the same reticulate
covering as the radial canals (Fig. 11.2, 11.4-5
). Where the canals penetrate the LV, lateral canals are developed.
Lateral openings extend concentrically from the radial canals into the
inter-radial areas. The inter-radial areas are covered by the same reticulate
covering as the radial canals (Fig. 11.2, 11.4-5 ![]() ). P1 has a round oscule in the
LV and P2 has an elongate oval oscule that is connected to the rim (at least in
young forms; Figs. 10.3-4
). P1 has a round oscule in the
LV and P2 has an elongate oval oscule that is connected to the rim (at least in
young forms; Figs. 10.3-4 ![]() ,
11.1, 11.6
,
11.1, 11.6 ![]() ). A fine mesh extends across part of the
opening for P2 suggesting it may have been covered by a reticulate mesh (Fig.
11.5
). A fine mesh extends across part of the
opening for P2 suggesting it may have been covered by a reticulate mesh (Fig.
11.5 ![]() ).
).
From the arrangement of the myocardinal yoke with the interior surface of the body chamber, the AM must have been connected to the interior of the body cavity, much as in Caribbea.
|
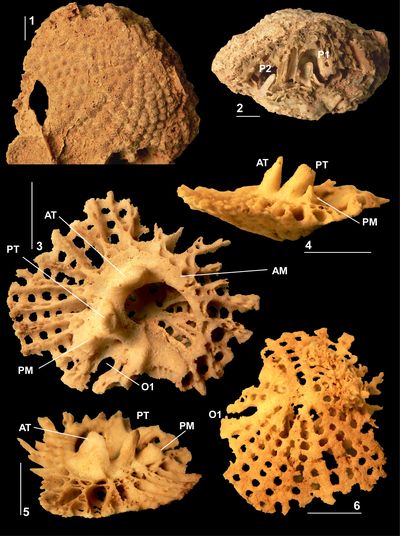
|
Figure
9:
Laluzia
peruviana (Gerth, 1928), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 9.1 (NS12: USNM 547521),
incomplete RV showing pillars, sockets for teeth, and enclosed socket for
posterior myophore. 9.2 (NS40: USNM 547549),
juvenile RV, showing sockets for teeth and posterior myophore and infolds. 9.3
(NS51: USNM
547530), incomplete
RV, adapical view, showing infolds. 9.4 (NS52: USNM 547531),
incomplete RV, adapical view, showing infolds. 9.5 (NS34: USNM 547522),
articulated shell, showing the anterior tooth fitting into its socket and the
anterior myophore that would attach directly onto the inner shell layer of the
RV. 9.6 (NS49: USNM 547529)
articulated shell showing longitudinal ribs on RV. 9.7 (NS48: USNM 547526),
RV, adapical view showing infolds. P0, ligamentary infolding; P1 and P2,
first and second pillars, ATS, anterior tooth socket; PTS, posterior tooth
socket, PMS, posterior myophore socket;
BCL, body cavity lining. Scale bar = 10
mm. |
|
|
Figure
10:
Laluzia
peruviana (Gerth, 1928), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 10.1 (NS45: USNM 547525),
LV, adapical view showing myocardinal arrangement and buttresses. 10.2-5 (NS66:
USNM
547533), LV,
various views showing myocardinal arrangement and buttresses: note the tunnels
through the myocardinal yoke visible in 10.2 and 10.5; 10.3,
abapical view showing radial furrows. 10.6 (NS41: USNM 547524),
LV, oblique view showing myocardinal arrangement and buttresses. O1 and O2,
oscules for first and second pillars. AT, anterior tooth; PT, posterior tooth;
AM, anterior myophore; PM, posterior myophore. Scale bar = 10 mm. |
|
|
Figure
11:
Laluzia
peruviana (Gerth, 1928), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 11.1-2 (NS36: USNM 547523),
LV: 11.1,
adapical view showing myocardinal arrangement and buttresses; 11.2,
abapical view showing pores and porules. 11.3-4 (NS11: USNM 547520),
incomplete LV; 11.3,
adapical view showing myocardinal arrangement and buttresses; 11.4,
abapical view showing pores and porules. 11.5-6 (NS41: USNM 547524),
LV (same as Fig. 9.6): 11.5,
abapical view showing pores and porules; 11.6,
adapical view showing myocardinal arrangement and buttresses. O1 and O2, oscules
for first and second pillars, AT, anterior tooth; PT, posterior tooth; AM,
anterior myophore; PM, posterior myophore. Scale bar = 10 mm. |
Discussion. Grubić
(2004, p. 151) stated that the type specimen of Pironaea
peruviana, which came from the middle member of the La Mesa Formation of
Peru (Gerth, 1928; Philip & Jaillard,
2004), is missing
from the collections at Naturalis, Leiden. I also could not find it in the same
collection in 2010 although I made a thorough search. New material, collected
from the upper member of the La Mesa Formation, Peru, was described under the
name Praebarrettia sparcilirata (Whitfield) by Philip and Jaillard
(2004). They considered that their material was synonymous not only with 'Pironaea'
peruviana, but also with Praebarrettia
sparcilirata and Praebarrettia porosa
Palmer. The age of these deposits, based on ammonites and bivalves, is
either upper Campanian or lower Maastrichtian (Philip & Jaillard,
2004; Dhondt & Jaillard,
2005). Philip and Jaillard
(2004, Pl. 3, figs. 1-2) illustrated a RV that conforms to the population of L.
peruviana from Puerto Rico. When plotted on scatter plots for myocardinal
arrangements (Fig. 8 ![]() ), the upper La Mesa specimen plots in the same field as
the Puerto Rican and Cardenas specimens. Although no LV was figured, it was
described (Philip & Jaillard,
2004, p. 11) as follows:
"Typical
radial canals covered with a thin, more or less continuous reticulate layer of
small pores (0.3-0.5 mm.). Transverse furrows described by Dommelen (1971) not observed in our specimens". This description is also comparable
with the material from Puerto Rico. I therefore consider that the type specimen
of Pironaea peruviana illustrated by Gerth
(1928) and the material described by Philip and Jaillard
(2004) as
Praebarrettia sparcilirata (Whitfield)
are the same species and are identical to the material described here from
Puerto Rico; all are assigned to Laluzia
peruviana here. Pons
et al. (2013)
described a series of specimens from the C�rdenas Formation in Mexico as Praebarrettia
sparcilirata (Whitfield) sensu
lato and recognized two successive morphotypes in the early Maastrichtian.
Their smaller morphotype is the earlier form and corresponds to Laluzia
armini G�tz & Mitchell, whereas their larger morphotype
is equivalent to L. peruviana. In
contrast to Philip and Jaillard
(2004) and Pons et
al. (2013), I recognize both Praebarrettia
porosa Palmer and P.
sparcilirata as separate species and maintain them in a distinct genus,
based on differences in their myocardinal-pillar arrangements and pore systems (Dommelen,
1971; G�tz & Mitchell,
2009; Fig. 8
), the upper La Mesa specimen plots in the same field as
the Puerto Rican and Cardenas specimens. Although no LV was figured, it was
described (Philip & Jaillard,
2004, p. 11) as follows:
"Typical
radial canals covered with a thin, more or less continuous reticulate layer of
small pores (0.3-0.5 mm.). Transverse furrows described by Dommelen (1971) not observed in our specimens". This description is also comparable
with the material from Puerto Rico. I therefore consider that the type specimen
of Pironaea peruviana illustrated by Gerth
(1928) and the material described by Philip and Jaillard
(2004) as
Praebarrettia sparcilirata (Whitfield)
are the same species and are identical to the material described here from
Puerto Rico; all are assigned to Laluzia
peruviana here. Pons
et al. (2013)
described a series of specimens from the C�rdenas Formation in Mexico as Praebarrettia
sparcilirata (Whitfield) sensu
lato and recognized two successive morphotypes in the early Maastrichtian.
Their smaller morphotype is the earlier form and corresponds to Laluzia
armini G�tz & Mitchell, whereas their larger morphotype
is equivalent to L. peruviana. In
contrast to Philip and Jaillard
(2004) and Pons et
al. (2013), I recognize both Praebarrettia
porosa Palmer and P.
sparcilirata as separate species and maintain them in a distinct genus,
based on differences in their myocardinal-pillar arrangements and pore systems (Dommelen,
1971; G�tz & Mitchell,
2009; Fig. 8 ![]() herein).
herein).
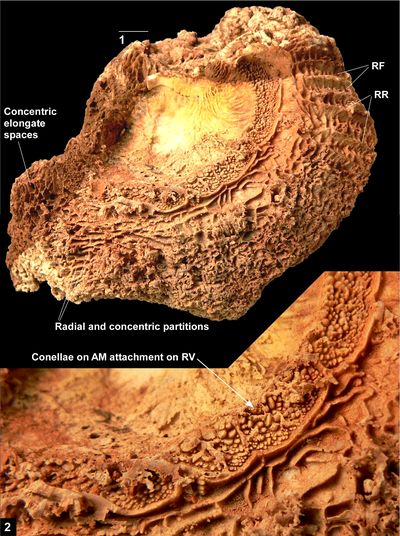
Pironaea sp. cf. P.
peruviana from Cuba (Vermunt, 1937), on which Grubić
(2004) erected Gloria vermunti, shows a very similar morphology
(Fig. 12 ![]() ) and has a
fine costation developed on the exterior surface of the RV. Pironaea
sp. cf. P. peruviana is associated with Caribbea
muellerriedi (Vermunt, 1937), and also seems to be conspecific with
the Puerto Rican material of L. peruviana.
) and has a
fine costation developed on the exterior surface of the RV. Pironaea
sp. cf. P. peruviana is associated with Caribbea
muellerriedi (Vermunt, 1937), and also seems to be conspecific with
the Puerto Rican material of L. peruviana.
|
|
Figure
12: New
photograph of Pironaea sp. cf. P.
peruviana from
Cuba (Vermunt, 1937), NLM.P240-1953, RV, transverse cross-section, showing short rounded
infolds and position of posterior myophore socket. Scale bar = 10 mm. |
Laluzia armini shows a similar myocardinal-pillar arrangement to L. peruviana but differs in the nature of the primary and secondary infolds. In L. armini, the primary and secondary infolds are narrow and comparatively elongate. In contrast, the infolds of L. peruviana are broad and short. Because the infold morphology in populations overlaps little, the two species appear to be separate. It is notable that L. peruviana occurs in both the Cardenas Formation (Mitchell, pers. obs.) and El Rayo Formation, where it is associated with a highly diverse, normal-shelf rudist assemblage; yet L. armini occurs in a low-diversity rudist association in the Cardenas Formation at La Luz which was interpreted as a relatively deep-water low-light environment (G�tz & Mitchell, 2009). It seems likely, therefore, that the two species may have been adapted to different environments or have been ancestor (L. armini) and descendant (L. peruviana).
'Pironaea' corrali Palmer, 1933, is a much more difficult form to interpret. The RV has a small number of infolds that range from non-moniliform to submoniliform. On the holotype the LV buttresses are weathered, and it is not possible to determine the myocardinal arrangement. I have not been able to work out the form of the myocardinal arrangement in any specimens of this species that I have seen (including material in Naturalis, The Netherlands, and the National Natural History Museum, Havana), and therefore its affinity and phylogenetic position remain unclear. New material will have to be analysed to determine the affinity of this form, and this I consider is one of the outstanding problems in understanding New World hippuritids.
Geographical and Stratigraphical Distribution. Laluzia peruviana is moderately widely distributed in lower Maastrichtian rudist assemblages of the American region. It is recorded from: the El Mesa Formation (upper Campanian, or more likely lower Maastrichtian) of Peru (Philip & Jaillard, 2004); the Habana Formation sensu lata (the specific occurrence which I regard as likely to be of early Maastrichtian age) from Pinar del Rio, Cuba (Vermunt, 1937), the El Rayo Formation (lower Maastrichtian) of Puerto Rico (recorded here), and the Cardenas Formation, near Rayon, San Luis Potosi State, Mexico (Myers, 1968; Myers Collection, Texas Memorial Museum, examined by SFM in 2009; Pons et al., 2013).
The only other described species, Laluzia armini, occurs in the Cardenas Formation (lower Maastrichtian) at La Luz, State of San Luis Potosi, Mexico (G�tz & Mitchell, 2009). Consequently, Laluzia appears to be a good biostratigraphic marker for the early Maastrichtian of the Central American region but could also be present in some upper Campanian assemblages of Peru.
Barrettia Clade
Diagnosis. Barrettiinae that have pallial canals in the inner layers of their LVs and a pore system in the LVs.
Remarks. Pallial canals are also present in the LV inner shell layers of Torreites. The absence of pallial canals in the LV of the Caribbea clade could, therefore, be a derived character. The clade includes three genera: Barrettia Woodward, 1862, Whitfieldiella Mitchell, 2010, and Parastroma Douvill�, 1926.
Genus Parastroma Douvill�, 1926
Type species. Parastroma sanchezi Douvill�, 1926, from the Campanian of Arroyo Hondo, Camag�ey Province, Cuba.
Diagnosis. A hippuritid with two primary infolds (P1 and P2) but with the multiple infolds represented by incipient rays. The PT and AT have ridges that fit into channeled grooves (sockets) on the CT and inner shell layer/PM lamella of the RV. The PM fits into a socket formed from the inner shell material of the RV; the anterior myophore is attached to a platform developed in the inner shell layer of the RV. The inner shell layer is composed of three elements of variable development: radial partitions with or without connellae, inter-radial partitions with or without connellae, and tabulae. Pallial canals are present in the inner layer of the LV. Vertical canals penetrate the outer zone of the LV and above each canal a sieve plate with a denticulate pore system is developed.
Discussion. Parastroma is distinguished from the other multiple-folded hippuritids by its lack of well-defined rays and instead contains only incipient rays. Interestingly, Parastroma shares its arrangement of myocardinal angles with Whitfieldiella but has a pore system similar to that of Barrettia. Until material is discovered from the Santonian (or older) it is not possible to sort out the evolutionary history of this clade at the present time. However, the silicified material from Puerto Rico allows the first detailed description of a species belonging to this clade.
Parastroma guitarti (Palmer, 1933)
(Figs. 13.1-2 ![]() ,
14.1-6
,
14.1-6 ![]() ,
15.1-3
,
15.1-3 ![]() ,
16.1-3
,
16.1-3 ![]() ,
17.1-4
,
17.1-4 ![]() )
)
*. 1933 Orbigna guitarti Palmer n. sp.: Palmer, p. 96, Pl. 1, fig. 1; Pl. 2, fig. 2; Pl. 3, fig. 1. (corrected to Orbignya in pen by the author in offprints).
v. 1937 Parastroma guitarti (Palmer); Mac Gillavry, p. 126, Pl. 5, figs. 1, 5; Pl. 10, figs. 5-9.
v. 1971 Parastroma guitarti (Palmer); Dommelen, p. 94, Figs. 8B, 31(1-5); Pl. 11, figs. 1-2; Pls. 12-15.
v. 2004 Parastroma guitarti (Palmer); Grubić, p. 168, Pl. 9, fig. 2.
Types. Palmer (1933) figured three specimens (syntypes) of the species from Sancti Spiritus, Santa Clara Province, Cuba. His Pl. 1, fig. 1, is a view of a specimen in profile, his Pl. 2, fig. 2, is a view of a specimen showing the myocardinal arrangement, and his Pl. 3, fig. 1, shows the pore system. The specimen figured in his Pl. 2, fig. 2, shows the distinctive features of the species and is clearly the most desirable to use in stabilizing the species concept; but the whereabouts of this specimen is not known and it cannot be designated as lectotype here.
Material. Including NS37 (USNM 547535) (locality 1202-10), NS38 (USNM 547536) (locality 1202-25), NS42 (USNM 547537) (locality 1202), NS43 (locality 1202), NS44 (USNM 547538) (locality 1202-3), NS46 (USNM 547539) (locality 1202), NS47 (USNM 547540) (locality 2000), NS49 (USNM 547541) (locality 1202-5), NS50 (USNM 547542) (locality 1202), NS60 (USNM 547543) (locality 1202), NS61 (USNM 547544) (locality 1202), NS62 (USNM 547545) (locality 1202), NS63 (USNM 547546) (locality 1202), and NS72 (USNM 547547) (locality 1202, very nice pores). All from the El Rayo Formation, Puerto Rico.
Diagnosis. A species of Parastroma in which deep conical folds (concentric spaces) are present in the complex inner shell layer of the RV and the crests of these folds are sharp with reduced or without conellae. LV buttresses contain a single line of pallial canals, except where buttresses bifurcate.
Description. The RV is large
and generally cylindro-conical, increasing rapidly in size. Its external surface
is marked by regular narrow furrows with flat-faced to gently rounded costae
between. The outer shell layer is thin, but its structure is not preserved in
the silicified material. The complex inner shell layer is composed of irregular,
broadly rectangular concentric spaces (Figs. 13.1-2 ![]() ,
14.1-2
,
14.1-2 ![]() ), with each space
being open abapically and narrowing adapically to a point. The abapical tops of
the walls of the concentric spaces are sharp or very weakly beaded (conellae-bearing).
Tabulae insertion in the inner shell layer produces a stack of cones giving the
typical chevron patterns seen in tangential and radial longitudinal sections. In
juveniles the body cavity occupies about one third of the diameter, but the
proportionate size of the body cavity decreases with growth. The apertural
surface is represented by radially directed ribs and furrows that are formed by
the variable height of the chevron structure, and commonly the radial partitions
correspond with the radial ribs (Fig. 13.1
), with each space
being open abapically and narrowing adapically to a point. The abapical tops of
the walls of the concentric spaces are sharp or very weakly beaded (conellae-bearing).
Tabulae insertion in the inner shell layer produces a stack of cones giving the
typical chevron patterns seen in tangential and radial longitudinal sections. In
juveniles the body cavity occupies about one third of the diameter, but the
proportionate size of the body cavity decreases with growth. The apertural
surface is represented by radially directed ribs and furrows that are formed by
the variable height of the chevron structure, and commonly the radial partitions
correspond with the radial ribs (Fig. 13.1 ![]() ). P0
is not present; P1 is
represented by an isolated circular body of outer shell material, and P2 is
represented by an isolated radially elongated body of outer shell material.
Consequently, the two pillars are only connected to the outer shell at the point
where they developed in juveniles. The two pillars rise high above the apertural
face so that they can reach toward the openings on the LV (Fig.
15.2
). P0
is not present; P1 is
represented by an isolated circular body of outer shell material, and P2 is
represented by an isolated radially elongated body of outer shell material.
Consequently, the two pillars are only connected to the outer shell at the point
where they developed in juveniles. The two pillars rise high above the apertural
face so that they can reach toward the openings on the LV (Fig.
15.2 ![]() ). The
sockets for the PT and AT are embayments on the margin of the body cavity (Fig.
14.1-2
). The
sockets for the PT and AT are embayments on the margin of the body cavity (Fig.
14.1-2 ![]() ). The socket for the PM is tear-shaped and fully embedded within the
complex inner shell layer being situated dorso-posteriorly of the PT and P1 (Fig.
13.1
). The socket for the PM is tear-shaped and fully embedded within the
complex inner shell layer being situated dorso-posteriorly of the PT and P1 (Fig.
13.1 ![]() ). The socket for the AM is represented by a broad platform developed
in the inner margin of the complex inner shell layer that extends from the area
opposite the dorsal side of the AT around almost half the circumference of the
body cavity as far as P2 (Figs. 13.1-2
). The socket for the AM is represented by a broad platform developed
in the inner margin of the complex inner shell layer that extends from the area
opposite the dorsal side of the AT around almost half the circumference of the
body cavity as far as P2 (Figs. 13.1-2 ![]() ,
14.1-2
,
14.1-2 ![]() ). The surface of the AM socket is
covered with strongly developed coarse connellae (Fig. 13.2
). The surface of the AM socket is
covered with strongly developed coarse connellae (Fig. 13.2 ![]() ); similar connellae
are seen on the surface of the PM in specimens from Cuba preserved in the
collections of the USNM in Washington and the National Natural History Museum in
Havana. Such structures would thereby present a salient vertical micro-relief
for adductor muscle insertion similarly to those presented by the laminar
vertical ridges shown in the RV AM of an exceptionally well-preserved specimen
of Pseudovaccinites gosaviensis (Douvill�)
illustrated in Skelton (2018, Fig. 10a-b). Such rotations of the
adductor muscle insertion surfaces can be explained as a constructional
adjustment to overcome the constraint on shell accretion normal to insertion
surfaces due to the histological mode of muscle insertion (Skelton,
2018, p.
13-15).
); similar connellae
are seen on the surface of the PM in specimens from Cuba preserved in the
collections of the USNM in Washington and the National Natural History Museum in
Havana. Such structures would thereby present a salient vertical micro-relief
for adductor muscle insertion similarly to those presented by the laminar
vertical ridges shown in the RV AM of an exceptionally well-preserved specimen
of Pseudovaccinites gosaviensis (Douvill�)
illustrated in Skelton (2018, Fig. 10a-b). Such rotations of the
adductor muscle insertion surfaces can be explained as a constructional
adjustment to overcome the constraint on shell accretion normal to insertion
surfaces due to the histological mode of muscle insertion (Skelton,
2018, p.
13-15).
The LV is generally gently to strongly domed (Fig.
15.1-2, 15.4 ![]() ). The
myocardinal yoke varies in development around the valve; it forms the support
for the PM, teeth and AM (Figs. 14.3
). The
myocardinal yoke varies in development around the valve; it forms the support
for the PM, teeth and AM (Figs. 14.3 ![]() ,
15.3
,
15.3 ![]() ). The MP is a low blade-like,
radially elongated plate that is situated on a strong buttress supporting the
myocardinal yoke that extends along the dorsal margin of the oscule for P1 (Figs.
14.3
). The MP is a low blade-like,
radially elongated plate that is situated on a strong buttress supporting the
myocardinal yoke that extends along the dorsal margin of the oscule for P1 (Figs.
14.3 ![]() ,
15.3
,
15.3 ![]() ). The myophore expands rapidly adapically giving it concave sides
when viewed in concentric profile (Fig. 15.4
). The myophore expands rapidly adapically giving it concave sides
when viewed in concentric profile (Fig. 15.4 ![]() ), but convex sides when viewed in
radial profile (Fig. 15.5
), but convex sides when viewed in
radial profile (Fig. 15.5 ![]() ). It is about half the length of the AT and PT
(Fig. 15.4-5
). It is about half the length of the AT and PT
(Fig. 15.4-5 ![]() ). Either the PT or the AT may be slightly larger than the other; both are
concentrically expanded and may be more-or-less rectangular or crudely pointed (Figs.
14.3
). Either the PT or the AT may be slightly larger than the other; both are
concentrically expanded and may be more-or-less rectangular or crudely pointed (Figs.
14.3 ![]() ,
15.4-5
,
15.4-5 ![]() ,
17.1-2
,
17.1-2 ![]() ). The AM curves around the dorsal side of the AT and forms
an extensive platform that encircles about half the myocardinal yolk (Figs.
14.3
). The AM curves around the dorsal side of the AT and forms
an extensive platform that encircles about half the myocardinal yolk (Figs.
14.3 ![]() ,
15.3
,
15.3 ![]() ). The AM is flat or gently inwardly facing. A strong buttress extends
from the PT on the interior side of the oscule (O1) for P1 and then extends
between the openings for P1 (O1) and P2 (O2) and bifurcates twice passing
towards the valves margin (Figs. 14.3
). The AM is flat or gently inwardly facing. A strong buttress extends
from the PT on the interior side of the oscule (O1) for P1 and then extends
between the openings for P1 (O1) and P2 (O2) and bifurcates twice passing
towards the valves margin (Figs. 14.3 ![]() ,
15.3
,
15.3 ![]() ). A weaker buttress extends along
the anterior side of O2 and bifurcates at least once passing towards the valve's margin
(Fig. 14.3
). A weaker buttress extends along
the anterior side of O2 and bifurcates at least once passing towards the valve's margin
(Fig. 14.3 ![]() ). The inner parts of the buttresses supporting the
PM, extending between O1 and O2, and on the anterior side of O2 lack pallial
canals; canals only appearing level with the tips of the pillars. Small pallial
canals pass below the myocardinal yoke at the end of the radial furrows for O1,
O2 and the other radial furrows. The earliest formed radial canals appear to
open out into the cavity inside the yoke; this however appears to be because the
original shell material was very thin and has not been silicified; originally
they appear to have been blind. Extensive buttresses are developed around the
remainder of the shell and may bifurcate once or twice passing towards the
valves margin (Figs. 14.3
). The inner parts of the buttresses supporting the
PM, extending between O1 and O2, and on the anterior side of O2 lack pallial
canals; canals only appearing level with the tips of the pillars. Small pallial
canals pass below the myocardinal yoke at the end of the radial furrows for O1,
O2 and the other radial furrows. The earliest formed radial canals appear to
open out into the cavity inside the yoke; this however appears to be because the
original shell material was very thin and has not been silicified; originally
they appear to have been blind. Extensive buttresses are developed around the
remainder of the shell and may bifurcate once or twice passing towards the
valves margin (Figs. 14.3 ![]() ,
15.4
,
15.4 ![]() ). In larger specimens
(Fig. 16.3
). In larger specimens
(Fig. 16.3 ![]() ), further
bifurcations of the buttresses occur passing towards the valve margin, but the
number of divisions becomes progressively fewer with growth. The area inside the
myocardinal yoke is composed of compact shell material (Figs. 14.3
), further
bifurcations of the buttresses occur passing towards the valve margin, but the
number of divisions becomes progressively fewer with growth. The area inside the
myocardinal yoke is composed of compact shell material (Figs. 14.3 ![]() ,
15.34
,
15.34 ![]() ,
14.3
,
14.3 ![]() ), with
holes only evident where silicification has been incomplete. The adapical base
of the myocardinal yoke contains a single row of pallial canals extending around
the base of the AM, the PT and the PM (Figs. 15.4
), with
holes only evident where silicification has been incomplete. The adapical base
of the myocardinal yoke contains a single row of pallial canals extending around
the base of the AM, the PT and the PM (Figs. 15.4 ![]() ,
16.1
,
16.1 ![]() ). The pallial canals of
the myocardinal yoke are blind at their inner ends, and expand in diameter as
they pass both radially outwards and abapically to open in a circle marginal to
the base of the cardinal apparatus. Abapically of this row of pallial canals
around the length of the AM platform are a series of larger tunnel canals that
open out into the interior of the myocardinal yoke (Fig. 16.1-2
). The pallial canals of
the myocardinal yoke are blind at their inner ends, and expand in diameter as
they pass both radially outwards and abapically to open in a circle marginal to
the base of the cardinal apparatus. Abapically of this row of pallial canals
around the length of the AM platform are a series of larger tunnel canals that
open out into the interior of the myocardinal yoke (Fig. 16.1-2 ![]() ). The
buttresses are penetrated by a series of pallial canals that are blind at their
adapical end, but form a linear array of openings along the abapical side of
each buttress (Figs. 14.3
). The
buttresses are penetrated by a series of pallial canals that are blind at their
adapical end, but form a linear array of openings along the abapical side of
each buttress (Figs. 14.3 ![]() ,
15.3
,
15.3 ![]() ,
16.3
,
16.3 ![]() ). Along each buttress only a single layer
of pallial canal openings is present except where buttresses branch, where two
rows may be developed for a short distance (Figs. 14.3
). Along each buttress only a single layer
of pallial canal openings is present except where buttresses branch, where two
rows may be developed for a short distance (Figs. 14.3 ![]() ,
15.3
,
15.3 ![]() ,
16.3
,
16.3 ![]() ). Between the
buttresses radial grooves are developed on the abapical side of the LV,
which have broad, rounded inner ends adjacent to the myocardinal yoke (Figs.
14.3
). Between the
buttresses radial grooves are developed on the abapical side of the LV,
which have broad, rounded inner ends adjacent to the myocardinal yoke (Figs.
14.3 ![]() ,
15.3
,
15.3 ![]() ,
16.3
,
16.3 ![]() ). The tops of P1 and P2 lie at the interior ends of two of the
more prominent radial grooves. The grooves extend under the yoke and pass into
radial canals. Along the adapical side of each groove is a linear series of
rounded double-canals that penetrate the LV (Fig. 15.3, 15.6
). The tops of P1 and P2 lie at the interior ends of two of the
more prominent radial grooves. The grooves extend under the yoke and pass into
radial canals. Along the adapical side of each groove is a linear series of
rounded double-canals that penetrate the LV (Fig. 15.3, 15.6 ![]() ); these extend
along the entire length of the groove/radial canal. Adapically, these canals
expand to emerge in a low rectangular field on the adapical surface of the LV (Fig.
17.4
); these extend
along the entire length of the groove/radial canal. Adapically, these canals
expand to emerge in a low rectangular field on the adapical surface of the LV (Fig.
17.4 ![]() ). The rectangular field is overlain by a low to high domed sieve
sheet (Fig. 15.1-2
). The rectangular field is overlain by a low to high domed sieve
sheet (Fig. 15.1-2 ![]() ) with different scales of
trabeculae that gives rise to the
fine reticulate pore system (Fig. 17.3-4
) with different scales of
trabeculae that gives rise to the
fine reticulate pore system (Fig. 17.3-4 ![]() ). The sieve sheets are only separated
by the narrow walls of the rectangular field, and not by an imperforate zone.
Denticulate margins to the pores are present where the trabeculae have been
broken away or were not preserved. The pores and sieve sheets are uniform across
the entire surface of the LV. The two pillars have openings above them (oscules)
in the LV; the circular oscule for P1 seems to have been covered by a fine
reticulate mesh similar to the other openings as part of it is preserved in
places; details for P2 are not as clear and it may or may not have been covered
by a reticulate mesh.
). The sieve sheets are only separated
by the narrow walls of the rectangular field, and not by an imperforate zone.
Denticulate margins to the pores are present where the trabeculae have been
broken away or were not preserved. The pores and sieve sheets are uniform across
the entire surface of the LV. The two pillars have openings above them (oscules)
in the LV; the circular oscule for P1 seems to have been covered by a fine
reticulate mesh similar to the other openings as part of it is preserved in
places; details for P2 are not as clear and it may or may not have been covered
by a reticulate mesh.
|
|
Figure
13:
Parastroma
guitarti (Palmer, 1933), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 13.1-2 (NS42: USNM 547537),
RV, adapical view: 13.1,
inner shell layer construction; 13.2,
detail showing connellae in anterior myophore attachment area. RF, radial
furrows; RR radial ribs. Scale bar = 10 mm. |
|
|
Figure
14:
Parastroma
guitarti (Palmer, 1933), El Rayo Formation (lower Maastrichtian), Puerto Rico.
14.1 (NS37: USNM
547535), RV with
teeth of LV in place. 14.2 (NS63: USNM 547546),
RV with partial LV in place. 14.3 (NS62: USNM 547545),
LV, adapical view showing myocardinal arrangement and buttresses with pallial
canals. AT, anterior tooth; PT, posterior tooth; AM, anterior myophore, PM,
posterior myophore, O1 and O2, oscules for first and second pillars; (L), deep
groove corresponding to ligamental infold. Scale bar = 10 mm. |
|
|
Figure
15:
Parastroma
guitarti (Palmer, 1933), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 15.1-2 (NS50: USNM 547542),
articulated shell: 15.1,
LV, abapical view showing domed sieve sheets; 15.2,
view of flank showing pillars extending up into LV. 15.3-6 (NS38: USNM
547536), LV various views: 15.3,
adapical showing myocardinal elements, rounded double-canals, and buttresses
with single rows of pallial canals; 15.4,
flank view showing short PM; 15.5,
oblique view showing pallial canals in myocardinal yoke; 15.6,
abapical view showing rounded double-canals (sieve sheets not preserved). AT,
anterior tooth; PT, posterior tooth; Am, anterior myophore; PT, posterior
myophore; P1 and P2, first and second pillars; O1, oscule for first pillar.
Scale bar = 10 mm. |
|
|
Figure
16:
Parastroma
guitarti (Palmer, 1933), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 16.1-2 (NS60: USNM 547543),
fragment of LV (viewed from outside yoke) showing tunnel canals and pallial
canals. 16.2,
fragment of LV (viewed from inside yoke) showing tunnel canals. 16.3 (NS44:
USNM
547538), LV,
adapical view showing single rows of pallial canals along each buttress. Scale
bar = 10 mm. |
Discussion. The pore system of P. guitarti
has been previously described by Palmer (1933), Mac Gillavry
(1937) and Dommelen (1971). Dommelen's illustrations show only
the coarser trabeculae preserved in the pore system. The more poorly preserved
parts of the pore system illustrated here, which show only the coarser
trabeculae in each sieve sheet, resemble the illustrations of Dommelen,
whereas the better preserved areas in the silicified material show areas where
virtually all of the finest trabeculae are preserved (Fig. 17.3-4 ![]() ).
).
The pallial canals in the buttresses of P. guitarti are aligned in single rows that are only double where the buttresses branch. This contrasts with other multiple-rayed hippuritids. In Parastroma trechmanni (Chubb), two rows of pallial canals have been illustrated (e.g., Dommelen, 1971, Pl. 20) in agreement with extensive material collected from Jamaica in the University of the West Indies Geological Museum (UWIGM) collection. Other species of multiple-ray hippuritids, including Barrettia (e.g., Dommelen, 1971, Pl. 9, fig. 1; Pl. 10, fig.1), Whitfieldiella (material in UWIGM) and Parastroma sanchezi Douvill� (material in the National Natural History Museum, Havana, Cuba), have three or more rows of pallial canals present within the buttresses. It appears that during the evolution of the lineage of Parastroma the number of rows of pallial canals in buttresses decreases progressively: P. sanchezi (3 or 4 rows of pallial canals) to P. trechmanni (2 rows of pallial canals) to P. guitarti (1 row of pallial canals).
|
|
Figure
17:
Parastroma
guitarti (Palmer, 1933), El Rayo
Formation (lower Maastrichtian), Puerto Rico. 17.1-2 (NS45: USNM 547525),
LV: 17.1,
adapical view showing myocardinal elements, rounded double-canals, and
buttresses with single rows of pallial canals; 17.2,
oblique view showing pallial canals in myocardinal yoke. 17.3 (NS72: USNM
547547), LV,
abapical view, showing details of well-preserved sieve plates divided into pores
and porules by trabeculae. 17.4 (NS43: USNM 547550),
LV, abapical view, showing details of well-preserved sieve plates divided into
pores and porules by trabeculae. AT, anterior tooth; PT, posterior tooth; AM,
anterior myophore. Scale bar = 10 mm. |
This study indicates that a distinctive group of hippuritid rudists, the
Barrettiinae, evolved in the Upper Cretaceous of the New World. The group was characterized
by the development of distinctive socket morphologies for the two
teeth and suggests a monophyletic subfamily. This subfamily can be divided into
two distinctive clades, those that have pallial canals in the left valve (Barrettia,
Whitfieldiella and Parastroma)
and those that lack them (Caribbea, Praebarrettia
and Laluzia). The former group appeared in the late Santonian and ranged
into the early Maastrichtian, whereas the latter group appeared in the late
Campanian or early Maastrichtian and ranged up to the late Maastrichtian (Fig. 18 ![]() ). The lower Campanian of the New World also saw the appearance of members of
the Hippuritinae (Vaccinites) and the
Torreitinae (Torreities), with
previous Santonian ages now generally referred to the early Campanian (Mitchell,
2020), but neither subfamily persisted into the Maastrichtian of the New World.
). The lower Campanian of the New World also saw the appearance of members of
the Hippuritinae (Vaccinites) and the
Torreitinae (Torreities), with
previous Santonian ages now generally referred to the early Campanian (Mitchell,
2020), but neither subfamily persisted into the Maastrichtian of the New World.
|
|
Figure
18:
Stratigraphic distribution of selected hippuritid rudists from the Upper
Cretaceous of the Old World (from Steuber, et al.,
2016) and New World showing migration events of Old World taxa into the New
World, and the two separate hippuritid radiations in the New World. |
The pore systems are now known in at least one species of each New World barrettine genus: Whitfieldiella sp. (Mitchell, 2010), Barrettia monilifera (Mitchell, 2010), Parastroma guitarti (Palmer, 1933; Mac Gillavry, 1937; Dommelen, 1971; herein), Laluzia peruviana (herein), Caribbea muellerriedi (Mitchell, 2010; herein), and Praebarrettia sparcilirata (Dommelen, 1971). This demonstrates that pores are different in each genus and validates the separation of species based on features of the myocardinal arrangement as suggested by Mitchell (2010).
This work indicates that continued provincialization of the rudists of
the Americas occurred even during the Late Cretaceous. While cosmopolitan and
endemic forms were present (Fig. 18 ![]() ) in the late Turonian (Hippurites), early to middle Campanian (Vaccinites and Torreites),
by the Maastrichtian the hippuritids of the American region were represented by
endemic radiations (Barrettia and Caribbea
and their allies), and cosmopolitan forms had disappeared.
) in the late Turonian (Hippurites), early to middle Campanian (Vaccinites and Torreites),
by the Maastrichtian the hippuritids of the American region were represented by
endemic radiations (Barrettia and Caribbea
and their allies), and cosmopolitan forms had disappeared.
Jann Thompson and Brian Huber facilitated my trip to the Smithsonian Institution for two weeks to study and sort the rudist collections in 2009. Peter Skelton accompanied me on this trip and I am extremely grateful to him for taking photographs. I thank: the late Ann Molineux (former Non-Vertebrate Paleontology Collections Manager, Texas Natural Science Center, The University of Texas at Austin) who facilitated my trip to the Texas Memorial Museum in 2009, which was supported by the National Science Foundation under Grant No. 0646468 awarded to the Texas Natural Science Center at the University of Texas; Dr Jon Todd facilitated me in The Natural History Museum, London; Dr Reinaldo Rojas-Consuegra kindly allowed access and permission to photograph the collections in the National Natural History Museum, Havana, Cuba; Dra Mar�a del Carmen Perrillat and Violeta A. Romero May�n are thanked for allowing access to the type collections of the Museum of Paleontology, Institute of Geology, Universidad Nacional Aut�noma de M�xico, Ciudad Universitaria, M�xico; Mr W.J. Wildenberg allowed free access to work through the extensive Caribbean rudist collections at Naturalis, Leiden, The Netherlands. Hern�n Santos facilitated me in the field in Puerto Rico and in the Geological Museum, University of Puerto Rico, Mayag�ez. Mr Ian Brown and Dr Sherene James-Williamson allowed access to the collections at the University of the West Indies Geological Museum, Kingston, Jamaica. I thank Peter Skelton for his thorough and careful review of the manuscript and for making suggestions that greatly improved it. The reviews of Robert Scott and Thomas Steuber also helped correct errors and improve the manuscript.
Astre G. (1931).- Sur une faune de Rudistes du Cr�tac� des plateaux sahariens au sud de l'Atlas marocain.- Bulletin de la Soci�t� d'Histoire naturelle de Toulouse, vol. 61, no. 2, p. 345-380.
Barrande J. (1881).- Syst�me Silurien du centre de la Boh�me. Classe des Mollusques. Ordre des Ac�phal�s.- chez l'auteur et �diteur, Prague, vol. VI, 342 p.
Chubb L.J. (1956).- Some rarer rudists from Jamaica, B.W.I.- Palaeontographica Americana, Ithaca - NY, vol. 4, p. 2-31.
Chubb L.J. (1971).- Rudists of Jamaica.- Palaeontographica Americana, Ithaca - NY, vol. 7, p. 157-257.
Dechaseaux C. (1952).- Classe des lamellibranches (Lamellibranchiata Blainville 1816). In: Piveteau J. (ed.), Trait�́ de pal�ontologie.- Masson, Paris, p. 220-364.
Dhondt A.V. & Jaillard �. (2005).- Cretaceous bivalves from Ecuador and northern Peru.- Journal of South American Earth Sciences, vol. 19, p. 325-342.
Dommelen H. van (1971).- Ontogenetic, phylogenetic and taxonomic studies of the American species of Pseudovaccinites and of Torreites and the multiple-fold hippuritids.- Thesis, University of Amsterdam, 125 p.
Douvill� H. (1891).- �tudes sur les rudistes. R�vision des principales esp�ces d'Hippurites.- M�moires de la Soci�t� g�ologique de France (Pal�ontologie), Paris, no. 6, vol. 1, p. 1-31 (3 Pls.).
H. (1910).- �tudes sur les rudistes : Rudistes de Sicile, d'Alg�rie, d'�gypte, du Liban et de la Perse.- M�moires de la Soci�t� g�ologique de France (Pal�ontologie), Paris, no. 41, t. XVIII, fasc. no. 1, 84 p. (7 Pls.).
Douvill� H. (1926).- Quelques fossiles du Cr�tac� sup�rieur de Cuba.- Compte rendu sommaire et Bulletin de la Soci�t� g�ologique de France (4e S�rie), Paris, vol. 26, p. 127-138.
Fischer P. (1887).- Manuel de conchyliologie et de pal�ontologie conchyliologique ou histoire naturelle des mollusques vivants et fossiles.- Paris, 1369 p. (23 Pls.).
Gerth H. (1928).- Neue Faunen der Oberen Kreide mit Hippuriten aus Nordperu.- Leidsche geologische Mededeelingen, Leiden, vol. 5, p. 231-241.
G�tz S. & Mitchell S.F. (2009).- The Laluzia armini (gen. et spec. nov.) ecosystem: understanding a deeper-water rudist lithosome from the Early Maastrichtian of Mexico.- Facies, Erlangen, vol. 55, p. 539-551.
Gray J.E. (1848).- On the arrangement of the Brachiopoda.- Annals and Magazine of Natural History (serie 2), London, vol. 2, p. 435-440.
Grubić A. (1979).- Torreites milovanovici sp. nov. from Oman, T. coxi sp. nov. and T. chubbi sp. nov. from Jamaica, a new description of the genus Torreites Palmer, and a reference to the significance of its palaeogeographic distribution.- Zavod za geoloska i geofizicka Istrazivanja, Vesnik Geologija, Beograd, vol. 37, p. 81-95 (2 Pls).
Grubić A. (2004).- Revision of the rudists subfamily Barrettiinae Chubb.- Bulletin T. CXXVIII de l'Acad�mie serbe des Sciences et des Arts, Classe des Sciences math�matiques et naturelles, Sciences naturelles, Beograd, no. 42, p. 139-197.
Gunter G.C. (2002).- Two very fossiliferous rudist sites from the Marchmont Inlier, northwestern Jamaica.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 36, p. 21-26.
Gunter G.C. & Mitchell S.F. (2005).- The lithostratigraphy of the Maldon Inlier, northwestern Jamaica. Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 38, p. 1-10.
Ifrim C., Stinnesbeck W. & Schafhauser A. (2005).- Maastrichtian shallow-water ammonites of northeastern Mexico.- Revista Mexicana de Ciencias Geol�gicas, Mexico City, vol. 22, p. 48-64.
Kauffman E. & Sohl N.F. (1974).- Structure and evolution of Antillean Cretaceous rudist frameworks.- Verhandlungen der naturforschenden Gesellschaft in Basel, vol. 84, p. 399-467.
Lamarck J.B. (1801).- Syst�me des animaux sans vert�bres ou tableau g�n�ral des classes, des ordres et des genres de ces animaux.- Paris, 432 p.
Lupu D. (1975).- Faune s�nonienne � rudistes de la province de Pinar del Rio (Cuba).- Dari de Seama ale Sedintelor (3 Paleontologie), vol. 61, p. 223-254.
Mac Gillavry H.J. (1937).- Geology of the province of Camag�ey, Cuba, with revisional studies in rudist paleontology.- Geographische en geologische Mededeelingen, Utrecht, vol. 14, 168 p.
Matthews S.C. (1973).- Notes on open nomenclature and on synonymy lists.- Palaeontology, vol. 16, p. 713-719.
Meneghini G. (1868).- Il nuovo genere Pironaea.- Atti della Societ� Italiana di Science Naturali, vol. 11, p. 402.
Milovanović B. (1938).- Anatomie compar�e et relations phylog�n�tiques des Lapeirouseinae.- Bulletin de l'Acad�mie serbe des Sciences, Classe des Sciences math�matiques et naturelles, Sciences naturelles, Belgrade, vol. 4, p. 129.
Mitchell S.F. (1999).- Stratigraphy of the Guinea Corn Formation (Upper Cretaceous) at its type locality in the Rio Minho between Grantham and Guinea Corn, northern Clarendon, Jamaica.- Journal of the Geological Society of Jamaica, Kingston, Jamaica, vol. 33, p. 1-12.
Mitchell S.F. (2010).- Revision of three large species of Barrettia from Jamaica.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 41, p. 1-16.
Mitchell S.F. (2013).- Revision of the Antillocaprinidae Mac Gillavry (Hippuritida, Bivalvia) and their position within the Caprinoidea d'Orbigny.- Geobios, Villeurbanne, vol. 46, p. 423-446.
Mitchell S.F. (2020).- Cretaceous geology and tectonic assembly of Jamaica. In: Davison I., Hull J.N.F. & Pindell J. (eds.), The basins, orogens and evolution of the southern Gulf of Mexico and northern Caribbean.- Geological Society of London, SP504, 41 p., https://doi.org/10.1144/SP504-2019-210
Mitchell S.F. & Skelton P.W. (2013).- A new rudist bivalve, Polytorreites gen. nov., from the Campanian of Puerto Rico demonstrating iterative evolution of American multiple-ray Hippuritidae.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 45, p. 119-123.
Mitchell S.F., Mart�nez-Col�n M., Ramsook R. & Santos H. (2011).- A primitive tube-bearing antillocaprinid rudist bivalve, Parasarcolites sohli sp. nov, from Jamaica and Puerto Rico, West Indies.- Cretaceous Research, vol. 34, p. 149-153.
Moore R.C. (ed., 1969).- Treatise on Invertebrate Paleontology, Part N, vol. 2 Mollusca, part 6 Bivalvia.- Geological Society of America and University of Kansas, Lawrence - KS, 1224 p.
M�llerried F.K.G. (1930).- Un Hippurites de la regi�n de Cardenas, S.L.P.- Anales del Instituto de Biolog�a, Mexico City, vol. 1, p. 165-168.
Munujos H., Pons J.M. & Vicens E. (2016).- The rudist bivalve Pironaea milovanovici K�hn, 1935, a multiple-fold Hippuritidae, from south-eastern Spain. Taxonomic implications. Pore and canal system constructional morphology.- Cretaceous Research, vol. 63, p. 122-141.
Munujos H., Pons J.M. & Vicens E. (2020).- The Pironaea species (Bivalvia: Hippuritidae), other than P. milovanovici K�hn, from SE Spain, E Serbia and E and SE Anatolia, and a revision of the genus Pironaea from the Upper Cretaceous of Eurasia.- Cretaceous Research, vol. 116, article 104554, 25 p.
Myers R.L. (1968).- Biostratigraphy of the Cardena Formation (Upper Cretaceous) San Luis Potos�, M�xico.- Paleontolog�a mexicana, Mexico City, vol. 24, 89 p. (16 Pls.).
Newell N.D. (1965).- Classification of the Bivalvia.- American Museum Novitates, New York, no. 2206, p. 1-25.
Olsson A.A. (1934).- Contributions to the paleontology of northern Peru: The Cretaceous of the Amotape region.- Bulletins of American Paleontology, Ithaca - NY, vol. 20, p. 1-105 (2 Pls.).
Orbigny A. d' (1847).- Sur les Brachiopodes ou Palliobranches (deuxi�me m�moire).- Comptes rendus hebdomadaires des S�ances de l'Acad�mie des Sciences, Paris, vol. 25, p. 266-269.
Palmer R.H. (1933).- Nuevos rudistas de Cuba.- Revista de Agricultura, Comercio y Trabajo, vol. 14, p. 95-125.
Parona C.F. (1900).- Sopra alcune rudiste senoniane dell'Appennino meridionale.- Memorie dell'Accademia delle Scienze di Torino, Classe di Scienze fisiche, matematiche e naturali, vol. 50, p. 1-23 (2 Pls.).
Philip J. & Jaillard �. (2004).- Revision of the Upper Cretaceous rudists from northwestern Peru.- Journal of South American Earth Sciences, vol. 17, p. 39-48.
Philip J. & Platel J.-P. (1994).- Praetorreites, nouveau genre de rudiste du Campanien d'Oman.- Geobios, Villeurbanne, vol. 27, no. 3, p. 303-319.
Pons J.M., Vicens E. & Garc�a-Barrera P. (2019).- Campanian and Maastrichtian hippuritid rudists (Hippuritida, Bivalvia) of the Chiapas Central Depression (southern Mexico) and implications for American multiple-fold hippuritid taxonomy.- Journal of Paleontology, vol. 93, p. 291-311.
Pons J.M., Vicens E., Oviedo A., Aguilar J., Garc�a-Barrera P. & Alenc�ster G. (2013).- The Rudist fauna of the C�rdenas Formation, Maastrichtian, San Luis Potos�.- Journal of Paleontology, vol. 87, p. 726-754.
Pons J.M., Vicens E., Pichardo Y., Aguilar J., Oviedo A., Alenc�ster G. & Garc�a-Barrera P. (2010).- A new Early Campanian Rudist fauna from San Luis Potosi in Mexico and its taxonomic and stratigraphic significance.- Journal of Paleontology, vol. 84, p. 974-995.
Schafhauser A. (2006).- The C�rdenas Formation in East-Central Mexico (Maastrichtian): Stratigraphy, depositional environment and rudist decline.- PhD thesis, Karlsruhe University, 106 p. URL: http://digbib.ubka.uni-karlsruhe.de/volltexte/1000004246
Schafhauser A., G�tz S & Stinnesbeck W. (2007).- Rudist decline in the Maastrichtian Cardenas Formation (East-central Mexico).- Pal�ogeography, Pal�oclimatology, Pal�oecology, vol. 251, p. 210-221.
Schumann D. (2010).- The morphology and function of the upper valve of Vaccinites vesiculosus (Woodward).- Turkish Journal of Earth Sciences, Ankara, vol. 19, p. 791-798.
S�nesse P. (1946).- Sur la forme ancestrale des Hippurites et les Rudistes qui l'accompagnent dans les Corbi�res.- Bulletin de la Soci�t� Linn�enne de Lyon, vol. 4, p. 85-95.
Skelton P.W. (2013).- Rudist classification for the revised Bivalvia volumes of the Treatise on Invertebrate Paleontology.- Caribbean Journal of Earth Science, Kingston, Jamaica, vol. 45, p. 9-33.
Skelton P.W. (2018).- Part N, Volume 1, Chapter 26A: Introduction to the Hippuritida (rudists): Shell structure, anatomy, and evolution.- Treatise Online, Lawrence - KS, vol. 104, p. 1-37.
Sohl N.F. (1992).- Upper Cretaceous gastropods (Fissurellidae, Haliotidae, Scissurellidae) from Puerto Rico and Jamaica. Journal of Paleontology, vol. 66, p. 414-434.
Sohl N.F. (1998).- Upper Cretaceous trochacean gastropods from Puerto Rico and Jamaica.- Palaeontographica Americana, Ithaca - NY, vol. 60, p. 1-109.
Sohl N.F. & Kollmann H.A. (1985).- Cretaceous actaeonellid gastropods from the Western Hemisphere.- U.S. Geological Survey Professional Paper, no. 1304, 104 p. (23 Pls.).
Steuber T., Mitchell S. F., Buhl D., Gunter G. & Kasper H.U. (2002).- Catastrophic extinction of Caribbean rudist bivalves at the Cretaceous-Tertiary boundary. Geology, vol. 30, p. 999-1002.
Steuber T., Scott R.W., Mitchell S.F. & Skelton P.W. (2016).- Part N, Revised, Volume 1, Chapter 26C: Stratigraphy and diversity dynamics of Jurassic�Cretaceous Hippuritida (rudist bivalves). Treatise Online, vol. 81, p. 1�17.
Trechmann C.T. (1924).- The Cretaceous limestones of Jamaica and their Mollusca.- Geological Magazine, London, vol. 61, p. 385-410.
Vermunt L.W.J. (1937).- Cretaceous rudists of Pinar del Rio province, Cuba.- Journal of Paleontology, Tulsa - OK, vol. 11, p. 261-275.
Whitfield R.P. (1897).- Observations on the genus Barrettia Woodward, with descriptions of two new species.- Bulletin of the American Museum of Natural History, New York, vol. 9, p. 233-246.
Woodward S.P. (1862).- Some account of Barrettia, a new and remarkable fossil shell from the hippurite limestone of Jamaica.- The Geologist: a Popular Illustrated Monthly Magazine of Geology, London, vol. V (October 1862), p. 372-377 (Pls. XX-XXI).
APPENDIX. MORPHOLOGICAL TERMS APPLICABLE TO HIPPURITID RUDISTS
Anterior Myophore (AM). Arc shaped muscle platform developed from the Myocardinal Yoke in the LV. The length, shape and orientation of the AM differ in different hippuritids.
Anterior Myophore Attachment (AMA). The Anterior Myophore may attach directly onto the wall of the body cavity or it may attach onto an inclined ledge or surface formed in the Inner Layer shell of the RV.
Anterior Tooth (AT). Developed on the myocardinal arc of the LV.
Apertural Face. The internal surface of the RV that may be gently concave, flat of gently convex. It may be unspecialized in early forms but becomes highly complex in advanced forms.
Apertural Radial Ribs. Positive features on the apertural surface of Parastroma which resemble the rays in ray-bearing complex multiple-fold hippuritids but lack any form of rays. They are formed by the variable height of the concentric and/or radial partitions.
Apertural Radial Furrows. Negative features on the apertural surface of Parastroma which resemble the inter ray spaces in ray-bearing complex multiple-fold hippuritids but lack any form of rays. They are formed by the variable height of the concentric and/or radial partitions.
Base (Dommelen, 1971, p. 45). Unpinched attachment of Ray to outer shell layer.
Beads (Woodward, 1862, p. 374). Wide parts of rays between constrictions; they may be regular or irregular.
Body Cavity (BC). Cavity formed in the RV in which the animal was situated. The corresponding cavity in the LV is much reduced or absent. Same as Visceral Cavity (CV).
Body Chamber Layer (BCL). A continuous layer lining the body cavity of the RV. This is directly equivalent to the inner shell layer in more primitive hippuritids but in more advanced forms the inner and outer shell layers are separated by the infolds (rays) and partitions formed from extensions of the inner shell layer. The Body Chamber Layer may rise to form a prominent wall on the apertural face.
Buttress. Support between the LV and the myocardinal yoke.
Central Tooth (CT). The reduced Central Tooth of the RV is represented by a support for receiving the two teeth of the LV; there is no socket in the LV other than for the gap between the two teeth.
Complex Inner Shell Layer. Situation where the inner shell layer of the RV is composed of a Body Chamber Layer separated by the Outer Shell Layer by a series of cavities formed by interactions between rays (when present), various radial and/or concentric partitions and horizontal tabulae.
Concentric Elongate Spaces. Regular or irregular concentrically elongated spaces developed between radial and concentric partitions in Parastroma.
Concentric Partitions. Concentric wall-like elements representing part of the Complex Inner Shell Layer of hippuritids between the Body Cavity Lining and the Outer Shell Layer.
Connellae. Small conical structures developed on tabulae or on the myocardinal areas of some complex hippuritids such as Parastroma. Include two types: Myocardinal Connellae and Tabular Connellae.
Costae. Positive features on the outer surface of the RV.
Cross-Yoke Tunnels. Tunnels that penetrate the myocardinal yoke. They may be small or large.
Cycles of Infolds (Grubić, 2004). Scheme where rays are inserted successively and considered by Grubić (2004) to have taxonomic value at the genus level. Each lineage, however, tends to show an increase in number of rays and cycles of rays during both ontogeny and lineage evolution. The concept is therefore of little value in determining generic affinities.
Degree of Pinching (Dommelen, 1971, p. 45). Ratio between minimum diameter (pinched area between beads) and maximum diameter (i.e., a bead) of a ray.
Dendritic Radial Muri. Irregular branching radial muri developed on surfaces of funnel plates.
Denticulae (Dommelen, 1971, Pl. 3). Minute teeth developed on margins of the pore walls, probably the broken terminations of trabeculae.
Double Vertical Canal. A vertical canal with a figure-of-eight shape that penetrates vertically through the LV of multiple fold hippuritid rudists.
Funnel Plates. Plates developed in the non-compact Outer Shell Layer that are separated by radial muri or pustules in the RV of some hippuritids. The formation is analogous to the shell layer developed in radiolitid rudists.
Infoldings. See Rays.
Interradial Plate (Dommelen, 1971, p. 94). Plate developed in LV of Parastroma between adjacent Radial Spaces and bearing pallial canals. Same as buttresses and radial walls.
Inter-Radial Spaces (Whitfield, 1897, p. 235). Inter-ray spaces or Quadrangular Pits.
Inter-Ray Partitions. Partitions between infolded rays.
Inter-ray pits. Pits developed in the inter-ray spaces by inter-ray partitions in some multiple-folded hippuritids.
Inter-Ray Spaces. The area between rays which is characterized by a depression on the surface of the RV. May be broken up by inter-ray partitions to form inter-ray quadrangular pits.
Knob (Dommelen, 1971, p. 45). Terminal bead in a ray.
Lacuna (Whitfield, 1897, p. 234). The "first bead of the ray" or knob, obsolete.
Left Valve (LV). The operculate or Free Valve that carries the pore system in hippuritid rudists.
Limbal Zone. Marginal zone of the LV. In adult specimens of some species it may have different kinds of pore developed compared to the rest of the LV.
Marginal Canals. Canals on the margin of the LV that open directly onto the apertural surface and not into the Radial Furrows.
Moniliform Rays (Woodward, 1862, p. 373). Rays with more than one constriction and with regular sized and spaced beads.
Multiple-Fold (Dommelen, 1971, p. 37). Term for Barrettia and alike forms.
Multiple-Ray (Dommelen, 1971, p. 37). Term for Barrettia and alike forms.
Muri. Radial features developed between the funnel plates in non-compact (cellular) outer shell layers in certain hippuritid rudists.
Myocardinal Connellae. Conical to irregular features developed on the surfaces of the sockets for the PM and AM in some complex hippuritids such as Parastroma.
Myocardinal Yoke. Thickened arc, circular or elliptical structure that supports the myocardinal (teeth and myophore) elements in the LV.
Non-Moniliform Rays (Dommelen, 1971, p. 45). Rays with more than one constriction and with only occasional beads.
Orimental Folds. Incipient folds that do not involve a tight invagination of the cortical layer of the shell.
Oscules. Holes developed in the LV above the pillars.
Outer Shell Layer. Calcitic shell layer of hippuritids.
Pallial Canals. Canals penetrating the inner shell layer. Seen in the LV of certain hippuritids in the New World. Contain tabulae or are blind suggesting that they are related to low density construction rather than having direct physical applications (cf. Cross-Yoke Tunnels).
Partitions. Wall-like elements representing part of the Complex Inner Shell Layer of hippuritids between the Body Cavity Lining and the Outer Shell Layer.
Pillars. The two or three principal rays or infolds (P0, P1 and P2).
Polygonal Field. Polygonal area with 4, 5 or 6 sides on the surface of the LV corresponding to a single or pair of vertical canals in multiple fold hippuritids. The Polygonal Field is defined by raised ridges that support the walls of the sieve sheet.
Pore System. The arrangement of pores on the LV. The pore system is regarded as having high taxonomic value at the genus level and has been used to define various genera.
Pores. Openings connecting the outside world with the radial furrows or lateral/vertical canals. Pores include simple, linear, polygonal, and dendritic.
Porules (Dommelen, 1971, p. 38). Small divisions in pores formed by tabulae.
Posterior Myophore (PM). Elliptical or forked muscle platform developed from the Myocardinal yoke in the LV. The position of the PM is different in different hippuritids and has value in distinction at the genus level.
Posterior Myophore Socket (PMS). Cavity for the PM that may be connected to the body cavity or separated from the body cavity by a lamina.
Posterior Tooth (PT). Tooth built on the myocardinal yoke of the LV.
Primary Infoldings. The two or three pillars.
Pustules. Bleb-like or irregular features developed between the funnel plates in non-compact (cellular) outer shell layers in certain hippuritid rudists.
Quadrangular Pits or Depressions (Whitfield, 1897, p. 235). Inter-ray pits.
Radial Canals (Dommelen, 1971, p. 70). Radially directed canals in the LV that exit at the RV knob of multiple-folded hippuritids or at the shell margin in simple hippuritids. They terminate at the shells outer margin and are overlain by the pore system. Some radial canals originated at the apex of the RV, others are intercalated as the valve increases in size. Where poorly preserved, the pore system is not preserved and only the radial furrows are visible.
Radial Grooves. Grooves on the abapical side of the LV that correspond to the rays in multiple fold hippuritids. Same as radial spaces.
Radial Partitions. Radially orientated wall-like elements representing part of the Complex Inner Shell Layer of hippuritids between the Body Cavity Lining and the Outer Shell Layer.
Radial Space (Dommelen, 1971, p. 95). Space above ray or apertural ridge in hippuritid rudists.
Radial Walls/Ribs (Dommelen, 1971, Pl. 11). Walls developed in LV of Parastroma between adjacent Radial Spaces and bearing pallial canals. Same as Buttresses.
Rays (Woodward, 1862, p. 372). Infoldings of the outer shell layer of the RV; the term used by Woodward (1862) to describe Barrettia. Two infoldings (P1 and P2) are present in all hippuritids, in many a third (P0 of Ligamental) is developed, whereas in multiple-folded hippuritids several to hundreds of infolds may be present.
Reticulum (Dommelen, 1971, p. 70). Pore covering in hippuritid rudists.
Riblets. Thin positive features on the exterior surface of the RV of hippuritid rudists.
Right Valve (RV). The lower or fixed valve.
Secondary Infoldings. Rays other than the primary three infoldings, in multiple-ray hippuritids.
Sieve Pustule. Pustule shaped covering separated from neighbouring pustules by a wide imperforate zone over a single or double vertical canal that bears various pores in multiple hippuritids such as Whitfieldiella.
Sieve Sheet/Plate. A sheet bearing pores overlaying a Polygonal Field in multiple fold hippuritids.
Simple Rays. Triangular to parallel-sided to concave sided infolds.
Stem (Dommelen, 1971, p. 45). Interval between the knob and the base of a ray in non-simple rays.
Submoniliform Rays (Dommelen, 1971, p. 45). Rays with more than one constriction (i.e., with a stem) and with only irregularly formed beads.
Sulcus. Furrow or groove on the exterior surface of the RV.
Tabulae Connellae. Irregularly distributed or aligned cones on the tabulae some complex hippuritids such as Parastroma.
Tabulae. Broadly horizontal layers in the Complex Inner Shell Layer or the Body Cavity.
Trabeculae (Dommelen, 1971, p. 38). Division of pores into porules.
Transverse Furrows (Dommelen, 1971, p. 70). Concentrically orientated furrows connecting the vertical canals to the sieve plates in Praebarrettia.
Transverse Walls (Whitfield, 1897, p. 235). Inter-ray partitions.
Tunnel Canals. Canals that pass through the Myocardinal Yoke.
Vertical Canals. Canals that penetrate vertically through the LV of multiple fold hippuritid rudists. They may be single or double.
Vesicular. Cell-like fill of Body Cavity by anastomosing tabulae.
Visceral Cavity (CV). Cavity formed in the RV in which the animal was situated. The corresponding cavity in the LV is much reduced or absent. Same as Body Cavity (BC).