◄ Carnets Geol. 20 (16) ►
![]()
Outline:
[1. Introduction]
[2. Geological and palaeontological framework]
[3. Systematic palaeontology]
[4. General discussion and concluding remarks] and ...
[Bibliographic references]
Dipartimento di Scienze della Terra, Universitý di Pisa, via Santa Maria 53, 56126 Pisa (Italy)
Museo di Storia Naturale, Universitý di Pisa, via Roma 79, 56011 Calci (Italy)
Gruppo Avis Mineralogia e Paleontologia Scandicci, p.za Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (Italy)
Museo di Storia Naturale, Universitý di Pisa, via Roma 79, 56011 Calci (Italy)
Gruppo Avis Mineralogia e Paleontologia Scandicci, p.za Vittorio Veneto 1, Badia a Settimo, 50018 Scandicci (Italy)
Studio Tecnico Geologia e Paleontologia, via F.lli Rosselli 4, S. Casciano Val di Pesa, 50026 Firenze (Italy)
Published online in final form (pdf) on October 14, 2020
DOI 10.2110/carnets.2020.2016
![]()
[Editor: Bruno Granier;
language editor: Stephen Carey]
![]()
Nowadays, the living species of the terrapin genus Mauremys (Testudinoidea: Geoemydidae) are mostly found in eastern Asia, but three of them inhabit the Western Palearctic ecozone. In Italy, occurrences of living individuals of Mauremys are interpreted as records of alien species; however, a growing fossil record demonstrates that this genus has inhabited Italy as recently as the Late Pleistocene. We report on a new fossil specimen of Mauremys from the Upper Pliocene (Piacenzian) marginal-marine deposits of Tuscany (central Italy). This find, consisting of a partial plastron and a loose neural, represents the second authentic report of Mauremys from the Italian Pliocene, as well as the first one from the Piacenzian of Italy. Therefore, it is a significant fossil that fills a gap in the chronostratigraphic distribution of Italian fossil Mauremys, helping ñ together with the Lower Pliocene holotype of Mauremys portisi ñ to bridge the rich Miocene and Pleistocene segments of this record. Moreover, two unusual scars observed on the external surface of the studied plastron are here referred to the ichnospecies Thatchtelithichnus holmani. These traces represent one of the few records worldwide of this rarely identified ichnospecies, as well as its geologically youngest published occurrence. Hypotheses regarding the origin of the Thatchtelithichnus traces are reevaluated, and an origin as attachment scars of aquatic ectoparasites (possibly ticks, leeches, or flukes) is reaffirmed as probable in cases of traces occurring on the exterior of the plastral bones of turtles.
• Geoemydidae;
• terrapin;
• turtle shell;
• parasitism;
• Piacenzian;
• Tuscany;
• palaeoherpetology;
• ichnology
Collareta A., Casati S., Zuffi M.A.L. & Di Cencio A. (2020).- First authentic record of the freshwater turtle Mauremys from the Upper Pliocene of Italy, with a new occurrence of the rarely reported ichnotaxon Thatchtelithichnus holmani.- Carnets Geol., Madrid, vol. 20, no. 16, p. 301-313.
PremiËre dÈcouverte attestÈe de la tortue d'eau douce Mauremys dans le PliocËne supÈrieur d'Italie, avec un nouveau signalement de Thatchtelithichnus holmani, un ichnotaxon rarement mentionnÈ.- De nos jours, les espËces du genre de tortues d'eau douce Mauremys (Testudinoidea : Geoemydidae) vivent principalement en Asie orientale, mais trois d'entre elles habitent l'Ècozone palÈarctique occidentale. En Italie, les dÈcouvertes d'individus vivants de Mauremys sont interprÈtÈes comme des signalements d'espËces ÈtrangËres ; cependant, un nombre croissant de fossiles montre que ce genre Ètait prÈsent en Italie dËs le PlÈistocËne supÈrieur. Nous signalons un nouveau spÈcimen fossile de Mauremys provenant de dÈpÙts marginaux marins du PliocËne supÈrieur (Plaisancien) de Toscane (Italie centrale). Cette dÈcouverte, comprenant un plastron partiel et une neurale dÈsarticulÈe, reprÈsente le second signalement confirmÈ de Mauremys dans le PliocËne italien et le premier dans le Plaisancien d'Italie. Ainsi, ce spÈcimen reprÈsente un fossile significatif puisque, avec l'holotype zanclÈen de Mauremys portisi, il comble la lacune entre les segments miocËne et plÈistocËne dans la distribution chronostratigraphique des fossiles italiens de Mauremys. En outre, deux cicatrices insolites observÈes sur la surface externe du plastron ÈtudiÈ sont attribuÈes ici ý l'ichno-espËce Thatchtelithichnus holmani. Il s'agit d'un des rares signalements de cette ichno-espËce ý l'Èchelle planÈtaire et aussi de son occurrence stratigraphique la plus jeune. Les hypothËses concernant l'origine des traces de type Thatchtelithichnus sont reconsidÈrÈes ý la lumiËre de notre dÈcouverte et leur interprÈtation en tant que traces cicatricielles d'attachement d'ectoparasites aquatiques (probablement des tiques, des sangsues ou des douves) est rÈaffirmÈe comme probable dans le cas de traces situÈes sur la face externe des plaques plastrales des tortues.
• Geoemydidae ;
• terrapËne ;
• carapace de tortue ;
• parasitisme ;
• Plaisancien ;
• Toscane ;
• palÈoherpÈtologie ;
• ichnologie
The extant terrapin genus Mauremys belongs to Geoemydidae, a family of testudinoid cryptodirans consisting of approximately 60 species in 25 genera (Barth et al., 2004). Currently, the living species of Mauremys exhibit a strongly disjunct distribution; they are mostly found in eastern Asia, but three of them ñ Mauremys caspica, Mauremys leprosa, and Mauremys rivulata - are distributed in the Western Palearctic ecozone. Two of these species, M. leprosa and M. rivulata, are found along the coasts of the Mediterranean Sea, the former in western North Africa (Morocco to western Libya) and the Iberian Peninsula (Keller & Busack, 2001), and the latter along the Mediterranean coast of the Balkan Peninsula, on several islands of the Eastern Mediterranean, in the coastal regions of western and southern Turkey, and along the Levantine coast (Barth et al., 2004). In Italy, where the only native freshwater turtles are Emys orbicularis and Emys trinacris, discoveries of living individuals of Mauremys are interpreted as records of alien species (Panzeri et al., 2014). However, a growing fossil record demonstrates that terrapins of the genus Mauremys have inhabited Italy as recently as the Late Pleistocene (Chesi et al., 2007) and the Italian Peninsula might even have been the route of a dispersal of Mauremys from the Mediterranean into the North African and Arabian domains in Late Miocene times (Chesi et al., 2009; Georgalis et al., 2020).
The currently monotypic ichnogenus Thatchtelithichnus was erected by Zonneveld et al. (2015) for circular to subcircular, ring-shaped grooves incised on the external surface of plastral bones of geoemydid turtles from the Eocene of Wyoming, USA. These traces were originally interpreted as the attachment scars of aquatic ectoparasites (possibly ticks, leeches, or spirorchid liver flukes) that infected the turtle while the latter was still living (Zonneveld et al., 2015). More recently, Wisshak et al. (2019) reinterpreted Thatchtelithichnus as a bioclaustration structure and, as such, not a valid ichnotaxon. Later, Zonneveld and Bartels (2020) provided new arguments in support of both the origin of Thatchtelithichnus by emplacement of ectoparasites and its ichnotaxonomic validity. In spite of their highly idiosyncratic aspect, to date, Thatchtelithichnus traces have only been reported by Zonneveld et al. (2015) and Zonneveld and Bartels (2020).
In the present work, we report on a new fossil record of Mauremys from the famous fossiliferous Pliocene deposits of the Valdelsa Basin (Tuscany, Italy), which has previously yielded significant remains of continental and marine vertebrates (e.g., Azzaroli, 1992; Abbazzi et al., 2008; Dominici et al., 2018; Collareta et al., 2020). This new specimen is here described and illustrated, and its palaeontological implications are discussed in the broader framework of the Italian fossil record of Mauremys. Furthermore, two unusual traces observed on this specimen are here characterised and referred to the only ichnospecies that is currently included in Thatchtelithichnus, i.e., Thatchtelithichnus holmani. This record is also discussed and its palaeobehavioural significance is evaluated.
The fossil terrapin described herein was
discovered by three of us (A.C., S.C. and A.D.C.) at an active quarry at La
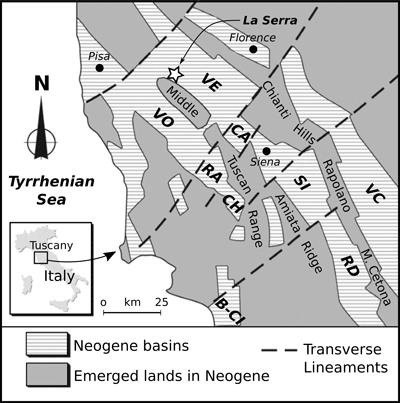
Serra village near San Miniato (Pisa Province, Tuscany, central Italy) (Fig. 1 ![]() ).
The GPS geographic coordinates of this quarry are 43∞ 39′ 28″ N; 10∞
48′ 57″ E. La Serra quarry lies at the northwestern corner of the
Valdelsa Basin, a 25 ◊ 60 km, NW-SE elongated extensional basin filled with some 1000 m of
Upper Neogene and Quaternary deposits (Benvenuti et al.,
2014). The sedimentary succession exposed at
this site has been referred by Dominici et al.
(2018) to the S6 Synthem recognised in the
Valdelsa Basin by Benvenuti et al. (2014). The S6 Synthem is a large-scale
depositional sequence that consists mostly of fluvial coarse-grained sandstones,
intertidal or coastal lagoon deposits, and shoreface and delta-front sandstones
(Benvenuti et al., 2014). At La Serra
quarry, a 25-m-thick stratigraphic section was measured by Garassino
et al. (2012), who recognised the following
informal units:
).
The GPS geographic coordinates of this quarry are 43∞ 39′ 28″ N; 10∞
48′ 57″ E. La Serra quarry lies at the northwestern corner of the
Valdelsa Basin, a 25 ◊ 60 km, NW-SE elongated extensional basin filled with some 1000 m of
Upper Neogene and Quaternary deposits (Benvenuti et al.,
2014). The sedimentary succession exposed at
this site has been referred by Dominici et al.
(2018) to the S6 Synthem recognised in the
Valdelsa Basin by Benvenuti et al. (2014). The S6 Synthem is a large-scale
depositional sequence that consists mostly of fluvial coarse-grained sandstones,
intertidal or coastal lagoon deposits, and shoreface and delta-front sandstones
(Benvenuti et al., 2014). At La Serra
quarry, a 25-m-thick stratigraphic section was measured by Garassino
et al. (2012), who recognised the following
informal units:
"Unit 1" (from the base of the section to 8 m above it). This unit consists of mixed sand and silt, cross-stratified sand, and massive clay. "Unit 1" was interpreted by Garassino et al. (2012) as representing an estuarine or deltaic environment with interspersed marshes and ponds.
"Unit 2" (from 8 to 14 m). This is a thick sand body. According to Garassino et al. (2012), it testifies to a barrier island environment, where both submarine longshore dunes and subaerial aeolian dunes were active.
"Unit 3" (from 14 to 19.5 m). This unit consists of intercalated silt and clay. "Unit 3" was referred by Garassino et al. (2012) to a sheltered lagoon depositional setting.
"Unit 4" (from 19.5 m to the top of the section at ~25 m). This unit consists of highly fossiliferous clayey sand (including an exceptionally rich decapod assemblage, as well as almost ubiquitous molluscs and rarer dental and dermal remains of elasmobranch cartilaginous fishes; Collareta et al., in press). It was interpreted by Garassino et al. (2012) as a shallowing-upward deposit that formed in a low-energy, shallow-marine palaeoenvironment across offshore transition and lower-shoreface settings.
Garassino et al. (2012) reported that fossils of continental vertebrates (i.e., indeterminate remains of cervids and turtles) occur in "Unit 2" ~10 m above the base of their stratigraphic log, in a bed that also features abundant woody remains. The terrapin fossil described herein instead came from a bed of deltaic/estuarine sands of "Unit 1" ~7 m above the base of the section measured by Garassino et al. (2012).
On the basis of sparse data on the ostracod and calcareous nannoplankton content of some samples, Garassino et al. (2012) regarded the sediments of La Serra quarry as not younger than the Early Pliocene (i.e., Zanclean) in age. More recently, building upon basin-wide correlations, biochronological considerations, and biostratigraphic analyses on several outcrops from throughout the Valdelsa successions, Benvenuti et al. (2014) and Dominici et al. (2018) referred the S6 Synthem to the upper Piacenzian-lower Gelasian ñ an interpretation supported by the Piacenzian age of the underlying S5 Synthem, as well as by the detection of the the Gauss-Matuyama reversal at Montopoli, some 6 km north of La Serra (Lindsay et al., 1980). In particular, Dominici et al. (2018) assigned a latest Piacenzian age to the invertebrate accumulation observed in the "Unit 4" deposits exposed at La Serra. Considering that the latter is stratigraphically higher than "Unit 2", a late Piacenzian age can be inferred for the terrapin fossil described herein.
From a palaeontological point of view, besides a rich record of marine vertebrates (e.g., Capellini, 1905; Bianucci, 1996; Danise & Dominici, 2014; Collareta et al., 2016, 2017, 2020; Dominici et al., 2018; Collareta, 2020), remains of continental vertebrates have been abundantly documented from the Pliocene deposits of the Valdelsa Basin (Azzaroli, 1992; Benvenuti & Dominici, 1992; Dominici, 1994; Benvenuti et al., 1995a, 1995b, 2007; Dominici et al., 1995; Rustioni & Mazza, 2001; Abbazzi et al., 2008). These fossils have mostly been referred to the Triversa and Montopoli Faunal Units, which correspond to the Upper Pliocene-lowermost Pleistocene Mammal Neogene (=MN) zones 16a and 16b (e.g., Esu & Ciangherotti, 2004).
|
Figure 1:
Location of La Serra, where the fossil terrapin
described herein was found (white star), and schematic regional geological map.
B-CI = Baccinello-Cinigiano Basin; CA = Casino Basin; RD = Radicofani Basin; SI
= Siena Basin; VC = Val di Chiana Basin; VE = Valdelsa Basin; VO-RA-CH =
Volterra-Radicondoli-Chiusdino basins. After Collareta et al.
(in press). |
TESTUDINES Batsch, 1788
CRYPTODIRA Cope, 1868
TESTUDINOIDEA Gray, 1825
GEOEMYDIDAE Theobald, 1868
Mauremys Gray, 1869
Type species. Emys fuliginosus Gray, 1860
Mauremys sp.
Referred material and repository. One loose neural plate and one partial plastron, thought to originate from the same individual, currently kept at Badia a Settimo (Scandicci, Italy) in the permanent exhibition of "Gruppo AVIS Mineralogia e Paleontologia Scandicci" (=GAMPS) under accession number GAMPS-00925.
Occurrence. Upper Pliocene (upper Piacenzian) dune sand exposed at La Serra, Pisa Province, Tuscany, central Italy.
Descriptive remarks. Among
several minute carapace bone fragments, a single element was
determined anatomically thanks to its significant completeness degree. This
element (Fig. 2 ![]() )
consists of a bony plate that lacks its left anterolateral
corner. It is hexagonal in outline and displays a short anterolateral margin. It
is significantly wider than long (estimated maximum transverse width: 25 mm;
maximum anteroposterior length: 18 mm), with a slightly concave anterior margin
that is slightly shorter than the slightly convex posterior margin. The dorsal surface of
the plate is gently convex upwards; on its anterior half, it shows traces of a
sagittal keel, whereas a near-straight epidermal sulcus, mostly lined by
indurated sediment, extends transversely at about one third of the
anteroposterior length of the plate (measured from its posterior margin).
Viscerally, remains of the attachments of the thoracic vertebrae are clearly
observable. The plate exhibits its minimum dorsoventral thickness (about 1.5 mm)
at its periphery, whereas it significantly thickens sagittally. Based on its
general morphology, as well as on the observation of remains of the attachments
of the thoracic vertebrae, this carapace element is interpreted as a neural
plate. This disjunct neural is attributed to Mauremys on the basis of the
co-occurrence of the following characters: plate hexagonal in outline, wider
than long; anterolateral margins short; anterior and posterior margins
anteriorly concave, displaying different lengths; and medial keel and sagittal
thickening present (Ernst & Barbour,
1989; Hervet,
2000; Li
et al., 2015; Georgalis
et al., 2019). In light of
the shape and proportions of this bone, which are strongly reminiscent of a
mid-position neural plate, the epidermal sulcus that ornaments the dorsal
surface might be interpreted as dividing the third vertebral scute (V3) from the
fourth one (V4). In the extant species Mauremys leprosa, V3-V4 is
observed on the fifth neural (Ne5), as is also the case for the extinct species Mauremys
massiliensis, Mauremys thaninensis, Mauremys campanii, and Mauremys
gaudryi, among others (Hervet, 2000,
2003; Claude
et al., 2007; Chesi, 2009; Chesi
et al., 2009; Vlachos
et al., 2019). Given these
observations, the neural plate from La Serra is here interpreted as Ne5.
)
consists of a bony plate that lacks its left anterolateral
corner. It is hexagonal in outline and displays a short anterolateral margin. It
is significantly wider than long (estimated maximum transverse width: 25 mm;
maximum anteroposterior length: 18 mm), with a slightly concave anterior margin
that is slightly shorter than the slightly convex posterior margin. The dorsal surface of
the plate is gently convex upwards; on its anterior half, it shows traces of a
sagittal keel, whereas a near-straight epidermal sulcus, mostly lined by
indurated sediment, extends transversely at about one third of the
anteroposterior length of the plate (measured from its posterior margin).
Viscerally, remains of the attachments of the thoracic vertebrae are clearly
observable. The plate exhibits its minimum dorsoventral thickness (about 1.5 mm)
at its periphery, whereas it significantly thickens sagittally. Based on its
general morphology, as well as on the observation of remains of the attachments
of the thoracic vertebrae, this carapace element is interpreted as a neural
plate. This disjunct neural is attributed to Mauremys on the basis of the
co-occurrence of the following characters: plate hexagonal in outline, wider
than long; anterolateral margins short; anterior and posterior margins
anteriorly concave, displaying different lengths; and medial keel and sagittal
thickening present (Ernst & Barbour,
1989; Hervet,
2000; Li
et al., 2015; Georgalis
et al., 2019). In light of
the shape and proportions of this bone, which are strongly reminiscent of a
mid-position neural plate, the epidermal sulcus that ornaments the dorsal
surface might be interpreted as dividing the third vertebral scute (V3) from the
fourth one (V4). In the extant species Mauremys leprosa, V3-V4 is
observed on the fifth neural (Ne5), as is also the case for the extinct species Mauremys
massiliensis, Mauremys thaninensis, Mauremys campanii, and Mauremys
gaudryi, among others (Hervet, 2000,
2003; Claude
et al., 2007; Chesi, 2009; Chesi
et al., 2009; Vlachos
et al., 2019). Given these
observations, the neural plate from La Serra is here interpreted as Ne5.
|
Figure
2:
GAMPS-00925, Mauremys sp. from the Upper Pliocene of La
Serra (Tuscany, central Italy), posterior neural plate in A) dorsal (external)
and B) ventral (visceral) views. |
Some plastral bones were found in
association with the aforementioned neural plate (Fig. 3 ![]() ).
These include the slightly damaged left and right hypoplastra, as well as the
substantially complete left and right xiphiplastra. Although disarticulated,
these four bones can be easily recombined along their mutual sutures to
reconstruct the original aspect of the middle and posterior portions of the
plastron, whose ventral surface was almost perfectly flat. The anterior margin
of the hypoplastra is only partially preserved, but it seems to have been more
or less straight and transversely oriented; furthermore, its morphology
demonstrates that a hyo-hypoplastral hinge was not present. The right
hypoplastron is 48.5 mm long (measurement taken along the medial margin of the
plate) and has a maximum transverse width of 42 mm posterior to the partly
preserved inguinal process. The inguinal process occurs at about mid-length of
the hypoplastron. Posterior to this level, the posterolateral margin of the
hypoplastron is weakly bowed laterally. In ventral view, the abdominal-femoral
sulcus crosses the hypoplastra horizontally in its median tract, whereas its
distal portions are anterolaterally directed. The abdominal-femoral sulcus
terminates just posterior to the inguinal process, where a strongly reduced,
barely discernible inguinal notch occurs. The anterior margin of the
xiphiplastra is gently bowed anteriorly and, in dorsal view only, it displays an
anteriorly convex tract close to the anterolateral corner of the bone. The right
xiphiplastron exhibits a medial length of 32.5 mm and a transverse width of 40.5
mm. In ventral view, the lateral margins of the xiphiplastra distinctly converge
backwards; they exhibit obvious, broad, fan-shaped notches in occurrence of the
distal terminations of the femoral-anal sulcus. The latter consists of two
near-straight, anteromedially projecting segments that meet each other
sagittally ñ i.e., on each xiphiplastron, the femoral-anal sulcus runs nearly perpendicular to the
posterolateral margin of the bone. On the right xiphiplastron, the femoral-anal
sulcus meets the medial margin of the plate 18 mm anterior to the apex of the
anal notch. The latter is formed by the converging posterior margins of the
xiphiplastra. The anal notch is narrow and deep, its apex being located 11 mm
anterior to the level of the caudal terminations of the posterolateral processes
of the constituent plates, which are rather sharply pointed backwards. Facing
each other, the posterior margins of the xiphiplastra are slightly concave
posteromedially and meet at an angle of about 90∞. On the whole, the posterior
lobe of the plastron is long and displays a distinctly rounded outline. The
visceral view of the posterior lobe shows that the femoral and anal scutes
covered the dorsomarginal portions of both the hypoplastra and xiphiplastra, as
reflected by the presence of a ridged elevation that runs close to the outer margins
of these plates. This elevation is posterior to the inguinal processes and extends
all around the hypoplastra and xiphiplastra, to disappear only near the apex of
the anal notch. The studied plastron belongs to the genus Mauremys on the
basis of the following combination of characters: absence of a hyo-hypoplastral
hinge; inguinal notch strongly reduced; posterior lobe long and well-rounded;
femoral scutes overlapping the dorsomarginal portions of the hypoplastra;
femoral-anal sulcus running nearly perpendicular to the lateral edge of the
plate; lateral margins of the xiphiplastra distinctly converging posteromedially;
anal scutes relatively short; and anal notch narrow and deep (Ernst &
Barbour, 1989; Hervet, 2003,
2004; Claude
et al., 2007; Chesi, 2009; Chesi
et al., 2009; GÛmez de Soler et al.,
2012; Li et
al., 2015; Vlachos et al.,
2015, 2019; Vlachos
& Delfino, 2016).
).
These include the slightly damaged left and right hypoplastra, as well as the
substantially complete left and right xiphiplastra. Although disarticulated,
these four bones can be easily recombined along their mutual sutures to
reconstruct the original aspect of the middle and posterior portions of the
plastron, whose ventral surface was almost perfectly flat. The anterior margin
of the hypoplastra is only partially preserved, but it seems to have been more
or less straight and transversely oriented; furthermore, its morphology
demonstrates that a hyo-hypoplastral hinge was not present. The right
hypoplastron is 48.5 mm long (measurement taken along the medial margin of the
plate) and has a maximum transverse width of 42 mm posterior to the partly
preserved inguinal process. The inguinal process occurs at about mid-length of
the hypoplastron. Posterior to this level, the posterolateral margin of the
hypoplastron is weakly bowed laterally. In ventral view, the abdominal-femoral
sulcus crosses the hypoplastra horizontally in its median tract, whereas its
distal portions are anterolaterally directed. The abdominal-femoral sulcus
terminates just posterior to the inguinal process, where a strongly reduced,
barely discernible inguinal notch occurs. The anterior margin of the
xiphiplastra is gently bowed anteriorly and, in dorsal view only, it displays an
anteriorly convex tract close to the anterolateral corner of the bone. The right
xiphiplastron exhibits a medial length of 32.5 mm and a transverse width of 40.5
mm. In ventral view, the lateral margins of the xiphiplastra distinctly converge
backwards; they exhibit obvious, broad, fan-shaped notches in occurrence of the
distal terminations of the femoral-anal sulcus. The latter consists of two
near-straight, anteromedially projecting segments that meet each other
sagittally ñ i.e., on each xiphiplastron, the femoral-anal sulcus runs nearly perpendicular to the
posterolateral margin of the bone. On the right xiphiplastron, the femoral-anal
sulcus meets the medial margin of the plate 18 mm anterior to the apex of the
anal notch. The latter is formed by the converging posterior margins of the
xiphiplastra. The anal notch is narrow and deep, its apex being located 11 mm
anterior to the level of the caudal terminations of the posterolateral processes
of the constituent plates, which are rather sharply pointed backwards. Facing
each other, the posterior margins of the xiphiplastra are slightly concave
posteromedially and meet at an angle of about 90∞. On the whole, the posterior
lobe of the plastron is long and displays a distinctly rounded outline. The
visceral view of the posterior lobe shows that the femoral and anal scutes
covered the dorsomarginal portions of both the hypoplastra and xiphiplastra, as
reflected by the presence of a ridged elevation that runs close to the outer margins
of these plates. This elevation is posterior to the inguinal processes and extends
all around the hypoplastra and xiphiplastra, to disappear only near the apex of
the anal notch. The studied plastron belongs to the genus Mauremys on the
basis of the following combination of characters: absence of a hyo-hypoplastral
hinge; inguinal notch strongly reduced; posterior lobe long and well-rounded;
femoral scutes overlapping the dorsomarginal portions of the hypoplastra;
femoral-anal sulcus running nearly perpendicular to the lateral edge of the
plate; lateral margins of the xiphiplastra distinctly converging posteromedially;
anal scutes relatively short; and anal notch narrow and deep (Ernst &
Barbour, 1989; Hervet, 2003,
2004; Claude
et al., 2007; Chesi, 2009; Chesi
et al., 2009; GÛmez de Soler et al.,
2012; Li et
al., 2015; Vlachos et al.,
2015, 2019; Vlachos
& Delfino, 2016).
|
Figure
3:
GAMPS-00925, Mauremys sp. from the Upper Pliocene
of La Serra (Tuscany, central Italy), left and right hypoplastra and
xiphiplastra in A) ventral (external) and B) dorsal (visceral) views. The
arrowheads superimposed on panel A indicate the location of two Thatchtelithichnus
holmani traces detailed in Figure 4 |
Based on comparisons with extant specimens of Mauremys kept at the Museo di Storia Naturale dell'Universitý di Pisa, and considering the dimensions of the preserved plastral bones, a total carapace length of about 175 mm can be reconstructed for GAMPS-00925. Such a size is well within the range of values reported by Ernst and Barbour (1989) for the living species of Mauremys, as well as close to the typical carapace length of the adults of the extant Mediterranean species M. leprosa (160-200 mm according to Hervet, 2000). Attribution of GAMPS-00925 at species level is precluded by the fragmentary nature of the studied material; nevertheless, this specimen can be distinguished from the extinct Italian species Mauremys campanii, M. portisi, and M. etrusca. In particular, it differs: i) from the Late Miocene M. campanii in the differing geometry of the abdominal-femoral sulcus, as well as by the more posterior position of the latter; ii) from the Early Pliocene M. portisi by the even more reduced inguinal notch; and iii) from the Early Pleistocene M. etrusca by the differing geometry of the abdominal-femoral sulcus, as well as by the more anterior position of the latter. In turn, the following features of the plastron establish the similarity of GAMPS-00925 to the extant M. leprosa: posterior lobe starting at the level of the distal terminations of the abdominal-femoral sulcus, at about mid-length of the hypoplastra; anal notch angular rather than rounded, depicting an angle of about 90∞; and posterolateral processes of the xiphiplastra posteriorly pointed. The fossil record of M. leprosa begin in the Lower Pliocene, the oldest remains of this species being known from the Algerian locality of AÔn Boucherit [Lapparent de Broin, 2000; but see Chesi et al. (2009) for a revised chronostratigraphical attribution of these finds].
Ichnological
observations. During examination of GAMPS-00925, two
unusual scars were observed on the ventral surface of the left hypoplastron and
xiphiplastron (Fig. 3.A ![]() ). In the following section, these scars are briefly
characterised.
). In the following section, these scars are briefly
characterised.
Thatchtelithichnus Zonneveld et al., 2015
Type ichnospecies. Thatchtelithichnus holmani Zonneveld et al., 2015
Thatchtelithichnus holmani Zonneveld et al., 2015
Descriptive remarks. The
ichnogenus Thatchtelithichnus was diagnosed by Zonneveld et al.
(2015) as follows: "Circular to subcircular
grooves with a central boss or platform, bored into a bone substrate. Grooves
penetrate into the cortical layer of the bone. The central platform may consist
of an unmodified platform or a rounded nubbin. Boring walls rounded, convex or
sharp and perpendicular to the external substrate surface". The diagnosis of Thatchtelithichnus
holmani, the sole ichnospecies included therein, was provided by the same
authors in these terms: "As for the ichnogenus. Non-penetrative borings into a
bone substrate, having a ring-like profile, and a central raised boss or
pedestal". These descriptions conform well to two scars observed on the
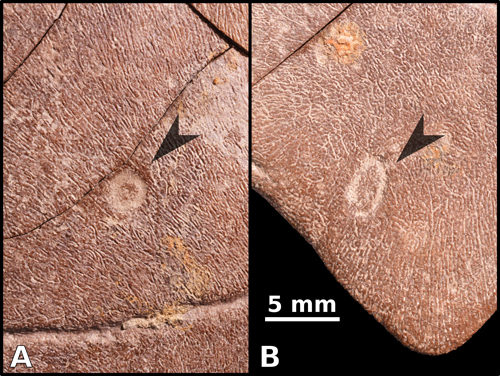
plastral scales of GAMPS-00925. The first
(Fig. 4.A ![]() ) is found on the ventral
surface of the left hypoplastron on the area that was originally covered by the
abdominal scute. It consists of a very shallow, circular depression, measuring
~3 mm in diameter, hosting at its centre a raised, rounded nubbin having a
diameter of ~1.5 mm. This trace is very similar to the holotype of T.
holmani as figured by Zonneveld et al. (2015: Fig.
9.1) and Zonneveld and Bartels
(2020: Fig. 1D-E,
but see also their Fig. 1F). Similar trace fossils, attributed by Zonneveld and Bartels
(2020) to T. holmani,
have also been reported from dinosaur long bones and ribs from the Jurassic of
Wyoming (i.e., the "rosettes" of Bader et al.,
2009), where they have been interpreted as pupation
chambers of dermestid beetles. A similarly shallow second scar
(Fig. 4.B
) is found on the ventral
surface of the left hypoplastron on the area that was originally covered by the
abdominal scute. It consists of a very shallow, circular depression, measuring
~3 mm in diameter, hosting at its centre a raised, rounded nubbin having a
diameter of ~1.5 mm. This trace is very similar to the holotype of T.
holmani as figured by Zonneveld et al. (2015: Fig.
9.1) and Zonneveld and Bartels
(2020: Fig. 1D-E,
but see also their Fig. 1F). Similar trace fossils, attributed by Zonneveld and Bartels
(2020) to T. holmani,
have also been reported from dinosaur long bones and ribs from the Jurassic of
Wyoming (i.e., the "rosettes" of Bader et al.,
2009), where they have been interpreted as pupation
chambers of dermestid beetles. A similarly shallow second scar
(Fig. 4.B ![]() )
is observed on the ventral surface of the left xiphiplastron on the
area that was originally covered by the anal scute. It consists of a rather
elongate depression, measuring ~4 ◊ 2.5 mm, with a central unmodified
platform that extends for a distance of less than 1 mm from the outer
edge of the trace. This scar is somewhat reminiscent of a large, roughly
elliptical specimen of T. holmani figured by Zonneveld
et al. (2015: Fig. 9.1).
)
is observed on the ventral surface of the left xiphiplastron on the
area that was originally covered by the anal scute. It consists of a rather
elongate depression, measuring ~4 ◊ 2.5 mm, with a central unmodified
platform that extends for a distance of less than 1 mm from the outer
edge of the trace. This scar is somewhat reminiscent of a large, roughly
elliptical specimen of T. holmani figured by Zonneveld
et al. (2015: Fig. 9.1).
|
Figure
4:
Detail of two Thatchtelithichnus holmani traces
found on the plastron-bottom of GAMPS-00925, indicated by superimposed arrowheads.
A) Circular groove found on the left hypoplastron. B) Elliptical groove found on
the left xiphiplastron. The scale bar applies to both panels. |
Scrutiny of the palaeoherpetological literature reveals
that fossils of Mauremys are known from several Upper Miocene to Upper Pleistocene sites in Italy
(Fig. 5 ![]() ;
Table 1). Most of these occurrences come
from Miocene or Pleistocene contexts; in particular, the Miocene segment of the
Italian record of Mauremys includes both Tortonian and Messinian
occurrences, whereas the Pleistocene fossils include Lower, Middle, and Upper Pleistocene instances. Prior to the present note, a single confirmed occurrence
of Mauremys was known from the Pliocene of Italy, consisting of the
holotype and only known specimen of Mauremys portisi from Valleandona (Asti Province, Piedmont; Sacco,
1889).
The stratigraphic occurrence of the M. portisi holotype is somewhat
debated; long regarded as "Astian" in age (e.g., Hervet,
2004; Chesi
et al., 2007), it was recently referred
to the zone MN 13 or 14, i.e., Lower Pliocene (Zanclean), by Chesi et al.
(2009).
A second, putative record of Mauremys from the Pliocene of Italy,
consisting of a partial carapace and associated plastron from Villafranca D'Asti
(Asti Province, Piedmont), was reported by Ambrosetti et al.
(1996)
as belonging to Mauremys sp. This specimen, referred to the Upper Pliocene zone MN 16a, was later reattributed to cf. Geoemyda sp. by Chesi
(2009). Therefore, the Late Pliocene terrapin
specimen from La Serra represents the second confirmed occurrence of Mauremys
in the Italian Pliocene, as well as the first authentic record of this genus
from the Piacenzian of Italy. As such, it is an important specimen that fills a
gap in the Italian chronostratigraphic distribution of fossil Mauremys,
and helps ñ together with the Lower Pliocene holotype of M. portisi ñ
bridging the rich Miocene and Pleistocene segments of this record. In light of
the above data, it is reasonable to hypothesise that terrapins of the genus Mauremys
continuously inhabited the Italian peninsula from the Late Miocene to the Middle
Pleistocene at least (the sole Late Pleistocene Italian record of Mauremys
comes from Sardinia; Chesi et al., 2007).
From a palaeoenvironmental point of view, the occurrence of Mauremys in
the Piacenzian deposits of La Serra does not imply by itself the nearby presence
of permanent freshwater ecosystems. Indeed, the extant Mediterranean species of Mauremys
can respond to prolonged conditions of drought by aestivating (Highfield,
2002) and they can even survive in brackish
waters, including coastal lagoons (Sidis & Gasith,
1985; Ernst &
Barbour, 1989; Arnold & Ovenden,
2002;
Vamberger et al., 2014). The
sediments that host the terrapin fossil described herein also feature mesohaline
and euryhaline taxa of foraminifers, ostracods, and gastropods, which suggest
a brackish lagoonal system, connected with a river mouth and sheltered seawards
by a chain of barrier islands (Garassino et al.,
2012).
Such a palaeoenvironmental reconstruction is consistent with the habitat
preferences of extant Mauremys spp., which are however particularly broad
(Ernst &
Barbour, 1989; Highfield, 2002).
;
Table 1). Most of these occurrences come
from Miocene or Pleistocene contexts; in particular, the Miocene segment of the
Italian record of Mauremys includes both Tortonian and Messinian
occurrences, whereas the Pleistocene fossils include Lower, Middle, and Upper Pleistocene instances. Prior to the present note, a single confirmed occurrence
of Mauremys was known from the Pliocene of Italy, consisting of the
holotype and only known specimen of Mauremys portisi from Valleandona (Asti Province, Piedmont; Sacco,
1889).
The stratigraphic occurrence of the M. portisi holotype is somewhat
debated; long regarded as "Astian" in age (e.g., Hervet,
2004; Chesi
et al., 2007), it was recently referred
to the zone MN 13 or 14, i.e., Lower Pliocene (Zanclean), by Chesi et al.
(2009).
A second, putative record of Mauremys from the Pliocene of Italy,
consisting of a partial carapace and associated plastron from Villafranca D'Asti
(Asti Province, Piedmont), was reported by Ambrosetti et al.
(1996)
as belonging to Mauremys sp. This specimen, referred to the Upper Pliocene zone MN 16a, was later reattributed to cf. Geoemyda sp. by Chesi
(2009). Therefore, the Late Pliocene terrapin
specimen from La Serra represents the second confirmed occurrence of Mauremys
in the Italian Pliocene, as well as the first authentic record of this genus
from the Piacenzian of Italy. As such, it is an important specimen that fills a
gap in the Italian chronostratigraphic distribution of fossil Mauremys,
and helps ñ together with the Lower Pliocene holotype of M. portisi ñ
bridging the rich Miocene and Pleistocene segments of this record. In light of
the above data, it is reasonable to hypothesise that terrapins of the genus Mauremys
continuously inhabited the Italian peninsula from the Late Miocene to the Middle
Pleistocene at least (the sole Late Pleistocene Italian record of Mauremys
comes from Sardinia; Chesi et al., 2007).
From a palaeoenvironmental point of view, the occurrence of Mauremys in
the Piacenzian deposits of La Serra does not imply by itself the nearby presence
of permanent freshwater ecosystems. Indeed, the extant Mediterranean species of Mauremys
can respond to prolonged conditions of drought by aestivating (Highfield,
2002) and they can even survive in brackish
waters, including coastal lagoons (Sidis & Gasith,
1985; Ernst &
Barbour, 1989; Arnold & Ovenden,
2002;
Vamberger et al., 2014). The
sediments that host the terrapin fossil described herein also feature mesohaline
and euryhaline taxa of foraminifers, ostracods, and gastropods, which suggest
a brackish lagoonal system, connected with a river mouth and sheltered seawards
by a chain of barrier islands (Garassino et al.,
2012).
Such a palaeoenvironmental reconstruction is consistent with the habitat
preferences of extant Mauremys spp., which are however particularly broad
(Ernst &
Barbour, 1989; Highfield, 2002).
|
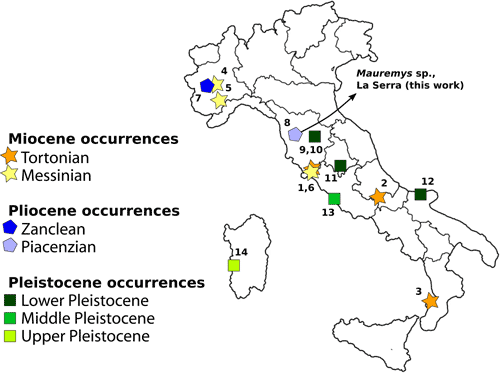
Figure 5:
Geographic and stratigraphic distribution of the Italian
fossil occurrences of Mauremys. Numbers associated with the records refer
to Table 1. |
Table 1: Geographic and stratigraphic distribution of the Italian
fossil occurrences of Mauremys. Numbers in the first column refer to Figure
5 ![]() .
.
| # | Taxon | Locality(ies) | Geological age | Updated literature reference |
| 1 | Mauremys campanii | Casteani, Montebamboli | Tortonian | Chesi et al., 2009 |
| 2 | Mauremys sp. | Scontrone | Tortonian | Patacca et al., 2013 |
| 3 | Mauremys sp. | Cessaniti | Tortonian | Georgalis et al., 2020 |
| 4 | Mauremys sp. | Moncucco Torinese | Messinian | Colombero et al., 2017 |
| 5 | cf. Mauremys sp. | S. Vittoria d'Alba | Messinian | Kotsakis & Mori, 1981 |
| 6 | Mauremys campanii | Pian Calcinaio | Messinian | Chesi et al., 2009 |
| 7 | Mauremys portisi | Valleandona | Zanclean | Chesi et al., 2009 |
| 8 | Mauremys sp. | La Serra | Piacenzian | This work |
| 9 | Mauremys etrusca | Restoni | Early Pleistocene | Kotsakis, 1980 |
| 10 | Mauremys etrusca | Colombaiolo | Early Pleistocene | Kotsakis, 1980 |
| 11 | Mauremys sp. | Torre Picchio | Early Pleistocene | Girotti et al., 2003 |
| 12 | Mauremys sp. | Cava Pirro, Cava Dell'Erba | Early Pleistocene | Delfino & Atzori, 2013 |
| 13 | Mauremys sp. | Grotta del Monte delle Gioie | Middle Pleistocene | Kotsakis & Barisone, 2003 |
| 14 | Mauremys sp. | San Giovanni di Sinis | Late Pleistocene | Chesi et al., 2007 |
The two specimens
of Thatchtelithichnus
holmani reported herein represent some of the few records worldwide of
this rarely reported ichnotaxon, as well as its geologically youngest published
occurrence. The present record stimulates some inferences regarding the Thatchtelithichnus
tracemaker(s). As mentioned above, Zonneveld et al.
(2015)
proposed that these borings were emplacement scars left by invertebrate
ectoparasites (or mesoparasites) while the host turtles were living. Wisshak
et al. (2019)
included Thatchtelithichnus in a list of
bioclaustration structures, which are not regarded as trace fossils. However, a
reanalysis of the type series of T. holmani performed by Zonneveld and Bartels
(2020) revealed that no part of this ichnotaxon involves bioclaustration; these
authors concluded that the genetic hypothesis formulated by Zonneveld
et al. (2015)
for explaining the Thatchtelithichnus scars remains the most likely one.
With respect to the ring-shaped traces observed by
Bader et al. (2009)
on dinosaur bones from the Jurassic of Wyoming and later attributed to T.
holmani by Zonneveld and Bartels
(2020),
an identification as incipient stages in the process of excavation of pupation
chambers by dermestid beetles has been proposed. Modern dermestid larvae (and
adults as well) feed on the desiccated flesh of carcasses (Hinton,
1945;
Timm, 1982)
and might even consume bone when other food sources are not available (Hefti
et al., 1980).
After some four weeks of feeding, the larvae pupate and create chambers in any
available compact surface, including desiccated flesh and bone (Hinton,
1945;
Timm, 1982).
When severe infestation occurs, bones might become figuratively honeycombed with
pupal chambers, their burrows sometimes extending towards the medullary cavity (Huchet
et al., 2013,
and references therein). Crucially, however, dermestids are not known to feed on
keratin, which constitutes the scutes that coat the plastral bones of turtles.
Among extant insects, only some species of tineid moths are specialised in the
consumption of the keratinous portions of carcasses, including the turtle shell
(Deyrup et al., 2005).
Members of the tineid genus Ceratophaga have been reported excavating
straight-sided, cylindrical pupation chambers into the astragali of African
bovids (Hill, 1987);
however, the morphology of tineid moth damage does not match that of Thatchtelithichnus.
Given these considerations, it is our contention that the Thatchtelithichnus
traces observed on the exterior of the plastral bones of turtles cannot be
interpreted as feeding marks or pupation chambers of insects (whether dermestid
beetles, tineid moths, or members of other hexapod lineages). For explaining
these scars, we concur with Zonneveld
et al. (2015)
and Zonneveld and Bartels (2020)
in inferring an origin as attachment traces of aquatic ectoparasites (one of
ticks, leeches, or flukes); this interpretation might also better explain
the variation in circularity of the Thatchtelithichnus traces found on
turtles (e.g., Fig. 4 ![]() and Zonneveld et al., 2015: Fig. 9.1). On the other hand, it should be noted that severe (yet
non-destructive) episodes of septicaemic cutaneous ulcerative disease (=SCUD, or
"shell rot"; see e.g., Granados et al., 2013)
that penetrate into the plastral scutes to skim the bone surface might also
locally produce shallow lesions on the plastral plates of extant freshwater
turtles (personal observation by M.A.L.Z.). Recognising that the Thatchtelithichnus
grooves represent the lesions of a widespread cutaneous disease such as SCUD
might also lead to the conclusion that neither Thatchtelithichnus nor
T. holmani should be regarded as valid ichnotaxa (Bertling
et al., 2006). However, the constancy of the ring-like aspect of the Thatchtelithichnus
scars and their similar occurrence on the plastron-bottom only in different
turtle taxa strongly evoke an origin as attachment traces of homotypic organisms
rather than as byproducts of extensive, shell-degrading bacterial infections [see
also Zonneveld et al. (2015)].
In conclusion, the ectoparasitic hypothesis currently seems the most
parsimonious for explaining the observation of Thatchtelithichnus-like structures on the exterior of the plastral bones of fossil
chelonians. Nevertheless, additional actualistic observations on living aquatic
turtles are clearly needed for confirming or disproving this tantalising
conjecture.
and Zonneveld et al., 2015: Fig. 9.1). On the other hand, it should be noted that severe (yet
non-destructive) episodes of septicaemic cutaneous ulcerative disease (=SCUD, or
"shell rot"; see e.g., Granados et al., 2013)
that penetrate into the plastral scutes to skim the bone surface might also
locally produce shallow lesions on the plastral plates of extant freshwater
turtles (personal observation by M.A.L.Z.). Recognising that the Thatchtelithichnus
grooves represent the lesions of a widespread cutaneous disease such as SCUD
might also lead to the conclusion that neither Thatchtelithichnus nor
T. holmani should be regarded as valid ichnotaxa (Bertling
et al., 2006). However, the constancy of the ring-like aspect of the Thatchtelithichnus
scars and their similar occurrence on the plastron-bottom only in different
turtle taxa strongly evoke an origin as attachment traces of homotypic organisms
rather than as byproducts of extensive, shell-degrading bacterial infections [see
also Zonneveld et al. (2015)].
In conclusion, the ectoparasitic hypothesis currently seems the most
parsimonious for explaining the observation of Thatchtelithichnus-like structures on the exterior of the plastral bones of fossil
chelonians. Nevertheless, additional actualistic observations on living aquatic
turtles are clearly needed for confirming or disproving this tantalising
conjecture.
Our gratitude to Simone Farina (Museo di Storia Naturale, Universitý di Pisa) for providing access to the specimens of extant Mauremys under his care. Constructive comments by Stephen J. Godfrey (Calvert Marine Museum) and Evangelos Vlachos (CONICET & Museo PaleontolÛgico Egidio Feruglio) greatly contributed to shape this work into its present form ñ thank you very much for your support! Not least, thanks are due to Bruno Granier (UniversitÈ de Bretagne Occidentale) and to Stephen Carey (Federation University Australia) for their valuable and thorough editorial support.
Abbazzi L., Benvenuti M., Ceci M.E., Esu D., Faranda C., Rook L. & Tangocci F. (2008).- The end of the Lago-Mare time in the SE Valdelsa Basin (central Italy): interference between local tectonism and regional sea-level rise.- Geodiversitas, Paris, vol. 30, p. 611-639.
Ambrosetti P., Azzaroli A., Ficcarelli G., Masini F., Sala B. & Torre D. (1996).- Vertebrati. Il Quaternario, Roma, vol. 9, p. 70-73.
Arnold N.E. & Ovenden D.W. (2002).- A field guide to the reptiles and amphibians of Britain and Europe.- Collins, London, 288 p.
Azzaroli A. (1992).- The cervid genus Pseudodama n.g. in the Villafranchian of Tuscany.- Palaeontographia Italica, Pisa, vol. 79, p. 1-41.
Bader K.S., Hasiotis S.T. & Martin L.D. (2009).- Application of forensic science techniques to trace fossils on dinosaur bones from a quarry in the Upper Jurassic Morrison Formation, northeastern Wyoming.- Palaios, Broken Arrow, vol. 24, p. 140-158.
Barth D., Bernhard D., Fritzsch G. & Fritz U. (2004).- The freshwater turtle genus Mauremys (Testudines, Geoemydidae)- a textbook example of an eastñwest disjunction or a taxonomic misconcept?.- Zoologica Scripta, London, vol. 33, p.213-221.
Batsch A.J.G.C (1788).- Versuch einer Anleitung, zur Kenntniþ und Geschichte der Thiere und Mineralien, f¸r akademische Vorlesungen entworfen, und mit den nˆthigsten Abbildungen versehen. Erster Theil. Allgemeine Geschichte der Natur; besondre der S‰ugthiere, Vˆgel, Amphibien und Fische.- Akademische Buchhandlung, Jena, 528 p.
Benvenuti M. & Dominici S. (1992).- Facies analysis, paleoecology and sequence stratigraphy in a Pliocene siliciclastic succession, San Miniato (Pisa, Italy).- Bollettino della Societý Paleontologica Italiana, Modena, vol. 31, p. 241-259.
Benvenuti M., Bertini A., Conti C., Dominici S. & Falcone D. (1995a).- Analisi stratigrafica e paleoambientale integrata del Pliocene dei dintorni di San Miniato.- Quaderni del Museo di Storia naturale di Livorno, Leghorn, vol. 14, p. 29-49.
Benvenuti M., Dominici S. & Rook L. (1995b).- Inquadramento stratigrafico-deposizionale delle faune a mammiferi villafranchiane (Unitý faunistiche Triversa e Montopoli) del Valdarno inferiore nella zona a sud dell'Arno (Toscana).- Il Quaternario, Roma, vol. 8, p. 457-464.
Benvenuti M., Bertini A., Conti C., & Dominici S. (2007).- Integrated analyses of litho-and biofacies in a Pliocene cyclothemic, alluvial to shallow marine succession (Tuscany, Italy).- Geobios, Villeurbanne, vol. 40, p. 143-158.
Benvenuti M., Del Conte S., Scarselli N. & Dominici S. (2014).- Hinterland basin development and infilling through tectonic and eustatic processes: latest Messinian-Gelasian Valdelsa Basin, Northern Apennines, Italy.- Basin Research, vol. 26, p. 387-402.
Bertling M., Braddy R., Bromley R.G., Demathieu G., Genise J., Mikul·ö R., Nielsen J., Nielsen K., Rindsberg A., Schlirf M. & Uchman A. (2006).- Names for trace fossils: a uniform approach.- Lethaia, Stockholm, vol. 39, p. 265-286.
Bianucci G. (1996).- A new record of baleen whale from the Pliocene of Tuscany (Italy).- Atti della Societý Toscana di Scienze Naturali, Memorie (Serie A), Pisa, vol. 102, p. 101-104.
Capellini G. (1905).- Balene fossili toscane. III Idiocetus Guicciardini.- Memorie della Reale Accademia delle Scienze dell'Istituto di Bologna, vol. 6, p. 71-80.
Chesi F. (2009).- Il registro fossile italiano dei cheloni.- Ph.D. Thesis, Universitý degli Studi di Firenze, Florence, 178 p.
Chesi F., Delfino, M. Abbazzi L. & Carboni L. (2007).- New fossil vertebrate remains from S. Giovanni di Sinis (Late Pleistocene, Sardinia): the last Mauremys (Reptilia, Testudines) in the central Mediterranean.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Palaeontology and Stratigraphy), Milan, vol. 113, p. 287-297.
Chesi F., Delfino M. & Rook L. (2009).- Late Miocene Mauremys (Testudines, Geoemydidae) from Tuscany (Italy): evidence of terrapin persistence after a mammal turnover.- Journal of Paleontology, Lawrence, vol. 83, p. 379-388.
Claude J., Suteethorn V. & Tong H. (2007).- Turtles from the late Eoceneñearly Oligocene of the Krabi Basin (Thailand).- Bulletin de la SociÈtÈ GÈologique de France, Paris, vol. 178, p. 305-316.
Collareta A. (2020).- Discovery of complemental males in a Pliocene accumulation of Chelonibia testudinaria (Linnaeus, 1758), with some notes on the evolution of androdioecy in turtle barnacles.- Neues Jahrbuch f¸r Geologie und Pal‰ontologie-Abhandlungen, Stuttgart, vol. 297, p. 193-203.
Collareta A., Bosselaers M. & Bianucci G. (2016).- Jumping from turtles to whales: a Pliocene fossil record depicts an ancient dispersal of Chelonibia on mysticetes.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Palaeontology and Stratigraphy), Milan, vol. 122, p. 35-44.
Collareta A., Casati S., Catanzariti R. & Di Cencio A. (2017).- First record of the knifetooth sawfish Anoxypristis (Elasmobranchii: Rhinopristiformes) from the Pliocene of Tuscany (central Italy).- Neues Jahrbuch f¸r Geologie und Pal‰ontologie-Abhandlungen, Stuttgart, vol. 284, p. 289-297.
Collareta A., Merella M., Casati S. & Di Cencio A. (in press).- Did titanic stingrays wander the Pliocene Mediterranean Sea? Some notes on a giant-sized myliobatoid stinger from the Piacenzian of Italy.- Neues Jahrbuch f¸r Geologie und Pal‰ontologie-Abhandlungen, Stuttgart.
Collareta A., Merella M., Mollen F.H., Casati S. & Di Cencio A. (2020).- The extinct catshark Pachyscyllium distans (Probst, 1879) (Elasmobranchii: Carcharhiniformes) in the Pliocene of the Mediterranean Sea.- Neues Jahrbuch f¸r Geologie und Pal‰ontologie-Abhandlungen, Stuttgart, vol. 295, p. 129-139.
Colombero S., Alba D.M., D'Amico C., Delfino M., Esu D., Giuntelli P., Harzhauser M., Mazza P.A.P., Mosca M., Neubauer T.A., Pavia G., Pavia M., Villa A. & Carnevale G. (2017).- Late Messinian mollusks and vertebrates from Moncucco Torinese, north-western Italy. Paleoecological and paleoclimatological implications.- Palaeontologia Electronica, vol. 20, article #20.1.10A.
Cope E.D. (1868).- On the origin of genera.- Proceedings of the Academy of Natural Sciences of Philadelphia, Philadelphia, vol. 1868, p. 242-300.
Danise S. & Dominici S. (2014).- A record of fossil shallow-water whale falls from Italy.- Lethaia, Stockholm, vol. 47, p. 229-243.
Delfino M. & Atzori M. (2013).- An update on the early Pleistocene herpetofauna from Pirro Nord.- Palaeontographica Abteilung A, Pal‰ozoologie-Stratigraphie, Stuttgart, vol. 298, p. 19-29.
Deyrup M., Deyrup N.D., Eisner M. & Eisner T. (2005).- A caterpillar that eats tortoise shells.- American Entomologist, Annapolis, vol. 51, p. 245-248.
Dominici S. (1994).- Regressive-transgressive cycles from the Pliocene of the San Miniato area (Tuscany, Italy): paleoecology and sequence stratigraphy.- Bollettino della Societý Paleontologica Italiana, Modena, vol. 33 (supplement 2), p. 117-126.
Dominici S., Rook L., Benvenuti M. & Abbazzi L. (1995).- Tapir remains in paralic deposits of Pliocene age in lower Valdarno (Tuscany, Italy): Facies analysis and taphonomy.- Geobios, Villeurbanne, vol. 28, p. 131-135.
Dominici S., Danise S. & Benvenuti M. (2018).- Pliocene stratigraphic paleobiology in Tuscany and the fossil record of marine megafauna.- Earth-Science Reviews, Amsterdam, vol. 176, p. 277-310.
Ernst C.H. & Barbour R.W. (1989).- Turtles of the world.- Smithsonian Institution Press, Washington - DC, 328 p.
Esu D., & Ciangherotti A. (2004).- Palaeoecologic and palaeobiogeographic character of Middle Pliocene non-marine mollusc faunas from north-western Italy.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Palaeontology and Stratigraphy), Milan, vol. 110, p. 517-530.
Garassino A., Pasini G., De Angeli A., Charbonnier S., Famiani F., Baldanza A. & Bizzarri R. (2012).- The decapod community from the Early Pliocene (Zanclean) of "La Serra" quarry (San Miniato, Pisa, Toscana, central Italy): sedimentology, systematics, and palaeoenvironmental implications.- Annales de PalÈontologie, Paris, vol. 98, p. 1-61.
Georgalis G.L., Villa A., Martin I., Vasilyan D. & Delfino M. (2019).- Fossil amphibians and reptiles from the Neogene locality of Maramena (Greece), the most diverse European herpetofauna at the Miocene/Pliocene transition boundary.- Palaeontologia Electronica, vol. 23, article #22.3.68, 99 p.
Georgalis G.L., Insacco G., Rook L., Spadola F. & Delfino M. (2020).- Turtle remains from the late Miocene of the Cessaniti area, southern Italy- insights for a probable Tortonian chelonian dispersal from Europe to Africa.- Swiss Journal of Palaeontology, vol. 139, article #1, 13 p.
Girotti O., Capasso Barbato L., Esu D., Gliozzi E., Kotsakis T., Martinetto E., Petronio C., Sardella R. & Squazzini E. (2003).- The section of Torre di Picchio (Terni, Umbria, central Italy): a Villafranchian site rich in vertebrates, molluscs, ostracods and plants.- Rivista Italiana di Paleontologia e Stratigrafia (Research in Palaeontology and Stratigraphy), Milan, vol. 109, p. 77-98.
GÛmez de Soler B., Campeny Vall-Llosera G., Van Der Made J., Oms O., AgustÌ J., Sala R., Blain H.-A., Burjachs F., Claude J., GarcÌa Catal·n S., Riba D. & Rosillo R. (2012).- A new key locality for the Pliocene vertebrate record of Europe: the Camp dels Ninots maar (NE Spain).- Geologica Acta, Barcelona, vol. 10, p. 1-17.
Granados J.L., Moreno O.G. & Brieva C.I. (2013).- Lesiones ulcerativas cut·neas en tortugas dulceacuÌcolas.- Revista de la Facultad de Medicina Veterinaria y de Zootecnia, Bogot·, vol. 60, p. 49-60.
Gray J.E. (1825).- A synopsis of the genera of reptiles and amphibia, with a description of some new species.- Annals of Philosophy, Edinburgh, vol. 10, p. 193-217.
Gray J.E. (1860).- Description of a new species of Emys lately living in the gardens of the Zoological Society.- Proceedings of the Zoological Society of London, London, vol. 1860, 232-233.
Gray J.E. (1869).- Description of Mauremys laniaria, a new freshwater tortoise.- Proceedings of the Zoological Society of London, London, vol. 1869, p. 499-500.
Hefti E., Trechsel U., R¸fenacht H. & Fleisch H. (1980).- Use of dermestid beetles for cleaning bones.- Calcified Tissue International, London, vol. 31, p. 45-47.
Hervet S. (2000).- Tortues du Quaternaire de France : CritËres de dÈtermination, rÈpartitions chronologique et gÈographique.- MÈsogÈe, Marseille, vol. 58, p. 3-47.
Hervet S. (2003).- Le groupe "Palaeochelys sensu latoñMauremys" dans le contexte systÈmatique des Testudinoidea aquatiques du Tertiaire d'Europe occidentale. Apports ý la biostratigraphie et ý la palÈobiogÈographie.- Ph.D. Thesis, MusÈum Nationale d'Histoire Naturelle, Paris, 406 p.
Hervet S. (2004).- SystÈmatique du groupe "Palaeochelys sensu latoñMauremys" (Chelonii, Testudinoidea) du Tertiaire d'Europe occidentale : Principaux rÈsultats.- Annales de PalÈontologie, Paris, vol. 90, p. 13-78.
Highfield A.C. (2002).- An introduction to the Mauremys turtles of the Mediterranean.- Tortoise Trust online article, available from: http://www.tortoisetrust.org/articles/mauremys.html
Hill A.P. (1987).- Damage to some fossil bones from Laetoli. In: Leakey M.D & Harris J.M. (eds.), Laetoli: a Pliocene site in Northern Tanzania.- Clarendon Press, Oxford, p. 543-545.
Hinton H.E. (1945).- A monograph of the beetles associated with stored products, I.- British Museum of Natural History, London, 443 p.
Huchet J.B., Le Mort F., Rabinovich R., Blau S., Coqueugniot H. & Arensburg B. (2013).- Identification of dermestid pupal chambers on southern Levant human bones: inference for reconstruction of Middle Bronze Age mortuary practices.- Journal of Archaeological Science, New York, vol. 40, p. 3793-3803.
Keller C. & Busack S.D. (2001).- Mauremys leprosa (Schweigger, 1812)- Maurische Bachschildkrˆte.- Handbuch der Reptilien und Amphibien Europas, Strasbourg, vol. 3, p. 57-88.
Kotsakis T. (1980).- RÈvision des tortues (Emydidae, Testudinidae, Trionychidae) du Plio-PlÈistocËne de Valdarno supÈrieur (Toscane, Italie).- Quaternaria, Rome, vol. 22, p. 11-37.
Kotsakis T. & Barisone G. (2003).- Cenni sui vertebrati fossili di Roma.- Memorie descrittive della Carta Geologica d'Italia, Rome, vol. 80, p. 115-143.
Kotsakis T. & Mori D. (1981).- Note di paleoerpetologia piemontese. I. I Cheloni delMessiniano superiore di Santa Vittoria d'Alba (Cuneo).- Rivista Piemontese di Storia Naturale, Turin, vol. 2, p. 89-98.
Lapparent de Broin F. de (2000).- African chelonians from the Jurassic to the present: phases of development and preliminary catalogue of the fossil record.- Palaeontologia Africana, Johannesburg, vol. 36, p. 43-82.
Li L., Mo J., Huang Z., Tong H. (2015).- First fossil turtle from Hainan Island, China.- Research & Knowledge, Mahasarakham, vol. 1, p. 48-52.
Lindsay E.H., Opdyke N.D. & Johnson N.M. (1980).- Pliocene dispersal of the horse Equus and late Cenozoic mammalian dispersal events.- Nature, London, vol. 287, p. 135-138.
Panzeri M., Mori E., Mazza G. & Menchetti M. (2014).- Records of introduced stripe-necked terrapins (Mauremys species) in Italy.- Acta Herpetologica, Florence, vol. 9, 227-230.
Patacca E., Scandone P. & Carnevale G. (2013).- The Miocene vertebrate-bearing deposits of Scontrone (Abruzzo, Central Italy): stratigraphic and paleoenvironmental analysis.- Geobios, Villeurbanne, vol. 46, p. 5-23.
Rustioni M. & Mazza P.A.P. (2001).- Taphonomic analysis of Tapirus arvernensis remains from the lower Valdarno (Tuscany, Central Italy).- Geobios, Villeurbanne, vol. 34, p. 469-474.
Sacco F. (1889).- I Cheloni astiani del Piemonte.- Memorie della Reale Accademia delle Scienze di Torino (Serie 2), vol. 39, p. 427-461.
Sidis I. & Gasith A. (1985).- Food habits of the Caspian terrapin (Mauremys caspica rivulata) in unpolluted and polluted habitats in Israel.- Journal of Herpetology, Columbus, vol. 19, p. 108-115.
Theobald W. (1868).- Catalogue of reptiles in the Museum of the Asiatic Society of Bengal.- Journal of the Asiatic Society, Calcutta, Extra number, 88 p.
Timm R.M. (1982).- Dermestids.- Field Museum of Natural History Bulletin, Chicago, vol. 53, p. 14-18.
Vamberger M., Stuckas H., Ayaz D., Lymberakis P., äirok˝ P. & Fritz U. (2014).- Massive transoceanic gene flow in a freshwater turtle (Testudines: Geoemydidae: Mauremys rivulata).- Zoologica Scripta, London, vol. 43, p. 313-322.
Vlachos E. & Delfino M. (2016).- Food for thought: Sub-fossil and fossil chelonian remains from Franchthi Cave and Megalopolis confirm a glacial refuge for Emys orbicularis in Peloponnesus (S. Greece).- Quaternary Science Reviews, Amsterdam, vol. 150, p. 158-171.
Vlachos E., Kotsakis T. & Delfino M. (2015).- The chelonians from the latest Mioceneñearliest Pliocene localities of Allatini and Pylea (East Thessaloniki, Macedonia, Greece).- Comptes Rendus Palevol, Paris, vol. 14, p. 187-205.
Vlachos E., Sterli J., Vasileiadou K. & Syrides G. (2019).- A new species of Mauremys (Testudines, Geoemydidae) from the late MioceneñPliocene of Central Macedonia (northern Greece) with exceptionally wide vertebral scutes.- Papers in Palaeontology, London, vol. 5, p. 177-195.
Wisshak M., Knaust D. & Bertling M. (2019).- Bioerosion ichnotaxa: review and annotated list.- Facies, Erlangen, vol. 65, article #24, 39 p.
Zonneveld J.P. & Bartels W.S. (2020).- Ichnologic note: in defence of Thatchtelithichnus Zonneveld, Bartels, Gunnell and McHugh, 2015.- Ichnos, Philadelphia, vol. 27, no. 2, p. 152-155.
Zonneveld J.P., Bartels W.S., Gunnell G.F. & McHugh L.P. (2015).- Borings in early Eocene turtle shell from the Wasatch Formation, South Pass, Wyoming.- Journal of Paleontology, Lawrence, vol. 89, p. 802-820.