◄ Carnets Geol. 20 (20) ►
![]()
Outline:
[1. Introduction]
[2. Material and method]
[3. Examples of primary biocrystals and replaced biocrystals]
[4. Gravitational cement from the lowermost Cretaceous (lower Valanginian) of E France]
[5. Endostromatolites from the uppermost Jurassic of SE Spain]
[6. Endostromatolites and gravitational cement from the uppermost Jurassic of W Ukraine]
[7. Conclusions] and ... [Bibliographic references]
Dépt. STU, Fac. Sci. Tech., UBO, 6 avenue Le Gorgeu, CS 93837, F-29238 Brest (France)
Published online in final form (pdf) on November 20, 2020
DOI 10.2110/carnets.2020.2020
![]()
[Editor: Michel Moullade; language editor:
Phil Salvador; technical editor: Bruno Granier]
![]()
Following a brief summary of some fundamentals in carbonate sedimentology (sedimentary petrography) that highlights the significance of organic matter, some examples of biocrystals in carbonate grains/particles, such as bioclasts or ooids, are provided as an introductory chapter to a discussion on gravitational cements versus endostromatolites. The gravitational cements, either marine (fibrous) or continental (dog-tooth), are made of hyaline (i.e., translucent) sparitic crystals whereas endostromatolites are made of colored sparitic crystals and/or micrite. Gravitational cements forms in the vadose zone whereas endostromatolites grow in small rock cavities in the marine phreatic zone. As such the latter can grow centripetally in all directions (not only downward).
asymmetrical cement;
geopetal infills;
endostromatolites;
microbes;
vadose zone;
phreatic zone
Granier B. (2020).- The biosignature of sparite permits the distinction between gravitational cement and endostromatolites.- Carnets Geol., Madrid, vol. 20, no. 20, p. 407-419.
La biosignature de la sparite permet de distinguer un ciment gravitationnel des endostromatolithes.- Après à un bref rappel de quelques principes fondamentaux de la sédimentologie (pétrographie sédimentaire) des carbonates qui met en évidence l'importance de la matière organique, quelques exemples de biocristaux dans les grains calcaires, tels que les bioclastes ou les ooïdes, sont présentés dans un chapitre d'introduction à une discussion portant sur la distinction entre ciments gravitationnels et endostromatolithes. Les ciments gravitationnels, qu'ils soient marins (fibreux) ou continentaux (en "dents de chien"), sont constitués de cristaux sparitiques hyalins (c'est-à-dire translucides) tandis que les endostromatolithes sont constitués de cristaux sparitiques colorés et /ou de micrite. Les ciments gravitationnels se forment dans la zone vadose alors que les endostromatolithes poussent probablement dans de petites cavités de la roche dans la zone phréatique marine. En tant que tels, ces derniers peuvent pousser de manière centripète dans toutes les directions (et pas seulement vers le bas).
ciment assymétrique ;
remplissages géopétaux ;
endostromatolithes ;
microbes ;
zone vadose ;
zone phréatique
In his PhD thesis, the present author (Granier, 1987: p. 96-97, Pl. 14, fig. a; Pl. 15, fig. d; Pl. 16, figs. c-d; Pl. 17, figs. a-b; Pl. 51, figs. h-i) described from uppermost Jurassic strata in "Sierra Helada (Serra Gelada, Alicante, Spain) "des ciments [sic] asymétriques" [asymetric cements] made of layers of radiating fibrous sparitic crystals with an amber yellow color and micritic laminae or bulges. Granier (1987) pointed out that these structures present in former rock cavities grew in various directions, i.e., they were not necessarily formed under gravitational dynamics, and that, accordingly, they should not be called "microstalactites". He interpreted them to be of microbial origin and more specifically "endostromatolites" sensu Monty (1980, 1982) because these structures have grown up in cavities. It appears that, in the scientific literature, such features are commonly erroneously interpreted as pendant or microstalactitic cement (e.g., "gravitational cement" in Flügel, 2004: Pl. 34, fig. 6), and even as microstalacmitic (!) features (e.g., Arnaud-Vanneau, 1980; Arnaud, 1981: vol. 3, Figs. IV, VII, Pls. 33-34). Recent publications document limestones exhibiting similar features from Italy (Amadio et al., 2018) with "dendritic shrub-like fabric" and Algeria (Belkhedim et al., 2019) with "asymmetric fibrous cements (AFC)". In the present author's view, all these structures, being partly of a microbial nature, should not be considered as "cements" (i.e., a word the use of which should be restricted to crystals resulting of the purely chemical precipitation from a brine in voids) but as microbial crusts growing in cavities, i.e., "endostromatolites", as shown through the following simple and naturalistic approach.
The studied material is from the present author's collection (currently stored at University of Western Brittany in Brest). It comprises a set of thin sections from various origins (France, Iran, Spain, and Ukraine), which are used either as references (e.g., bioclasts and ooids) for comparison or as case specimens illustrating either endostromatolites (Spain) or gravitational cements (France), or both of them (Ukraine).
The significance of organic matter in carbonate sedimentology is still noticeably undervalued, which brings us to summarize some essentials on carbonate petrography under a standard microscope with focus on biocrystals. Obviously, small amounts of iron oxides or hydroxides, for example, may contribute to coloring quartz or calcite crystals, but organic matter may contribute too, even more significantly, particularly in biocrystals. With the support of modern techniques (SEM, AFM, EDS-energy dispersive spectroscopy, NanoSIMS, TOF-SIMS, microXANES, fluorescence X, high resolution episcopic microscopy, synchrotron XR fluorescence), recent publications (e.g., Dauphin, 2005, 2016; Stolarski & Mazur, 2005; Gorzelak et al., 2012; Cuif et al., 2018, and older references therein) document how organic matter is incorporated within biocrystals during biomineralization processes. Although none of these techniques are available to the present author, the presence of organic matter within biocrystals, i.e., the biosignature of sparite, can be revealed by their natural colors with transmitted light or with reflected light using the "white card" technique (e.g., Granier, 2017) on a standard microscope.
According to Dunham (1962), most limestones are basically either made of components bound together at deposition, i.e., boundstones, or made of components not bound together at deposition and ranging from mudstone, wackestone, packstone, and grainstone with respect to the arrangement and amount of micrite (former calcareous mud), allochems (calcareous grains), cement and residual porosity. It is worth mentioning that this cardinal classification lumps together all "recrystallized" limestones (e.g., dolomitized limestones) in the broad category of "crystalline limestones" (Dunham, 1962). Micrite refers to the microscopic appearance with transmitted light of crystals smaller than 10 µm in size, i.e., less than one third or half the thickness of the ca. 25 µm thick rock slice affixed on the glass slide, the whole forming the petrographic thin section. Accordingly, micrite is dark, not translucent, in transmitted light. Allochems, as well as components bound together at deposition, can be partly or fully micritic but some also comprise yellowish sparite parts whereas others may comprise hyaline (i.e., translucent) sparite parts. The yellowish sparite is either 1) primary, i.e., consisting of genuine biocrystals that constitute calcitic skeletons, tests or shells, or 2) secondary, i.e., resulting from the mosaic replacement of primary biocrystals of aragonite or high-Mg calcite. As summarized by Granier et al. (2016), "Replacement of aragonite by calcite (i.e., calcitization) commonly results in a mosaic of crystals of a similar size, with a brownish pseudopleiochroism due to the preservation of the original organic matter of the bioclast". The color and the pseudopleiochroism are directly related to the organic matter contents of these calcite crystals. In contrast, hyaline sparite is a translucent cement resulting from the direct precipitation of calcite from a brine in vugs to form palisadic or drusic cements, or it could be the mosaic replacement of a primary aragonite or high-Mg calcite cement. Consequently, most hyaline calcite crystals that result from chemical precipitation should be treated as cement sensu stricto, whereas most colored calcite crystals are either primary biocrystals or recrystallized biocrystals and accordingly they should NOT be treated as cement.
|
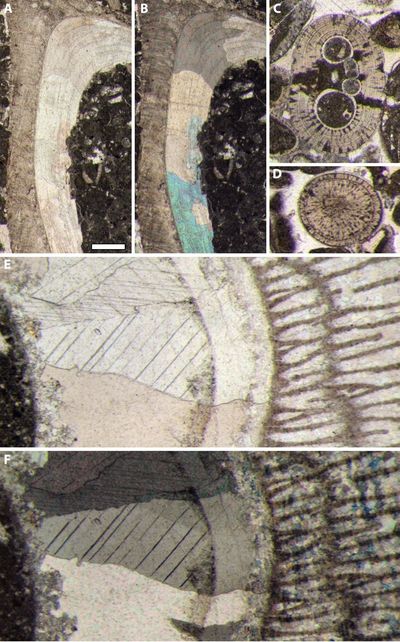
Figure 1:
A-B) Echinoderm (crinoid) remains --colored part on the left-hand side-- with
its syntaxial overgrowth --translucent part on the right-hand side--. The whole
feature is behaving like a single crystal. Thin section B.12566,
exact location unknown, Brittany (France), Paleozoic. A) transmitted light; B)
cross polars. C-D) Rudist shell with a coarsely fibrous primary calcitic part to
the left and a secondary calcitic part to the right. The latter consists of a
mosaic of calcite crystals that replaced the original aragonitic part. Ghosts of
the primary structure are locally preserved, such as the vertical lining in the
middle of both pictures (white arrowheads). Thin section HL
74 (B) 45x60, Serra Gelada, L'Alfàs del
Pi, Alicante (Spain), "Calcaires à Rudistes et à Huîtres", lower
Albian. C) transmitted light;
D) cross polars. All photos same scale bar = 250 µm. |
|
Figure
2:
A-B) Rudist shell with a colored primary calcitic part to the left and a
secondary calcitic part to the right. The latter consists of a mosaic of calcite
crystals that replaced the original aragonitic part. Ghosts of the primary
structure, i.e., many linings, are locally preserved. Thin section HL 42 bis (A) 45x60, Serra Gelada,
L'Alfàs del
Pi, Alicante (Spain), "Calcaires à Rudistes et à Huîtres", lower
Albian. A) transmitted light;
B) cross polars. C-D) Radial fibrous sparitic marine ooids. Thin section BR
2530, Golian, Shirwan, North Khorasan (Iran), Tirgan Formation, upper
Barremian. Transmitted light. E-F) Rudist shell (Eoradiolites) with a
secondary calcitic part to the left and a primary calcitic cellular part to the
right. The cellular part (right) consists of walls made of yellowish primary
calcite and vugs cemented by a translucent drusy sparitic cement. The secondary
calcitic part (left) consists of a mosaic of calcite crystals that replaced the
original aragonitic part. Ghosts of the primary structure are locally preserved,
such as the irregular curved lining in the middle of both pictures. Thin section
HL 74 (A) 45x60,
Serra Gelada, L'Alfàs del Pi, Alicante (Spain), "Calcaires à Rudistes et
à Huîtres", lower Albian: E) transmitted light; F) cross polars. All
photos same scale bar = 250 µm. |
A
short list of seminal books helping on identification of bioclasts in
sedimentary petrography includes Bøggild (1930), Miík
(1966), Majewske
(1969), Horowitz and Potter
(1971), Samuel et al.
(1972), Flügel (1982, 2004,
2010), or the more recent Dias-Brito
et al. (2017), amongst many others. They provide illustrations of 1)
primarily calcitic bioclasts (e.g., echinoids, Fig.
1.A-B ![]() herein), 2)
primarily aragonitic bioclasts (e.g., gastropods), eventually secondarily
replaced (e.g., bivalves, Fig. 4.E
herein), 2)
primarily aragonitic bioclasts (e.g., gastropods), eventually secondarily
replaced (e.g., bivalves, Fig. 4.E ![]() herein) or dissolved and later
secondarily cemented (e.g., gastropods, Fig. 4.B
herein) or dissolved and later
secondarily cemented (e.g., gastropods, Fig. 4.B ![]() herein), and 3) dual
calcitic-aragonitic bioclasts (e.g., serpulids or rudists, Figs.
1.C-D
herein), and 3) dual
calcitic-aragonitic bioclasts (e.g., serpulids or rudists, Figs.
1.C-D ![]() ,
2.A-B, .E-F
,
2.A-B, .E-F ![]() herein). Primary
biocrystals and replaced biocrystals are amber yellow colored
or "cloudy" due to their residual organic matter content. A few
examples are documented and illustrated herein.
herein). Primary
biocrystals and replaced biocrystals are amber yellow colored
or "cloudy" due to their residual organic matter content. A few
examples are documented and illustrated herein.
|
|
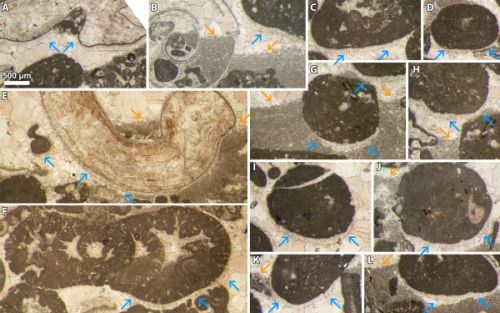
Figure
3:
A) Polished slab of an oncoid floastone to bindstone facies from Serra Gelada,
L'Alfàs del Pi, Province of Alicante (Spain), "Aloisalthella
Limestones", Tithonian-lower Berriasian. White arrowheads point to
stratiform binding micritic films, red arrowhead points to a patch of
red-colored micrites corresponding to upper Valanginian lower Aptian pelagic
mud that has percolated downward into the porous network. Sample SH
19; B) Detail of a colored fibrous sparitic cauliflower-like structure
growing in a cavity of the "Aloisalthella Limestones" (Granier,
1987, Pl. 15, fig. d). Thin section SHJ 1B, Serra
Gelada, L'Alfàs del Pi, Province of Alicante (Spain); C) Polished slab of the
beachrock facies from Crozet, Ain Department (France), Chambotte Formation,
lower Valanginian. Sample 65; D) Polished slab of a coquina facies from Bukivna/Буківна,
Ivano-Frankivsk Oblast (Ukraine), "Niźniower
Kalkstein", Tithonian-lower Berriasian. The yellow arrowhead points to a
former shelter cavity below a bivalve shell, now fully cemented. Sample F3.
Thick black arrowheads are upward-oriented; scale bars: red = 1 cm, white = 500
µm. |
Radial
ooids - samples BR2530 (Fig. 2.C-D ![]() ): The material was originally collected at
Golian, 10 km south of Shirwan, North Khorasan Province, Iran (GPS coordinates:
37°14'13.8"N 57°54'17.3"E). The microfacies of this rock sample from
the lower part of the Tirgan Formation, upper Barremian, is typically a
floatstone of Balkhania balkhanica Mamontova and Pseudoactinoporella
iranica Bucur et al. with a bioclastic-oolitic grainstone
matrix. Nobody ever questioned the role of organic matter in
bioclasts whereas fierce debates have taken place regarding its role in ooids (e.g.,
Dangeard, 1936; O'Reilly et al.,
2017; Diaz
& Eberli, 2019, and references therein) and in microbial crusts or
stromatoids sensu Kalkowsky (1908). Microbes and mucilage (i.e.,
extracellular polymeric substances or EPS) were well documented in modern and
fossil stromatolites and oncoids. On the opposite side until recently (e.g.,
Brehm et al., 2004; O'Reilly et al.,
2017; Diaz
& Eberli, 2019, and references therein), their role in the
ooid factory was still questioned. Iranian ooids are calcitic with a radial
fibrous fabric suggesting they were originally formed as high-magnesium calcite.
Their cortices are light brown, which implies that organic matter is embedded in
their crystal framework.
): The material was originally collected at
Golian, 10 km south of Shirwan, North Khorasan Province, Iran (GPS coordinates:
37°14'13.8"N 57°54'17.3"E). The microfacies of this rock sample from
the lower part of the Tirgan Formation, upper Barremian, is typically a
floatstone of Balkhania balkhanica Mamontova and Pseudoactinoporella
iranica Bucur et al. with a bioclastic-oolitic grainstone
matrix. Nobody ever questioned the role of organic matter in
bioclasts whereas fierce debates have taken place regarding its role in ooids (e.g.,
Dangeard, 1936; O'Reilly et al.,
2017; Diaz
& Eberli, 2019, and references therein) and in microbial crusts or
stromatoids sensu Kalkowsky (1908). Microbes and mucilage (i.e.,
extracellular polymeric substances or EPS) were well documented in modern and
fossil stromatolites and oncoids. On the opposite side until recently (e.g.,
Brehm et al., 2004; O'Reilly et al.,
2017; Diaz
& Eberli, 2019, and references therein), their role in the
ooid factory was still questioned. Iranian ooids are calcitic with a radial
fibrous fabric suggesting they were originally formed as high-magnesium calcite.
Their cortices are light brown, which implies that organic matter is embedded in
their crystal framework.
Rudist
shells - samples HL (Figs. 1.C-D ![]() ,
2.A-B, .E-F
,
2.A-B, .E-F ![]() ): The material was originally
collected at Morro de Sant Jordi, Parc Natural de la Serra Gelada, L'Alfàs del
Pi, Province of Alicante, Spain (GPS coordinates: 38°34'03.0"N
0°03'21.8"W).
Per definition rudist floatstones with wackestone to bacinella bindstone
(Granier, 2012, and in prep.) matrices from the "Calcaires à
Rudistes et à Huîtres" (Granier, 1987), lower Albian, yield
numerous bivalve shells. In both samples (HL42bis: Requienidae, and HL74: Requienidae and
Eoradiolites gr. davidsoni Hill), the originally aragonitic inner layer of the rudist shells is replaced
by a mosaic of large calcite crystals that retain few organic ghosts of the
primary structure (Figs. 1.C-D
): The material was originally
collected at Morro de Sant Jordi, Parc Natural de la Serra Gelada, L'Alfàs del
Pi, Province of Alicante, Spain (GPS coordinates: 38°34'03.0"N
0°03'21.8"W).
Per definition rudist floatstones with wackestone to bacinella bindstone
(Granier, 2012, and in prep.) matrices from the "Calcaires à
Rudistes et à Huîtres" (Granier, 1987), lower Albian, yield
numerous bivalve shells. In both samples (HL42bis: Requienidae, and HL74: Requienidae and
Eoradiolites gr. davidsoni Hill), the originally aragonitic inner layer of the rudist shells is replaced
by a mosaic of large calcite crystals that retain few organic ghosts of the
primary structure (Figs. 1.C-D ![]() ,
2.A-B, .E-F
,
2.A-B, .E-F ![]() ).
).
Bivalve
shell - sample 65 (Fig. 4.E ![]() ): The material was originally collected on the side
of the parking area near the Fierney cable-car station of Crozet, Ain Department,
France (GPS coordinates: 46°17'03.3"N 6°00'22.0"E). The bioclastic
grainstone facies from the uppermost part of the Chambotte Formation (Böker,
1994; Granier, 2019a), lower Valanginian, yield numerous large benthic
foraminifers, including Pfenderina neocomiensis (Pfender) and Pseudocyclammina
lituus (Yokoyama) (Fig. 4.F
): The material was originally collected on the side
of the parking area near the Fierney cable-car station of Crozet, Ain Department,
France (GPS coordinates: 46°17'03.3"N 6°00'22.0"E). The bioclastic
grainstone facies from the uppermost part of the Chambotte Formation (Böker,
1994; Granier, 2019a), lower Valanginian, yield numerous large benthic
foraminifers, including Pfenderina neocomiensis (Pfender) and Pseudocyclammina
lituus (Yokoyama) (Fig. 4.F ![]() ). Surprisingly
all primary aragonitic bioclasts were not dissolved to form micritic envelopes (e.g.,
Fig. 4.B
). Surprisingly
all primary aragonitic bioclasts were not dissolved to form micritic envelopes (e.g.,
Fig. 4.B ![]() ); some were replaced by a mosaic of calcite crystals that retains some
organic ghosts of the primary structure (Fig. 4.E
); some were replaced by a mosaic of calcite crystals that retains some
organic ghosts of the primary structure (Fig. 4.E ![]() ). It is suggested here that
the original amount of organic matter in the original biocrystals, as well as
its later degree of oxidation and the mesh density of the original aragonite
needles, could be clues to this structural difference.
). It is suggested here that
the original amount of organic matter in the original biocrystals, as well as
its later degree of oxidation and the mesh density of the original aragonite
needles, could be clues to this structural difference.
Gravitational
cement - sample 65 (Figs. 3.C ![]() , 4
, 4 ![]() ): Same as above for the "Bivalve shell -
sample 65", the material was originally collected on the side of the parking
area near the Fierney cable-car station of Crozet (GPS coordinates: 46°17'03.3"N 6°00'22.0"E), from the lower Valanginian part of the Chambotte Formation.
): Same as above for the "Bivalve shell -
sample 65", the material was originally collected on the side of the parking
area near the Fierney cable-car station of Crozet (GPS coordinates: 46°17'03.3"N 6°00'22.0"E), from the lower Valanginian part of the Chambotte Formation.
The
fabric is dominantly grain-supported with well-sorted and commonly coarse grains
(Fig. 3.C ![]() ). It is a beach-rock facies as evidenced by the fibrous (hence marine)
gravitational cement that developed in intergranular cavities below the largest
grains (Fig. 4.B
). It is a beach-rock facies as evidenced by the fibrous (hence marine)
gravitational cement that developed in intergranular cavities below the largest
grains (Fig. 4.B ![]() , blue arrowheads). This gravitational cement documents a marine
vadose setting where beach-rocks form. Some micrite is also partly infilling
cavities, including some micritic envelopes following the leaching of the
primarily aragonitic bioclasts (Fig. 4.B
, blue arrowheads). This gravitational cement documents a marine
vadose setting where beach-rocks form. Some micrite is also partly infilling
cavities, including some micritic envelopes following the leaching of the
primarily aragonitic bioclasts (Fig. 4.B ![]() ). A thin isopachous palisadic cement is
locally observed (Fig. 4.B
). A thin isopachous palisadic cement is
locally observed (Fig. 4.B ![]() ), which predates the leaching of aragonite. It
suggests an initial marine phreatic setting for diagenesis, hence the occurrence
of a short episode of (subaqueous) hardground formation preceding emersion and (subaerial)
beach-rock formation. Not all the aragonitic bioclasts were leached. As
documented above, a few were replaced by calcite (Fig. 4.E
), which predates the leaching of aragonite. It
suggests an initial marine phreatic setting for diagenesis, hence the occurrence
of a short episode of (subaqueous) hardground formation preceding emersion and (subaerial)
beach-rock formation. Not all the aragonitic bioclasts were leached. As
documented above, a few were replaced by calcite (Fig. 4.E ![]() ). The micrite, which
later percolated through the porous network (i.e., intergranular,
intragranular and moldic porosity), commonly displays a geopetal layout (Fig.
4.B, .E, .G-H, .J-L
). The micrite, which
later percolated through the porous network (i.e., intergranular,
intragranular and moldic porosity), commonly displays a geopetal layout (Fig.
4.B, .E, .G-H, .J-L ![]() , orange arrowheads). It postdates the leaching that in turn
postdates the gravitational cement. Finally, the remaining porosity was cemented
by a meteoric phreatic cement and then a deep-burial drusy sparitic cement.
, orange arrowheads). It postdates the leaching that in turn
postdates the gravitational cement. Finally, the remaining porosity was cemented
by a meteoric phreatic cement and then a deep-burial drusy sparitic cement.
|
|
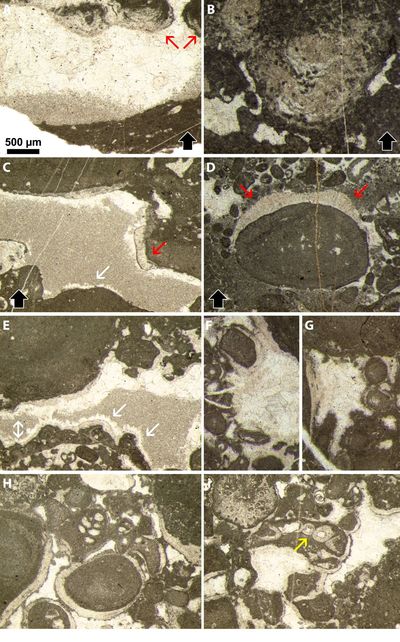
Figure 4:
Early gravitational cement (blue arrowheads) and late geopetal
micritic infills (orange arrowheads) from Crozet, Ain Department (France),
Chambotte Formation, lower Valanginian. A, D, I, L) Thin section Bb1995/65c.
B-C, K) Thin section BR 2594. E-F) Thin section
BR 2595. G-H, J) Thin section Bb2003/65c'''.
All photos same scale bar = 500 µm. |
Endostromatolites
- samples SHJ (Figs. 3.A-B ![]() , 5
, 5 ![]() ): The material was originally collected at Morro
de Sant Jordi, Parc Natural de la Serra Gelada, L'Alfàs del Pi, Province of
Alicante, Spain (GPS coordinates: 38°33'07.3"N 0°03'32.0"W). The
stratigraphic unit was improperly named "Mud mounds à Clypéines" (Granier,
1987), Tithonian-lower Berriasian. Although the fabrics are dominated by
microbial macrostructures (oncoids, algal mats, stromatolites, ...) and comprise
various microstructures (such as Bacinella, Cayeuxia, Ortonella,
Girvanella, Gakhumella huberi, coccoids, ... and microbial
crusts /stromatoids/ growing in cavities, hence endostromatolites), this unit
should have been interpreted as biostromes (Fig. 3.A
): The material was originally collected at Morro
de Sant Jordi, Parc Natural de la Serra Gelada, L'Alfàs del Pi, Province of
Alicante, Spain (GPS coordinates: 38°33'07.3"N 0°03'32.0"W). The
stratigraphic unit was improperly named "Mud mounds à Clypéines" (Granier,
1987), Tithonian-lower Berriasian. Although the fabrics are dominated by
microbial macrostructures (oncoids, algal mats, stromatolites, ...) and comprise
various microstructures (such as Bacinella, Cayeuxia, Ortonella,
Girvanella, Gakhumella huberi, coccoids, ... and microbial
crusts /stromatoids/ growing in cavities, hence endostromatolites), this unit
should have been interpreted as biostromes (Fig. 3.A ![]() ) rather than as bioherms.
In addition, a correct labeling should have been "Clypeina
Limestones" (Granier, 2019a) or better "Aloisalthella
Limestones", based on the latest systematics of the algal epynom (Granier
& Lethiers, 2019).
) rather than as bioherms.
In addition, a correct labeling should have been "Clypeina
Limestones" (Granier, 2019a) or better "Aloisalthella
Limestones", based on the latest systematics of the algal epynom (Granier
& Lethiers, 2019).
The
rock sample corresponds to a microbial boundstone (Fig. 3.A ![]() ), probably formed in
a very shallow-water setting. Some large cavities between grains were sites for
endostromatolitic growth. These commonly zoned
endostromatolites comprise yellowish fibrous sparitic crusts and micritic
crusts and bulges. Although they commonly look
like they were formed under some gravity control, there are
places where that was obviously not the case (Fig. 5.C-D
), probably formed in
a very shallow-water setting. Some large cavities between grains were sites for
endostromatolitic growth. These commonly zoned
endostromatolites comprise yellowish fibrous sparitic crusts and micritic
crusts and bulges. Although they commonly look
like they were formed under some gravity control, there are
places where that was obviously not the case (Fig. 5.C-D ![]() ). As already documented
by Granier (1987), the remainder of the cavities were then filled with
1)
dog-tooth like sparitic cements (Fig. 5.C, .E
). As already documented
by Granier (1987), the remainder of the cavities were then filled with
1)
dog-tooth like sparitic cements (Fig. 5.C, .E ![]() , white arrowheads), which are the
mark of a continental phreatic setting, 2) red-colored marine micrites
that have percolated downward through the remaining porous network
and a fracture network, and finally 3) a drusy sparitic cement, which
corresponds to the deep burial setting. At Serra Gelada, the Tithonian-lower Berriasian "Aloisalthella
Limestones" are topped by an angular unconformity (Granier, 1987,
Pl. 13, fig. d) in the form of a bored surface, a feature which suggests the
occurrence of at least one episode of emersion and erosion preceding a flooding.
The fracture network is post-early Valanginian in age (see Granier,
1987; Granier et al.,
1995). It was polyphasic,
as documented by several episodes of fracturing, phreatic cementation and red-colored
micrite infill (e.g., Granier, 1987, Pl.
16, fig. a), which have occurred over part of the late Valanginian earliest
Aptian time interval.
, white arrowheads), which are the
mark of a continental phreatic setting, 2) red-colored marine micrites
that have percolated downward through the remaining porous network
and a fracture network, and finally 3) a drusy sparitic cement, which
corresponds to the deep burial setting. At Serra Gelada, the Tithonian-lower Berriasian "Aloisalthella
Limestones" are topped by an angular unconformity (Granier, 1987,
Pl. 13, fig. d) in the form of a bored surface, a feature which suggests the
occurrence of at least one episode of emersion and erosion preceding a flooding.
The fracture network is post-early Valanginian in age (see Granier,
1987; Granier et al.,
1995). It was polyphasic,
as documented by several episodes of fracturing, phreatic cementation and red-colored
micrite infill (e.g., Granier, 1987, Pl.
16, fig. a), which have occurred over part of the late Valanginian earliest
Aptian time interval.
|
|
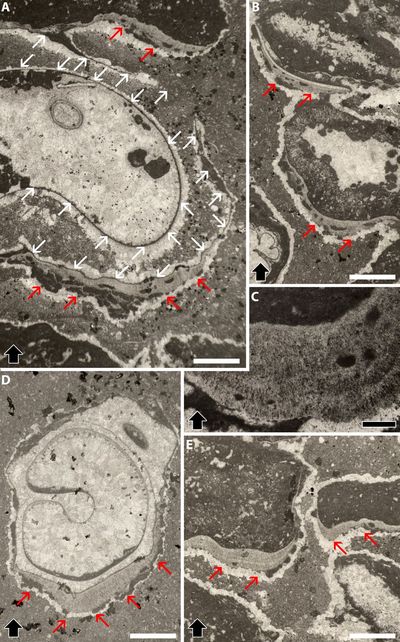
Figure
5:
Endostromatolites (red arrowheads) from Serra Gelada, L'Alfàs del Pi,
Province of Alicante (Spain), "Aloisalthella Limestones",
Tithonian-lower Berriasian. Thick black arrowheads in A-D are upward-oriented;
E-I are oblique or horizontal sections (i.e., oblique or parallel to
stratification). A) infill, cf. Granier, 1987, Pl.
51, fig. h. The red arrowheads point to non-strictly gravitational crusts. Thin
section SHJ 1B; B) Thin section SHJ1
b; C) The red arrowhead points to a non-gravitational crust whereas the
white arrowhead points to a dog-tooth like sparitic cement. Thin
section SHJ1B (uncovered); D) The red arrowheads
point to a non-gravitational crust. Thin section SHJ 2
(A); E) The white arrowheads point to the dog-tooth like
sparitic cement. Thin section SHJ 1B (?); F) Thin section SHJ1B
(covered); G) Thin section SHJ1B (covered);
H) Thin section SHJ 1B (?); I) The yellow arrowhead points to verticil of Aloisalthella
sulcata (Alth), the calcification of which is yellowish-colored as
the endostromatolitic crusts. Thin section SHJ 1B (?). All photos same scale bar
= 500 µm. |
Endostromatolites
and gravitational cement - sample F3 (Figs. 3.D ![]() , 6
, 6 ![]() ): The material was originally
collected near Bukivna/Буківна,
Ivano-Frankivsk Oblast/Province, Ukraine (GPS coordinates: 48°59'04.5"N
24°58'07.4"E).
The stratigraphic unit corresponds to the "Niźniower
Kalkstein" (Alth, 1882; Głażewski, 1937; Granier,
2019b, 2020), upper Tithonian-lower Berriasian, the stratum typicum of Actinoporella
podolica (Alth, 1878) (see Granier,
2019b), that of Aloisalthella
sulcata (Alth, 1882) (see Granier & Lethiers,
2019), and also that of some agglutinated foraminifers described by Cushman
and Głażewski (1949).
): The material was originally
collected near Bukivna/Буківна,
Ivano-Frankivsk Oblast/Province, Ukraine (GPS coordinates: 48°59'04.5"N
24°58'07.4"E).
The stratigraphic unit corresponds to the "Niźniower
Kalkstein" (Alth, 1882; Głażewski, 1937; Granier,
2019b, 2020), upper Tithonian-lower Berriasian, the stratum typicum of Actinoporella
podolica (Alth, 1878) (see Granier,
2019b), that of Aloisalthella
sulcata (Alth, 1882) (see Granier & Lethiers,
2019), and also that of some agglutinated foraminifers described by Cushman
and Głażewski (1949).
The
rock sample corresponds to a coquina (Fig. 3.D ![]() ), probably in a supratidal
wash-over setting. Its texture is that of floatstone to rudstone because the
allochems are large, including numerous gastropods, some corals, sponges, Arabicodium
sp. (i.e., a large Bryopsidalean alga), and Cayeuxia
microstructures. The related large cavities between grains or inside them were
sites for endostromatolitic growth (Fig. 6
), probably in a supratidal
wash-over setting. Its texture is that of floatstone to rudstone because the
allochems are large, including numerous gastropods, some corals, sponges, Arabicodium
sp. (i.e., a large Bryopsidalean alga), and Cayeuxia
microstructures. The related large cavities between grains or inside them were
sites for endostromatolitic growth (Fig. 6 ![]() ). Although the yellowish fibrous
sparitic layers commonly look like they were formed under
gravity control, that is not obvious in some places. These endostromatolites,
commonly zoned, also comprise micritic bulges, either embedded or developing on
the side of the crust. The emersion that affected the "Niźniow" limestones led to the dissolution of most
aragonitic shells, leaving behind micritic envelops, in a (?) continental vadose
setting. The development of non-isopachous, hence partly gravitational,
dog-tooth sparitic cement in all cavities, including in the micritic envelops,
means that the rock remained in a dominantly continental vadose setting (with at
least one episode in a phreatic setting). The micrite, which percolated later
through the porous network in a marine phreatic setting and almost completely
filled in the cavities, postdates the dog-tooth cementation and is probably the
mark of the Late Cretaceous marine transgression in the area.
). Although the yellowish fibrous
sparitic layers commonly look like they were formed under
gravity control, that is not obvious in some places. These endostromatolites,
commonly zoned, also comprise micritic bulges, either embedded or developing on
the side of the crust. The emersion that affected the "Niźniow" limestones led to the dissolution of most
aragonitic shells, leaving behind micritic envelops, in a (?) continental vadose
setting. The development of non-isopachous, hence partly gravitational,
dog-tooth sparitic cement in all cavities, including in the micritic envelops,
means that the rock remained in a dominantly continental vadose setting (with at
least one episode in a phreatic setting). The micrite, which percolated later
through the porous network in a marine phreatic setting and almost completely
filled in the cavities, postdates the dog-tooth cementation and is probably the
mark of the Late Cretaceous marine transgression in the area.
|
|
Figure 6:
Endostromatolites (red arrowheads) from Bukivna/Буківна,
Ivano-Frankivsk Oblast (Ukraine), "Niźniower
Kalkstein", Tithonian-lower Berriasian. A) White arrowheads point to the
micritic envelop of a former aragonitic gastropod shell. Thin section BR 3008.
B) As in the previous picture, the micritic infill of the interparticular and
moldic cavities postdates a phreatic cementation, that in turn postdates the
dissolution of aragonitic shells. Thin section BR 3005. C) Detail of a colored
fibrous sparitic crust growing in a cavity. Thin section BR 3008. D) Colored
fibrous sparitic crusts in a cavity below a gastropod shell. Thin section BR
3010. E) Colored fibrous sparitic crusts growing in cavities. Thin section BR
3006. Red arrowheads point to endostromatolites (crusts); thick black arrowheads
are upward-oriented; scale bars: white = 1 mm, black = 250 µm. |
As
documented above, sparite crystals forming gravitational cements, whether they
are 1) fibrous in shape and the signature of a vadose marine setting (e.g.,
in the French sample, Figs. 3.C ![]() , 4
, 4 ![]() ) or
2) dog-tooth shaped and the signature of
a vadose continental setting (e.g., in the Ukrainian sample, Figs.
3.D
) or
2) dog-tooth shaped and the signature of
a vadose continental setting (e.g., in the Ukrainian sample, Figs.
3.D ![]() , 6
, 6 ![]() ), are hyaline/translucent. In contrast, sparitic biocrystals that form
bioclasts are commonly colored by organic matter, which is also the case of
sparitic crystals forming colored crusts in some rock cavities and that should
be assigned a biogenic origin. Such individual colored crusts are here called
endostromatoids whereas the assembly of colored sparitic and/or clotted micritic
crusts in a similar setting is called endostromatolite, not cement. Besides
being colored, contrary to the crystals forming gravitational cements that are
always hyaline, endostromatolitic biocrystals are not necessarily growing
gravitationally as documented herein (e.g., Fig. 5
), are hyaline/translucent. In contrast, sparitic biocrystals that form
bioclasts are commonly colored by organic matter, which is also the case of
sparitic crystals forming colored crusts in some rock cavities and that should
be assigned a biogenic origin. Such individual colored crusts are here called
endostromatoids whereas the assembly of colored sparitic and/or clotted micritic
crusts in a similar setting is called endostromatolite, not cement. Besides
being colored, contrary to the crystals forming gravitational cements that are
always hyaline, endostromatolitic biocrystals are not necessarily growing
gravitationally as documented herein (e.g., Fig. 5 ![]() ). Because no specific
laboratory investigation of the organic contents of such endostromatolites has
hitherto been attempted, it is felt that the features given above should serve
as a preliminary guide to distinguish between endostromatolites (e.g.,
"gravitational cement" in Flügel, 2004: Pl. 34, fig. 6) and
genuine gravitational cement
(e.g., "pendant (or gravitational) cement" in Flügel,
2004: Pl. 33, fig. 4).
). Because no specific
laboratory investigation of the organic contents of such endostromatolites has
hitherto been attempted, it is felt that the features given above should serve
as a preliminary guide to distinguish between endostromatolites (e.g.,
"gravitational cement" in Flügel, 2004: Pl. 34, fig. 6) and
genuine gravitational cement
(e.g., "pendant (or gravitational) cement" in Flügel,
2004: Pl. 33, fig. 4).
The manuscript also benefited from the constructive reviews of Bogusław Kołodziej and George Pleș. The author would like to thank Phil Salvador for his appreciated help with the original (English) text and Robert W. Scott for the final touch. He would like to acknowledge the support of Alin Oprișa, Volodymyr Khomyn, Lev Monchak, and Natalia Shkolna for fieldwork in Ukraine (2019), and that of Zinat Hasanzade, Javad Pakravan, and Morteza Taherpour Khalil Abad for fieldwork in Iran (2017).
Alth A. von (1878).- O galicyjskich gatunkach skamieniałych otwornie rodzaju Gyroporella Gümb.- Rozprawy i Sprawozdania z Posiedzeń Wydziału Matematyczno-Przyrodniczego Akademii Umiejętności, Krakowie, t. V, p. 71-112 (Pls. VI-VII).
Alth A. von (1882).- Die versteinerungen des Niźniower Kalksteines.- Beiträge zur Paläontologie Österreich-Ungarns und des Orients, Wien, Band I, p. 183-332.
Amodio S., Barattolo F. & Riding R. (2018).- Early Cretaceous dendritic shrub-like fabric in karstified peritidal carbonates from southern Italy.- Sedimentary Geology, vol. 373, p. 134-146.
Arnaud H. (1981).- De la plate-forme urgonienne au bassin vocontien : Le Barrémo-Bédoulien des Alpes occidentales entre l'Isère et le Büech (Vercors méridional, Diois oriental et Dévoluy).- Thèse, Grenoble; Géologie Alpine, Grenoble, Mémoire H.S., no. 12, 804 p.
Arnaud-Vanneau A. (1980).- Micropaléontologie, paléoécologie et sédimentologie d'une plate-forme carbonatée de la marge passive de la Téthys : L'Urgonien du Vercors septentrional et de la Chartreuse (Alpes occidentales).- Thèse, Grenoble; Géologie Alpine, Grenoble, Mémoire H.S., no. 11, 874 p.
Belkhedim S., Munnecke A., Benhamou M., Nemra A. & Sadji R. (2019).- Challenging asymmetric cements as indicators of vadose diagenesis: "pseudo-gravitational" cements from the lower Pliensbachian of the Traras Mountains in NW Algeria.- Facies, Erlangen, vol. 65, no. 2, paper 12, 23 p.
Böker E. (unpublished, 1994).- Interprétation des milieux de dépôt et recherche des discontinuités sédimentaires dans une coupe de la Haute-Chaîne à la limite Berriasien-Valanginien.- Mémoire de Diplôme d'Ingénieur-Géologue, Université de Genève, 43 p. (31 Pls.).
Bøggild O.B. (1930).- The shell structure of the mollusks.- Det Kongelige Danske Videnskabernes Selskabs Skrifter, Naturvidenskabelige og Mathematiske Afdeling, Kjobenhavn, Raekke 9, II.2, p. 231-325. (Pls. IXV).
Brehm U., Palinska K.A. & Krumbein W.E. (2004).- Laboratory cultures of calcifying biomicrospheres generate ooids - A contribution to the origin of oolites.- Carnets Geol., Madrid, vol. 4, no. L03, 6 p. DOI: 10.4267/2042/309
Cuif J.-P., Dauphin Y., Luquet G., Medjoubi K., Somogyi A. & Perez-Huerta A. (2018).- Revisiting the organic template model through the microstructural study of shell development in Pinctada margaritifera, the Polynesian pearl oyster.- Minerals, Basel, vol. 8, paper 370, 20 p.
Cushman A. & Głazewski K. (1949).- 324. Jurassic Foraminifera fro the Nizniow Limestone of Podole, Poland.- Contributions from the Cushman Laboratory for Foraminiferal Research, Sharon, vol. 25, part 1, p. 1-11.
Dangeard L. (1936).- Étude des calcaires par coloration et décalcification. Application à l'étude des calcaires oolithiques.- Bulletin de la Société géologique de France (5e Série), Paris, t. VI, p. 237-246 (Pl. XVI).
Dauphin Y. (2005).- Biomineralization. In: King R.B. (Ed.), Encyclopedia of inorganic chemistry, Second edition, Volume I.- John Wiley & Sons, Chichester, p. 391-404.
Dauphin Y. (2016).- Biomineralization. In: Encyclopedia of inorganic and bioinorganic chemistry, Online.- John Wiley & Sons, Chichester, 15 p.
Dias-Brito D. (ed.), Granier B., Mettraux M. & Brito H. (2017).- Guia petrográfico dos carbonatos do Brasil.- Obra 4, UNESPetro, Rio Claro, 232 p.
Diaz M.R. & Eberli G.P. (2019).- Decoding the mechanism of formation in marine ooids: A review.- Earth-Science Reviews, vol. 190, p. 536-556.
Dunham R.J. (1962).- Classification of carbonate rocks according to depositional texture. In: Ham W.E. (ed.), Classification of carbonate rocks.- American Association of Petroleum Geologists, Memoir, Tulsa - OK, vol. 1, p. 108-121.
Flügel E. (1982).- Microfacies analysis of limestones.- Springer-Verlag, Berlin, 633 p.
Flügel E. (2004).- Microfacies of carbonate rocks. Analysis, interpretation and application.- Springer-Verlag, Berlin, xxxviii + 976 p.
Flügel E. (2010).- Microfacies of carbonate rocks. Analysis, interpretation and application.- 2nd edition, Springer-Verlag, Berlin, xxiii + 984 p.
Głażewski K. (1937).- Jura w Horodence i rozmieszczenie jury na Podolu.- Annales Societatis Geologorum Poloniae, Kraków, tom XIII, p. 98-103.
Gorzelak P., Stolarski J., Mazur M. & Meibom A. (2012).- Micro- to nanostructure and geochemistry of extant crinoidal echinoderm skeletons.- Geobiology, vol. 11, no. 1, p. 29-43.
Granier B. (1987).- Le Crétacé inférieur de la Costa Blanca entre Busot et Altea, Alicante (Espagne) : Biostratigraphie, sédimentologie, évolution tectono-sédimentaire.- PhD Thesis, Université Pierre et Marie Curie, Paris; Mémoires des Sciences de la Terre, no. 87-49, 281 p. (54 Pls.)
Granier B. (2012).- Bacinella, a discrete type of Mesozoic calcimicrobial structures. In: Gawlick H.J. & Missoni S. (chairpersons), Sedimentology in the hearth of the Alps.- CD-ROM, 29th IAS Meeting of Sedimentology, 10th-13th September, Schladming, p. 91 (abstract).
Granier B. (2017).- New stratigraphic and genetic model for the dolomitic Cretaceous Pinda reservoirs in Angola. Part I - The Pinda of Angola, an integrated lithostratigraphic approach.- Carnets Geol., Madrid, vol. 17, no. 4, p. 105-127. DOI: 10.4267/2042/62283
Granier B. (2019a).- Dual biozonation scheme (benthic foraminifera and "calcareous" green algae) over the Jurassic-Cretaceous transition. Another plea to revert the system boundary to its historical Orbigny's and Oppel's definition.- Cretaceous Research, vol. 93, p. 245-274.
Granier B. (2019b).- Observations on some Actinoporellas (Chlorophyta, Polyphysaceae). Revision of the Jacques Emberger Collection. Part 1.- Carnets Geol., Madrid, vol. 19, no. 7, p. 113-139. DOI: 10.4267/2042/70196
Granier B. (2020).- Introduction to thematic issue, "The transition of the Jurassic to the Cretaceous: an early XXIth century holistic approach".- Cretaceous Research, vol. 114, article 104530, 8 p.
Granier B., Clavel B. & Charollais J. (2016).- Comments on "Estimating the impact of early diagenesis on isotope records in shallow-marine carbonates: A case study from the Urgonian platform in western Swiss Jura" by A. Godet et al. [Palæogeography Palæoclimatology Palæoecology 454 (2016) 125-138].- Carnets Geol., Madrid, vol. 16, no. 17, p. 417-429. DOI: 10.4267/2042/61385
Granier B. & Lethiers A. (2019).- Aloisalthella, a new genus of fossil Polyphysacean green algae (Chlorophyta, Dasycladales), with notes on the genus Clypeina (Michelin, 1845).- Palaeontologia Electronica, no. 22.2.45A, 20 p. DOI: 10.26879/923
Granier B., Virgone A., Busnardo R. & Bulot L.G. (1995).- Des calpionelles dans l'Hauterivien supérieur. Découverte exceptionnelle à Busot (Alicante, Espagne).- Comptes-Rendus de l'Académie des Sciences (série II a), Paris, t. 321, p. 1179-1186.
Horowitz A.S. & Potter P.E. (1971).- Introductory petrography of fossils.- Springer-Verlag, Berlin, 302 p.
Kalkowsky E. (1908).- 3. Oolith und stromatolith im norddeutschen Buntsandstein.- Zeitschrift der deutschen geologischen Gesellschaft, Berlin, Band 60, Heft I, p. 68-125 (Pls. IV-XI). URL: https://archive.org/details/zeitschriftderd601908deut
Majewske O.P. (1969).- Recognition of Invertebrate fossil fragments in rocks and thin sections.- International Sedimentary Petrographical Series, Leiden, vol. XIII, 101 p.
Miík M. (1966).- Microfacies of the Mesozoic and Tertiary Limestones of the West Carpathians.- Slovenská Akadémia Vied, Bratislava, 280 p.
Monty C.L.V. (1980).- Miocene stromatolitic biostromes, Benejuzar, S.E. Spain.- Résumés, 26e Congrès Géologique International, Paris (7-17 juillet 1980), vol. II, section 6, p. 522.
Monty C.L.V. (1982).- Cavity and fissure dwelling stromatolites (endostromatolites) from Belgian Devonian mud mounds.- Annales de la Société géologique de Belgique, Bruxelles, vol. 105, p. 343-344.
O'Reilly S.S., Mariotti G., Winter A.R., Newman S.A., Matys E.D., McDermott F., Pruss S.B., Bosak T., Summons R.E. & Klepac-Ceraj V. (2017).- Molecular biosignatures reveal common benthic microbial sources of organic matter in ooids and grapestones from Pigeon Cay, The Bahamas.- Geobiology, vol. 7, no. 1, p. 107-130.
Samuel O., Borza K. & Köhler E. (1972).- Microfauna and lithostratigraphy of the Paleogene and adjacent Cretaceous of the middle Váh Valley (West Carpathian).- Geologický ústav Dionýza túra, Bratislava, 246 p. (CLXXX Pls.)
Stolarski J. & Mazur M. (2005).- Nanostructure of biogenic versus abiogenic calcium carbonate crystals.- Acta Palaeontologica Polonica, Warszawa, vol. 50, no. 4, p. 847-865.
Tucker M.E. (1990).- 3.8.1 Skeletal carbonates. In: Briggs D.E.G. & Crowther P.R. (eds.), Palaeobiology. A synthesis.- Blackwell Scientific Publications, Oxford, p. 247-250.