◄ Carnets Geol. 21 (1) ►
![]()
Outline:
[1. Introduction]
[2. Material]
[3. Historical summary regarding the nature of Bacinella]
[4. The corroding ability of the organisms that built bacinella structures]
[5. An alternative hypothesis regarding the origin of bacinella structures]
[6. Soft, firm or hard bacinellastructures]
[7. Bacinella structures and the question of bacinella biostromes versus bacinella bioherms]
[8. Conclusions] [Bibliographic references] and ...
[Plates]
2 impasse Charles Martel, 29217 Plougonvelin (France)
Published online in final form (pdf) on January 21, 2021
DOI 10.2110/carnets.2021.2101
![]()
[Editor: Michel Moullade; language editor:
Phil Salvador; technical editor: Bruno R.C. Granier]
![]()
The controversy surrounding Bacinella irregularis Radoičić, 1959, and Lithocodium aggregatum Elliott, 1956, has involved both sedimentologists and paleontologists. Arguments to exclude them from the Codiaceae are reviewed. The same arguments may be advanced to exclude their ascription to most other organisms. The only hypothesis resisting all elements of disproof is that of a consortium involving microbial communities. Accordingly, these structures are treated here as biosedimentary structures, namely bacinella structures, and not as taxa. One of the features specific to the organisms that built these structures is their strong corroding ability, as shown by the examples of fossils partly or fully cannibalized. This represents one state of corrosion beyond surficial etching and borings. Besides their capacity to corrode calcareous substrates, these microbial organisms formed nodules or oncoids and even built biostromes, which, depending on their stage of development, may have formed soft, firm or even hard substrates. However, to our knowledge, microbial communities contributing to bacinella structures never built up any bioherm. To complete this summary review, a building-block model that coherently takes into account most architectural variations is presented in the form of a figure sketch. The endolithic foraminifer Troglotella incrustans, which is commonly found associated with bacinella structures, is not an encruster or a borer. Lastly, contrary to certain erroneous hypotheses, no major bacinella episode has been ever found coeval with any significant Oceanic Anoxic Event.
• Bacinella;
• Lithocodium;
• Troglotella;
• Jurassic;
• Cretaceous
Granier B.R.C. (2021).- Bacinella, a discrete type of Mesozoic calcimicrobial structure.- Carnets Geol., Madrid, vol. 21, no. 1, p. 1-25.
Bacinella, un type particulier de structure calcimicrobienne mésozoďque.- La controverse entourant Bacinella irregularis Radoičić, 1959, et Lithocodium aggregatum Elliott, 1956, a fait s'opposer sédimentologues et paléontologues. Les arguments pour les exclure des Codiaceae sont passés en revue. Ces męmes arguments peuvent également ętre avancés pour réfuter leur attribution ŕ la plupart des autres groupes d'organismes. La seule hypothčse qui résiste alors ŕ tous ces éléments de réfutation est celle d'une association impliquant des communautés microbiennes. En conséquence, ces structures sont ici traitées comme des structures biosédimentaires, ŕ savoir des structures bacinellae, et non comme des taxons. L'une des caractéristiques propres aux organismes responsables de ces structures bacinellae est leur forte capacité de corrosion, comme en témoignent des exemples de fossiles partiellement ou totalement cannibalisés. Cette cannibalisation représente un état de corrosion au-delŕ de l'altération de surface et des perforations. Outre leur capacité ŕ corroder les substrats calcaires, ces organismes microbiens sont capables de former des nodules ou des oncoďdes, voire męme des biostromes qui, selon leur stade de développement, peuvent constituer des substrats mous, fermes, voire durs. Cependant, dans l'état actuel de nos connaissances, les communautés microbiennes contribuant ŕ ces structures bacinellae n'ont jamais construit de bioherme. Pour compléter ce tour d'horizon synthétique, un modčle de construction rassemblant de maničre cohérente la plupart des variantes architecturales est présenté. Quant au foraminifčre endolithique Troglotella incrustans, fréquemment associé aux structures bacinellae, ce n'est ni un organisme encroűtant, ni un perforant. Enfin, contrairement ŕ certaines hypothčses erronées, aucun épisode majeur ŕ bacinellae n'a jamais été observé ŕ la suite d'un Événement Anoxique Océanique significatif.
• Bacinella ;
• Lithocodium ;
• Troglotella ;
• Jurassique ;
• Crétacé
The controversial nature of Bacinella Radoičić, 1959, and that of its "allied genus" Lithocodium Elliott, 1956, are still the subject of endless discussions about their systematic affinities (Elliott, 1963; Segonzac & Marin, 1972; Banner et al., 1990; Schmid & Leinfelder, 1995, 1996; Schlagintweit et al., 2011a, 2011b; Schlagintweit & Bover-Arnal, 2012, 2013). The present author, who was a longtime follower of André F. Maurin's views (Maurin et al., 1985), never questioned their microbial nature (Granier, 1987) and did not contributed much to the debate until the 29th IAS Meeting of Sedimentology in Schladming. Except for a short report on Bacinella presented on the occasion of this meeting (Granier, 2012a), he never gathered any evidence supporting this assumption. In that connection the scope of this article is to address most significant aspects of these fossil structures. Specifically, it is a mixture of arguments based on paleophycology and carbonate sedimentology, with a discussion on erroneous hypotheses regarding some Bacinella temporal occurrences made by some authors, mostly geochemists (Pittet et al., 2002; Immenhauser et al., 2005; Föllmi, 2008; Rameil et al., 2010; Huck et al., 2010).
With respect to bacinella structures, the present author revised material used for his PhD thesis (Granier, 1987). It consists of Berriasian specimens from Puig Campana (Province of Alicante, Spain) and lower Albian specimens from Serra Gelada/Sierra Helada (Province of Alicante, Spain; the spelling Serra Gelada will be used in the remainder of this paper). The middle Berriasian material comes from the unit 3, i.e., "Neotrocholina Oolites" of the "coupe de la brčche de Roland" (op. cit.: Fig. 5) on the western flank of the Puig, whereas the Albian material comes from several sections located in the Parc Natural de la Serra Gelada that span unit 5, i.e., the "Limestones with Rudists and Oysters":
As for Troglotella, an endolithic foraminifer commonly associated with bacinella structures, and examples of borings, the material presented comes from various locations (Algeria, Brazil, United Arab Emirates) that are documented in Table 1.
Table 1: List of material studied (thin sections).
| Sample, thin section | Lithostratigraphic unit, chronostratigraphic unit | GPS coordinates | Locality, country |
| PC 16463 (28/03/1982) |
Aloisalthella
Limestones, Tithonian-lower Berriasian |
ca. 38°35'21.5"N 0°12'27.4"W | Puig Campana, Finestrat, Alicante, Spain |
| Brčche 7 / ZC16547 (25/03/1983) |
Neotrocholina Oolites,
middle Berriasian |
ca. 38°35'51.0"N 0°12'11.0"W | Puig Campana, Finestrat, Alicante, Spain |
| Brčche 29 / ZC16635 (25/03/1983) |
Coscinoconus
Limestones, upper Berriasian |
ca. 38°35'54.1"N 0°12'15.7"W | Puig Campana, Finestrat, Alicante, Spain |
| Dj3a (19/05/1985) | Coral Biolitites, middle Albian ("Biolithites de Madréporaires") | ca. 38°31'34.6"N 0°24'35.1"W | Cabeço d'Or, Busot, Alicante, Spain |
| HL 232 (25/07/1984) |
Aloisalthella
Limestones, Tithonian-lower Berriasian |
38°33'06.3"N 0°03'39.1"W | Serra Gelada, L'Alfŕs del Pi, Alicante, Spain |
| Carabiniers 14 & 15 / HL 114, HL 115 45x60 & HL HL 115bis (11/06/1984) |
Limestones
with Rudists and Oysters, lower Albian |
38°34'03.3"N 0°03'22.5"W | Serra Gelada, L'Alfŕs del Pi, Alicante, Spain |
| Relais 24 / HL 740 (01/05/1985) |
Limestones
with Rudists and Oysters, lower Albian |
38°33'20.2"N 0°03'43.6"W | Serra Gelada, L'Alfŕs del Pi, Alicante, Spain |
| TC 029 & TO 18 | Kimmeridgian | Saharan Atlas, Algeria | |
| 3020 & 3022, USNM 43016 & USNM 43019 | upper Tithonian-Berriasian | Blake Escarpment, offshore E Florida, U.S.A. |
|
| US34, 8954' | Arab Formation, Kimmeridgian | Umm Shaif, offshore Abu Dhabi, U.A.E. | |
| US 221D, 6157'1" | Kharaib Formation (member no. 3), lowermost Aptian | Umm Shaif, offshore Abu Dhabi, U.A.E. | |
| Cafuz 6 / Cafuz 6.2-A | Riachuelo Formation, ?lower-middle Albian | ca. 10°47'41.6"S 37°16'52.8"W | Fazenda Cafuz, Sergipe Basin, Brazil |
Elliott (1963) and later Segonzac and Marin (1972) regard Bacinella and Lithocodium either as one single organism or as structures built by a single organism. For instance, as early as 1963, Elliott (op. cit.: p. 295) states "it seems likely that Lithocodium and Bačinella may be constructed by the same organism". In contrast, other authors may have regarded Bacinella and Lithocodium as discrete organisms and may even have erected more new species. Although, according the International Code of Botanical Nomenclature (or the International Code of Zoological Nomenclature), Lithocodium should have priority over Bacinella (e.g., Banner et al., 1990; Koch et al., 2002), the popularity of Bacinella exceeds by far that of Lithocodium. Note that Schlagintweit and Bover-Arnal (2013), as Elliott (1963) before, use the original spelling of Bačinella with a caron or inverted circumflex (ˇ) placed over the (c), which means that the correct spelling of Bačinella should be Batchinella, not Bacinella.
Schlagintweit
and Bover-Arnal (2013) reviewed all the Bacinella
representatives to date but, except for Lithocodium aggregatum, their
list does not include any other Lithocodium representatives. However, it
looks like Lithocodium japonicum Endo, 1961, and Lithocodium
morikawai Endo, 1961, both from the Torinosu Limestone (Upper
Jurassic of Japan), are "nearly identical" with Lithocodium
aggregatum. They also summarize and discuss various ascriptions of similar
structures to green algae, red algae, foraminifers, sponges, microbial
structures (Maurin et al., 1985), and consortia of microbes or
microbes with other organisms. For instance, as already pointed out by Cherchi
and Schroeder (2006) and by Schlagintweit and Bover-Arnal
(2013), some Bacinella-like
structures may have mistakenly been considered as an incrusting foraminifer with
agglutinated alveolar walls, similar to Coscinophragma (Schmid
& Leinfelder, 1995, 1996). Reciprocally, as documented here, some
foraminifers may have mistakenly been interpreted as representatives of Lithocodium
(e.g., L. regularis Johnson, 1968: Pl. 2, fig.
3; Pl. 1 ![]() , figs. C-D
herein; op. cit.: Pl. 4, figs. 6-7; Pl. 1
, figs. C-D
herein; op. cit.: Pl. 4, figs. 6-7; Pl. 1 ![]() , figs. A-B herein). Finally,
Schlagintweit and Bover-Arnal
(2013) also contribute to the debate
and put forward their own interpretations.
, figs. A-B herein). Finally,
Schlagintweit and Bover-Arnal
(2013) also contribute to the debate
and put forward their own interpretations.
The name Lithocodium, created by Elliott (1956, 1957) and made from the combination of the ancient Greek λίθος, lithos (stone), and the name of a living alga Codium, is misleading. Starting with Elliott (1956) and until recently, many authors ascribed it to the Codiaceae. However, this assumption is not tenable (Granier, 2011): Codiaceae are green giant unicellular algae. Their cortex is composed of utricules and their medulla of a mesh of siphons. These siphons are not septate (Schlagintweit et al., 2011a, 2011b). Unlike to calcimicrobial structures, Codiaceae are not known as encrusters in the fossil record and modern species are either lightly or not mineralized: If they were green algae, their calcareous coating would most probably be aragonitic, not calcitic (Granier, 2012c). Additionally, Codiaceae are not known to corrode any calcareous substrate. Actually, most remaining options (green algae, red algae, foraminifers or sponges) are hardly defendable hypotheses, which leaves us with the microbial interpretation and/or that of structures built by more than one organism, including some microbes.
Because one cannot refer to micropaleontology and/or to paleophycology to name such biosedimentary structures (note: similarly, coated grains, stromatolites, or reefs are not given binomial Linnean names), there is also no need to refer to taxonomy when dealing with them. Similarly, the use of "bacinellid structures" (initiated by Maurin et al., 1985) should be abandoned because the suffix –id may have a taxonomic connotation (at the level of the family for animals). The use of "bacinelloid structures" is no better either because both suffixes –id and -oid are derived from the same ancient Greek word είδος, eidos (looking like). It is suggested here that both generic names Bacinella and Lithocodium be abandoned, and that these biosedimentary structures be referred to as "bacinella structures" (pronounced "batchinella", and with no first capital as in Bacinella) or bacinellae (plural).
Cherchi
and Schroeder (2013) regard the type-material of Lithocodium
aggregatum Elliott, 1956, as "a
calcimicrobial crust, infested by boring sponges (ichnotaxon Entobia)".
Plate 2 ![]() , figs. A-C, documents an oyster shell with genuine Entobia cavities
for comparison with Elliott's material. The latter display typical
scalloped edges and are partly filled with calcareous chips, which are carving
by-products. A dense boring network affecting another oyster shell is also
illustrated in Plate 2
, figs. A-C, documents an oyster shell with genuine Entobia cavities
for comparison with Elliott's material. The latter display typical
scalloped edges and are partly filled with calcareous chips, which are carving
by-products. A dense boring network affecting another oyster shell is also
illustrated in Plate 2 ![]() , figs. D-E. In both cases similarities with bacinella
structures are quite superficial. The bifurcating
"tubules" of Lithocodium grow up
from basal "vesicles" toward the external surface of the crust
whereas,
in genuine Entobia borings, "exploratory threads" or "intercameral
canals" radiate in all directions from the clionid "chamber", but
not necessarily toward the external surface.
, figs. D-E. In both cases similarities with bacinella
structures are quite superficial. The bifurcating
"tubules" of Lithocodium grow up
from basal "vesicles" toward the external surface of the crust
whereas,
in genuine Entobia borings, "exploratory threads" or "intercameral
canals" radiate in all directions from the clionid "chamber", but
not necessarily toward the external surface.
Demospongiae
spicules are locally found in bacinella structures. Because these
spicules were originally siliceous, although silica was leached and the moldic
cavity later cemented by calcite, their shapes are still preserved. In many
cases it could be difficult to discriminate spicule sections from sections of bacinella
"tubules".
In Plate 3 ![]() , figs. C, E, fusiform shapes perpendicular to "tubules"
have been proven to be spicules. In Plate 3
, figs. C, E, fusiform shapes perpendicular to "tubules"
have been proven to be spicules. In Plate 3 ![]() , fig. D, some circular shapes could
also be sponge spicules. Because sponge spicules 1) form the skeleton of
siliceous sponges, which could be infested by the organisms that built bacinella
structures (e.g., Cherchi
& Schroeder, 2006), 2) could be part of the agglutinated grains
forming the test of Orbitolinidae (and sometimes called "calcite eyes",
e.g., Pl. 3
, fig. D, some circular shapes could
also be sponge spicules. Because sponge spicules 1) form the skeleton of
siliceous sponges, which could be infested by the organisms that built bacinella
structures (e.g., Cherchi
& Schroeder, 2006), 2) could be part of the agglutinated grains
forming the test of Orbitolinidae (and sometimes called "calcite eyes",
e.g., Pl. 3 ![]() , figs. A, D), or 3) were commonly dispersed in the muddy
matrix, no direct relationship between siliceous sponges and bacinella
structures can be established.
, figs. A, D), or 3) were commonly dispersed in the muddy
matrix, no direct relationship between siliceous sponges and bacinella
structures can be established.
Schlagintweit and coauthors (Schlagintweit et al., 2011a, 2011b; Schlagintweit & Bover-Arnal, 2012, 2013) consider that both Bacinella Radoičić, 1959, and Lithocodium Elliott, 1956, are Ulotrichalean algae that, in addition to growing in pre-existing cavities (including borings), could have "excavated complex biogenic branching galleries into biogenic hard substrates" at some stage of their development. According to Banner et al. (1990), "Where relatively large biogenic skeletal fragments have been previously bored, the medullary filaments of encrusting L. aggregatum may fill these borings so closely that the impressions may be gained, erroneously, that the encrusting alga had, itself, created the borings!"
If Ulotrichales and clionids are plausible organisms for the origin of some marginal borings in any calcareous substrate (e.g., corals, mollusk shells, and microbial crusts), both hypotheses should be rejected because they do not provide consistent explanations for:
Because no other hypothesis can explain the facts reported above, the hypothesis that these structures are of microbial nature provides a robust justification for the abandonment of the binomial Linnean classification and for the use of "bacinella structures" when referring to them.
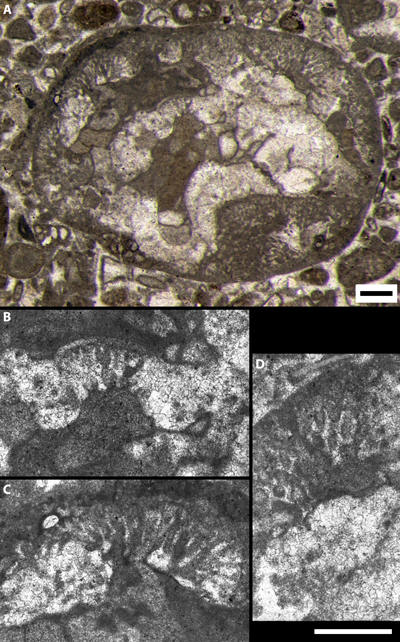
|
Figure 1:
Orbitolinidae
corroded by bacinella structures (excerpt of Granier, 1987: Pl.
50, fig. g), "Limestones with Rudists and Oysters", lower Albian,
Serra Gelada, L'Alfŕs del Pi, Alicante (Spain). Sample Relais 24, thin section
HL 740 01/05/1985. Thick white arrow is upward-oriented. White scale bar = 250
µm. |
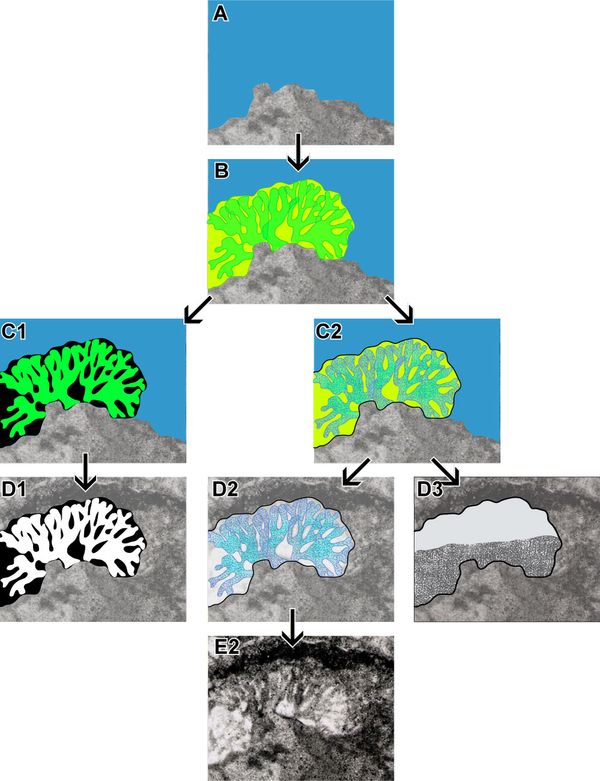
An accreted bacinella nodule from
the middle Berriasian "Neotrocholina Oolites" in the
"coupe de la brčche de Roland" (Puig Campana, Finestrat,
Alicante, Spain) is quite singular because next to bacinella "bubbles"
either empty, partly filled or fully filled by internal sediment,
other "envelopes" enclose bushy
filamentous structures similar to small cayeuxia structures (Granier, 1987: Pl. 52, fig.
e; 2003, Fig. 6; Pl. 5 ![]() , fig. C herein).
This
nodule shows various degrees of preservation and it looks like bacinella
"bubbles"
represent specimens where filaments were either loosely mineralized or not
mineralized. Fig.
2
, fig. C herein).
This
nodule shows various degrees of preservation and it looks like bacinella
"bubbles"
represent specimens where filaments were either loosely mineralized or not
mineralized. Fig.
2 ![]() presents a conceptual model of bacinella
building with three proven or probable end-products.
presents a conceptual model of bacinella
building with three proven or probable end-products.
|
Figure 2:
Conceptual model of bacinella growth with
three proven or probable end-products. This model is based on sample Brčche 7,
thin section ZC16547, "Neotrocholina Oolites", middle Berriasian, Puig
Campana,
Alicante (Spain). See description in the text. |
Some
soft bushy filaments (Fig. 2.B ![]() )
grow on a calcareous substrate (Fig. 2.A
)
grow on a calcareous substrate (Fig. 2.A ![]() ),
e.g., a bacinella oncoid in the case studied. Departing from that there are 3 options:
),
e.g., a bacinella oncoid in the case studied. Departing from that there are 3 options:
To summarize, tubules in micrite look like the so-called "Lithocodium",
empty or partly filled "envelopes" look like bacinella "bubbles",
and calcified bushy filaments look like small cayeuxia structures, as
illustrated in Plate 5 ![]() , figs. A-D.
, figs. A-D.
Bioturbations may have affected large
bacinella structures. Plate 4 ![]() ,
figs. A-B, shows two burrows. Their occurrence in a
bacinella
mesh suggests that the latter was forming a soft to firm substrate.
,
figs. A-B, shows two burrows. Their occurrence in a
bacinella
mesh suggests that the latter was forming a soft to firm substrate.
Plate 4 ![]() ,
fig. G, and Plate 6
,
fig. G, and Plate 6 ![]() , fig. I, (Granier,
1987: Pl. 51, fig. c) show pholadid boring, i.e.,
Gastrochaenolites ichnosp., in spongiostromata to bacinella structures.
Such occurrences suggest that, at some stage, the bacinella structures
were forming firm to hard substrates.
, fig. I, (Granier,
1987: Pl. 51, fig. c) show pholadid boring, i.e.,
Gastrochaenolites ichnosp., in spongiostromata to bacinella structures.
Such occurrences suggest that, at some stage, the bacinella structures
were forming firm to hard substrates.
Plate 6 ![]() , fig. B, shows a tubular and flexuous boring affecting cayeuxia to
spongiostromata to bacinella structures,
and that is often ascribed to Troglotella. Many authors (Werner,
1986; Schlagintweit, 1991,
2008, 2010; Leinfelder et al.,
1993; Schmid & Leinfelder,
1995, 1996; Kołodziej,
1997; Cherchi & Schroeder,
2000) consider that this endolithic foraminifer is
also able to bore its own cavities. As a matter of fact, it has been observed
in borings affecting echinoderm remains (Pl. 6
, fig. B, shows a tubular and flexuous boring affecting cayeuxia to
spongiostromata to bacinella structures,
and that is often ascribed to Troglotella. Many authors (Werner,
1986; Schlagintweit, 1991,
2008, 2010; Leinfelder et al.,
1993; Schmid & Leinfelder,
1995, 1996; Kołodziej,
1997; Cherchi & Schroeder,
2000) consider that this endolithic foraminifer is
also able to bore its own cavities. As a matter of fact, it has been observed
in borings affecting echinoderm remains (Pl. 6 ![]() ,
fig. D),
corals (Pl. 6
,
fig. D),
corals (Pl. 6 ![]() ,
figs. C,
E), and various microbial structures, including cayeuxia structures (Pl. 6
,
figs. C,
E), and various microbial structures, including cayeuxia structures (Pl. 6 ![]() ,
figs. A-B, F); more examples are document by Schlagintweit
(2012). It
is also commonly observed inside bacinella structures (Pl. 6
,
figs. A-B, F); more examples are document by Schlagintweit
(2012). It
is also commonly observed inside bacinella structures (Pl. 6 ![]() ,figs. G-H). In
most cases, the foraminifer wall is never attached to the cavity wall, a feature
also reported from borings genuinely made by other foraminifers (e.g., Cherchi
& Schroeder, 1991: Figs. 4-6; Vénec-Peyré,
1987: Figs. 8-9; 1993: Pl. 1, figs. 1-8; Pl. 3, figs. 2-7). This feature suggests that, contrary
to its specific epiteth, Troglotella incrustans
should not be consider as an encrusting foraminifer.
,figs. G-H). In
most cases, the foraminifer wall is never attached to the cavity wall, a feature
also reported from borings genuinely made by other foraminifers (e.g., Cherchi
& Schroeder, 1991: Figs. 4-6; Vénec-Peyré,
1987: Figs. 8-9; 1993: Pl. 1, figs. 1-8; Pl. 3, figs. 2-7). This feature suggests that, contrary
to its specific epiteth, Troglotella incrustans
should not be consider as an encrusting foraminifer.
According
to Leinfelder et al. (1993), their "Bullopora aff. laevis",
i.e., Troglotella incrustans Wernli
& Fookes, 1992, bores from a shell outer surface inwards whereas,
according to Cherchi and Schroeder
(2000), their "Troglotella
n. sp." bores
from a cavity inside the shell outwards. Because Troglotella has an imperforate
test, as is the case for all agglutinating foraminifers, the first option is not
acceptable. Therefore, the reconstruction of Troglotella
boring activity by Leinfelder et al. (1993: Fig. 7) is not valid
because pseudopods are concentrated at the buccal plug of a single distal
foramen (Pl. 6 ![]() ,
fig. A), and are not distributed all over the test outer
surface, as is the case for the hyaline tests of perforate foraminifers.
,
fig. A), and are not distributed all over the test outer
surface, as is the case for the hyaline tests of perforate foraminifers.
Cherchi and Schroeder (2000) also noted that "La morfologia della cavitŕ tubiforme entro la quele viveva Troglotella si adatta perfettamente alla morfologia esterna del guscio" (i.e., The morphology of the tubular cavity within which Troglotella lived adapts perfectly to the external morphology of the test). However, in the present author's opinion, the opposite is true: The external morphology of the Troglotella test is 'more or less' perfectly adapted to the morphology of the tubular boring within which it lived. The arrangement of the chambers is uniserial in tubular cavities but becomes irregular in other cavities. Cherchi and Schroeder (2000) believe that this "stadio irregolare" (i.e., irregular growth stage) could have represented a final growth stage. However, when considering bacinella structures only, it looks like the "stadio irregolare" is the most common stage and the uniserial arrangement the exception. This fact has been documented many times (Leinfelder et al., 1993: Pl. 42, figs. 2-7; Schmid & Leinfelder, 1995: Figs. 1-5; Schmid & Leinfelder, 1996: Figs. 3.A-B, 4.A-B, 6.A-B; Pl. 1, figs. 1-4; Pl. 2, figs. 1-5; Cherchi & Schroeder, 2000: Pl. 1, fig. 8; Bucur et al., 2010: Pl. 25, figs. 4, 6-7; Pl. 62, fig. 10; Schlagintweit et al., 2011a, 2011b: Fig. 9.b, 9.d-f; Bucur & Săsăran, 2011: Pl. 1, fig. 4; Pl. 2, fig. 5; Pl. 4, fig. 12; Pl. 5, figs. 4-6; Schlagintweit, 2012: Figs. 1.a-b, 3.a-e, 4.a-f, 5.a-j, 6.a-d, 7.a-b, 8.a-d, 9.a-b, 12.a-b; Pleş et al., 2013: Fig. 11.a).
The
observations set out above show that Troglotella
never was a boring foraminifer. Rather, it merely was an endolithic foraminifer.
Its shizonte or its gamonte penetrated in preexisting cavities, including
borings (Pl. 6 ![]() , figs. A-F, I) or vugs left after the organic matter decay
in bacinella crusts.
, figs. A-F, I) or vugs left after the organic matter decay
in bacinella crusts.
Records of bacinella structures were mostly coined as oncoids, i.e., centimeter-sized aggregated and coated grains (Dahanayake et al., 1976) but never as metric to decametric geobodies. However, they may well form biostromes and, as early as 1984 (unpublished), the present author described some peculiar strata in SE Spain as consisting of "grey to whitish micritic limestones forming massive and thick beds (...), characterized by the presence of giant oysters (...) and bio-sedimentary structures of bacinella type, with diffuse contours or, on the contrary, forming the bulk of the rock, therefore true [sic] stromatolites" [i.e., "des calcaires micritiques gris ŕ blanchâtres, en 'bancs compacts' (…), caractérisés par la présence d'huîtres géantes (…) et de structures bio-sédimentaires de type bacinella, diffuses ou, au contraire, formant l'essentiel de la roche, soit alors de véritables stromatolithes" (Granier, 1987)]. Obviously, the current definition of "stromatolites" (see discussion in Riding, 2011a, 2011b) does not fit well with the material studied and the term "microbialites" looks definitely more appropriate today to describe these large bacinella geobodies (Granier, 2003).
|
Figure 3:
A) The "coupe
des carabiniers" in L'Alfŕs del Pi (Granier, 1984: Fig. 9;
1987: Fig. 25; 2003: Fig. 2) spans the lower Albian "Cylindroporella
Oolites" and "Limestones with Rudists and Oysters". The latter
comprises a bacinella biostrome high-lighted in a blue color in the
figure. Caption: a. ooids, b. Orbitolinidae, c. corals, d. rudists, e. oysters,
f gastropods, g. bacinella
structures, h. echinoderm remains; B) Photo of the bacinella
biostrome with vague marbled laminations seen at the outcrop (excerpt of Granier,
1984: Pl. 5, fig. a). |
Per
definition, the lower Albian "Limestones with Rudists and Oysters" (Granier,
1987), which consist of rudist and oyster floatstones with wackestone to bacinella
boundstone matrices, yield numerous bivalve shells. As reported above smaller
shells are commonly partly or fully corroded by the organisms that built bacinella
structures (Pl. 4 ![]() ,
figs. C-D,
F, H-I).
,
figs. C-D,
F, H-I).
To our knowledge, the Spanish discovery represents the first genuine record ever illustrated of a "Bacinella biostrome" (Granier, 1987: Pl. 18, fig. a). In retrospect this minor observation is more important in light of later observations.
Subsequently bacinella biostromes have
also been reported from outcrops in Oman (Pittet et al.,
2002, p.
561). If Bacinella may well have formed biostromes, they never formed
"buildups", contrary to the opinion of Koch et al.
(2002),
Immenhauser et al. (2005)
and Rameil et al. (2010).
Actually, when Koch et al. (2002)
consider "a vertical pile of Lithocodium encrustation sequences and
associated rudist-lenses separated from each other by bioclastic sands" as
a "Lithocodium (syn. Bacinella irregularis)-reef-mound"
or as a "patch-reef", they make the mistake of describing a pile of
superimposed biostromes (hence with almost no relief above the surrounding sea
flour) as a buildup, which would have formed a topographic structure. A modern
analog to bacinella biostromes could be patchy mussel banks on muddy
shores. However, Koch et al. (2002) were
right when they demonstrated the importance of early diagenesis and
differential compaction. It is suggested here that bioturbation and a lesser
density of bacinella in the muddy inter-patch areas could have played a
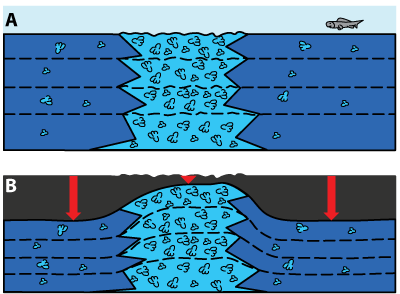
role too, with a higher dewatering (Fig. 4 ![]() ) between the patches.
) between the patches.
|
Figure 4:
A) Pile of bacinella biostromes and
scattered bacinella nodules
in light blue color before dewatering and mechanical, then chemical compaction; B)
same as above after compaction. This sketch illustrates
differential compaction (red arrows). The dotted lines represent time lines. |
Differential compaction as a factor in the creation of pseudo-buildups (actually piles of biostromes, not bioherms) is neglected by Immenhauser et al. (2005), whereas modern differential erosion is neglected by both Immenhauser et al. (2005) and Rameil et al. (2010) when describing surface exposures.
On the basis of a supposed contemporaneity of the erroneously called "Bacinella blooms" with the Oceanic Anoxic Event OAE1a, some authors (Pittet et al., 2002; Immenhauser et al., 2005; Föllmi, 2008; Rameil et al., 2010; Huck et al., 2010) concluded that the two events were related. However, the contemporaneity hypothesis is contradicted by rigorous biostratigraphic (e.g., ammonites) and sequence stratigraphic data (for detailed discussion, see Granier, 2012b, 2014; Granier & Busnardo, 2013).
To summarize:
In 1984, the present author attended the fieldwork seminar organized by the late A.F. Maurin and the late C.L.V. Monty in Santa Pola, Alicante (Spain) for Total C.F.P. On this occasion, he was introduced to an innovative approach to carbonate sedimentology. In 2012, he benefited from a Smithsonian Fellowship allowing him to investigate the J. Harlan Johnson Collection stored on the premises of the Smithsonian Institution. He is grateful for the support provided by the staff of the Department of Paleobiology at the Smithsonian National Museum of Natural History, particularly William A. DiMichele and Jonathan G. Wingerath for having facilitated his work there. In 2015, he was the laureate of a Franco-Brazilian professorship at the UNESPetro in Rio Claro (Săo Paulo State, Brazil) that enabled him to work on Brazilian material. He would like to thank the staff of the UNESPetro, particularly Dimas Dias-Brito for his hospitality. Phil Salvador helped polishing the English text. The reviews of Ioan I. Bucur and George Pleş also helped to improve the manuscript.
Banner F.T., Finch E.M. & Simmons M.D. (1990).- On Lithocodium Elliott (calcareous algae); its paleobiological and stratigraphical significance.- Journal of Micropalaeontology, London, vol. 9, no. 1, p. 21-36.
Bathurst R.C.G. (1966).- Boring algae, micrite envelopes and lithification of molluscan biosparites.- Geological Journal, vol. 5, no. 1, p. 15-32.
Bucur I.I., Săsăran E., Balica C., Beleş D., Bruchental C., Chendeş C., Chendeş O., Hosu A., Lazăr D.F., Lăpădat A., Marian A.V., Mircescu C., Turi V. & Ungureanu R. (2010).- Mesozoic carbonate deposits from some areas of the Romanian Carpathians - Case studies.- Cluj University Press, 203 p.
Bucur I.I. & Săsăran E. (2011).- Upper Jurassic-Lower Cretaceous algae of Haghimas Mountains (Lacul Roşu-Cheile Bicazului area). In: Bucur I.I. & Săsăran E. (eds.), Calcareous algae from Romanian carpathians. Field Trip Guidebook.- 10th International Symposium on Fossil Algae, Cluj-Napoca (12-18 September 2011), Cluj University Press, p. 57-96.
Cherchi A. & Schroeder R. (1991).- Perforations branchues dues ŕ des Foraminifčres cryptobiotiques dans des coquilles actuelles et fossiles.- Comptes Rendus de l'Académie des Sciences (Série II), Paris, t. 312, p. 111-115.
Cherchi A. & Schroeder R. (2000).- Foraminiferi criptobionti del Cenomaniano inferiore della Montagna di Tauch (Corbičres, Francia meridionale).- Giornale di Geologia (ser. 3a), Bologna, vol. 62, supplemento, p. 41-46.
Cherchi A. & Schroeder R. (2006).- Remarks on the systematic position of Lithocodium Elliott, a problematic microorganism from the Mesozoic carbonate platforms of the Tethyan realm.- Facies, Erlangen, vol. 52, p. 435-440.
Cherchi A. & Schroeder R. (2013).- Revision of the holotype of Lithocodium aggregatum Elliott, 1956 (Lower Cretaceous, Iraq): New interpretation as sponge-calcimicrobe consortium.- Facies, Erlangen, vol. 59, no. 1, p. 49-57.
Dahanayake K., Champetier Y. & Hilly J. (1976).- Oncolithes et pseudoncolithes : Relation entre la disposition séquentielle et la participation de Bacinella irregularis Radoičić aux structures oncolithiques.- Bulletin de la Société Géologique de France, Paris, (7e série), t. XVIII, no. 6, p. 1665-1669.
Elliott G.F. (1956).- Further records of fossil calcareous algae from the Middle East.- Micropaleontology, New York, vol. 2, no. 4, p. 327-334 (2 Pls.).
Elliott G.F. (1957).- Algues calcaires codiacées fossiles d'Iraq, nouvelles et peu connues.- Bulletin de la Société Géologique de France (6e série), Paris, t. VI, p. 789-795 (Pl. XXV).
Elliott G.F. (1963).- Problematic microfossils from the Cretaceous and Palaeocene of the Middle East.- Paleontology, vol. 6, part 2, p. 293-300 (Pls. 46-47).
Endo R. (1961).- Calcareous algae from the Jurassic Torinosu Limestone of Japan. In: Commemorative Volume dedicated to Professor Riuji Endo.- The Science Reports of the Saitama University, Series B (Biology and Biology and Earth Sciences), p. 53-75 (17 Pls.).
Föllmi K.B. (2008).- A synchronous, middle Early Aptian age for the demise of the Helvetic Urgonian platform related to the unfolding oceanic anoxic event 1a ("Selli event"). Comment on the article "Sur la présence de grands foraminifčres d'âge aptien supérieur dans l'Urgonien de la Nappe du Wildhorn (Suisse centrale). Note préliminaire" by R. Schroeder, K. Schenk, A. Cherchi & B. Schwizer, Revue de Paléobiologie, 2007, 665-669.- Revue de Paléobiologie, Genčve, vol. 27, no. 2, p. 461-468.
Granier B. (1984), unpublished.- Recherches géologiques sur la Sierra Helada, Province d'Alicante, Espagne.- Rapport de stage, Total C.F.P., Direction Exploration, Département TEP/DE/REN-MCA, 50 p. (12 Pls.).
Granier B. (1987).- Le Crétacé inférieur de la Costa Blanca entre Busot et Altea, Alicante (Espagne) : Biostratigraphie, sédimentologie, évolution tectono-sédimentaire.- Thčse, Docteur de l'Université Paris VI (nouveau régime), 23 Novembre 1987; Mémoires des Sciences de la Terre, Université Pierre et Marie Curie, Paris, 87-49, vol. I (text): 281 p. ; vol. II (plates): 54 Pls.
Granier B. (2003).- Second day : Cretaceous calcareous algae and microbial carbonates from the Prebetic Zone (Betic Cordillera, E Spain).- Field Trip Guide Book, 8th International Symposium on Fossil Algae, Granada, September 18th-20th, p. 25-34.
Granier B. (2011).- Cretaceous benthic calcareous algae.- Paleontologia: Cenários de Vida, Rio de Janeiro, vol. 3, p. 199-210.
Granier B. (2012a).- Bacinella, a discrete type of Mesozoic calcimicrobial structures. In: Gawlick H.J. & Missoni S. (chairpersons), Sedimentology in the hearth of the Alps.- CD-ROM, 29th IAS Meeting of Sedimentology, 10th-13th September, Schladming, p. 91 (abstract).
Granier B. (2012b).- Geochemical correlations of Shu'aiba sections: Stop trusting the trends! In: Gawlick H.J. & Missoni S. (chairpersons), Sedimentology in the hearth of the Alps.- CD-ROM, 29th IAS Meeting of Sedimentology, 10th-13th September, Schladming, p. 361 (abstract).
Granier B. (2012c).- The contribution of calcareous green algae to the production of limestones: A review.- Geodiversitas, Paris, vol. 34, no. 1, p. 35-60.
Granier B. (2014).- Comment on "Early Aptian paleoenvironmental evolution of the Bab Basin at the southern Neo-Tethys margin: Response to global-carbon cycle perturbations across Ocean Anoxic Event 1a" by K. Yamamoto et al.- Geochemistry, Geophysics, Geosystems, vol. 15, no. 5, p. 2086-2090.
Granier B. & Busnardo R. (2013).- New stratigraphic data on the Aptian of the Persian Gulf. In: Skelton P., Granier B. & Moullade M. (eds.), Special issue: Spatial patterns of change in Aptian carbonate platforms and related events.- Cretaceous Research, vol. 39, p. 170-182.
Johnson J.H. (1968).- Lower Cretaceous algae from the Blake Escarpment, Atlantic Ocean, and from Israel.- Professional Contributions of the Colorado School of Mines, Golden, no. 5, vi + 46 p.
Huck S., Rameil N., Korbar T., Heimhofer U., Wieczorek T.D. & Immenhauser A. (2010).- Latitudinally different responses of Tethyan shoal-water carbonate systems to the Early Aptian oceanic anoxic event (OAE 1a).- Sedimentology, vol. 57, no. 7, p. 1585-1614.
Immenhauser A., Hillgärtner H. & Bentum E. van (2005).- Microbial-foraminiferal episodes in the Early Aptian of the southern Tethyan margin: ecological significance and possible relation to oceanic anoxic event 1a.- Sedimentology, vol. 52, p. 77-99.
Koch R., Moussavian E., Ogorelec B., Skaberne D. & Bucur I.I. (2002).- Development of a Lithocodium (syn. Bacinella irregularis)-reef-mound. A patch reef within Middle Aptian lagoonal limestone sequence near Nova Gorica (Sabotin Mountain, W-Slovenia).- Geologija, Ljubljana, Knjiga 45/1, p. 71-90.
Kołodziej B. (1997).- Boring Foraminifera from exotics of the Štramberk-type Limestones (Tithonian-Lower Berriasian, Polish Carpathians).- Annales Societatis Geologorum Poloniae, vol. 67, no. 2-3, p. 249-256.
Leinfelder R.R., Nose M., Schmid D.U. & Werner W. (1993).- Microbial crusts of the Late Jurassic: Composition, palaeoecological significance and importance in reef construction.- Facies, Erlangen, vol. 29, p. 195-230.
Maurin A.F., Bernet-Rollande M.C., Monty C.L.V. & Nazhat S. (1985).- The microbial nature of bacinellid textures. Sedimentological bearings.- 6th European Regional Meeting of Sedimentology, Lerida, p. 285-287.
Pittet B., Buchem F.S.P. van, Hillgärtner H., Razin P., Grötsch J. & Droste H. (2002).- Ecological succession, palaeoenvironmental change, and depositional sequences of Barremian-Aptian shallow-water carbonates in northern Oman.-Sedimentology, vol. 49, p. 555-581.
Pleş G., Mircescu C.V., Bucur I.I. & Săsăran E. (2013).- Encrusting micro-organisms and microbial structures in Upper Jurassic limestones from the Southern Carpathians (Romania).- Facies, Erlangen, vol. 59, p. 19-48.
Radoičić R. (1959).- Nekoliko problematičnih mikrofosila iz dinarske krede.- Vesnik Zavoda za Geološka i Geofizička Istraživanja nr Srbije, Beograd, t. XVII, p. 87-92 (Pls. I-III).
Rameil N., Immenhauser A., Warrlich G., Hillgärtner H. & Droste H.J. (2010).- Morphological patterns of Aptian Lithocodium-Bacinella geobodies: relation to environment and scale.- Sedimentology, vol. 57, no. 3, p. 883-911.
Riding R. (2011a).- Microbialites, stromatolites and thrombolites. In: Reitner J. & Thiel V. (eds.), Encyclopedia of Geobiology.- Encyclopedia of Earth Science Series, Springer, Heidelberg, p. 635-654.
Riding R. (2011b).- The nature of stromatolites: 3,500 million years of history and a century of research. In: Reitner J., Quéric N.-V. & Arp G. (eds.), Advances in stromatolite geobiology.- Lecture Notes in Earth Sciences, Heidelberg, vol. 131, p. 29-74.
Schlagintweit F. (1991).- Allochthone Urgonkalke im Mittleren Abschnitt der Nördlichen Kalkalpen: Fazies, Paläontologie und Paläogeographie.- Münchner Geowissenschaftliche Abhandlungen Reihe A, Band 20, 120 p.
Schlagintweit F. (2008).- Bioerosional structures and pseudoborings from Late Jurassic and Late Cretaceous-Paleocene shallow-water carbonates (Northern Calcareous Alps, Austria and SE France) with special reference to cryptobiotic foraminifera.- Facies, Erlangen, vol. 54, no. 3, p. 377-402.
Schlagintweit F. (2010).- Taxonomic revision of Late Jurassic "Lithocodium aggregatum Elliott" sensu Schmid & Leinfelder, 1996.- Jahrbuch der Geologischen Bundesanstalt, Wien, Band 150, Heft 3-4, p. 393-406.
Schlagintweit F. (2012).- New insights into Troglotella incrustans Wernli & Fookes, 1992, a fascinating Upper Jurassic-Upper Cretaceous foraminifer.- Studia Universitatis Babes-Bolyai Geologia, Cluj napoca, vol. 57, no. 2, p. 17-26.
Schlagintweit F. & Bover-Arnal T. (2012).- The morphological adaptation of Lithocodium aggregatum Elliott (calcareous green alga) to cryptic microhabitats (Lower Aptian, Spain): An example of phenotypic plasticity.- Facies, Erlangen, vol. 58, no. 1, p. 37-55.
Schlagintweit F. & Bover-Arnal T. (2013).- Remarks on Bačinella Radoičić, 1959 (type species B. irregularis) and its representatives.- Facies, Erlangen, vol. 59, no. 1, p. 59-73.
Schlagintweit F., Bover-Arnal T. & Salas R. (2011a).- New insights into Lithocodium aggregatum Elliott 1956 and Bacinella irregularis Radoičić 1959 (Late Jurassic-Lower Cretaceous): Two ulvophycean green algae (?Order Ulotrichales) with a heteromorphic life cycle (epilithic/euendolithic).- Facies, Erlangen, vol. 56, no. 4, p. 509-547.
Schlagintweit F., Bover-Arnal T. & Salas R. (2011b).- Erratum to: New insights into Lithocodium aggregatum Elliott 1956 and Bacinella irregularis Radoičić 1959 (Late Jurassic-Lower Cretaceous): Two ulvophycean green algae (?Order Ulotrichales) with a heteromorphic life cycle (epilithic/euendolithic).- Facies, Erlangen, vol. 56, no. 4, p. 635-673.
Schlagintweit F., Bucur I.I, Rashidi K. & Saberzadeh B. (2013).- Bioerosive structures in orbitolinid foraminifera: examples from the Lower Cretaceous of central Iran.- Studia UBB Geologia, Cluj-Napoca, vol. 58, no. 2, p. 5-12.
Schmid D.U. & Leinfelder R.R. (1995).- Lithocodium aggregatum Elliott n'est pas une algue mais un foraminifčre encroűtant, commensalisé par le foraminifčre Troglotella incrustans Wernli et Fookes.- Comptes Rendus de l'Académie des Sciences (Série IIa), Paris, t. 320, p. 531-538.
Schmid D.U. & Leinfelder R.R. (1996).- The Jurassic Lithocodium aggregatum - Troglotella incrustans foraminiferal consortium.- Paleontology, vol. 39, part 1, p. 21-52.
Segonzac G. & Marin P. (1972).- Lithocodium aggregatum Elliott et Bacinella irregularis Radoičić de l'Aptien de Teruel (Espagne) : Deux stades de croissance d'un seul et męme organisme incertae sedis.- Bulletin de la Société Géologique de France, Paris, (7e série), t. XIV, p. 331-335 (Pl. XI).
Vénec-Peyré M.-T. (1987).- Boring Foraminifera in French Polynesian coral reefs.- Coral Reefs, vol. 5, no. 3, p. 205-212.
Vénec-Peyré M.-T. (1993).- Mise en évidence d'un mode de vie endolithe chez les foraminifčres Gypsina globulus (R.) et Cribrobaggina reniformis (H.-A- et E.) dans les récifs de Polynésie. Révision taxinomique de G. globulus.- Revue de Micropaléontologie, Paris, vol. 36, no. 1, p. 67-75.
Werner W. (1986).- Palökologische und biofazielle Analyse des Kimmeridge (Oberjura) von Consolaçao, Mittelportugal.- Zitteliana, Munich, Band 13, p. 1-109.
Wernli R. & Fookes E. (1992).- Troglotella incrustans n. gen., n. sp., un étrange et nouveau foraminifčre calcicavicole du complexe récifal kimméridgien de Saint-Germain-de-Joux (Ain, France).- Bollettino della Societa Paleontologica Italiana, Modena, vol. 31, p. 95-103.
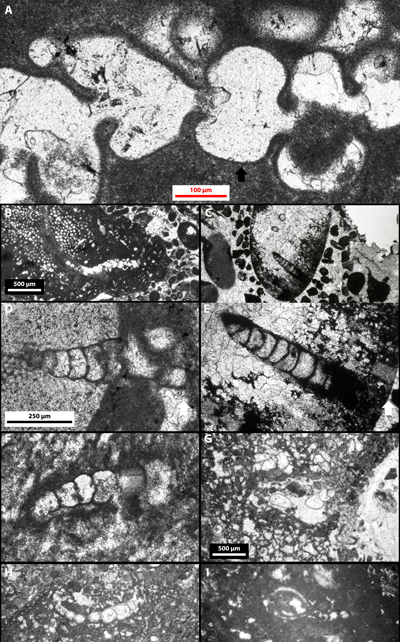
Plate 1: Lithocodium regularis Johnson, 1968 (upper Tithonian-Berriasian, Blake Escarpment, offshore E Florida, U.S.A.), probably a ? Everticyclammina sp. : A-B) excerpts of the original publication (excerpt of Johnson, 1968: Pl. 4, figs. 6-7), thin section Sheridan's 12S (not found); C) excerpt of the original publication (excerpt of Johnson, 1968: Pl. 2, fig. 3), thin section 3022, USNM 43019; D) unpublished, thin section 3020, USNM 43016; E) new microphotograph of Pl. 2, fig. 3, thin section 3022, USNM 43019. Yellow arrows point to miliolid foraminifers that were incorporated in the larger agglutinated foraminiferal test. These specimens bearing U.S. National Museum labels are stored at the Smithsonian Institution in Washington D.C. (U.S.A.). White scale bar (A-C) = 500 µm; black scale bar (D-E) = 250 µm. |
|
|
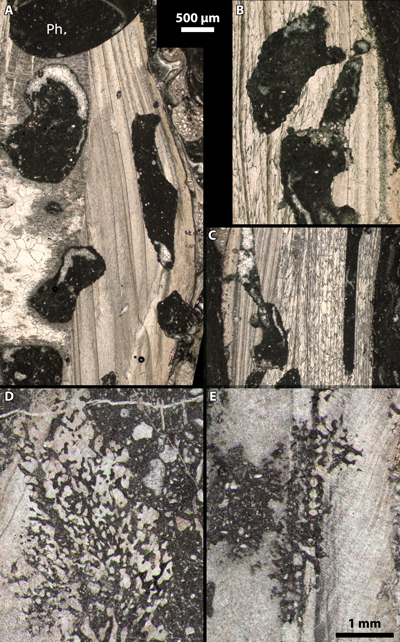
Plate 2: Examples of clionid borings, i.e., Entobia ichnosp., in oyster shells: A-C) Ph.: pholadid boring, i.e., Gastrochaenolites ichnosp., and numerous clionid borings. Riachuelo Formation, ?lower-middle Albian, Fazenda Cafuz, Sergipe Basin (Brazil). Sample Cafuz 6, thin section Cafuz 6.2-A; D-E) "exploratory threads" drilled by clionids. "Coral Biolitites", middle Albian, Cabeço d'Or/Cabezon de Oro, Busot, Alicante (Spain). Sample / thin section Dj3a 19/05/1985. White scale bars (A-C) = 500 µm; black scale bar (D-E) = 1 mm. |
|
|
Plate 3: "Limestones with Rudists and Oysters", lower Albian, Serra Gelada, Alicante (Spain). Yellow arrowheads point to proven (e.g., Pl. 3, figs. C, E) or probable sponge spicules (e.g., Pl. 3, figs. A, D). A) patchy distribution of bacinella structures inside and outside corroded Orbitolinidae; B) crusts displaying the Lithocodium-like pattern; C) sponge spicules (yellow arrows) within a Lithocodium-like crust. The white arrow points to a small corroded calcitic bioclast embedded in the bacinella structure; D) detail of Plate 3, fig. A. Detail of bacinella structures inside and outside corroded Orbitolinidae; E) detail of Plate 3, fig. B. Sponge spicule (yellow arrows) within a Lithocodium-like crust; F-G) cannibalized Orbitolinidae; H) detail of Plate 3, fig. I; I) bacinella structures corroding bivavia shells. Geopetal internal sediment is a micrite with slightly coarser crystals than those of the surrounding matrix; J) detail of Plate 3, fig. I. A-G) sample Relais 24, thin section HL 740 01/05/1985; H-J) sample Carabiniers 14, thin section HL 114 11/06/1984. The thick white arrow is upward-oriented. Black scale bars = 1 mm; white scale bars = 500 µm. |
|
|
Plate 4: A-F, H-I) Photo and photomicrographs of bacinella boundstones from the "Limestones with Rudists and Oysters", lower Albian, Serra Gelada, L'Alfŕs del Pi, Alicante (Spain): A) detail of Plate 4, fig. E. Burrow (excerpt of Granier, 1984: Pl. 5, fig. d); B) detail of Plate 4, fig. E. Burrow partly filled with coprolites (excerpt of Granier, 1984: Pl. 5, fig. c); C) detail of Plate 4, fig. E. Micritic infill of the cavity of a corroded bioclast (? rudist) embedded in the bacinella structure (excerpt of Granier, 1984: Pl. 6, fig. a); D) detail of Plate 3, fig. A. Micritic infill of the cavity of a corroded (?) gastropod shell embedded in the bacinella structure (excerpt of Granier, 1984: Pl. 6, fig. b; 2003: Fig. 8); E) photo of the thin section HL 115 45x60 (excerpt of Granier, 1987: Pl. 23, fig. j; 2003: Fig. 4); F) corroded rudist shell embedded in the bacinella structure (excerpt of Granier, 1984: Pl. 6, fig. d; 2003: Fig. 7); H) detail of Plate 4, fig. F; I) detail of Plate 4, fig. F. Yellow arrows point to the ghost structure of a rudist shell cannibalized by the organisms that built bacinella structures. G) Photomicrograph of a pholadid (orange arrows) boring, i.e., Gastrochaenolites ichnosp. (blue arrows) in a spongiostromata to bacinella structure from the "Aloisalthella Limestones", Tithonian-lower Berriasian, Serra Gelada, L'Alfŕs del Pi, Alicante (Spain). A-E) sample Carabiniers 15, thin section HL 115 45x60 11/06/1984; F, H-I) sample Carabiniers 15, thin section HL 115bis 11/06/1984; G) thin section HL 232 25/07/1984. Thick arrows are upward-oriented. Red scale bar (E) = 1 mm; white scale bar (A-D, F-G) = 500 µm; black scale bar (H-I) = 250 µm. |
|
|
Plate 5: "Neotrocholina Oolites", middle Berriasian, Puig Campana, Alicante (Spain): A) nodule with bacinella and small cayeuxia structures; B) detail of Plate 5, fig. A; C) detail of Plate 5, fig. A (detail of Pl. 52, fig. e, in Granier, 1987; 2003: Fig. 6); D) detail of Plate 5, fig. A. A-D) sample Brčche 7, thin section ZC16547 25/03/1983. Black scale bar (A) and white scale bar (B-D) = 250 µm. |
|
|
Plate 6: A-H) Troglotella incrustans Wernli & Fookes, 1992: A) detail of Pl. 6, fig. B; B) "Coscinoconus Limestones", upper Berriasian, Puig Campana, Finestrat, Alicante (Spain); C) borings in an eroded piece of coral from the Arab Formation, Kimmeridgian, offshore Abu Dhabi (U.A.E.); D) tubular boring in an echinoderm remain from the Kimmeridgian of the Saharan Atlas (Algeria);; E) detail of Plate 6, fig. C; F) tubular boring in a cayeuxia structure from the Kimmeridgian of the Saharan Atlas (Algeria); G) large cavity and H) tubular boring in a bacinella mesh from Kharaib (3) Formation, lowermost Aptian, offshore Abu Dhabi (U.A.E.); I) Photomicrograph of a pholadid boring, i.e., Gastrochaenolites ichnosp., in a spongiostromata to bacinella structure from the "Aloisalthella Limestones", Tithonian-lower Berriasian, Puig Campana, Finestrat, Alicante (Spain). A-B) sample ZC16635/thin section Brčche 29 25/03/1983; C, E) Umm Shaif 34, 8954'; D) TC 029; F) TO 18; G-H) Umm Shaif 221D, 6157'1"; I) sample/thin section PC 16463 28/03/1982. Red scale bar (A) = 100 µm; white scale bar (B-C, G-I) = 500 µm and black scale bar (D-F) = 250 µm. |
|