◄ Carnets Geol. 20 (17) ►
![]()
Outline:
[1. Introduction]
[2. Geological setting]
[3. Material and methods]
[4. Systematic palaeontology]
[5. Discussion and conclusion]
[Bibliographic references] and ...
[Plate]
Office National des Mines, 24, rue de l'énergie, 2035 La Charguia, Tunis (Tunisia)
(corresponding author) University of Catania. Department of Biological, Geological and Environmental Science, Corso Italia 55, 95129 Catania (Italy)
Office National des Mines, 24, rue de l'énergie, 2035 La Charguia, Tunis (Tunisia)
Published online in final form (pdf) on October 14, 2020
DOI 10.2110/carnets.2020.2017
![]()
[Editor: Bruno Granier;
language editor: Stephen Eagar]
![]()
Micropalaeontological analyses were performed on two stratigraphical sections sampled in upper Messinian deposits outcropping in eastern Tunisia, allowing us to identify some sedimentary levels with high concentrations of fresh or brackish water ostracods, which can be referred to the Lago-Mare fauna. Some of these species can be considered Paratethysian, or rather as species that spread in the Mediterranean area starting from the Paratethys areas, while others show Paratethysian affinity. The Lago-Mare fauna is little known in the south Mediterranean regions and the present article provides new data on its geographic distribution.
Of the two sections sampled, the first one, the Wadi El Kebir section, is located in the south eastern part of the Cape Bon Peninsula and shows horizons dominated by Cyprideis agrigentina and Cyprideis ex C. torosa group; the second one, the Salakta section, is located in the Sahel region and shows a level with a very rich Lago-Mare ostracod fauna consisting essentially of Amnicythere propinqua, Mediocytherideis punctata, and Ilyocypris gibba.
• Ostracods;
• upper Messinian;
• Lago-Mare facies;
• Tunisia;
• palaeoenvironmental evolution
Temani R., Sciuto F. & Ammar H.K. (2020).- Messinian Lago-Mare ostracods from Tunisia.- Carnets Geol., Madrid, vol. 20, no. 17, p. 315-331.
Ostracodes du Lago-Mare messinien en Tunisie.- Des analyses micropaléontologiques ont été réalisées sur deux coupes stratigraphiques échantillonnées dans des dépôts du Messinien supérieur affleurant en Tunisie orientale. Elles nous ont permis d'identifier certains niveaux sédimentaires présentant de fortes concentrations en ostracodes d'eaux douce ou saumâtre, qui peuvent être rapportés à la faune de faciès Lago-Mare. Certaines de ces espèces peuvent être considérées comme paratéthysiennes ou plutôt comme des espèces ayant migré en Mer Méditerranéenne depuis les régions paratéthysiennes, alors que d'autres présentent une affinité paratéthysienne. La faune de faciès Lago-Mare est peu connue dans les régions sud-méditerranéennes et cet article fournit de nouvelles données sur sa répartition géographique.
Des deux sections étudiées, la première, la coupe de Wadi El Kebir, située dans la partie sud-est de la péninsule du Cap Bon, comporte des niveaux dominés par Cyprideis agrigentina et Cyprideis ex gr. C. torosa, tandis que la seconde, la coupe de Salakta, située dans la région du Sahel, comporte un niveau doté d'une très riche faune d'ostracodes de faciès Lago-Mare, essentiellement constituée des espèces Amnicythere propinqua, Mediocytherideis punctata et Ilyocypris gibba.
• Ostracodes ;
• Messinien supérieur ;
• faciès Lago-Mare ;
• Tunisie ;
• évolution paléoenvironmentale
At the end of the Miocene, the Mediterranean basin underwent one of the most disturbing environmental crisis that occurred in geological time. During this event, called Messinian Salinity Crisis (MSC) (Hsü et al., 1973; Clauzon et al., 1996; Gliozzi et al., 2007; Garcia-Castellanos et al., 2009; Grossi et al., 2011; Krijgsman et al., 2018; G. Mascle & J. Mascle, 2019; Ben Moshe et al., 2020, inter alia), started 5.97 Ma ago (Manzi et al., 2013), large thicknesses of evaporites were deposited on the bottom of Mediterranean Sea (Ruggieri, 1967; Benson, 1978; Cita et al., 1978; G. Mascle & J. Mascle, 2019, inter alia). The causes that led to the deposition of evaporitic salts can be correlated to the more or less total closure of the connection between the Atlantic and Mediterranean seas.
According to an old hypothesis, the Mediterranean Sea dried up completely, due to the closure of the Strait of Gibraltar, leading to the deposition of evaporitic salts in deep hypersaline basins (Hsü et al., 1973; Benson 1973a, 1973b; Cita et al., 1978, inter alia). More recently, researchers have argued that the Strait of Gibraltar did not close entirely and that the Mediterranean Sea did not completely dry up; on the contrary, they suggest the presence of shallow saline water sedimentation on the floor of deep depressions, with a recurrent feeding mechanism of these depressions, including alternation of dewatering and filling events, or a system of basins located at different altitudes but connected by waterfalls (Roveri et al., 2014; Krijgsman et al., 2018; G. Mascle & J. Mascle, 2019).
The temporary Mediterranean disconnection from the Atlantic Ocean would be the result of a tectonic uplift of the Gibraltar threshold that controls the inflow of water required to compensate for its hydrological deficit (Cita et al., 1978; Garcia-Castellanos et al., 2009, inter alia).
According to Ben Moshe et al., 2020, during the third stage of the MSC (Lago-Mare event, 5.55-5.33Ma) the Mediterranean Sea level fluctuated repeatedly. Several parts of the Mediterranean Basin emerged and were affected by intense continentalization, with strong subaerial erosion phenomena, chemical dissolution of the previous Messinian evaporites, and, in the widespread lower basin area, deposition of terrigenous sedimentary facies characterized by brackish to a freshwater fauna (Orszag-Sperber et al., 2000; Rouchy et al., 2007; Gliozzi et al., 2007). This sedimentary facies, called "Lago-Mare" (sensu Ruggieri, 1967), occurs discontinuously but widely in the Mediterranean Basin from the Eastern to the Western Basin (Orszag-Sperber et al., 2000; Rouchy & Caruso, 2006; Gliozzi et al., 2007; Grossi et al., 2011; Faranda et al., 2013, inter alia).
According to the studies undertaken in Sicily and in many geographical areas of the Central Mediterranean Basin, the sediments deposited during the Lago-Mare event are represented by two different facies: the first consists of marls containing oligohaline faunas ("Congeria marls") (Di Geronimo et al., 1989; Sciuto et al., 2018), the second consists of reddish arkosic sands ("Arenazzolo") containing brackish to freshwater ostracods (Bonaduce & Sgarella, 1999; Roveri et al., 2008; Sciuto et al., 2018). Part of this fauna points to original mesohaline to hyperhaline shallow-water paleoenvironments with low oxygen content (Grossi et al., 2015) seemingly produced by overflows from the Pannonian-Pontian Paratethysian waters (Gliozzi et al., 2007; Stoica et al., 2016).
Only a few detailed studies have been carried out on the Messinian ostracods of Tunisia, (Gulf of Gabes, South Tunisia; Bonaduce et al., 1992) and even fewer on the "Lago-Mare" fauna. General studies have been performed on the Messinian facies (El Euch-El Koundi et al., 2009; Abdi et al., 2014; Frigui et al., 2016). Therefore, the main purpose of the present contribution focuses on the ostracod fauna collected from two stratigraphic sequences out-cropping in some localities in Eastern Tunisia, is to verify the presence of both Lago-Mare faunas and Paratethysian species therein.
The acquisition of this data will allow us to understand and to interpret the paleoecologic and paleoclimatic evolution of this area during the post evaporitic phase of the late Messinian.
The Messinian facies are relatively rare in Tunisia. The evaporitic ones are only known offshore, the terrigenous ones are only known in the Cap Bon Peninsula in northern Tunisia (Burollet, 1951; Colleuil, 1976; Ben Salem, 1992; Frigui et al., 2016; Temani et al., 2018, 2019) and in the Sahel region (Besème & Kamoun, 1988; Kamoun et al., 2001; Moissette et al., 2010; Abdi et al., 2014; Frigui et al., 2016; Temani et al., 2018, 2019). They are usually squeezed between the upper Tortonian coastal to continental deposits (Saouaf and Somaa Formations) and the Lower Pliocene marine marls.
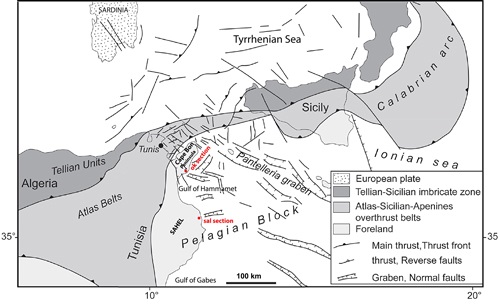
The Cape Bon Peninsula (Fig. 1
![]() )
is almost entirely
constituted by the Jebel Abderrahmane anticline. The oldest series exposed is
the Middle Eocene Souar Formation (Burollet, 1956; Abdi
et al., 2014). The overlying Neogene deposits consist of marine and terrestrial
siliciclastic facies and carbonates (Burollet, 1956; Colleuil,
1976; Demarcq et al., 1976; Ben
Salem, 1992; Bédir et al.,
1996; Mannaï-Tayech, 2006).
)
is almost entirely
constituted by the Jebel Abderrahmane anticline. The oldest series exposed is
the Middle Eocene Souar Formation (Burollet, 1956; Abdi
et al., 2014). The overlying Neogene deposits consist of marine and terrestrial
siliciclastic facies and carbonates (Burollet, 1956; Colleuil,
1976; Demarcq et al., 1976; Ben
Salem, 1992; Bédir et al.,
1996; Mannaï-Tayech, 2006).
The Sahel area in Eastern Tunisia (Fig. 1
![]() ) extends
from the "North-South Axis" eastward to the Mediterranean Sea. It is a flat
foreland to the east of the Alpine Domain of Tunisia (Fig. 1
) extends
from the "North-South Axis" eastward to the Mediterranean Sea. It is a flat
foreland to the east of the Alpine Domain of Tunisia (Fig. 1
![]() ). The outcropping
sedimentary series range from the Upper Miocene to the Quaternary (Burollet,
1956; Demarcq
et al., 1967; Kamoun, 1981;
Besème & Kamoun, 1988;
Bédir, 1995; Gaaloul,
1995; Ben Youssef et al.,
2002; Frigui, 2003; Abdi et al.,
2014).
). The outcropping
sedimentary series range from the Upper Miocene to the Quaternary (Burollet,
1956; Demarcq
et al., 1967; Kamoun, 1981;
Besème & Kamoun, 1988;
Bédir, 1995; Gaaloul,
1995; Ben Youssef et al.,
2002; Frigui, 2003; Abdi et al.,
2014).
|
Figure 1:
Structural scheme of the Central Mediterranean area with the
location of the OK Section and SAL Section (modified after many authors). |
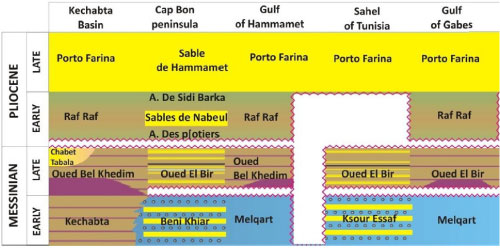
In these areas, the Messinian deposits (Fig. 2 ![]() )
are represented by two
lithostratigraphic units: the Beni Khiar Fm. and the Oued El Bir Fm. (Colleuil,
1976; Fournié, 1978; Bismuth,
1984; Bédir
et al., 1996; Moissette et
al., 2010; Frigui
et al., 2016). The Beni Khiar Fm., with its lateral offshore
equivalent, the Melqart Fm. (Bonaduce et
al., 1988), consists of oolitic and bioclastic limestones alternating with
sandy and marly layers. The Oued El Bir Fm., or Oued bel Khedim Fm. in offshore
wells, consists of sands, sandstones, and sometimes chalky clays. The Beni Khiar
Fm. was described by Colleuil (1976) who proposed a Tortonian age for its
lower part (Somaa and Beni Khiar formations and a Messinian age for its
uppermost part (Oued El Bir Fm.). Ben Salem (1998) and Hooyberghs
and Ben Salem (1999) distinguished continental siliciclastic
deposits (Somaa Fm.) followed by marine deposits represented by sandstones and
clays (Beni Khiar Fm.) referred to the lower Messinian (N17 biozone of Blow).
The oolitic limestones follow. The sedimentary sequence ends with lagoonal and
lacustrine siliciclastic deposits (Oued El Bir Fm.) attributed to the upper
Messinian (Fig. 2
)
are represented by two
lithostratigraphic units: the Beni Khiar Fm. and the Oued El Bir Fm. (Colleuil,
1976; Fournié, 1978; Bismuth,
1984; Bédir
et al., 1996; Moissette et
al., 2010; Frigui
et al., 2016). The Beni Khiar Fm., with its lateral offshore
equivalent, the Melqart Fm. (Bonaduce et
al., 1988), consists of oolitic and bioclastic limestones alternating with
sandy and marly layers. The Oued El Bir Fm., or Oued bel Khedim Fm. in offshore
wells, consists of sands, sandstones, and sometimes chalky clays. The Beni Khiar
Fm. was described by Colleuil (1976) who proposed a Tortonian age for its
lower part (Somaa and Beni Khiar formations and a Messinian age for its
uppermost part (Oued El Bir Fm.). Ben Salem (1998) and Hooyberghs
and Ben Salem (1999) distinguished continental siliciclastic
deposits (Somaa Fm.) followed by marine deposits represented by sandstones and
clays (Beni Khiar Fm.) referred to the lower Messinian (N17 biozone of Blow).
The oolitic limestones follow. The sedimentary sequence ends with lagoonal and
lacustrine siliciclastic deposits (Oued El Bir Fm.) attributed to the upper
Messinian (Fig. 2 ![]() )
(Moissette et
al., 2010; Abdi et al.,
2014; Frigui et
al., 2016).
)
(Moissette et
al., 2010; Abdi et al.,
2014; Frigui et
al., 2016).
|
Figure
2:
Stratigraphic schema of the investigated region (modified after
Frigui et al., 2016). |
Two stratigraphic sequences were sampled for ostracod faunas: the Wadi El Kebir section and the Salakta section.
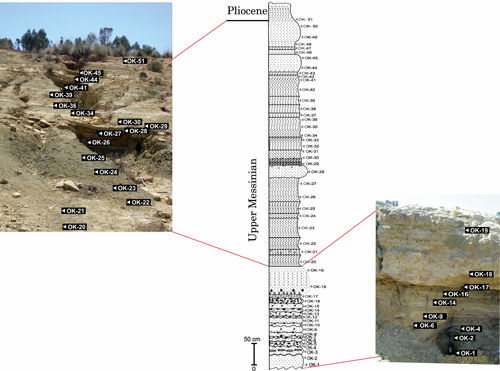
The Wadi El Kebir (OK) section (Fig. 3 ![]() )
crops out in the eastern side of
the Wadi El Kebir dam, in the Nabeul region, south eastern part of the Cape Bon
Peninsula. It consists of about 8.5 m thick siliciclastic deposits. It can be
divided into two lithostratigraphic units. The first one, 2.5m thick, is mostly
sandy and capped at the top by sandstone sediments (70 cm) that are very rich in
gastropods. This horizon shows bioturbation at the base, with bryozoa, algae,
and echinoderms debris at the top. The second unit begins with centimetric
sandy-clayey layers followed by laminated clays interbedded in sandstones. The
top part (80 cm thick) is mostly sandy and sometimes shows clay levels with
sandstones. These deposits correspond to the upper Messinian "Oued El Bir Fm." (Fig. 2
)
crops out in the eastern side of
the Wadi El Kebir dam, in the Nabeul region, south eastern part of the Cape Bon
Peninsula. It consists of about 8.5 m thick siliciclastic deposits. It can be
divided into two lithostratigraphic units. The first one, 2.5m thick, is mostly
sandy and capped at the top by sandstone sediments (70 cm) that are very rich in
gastropods. This horizon shows bioturbation at the base, with bryozoa, algae,
and echinoderms debris at the top. The second unit begins with centimetric
sandy-clayey layers followed by laminated clays interbedded in sandstones. The
top part (80 cm thick) is mostly sandy and sometimes shows clay levels with
sandstones. These deposits correspond to the upper Messinian "Oued El Bir Fm." (Fig. 2 ![]() )
(Said Benzarti et al., 2010; Moissette
et al., 2010; Frigui et
al., 2016; present paper). Fifty-one samples were taken from the Wadi El Kebir section.
)
(Said Benzarti et al., 2010; Moissette
et al., 2010; Frigui et
al., 2016; present paper). Fifty-one samples were taken from the Wadi El Kebir section.
The Salakta section (SAL)
(Fig. 5 ![]() )
is nearly 10 m thick and is located
about 3 km north of the village of Salakta. The lower levels consist of 2 m
thick silty marls followed
by 3.5 m of bioclastic sand interbedded with centrimetric marl levels. This sand
shows fragments of bivalves and bioturbations in some levels. Moving upwards,
one finds 1.5 m of yellow sandstone displaying abundant gasteropod molds,
followed by 2m of yellow fine sands containing bioclasts and Pectinids and
grading up to calcareous sandstones. The whole is covered by 1m of green marl
containing broken and complete oyster shells and 50 cm of fine sand. Thirty-one
samples were taken from the Salakta section. The deposits correspond to the
upper Messinian "Oued El Bir Fm."
)
is nearly 10 m thick and is located
about 3 km north of the village of Salakta. The lower levels consist of 2 m
thick silty marls followed
by 3.5 m of bioclastic sand interbedded with centrimetric marl levels. This sand
shows fragments of bivalves and bioturbations in some levels. Moving upwards,
one finds 1.5 m of yellow sandstone displaying abundant gasteropod molds,
followed by 2m of yellow fine sands containing bioclasts and Pectinids and
grading up to calcareous sandstones. The whole is covered by 1m of green marl
containing broken and complete oyster shells and 50 cm of fine sand. Thirty-one
samples were taken from the Salakta section. The deposits correspond to the
upper Messinian "Oued El Bir Fm."
For each sample, 250g of sediments were washed using diluted hydrogen peroxide for their disaggregation and sieved through standard sieves (63/125/250/500 µm). Residuals ≥250 mm were picked out completely and used for detailed taxonomic investigations. From the 125 µm sieve-residual 0.2 g/sample were picked and then quartered when necessary.
SEM micrographs were obtained through a LMU Tescan Vega II Scanning Electron Microscope at the Electronic Microscopy Laboratory of the University of Catania.
The specimens are deposited in the paleontological and sedimentological laboratory of the Geological Survey of the National Office of Mines of Tunisia.
Wadi El Kebir (OK) section (Figs. 3 ![]() - 4
- 4 ![]() ):
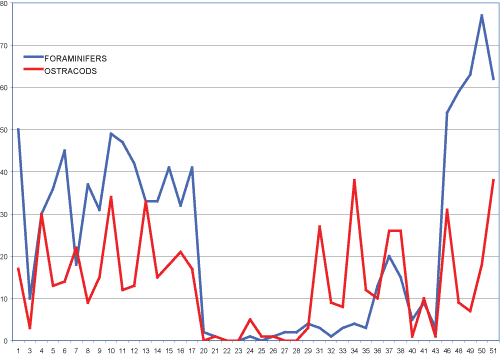
31 ostracod taxa were identified in the OK section. The ostracod fauna is
rare and not referable to a purely marine environment and dispersed along the
entire section and is only represented,
sometimes abundantly, by species of the genus Cyprideis
Jones, 1857.
):
31 ostracod taxa were identified in the OK section. The ostracod fauna is
rare and not referable to a purely marine environment and dispersed along the
entire section and is only represented,
sometimes abundantly, by species of the genus Cyprideis
Jones, 1857.
|
Figure
3:
The sedimentary succession cropping out at OK section and the
corresponding stratigraphical log with sampling location. |
|
Figure 4:
Distribution of ostracods and foraminifers along the OK section
(abscissa axis=samples; ordinate axis=number of specimens). |
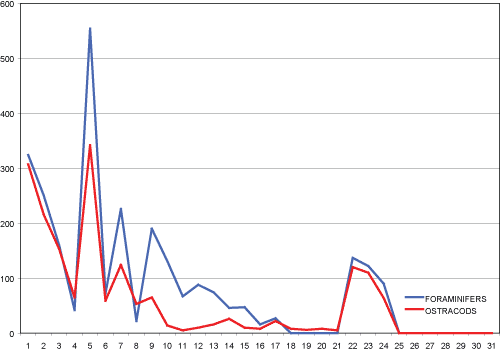
Salakta (SAL) section
(Figs. 5 ![]() - 6
- 6 ![]() ):
53 ostracod taxa were identified in the SAL section. The ostracod fauna
is not referable to a purely marine environment and is concentrated in a precise
stratigraphic interval between samples 9 and 17 (Fig. 4
):
53 ostracod taxa were identified in the SAL section. The ostracod fauna
is not referable to a purely marine environment and is concentrated in a precise
stratigraphic interval between samples 9 and 17 (Fig. 4 ![]() ). Ostracod association
in this level consist of such
species widespread and abundant
as Candona and Cyprideis associated
with Phlyctenophora farkasi (Zalanyi, 1913) and I. gibba, followed
by Amnicythere
propinqua (Livental, 1929), Mediocytherideis
punctata (Ligios
et al., 2008) and species belonging to the
genera Cytherois, Cypria, and
Ilyocypris. Foraminifers are rare. In
this group of samples were found charophyte gyrogonites,
the best preserved of which are referable to Chara hispida Linnaeus,
1753, and C.
vulgaris Linnaeus, 1753.
). Ostracod association
in this level consist of such
species widespread and abundant
as Candona and Cyprideis associated
with Phlyctenophora farkasi (Zalanyi, 1913) and I. gibba, followed
by Amnicythere
propinqua (Livental, 1929), Mediocytherideis
punctata (Ligios
et al., 2008) and species belonging to the
genera Cytherois, Cypria, and
Ilyocypris. Foraminifers are rare. In
this group of samples were found charophyte gyrogonites,
the best preserved of which are referable to Chara hispida Linnaeus,
1753, and C.
vulgaris Linnaeus, 1753.
|
Figure
5:
The sedimentary succession cropping out at SAL Section and the
corresponding stratigraphical log with sampling location. |
|
Figure 6:
Distribution of foraminifers and ostracods along the SAL Section
(abscissa axis=samples; ordinate axis=number of specimens). |
The distribution of Miocyprideis polita Bonaduce et al., 1992, is particularly meaningful; it is distributed, sometimes abundantly, in the lower and upper portion of the section, but it is practically absent between samples 9 and 21.
Ostracod species are particularly important for the late Messinian paleogeography and paleo-environment. They are well represented in some stratigraphic levels of the two sections and are listed systematically (according to Meisch et al., 2019).
They were found for the first time in this studied sector of the Mediterranean area.
Class OSTRACODA Latreille, 1806
Subclass PODOCOPA Sars, 1866
Order PODOCOPIDA Sars, 1866
Family CANDONIDAE Kaufmann, 1900
Genus Phlyctenophora Brady, 1880
Type species: Phlyctenophora zealandica Brady, 1880.
Phlyctenophora farkasi (Zalanyi, 1913)
(Pl. 1
![]() ,
fig. 1)
,
fig. 1)
2006 Phlyctenophora farkasi (Zalanyi), Olteanu, p. 20.
2007 Phlyctenophora farkasi, Pezely & Sremac, p. 83.
2008 Phlyctenophora farkasi (Zalanyi, 1913), Faranda et al., p. 301, Pl. 5, fig. 4
2013 Phlyctenophora farkasi (Zalanyi, 1913), Faranda et al., p. 846, Figs. 6 (u), 7 (e, f, h).
P. farkasi is reported always from shallow marine water and transitional environments, such as marshes, lagoons, and estuaries, often associated with Neomonoceratina laskarevi (Krstic & Pietrzeniuk, 1972). The species has been found in the Upper Miocene of central Crete (Faranda et al., 2008), the Badenian (Langhian-lower Serravallian) of the North Croatian Basin (Bakrač et al., 2010), Transylvania (Olteanu, 2006), and Medvednica Mt. (Croatia) (Pezelj & Sremac, 2007). In the Messinian the species is reported from the Adana Basin in southern Turkey (Faranda et al., 2013). The species does not appear to be reported in Italy. The genus is reported from the Upper-Middle Miocene of Buonfornello (Aruta, 1982).
P. farkasi is stratigraphically referred to the higher part of the upper Badenian in the Carpathian area (Olteanu, 2006) and in the Pokupsko area (Banovina, Croatia) where it is recognized the NO10 Zone Carinocythereis carinata - Phlyctenophora farkasi of the upper Badenian (Hajek-Tadesse & Prtoljan, 2011). According to Olteanu (2006), the species would be part of an association that would represent the faunal transition between the Paratethysian region fauna and those of the Mediterranean area.
Family ILYOCYPRIDIDAE Kaufmann, 1900
Subfamily Ilyocypridinae Kaufmann, 1900
Genus Ilyocypris Brady & Norman, 1889
Type species: Cypris gibba Ramdohr, 1808
Ilyocypris gibba (Ramdohr, 1808)
(Pl. 1
![]() ,
fig. 2)
,
fig. 2)
1808 Cypris gibba Ramdohr, p. 91, Pl. 3, figs. 13-14, 17;
1965 Ilyocypris gibba (Ramdohr); Devoto, p. 345, Fig. 50.
1979 Ilyocypris gibba (Ramdohr); Carbonnel & Peypouquet, p. 195, Pl. 1, fig. 2.
1998 Ilyocypris gibba (Ramdohr); Gliozzi & Mazzini, p. 80, Pl. 2, fig. A.
1999 Ilyocypris gibba (Ramdohr); Mazzini et al., p. 297, Pl. 2, fig. 5.
2000 Ilyocypris gibba (Ramdohr); Meisch, p. 245, Fig. 104.
2005 Ilyocypris gibba (Ramdohr); Rodriguez-Làzaro & Martin-Rubio, p. 40, Pl. 1, figs. 1-3, 7.
2006 Ilyocypris gibba (Ramdohr); Rossetti et al., p. 124, Fig. 2 (I-K).
2006 Ilyocypris gibba (Ramdohr); Pieri et al., p. 5.
2008 Ilyocypris gibba (Ramdohr); Akdemir, p. 109, Fig. 3.
2008 Ilyocypris gibba (Ramdohr); Beker et al., p. 12, Pl. 1, figs. 10-11.
2014 Ilyocypris gibba (Ramdohr); Uçak et al., p. 4.
2015 Ilyocypris gibba (Ramdohr); Sciuto et al., p. 50, Pl. 1, U.
Remarks: I. gibba (Ramdohr) is a Holarctic species, known from a very wide area in Europe and Asia, as well as from East Africa and North America (Henderson, 1990). In Sicily it has been recorded in Recent deposits by Pieri et al. (2006) and in Pleistocene sediments by Sciuto et al. (2015). The stratigraphical distribution of I. gibba is wide, ranging from the Tortonian to the Recent (Beker et al., 2008). The species is widespread in all freshwater environments, in a wide temperature range.
Family CYTHERIDEIDAE Sars, 1925
Genus Cyprideis Jones, 1857
Type species: Candona torosa Jones, 1850, by subsequent designation of Jones (1857).
Cyprideis agrigentina Decima, 1964
(Pl. 1
![]() ,
fig. 3)
,
fig. 3)
1964 Cyprideis pannonica agrigentina; Decima, p. 108-111, Pl. 29, figs. 4-8; Pl. 30, figs. 1-10; Pl. 31, fig. 1.2; Pl. 37, figs. 16-21.
1964 Cyprideis pannonica pseudoagrigentina; Decima, p. 111-113, Pl. 31, figs. 3-7; Pl. 32, figs. 1-2; Pl. 38, figs. 1-2.
1978 Cyprideis pannonica (Méhes); Benson, p. 780, Pl. 2, figs. 4-8.
1999 Cyprideis "agrigentina" Decima; Bonaduce & Sgarrella, p. 84-86, Pl. 1, fig. 1.
2007 Cyprideis agrigentina Decima; Rouchy et al., p. 392-393, 400, 407, 410-411, Pl. 4, figs. 1-2.
2008 Cyprideis agrigentina Decima; Gross et al., p. 133, 135, 137-140.
2008 Cyprideis agrigentina Decima; Trenkwalder et al., p. 94.
2018 Cyprideis agrigentina Decima; Sciuto et al., p. 11, Fig. 6.9
Remarks: According to Gliozzi (1999) the specific attribution of Cyprideis is rather complex. Indeed, the morphological characters of the carapace can be too diverse to justify the adoption of the term C. ex gr. pannonica by Gross et al. (2008) to indicate rather small and nearly smooth specimens of the genus found in upper Sarmatian and lower Pannonian sediments of the central Paratethys. This character is so distinct that Ligios and Gliozzi (2012) continue to propose to create the "C. torosa group", including species such as C. agrigentina Decima, 1964, C. ruggierii Decima, 1964, C. torosa (Jones, 1850), and in part C. crotonensis Decima, 1964, and C. calabra Decima, 1964 (Ligios & Gliozzi, 2012).
In this paper, we have decided to continue to use the specific name "agrigentina". We have identified the species through the observation of the internal characters of the carapace and particularly muscle scars, hinge, and duplicature. All the others specimens of "Cyprideis" have been grouped into the "C. torosa group" according to Ligios and Gliozzi (2012).
C. agrigentina is widespread in all the brackish Mediterranean domain during the "Lago-Mare" phase of the Messinian Salinity Crisis from the end of the evaporitic phase (about 5.5 Ma) to the Messinian/Zanclean boundary (5.33 Ma) (Cosentino et al., 2007; Gross et al., 2008; Guerra-Merchán et al., 2010; Cipollari et al., 2013). It seems to occur more frequently in the mesohaline high mesohaline facies, where it creates oligotypic assemblages together with Ammonia tepida (Bonaduce & Sgarrella, 1999; Grossi & Gennari, 2008; Guerra-Merchán et al., 2010), while in the oligo-mesohaline environment it seems to be vicariant with Cyprideis anlavauxensis (Grossi & Gennari, 2008).
C. agrigentina is reported from the Messinian of Eraclea Minoa (Decima, 1964), the Pannonian Stage of the northern Vienna Basin (Kováč et al., 1998), the Upper Miocene of eastern Anatolia (Nazik et al., 2008), the Pliocene of Almeria (Addicott et al., 1978), the Sarmatian of Turkey, the lower Sarmatian of Romania (Radu & Stoica, 2005), the Upper Miocene deposits of Anatolia (Şafak et al., 1999) and the lower Pannonian of Hungary (Kollmann, 1960). The majority of Cyprideis species live in brackish (meso-brachyhaline), euryhaline, mainly mesohaline (5-18) environments but also oligohaline and hyperhaline (Gross, 2004, inter alia). Gliozzi et al. (2007) consider C. agrigentina as a Lago-Mare species with Paratethysian affinity. Unlike C. torosa, C. agrigentina cannot be considered as a paleosalinometer for the MSC (Grossi et al., 2015).
Cyprideis ex C. torosa (Jones, 1850) group Ligios & Gliozzi (2012)
(Pl. 1
![]() ,
fig. 4)
,
fig. 4)
1964 Cyprideis torosa (Jones); Decima, Pl. 11, figs. 3-8c; Pl. 12, figs. 1-8d; Pl. 15, figs. 11-15.
2002 Cyprideis torosa (Jones, 1850); Wouters, Pl. 3, figs. 1a-4d.
2005 Cyprideis torosa (Jones, 1850); Matzke-Karasz & Witt, Pl. 3, figs. 8-11.
2007 Cyprideis torosa (Jones, 1850); Medley et al., Pl. 1, fig. e.
2012 Cyprideis torosa (Jones, 1850); Lucena-Moya et al., p. 6.
2011 Cyprideis torosa; Frenzel et al., p. 59.
2013 Cyprideis torosa (Jones, 1850); Valls et al., Fig 3, G-I.
2014 Cyprideis torosa (Jones, 1850); Chekhovskaya et al., p. 213, Pl. 2, fig. 7.
2015 Cyprideis torosa (Jones, 1850); Altınsaçlı et al., p. 379.
2015 Cyprideis torosa (Jones, 1850); Schornikov, Pl. 1, figs. 15-18.
2016 Cyprideis torosa (Jones, 1850); Bejaoui et al., Fig. 7, G-I.
2016 Cyprideis torosa (Jones); Baak et al., Figs. 4, 18.
2018 Cyprideis ex C. torosa (Jones, 1850) group; Sciuto et al., p. 11, Fig. 6.10-12
Remarks: Following Ligios and Gliozzi (2012), who point to the remarkable similarity of C. agrigentina, C. ruggierii, C. torosa, and in part C. crotonensis and C. calabra, we included these species in a comprehensive informal "C. torosa group". Wouters (2016) also confirms that Cyprideis torosa Jones, 1850, is a single, highly variable, polymorphic, and widely distributed species, with locally different populations. This group includes euryhaline and eurythermal species that can live from freshwater to hypersaline water (sebkha) (Athersuch et al., 1989; Boomer et al., 1996; Bejaoui et al., 2016). It has been reported from western and southern Europe, i.e. the Mediterranean coasts, including the Mediterranean Isles, and from the Atlantic coasts of west and northwest Europe. The species is also known from North Africa (Bejaoui et al., 2016), Eurasia, central and southwest Asia, (Black Sea, Caspian Sea, Lake Aral, and Lake Issyk Kul), and China (Wouters, 2002). It was also found in hypersaline environment at Santa Pola, a coastal salt marsh of the western Mediterranean (Mezquita et al., 2011), in brackish estuaries and lagoons of mainland Portugal (Cabral et al., 2016), and in coastal mesohaline lagoons in Turkey (Altinsaçli et al., 2015). Fossil specimens are reported from the Miocene to Recent (Meisch, 2000).
Family LEPTOCYTHERIDAE Hanai, 1957
Subfamily Leptocytherinae Hanai, 1957
Genus Leptocythere Sars, 1925
Type species: Cythere pellucida Baird, 1850.
Amnicythere propinqua (Livental, 1929)
(Pl. 1
![]() ,
fig. 5)
,
fig. 5)
1929 Cythere propinqua Livental, p. 20, Pl. 1, figs. 21-22.
1996 Leptocythere cymbula Livental, 1929; Boomer et al., p. 81, Fig. 4 A-H.
1999 Leptocythere propinqua Livental; Gliozzi, p. 199, Pl. 1, fig. c.
2004 Amnicythere cymbula Olteanu, p. 4.
2007 Amnicythere propinqua (Livental, 1929); Gliozzi et al., p. 331.
2008 Amnicythere propinqua (Livental, 1929); Gliozzi & Grossi, p. 290.
2008 Amnicythere propinqua (Livental, 1929); Grossi & Gennari, p. 77.
2014 Amnicythere cymbula (Livental, 1929), Chekhovskaya et al., p. 213, Pl. 1, fig. 8.
2016 Amnicythere propinqua Livental; Stoica et al., p. 859, Pl. 4, figs. 1-11.
2018 Amnicythere propinqua Livental; Williams et al., p. 56, Fig. 12 (1).
Remarks: A. propinqua is typical of shallow and oligo mesohaline waters. It is reported from the Upper Miocene strata (Pontian) of the Taman Peninsula (Azov Sea) (Stoica et al., 2016), from the Upper Pliocene and post Pliocene of the Caspian and Black Sea regions (Boomer et al., 1996), from the Upper Pleistocene and Holocene of the northern Caspian Sea, where it is commonly reported at depth of 2.5-10 m with salinity of 7-13.5 (Chekhovskaya et al., 2014), and from the Holocene of the Black Sea, where it is considered to be part of the Ponto-Caspian (brackish) assemblages (Williams et al., 2018).
In the Mediterranean region A. propinqua is reported from Lago-Mare events in the northern and central Apennines (Gliozzi et al., 2007; Gliozzi & Grossi, 2008; Grossi & Gennari, 2008), in the upper Messinian of Aléria Basin (Corsica) (Carbonnel, 1978), and in the upper Messinian of the Moncucco quarry (Torino Hill) (Trenkwalder et al., 2008).
The genus Amnicythere is widespread in the Miocene in the Paratethysian with Loxoconchissa, Loxocorniculina, Pontoniella, and Zalanyiella. These taxa migrated to the Mediterranean domain during the Messinian Lago-Mare event (Grossi & Gennari, 2008), and A. propinqua is therefore indicated as Paratethysian (Gliozzi et al., 2005; Gliozzi & Grossi, 2008).
Subfamily Mediocytherideisinae Mandelstam, 1960
Genus Mediocytherideis Mandelstam, 1956
Type species: Cytherideis apatoica Schweyer, 1949, by original designation.
Mediocytherideis (Sylvestra) punctata Ligios et al., 2008
(Pl. 1
![]() ,
fig. 6)
,
fig. 6)
2008 Mediocytherideis (Sylvestra) punctata Ligios et al., p. 156, Pl. 4, figs. 1-10; Pl. 5, figs. 1,2.
2012 Mediocytherideis (Sylvestra) punctata Ligios et al.; Ligios et al., p. 357
The reports of all the species belonging to the genus Mediocytherideis refer to brackish environments (Ligios et al., 2008). The genus is reported from the Upper Miocene sediments of Lake Pannon, from where it spread, alongside other taxa, to Recent Caspian Sea (Cziczer et al., 2009), and from the Upper Miocene of the Strymon Basin (northern Greece) (Grossi et al., 2015). Mediocytherideis is considered as a Paratethysian genus, while the species M. punctata is a Mediterranean species with Paratethysian affinity (Gliozzi et al., 2007; Ligios et al., 2008).
The characters of the microfauna found along the two sections analysed in the present paper highlight that, during the late Messinian post-evaporitic phase, thin basins of tectonic origin formed above the evaporitic and pre-evaporitic substratum also in the North African region (Rouchy et al., 2001; Rouchy & Caruso, 2006; Sciuto et al., 2018, inter alia). These basins, which could at first be assimilated to open lagoons, were, at certain times of their evolution, isolated from the sea and colonized by brackish and even fresh-water species when river inputs prevailed (Lago-Mare fauna); in the absence of fluvial inputs, conditions of hypersalinity, like sebka, would have been established.
The shallow-water character of these basins is also indicated by the
finding of charophyte gyrogonites, the best preserved of
which are referable to Chara hispida Linnaeus, 1753
(Pl. 1
![]() ,
fig. 15). C. hispida
lives in oligotrophic, freshwater, shallow-lake environments, peatland,
mud-calcareous gjtzia (i.e., floating islands); it can
occasionally be found also in brackish waters. Therefore, it may be
characterized as tolerant to salinity at depth from about 0.5 m to about 3.5 m (Barinova
et al., 2014, and references therein).
,
fig. 15). C. hispida
lives in oligotrophic, freshwater, shallow-lake environments, peatland,
mud-calcareous gjtzia (i.e., floating islands); it can
occasionally be found also in brackish waters. Therefore, it may be
characterized as tolerant to salinity at depth from about 0.5 m to about 3.5 m (Barinova
et al., 2014, and references therein).
According to Gliozzi et al. (2007) and Ligios et al. (2008), Amnicythere propinqua, Cyprideis agrigentina, and Mediocytherideis punctata can be considered as species with Paratethysian affinity.
The finding of species belonging to the genus Miocyprideis in the lower part of the SAL section, would indicate warm climatic conditions for this stratigraphic level. Indeed, Recent species of the genus Miocyprideis live only in warm tropical shallow waters of the Indo-Pacific region (Maddocks, 1995, inter alia) and in the Atlantic coasts (Carbonnel, 1986). In the Recent Mediterranean the genus is not reported and its disappearance could be linked to Plio-Pleistocene cooling.
Finally, the discovery, both in Sicily and Tunisia, of C. torosa, suggests that the Sicilian Channel high played a modest role in preventing the migration of the non marine fauna from NE to SW in the paleomediterranean area during the Lago-Mare event (Sciuto et al., 2018; present paper).
The authors are grateful to the Editor and to the anonymous referees for the suggestions on the manuscript. Special thanks are also due to Mr. Alfio Viola (Electronic Microscopy Laboratory, Earth Science Section, Catania University) for SEM assistance. Palaeoecological Research Group contribution no. 463. This research was supported by the University of Catania, Progetto PiaCeRi: "Biodiversità e paleobiodiversità di invertebrati e macroalghe di ambienti marini" - Piano Incentivi per la Ricerca di Ateneo 2020-22 linea di intervento 2.
Abdi A.M., Seaquist J., Tenenbaum D.E., Eklundh L. & Ardö J. (2014).- The supply and demand of net primary production in the Sahel.- Environmental Research Letters, Bristol, vol. 9, article 094003, 11 p.
Addicott W.O., Snavely P.D. Jr, Bukry D. & Poore R.Z. (1978).- Neogene stratigraphy and paleontology of southern Almeria Province, Spain: An overview.- Geological Survey Bulletin, Washington - DC, vol. 1454, p. 1-49.
Akdemir D. (2008).- Differences in Ostracoda (Crustacea) assemblages between two maar lakes and one sinkhole lake in the Konja region of Turkey.- Turkish Journal of Zoology, Ankara, vol. 32, p. 107-113.
Altinsaçli S., Altinsaçli S. & Paçal F.P. (2015).- Diversity, species composition and habitat preferences of Ostracoda (Crustacea) in seven karstic springs of varying salinity in Milas (Muğla, Turkey).- International Journal of Fisheries and Aquatic Studies, Delhi, vol. 3, p. 374-390.
Aruta L. (1982).- Gli ostracodi del Saheliano (Miocene medio-superiore) di C. Pestavecchia (Bonfornello, Palermo).- Bollettino della Società Paleontologica Italiana, Milano, vol. 21, no. 1, p. 113-132.
Aruta L. & Ruggieri G. (1980).- Nuovo ostracode marino del Saheliano dell'Italia meridionale.*- Bollettino della Società Paleontologica Italiana, Milano, vol. 19, no. 1, p. 21-24.
Athersuch J., Horne D.J. & Whittaker J.E. (1989).- Marine and brackish water ostracods.- Synopses of the British Fauna, Leiden, vol. 43, 343 p.
Baak C.G.C. van, Stoica M., Grothe A., Aliyeva E. & Krijgsman W. (2016).- Mediterranean-Paratethys connectivity during the Messinian salinity crisis: The Pontian of Azerbaijan.- Global and Planetary Change, vol. 141, p. 63-81.
Baird W. (1850).- The natural history of the British Entomostraca.- Ray Society, London, 364 p.
Bakrač K., Hajek-Tadesse V., Miknić M., Grizelj A., Hećimović I. & Kovačić M. (2010).- Evidence for Badenian local sea level changes in the proximal area of the North Croatian Basin.- Geologia Croatica, Zagreb, vol. 63, no. 3, p. 259-269.
Barinova S., Romanov R. & Solak C.N. (2014).- New record of Chara hispida (L.) Hartm. (Streptophyta: Charophyceae, Charales) from the Işıklı Lake (Turkey) and critical checklist of Turkish charophytes.- Natural Resources and Conservation, vol. 2, p. 33-42.
Bédir M. (1995).- Mécanismes géodynamiques des bassins associés aux couloirs de décrochements de la marge atlasique de la Tunisie. Séismo-stratigraphie, séismo-tectonique et implications pétrolières.- Thèse d'État, Faculté des Sciences de Tunis, Université de Tunis El Manar II, 407 p.
Bédir M., Tlig S., Bobier C.L. & Issaoui N. (1996).- Sequence stratigraphy, Basin dynamics and petroleum geology of Miocene from the eastern Tunisia.- American Association of Petroleum Geologists, Bulletin, Tulsa - OK, vol. 80, no. 1, p. 63-81.
Bejaoui S., Sciuto F., Karoui-Yaakoub N., Reitano A. & Ali Nebiha B.H. (2016).- Pleistocene microfauna evolution along the southern side of the Bizerte Lake (Sidi Mansour section, northeast Tunisia): Palaeoenvironmental interpretation.- Annales de Paléontologie, Paris, vol. 102, p. 151-159.
Beker K., Tunoglu C. & Ertekin I.K. (2008).- Pliocene-Lower Pleistocene Ostracoda fauna from Insuyu Limestone (Karapinar-Konia/Central Turkey) and its paleoenvironmental implications.- Geological Bulletin of Turkey, Ankara, vol. 51, no. 1, p. 1-31.
Ben Moshe L., Ben-Avraham Z., Enzel Y. & Schattner U. (2020).- Estimating drawdown magnitudes of the Mediterranean Sea in the Levant basin during the Lago Mare stage of the Messinian Salinity Crisis.- Marine Geology, vol. 427, article 106215, 13 p.
Ben Salem H. (1992).- Contribution à la connaissance de la géologie du Cap Bon : Stratigraphie, tectonique et sédimentologie.- Thèse de 3ème Cycle, Faculté des Sciences de Tunis, Université de Tunis El Manar II, 203 p.
Ben Salem H. (1998).- Les formations miocènes post-Saouaf des environs de Nabeul (Cap Bon) et leurs équivalents offshore et en Tunisie sud atlasique.- Notes du Service Géologique de Tunisie, Tunis, vol. 64, p. 123-128.
Ben Youssef M., Ghanmi M., Gaied M.S., Rabia M.C. & Razgallah S. (2002).- Enregistrements biosédimentaires des événements géodynamiques à la fin du Miocène (Messinien) en Tunisie nord-orientale : Dualité entre l'eustatisme et la tectonique. In: 3ème Congrès Français de Stratigraphie - Lyon, p. 41-42.
Benson R.H. (1973a).- An ostracodal view of the Messinian Salinity Crisis. In: Drooger C.W. (ed.), Messinian events in the Mediterranean.- North-Holland Publishing Company, Amsterdam, p. 235-242.
Benson R.H. (1973b).- Psychrospheric and continental ostracoda from ancient sediments in the floor of the Mediterranean. In: Ryan W.B.F., Hsü K.J., Cita M.B, Dumitrica P., Lort J.M., Mayne W., Nesteroff W.D., Pautot G., Stradner H. & Wezel F.C. (eds.).- Initial Reports of the Deep Sea Drilling Project, Washington - DC, vol. XIII, § 36.2, p. 1002-1008.
Benson R.H. (1978).- The paleoecology of the ostracodes of DSDP Leg 42A. In: Hsü K.J., Montadert L., Garrison R.G., Fabricius F.H., Kidd R.B., Müller C., Cita M.B, Bizon G, Wright R.C., Erickson A.J., Bernouilli D. & Mélières F. (eds.).- Initial Reports of Deep Sea Drilling Project, Washington - DC, vol. XLII, § 35, p. 777-787.
Besème P. & Kamoun Y. (1988).- Le Messinien marin de Ksour Essaf (Sahel, Tunisie orientale) : Une étude stratigraphique, sédimentologique et paléoécologique.- Revue des Sciences de la Terre, Borj Cedua, vol. 8, p. 129-142.
Bismuth H. (1984).- Les unités lithostratigraphiques du Miocène en Tunisie orientale.- Société des Sciences de la Terre de Tunisie, Tunis, 3 p.
Bonaduce G., Masoli M. & Pugliese N. (1988).- Remarks on the benthic Ostracoda on the Tunisian shelf. In: Hanai T., Ikeya N. & Ishizaki K. (eds.), Evolutionary biology of Ostracoda its fundamentals and applications. Proceedings of the Ninth International Symposium on Ostracoda, Japan (29 July-2 August 1985).- Developments in Palaeontology and Stratigraphy, vol. 11, p. 449-466.
Bonaduce G. & Sgarrella F. (1999).- Paleoecological interpretation of the latest Messinian sediments from southern Sicily (Italy).- Memorie della Società Geologica Italiana, Roma, vol. 54, p. 83-91.
Bonaduce G., Ruggieri G., Russo A. & Bismuth H. (1992).- Late Miocene ostracods from the Ashtart 1 well (Gulf de Gabès, Tunisia).- Bollettino della Società Paleontologica Italiana, Milano, vol. 31, no. 1, p. 3-93.
Boomer I., Whatley R. & Aladin N.V. (1996).- Aral Sea Ostracoda as environmental indicators.- Lethaia, Oslo, vol. 29, p. 77-85.
Brady G.S. (1880).- Report on the Ostracoda dredged by H.M.S. Challenger during the Years 1873-1876.- Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873-76. Zoology 1 (part 3), 184 p. (45 Pls.). URL: http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-03/README.htm
Brady G.S. & Norman A.M. (1889).- A monograph of the marine and freshwater Ostracoda of the North Atlantic and of Northwestern Europe. Section 1, Podocopa.- Scientific Transactions of the Royal Dublin Society (Series II), vol. IV, p. 63-270.
Burollet P.F. (1951).- Étude géologique des bassins mio-pliocènes du Nord-Est de la Tunisie : Région entre Mateur, Ferryville et Porto-Farina.- Annales des Mines et de la Géologie, Tunis, vol. 8, p. 94.
Burollet P.F. (1956).- Contribution à l'étude stratigraphique de la Tunisie centrale.- Annales des Mines et de la Géologie, Tunis, vol. 18, 350 p.
Butler R.W.H., Grasso M. & Lickorish H. (1995).- Plio-Quaternary megasequence geometry and its tectonic controls within the Maghrebian thrust belt of south-central Sicily.- Terra Nova, vol. 7, p. 171-178.
Cabral M.C., Fatela F., Lopes V., Freitas M.C. & Andrade C. (2016).- Cyprideis torosa (Jones, 1850) in mainland Portugal: What do we know?- Journal of Micropalaeontology, London, vol. 36, no. 1, p. 94-112.
Carbonnel G. (1978).- La zone à Loxoconcha djaffarovi Schneider (Ostracoda, Miocène supérieur) ou le Messinien de la vallée du Rhône.- Revue de Micropaléontologie, Paris, vol. 21, no. 3, p. 106-118.
Carbonnel G. (1986).- Ostracodes tertiaires, Paléogène à Néogène, du Basin sénégalo-guinéen.- Documents du Bureau de Recherches Géologiques et Minières, Orléans, vol. 101, p. 34-231.
Carbonnel G. & Peypouquet J.-P. (1979).- Les ostracodes des series du Bassin de l'Omo.- Bulletin de l'Institut Géologique du Bassin d'Aquitaine Bordeaux, vol. 25, p. 167-199.
Chekhovskaya M.P., Stepanova A. Yu., Khusid T.A., Matul A.S. & Rakowski A.Z. (2014).- Late Pleistocene-Holocene ostracod assemblages of the Northern Caspian Sea shelf.- Oceanology, vol. 54, no. 2, p. 212-221.
Cipollari P., Cosentino D., Radeff G., Schildgen T.F., Faranda C., Grossi F., Gliozzi E., Smedile A., Gennari R., Darbas G.¸ Dudas F.Ö., Gürbüz K., Nazik A. & Hechtler H. (2013).- Easternmost Mediterranean evidence of the Zanclean flooding event and subsequent surface uplift: Adana Basin, southern Turkey. In: Robertson A.H.F., Parlak O. & Ünlügenç U.C. (eds.), Geological Development of Anatolia and the Easternmost Mediterranean Region.- Geological Society London, Special Publications, vol. 372, p. 473-493.
Cita M.B., Wright R.C., Ryan W.B.F. & Longinelli A. (1978).- Messinian Paleoenvironments. In: Hsü K.J., Montadert L., Garrison R.G., Fabricius F.H., Kidd R.B., Müller C., Cita M.B, Bizon G, Wright R.C., Erickson A.J., Bernouilli D. & Mélières F. (eds.).- Initial Reports of Deep Sea Drilling Project, Washington - DC, vol. XLII, § 53, p. 1003-1035.
Clauzon G., Suc J.-P., Gautier F., Berger A. & Loutre M.F. (1996).- Alternate interpretation of the Messinian Salinity Crisis: Controversy resolved?- Geology, Boulder - CO, vol. 24, no. 4, p. 363-366.
Colleuil B. (1976).- Étude stratigraphique et néotectonique des formations néogènes et quaternaires de la région de Nabeul-Hammamet (Cap-Bon, Tunisie).- Diplôme d'Études Supérieures de Sciences Géologiques, Nice, 94 p.
Cosentino D., Gliozzi E. & Pipponzi G. (2007).- The late Messinian Lago-Mare episode in the Mediterranean Basin: Preliminary report on the occurrence of Paratethyan ostracod fauna from central Crete (Greece).- Geobios, Villeurbanne, vol. 40, p. 339-349.
Cziczer I., Magyar I., Pipík R., Böhme M., Coric S., Bakrac K., Süto-Szentai M., Lantos M., Babinszki E. & Müller P. (2009).- Life in the sublittoral zone of long-lived Lake Pannon: Paleontological analysis of the Upper Miocene Szák Formation, Hungary.- International Journal of Earth Sciences (Geologische Rundschau), vol. 98, no. 7, p. 17411766.
Decima A. (1964).- Ostracodi del gen. Cyprideis Jones del Neogene e del Quaternario italiani.- Palaeontographia Italica, Pisa, vol. 57, p. 81-133.
Demarcq G., Meon-Vilain H., Miguet R. & Kujawski H. (1976).- Un bassin paralique néogène : Celui de Skanes-Monastir (Tunisie centrale).- Notes du Service Géologique de Tunisie, Tunis, vol. 42, p. 97-147.
Devoto G. (1965).- Lacustrine Pleistocene in the Lower Liri Valley.- Geologica Romana, vol. 4, p. 291-368.
Di Geronimo I., Esu D. & Grasso M. (1989).- Gli strati a "congerie" del Messiniano superiore del margine nord occidentale ibleo. Caratteristiche faunistiche e possibili implicazioni paleogeografiche e paleoclimatiche.- Atti Accademia Peloritana dei Pericolanti, Messina, vol. 68, no. 1, p. 129-150.
Doruk N. (1973).- On Chrysocythere paradisus sp. nov.- Stereo-Atlas of Ostracod Shells, London, vol. 1, p. 89-92.
El Euch-El Koundi N., Ferry S., Suc J.-P., Clauzon G., Melinte-Dobrinescu M.C., Gorini C., Safra A. & Zargouni F. (2009).- Messinian deposits and erosion in northern Tunisia: Inferences on Strait of Sicily during the Messinian Salinity Crisis.- Terra Nova, vol. 21, p. 41-48,
Faranda C., Cipollari P., Cosentino D., Gliozzi E. & Pipponzi G. (2008).- Late Miocene ostracod assemblages from eastern Mediterranean coral reef complexes (central Crete, Greece).- Revue de Micropaléontologie, Paris, vol. 51, p. 287-308.
Faranda C., Gliozzi E., Cipollari P., Grossi F., Darbaş G., Gürbüz K., Nazik A., Gennari R. & Cosentino D. (2013).- Messinian paleoenvironmental changes in the easternmost Mediterranean Basin: Adana Basin, southern Turkey.- Turkish Journal of Earth Science, Ankara, vol. 22, p. 839-863.
Fournié D. (1978).- Nomenclature lithostratigraphique des séries du Crétacé inférieur au Tertiaire de la Tunisie.- Bulletin du Centre de Recherches Exploration Production elf-Aquitaine, Pau, vol. 2, p. 97-14.
Frenzel P., Schulze I., Pint A., Boomer I. & Feike M. (2011).- Salinity dependant morphological variation in Cyprideis torosa.- Joannea Geologie und Paläontologie, Graz, vol. 11, p. 59-61.
Frigui M. (2003).- Le Néogène aux environs de la région de Mahdia : Enregistrements tectono-eustatiques et conséquences paléogéographiques.- D.E.A., Faculté des Sciences de Tunis, Université de Tunis El Manar II, 104 p.
Frigui M., Ben Youssef M. & Ouaja M. (2016).- Evidences of "Lago-Mare" episode around the Messinian-Pliocene boundary in eastern Tunisia (central Mediterranean).- Journal of African Earth Sciences, vol. 123, p. 57-74.
Gaaloul N. (1995).- Les environnements siliciclastiques du Néogène du Sahel de la Tunisie. Palynologie et biosédimentologie.- Thèse de spécialité, Faculté des Sciences de Tunis, Université de Tunis El Manar II, 265 p.
Garcia-Castellanos D., Estrada F., Jiménez-Munt I., Gorini C., Fernàndez M., Vergés J. & De Vicente R. (2009).- Catastrophic flood of the Mediterranean after the Messinian salinity crisis.- Nature, vol. 462, p. 778-782.
Gliozzi E. (1999).- A late Messinian brackish water ostracod fauna of Paratethyan aspect from Le Vicenne Basin (Abruzzi, central Apennines, Italy).- Palæogeography, Palæoclimatology, Palæoecology, vol. 151, p. 191-208.
Gliozzi E., Ceci M.E., Grossi F. & Ligios S. (2007).- Paratethyan ostracod immigrants in Italy during Late Miocene.- Geobios, Villeurbanne, vol. 40, p. 325-337.
Gliozzi E. & Grossi F. (2008).- Late Messinian Lago-mare ostracod palaeoecology: A correspondence analysis approach.- Palæogeography, Palæoclimatology, Palæoecology, vol. 264, p. 288-295.
Gliozzi E. & Mazzini I. (1998).- Paleoenvironmental analysis of the 250.000 years Quaternary sediment core of Valle di Castiglione (Latium, Italy) using ostracods. In: Crasquin-Soleau S., Braccini E. & Lethiers F. (eds.), What about Ostracoda!- Bulletin du Centre Recherche elf Exploration Production, Pau, Mémoire 20, p. 70-90.
Gliozzi E., Rodriguez-Lazaro J., Nachite D., Martin-Rubio M. & Bekkali R. (2005).- An overview of Neogene brackish leptocytherids from Italy and Spain: Biochronological and palaeogeographical implications.- Palæogeography, Palæoclimatology, Palæoecology, vol. 225, p. 283-301.
Gross M. (2004).- Zur Ostracodenfauna (Crustacea), Paläoökologie und Stratigrafie der Tongrube Mataschen (Unter-Pannonium, Steirisches Becken, Österreich).- Joannea Geologie and Paläontologie, Graz, vol. 5, p. 49-129.
Gross M., Minati K., Danielopol L. & Piller W.E. (2008).- Environmental changes and diversification of Cyprideis in the Late Miocene of the Styrian Basin (Lake Pannon, Austria).- Senckenbergiana lethaea, Frankfurt am Main, vol. 88, p. 161-181.
Grossi F. & Gennari R. (2008).- Palaeoenvironmental reconstruction across the Messinian Zanclean boundary by means of ostracods and foraminifers: The Montepetra borehole (Northern Apennine, Italy).- Atti del Museo Civico di Storia Naturale di Trieste, vol. 53 (Suppl.), p. 67-88.
Grossi F., Gliozzi E., Anadónc P., Castorina F. & Voltaggio M. (2015).- Is Cyprideis agrigentina Decima a good paleosalinometer for the Messinian Salinity Crisis? Morphometrical and geochemical analyses from the Eraclea Minoa section (Sicily).- Palæogeography, Palæoclimatology, Palæoecology, vol. 419, p. 75-89.
Grossi F., Gliozzi E. & Cosentino D. (2011).- Paratethyan ostracod immigrants mark the biostratigraphy of the Messinian Salinity Crisis.- Joannea Geologie und Paläontologie, Graz, vol. 11, p. 66-68.
Guerra-Merchán A., Serrano F., Garcés M., Gofas S., Esu D., Gliozzi E. & Grossi F. (2010).- Messinian Lago-Mare deposits near the Strait of Gibraltar (Malaga Basin, S Spain).- Palæogeography, Palæoclimatology, Palæoecology, vol. 285, p. 264-276.
Hajek-Tadesse V. & Prtoljan B. (2011).- Badenian Ostracoda from the Pokupsko area (Banovina, Croatia).- Geologica Carpathica, Bratislava, vol. 62, no. 5, p. 447-461.
Hanai T. (1957).- Studies on the Ostracoda from Japan. 1 - Subfamily Leptocytherinae, new Subfamily.- Journal of the Faculty of Science, Tokyo University Section, vol. 2, no. 10. p. 431-468.
Henderson P.A. (1990).- Freshwater Ostracoda. In: Kermack D.M. & Barnes R.S.K. (eds.), Synopses of the British Fauna.- The Linnean Society of London, vol. 42, p. 228.
Hooyberghs H.J.F. & Ben Salem H. (1999).- Biostratigraphie des foraminifères planctoniques de la formation Saouaf (Tortonien) dans le synclinal de Takelsa (Cap Bon, Tunisie).- Notes du Service Géologique de Tunisie, Tunis, vol. 66, p. 113-123.
Hsü K.J., Ryan W.F.B. & Cita M.B. (1973).- Late Miocene desiccation of the Mediterranean.- Nature, vol. 242, p. 240-244.
Jones T.R. (1850).- Description of Entomostraca of the Pleistocene beds of Newbury, Copford, Clacton and Grays.- Annals and Magazine of Natural History, London, vol. 2, no. 6, p. 25-28.
Jones T.R. (1857).- A monograph of the Tertiary Entomostraca of England.- Palaeontographical Society of London, vol. 9, no. 2, p. 1-68.
Kamoun Y. (1981).- Néotectonique dans la région de Monastir-Mahdia (Tunisie orientale).- Thèse de Doctorat 3ème cycle, Université Paris Sud, 175 p.
Kamoun Y., Hooyberghs H.J.F., Ben Youssef M., Zouari H. & Gaaloul N. (2001).- Signature de la transgression du Messinien inférieur en Tunisie orientale. Coupe de l'oued Macera.- Notes du Service Géologique de Tunisie, Tunis, vol. 68, p. 5-14.
Kaufmann A. (1900).- Neue Ostracoden aus der Schweiz.- Zoologischer Anzeiger, vol. 23, p. 131-133.
Kollmann K. (1960).- Cytherideinae und Schulerideinae n. subfam. (Ostracoda) aus dem Neogen des Ostl. Oesterreich.- Mitteilungen des Geologischen Gesellschaft in Wien, vol. 51, p. 89-155.
Kováč M., Barath I., Kovačova-Slamkova M., Pipík R., Hlavaty I. & Hudačkova N. (1998).- Late Miocene Paleoenvironments and Sequence Stratigraphy: Northern Vienna Basin.- Geologica Carpathica, Bratislava, vol. 49, p. 445-458.
Krijgsman W., Capella W., Simon, D., Hilgen F.J., Kouwenhoven T.J., Meijer P.T., Sierro F.J., Tulbure M.A., Berg B.C.J. van den, Schee M. van der & Flecker R. (2018).- The Gibraltar Corridor: Watergate of the Messinian Salinity Crisis.- Marine Geology, vol. 403, p. 238-246.
Krstic N. & Pietrzeniuk E. (1972).- Paijenborchella (Eopaijenborchella) laskarevi, eine Ostracoden art aus dem Oberen Torton des Pannonischen Beckens.- Geologie mit Beiheften, Berlin, vol. 21, no. 1, p. 100-109.
Latreille P.A. (1806).- Genera crustaceorum et insectorum.- A. Koenig, Paris, 303 p.
Ligios S., Bossio A. & Gliozzi E. (2008).- New species of Mediocytherideis (Ostracoda, Mediocytherideisinae) in the brackish Messinian of Italy.- Bollettino della Società Paleontologica Italiana, Milano, vol. 47, no. 2, p. 147-167.
Ligios S. & Gliozzi E. (2012).- The genus Cyprideis Jones, 1857 (Crustacea, Ostracoda) in the Neogene of Italy: A geometric morphometric approach.- Joannea Geologie und Paläontologie, Graz, vol. 11, p. 110-112.
Linnaeus C. (1753).- Species plantarum, exhibentes plantas rite cognitas ad genera relatas cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum systema sexuale digestas.- Holmiae: Impensis Laurentii Salvii, Stokholm, 824 p. DOI: https://doi.org/105962/bhl.title.699
Livental V.E. (1929).- Ostracoda from the Akchagyl and Apscheron strata along the Babazansk section.- publisher unknown, Bacu, 420 p. [in Russian].
Lucena-Moya P., Abraín R., Pardo I., Hermida B. & Domínguez M. (2012).- Invertebrate species list of coastal lagoons in the Balearic Islands.- Transitional Waters Bulletin, Lecce, vol. 4, p. 1-11.
Maddocks R.F. (1995).- Mangrove-mud Ostracoda of Nosy Be, Madagascar. In: Riha J. (ed.), Ostracoda and biostratigraphy.- A.A. Balkema, Rotterdam, p. 351-363.
Mandelstam M.I. (1956).- Rod Mediocytherideis gen. nov. In: Mandelstam M.I., Schneyder G.F. & Zanina I.E. (eds.), Order Ostracoda. New families and genera of Invertebrates.- Vses. Naukno Issled. Geol. Inst. (VSEGEI), vol. 12, p. 137-138 [in Russian].
Mandelstam M.I. (1960).- Subfamily Mediocytherideinae subfam. nov. In: Tschernysheva M.I. (ed.), Trilobitomorpha and Crustacea.- SSSR, Osnovy Palentologii, 127 p.
Mannaï-Tayech B. (2006).- Les séries silicoclastiques miocènes du Nord-Est au Sud-Ouest de la Tunisie : Une mise au point.- Geobios, Villeurbanne, vol. 39, p. 71-84.
Manzi V., Gennari R., Hilgen F., Krijgsman W., Lugli S., Roveri M. & Sierro F.J. (2013).- Age refinement of the Messinian salinity crisis onset in the Mediterranean.- Terra Nova, vol. 25, p. 315322.
Mascle G. & Mascle J. (2019).- The Messinian salinity legacy: 50 years later.- Mediterranean Geoscience Reviews, vol. 1, p. 5-15.
Matzke-Karasz R. & Witt W. (2005).- Ostracods of the Paratethyan Neogene Kılıç and Yalakdere Formations near Yalova (İzmit Province, Turkey).- Zitteliana, München, vol. A45, p. 115-133.
Mazzini I., Anadon P., Barbieri M., Castorina F., Ferreli L., Gliozzi E., Mola M. & Vittori E. (1999).- Late Quaternary sea-level changes along the Tyrrhenian coast near Orbetello (Tuscany, central Italy): Palaeoenvironmental reconstruction using ostracods.- Marine Micropaleontology, vol. 37, p. 289-311.
Medley P., Tibert N.E., Patterson W.P., Allen C.H., Greer L. & Colin J.-P. (2007).- Paleosalinity history of middle Holocene lagoonal and lacustrine deposits in the Enriquillo Valley, Dominican Republic based on pore morphometrics and isotope geochemistry of Ostracoda.- Micropaleontology, Flushing - NY, vol. 53, p. 409-419.
Meisch C. (2000).- Freshwater Ostracoda of Western and Central Europe.- Süswasserfauna von Mitteleuropa, Band 8/3, 522 p.
Meisch C., Smith R.J. & Martens K. (2019).- A subjective global checklist of the extant non-marine Ostracoda (Crustacea).- European Journal of Taxonomy, vol. 492, p. 1-135.
Mezquita F., Aguilar-Alberola J.A., Carbonell E., Escrivà A., Rueda J., Schmit O. & Zamora L. (2011).- Introducciò als ostracodes (Crustacea: Ostracoda) castellonencs.- Actes del 1r Congrés sobre Fauna Castellonenca, p. 37-46.
Moissette P., Cornée J.-J., Mannaï-Tayech B., Rabhi M., André J.-P., Koskeridou E. & Meon H. (2010).- The western edge of the Mediterranean Pelagian Platform: A Messinian mixed siliciclastic-carbonate ramp in northern Tunisia.- Palæogeography, Palæoclimatology, Palæoecology, vol. 285, p. 85-103.
Nazik A., Turkmen B., Koc C., Aksoy E., Avşar N. & Yayik H. (2008).- Fresh and Brackish Water Ostracods of Upper Miocene Deposits, Arguvan/Malatya (Eastern Anatolia).- Turkish Journal of Earth Sciences, Ankara, vol. 17, p. 481-495.
Olteanu R. (2006).- Paleoecologia ecosistemelor salmastre din Bazinul Dacic.- GeoEcoMar, Bucuresti, 89 p.
Orszag-Sperber F., Rouchy J.-M. & Blanc-Valleron M.-M. (2000).- La transition Messinien Pliocène en Méditerranée orientale (Chypre) : La période du Lago-Mare et sa signification.- Comptes Rendus de l'Académie des Sciences (Series IIA - Earth and Planetary Science), Paris, vol. 331, no. 7, p. 483-490.
Pezelj D. & Sremac J. (2007).- Badenian marginal marine environment in the Medvednica Mt. (Croatia).- Joannea Geologie und Paläontologie, Graz, vol. 9, p. 83-84.
Pieri V., Martens K., Naselli-Flores L., Marrone F. & Rossetti G. (2006).- Distribution of recent ostracods in island waters of Sicily (Southern Italy).- Journal of Limnology, Verbania Pallanza, vol. 65, no. 1, p. 1-8.
Radu E. & Stoica M. (2005).- Lower Sarmatian microfauna from the hydrogeological Wellfa Hârlău (Iaşi county).- Acta Palaeontologica Romaniae, Bucarest, vol. 5, p. 413-421.
Ramdohr F.A. (1808).- Ueber die gattung Cypris Müller und drei zu derselben gehörige neue Arten.- Geselschaft Naturforschung Freunde Magazin neuesten Entdeckungen gesammter Naturkunde, Berlin, Jarhgang 2, p. 83-93.
Rodriguez-Làzaro J. & Martin-Rubio M. (2005).- Pliocene Ilyocyprididae (Ostracoda) from the Ebro Basin (N Spain).- Revue de Micropaléontologie, Paris, vol. 48, p. 37-49
Rossetti G., Martens K., Meisch C., Tavernelli S. & Pieri V. (2006).- Small is beautiful: Diversity of freshwater ostracods (Crustacea, Ostracoda) in marginal habitats of the province of Parma (Northern Italy).- Journal of Limnology, Verbania Pallanza, vol. 65, no. 2, p. 121-131.
Rouchy J.-M. & Caruso A. (2006).- The Messinian salinity crisis in the Mediterranean Basin: A reassessment of the data and an integrated scenario.- Sedimentary Geology, Tulsa - OK, vol. 188-189, p. 35-67.
Rouchy J.-M., Caruso A., Pierre C., Blanc-Valleron M.M. & Bassetti M.A. (2007).- The end of the Messinian salinity crisis: Evidences from the Chelif Basin (Algeria).- Palæogeography, Palæoclimatology, Palæoecology, vol. 254, p. 386-417.
Rouchy J.-M., Orszag-Sperber F., Blanc-Valleron M.-M., Pierre C., Rivière M., Combourieu-Nebout N. & Panayides I. (2001).- Paleoenvironmental changes at the MessinianPliocene boundary in the eastern Mediterranean (southern Cyprus basins): Significance of the Messinian Lago-Mare.- Sedimentary Geology, Tulsa - OK, vol. 145, no. 12, p. 93-117.
Roveri M., Manzi V., Gennari R., Iaccarino S. & Lugli S. (2008).- Recent advances in the Messinian stratigraphy of Italy and their Mediterranean-scale implication.- Bollettino della Società Paleontologica Italiana, Milano, vol. 47, p. 71-85.
Ruggieri G. (1967).- The Miocene and later evolution of the Mediterranean Sea. In: Adams C.G. & Ager D.V. (eds.), Aspects of Tethyan Biogeography.- Systematics Association, Publication 7, London, p. 283-290.
Ruggieri G. & Russo A. (1980).- Due nuovi generi di Osrracodi marini del Miocene superiore italiano.- Bollettino della Società Paleontologica Italiana, Milano, vol. 19, no. 1, p. 25-35.
Şafak Ü., Avşar N. & Meriç E. (1999).- Batı Bakırköy (İstanbul) Tersiyer Çökellerinin Ostrakod ve Foraminifer Topluluğu.- Maden Tetkik ve Arama Dergisi (Bulletin of the Mineral Research and Exploration), Ankara, vol. 121, p. 17-31.
Said Benzarti R., Colin J.-P., Gargouri S., Ben Ismail K., Torkhani H., Temani R., Mekki M., Ben Abdeladhim F. & Ayadi S. (2010).- Livret guide d'excursion. In: Said Benzarti R. (ed.), ROLF 23 (23ème Réunion des Ostracodologistes de Langue Française).- Tunis, 30 p.
Sars G.O. (1866).- Oversight af Norges marine Ostracoder.- Forhandlinger i Videnskabs Selskabet i Christiania, vol. 8, p. 1-130.
Sars G.O. (1925).- An account of the Crustacea of Norway with short descriptions and figures of all the species.- Bergen Museum, vol. 9, p. 73-208.
Schornikov E. (2015).- The taxonomic position of the Cyprideis species (Ostracoda, Cytherideidae) that occurs in Lake Issyk-Kul, NE Kyrgyzstan.- Micropaleontology, Flushing - NY, vol. 61, p. 37-48.
Schweier A.V. (1949).- On the Pliocene ostracoda of the northern Caucasus and lower Volga region with some new data on the systematics of fossil ostracods.- Trudy Vsesoyuznogo neftyanogo nauchno-issledovatelskogo geologorazvedochnogo instituta (VNIGRI) (Novaya Seriya), Leningrad, Vypusk 30, p. 7-68 [in Russian].
Sciuto F., Baldanza A., Temani R. & Privitera G. (2018).- New reports of Paratethyan ostracods affinity from the Mediterranean Basin (Sicily, Italy).- Palaeontologia Electronica, article 21.1.12A, 19 p.
Sciuto F., Rosso A., Sanfilippo R. & Maniscalco R. (2015).- New faunistic data on the Pleistocene environmental evolution of the south-western edge of the Hyblean plateau (SE Sicily).- Carnets Geol., Madrid, vol. 15, no. 5, p. 41-57. DOI: https://doi.org/10.4267/2042/56401
Stoica M., Krijgsman W., Fortuin A. & Gliozzi E. (2016).- Paratethyan ostracods in the Spanish Lago-Mare: More evidence for interbasinal exchange at high Mediterranean Sea level.- Palæogeography, Palæoclimatology, Palæoecology, vol. 441, p. 854-870.
Temani R., Ammar H. K. & Sciuto F. (2018).- New reports of Messinian Lago-Mare episodes from Tunisia: Ostracods and paleoenvironmental implications. In: Zhang Z., Khélifi N., Mezghani A. & Heggy E. (eds.), Patterns and mechanisms of climate, paleoclimate and paleoenvironmental changes from low-latitude regions.- Advances in Science, Technology & Innovation, p. 69-71.
Temani R., Sciuto F. & Ammar H.K. (2019).- Some reports of Paratethyan ostracods from the Central Mediterranean area.- International Journal of Paleobiology & Paleontology, Troy MI, vol. 2, no. 1, p. 1-3.
Trenkwalder S., Violanti D., D'Atri A., Lozar F., De la Pierre F. & Irace A. (2008).- The Miocene/Pliocene boundary and the Early Pliocene micropalaeontological record: New data from the Tertiary Piedmont Basin (Moncucco quarry, Torino Hill, Northwestern Italy).- Bollettino della Società Paleontologica Italiana, Milano, vol. 47, p. 87-103.
Uçak S., Külköylüoğlu O., Akdemir D. & Başak E. (2014).- Distribution, diversity and ecological characteristic of freshwater Ostracoda (Crustacea) in shallow aquatic bodies of the Ankara Region Turkey.- Wetlands, vol. 34, p. 309-324.
Valls L., Rued J. & Mesquita-Joanes F. (2013).- Dynamics of Ostracoda (Crustacea) assemblages in a Mediterranean pond system (Raco' de l'Olla, Albufera Nat. Park) with focus on the exotic species Candonocypris novaezelandiae (Baird, 1843).- Annales de Limnologie - International Journal of Limnology, Toulouse, vol. 49, p. 237-247.
Williams L.R., Hiscott R.N., Aksu A.E., Bradley L.R., Hornec D.J. & Stoica M. (2018).- Holocene paleoecology and paleoceanography of the southwestern Black Sea shelf revealed by ostracod assemblages.- Marine Micropaleontology, vol. 142, p. 48-66.
Wouters K. (2002).- On the distribution of Cyprideis torosa (Jones) (Crustacea, Ostracoda) in Africa, with the discussion of a new record from the Seychelles.- Bulletin de l'Institut Royal des Sciences Naturelles de Belgique (Biologie), Bruxelles, vol. 72, p. 131-140.
Wouters K. (2016).- On the modern distribution of the euryhaline species Cyprideis torosa (Jones, 1850) (Crustacea, Ostracoda).- Journal of Micropalaeontology, London, vol. 36, p. 21-30.
Zalanyi B. (1913).- Miocäne Ostrakoden aus Ungarn.- Mitteilungen aus dem Jahrbuchder Kgl. Ungarischen Geologischen Reichanstalt, Budapest, Band XXI, p. 81-152 (Pls. V-IX).
|
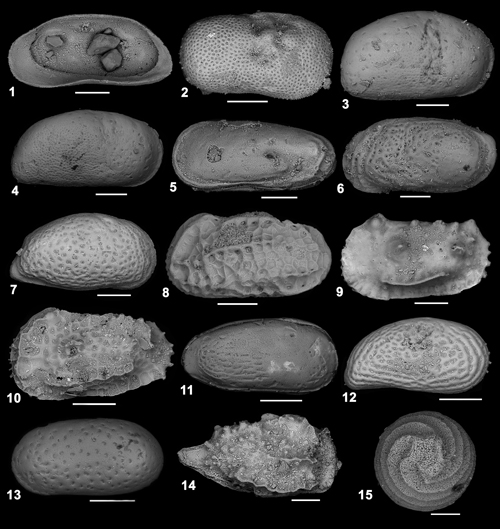
fig. 1 - Phlyctenophora farkasi
(Zalanyi, 1913). Right valve internal view (Scale bar 200 µm); |
|
|