◄ Carnets Geol. 21 (7) ►
![]()
Sommaire
[1. Introduction]
[2. Contexte géologique et transition Jurassique/Crétacé dans la région du Mont Liban]
[3. Étude paléontologique (Y.A.)]
[4. Répartition et âge de la faune de brachiopodes de Zeghrine]
[5. Conclusions]
[Références bibliographiques]
et ...
[Planche]
29 impasse des mésanges, F-01700, Beynost (France)
6D avenue Général de Gaulle, F-05100 Briançon (France)
2 impasse Charles Martel, 29217 Plougonvelin (France)
17 rue Jean Baptiste Perret, F-69660 Collonges-au-Mont-d'Or (France)
Manuscrit en ligne depuis le 24 mars 2021
DOI 10.2110/carnets.2021.2107
![]()
[Éditeur scientifique : Michel Moullade ; éditeur technique : Bruno R.C. Granier]
![]()
Les gisements jurassiens français ou suisses du Crétacé basal (Valanginien ou Hauterivien) recčlent de nombreuses espčces de brachiopodes parmi lesquelles 3 espčces inconnues au Liban : Lamellaerhynchia desori (Loriol in Pictet & Campiche, 1872), Sulcirhynchia valangiensis (Loriol, 1864) et Terebratulina arzierensis (Loriol, 1864). Le site fossilifčre de la Formation de Salima ŕ Zeghrine, une localité proche de Bikfaya (Mont Liban), recčle une association constituée de Belothyris pseudojurensis (Leymerie, 1842), Lamellaerhynchia hauteriviensis Burri, 1953, Loriolithyris valdensis (Loriol, 1868), Lor. latifrons (Pictet, 1872), Sellithyris carteroniana (Orbigny, 1847) et Terebratulina biauriculata Orbigny, 1850, toutes également présentes dans les localités jurassiennes. Sur la base de l'étude de son association de brachiopodes, la Formation de Salima est par conséquent attribuée au Valanginien indifférencié.
• Liban ;
• Salima ;
• Bikfaya ;
• Jurassique ;
• Crétacé ;
• Valanginien ;
• brachiopodes
Alméras Y., Ferry S., Granier B.R.C. & Merran Y. (2021).- Brachiopodes jurassiens de l'intervalle Valanginien - Hauterivien. Leur contribution ŕ la datation de la Formation de Salima au Mont Liban.- Carnets Geol., Madrid, vol. 21, nº 7, p. 163-179.
Jurassian brachiopods of the Valanginian - Hauterivian interval. Their contribution to the dating of the Salima Formation in Mount Lebanon.- The Jurassian French or Swiss outcrops of the lowermost Cretaceous (Valanginian or Hauterivian) yield numerous brachiopod species including 3 species: Lamellaerhynchia desori (Loriol in Pictet & Campiche, 1872), Sulcirhynchia valangiensis (Loriol, 1864), and Terebratulina arzierensis (Loriol, 1864), which are not found in Lebanon. The fossiliferous site of the Salima Formation at Zeghrine, a locality close to Bikfaya (Mount Lebanon), yields an assemblage made of Belothyris pseudojurensis (Leymerie, 1842), Lamellaerhynchia hauteriviensis Burri, 1953, Loriolithyris valdensis (Loriol, 1868), Lor. latifrons (Pictet, 1872), Sellithyris carteroniana (Orbigny, 1847), and Terebratulina biauriculata Orbigny, 1850; all are also present in Jurassian localities. On the basis of the study of its brachiopod assemblage, the Salima Formation is therefore ascribed an undifferentiated Valanginian age.
• Lebanon;
• Salima;
• Bikfaya;
• Jurassic;
• Cretaceous;
• Valanginian;
• Brachiopoda
L'étude des faunes de brachiopodes présentée ici a été envisagée afin de répondre ŕ un questionnement concernant l'âge d'une formation géologique au Liban. Aprčs un résumé de l'état actuel des connaissances stratigraphiques sur ce pays du Proche-Orient au passage Jurassique/Crétacé, l'étude paléontologique essentiellement axée sur les faunes des gisements jurassiens français ou suisses du Crétacé basal (Valanginien ou Hauterivien) sert d'introduction ŕ une discussion sur l'âge de la Formation de Salima et sur sa position clef dans le contexte géologique régional.
La coupe-type de la
Formation de Salima (Figs. 1 ![]() -
2
-
2 ![]() ) ou J7 des cartes géologiques du Liban, située
ŕ Zeghrine, une localité proche de Bikfaya (Gouvernorat du Mont Liban), et décrite
ŕ l'origine par Wetzel et Dubertret
(1951), a ensuite été
revisitée par Toland (2000), Noujaim-Clark et BouDagher-Fadel
(2001), Ferry et al.
(2007), Granier et al. (2016)
et Maksoud et al. (2020). Granier et al.
(2016)
amendent significativement cette unité lithostratigraphique en la restreignant
ŕ son seul "membre supérieur" calcaire, supposé crétacé et plus
précisément valanginien inférieur (Fig. 5
) ou J7 des cartes géologiques du Liban, située
ŕ Zeghrine, une localité proche de Bikfaya (Gouvernorat du Mont Liban), et décrite
ŕ l'origine par Wetzel et Dubertret
(1951), a ensuite été
revisitée par Toland (2000), Noujaim-Clark et BouDagher-Fadel
(2001), Ferry et al.
(2007), Granier et al. (2016)
et Maksoud et al. (2020). Granier et al.
(2016)
amendent significativement cette unité lithostratigraphique en la restreignant
ŕ son seul "membre supérieur" calcaire, supposé crétacé et plus
précisément valanginien inférieur (Fig. 5 ![]() ) dans l'une de leurs trois hypothčses
de travail, i.e., leur option favorite "A" dans leur Fig. 8 (op.
cit.). Son "membre inférieur" marneux est alors, quant ŕ
lui, attribué au Jurassique et plus précisément au Tithonien comme
l'avaient déjŕ pressenti Bischoff (1964,
1990a, 1990b) et
correctement indiqué Toland (2000), et non au Berriasien comme erronément
rapporté par Noujaim-Clark et BouDagher-Fadel
(2001). En se
fondant sur une récolte récente de brachiopodes de la Formation de Salima, Maksoud
et al. (2020) confirment son attribution au Valanginien ; toutefois ils
optent pour un âge Valanginien supérieur alors que Granier et al.
(2016) faisaient plutôt l'hypothčse d'un âge Valanginien inférieur. Ŕ
la lueur de datations palynologiques récentes des Marnes d'Arzier en Suisse (Monteil
in Jan du Chęne et al., 2016), une formation connue pour
ses gisements historiques de brachiopodes (e.g., Loriol, 1862,
1864, 1868, 1872), l'étude de matériel libanais inédit provenant d'une récolte
plus ancienne effectuée par deux d'entre nous (S.F., Y.M.) nous permet
aujourd'hui de revenir sur l'attribution par Maksoud et al.
(2020) de la Formation de Salima amendée au seul Valanginien supérieur, une
attribution sensée ętre mieux renseignée et plus précise.
) dans l'une de leurs trois hypothčses
de travail, i.e., leur option favorite "A" dans leur Fig. 8 (op.
cit.). Son "membre inférieur" marneux est alors, quant ŕ
lui, attribué au Jurassique et plus précisément au Tithonien comme
l'avaient déjŕ pressenti Bischoff (1964,
1990a, 1990b) et
correctement indiqué Toland (2000), et non au Berriasien comme erronément
rapporté par Noujaim-Clark et BouDagher-Fadel
(2001). En se
fondant sur une récolte récente de brachiopodes de la Formation de Salima, Maksoud
et al. (2020) confirment son attribution au Valanginien ; toutefois ils
optent pour un âge Valanginien supérieur alors que Granier et al.
(2016) faisaient plutôt l'hypothčse d'un âge Valanginien inférieur. Ŕ
la lueur de datations palynologiques récentes des Marnes d'Arzier en Suisse (Monteil
in Jan du Chęne et al., 2016), une formation connue pour
ses gisements historiques de brachiopodes (e.g., Loriol, 1862,
1864, 1868, 1872), l'étude de matériel libanais inédit provenant d'une récolte
plus ancienne effectuée par deux d'entre nous (S.F., Y.M.) nous permet
aujourd'hui de revenir sur l'attribution par Maksoud et al.
(2020) de la Formation de Salima amendée au seul Valanginien supérieur, une
attribution sensée ętre mieux renseignée et plus précise.
|
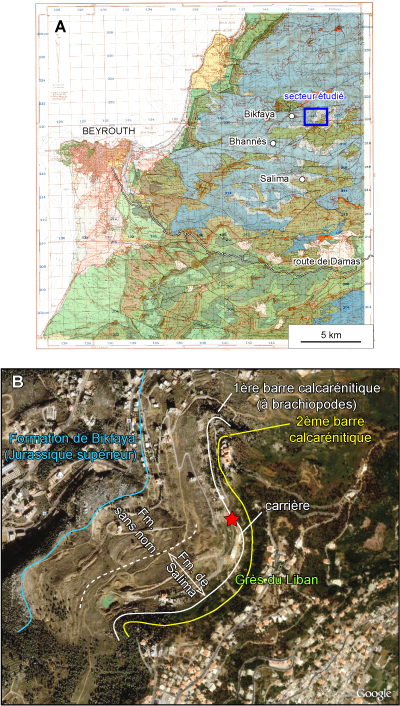
Figure 1
:
Localisation de la coupe de Zeghrine : A) reproduction de la carte géologique
de Beyrouth au 50.000e (Dubertret, 1951) avec la position de
localités (Bhannčs, Bikfaya et Salima) ayant donné leur nom ŕ une unité
lithostratigraphique (respectivement notées ßJ6, J6a et J7). Bkâatouta, qui a
donné son nom ŕ une nouvelle unité, membre inférieur des Grčs du Liban (notés
C1, est hors champ et ne figure pas sur cette carte) ; B) imagerie satellitaire
de la Formation de Salima sensu Dubertret dans sa localité-type,
ici divisée en une formation sans nom pour sa partie inférieure et la
Formation de Salima amendée pour sa partie supérieure. Le tracé bleu marque
le toit des calcaires micritiques blancs de la Formation de Bikfaya, le tracé
blanc en pointillé la position approximative du toit de la formation sans nom,
le tracé blanc continu le toit des calcaires oolithitiques de la Formation de
Salima amendée, le tracé jaune le toit des calcarénites du Membre de Bkâatouta,
unité lithostratigraphique nouvellement définie ŕ la base des Grčs du Liban
(Figs. 2.A-C |
|
Figure 2
:
La coupe de Zeghrine : A) coupe lithologique ; B) vue panoramique de la coupe ;
C), vue des deux barres calcarénitiques orangées avec en 1 le toit de la
Formation de Salima et en 2 le toit du Membre de Bkâatouta, unité
lithostratigraphique nouvellement définie ŕ la base des Grčs du Liban (Fig.
3 |
|
Figure 3
:
Toit des calcarénites du Membre de Bkâatouta, unité nouvellement définie ŕ
la base des Grčs du Liban, ici sur la coupe de Zeghrine. A) vue du passage des
calcaires oolithiques aux grčs. Des brčches ŕ galets anguleux sont intercalées
dans les grčs ŕ stratifications obliques. B) détail de la partie sommitale de
ce membre avec une colonie de coraux branchus. |
Ordre Rhynchonellida Kuhn, 1949
Super-famille Rhynchonellacea Gray, 1848
Famille Rhynchonellidae Gray, 1848
Sous-famille Cyclothyridinae Makridin, 1955
Genre Sulcirhynchia Burri, 1953
1953. Sulcirhynchia n. gen., Burri, p. 271.
1956. Sulcirhynchia Burri, Burri, p. 642.
1956. Sulcirhynchia Burri, Owen, p. 168.
1965. Sulcirhynchia Burri, Ager, p. H619.
1973. Sulcirhynchia Burri, Owen, p. 124.
Espčce-type : Rhynchonella valangiensis Loriol, 1864.
Sulcirhynchia valangiensis (Loriol, 1864)
(Pl. 1 ![]() , figs.
1-3)
, figs.
1-3)
1864. Rhynchonella valangiensis de Loriol, p. 442 ; Pl., figs. 14-17.
1868. Rhynchonella valangiensis de Loriol, Loriol, p. 57 ; Pl. 5, figs. 6-8.
1872. Rhynchonella valangiensis de Loriol, Pictet & Campiche, p. 14 ; Pl. 195, figs. 9-12.
1913. Rhynchonella valangiensis de Loriol, Jacob & Fallot, p. 51 ; Pl. 7, figs. 1-14.
? 1930. Rhynchonella valangiensis de Loriol, Roch, p. 321 et 348.
1953. Sulcirhynchia valangiensis (de Loriol), Burri, p. 272 ; Pl. 15, fig. 1.
1954. Sulcirhynchia valangiensis (de Loriol), Burri, p. 419-425.
1956. Sulcirhynchia valangiensis (de Loriol), Burri, p. 643 ; Pl. 6, fig. 1.
? 1960. Sulcirhynchia valangiensis (de Loriol), Popkhadzé, p. 147 ; Pl. 1, figs. 1-2.
Lectotype : Loriol, 1864, Pl., fig. 14. Marnes d'Arzier (Valanginien inférieur), Arzier (carričre de La Violette), canton de Vaud (Jura suisse) (désignation de Burri, 1956).
Matériel étudié. Arzier : 50 topotypes (coll. Alméras, FSL 706678 et 706679).
Morphologie. Les 50 topotypes d'Arzier
examinés comprennent des coquilles longues de 4,7 ŕ 17,3 mm, de contour
triangulaire, le plus souvent aussi large que long. Elles sont assez épaisses
et antérieurement bilobées (Pl. 1 ![]() , figs. 1-3). En effet, au stade adulte, leur
bord frontal sinueux est profondément échancré au milieu des deux valves (Pl. 1
, figs. 1-3). En effet, au stade adulte, leur
bord frontal sinueux est profondément échancré au milieu des deux valves (Pl. 1 ![]() ,
figs. 2-3). Cette échancrure, caractéristique de l'espčce, est ŕ peine ébauchée
ou męme absente (Pl. 1
,
figs. 2-3). Cette échancrure, caractéristique de l'espčce, est ŕ peine ébauchée
ou męme absente (Pl. 1 ![]() , fig. 1) chez les jeunes individus au contour arrondi
plus régulier. Valve dorsale plus bombée que la valve ventrale avec convexité
maximale en son milieu. Les spécimens jeunes (jusqu'ŕ une longueur d'environ
10 mm) demeurent rectimarginés. Commissures latérales des valves droites, se
relevant brusquement ŕ angle droit ŕ proximité du front, d'oů une
uniplication frontale s'accentuant au cours de la croissance des coquilles. La
morphogenčse de S. valangiensis a été décrite et figurée par Burri
(1954, Fig. 1, et 1956, Fig. 10).
, fig. 1) chez les jeunes individus au contour arrondi
plus régulier. Valve dorsale plus bombée que la valve ventrale avec convexité
maximale en son milieu. Les spécimens jeunes (jusqu'ŕ une longueur d'environ
10 mm) demeurent rectimarginés. Commissures latérales des valves droites, se
relevant brusquement ŕ angle droit ŕ proximité du front, d'oů une
uniplication frontale s'accentuant au cours de la croissance des coquilles. La
morphogenčse de S. valangiensis a été décrite et figurée par Burri
(1954, Fig. 1, et 1956, Fig. 10).
La costulation apparaît
vers 4 ŕ 6 mm ŕ partir du sommet du crochet. Les coquilles de 6,5 ŕ 17,3 mm
sont ornées de 27 ŕ 41 côtes simples, rayonnantes, fines et réguličres sur
la région cardinale et devenant plus fortes et plus aiguës aux approches du
front. Le nombre moyen de côtes sur la valve dorsale, calculé sur 40 spécimens,
est de 32,2. Les individus les plus petits, de 4,7 ŕ 6,5 mm, sont moins densément
costés.
Le développement du
foramen et l'incurvation du crochet (de dimensions moyennes, non cręté et
dressé) sont décrits et figurés par Burri (1954, Fig. 2, et 1956, Fig.
11). L'évolution du foramen au cours de la croissance présente des exceptions
par rapport ŕ la description de Burri. C'est ainsi qu'ŕ des tailles
adultes approximativement identiques, on peut observer des foramens ourlés, de
contour triangulaire (Pl. 1 ![]() , fig. 2) ou ovale
(Pl. 1
, fig. 2) ou ovale
(Pl. 1 ![]() , fig. 3), les plaques
deltidiales demeurant séparées. La coquille plus petite de la Pl. 1
, fig. 3), les plaques
deltidiales demeurant séparées. La coquille plus petite de la Pl. 1 ![]() , fig. 1,
montre un foramen circulaire avec des plaques deltidiales ŕ peines réunies par leur base.
, fig. 1,
montre un foramen circulaire avec des plaques deltidiales ŕ peines réunies par leur base.
Caractčres internes. Voir Burri, 1953, Fig. 2, Burri, 1954, Fig. 6, et Burri, 1956, Fig. 9.
Extension verticale et répartition géographique. Espčce abondante dans
les Marnes valanginiennes d'Arzier, carričres d'Arzier (localité-type dans le
canton de Vaud). Pour Loriol (1864), S. valangiensis "paraît
caractériser des couches supérieures de l'étage Valanginien". Burri
(1956) confirme l'âge Valanginien supérieur, contrairement ŕ Burri
(1953) ("base du Valanginien"). L'incertitude sur l'âge des Marnes d'Arzier
est maintenant levée, celles-ci étant désormais datées du Valanginien inférieur
(Walter, 1989, Fig. 4 ; 1991, p. 296 ; Jan du Chęne et al.,
2016) et męme Hauterivien en ce qui concerne celui donné par Lamellaerhynchia)
alors que les Marnes ŕ bryozoaires sont plus récentes (Valanginien supérieur).
Valanginien supérieur
(= Marnes ŕ bryozoaires) de Sainte-Croix. Męme âge ŕ Villers-le-Lac
(Limonite), ŕ Censeau, ŕ Vesancy prčs de Gex et ŕ Metabief. Calcaires de l'Auberson
dans le Jura vaudois. La répartition de S. valangiensis est connue le
long de la Chaîne du Jura, depuis la région de Neuchâtel jusqu'aux environs
de Bellegarde (Jacob & Fallot, 1913). Présence éventuelle (ŕ
confirmer) en Géorgie et au Daghestan (Popkhadzé, 1960).
Sulcirhynchia
valangiensis n'a pas été trouvé ŕ Zeghrine, ŕ l'Est de Beyrouth
(Liban).
Genre Lamellaerhynchia Burri, 1953
1953. Lamellaerhynchia n. gen., Burri, p. 274.
1956. Lamellaerhynchia Burri, Burri, p. 652.
1964. Lamellaerhynchia Burri, Ager, p. 111.
1965. Lamellaerhynchia Burri, Ager, p. H617.
1973. Lamellaerhynchia Burri, Owen, p. 126.
1974. Lamellaerhynchia Burri, Lobatscheva, p. 72.
1975. Lamellaerhynchia Burri, Calzada Badia, p. 15, 17.
Espčce-type : Terebratula rostriformis (Roemer, 1836) (= Terebratula multiformis Roemer, 1839 (partim !)).
Lamellaerhynchia desori (Loriol in Pictet & Campiche, 1872)
(Pl. 1 ![]() , fig. 4)
, fig. 4)
1872. Rhynchonella desori de Loriol, Pictet & Campiche, p. 16 ; Pl. 196, figs. 1-2.
1887. Rhynchonella desori de Loriol, Mallada, p. 151 ; Pl. 51, figs. 9-13.
1953. Lamellaerhynchia desori (de Loriol), Burri, p. 277 ; Pl. 15, fig. 3.
1956. Lamellaerhynchia desori (de Loriol), Burri, p. 657 ; Pl. 6, figs. 2-3.
? 1972. Monticlarella desori (Pictet), Smirnova, p. 28 ; Pl. 1, fig. 6.
Lectotype : Loriol in Pictet & Campiche, 1872, Pl. 196, fig. 2. Valanginien de Villers-le-Lac (désignation de Burri, 1953).
Matériel étudié. 10
ex. des Marnes d'Arzier (FSL 706680) (Valanginien inférieur) dont celui de la Pl. 1 ![]() , fig. 4, comparables au lectotype par leurs petites dimensions (longueur
ne dépassant pas 10 mm), leur uniplication frontale, les caractčres du crochet
non cręté, subdressé ŕ dressé et du petit foramen ovale séparant ŕ sa
base les plaques deltidiales. 12 ŕ 18 côtes simples sur la valve dorsale (12
ŕ 20 côtes selon Burri, 1956). Nvd moyen = 15,3.
, fig. 4, comparables au lectotype par leurs petites dimensions (longueur
ne dépassant pas 10 mm), leur uniplication frontale, les caractčres du crochet
non cręté, subdressé ŕ dressé et du petit foramen ovale séparant ŕ sa
base les plaques deltidiales. 12 ŕ 18 côtes simples sur la valve dorsale (12
ŕ 20 côtes selon Burri, 1956). Nvd moyen = 15,3.
Extension verticale
et répartition géographique. Espčce abondante dans
les Marnes d'Arzier (Valanginien inférieur), plus rare dans les Marnes ŕ Astieria
(Valanginien supérieur). Jura suisse (Arzier, Sainte-Croix) et Jura français
(Villers-le-Lac). Lamellaerhynchia desori a également été cité par
les auteurs dans les Marnes de l'Hauterivien inférieur. Cette extension
verticale s'explique (Burri, 1953) par le fait que les Marnes ŕ Astieria
étaient autrefois placées dans l'Hauterivien alors qu'elles sont actuellement
rapportées au Valanginien supérieur (Walter, 1989, Fig. 4). Il peut
aussi y avoir eu confusion avec des individus jeunes d'autres espčces de
l'Hauterivien. Présence possible dans le Valanginien de Crimée et d'Espagne (Mancha
Real).
Non observé ŕ Zeghrine.
Lamellaerhynchia hauteriviensis Burri, 1953
1953. Lamellaerhynchia hauteriviensis Burri, p. 279 ; Pl. 15, fig. 4.
1956. Lamellaerhynchia hauteriviensis Burri, Burri, p. 660 ; Pl. 8, figs. 1-2.
2020. Lamellaerhynchia hauteriviensis Burri, Maksoud et al., p. 6 ; Fig. 4.A-B.
Holotype : Burri, 1953, Pl. 15, fig. 4. Hauterivien inférieur (Zone ŕ Acanthodiscus radiatus), Le Landeron, Scheibenstand (S.A. Blatt, 134, coordonnées : 571,52/212,50).
Matériel étudié. Sainte-Croix : 6 ex. (coll. Alméras, FSL 706681). Zeghrine, ŕ l'Est de Beyrouth : 14 ex. (coll. Ferry, FSL 706682).
Morphologie. Espčce de dimensions
moyennes, de contour pentagonal arrondi plus large que long (Tabl. 1), avec
uniplication frontale débutant vers le milieu de la coquille. Cette
uniplication s'élčve au front au cours de la croissance, créant ainsi un
sinus ventral large, profond et bien délimité auquel correspond un pli médian
dorsal assez élevé (h/L = 0,47). La valve ventrale est faiblement renflée au
niveau de son crochet tandis que l'épaisseur maximale de la valve dorsale se
situe ŕ proximité immédiate ou au niveau du bord frontal.
Les côtes fortement
arrondies sont nombreuses : 18 ŕ 24 (Nvd moyen = 19,3 ŕ Sainte-Croix et 21,1
au Landeron). 5-6 côtes sur le pli dorsal (Tabl. 1). Quelques millimčtres sous
le crochet et l'umbo dorsal demeurent lisses. Crochet non cręté, subdressé,
quelquefois dressé. Foramen circulaire, plus rarement ovale. Plaques
deltidiales réunies et élevées (grande distance entre le foramen et le bord
cardinal).
Les calcaires
oolithiques de Zeghrine ont livré 14 coquilles uniplissées, de plus petites
dimensions, moins densément costées (Tabl. 1). Le stade postérieur lisse est
plus développé (tiers postérieur ŕ mi-longueur des coquilles) que chez L.
hauteriviensis du Landeron et de Sainte-Croix. Par contre, les caractčres
du crochet, du foramen et des plaques deltidiales sont identiques. A notre avis,
cette morphologie correspondrait au stade jeune du développement de L.
hauteriviensis dont les formes adultes se situent dans l'Hauterivien
inférieur.
Tableau 1 : Valeurs moyennes et intervalles de variation des dimensions et du nombre de côtes chez Lamellaerhynchia hauteriviensis Burri. Populations de la localité-type du Landeron, dans le Jura suisse occidental (données de Burri, 1953, Tabl. 3) (1), de Sainte-Croix (2) et de Zeghrine, au Liban (3).
| N | L | l | E | l/L | E/L | |
| 1 | 26 | 16,4 (14,2-19,3) | 18,1 (15,4-21,5) | 9,7 (7,3-11,4) | 1,10 (0,99-1,30) | 0,59 (0,47-0,73) |
| 2 | 6 | 15,2 (13,9-17,3) | 18,1 (16,1-20,1) | 8,9 (7,7-9,8) | 1,20 (1,16-1,21) | 0,58 (0,54-0,63) |
| 3 | 14 | 9,9 (7,0-11,3) | 10,9 (8,8 -12,2) | 5,9 (4,5-7,6) | 1,11 (0,99-1,26) | 0,60 (0,52-0,67) |
| h | ls | h/ls | h/L | Nvd | Nb |
| 21,1 (18-24) | |||||
| 7,2 (6,6-8,0) | 8,8 (7,6-10,2) | 0,83 (0,65-0,99) | 0,47 (0,44-0,54) | 19,3 (17- 22) | 5,3 (5-6) |
| 3,9 (2,5-5,5) | 7,5 (6,4-9,0) | 0,52 (0,39-0,64) | 0,39 (0,29-0,49) | 10,2 (9-11) | 4,4 (4-5) |
Caractčres internes. Voir Burri, 1953, Fig. 5, et Burri, 1956, Fig. 20.
Extension verticale
et répartition géographique. Hauterivien inférieur,
zone ŕ Acanthodiscus radiatus du Jura suisse et français. Le Landeron, bassin
N. Vingelz, environs de Twan, Le Cressier, Hauterive, Valangin, Villers-le-Lac,
l'Auberson, environs de Sainte-Croix et vallée du Nozon.
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban).
Ordre Terebratulida Waagen, 1883
Sous-ordre Terebratulidina Waagen, 1883
Super-famille Terebratulacea Gray, 1840
Famille Terebratulidae Gray, 1840
Sous-famille Sellithyridinae Muir-Wood, 1965
Genre Loriolithyris Middlemiss, 1968
1968. Loriolithyris gen. nov., Middlemiss, p. 176.
1975. Loriolithyris Middlemiss, Calzada Badia, p. 44.
1983. Loriolithyris Middlemiss, Calzada Badia, p. 143.
1983. Loriolithyris Middlemiss, Cooper, p. 200.
Espčce-type : Terebratula russillensis Loriol, 1866.
Loriolithyris valdensis (Loriol, 1868)
(Pl. 1 ![]() , figs.
7-12 ; Fig. 4
, figs.
7-12 ; Fig. 4 ![]() ; Tabl. 2)
; Tabl. 2)
1868. Terebratula valdensis de Loriol, p. 52 ; Pl. 4, figs. 9-12.
1872. Terebratula valdensis de Loriol in Pictet & Campiche, p. 66 ; Pl. 201, figs. 11-15.
? 1951. Terebratula valdensis de Loriol, Gigout, p. 360.
1966. Sellithyris valdensis (de Loriol), Bogdanova & Lobatcheva, p. 55 ; Pl. 5, fig. 7.
1968. Loriolithyris valdensis (de Loriol), Middlemiss, p. 182 ; Pl. A, fig. 5.
? 1972. Sellithyris valdensis (de Loriol), Smirnova, p. 81 ; Pl. 7, fig. 5.
1973. Loriolithyris valdensis (de Loriol), Dieni et al., p. 182 ; Pl. 36, figs. 9-10.
non 1974. Loriolithyris valdensis (de Loriol), Pajaud, p. 103 ; Pl. 2, fig. F.
1975. Loriolithyris valdensis (de Loriol), Calzada Badia, p. 45 ; Pl. 5, fig. 1.
non 1980. Loriolithyris valdensis (de Loriol), Middlemiss, p. 524 ; Pl. 55, figs. 5-9.
pars 1981. Loriolithyris valdensis (de Loriol), Middlemiss, p. 720.
1983. Loriolithyris valdensis (de Loriol), Cooper, Pl. 18, figs. 16-21 ; Pl. 64, figs. 23-24.
1984. Terebratula valdensis de Loriol, Bechon et al., Pl. 2, figs. 7, 9.
1988. Loriolithyris valdensis (de Loriol), Gaspard, p. 154 ; Pl. XVI, figs. 10-16 ; Pl. XVII, figs. 1-10. Figuration du lectotype.
Lectotype : Loriol, 1868, Pl. 4, fig. 9. Couche B du Valanginien inférieur des carričres d'Arzier (canton de Vaud) (désignation de Middlemiss, 1968, sans figuration de ce spécimen).
Matériel étudié. Arzier
(Vaud) : 17 ex. mesurés (Tabl. 2) (coll. Alméras, FSL 706683).
Villereversure (Ain), Calcaires ŕ Alectryonia : a) couche 2 base
: ex. Pl. 1 ![]() , figs. 7-10 ; 7 ex. mesurés (coll. Alméras, FSL 706684) ;
225 ex. déformés ou incomplčtement conservés, surtout des stades jeunes du développement
(coll. Alméras, FSL 706685) ; b) couche 2 sommet : 40 ex. mesurés
(coll. Alméras, FSL 706686) ; 95 ex. jeunes, déformés ou incomplets
(coll. Alméras, FSL 706687). Hauteville (Ain), carričre sous le
sanatorium de la Savoie : Couches ŕ Alectryonia (Valanginien supérieur),
2 ex., coll. Walter, FSL 706688).
, figs. 7-10 ; 7 ex. mesurés (coll. Alméras, FSL 706684) ;
225 ex. déformés ou incomplčtement conservés, surtout des stades jeunes du développement
(coll. Alméras, FSL 706685) ; b) couche 2 sommet : 40 ex. mesurés
(coll. Alméras, FSL 706686) ; 95 ex. jeunes, déformés ou incomplets
(coll. Alméras, FSL 706687). Hauteville (Ain), carričre sous le
sanatorium de la Savoie : Couches ŕ Alectryonia (Valanginien supérieur),
2 ex., coll. Walter, FSL 706688).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban)
: ex. Pl. 1 ![]() , figs. 11-12 ; 7 ex. jeunes ou incomplčtement conservés (coll. Ferry,
FSL 706689).
, figs. 11-12 ; 7 ex. jeunes ou incomplčtement conservés (coll. Ferry,
FSL 706689).
Morphologie. Les deux coquilles de Zeghrine, Pl. 1 ![]() , figs. 11-12, s'inscrivent parfaitement dans la morphogenčse de
Loriolithyris valdensis du Valanginien supérieur de
Villereversure, Pl. 1
, figs. 11-12, s'inscrivent parfaitement dans la morphogenčse de
Loriolithyris valdensis du Valanginien supérieur de
Villereversure, Pl. 1 ![]() , figs. 7-10. Les croissances en largeur et en épaisseur
de cette espčce dans les gisements de Villereversure et d'Arzier sont illustrés
sur la Figure 4
, figs. 7-10. Les croissances en largeur et en épaisseur
de cette espčce dans les gisements de Villereversure et d'Arzier sont illustrés
sur la Figure 4 ![]() . Celle-ci montre une faible dispersion des individus et permet de
relier les stades jeunes aux stades adultes du développement. 53 coquilles de
Villereversure ont fait l'objet d'une étude biométrique (Tabl. 2). 28
exemplaires de 3,3 ŕ 18,8 mm sont rectimarginés et leur contour est
subcirculaire (l/L = 0,85 ŕ 1,00). Les coquilles de plus grande taille, assez
épaisses, acquičrent un contour subpentagonal plus long que large, avec une
largeur maximale se situant vers le milieu de la longueur. La sulciplication
frontale apparaît dčs le tiers antérieur (Tabl. 2), créant sur la valve
dorsale deux plis saillants, arrondis, encadrant une dépression médiane
nettement creusée sur le bord antérieur des plus grands individus (Pl. 1
. Celle-ci montre une faible dispersion des individus et permet de
relier les stades jeunes aux stades adultes du développement. 53 coquilles de
Villereversure ont fait l'objet d'une étude biométrique (Tabl. 2). 28
exemplaires de 3,3 ŕ 18,8 mm sont rectimarginés et leur contour est
subcirculaire (l/L = 0,85 ŕ 1,00). Les coquilles de plus grande taille, assez
épaisses, acquičrent un contour subpentagonal plus long que large, avec une
largeur maximale se situant vers le milieu de la longueur. La sulciplication
frontale apparaît dčs le tiers antérieur (Tabl. 2), créant sur la valve
dorsale deux plis saillants, arrondis, encadrant une dépression médiane
nettement creusée sur le bord antérieur des plus grands individus (Pl. 1 ![]() , figs.
10, 12). La valve ventrale est antérieurement déprimée et faiblement relevée
en son milieu par un pli large et arrondi. Les commissures latérales,
subrectilignes sur leurs deux-tiers postérieurs, se relčvent fortement en
direction dorsale aux approches du front des spécimens adultes. Chez six
coquilles de 14,4 ŕ 19,5 mm, une planoplication ou une légčre uniplication
arrondie se substitue au stade sulciplissé.
, figs.
10, 12). La valve ventrale est antérieurement déprimée et faiblement relevée
en son milieu par un pli large et arrondi. Les commissures latérales,
subrectilignes sur leurs deux-tiers postérieurs, se relčvent fortement en
direction dorsale aux approches du front des spécimens adultes. Chez six
coquilles de 14,4 ŕ 19,5 mm, une planoplication ou une légčre uniplication
arrondie se substitue au stade sulciplissé.
Le crochet gros, non cręté,
court et dressé, est percé d'un grand foramen circulaire. Les plaques
deltidiales sont séparées chez les coquilles jeunes (mais pas uniquement) (27
exemplaires de Villereversure). Ces plaques se réunissent chez 26 autres
exemplaires en général de plus grandes dimensions. Le deltidium est alors
large mais presque toujours masqué. Enfin, les plaques deltidiales ne sont
visibles et élevées que chez deux spécimens de 16,5 et 22 mm.
Les exemplaires du Valanginien
d'Arzier possčdent les męmes caractéristiques. Leurs dimensions sont en
moyenne plus grandes étant donné le faible pourcentage de coquilles jeunes
mesurables. La situation de la naissance de la sulciplication frontale et la
profondeur de la dépression médiane dorsale sont comparables dans les deux
gisements d'Arzier et de Villereversure (Tabl. 2).
Loriolithyris
valdensis se distingue facilement
de Musculina acuta (Quenstedt) de l'Hauterivien par son crochet
court et recourbé, par son grand foramen et par son deltidium presque invisible
(crochet long et subdressé, deltidium élevé bien exposé chez M. acuta).
Tableau 2 : Valeurs moyennes et intervalles de variation des dimensions des coquilles chez Loriolithyris valdensis (Loriol). Populations d'Arzier (1) et de Villereversure (2). U1 et U2 = situation de la naissance de la sulciplication frontale ; p et d = profondeur et largeur du sinus médian dorsal. Autres paramčtres : voir Tabl. 1.
| N | L | l | E | l/L | E/L | |
| 1 | 17 | 17,0 (7,4-23,2) | 14,1 (6,5-18,2) | 8,9 (3,3-13,3) | 0,84 (0,75-0,91) | 0,52 (0,44-0,57) |
| 2 | 53 | 13,6 (3,3-25,0) | 11,3 (3,3-20,8) | 6,2 (1,5-13,0) | 0,84 (0,75-1,00) | 0,44 (0,36-0,57) |
| U1 | U2 | p | d | p/d | p/L | |
| 1 | 15,6 (9,0-18,4) | 0,79 (0,72-0,84) | 2,0 (0,5-3,0) | 6,6 (3,9-9,2) | 0,29 (0,12-0,41) | 0,10 (0,03-0,14) |
| 2 | 14,4 (9,6-19,7) | 0,78 (0,68-0,88) | 1,8 (0,7-3,0) | 5,4 (3,7-7,7) | 0,32 (0,13-0,53) | 0,09 (0,04 -0,16) |
Caractčres internes. Voir Bogdanova (1966, Fig. 19), Middlemiss (1968, Fig. 5) ainsi que Dieni et al. (1973, Fig. 7).
Extension verticale
et répartition géographique. Valanginien d'Arzier
(plus abondante dans la Couche B de Loriol, 1868 = Valanginien inférieur).
Valanginien supérieur de Sainte-Croix (Marnes ŕ bryozoaires), de
Villers-le-Lac (Doubs), Metabief et Vesancy, et Calcaires ŕ Alectryonia de
Hauteville (Ain). Présence ŕ confirmer dans le Valanginien de la Meseta
marocaine occidentale (arričre-pays de Casablanca, Mazagan et Safi) (Gigout,
1951). Valanginien supérieur du Centre-Est de la Sardaigne (Badde Funtana Morta,
Orosei) et du Levant espagnol (Cati, Castellón) (Calzada Badia,
1975).
Berriasien-Valanginien inférieur des Corbičres (Middlemiss,
1968).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban).
|
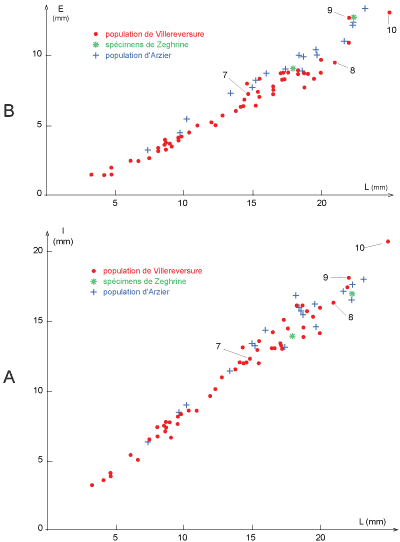
Figure 4
:
Dispersion des spécimens au cours de la croissance en largeur (A) et en épaisseur
(B) de Loriolithyris valdensis (Loriol). Coquilles d'Arzier
(croix bleues), de Villereversure, Ain (points rouges), et de Zeghrine, Liban
(astérisques vertes). Les numéros 7 ŕ 10 situent les exemplaires de
Villereversure représentés Pl. 1 |
Loriolithyris latifrons (Pictet, 1872)
(Pl. 1 ![]() , figs. 13-14)
, figs. 13-14)
1872. Terebratula latifrons Pictet, p. 67 ; Pl. 201, figs. 16-17.
1981. Loriolithyris russillensis latifrons (Pictet), Middlemiss, p. 718. Avec la synonymie.
2020. Loriolithyris latifrons (Pictet), Maksoud et al., p. 6 ; Fig. 4.C-F.
Lectotype : Pictet & Campiche, 1872, Pl. 201, fig. 16. Valanginien supérieur, Villers-le-Lac (Doubs) (désignation de Middlemiss, 1981).
Matériel étudié. Zeghrine, ŕ l'Est de Beyrouth (Liban) : 2 ex.,
Pl. 1 ![]() , figs. 13-14. La coquille
de la Pl. 1
, figs. 13-14. La coquille
de la Pl. 1 ![]() , fig. 14, est proche du lectotype, mais avec des dimensions un peu
inférieures. La sulciplication frontale n'apparaît qu'un peu plus tard au
cours de la croissance. Le spécimen de la Pl. 1
, fig. 14, est proche du lectotype, mais avec des dimensions un peu
inférieures. La sulciplication frontale n'apparaît qu'un peu plus tard au
cours de la croissance. Le spécimen de la Pl. 1 ![]() , fig. 13, représente un stade
jeune.
, fig. 13, représente un stade
jeune.
Extension verticale
et répartition géographique.
Valanginien supérieur de Villers-le-Lac (Doubs) et de Vesancy prčs de Gex (Ain).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban).
Genre Sellithyris Middlemiss, 1959
1959. Sellithyris Middlemiss, p. 113.
1965. Sellithyris Middlemiss, Muir-Wood, p. H793.
1966. Sellithyris Middlemiss, Bogdanova et Lobatcheva, p. 48.
1975. Sellithyris Middlemiss, Calzada Badia, p. 30.
1976. Sellithyris Middlemiss, Middlemiss, p. 39.
1983. Sellithyris Middlemiss, Cooper, p. 217.
Espčce-type : Terebratula sella J. de C. Sowerby, 1823.
Sellithyris carteroniana (Orbigny, 1847)
(Pl. 1 ![]() , fig. 15)
, fig. 15)
1847. Terebratula carteroniana d'Orbigny, p. 80 ; Pl. 507, figs. 1-5.
1981. Sellithyris carteroniana (d'Orbigny), Middlemiss, p. 711, avec la synonymie ŕ laquelle il faut ajouter :
1868. Terebratula carteroniana d'Orbigny, Loriol, p. 53.
1951. Terebratula carteroniana d'Orbigny, Gigout, p. 360 ; Pl. 9, figs. 11-14.
Holotype : Orbigny, 1847, Pl. 507, figs. 1-4. Valanginien, environs de Morteau (Doubs).
Matériel étudié.
Arzier (Vaud) : ex. Pl. 1 ![]() , fig. 15, et 4 autres spécimens (coll. Alméras,
FSL 706690).
, fig. 15, et 4 autres spécimens (coll. Alméras,
FSL 706690).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban) : 2 ex. en mauvais état de
conservation (coll. Ferry,
FSL 706691).
Famille Cancellothyrididae Thomson, 1926
Sous-famille Cancellothyridinae Thomson, 1926
Genre Terebratulina Thomson, 1926
1847. Terebratulina nov. gen. d'Orbigny, p. 56.
1851-1854. Terebratulina d'Orbigny, Davidson, p. 63.
1855. Terebratulina d'Orbigny, Deslongchamps E. & Deslongchamps E.E., p. 121.
1862-1885. Terebratulina d'Orbigny, Deslongchamps, p. 56.
1879. Terebratulina d'Orbigny, Douvillé, p. 15.
1965. Terebratulina d'Orbigny, Muir-Wood, p. H807.
Espčce-type : Anomia caputserpentis Linné, 1767, p. 1153 (non Linné, 1758, p. 703) = A. retusa Linné, 1758, p. 701.
Terebratulina arzierensis (Loriol, 1864)
(Pl. 1 ![]() , figs.
16-17)
, figs.
16-17)
1864. Terebratula (Terebratella) arzierensis Loriol, p. 441 ; Pl., figs. 18-20.
Lectotype : Loriol, 1864, Pl., fig. 12. Valanginien, carričres d'Arzier (Vaud).
Matériel étudié.
Arzier : ex. Pl. 1 ![]() , figs. 16-17 et 12 autres ex. (coll.
Alméras, FSL 706692).
, figs. 16-17 et 12 autres ex. (coll.
Alméras, FSL 706692).
Remarque. D'aprčs Pictet (in Pictet & Campiche, 1872), cette espčce est abondante dans la couche B du Valanginien inférieur des carričres d'Arzier.
Terebratulina biauriculata Orbigny, 1850
(Pl. 1 ![]() , figs.
18-19 ; Tabl. 3)
, figs.
18-19 ; Tabl. 3)
non 1841. Terebratula auriculata Roemer, p. 39 ; Pl. 7, fig. 9 (= Terebratulina chrysalis (Schlotheim, 1813), selon Schloenbach, 1866, p. 12, 65 ; voir aussi Siblik, 1963, p. 70).
1847. Terebratulina auriculata d'Orbigny, p. 58 ; Pl. 502, figs. 3-7.
1850. Terebratulina biauriculata d'Orbigny, p. 85 (avec référence au spécimen figuré par Orbigny, 1847, Pl. 502, figs. 3-7 (= T. auriculata) et refus de la mise en synonymie avec T. auriculata Roemer, 1841).
1863. (Terebratula) Terebratulina biauriculata d'Orbigny, Ooster, p. 34 ; Pl. 12, figs. 11-12.
1906. Terebratulina auriculata d'Orbigny, Ascher, p. 167 ; Pl. 14, fig. 13.
1907. Terebratulina biauriculata d'Orbigny, Karakasch, p. 218 ; Pl. 20, fig. 28.
1962. Terebratulina biauriculata d'Orbigny, Siblik, p. 277.
1963. Terebratulina biauriculata d'Orbigny, Siblik, p. 70 ; Pl. 2, fig. 2.
1974. Terebratulina biauriculata d'Orbigny, Pajaud, p. 95 et 112 ; Pl. 1, fig. A1-A2.
2020. Terebratulina biauriculata Orbigny, Maksoud et al., p. 6 ; Fig. 4.G-J.
Holotype : Orbigny, 1847, Pl. 502, figs. 3-7. Néocomien de La Couronne (Bouches-du-Rhône).
Matériel étudié. Zeghrine, ŕ l'Est de Beyrouth (Liban) : 36 ex. mesurés
(Tabl. 3) dont les 2
coquilles, Pl. 1 ![]() , figs. 18-19, et 19 ex. incomplčtement conservés et/ou déformés
(coll. Ferry, FSL 706693).
, figs. 18-19, et 19 ex. incomplčtement conservés et/ou déformés
(coll. Ferry, FSL 706693).
Remarque. Cette espčce a été créée par Orbigny (1847) sous le nom de Terebratulina auriculata. Elle a ensuite été désignée par le męme auteur (1850), et sans aucune justification notable, comme T. biauriculata. L'origine de ce changement réside en fait dans l'antériorité du nom d'auriculata détenue par Roemer (1841). Or, selon Schloenbach (1866), l'individu figuré sous ce nom par Roemer serait Terebratulina chrysalis (Schlotheim, 1813). Dans ces conditions, le nom valable pour les exemplaires figurés et décrits par Orbigny en 1847 est T. biauriculata (voir synonymie et également Siblik, 1963). Les spécimens de Zeghrine doivent donc ętre déterminés comme T. biauriculata.
Morphologie. Petites
coquilles déprimées, subcirculaires (Tabl. 3). Un seul exemplaire de 13 mm est
bien plus large que long (l/L = 1,12). Par comparaison avec la valve dorsale
presque plane, la valve ventrale est légčrement convexe dans sa moitié postérieure
; elle se creuse vers son quart antérieur d'une dépression large, peu profonde
et non délimitée latéralement ŕ laquelle correspond sur la valve dorsale un
faible renflement médian. Celui-ci détermine une commissure frontale large et
réguličrement arrondie, qui devient plus élevée chez les sujets âgés (Pl. 1 ![]() , fig. 19). Cette commissure frontale est également plus ou moins marquée
chez les petites coquilles jusqu'ŕ 8,3 mm (h/L = 0,12 ŕ 0,51). Seules trois
coquilles (sur 36) de 7,6-9,3 mm et 10,0 mm montrent une uniplication
naissante ŕ leur front. Les commissures latérales subrectilignes s'infléchissent
dorsalement ŕ l'approche du front uniplissé. Ornementation caractéristique
constituée de fines côtes rayonnantes dont certaines naissent par
intercalation ŕ des niveaux variés. Leur nombre sur le bord des coquilles est
compris entre 33 et 67 ; il augmenta avec la taille des spécimens (Nvd moyen =
44,9).
, fig. 19). Cette commissure frontale est également plus ou moins marquée
chez les petites coquilles jusqu'ŕ 8,3 mm (h/L = 0,12 ŕ 0,51). Seules trois
coquilles (sur 36) de 7,6-9,3 mm et 10,0 mm montrent une uniplication
naissante ŕ leur front. Les commissures latérales subrectilignes s'infléchissent
dorsalement ŕ l'approche du front uniplissé. Ornementation caractéristique
constituée de fines côtes rayonnantes dont certaines naissent par
intercalation ŕ des niveaux variés. Leur nombre sur le bord des coquilles est
compris entre 33 et 67 ; il augmenta avec la taille des spécimens (Nvd moyen =
44,9).
Crochet non cręté,
droit ŕ subdressé. Foramen circulaire (35 ex.). Un seul spécimen long de 14,9
mm possčde un foramen ovale, en contact avec l'umbo dorsal, les plaques
deltidiales demeurant séparées. Chez les autres spécimens, on observe des
plaques deltidiales séparées (8 ex. de 7,6 ŕ 14,9 mm), juste réunies par
leur base (7 ex. de 9,3 ŕ 11,9 mm) ou encore réunies (20 ex.) en un deltidium
peu élevé (9 ex. de 10,0 ŕ 13,5 mm) ou élevé (11 ex. de 9,3 ŕ 14,6 mm).
Tableau 3 : Valeurs moyennes et intervalles de variation des dimensions et du nombre de côtes chez Terebratulina biauriculata Orbigny de Zeghrine. U1 et U2 = situation de la naissance de l'uniplication frontale ; h et h/l = hauteur et hauteur relative du pli médian dorsal ; Nvd = nombre de côtes sur la valve dorsale. Autres paramčtres : voir Tabl. 1.
| L | l | E | l/L | E/L |
| 11,3 (7,6-14,9) | 10,7 (7,6-14,6) | 4,3 (2,6-7,5) | 0,94 (0,83-1,12) | 0,37 (0,26-0,55) |
| U1 | U2 | h | h/L | Nvd |
| 8,5 (6,9-10,4) | 0,74 (0,65-0,84) | 3,2 (1,0-6,6) | 0,27 (0,12-0,51) | 44,9 (33-67) |
Affinités. Dans
sa diagnose originale de Cruralina strambergensis du Valanginien
de Stramberg, Nekvasilovŕ (1978) admet une certaine ressemblance
morphologique avec Terebratulina biauriculata, en particulier avec le spécimen
de la Formation Hradistë d'Allemagne du Nord (château de Koniakau) figuré par
Ascher (1906, Pl. 14, fig. 13). Outre quelques différences dans les
structures internes (Nekvasilovŕ, 1978, Fig. 3), C. strambergensis
se différencie par son gros crochet plus court percé d'un trčs grand foramen
circulaire toujours en contact avec l'umbo dorsal ainsi que par l'absence
d'une uniplication frontale ŕ tous les stades du développement (les coquilles
de C. strambergensis sont toujours rectimarginées).
L'holotype de T.
biauriculata d'Alcide d'Orbigny n'a pas été retrouvé dans les
collections du Muséum National d'Histoire Naturelle de Paris (communication de
J. Drot ŕ Nekvasilovŕ, 1978). Un syntype est figuré par
RECOLNAT.
Les nombreux spécimens
des "Marno-calcaires ŕ Térébratulines" (Valanginien supérieur) de
l'arc de Castellane assurent l'identification et le statut de l'espčce (Cotillon
in Pajaud, 1974). Pajaud
(1974) place l'espčce dans ses
"Calcaires beiges berriasiens ŕ valanginiens" et Cotillon
(in Pajaud, 1974) dans le Valanginien supérieur. L'exemplaire de Gaspard
(1988)
est donné comme d'âge Barrémien.
Extension verticale
et répartition géographique. Valanginien supérieur
du Bassin du Sud-Est, France : La Couronne (Bouches-du-Rhône), "Marno-calcaires
ŕ Térébratulines" de l'arc de Castellane (Haute-Provence). Valanginien
des Alpes bernoises (Ooster, 1863), d'Allemagne du Nord (Ascher,
1906), de Crimée (Karakasch, 1907), des Carpathes centrales (Siblik,
1962), des Montagnes de Strŕzovsk en Tchéquie (Siblik,
1963).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban).
Sous-ordre Terebratellidina Muir-Wood, 1965
Super-famille Terebratellacea King, 1850
Famille Dallinidae Beecher, 1893
Sous-famille Kingeninae Elliott, 1948
Genre Belothyris Smirnova, 1960
1960. Belothyris gen. nov., Smirnova, p. 117.
1965. Belothyris Smirnova, Muir-Wood, p. H840.
1973. Belothyris Smirnova, Owen, p. 129.
Espčce-type : Belothyris plana Smirnova, 1960.
Belothyris pseudojurensis (Leymerie, 1842)
(Pl. 1 ![]() , figs. 20-22)
, figs. 20-22)
1842. Terebratula pseudojurensis Leymerie, p. 12 ; Pl. 15, figs. 5-6, ŕ l'exclusion de toute autre référence, exceptés les suivantes :
1907. Waldheimia pseudojurensis Leymerie, Deninger, p. 468.
1973. Belothyris pseudojurensis (Leymerie), Dieni et al., p. 203 ; Pl. 38, figs. 4-6 (sans la synonymie).
Synonymie. Les autres citations de pseudojurensis (Orbigny, 1847 ; Loriol, 1862, 1868 ; Walker, 1868 ; Pictet & Campiche, 1872 ; Davidson, 1874) ne se rapportent pas ŕ l'espčce de Leymerie. Il s'agirait plutôt de diverses espčces du genre Modestella Owen, avec en particulier M. gr. hippopoides (Pictet).
Holotype : Leymerie, 1842, Pl. 15, fig. 6. Hauterivien de Marolles (Aube). La désignation d'un néotype pour une espčce différente de pseudojurensis (Owen, 1970) n'est donc pas judicieuse.
Matériel étudié. Zeghrine, ŕ l'Est de Beyrouth (Liban) : ex.
Pl. 1 ![]() , figs. 20-22, et 35
exemplaires mesurés (coll. Ferry, FSL 706694).
, figs. 20-22, et 35
exemplaires mesurés (coll. Ferry, FSL 706694).
Valeurs moyennes et intervalles de variation des dimensions (en mm).
| L | l | E | l/L | E/L |
| 11,3 (8,9-14,0) | 9,2 (7,0-11,3) | 6,0 (4,5-7,6) | 0,82 (0,72-0,90) | E/L = 0,53 (0,46-0,62) |
Morphologie. Petites coquilles de 8,9
ŕ 14 mm, peu épaisses (E/L moyen = 0,53). Valves également et réguličrement
convexes en tous sens, avec le tiers antérieur de la valve dorsale dessinant
parfois un léger méplat médian. Contour triangulaire étroit (Pl. 1 ![]() , figs.
20-21) ou un peu plus large (Pl. 1
, figs.
20-21) ou un peu plus large (Pl. 1 ![]() , fig. 22), la largeur maximale se situant
vers le milieu de la longueur. Bord frontal tronqué (27 ex.) ou plus rarement
faiblement arrondi (8 ex.). Commissures toujours tranchantes, les valves se réunissant
suivant un angle aigu. Commissures latérales rectilignes. Commissure frontale
rectimarginée. Crochet court, non cręté, dressé. Foramen circulaire. Le
recourbement du crochet court masque le symphytium (8 ex.) ou bien celui-ci est
peu exposé (19 ex.). Les plaques deltidiales réunies en une pičce unique ne
sont visibles que sur 8 exemplaires. Le septum médian, visible ŕ travers le
test de 23 spécimens, s'étend sur la moitié de la longueur de la valve
dorsale.
, fig. 22), la largeur maximale se situant
vers le milieu de la longueur. Bord frontal tronqué (27 ex.) ou plus rarement
faiblement arrondi (8 ex.). Commissures toujours tranchantes, les valves se réunissant
suivant un angle aigu. Commissures latérales rectilignes. Commissure frontale
rectimarginée. Crochet court, non cręté, dressé. Foramen circulaire. Le
recourbement du crochet court masque le symphytium (8 ex.) ou bien celui-ci est
peu exposé (19 ex.). Les plaques deltidiales réunies en une pičce unique ne
sont visibles que sur 8 exemplaires. Le septum médian, visible ŕ travers le
test de 23 spécimens, s'étend sur la moitié de la longueur de la valve
dorsale.
Cette morphologie est
tout ŕ fait comparable ŕ celle des types figurés par Leymerie (1842),
mais avec des dimensions réduites d'environ de moitié : longueur moyenne :
11,3 mm pour 22 mm (intervalle de variation : 8,9-14 mm). Ces dimensions sont,
par contre, identiques ŕ celles des spécimens de Sardaigne données par Dieni
et al. (1973). Contrairement aux exemplaires de diverses localités
rapportés ŕ tort ŕ pseudojurensis par différents auteurs (voir
synonymie), il n'y a ni pli dorsal, ni dépression ventrale influant sur le
relief antérieur des valves chez la vraie pseudojurensis. Sur le contour
des coquilles, l'indentation frontale est également absente. L'espčce a été
rapportée au genre Belothyris Smirnova par Dieni et
al. (1973).
Extension verticale
et répartition géographique. Est du Bassin de Paris :
Hauterivien du département de l'Aube (Marolles, Vendeuvre, Avallon) (Leymerie,
1842). Hauterivien inférieur du Centre-Est de la Sardaigne (Puntŕ Doroné,
Lanaitto, vallée de l'Orudč, Dorgali) (Dieni et al.,
1973).
Valanginien de Zeghrine, ŕ l'Est de Beyrouth (Liban).
La faune de Brachiopodes de Zeghrine est surtout connue dans le Jura suisse et dans le Jura français, depuis les environs de Neuchâtel jusqu'ŕ Hauteville dans le Bugey (Jura méridional). Les principaux gisements en Suisse cités dans la littérature paléontologique se situent dans le Landeron, au Cressier, dans les environs d'Hauterive et de Valangin, la vallée de Nozon et pour le canton de Vaud, ŕ Arzier, Sainte-Croix et ŕ l'Auberson. Dans le Jura français, on peut citer les gisements de Villers-le-Lac, Metabief, Morteau (Doubs), de Censeau (Jura) ainsi que Vesancy prčs de Gex et Hauteville (Ain). Certaines espčces se retrouvent dans l'Est du Bassin de Paris (Belothyris pseudojurensis dans le département de l'Aube) ou en Haute-Provence, comme Terebratulina biauriculata dans les Marnes ŕ Térébratulines de l'arc de Castellane ou dans le département des Bouches-du-Rhône. Certains éléments de la faune ont également été signalés dans les Alpes bernoises, en Allemagne du Nord, en Crimée, dans les Carpathes centrales ou en Tchéquie (montagnes de Strŕzovsk) ainsi que dans le Centre-Est de la Sardaigne. La présence męme partielle de cette faune reste ŕ confirmer en Géorgie, au Daghestan, en Crimée, en Espagne et dans la Meseta marocaine occidentale (arričre-pays de Casablanca, Mazagan et Safi). Le lecteur pourra se reporter aux paragraphes extension verticale et répartition géographique des diverses espčces décrites dans ce travail.
L'âge global de la faune est Valanginien sensu lato (par exemple, les couches A et B d'Arzier in Loriol, 1862, 1864, 1868, datées du Valanginien inférieur --zones ŕ Pertransiens et ŕ Neocomiensiformis-- par Jan du Chęne et al., 2016) et męme Hauterivien en ce qui concerne celui donné par Lamellaerhynchia hauteriviensis. Si la faune débute bien dčs le Valanginien inférieur dans les Marnes d'Arzier, la plupart des espčces connaissent leur apogée dans les Calcaires ŕ Alectryonia, dans les Marnes ŕ bryozoaires ou encore dans les Marnes ŕ Astieria, formations toutes datées du Valanginien supérieur (zones ŕ Verrucosum, ŕ Trinodosum et ŕ Callidiscus pour Walter, 1989, Fig. 4). Les datations bien documentées plaident pour un âge Valanginien mais ne permettent pas distinguer le sous-étage.
Sur le plan paléoécologique, il faut noter l'abondance de coquilles jeunes ŕ Zeghrine. C'est le cas de Lamellaerhynchia hauteriviensis, de Loriolithyris valdensis, de Terebratulina biauriculata et de Belothyris pseudojurensis. La présence de plus grandes coquilles, voire d'adultes, exclut toute hypothčse de tri des coquilles par les courants avant leur ensevelissement. Ces observations témoignent de conditions environnementales peu favorables ŕ la fixation et au développement des organismes benthiques fixés comme les Brachiopodes : Forts apports de matériaux terrigčnes et hydrodynamisme assez faible ne permettant pas (ou peu) le renouvellement des apports trophiques. Une męme situation se retrouve dans le Valanginien de Villereversure (Ain) oů un abondant matériel a été collecté.
L'étude des
brachiopodes de Zeghrine et les extensions verticales des męmes espčces
connues par ailleurs (en particulier dans la couche 2 de Villereversure, Jura méridional)
permettent de dater la Formation de Salima du Valanginien sans plus de précision
(Fig. 5 ![]() ). En effet, aucune des deux hypothčses précédemment envisagées, i.e., un
âge Valanginien inférieur (Granier et al., 2016) ou un âge
Valanginien supérieur (Maksoud et al., 2020), ne peut ętre
confirmée ou définitivement rejetée.
). En effet, aucune des deux hypothčses précédemment envisagées, i.e., un
âge Valanginien inférieur (Granier et al., 2016) ou un âge
Valanginien supérieur (Maksoud et al., 2020), ne peut ętre
confirmée ou définitivement rejetée.
|
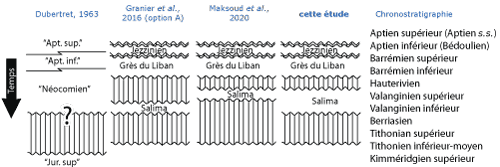
Figure 5
:
Les différentes interprétations des lacunes encadrant la Formation de Salima.
Cette unité lithostratigraphique qui n'apparaît pas dans la premičre
colonne correspondant ŕ l'interprétation de Dubertret
(1963) était
alors rapportée au Jurassique supérieur. La deuxičme colonne correspond ŕ
l'hypothčse préférée de Granier et al. (2016) proposant un
âge Valanginien inférieur et la troisičme colonne correspond ŕ l'hypothčse
d'un âge Valanginien supérieur avancée par Maksoud et al.
(2020). La quatričme colonne présente notre interprétation basée sur la
seule faune de brachiopodes de Zeghrine, interprétation plus ouverte que les
deux précédentes, mais ne les excluant pas. Légende : "Jur. sup." =
"Jurassique supérieur" ; "Apt. inf." = "Aptien inférieur"
; "Apt. sup." = "Aptien supérieur". |
Le premier auteur (Y.A.) remercie P. Binard pour son assistance efficace dans le montage de la planche photographique. L'ensemble des auteurs s'accorde pour adresser leurs remerciements ŕ Philippe Fauré et Sibelle Maksoud pour leurs suggestions, męme si toutes ces derničres n'ont pas nécessairement été prises en considération. Ainsi, les opinions exprimées dans la version publiée de cet article n'engagent que leurs auteurs et ne reflčtent pas nécessairement le point de vue des rapporteurs.
Ager D.V. (1964).- Further new rhynchonelloid brachiopods from the Texas Cretaceous.- Journal of Paleontology, Tulsa, vol. 38, nº 1, p. 109-112.
Ager D.V. (1965).- Mesozoic and Cenozoic Rhynchonellacea. In : Moore R.C. (ed.), Part H. Brachiopoda, 2.- Treatise on Invertebrate Paleontology, Geological Society of America, Boulder-CO ; University Kansas Press, Lawrence-KS, p. H597-H632.
Ascher E. (1906).- Die Gastropoden, Bivalven und Brachiopoden der Grödischter Schichten.- Beiträge zur Paläontologie und Geologie Osterreich-Ungarns und des Orients, Wien, Band XIX, p. 135-172.
Bechon F., Decrouez D. & Villoutreys O. de (1984).- Les collections du Département de Géologie et de Paléontologie des Invertébrés du Museum d'Histoire naturelle de Genčve. 7. La collection P.J. Pictet (Annelida, Bryozoa, Brachiopoda, Arthropoda, Bivalvia).- Revue de Paléobiologie, Genčve, vol. 3, nº 2, p. 281-296.
Bischoff G. (1964).- Ostracoden-Studien im Libanon, 3: Die Gattung Cytherelloidea im Oberen Jura und in der Unterkreide.- Senckenbergiana Lethaea, Frankfurt am Main, Band 45, p. 1-27.
Bischoff G. (1990a).- Ostracoden-Studien im Libanon, 4: The genus Hutsonia in the Late Jurassic.- Senckenbergiana Lethaea, Frankfurt am Main, Band 70, p. 397-429.
Bischoff G. (1990b).- Ostracoden-Studien im Libanon, 5: The genus Schuleridea in the Late Jurassic and Early Cretaceous.- Senckenbergiana Lethaea, Frankfurt am Main, Band 71, p. 85-137.
Bogdanova T.N. & Lobatcheva S.V. (1966).- Le Néocomien du Kopet-Dag. Livre 2 : La faune du Néocomien du Kopet-Dag.- Travaux de l'Institut de Recherches Géologiques pour toute l'Union (VSEGEI) (n.s.), Léningrad, vol. 130, 140 p.
Burri F. (1953).- Beiträge zur Systematik der Brachiopoden aus der untersten Kreide im westschweizerischen Juragebirge.- Eclogae geologicae Helvetiae, Bâle, vol. 46, nº 2, p. 269-285.
Burri F. (1954).- Die Morphogenese von Sulcirhynchia valangiensis (Brachiopoda).- Eclogae geologicae Helvetiae, Bâle, vol. 47, nº 2, p. 419-425.
Burri F. (1956).- Die Rhynchonelliden der Unteren Kreide (Valanginien-Barrémien) im westschweizerischen Juragebirge.- Eclogae geologicae Helvetiae, Bâle, vol. 49, nº 2, p. 599-701.
Calzada Badia S. (1975).- Braquiopodos infracretŕcicos del Levante espańol.- Trabajos del Museo geologico del Seminario de Barcelona (seccion de Biostratigrafia del C.S.I.C.), vol. 14, public. nº 149, 71 p.
Calzada Badia S. (1983).- Dos braquiopodos hauterivienses de San Mateo (Castellon, Espańa).- Boletín de la Real Sociedad Espańola de Historia Natural (Geologia), Madrid, vol. 81, nº 3-4, p. 141-151.
Cooper G.A. (1973).- Fossil and Recent Cancellothyridacea (Brachiopoda).- Scientific Report of Tohoku University (2nd ser., Geol.), spec. vol. 6, Hatai Memorial volume, p. 371-390.
Cooper G.A. (1983).- The Terebratulacea (Brachiopoda), Triassic to Recent: A study of the brachidia (loops).- Smithsonian Contributions to Paleobiology, Washington D.C., vol. 50, 445 p.
Davidson T. (1851-1854).- On the classification of the Brachiopoda.- Palaeontographical Society, London, vol. 7, 136 p.
Davidson T. (1874).- A monograph of the British Fossil Brachiopoda. vol. 4, pt. 1. Supplement to the Recent, Tertiary and Cretaceous species.- Palaeontographical Society, London, vol. 27, 72 p.
Deninger K. (1907).- Die mesozoischen Formationen auf Sardinien.- Neues Jarhrbuch für Mineralogie, Geologie und Paleontologie, Stuttgart, Band 23, p. 435-473.
Deslongchamps E. & Deslongchamps E.E. (1855).- Introduction ŕ l'histoire naturelle des Brachiopodes vivants et fossiles.- Mémoires de la Société Linéenne de Normandie, Caen, vol. 10, p. 69-271.
Deslongchamps E.E. (1862-1885).- Paléontologie française. Terrains jurassiques. Vol. 6 : Brachiopodes.- Masson édit., Paris, 448 p.
Dieni I., Middlemiss F.A. & Owen E.F. (1973).- The Lower Cretaceous Brachiopods of East-Central Sardinia.- Bolletino della Societŕ Paleontologica Italiana, Modena, vol. 12, nº 2, p. 166-216.
Douvillé H. (1879).- Note sur quelques genres de Brachiopodes (Terebratulidae et Waldheimiidae).- Bulletin de la Société géologique de France (sér. 3), Paris, t. VII, p. 251-277.
Dubertret L. (1951).- Carte géologique au 50.000e. Feuille de Beyrouth.- Institut Géographique National, Paris, 66 p. (XII Pls.).
Dubertret L. (1963).- A. Liban et Syrie. Chaîne des grands massifs côtiers et confins ŕ l'Est. In : Dubertret L. (ed.), Liban, Syrie, Jordanie.- Lexique Stratigraphique International (Asie), Paris, vol. III, fasc. 10 c1, p. 7-155.
Ferry S., Merran Y., Grosheny D. & Mroueh M. (2007).- The Cretaceous of Lebanon in the Middle East (Levant) context. In : Bulot L.G., Ferry S. & Grosheny D. (eds.), Relations entre les marges septentrionale et méridionale de la Téthys au Crétacé [Relations between the northern and southern margins of the Tethys ocean during the Cretaceous period].- Carnets Geol., Madrid, vol. 7, nº M02, Abstract 08 (CG2007_M02/08), p. 38-42. DOI : 10.4267/2042/8029
Gaspard D. (1988).- Sellithyridinae, Terebratulidae du Crétacé d'Europe occidentale - Dynamique des populations, systématique et évolution.- Cahiers de Paléontologie, Paris, 243 p.
Gigout M. (1951).- Études géologiques sur la Meseta marocaine occidentale (arričre-pays de Casablanca, Mazadan et Safi).- Notes et Mémoires du Service géologique du Maroc, Rabat, vol. 86, 507 p.
Granier B., Toland C., Gčze R., Azar D. & Maksoud S. (2016).- Some steps toward a new story for the Jurassic - Cretaceous transition in Mount Lebanon.- Carnets Geol., Madrid, vol. 16, no. 8, p. 247-269.
Jacob C. & Fallot P. (1913).- Étude sur les Rhynchonelles portlandiennes, néocomiennes et méso-crétacées du Sud-Est de la France.- Mémoires de la Société Paléontologique Suisse, Genčve, vol. 39, 82 p.
Jan du Chęne R., Monteil E., Charollais J., Granier B. & Clavel B. (2016).- Étude palynologique du sondage du Morand (commune de Montricher, canton de Vaud, Suisse). Lithologie, inventaire paléontologique et biostratigraphie du Crétacé inférieur du Jura vaudois.- Archives des Sciences, Genčve, vol. 68 (2015), p. 117-162.
Karakasch N.I. (1907).- Le Crétacé inférieur de la Crimée et sa faune.- Travaux de la Société Impériale des Naturalistes de Saint-Pétersbourg, vol. 32, nº 5, 482 p.
Leymerie A. (1842).- Suite du mémoire sur le terrain crétacé du département de l'Aube. 2čme partie : Partie paléontologique.- Mémoires de la Société géologique de France (1čre série), Paris, vol. 5, mém. nº 1, p. 1-34.
Lobatscheva S.V. (1974).- Sur le genre Cyclothyris (Brachiopoda).- Revue de Paléontologie, S.S.S.R., vol. 1, p. 72-82. Traduction : Paleontological Journal, Washington D.C., vol. 1, p. 67-77.
Loriol P. de (1862).- Description des animaux invertébrés fossiles contenus dans l'étage Néocomien moyen du Mont Salčve.- N. Georg édit., Genčve, 214 p.
Loriol P. de (1864).- Description de quelques brachiopodes crétacés.- Mémoires de la Société de Physique et d'Histoire naturelle de Genčve, vol. 17, nº 2, p. 437-447.
Loriol P. de (1868).- Monographie des couches de l'étage Valangien des carričres d'Arzier (Vaud).- Matériaux pour la Paléontologie Suisse (4čme sér.), Genčve, Livres 10 et 11, 110 p.
Maksoud S., Granier B., Gčze R., Alméras Y., Toland C. & Azar D. (2020).- The Jurassic/Cretaceous boundary in Lebanon. Revision of the Salima Formation.- Cretaceous Research, vol. 107, article 104268, p. 1-8.
Mallada L. (1887).- Sinopsis de las especies fosiles que se han encontrando en Espańa. III : Terreno mesozoico (Cretaceo inferior).- Commission del Mapa geologica de Espańa, Madrid, vol. 14, 171 p.
Middlemiss F.A. (1959).- English Aptian Terebratulidae.- Palaeontology, London, vol. 2, nº 1, p. 94-142.
Middlemiss F.A. (1968).- Brachiopodes du Crétacé inférieur des Corbičres orientales (Aude).- Annales de Paléontologie, Paris, Invertébrés, vol. 54, nº 2, p. 173-197.
Middlemiss F.A. (1976).- Lower Cretaceous Terebratulidina of Northern England and Germany and their geological background.- Geologische Jarhbuch, Hannover, A30, p. 21-104.
Middlemiss F.A. (1980).- Lower Cretaceous Terebratulidae from south-western Morocco and their biogeography.- Palaeontology, London, vol. 23, nº 3, p. 515-556.
Middlemiss F.A. (1981).- Lower Cretaceous Terebratulidae of the Jura region. 1. Revision of some species described by Pictet and de Loriol.- Eclogae geologicae Helvetiae, Bâle, vol. 74, nº 3, p. 701-733.
Muir-Wood H.M. (1965).- Mesozoic and cenozoic Terebratulidina. In : Moore R.C. (ed.), Part H. Brachiopoda, 2.- Treatise on Invertebrate Paleontology, Geological Society of America, Boulder-CO ; University Kansas Press, Lawrence-KS, p. H762-H816.
Muir-Wood H.M., Elliott G.F. & Hatai K. (1965).- Mesozoic and Cenozoic Terebratellidina. In : Moore R.C. (ed.), Part H. Brachiopoda, 2.- Treatise on Invertebrate Paleontology, Geological Society of America, Boulder-CO ; University Kansas Press, Lawrence-KS, p. H816-H857.
Nekvasilovŕ O. (1978).- Cancellothyridacea (Brachiopoda) from the Lower Cretaceous of Stramberg (Czechoslovakia).- Casopi pro Mineralogii a Geologi, Praha, vol. 23, nº 2, p. 171-182.
Noujaim Clark G. & Boudagher-Fadel M.K. (2001).- The larger benthic Foraminifera and stratigraphy of the Upper Jurassic/Lower Cretaceous of Central Lebanon.- Revue de Micropaléontologie, Paris, vol. 44, nº 3, p. 215-232.
Ooster W.A. (1863).- Pétrifications remarquables des Alpes suisses. Synopsis des Brachiopodes fossiles des Alpes suisses.- H. Georg édit., Genčve, 71 p.
Orbigny A. d' (1847-1850).- Paléontologie française. Animaux Mollusques et Rayonnés de France. Terrains crétacés. 4 : Brachiopodes.- Masson édit., Paris, 390 p.
Orbigny A. d' (1850).- Prodrome de Paléontologie stratigraphique universelle des Animaux Mollusques et Rayonnés (faisant suite ŕ un cours élémentaire de Paléontologie et de Géologie stratigraphique).- Masson édit., Paris, vol. 1, 394 p.
Owen E.F. (1956).- The Lower Cretaceous Brachiopods "Rhynchonella" gibbsiana (J. de C. Sowerby) and Sulcirhynchia hythensis sp. nov.- The Annals and Magazine of Natural History (ser. 12), London, vol. 9, p. 164-172.
Owen E.F. (1970).- A revision of the brachiopod subfamily Kingeninae Elliott.- Bulletin of the British Museum (Natural History) (Geology), London, vol. 19, nº 2, p. 27-83.
Owen E.F. (1973).- The distribution of Lower Cretaceous (Berriasian-Barremian) rhynchonelloid and terebratelloid brachiopods in the northern hemisphere. In : Casey R. & Rawson P.F. (ed.), The Boreal Lower Cretaceous.- Geological Journal, Special Issue, vol. 5, p. 121-130.
Pajaud D. (1974).- Rhynchonelles et Térébratules du Crétacé de la région de Castellane (Haute-Provence).- Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon, vol. 61, p. 91-116.
Pictet P.J. & Campiche M. (1872).- Description des fossiles du terrain crétacé des environs de Sainte-Croix. Classe des Mollusques Brachiopodes. 5čme partie (terminée par P. de Loriol).- Matériaux pour la Paléontologie Suisse, Genčve, 158 p.
Popkhadzé M.V. (1960).- Quelques brachiopodes crétacés inférieurs du Caucase.- Travaux de l'Institut de Paléobiologie, Académie des Sciences de la République de Géorgie, Tbilissi, vol. 5, p. 107-160.
Roch É. (1930).- Études géologiques dans la région méridionale du Maroc occidental.- Notes et Mémoires du Service des Mines du Maroc, Rabat, vol. 9, 542 p.
Roemer F.A. (1841).- Versteinerungen des norddeutschen Kreidegebirges.- Hahn édit., Hannover, 145 p.
Schloenbach U. (1866).- Ueber die Brachiopoden aus dem unteren Gault (Aptien) von Ahaus in Westphalen.- Zeitschrift der deutschen Geologischen Gesellschaft, Stuttgart, vol. 18, nº 2, p. 364-376.
Siblik M. (1962).- Brachiopodes du Jurassique et du Crétacé inférieur des Carpathes centrales.- Geologické Prŕce, Bratislava, vol. 26, p. 273-277.
Siblik M. (1963).- Découverte de brachiopodes dans les tuffites du Néocomien de Kosécké Rovné.- Geologické Prŕce, Bratislava, vol. 27, p. 69-74.
Smirnova T.N. (1960).- Sur une sous-famille nouvelle de Dallinidés du Crétacé inférieur.- Revue de Paléontologie, Moscou, vol. 2, p. 114-120.
Smirnova T.N. (1966).- Systematics of Early Cretaceous terebratulids of subfamily Cancellothyridinae.- Revue de Paléontologie, S.S.S.R., vol. 3, p. 31-41. Traduction : International Geology Review, U.S.A., vol. 9, nº 4, p. 540-548.
Smirnova T.N. (1972).- Brachiopodes du Crétacé inférieur de Crimée et du Caucase septentrional.- Nauka édit., Moscou, 143 p.
Toland C. (2000).- A sequence stratigraphic reference section for the Tithonian of Lebanon. In : Alsharhan A.S. & Scott R.W. (eds.), Middle East models of Jurassic/Cretaceous carbonate systems.- SEPM Special Publication, vol. 69, p. 53-64.
Walker J.F. (1868).- On the species of Brachiopoda, which occur in the Lower Greensand at Upware.- Geological Magazine, London, vol. 5, nº 51, p. 399-407.
Walter B. (1989).- Au Valanginien supérieur, une crise de la faune de bryozoaires : Indication d'un important refroidissement dans le Jura.- Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 74, p. 255-263.
Walter B. (1991).- Cavidés néocomiens (Bryozoa, Cyclostomata).- Geobios, Lyon, vol. 24, nº 3, p. 289-308.
Wetzel & Dubertret L. (1951).- Notice explicative de la Carte géologique du Liban au 200.000e.- Ministčre des Travaux Publics, Beyrouth, 62 p.
Planche
1 : |
|